charakterisierung eines in vivo modells zur untersuchung ... · aus dem lehrstuhl für anatomie ii...
TRANSCRIPT

Aus dem Lehrstuhl für Anatomie II
der
Friedrich-Alexander-Universität Erlangen-Nürnberg
Direktorin: Prof. Dr. Elke Lütjen-Drecoll
Charakterisierung eines in vivo Modells zur
Untersuchung der Wimpernbiologie und Trichomegalie:
Mäusewimpern Morphologie, Entwicklung,
Wachstumszyklus und Prolongation von Anagen durch
Bimatoprost
Inaugural-Dissertation
Zur Erlangung der Doktorwürde
der Medizinischen Fakultät
der
Friedrich-Alexander-Universität
Erlangen-Nürnberg
Vorgelegt von
Tobias Andreas Fuchs
aus
Regensburg

Gedruckt mit Erlaubnis der
Medizinischen Fakultät der Friedrich-Alexander-Universität
Erlangen Nürnberg
Dekan: Prof. Dr. med. Dr. h.c. Jürgen Schüttler
Referent: Prof. Dr. med. Elke Lütjen-Drecoll
Korreferent: Prof. Dr. med. Michael Eichhorn
Tag der mündlichen Prüfung: 20. Juli 2011

Meinen Eltern Inge und Bernhard Fuchs

Inhaltsverzeichnis
Inhaltsverzeichnis
1. Zusammenfassung und Abstract 1
1.1 Zusammenfassung 1
1.2 Abstract 2
2. Einleitung 4
2.1 Trichomegalie und Wimpernwachstum 4
2.2 PGF-2α Analoga 5
2.2.1 Chemie von PGF-2α Analoga 5
2.2.2 Haarwachstum stimulierende Potenz von PGF-2α 8
2.3 Ein Tiermodell zur Untersuchung des Wimpernwachstums 9
2.4 Einfluss von Bimatoprost auf das Wimpernwachstum bei
der Maus 10
3. Material und Methodik 11
3.1 Tiere 11
3.2 Probengewinnung 11
3.3 Bimatoprostbehandlung 12
3.3.1 Makroskopische Untersuchung 13
3.3.2 Histologische Untersuchung 14
3.4 Statistische Analyse 15
4. Ergebnisse 16
4.1 Morphologie des Augenlids 16
4.2 Morphologie der Wimper 18
4.2.1 Histologischer Aufbau des Wimpernfollikels 18
4.2.2 Makroskopische Beschreibung des Wimpernschaftes 18
4.3 Unterteilung des Wimpernwachstumszyklus 19
4.3.1 Telogen 20
4.3.2 Anagen-early 21
4.3.3 Anagen-middle 22
4.3.4 Anagen-late 22
4.3.5 Katagen-early 23

Inhaltsverzeichnis
4.3.6 Katagen-late 25
4.4 Morphogenese und erster Zyklus der Wimper 27
4.4.1 Makroskopie 27
4.4.2 Histologie 27
4.4.2.1 Morphogenese in Woche 0 post partum 27
4.4.2.2 Morphogenese in Woche 1 post partum 29
4.4.2.3 Beginn des ersten Wimpernzyklus in Woche 2 post
partum 30
4.4.2.4 Woche 3 post partum 30
4.4.2.5 Woche 4 post partum 31
4.4.2.6 Woche 5 post partum 31
4.4.2.7 Woche 6 post partum 32
4.5 Bimatoprostbehandlung 33
4.5.1 Makroskopische Untersuchung 33
4.5.2 Histologische Untersuchung 36
5. Diskussion 40
5.1 Vergleich der Morphologie der Wimpern der Maus mit
menschlichen Wimpern und mit Fellhaaren 40
5.2 Vergleich der Morphogenese und der frühen Entwicklung der
Wimpern mit der der Fellhaare 40
5.3 Vergleich des Wachstumszyklus und des Exogens der
Wimper mit der dorsaler Fellhaare 41
5.4 Effekte von Bimatoprostbehandlung 41
5.5 Zunahme der Wimperndicke und Hyperpigmentierung
durch Bimatoprost 42
5.6 Wimpernverlängerung durch Bimatoprost 43
6. Literaturverzeichnis 44
7. Abkürzungsverzeichnis 49
8. Verzeichnis der Vorveröffentlichungen 51
9. Danksagung 52

1
Zusammenfassung
1. Zusammenfassung und Abstract
1.1 Zusammenfassung
Hintergrund und Ziele: Hypertrichosis oder Alopezie der Wimpern ist sowohl
mit unterschiedlichen Krankheiten als auch mit bestimmten Therapeutika
assoziiert. Veränderungen der Wimpern können optisch störend und damit,
gerade weil die Gesichtsregion betroffen ist, psychisch belastend sein und zu
Schäden am Auge, zu Therapieabbrüchen und zu Non-Compliance führen. Da
die Mechanismen des Wimpernwachstums und die damit zugrundeliegenden
Ursachen der Wimpernhypotrichose oder Wimpernhypertrichose weitgehend
unklar sind, ist auch die Therapie limitiert. Ziel dieser Studie war es zunächst zu
untersuchen, ob die Maus ein Tiermodell für das Wimpernwachstum darstellt
und wie der Wimpernzyklus sich zu dem Haarzyklus der Maus verhält. Bei
menschlichen Augen induziert die Behandlung mit Prostaglandin F-2α, das zur
Augeninnendrucksenkung bei Glaukompatienten angewandt wird, eine
Hypertrichose der Wimpern. In einem zweiten Versuchsaufbau wurde dann die
pharmakologische Beeinflussung des Wimpernfollikels durch Behandlung mit
dem Prostaglandin F-2α Analog Bimatoprost untersucht.
Methoden: Obere Augenlider und definierte Abschnitte dorsaler Rückenhaut
wurden weiblichen 0,1,2,3,4,5 und 6 Wochen alten C57BL-6J Mäusen
entnommen (pro Gruppe=2-4). Die Wimpern der rechten Augenlider wurden
unter dem Stereomikroskop gezählt. Die linken Augenlider und die Haut wurden
in Epon eingebettet und 1µm dicke, mit Toluidinblau gefärbte Serienschnitte
(160 Schnitte; 160µm/Augenlid) zur histologischen Beschreibung der
Morphogenesestadien und des ersten Wachstumszyklus angefertigt.
Die rechten Augenlider von 4 Wochen alten weiblichen C57BL-6J Mäusen
wurden 14 Tage mit 0,03% Bimatoprost bzw. mit Vehikel behandelt (pro
Gruppe=10). Die Wimpern der entnommenen oberen Augenlider wurden
gezählt und der basale Durchmesser vermessen. Serienschnitte wurden
angefertigt (160 Schnitte; 160µm/Augenlid). Alle Wimpernfollikel innerhalb der
160 Serienschnitte wurden gezählt, einer Wachstumsphase zugeordnet, der
Durchmesser des Bulbus und der dermalen Papille vermessen und die Anzahl
der Melaningranula quantifiziert.

2
Zusammenfassung
Ergebnisse und Beobachtungen: Augenlider von Mäusen wiesen die
gleichen anatomischen Strukturen auf wie humane Augenlider. Die generellen
morphologischen Eigenschaften der Wimpern von Mäusen (Schaft, Follikel,
Morphogenese und Wachstumszyklus) waren mit denen der dorsalen Fellhaare
vergleichbar. Es fehlte jedoch der M. arrector pili. Nach der Morphogenese der
Mäusewimpern wurde ein synchroner Zyklus beobachtet, welcher früher
begann und schneller voranschritt, als bei den dorsalen Fellhaaren. Exogen
fand während jedes Zyklus in Katagen statt. Bimatoprostbehandlung
prolongierte signifikant die Dauer von Anagen und induzierte die Bildung
längerer und dickerer Wimpern. Es gab keine Anzeichen für eine durch
Bimatoprost induzierte de-novo Follikulogenese.
Praktische Schlussfolgerungen: Wimpern von Mäusen eignen sich
hervorragend als gut standardisierbares in-vivo-Modell zur quantitativen und
qualitativen Analyse der Morphologie, der Morphogenese, des
Wachstumszyklus und der Bestimmung des Zeitpunktes von Exogen. Anhand
dieses Wimpernmodells können molekulare Kontrollmechanismen des
Wachstumszyklus analysiert und neuartige Medikamente zur Therapie von
Wimpernfehlregulationen entwickelt werden. Die durch Bimatoprost induzierte
Hypertrichose der Wimpern beruht wahrscheinlich auf einer Verlängerung von
Anagen, so dass vermehrt Follikel mit zwei Wimpern pro Follikel verblieben.
1.2 Abstract
Background and intention: Hypertrichosis or alopecia of eyelashes can be
associated with various diseases or treatments. Eyelash disorders can be
optically disturbing and thus, due to the affection of the face region,
psychologically strain. In addition, changes in number and morphology of
eyelashes can result in eye damage, discontinuation of treatment or non-
compliance. As mechanisms of growth controls of eyelashes and the underlying
causes of eyelash hypo- or hypertrichosis are largely obscure, available therapy
is limited. The intention of this study was, to investigate whether the mouse can
serve as a model for eyelash morphogenesis and to compare the eyelash cycle
with the hair cycle of the mouse. In a second experimental setup changes in

3
Zusammenfassung
eyelash follicle biology were described following treatment with Bimatoprost, an
antiglaucoma drug, that induces hypertrichosis of eyelashes in a number of
patients.
Methods: Upper eyelids and defined regions of dorsal skin were excised from
female C57BL-6J mice at 0,1,2,3,4,5 and 6 weeks of age (2-4/age). Eyelashes
of right eyelids were counted under a stereomicroscope. Left eyelids were
embedded in epon and serial sections of 1µm (160 sections; 160µm/eyelid)
were prepared and stained with toluidineblue to describe histologically
morphogenesis and the first growth cycle.
Right eyelids of 4 week old female C57BL-6J mice were treated with 0,03%
bimatoprost or its vehicle (10/each). Eyelashes of excised eyelids were counted
and basal thickness was measured. Serial sections were prepared (160
sections; 160µm/eyelid). Eyelash follicles within this sections were counted,
growth phase was determined, diameter of the bulb and the dermal papilla was
measured and number of melanin granules was quantified.
Results and observations: Mouse eyelids exhibited the same anatomic
features as human eyelids. General morphological features of mouse
eyelashes, regarding to shaft, follicle, morphogenesis, and growth cycle, were
comparable to those of pelage hair follicles. Eyelashes had no m. arrector pili.
After morphogenesis of mouse eyelashes, a synchronized cycle was observed,
that initiated earlier and progressed faster, compared to pelage hairs. Eyelash
exogen occurred during each cycle during catagen. Treatment with bimatoprost,
significantly extended the duration of anagen and induced generation of longer
and thicker eyelashes. There was no evidence of bimatoprost-induced de-novo
folliculogenesis.
Conclusions: Mouse eyelashes offer an excellent in-vivo-model for the
quantitative and qualitative analysis of eyelash morphology, morphogenesis,
growth cycle and determination of exogen. This model will help to analyze the
molecular controls of eyelash growth and to develop novel drugs to treat
eyelash disorders. Hypertrichosis following treatment with bimatoprost is
presumably due to an extended anagen, leading to an increased number of two
shaft follicles.

4
Einleitung
2. Einleitung
2.1 Trichomegalie und Wimpernwachstum
Hypertrichosis der Wimpern oder Trichomegalie ist definiert als Längen-,
Dicken-, Steifigkeits- und Pigmentzunahme sowie die Neigung der Wimpern,
sich zu kringeln.[58] Dabei tritt Trichomegalie oft in Verbindung mit einer
steigenden Anzahl der Wimpern, einer additiven Reihe und erhöhtem Übergang
von Vellushaar zu terminalem Haar auf.[58] 1965 wurde Trichomegalie erstmals
als kongenitaler Symptomkomplex von Oliver-McFarlane beschrieben.[46] Heute
ist sowohl die Assoziation von Trichomegalie mit vielen Krankheitsbildern
bekannt, als auch das Auftreten von Trichomegalie als Nebenwirkung von
systemischen und lokalen Therapeutika. So tritt eine Trichomegalie bei
okulokutanem Albinismus Typ I,[47] sowie bei systemischen Erkrankungen wie
Dermatomyositis,[64] systemischem Lupus Erythematosus,[57] HIV Typ I[30] oder
lokalen Infektionen wie Uveitis[2] auf. Auch systemische Therapie mit Interferon-
alpha,[22] Topamirat,[58] Immunsuppressoren wie Cyclosporin[77] oder
Tacrolimus,[76] Therapie mit Anti-EGRF (epidermal growth factor receptor)
rekombinanten Antikörpern wie Cetuximab[31] oder EGFR-
Tyrosinkinaseinhibitoren wie Erlotinib[4] oder Gefitinib[47] kann Trichomegalie
verursachen. Auch lokale Applikation von Cyclosporin[51] als Augentropfen kann
zu Trichomegalie führen.
In den letzten Jahren wurde eine Trichomegalie durch Therapie des Glaukoms
mit lokalen PGF-2α Analoga beschrieben. Bei lokaler antiglaukomatöser
Therapie mit Latanoprost[14] und mit Bimatoprost[6],[20],[21],[63],[71] trat
Trichomegalie als unerwünschter Nebeneffekt auf (Abbildung 1).
Abbildung 1: Das
linke mit dem
PGF-2α Analogon
Bimatoprost be-
handelte Auge
zeigt unilaterale
Trichomegalie im
Gegensatz zum unbehandelten rechten Auge. Quelle: Tosti A., Pazzaglia M.,
Voudouris S. and Tosti G. (2004). "Hypertrichosis of the eyelashes caused by
bimatoprost." J Am Acad Dermatol 51: S149-50. [71]

5
Einleitung
Eine Fehlregulation des Wimpernwachstums kann optisch störend und damit,
gerade weil die Gesichtsregion betroffen ist, psychisch belastend sein und zu
Non-Compliance bis hin zum Therapieabbruch führen. Auch wurde ein
unnötiger Verbrauch des Medikaments aufgrund des erschwerten
Zugangsweges durch zu lange Wimpern beschrieben.[63]
Andererseits könnten gerade diese Trichomegalie induzierenden Substanzen
aufgrund der Haarwachstum stimulierenden Potenz als Medikamente zur
Therapie von Wimpernwachstumsstörungen verwendet werden.[8] So könnten
PGF-2α Analoga zum Beispiel zur Therapie der Alopezie von Wimpern,[35] wie
auch der Milphosis, einer lokalen Alopecia areata der Wimpern,[56] eingesetzt
werden. Zusätzlich zur psychischen Belastung sind bei diesen Erkrankungen,
infolge des Verlustes der Schutzfunktion der Wimpern vor Sonnenlicht oder
Partikeln, auch Schäden der Kornea möglich. Ursachen von Milphosis, dem
alleinigen Verlust der Wimpern oder Madarosis, einem kombinierten Wimpern
und Augenbrauenverlust sind beispielsweise Infektion, Trauma oder Allergie.[56]
Auch wurde Madarosis bei Autoimmunkrankheiten wie dem diskoiden Lupus
Erythematosus[62] und Milphosis bei endokrinologischen Erkrankungen wie
Hyperthyreose[29] oder bei kongenitalen Erkrankungen[11] beschrieben. Häufig
ist der Ausfall von Wimpern Folge von Chemotherapie[40] oder Botulinum
Injektion[32].
Der Mechanismus, durch den sowohl die Trichomegalie als auch der
Wimpernverlust verursacht wird, ist bisher nicht einheitlich geklärt. Auch der
Einfluss von Prostaglandinen wird widersprüchlich beschrieben. In einzelnen
klinischen Falldarstellungen wird eine erfolgreiche Therapie von Alopecia areata
der Wimpern durch lokale Applikation von PGF-2α Analoga dargestellt,[35],[37] in
anderen Studien konnte ein Wiederwachsen von Wimpern durch Therapie mit
PGF-2α Analoga nicht nachgewiesen werden.[54]
2.2 PGF-2α Analoga
2.2.1 Chemie von PGF-2α Analoga
Die bisher unter dem Terminus PGF-2α Analoga zusammengefasste Gruppe
chemischer Strukturen ist eine Sammelbezeichnung für Gewebshormone, die
ihre Namensgebung Ulf Svante von Euler verdanken, welcher diese 1934

6
Einleitung
erstmals aus der Prostata extrahierte.[17] 1960-1962 wurden sie von Sune
Bergström entsprechend ihrer Löslichkeit in Ethylether oder Phosphat in PGE
oder PGF unterteilt.[3] Weitere Prostaglandine wurden später u.a. 1973 von B.
Samuelsson entdeckt.[19] 1982 erhielten Sune Bergström, Bengt Samuelsson
und John Vane den Nobelpreis für ihre Entdeckungen.[17] Prostaglandine
werden auf einem der drei Hauptwege der Arachidonsäurekaskade, dem
sogenannten Cyclooxigenase-Pathway, synthetisiert (Fig. 1).
Fig. 1: Prostaglandinsynthese. Arachidonsäure wird durch eine Phospholipase aus
Phospholipiden der Zellmembran freigesetzt. Aus freier Arachidonsäure werden in
Abhängigkeit von den Enzymen der unterschiedlichen Zellen Prostaglandin H2 (PGH2)
Leukotriene und Epoxyeicosatriensäuren synthetisiert. PGH2 ist Substrat der
Prostaglandinsynthasen (PG-S), Thromboxansynthase (Tx-S) und
Prostacyclinsynthase (PGI2-S). Modifiziert aus Quelle Smith W.L., Marnett l.J. and
DeWitt D.L. (1991). „Prostaglandin and thromboxane biosynthesis.“ Pharmacol Ther
49: 153-79 [65]
Neben den zur Therapie von Glaukom eingesetzten Prostaglandin-Analoga,
zählt das zur Anregung des Wimpernwachstums eingesetzte Therapeutikum
Bimatoprost durch Substitution einer Ethylamid-Gruppe an C-1 zur chemischen
Subgruppe der Prostamide.[79] Parallel zur Synthese von Prostaglandin-G2 aus
Arachidonsäure bei den Prostaglandinen wird Prostamid-G2 aus Anandamid
(=Arachidonylethanolamid) durch eine Cyclooxygenase synthetisiert.[79]
Während Arachidonsäure durch beide Isoenzyme COX-1 und COX-2 zu PGG2
oxidiert werden kann, erfolgt die Oxidierung von Anandamid nur durch COX-

7
Einleitung
2.[79] Analog zur Prostaglandinsynthese dient Prostamid H2 als Substrat für
verschiedene Prostamide, darunter Prostamid-F2α (Fig. 2).[79] Bimatoprost bindet
an den Prostaglandin-FP-Rezeptor, interagiert aber möglicherweise mit dem
FP-altFP-Rezeptor Heterodimer, Splicing Varianten des FP-Rezeptors.[33]
Fig. 2: Prostaglandin- und Prostamidsynthese im Vergleich. Anandamid dient im
Gegensatz zur Arachidonsäure nicht den beiden Isoenzymen Cycloxygenase-1 und -2
(COX-1,-2) als Substrat, sondern nur COX-2. Modifiziert aus Quelle Woodward D. F.,
Liang Y. and Krauss A. H. (2008). "Prostamides (prostaglandin-ethanolamides) and
their pharmacology." Br J Pharmacol 153: 410-9. [79]
Bei einer steigenden Tendenz der Prävalenz des primären
Offenwinkelglaukoms aufgrund der demographischen Entwicklung, erfahren die
PGF-2α Analoga eine häufige Anwendung, um eine der weltweit häufigsten
Ursachen für Blindheit infolge der Degeneration des N. Optikus zu reduzieren.[5]
Dabei senken diese durch eine Erhöhung vor allem des uveoskleralen sowie
des trabekulären Kammerwasserabflusses,[25] u.a. in Folge von Aktivierung der
Matrix-Metalloproteinasen,[45] den Augeninnendruck. Die Innovation
Prostaglandine als Therapeutika für Glaukom zu verwenden entwickelten Bito

8
Einleitung
und Camras in den 1980igern.[25] Heute zählen zu den in der
antiglaukomatösen Therapie eingesetzten PGF-2α Analoga u.a. Prostaglandin
F2α-Isopropyl-Ester, Latanoprost, Travoprost, Bimatoprost und Tafluprost (Fig.
3).[25]
Fig. 3: Antiglaukomatöse Prostaglandin Derivate.
Prostaglandin F2α-Isopropyl-Ester, Latanoprost, Travoprost, Bimatoprost (Substitution
einer Ethylamid-Gruppe an C-1 und Substitution einer Phenylgruppe) und Tafluprost.
Modifiziert aus Quelle: Ishida N., Odani-Kawabata N., Shimazaki A. and Hara H.
(2006). "Prostanoids in the therapy of glaucoma." Cardiovasc Drug Rev 24: 1-10 [25]
2.2.2 Haarwachstum stimulierende Potenz von PGF-2α Analoga
Dass PGF-2α Analoga nicht nur Augeninnendruck senkende, sondern auch
Haarwachstum stimulierende Potenz aufweisen, wurde erstmals 1997
entdeckt.[8],[27] Sogar erfolgreiche Therapie von Wimpernausfall und diffuser

9
Einleitung
Verdünnung von Kopfhaaren wurde in klinischen Einzelstudien
beschrieben.[35],[37],[78] Die Potenz von PGF-2α Analoga, dünnes, kurzes,
unpigmentiertes Vellushaar in dickes, langes pigmentiertes terminales Haar zu
differenzieren, wurde beispielsweise in der Wangenregion[20],[21],[27] beschrieben
(Abbildung 2). Wachstum oder Ausdifferenzierung von Haaren, auch in
Regionen androgener Alopezie,[72] lässt darauf schließen, dass möglicherweise
Therapie von Alopecia areata nicht nur auf Wimpern beschränkt, sondern auch
in anderen Körperregionen möglich ist.
Abbildung 2: Pigmentierung des Vellushaares in
der malaren Region nach lokaler Applikation von
Bimatoprost (siehe Pfeil). Modifiziert aus Quelle:
Hart J. and Shafranov G. (2004). "Hypertrichosis
of vellus hairs of the malar region after unilateral
treatment with bimatoprost." Am J Ophthalmol
137: 756-7.[20]
So wurde beschrieben, dass PGF-2α in dorsalen Fellhaarfollikeln der Maus
Telogen zu Anagen-Konversion induzieren kann.[59] Follikel, lokalisiert an
unterschiedlichen Stellen des Körpers, werden jedoch durch unterschiedliche
molekulare Kontrollmechanismen gesteuert.[12],[66] Dies hat zur Folge, dass die
gleiche Substanz auf Haare unterschiedlicher Regionen unterschiedliche
Wirkungen haben kann. Beispielsweise induzieren Androgene zwar in der Bart-
und Achselregion während der Pubertät die Differenzierung von Vellushaar zu
terminalem Haar und dennoch ist die gleiche Substanz Ursache androgener
Alopezie.[18],[52] Auch für PGF-2α Behandlung wurden unterschiedliche
Reaktionen beschrieben. Während Alopecia areata in der Region der
Augenbrauen nicht erfolgreich mit Latanoprost therapiert werden konnte,[55]
wurde ein Wiederwachsen der Wimpern nach Alopezie infolge von
Latanoprosttherapie beschrieben.[35],[37]
2.3 Ein Tiermodell zur Untersuchung des Wimpernwachstums
Den klinischen Studien, die den wachstumsstimulierenden Effekt von PGF-2α
Analoga auf Wimpern oder sogar auf Haare anderer Regionen beschreiben,

10
Einleitung
steht nur eine geringe Zahl an tierexperimentellen Studien gegenüber. Während
sowohl die Morphogenese als auch der Wachstumszyklus dorsaler Hauthaare
von C75BL/6 Mäusen bereits intensiv erforscht wurde,[7],[49],[41],[43],[68] ist die
genaue Biologie des Wimpernfollikels, der Wimpernwachstumszyklus mit
seinen Kontrollmechanismen, sowie die Morphogenese der Wimpern
weitgehend unklar. Erst durch ein systematisches Beschreiben der Morphologie
des Wimpernzyklus können pathologische Fehlregulationen oder die Wirkung
von Pharmaka erkannt werden.
Ziel dieser Studie war es, die Wimpern von C57BL/6 Mäusen, dem bisher in der
Haarforschung am besten charakterisierten Tiermodell[7],[49],[41] zu beschreiben
und mit menschlichen Wimpern zu vergleichen. Dabei wurden die Morphologie
des Augenlids und der Wimpern, die Morphogenese, die Wachstumsstadien
und der Zeitpunkt des Exogen beschrieben und mit den am dorsalen Hauthaar
der Tiere erhobenen Befunden verglichen.
2.4 Einfluss von Bimatoprost auf das Wimpernwachstum bei der
Maus
Nachdem gezeigt werden konnte, dass die Morphologie und das Wachstum der
Mäusewimpern mit den menschlichen Wimpern gut vergleichbar ist, wurde der
Einfluss von Bimatoprost auf die Morphologie der Wimpern und ihrer Follikel
hinsichtlich der Anzahl, der Dicke, der Pigmentierung qualitativ und quantitativ
untersucht und die Modulation des Wimpernzyklus durch Bimatoprost
beschrieben.

11
Materialien und Methodik
3. Material und Methodik
3.1 Tiere
Es wurden weibliche C57BL/6J Mäuse von Harlan-Winkelman (Borchen,
Deutschland) verwendet, wobei Mäuse im Alter von drei bis sechs Wochen
direkt erworben wurden. Zur exakten Altersbestimmung jüngerer Tiere wurden
trächtige Mäuse gekauft und deren Würfe benutzt. Die Tiere lebten in Gruppen
von vier bis sechs bzw. die trächtigen Mäuse einzeln in einem Käfig, in dem ad
libidum Nahrung sowie Wasser zur Verfügung stand. Sowohl die Temperatur,
die Luftfeuchtigkeit, als auch der 12 Stunden Tag-Nacht Rhythmus wurden im
Laboratorium geregelt und überwacht. Alle Tierversuche wurden von der
Bewilligungsbehörde der Regierung von Mittelfranken genehmigt.
3.2 Probengewinnung
Zur Beschreibung der Morphologie der Wimper, der Entwicklung des Augenlids
und der Morphogenese der Wimpernfollikel im Vergleich zur Morphogenese des
dorsalen Fellhaares wurden unbehandelte Mäuse im Alter von 0,1,2,3,4,5, und
6 Wochen in sieben Gruppen von jeweils zwei bis vier Mäusen unterteilt. Mäuse
bis zu einem Alter von einer Woche wurden durch Dekapitation, die älteren
durch zervikale Dislokation getötet, um chirurgisch bilateral die oberen
Augenlider und dorsal, kaudal des Halses, mittig zwischen den beiden
Schulterblättern, eine Fläche von zwei cm2 Haut zu entnehmen (Fig. 4).
Die rechten oberen Augenlider wurden für vier Stunden in 4%
Paraformaldehyd-PBS fixiert und mit PBS gespült. Sie dienten der
makroskopischen Analyse. Dazu wurden die Wimpernschafte pro Augenlid in
jeder Altersgruppe (n=2-4 pro Gruppe) unter einem Stereomikroskop (Carl
Zeiss, Göttingen, Deutschland) dreimal gezählt und der Mittelwert gebildet.
Manche Augenlider wurden mithilfe einer Rasierklinge dünn geschnitten und auf
einen Objektträger aufgebracht, um die Wimpern unter einem Lichtmikroskop
(Carl Zeiss) betrachten oder Fotografien einziehen zu können (Software: Qwin,
Leica Microsystems GmbH, Wetzlar, Deutschland).
Die linken oberen Augenlider wurden eine Nacht in Ito-Lösung fixiert (2,5%
Glutaraldehyd, 2,5% Paraformaldehyd, 0,01% Pikrinsäure in 0,1 mM Cacodylat-

12
Materialien und Methodik
Puffer) und im Anschluss daran in Epon eingebettet. Von den Eponblöcken
wurden 1µm dicke Schnittserien (160 Schnitte je Wimper) hergestellt, jeder
zweite mit Toluidinblau (Sigma, Hannover, Deutschland) angefärbt, fotografiert
(Software: Qwin, Leica Microsystems GmbH) und die darin angeschnittenen
Papillen unter dem Lichtmikroskop (Carl Zeiss) einem Morphogenesestadium
bzw. einer Wachstumsphase zugeordnet. Die Wachstumsphase der
Wimpernfollikel wurde mithilfe einer Modifikation der Phaseneinteilung dorsaler
Fellhaare (siehe Kap. 4.3) lichtmikroskopisch bestimmt.
Um die Wimpernmorphogenese mit der Morphogenese der dorsalen Fellhaare
vergleichen zu können, wurde die dorsal entnommene Haut desselben Tieres
oder eines Tieres desselben Wurfes (1 Woche p.p.) analog zur Prozedur des
linken Augenlids präpariert und histologisch, lichtmikroskopisch die
Wachstumsphase der Haare in Serienschnitten bestimmt. Alle Chemikalien
wurden von Carl Roth (Karlsruhe, Deutschland) erworben, sofern nicht anders
vermerkt.
Fig. 4: Schemazeichnung der Probengewinnung.
Es wurden bilateral die oberen Augenlider (Pfeile schwarz) und
dorsal, kaudal des Halses, mittig zwischen den beiden
Schulterblättern, eine 2 cm² große Hautfläche zwischen den
Schulterblättern entnommen (Rechteck weiß).
3.3 Bimatoprostbehandlung
Um den Effekt von Bimatoprost hinsichtlich der Anzahl, der Länge, der Dicke
und der Pigmentierung der Wimpern zu evaluieren, wurden Wimpern von
Mäusen mit 0,03% Bimatoprost Augentropfen (Lumigan®; Allergan, Irvine, CA)
bzw. mit dem Vehikel (NaCl, Na2HPO4, Zitronensäure und Benzalkoniumchlorid
als Konservierungsmittel in gereinigtem Wasser; Allergan, Irvine CA) behandelt.
Vier Wochen alte Mäuse wurden in zwei Gruppen unterteilt, Vehikel (n=10) und

13
Materialien und Methodik
Bimatoprost (n=10). Täglich einmal morgens im Zeitraum von 14 Tagen wurden
mit der Pipette in das rechte Auge 3µl 0,03% Bimatoprost Augentropfen, bzw.
Vehikel appliziert. Die linken Augen blieben unbehandelt und dienten als
unbehandelte Kontrolle. Am 15. Tag, im Alter von sechs Wochen, wurden die
Tiere getötet und die oberen Augenlider entnommen. Als Basis-Kontrollgruppe
dienten unbehandelte Mäuse (n=10) derselben Altersgruppe (Fig. 5).
Fig. 5:
Therapieschema.
Schematische Dar-
stellung der Thera-
pie von Basis-
kontroll-, Vehikel-
und Bimatoprost-
gruppe.
3.3.1 Makroskopische Untersuchung
Die oberen Augenlider wurden eine Nacht in Ito-Lösung fixiert (2,5%
Glutaraldehyd, 2,5% Paraformaldehyd, 0,01% Pikrinsäure in 0,1 mM Cacodylat-
Puffer), mit PBS gespült und die Wimpern pro Augenlid unter dem
Stereomikroskop (Carl Zeiss, Göttingen, Deutschland) gezählt. Danach wurde
der temporale Anteil des Augenlids unter dem Stereomikroskop (Carl Zeiss)
exzidiert, mit einer Rasierklinge dünn getrimmt, auf einen Objektträger
aufgebracht und lichtmikroskopisch fotografiert (Mikroskop: Carl Zeiss;
Software: Qwin, Leica Microsystems GmbH, Wetzlar, Deutschland). Pro
Augenlid wurden drei Wimpern randomisiert aus jeder von drei Längengruppen
(kurz~250µm; mittel~450µm; lang~2500µm) nach technischen Gesichtspunkten
gewählt und der basal, direkt an das Epithel angrenzende Durchmesser dieser
Wimpernschafte, mit geeigneter Software gemessen (Qwin; Leica
Microsystems GmbH, Wetzlar, Deutschland).

14
Materialien und Methodik
3.3.2 Histologische Untersuchung
Der nasale Rest der Augenlider wurde in Epon eingebettet und je 160 mit
Toluidinblau (Sigma, Hannover, Deutschland) gefärbte Serienschnitte mit einer
Dicke von je 1µm angefertigt (160x1µm/Augenlid). Jeder zweite Schnitt einer
Serie wurde histologisch unter dem Lichtmikroskop fotografiert (Mikroskop: Carl
Zeiss; Software: Qwin, Leica Microsystems GmbH, Wetzlar, Deutschland). Der
Aufbau des Augenlids wurde beschrieben und Wimpernfollikel von den
Fellhaarfollikeln – von nun an als Haarfollikel bezeichnet – anhand ihrer
Lokalisation, der Wachstumsrichtung und ihrer Schnittachse differenziert. Alle
Wimpernfollikel mit sichtbarer dermaler Papille innerhalb des 160µm
Serienschnittes wurden auf den digital fotografierten Bildern nummeriert und
damit die totale Follikelanzahl pro 160µm evaluiert. Die Wachstumsphase jedes
nummerierten Follikels wurde mithilfe einer Modifikation der
Phasenbestimmung dorsaler Fellhaare lichtmikroskopisch bestimmt (siehe Kap.
4.3). Damit konnte der prozentuale Anteil jeder Wachstumsphase errechnet
werden. Der Durchmesser der dermalen Papille und des Bulbus wurde
lichtmikroskopisch bestimmt, indem die größte Messung der vertikalen
Verbindungslinie relativ zur Achse der Wimper als maximaler Durchmesser
definiert wurde (Fig. 6). Dazu wurde innerhalb des Serienschnittes der
maximale Durchmesser der dermalen Papille bzw. des Bulbus gesucht, dieser
Schnitt lichtmikroskopisch digital fotografiert und mit geeigneter Software die
Länge der Verbindungslinie gemessen (Mikroskop: Carl Zeiss; Software: Qwin,
Leica Microsystems GmbH, Wetzlar, Deutschland). Bei dieser Analyse wurden
nur Follikel berücksichtigt, deren Zentrum und damit der
dickste Anteil, innerhalb der 160µm lag.
Fig. 6: Follikel- und Papillendurchmesser.
Der maximale Durchmesser der Papille und des Follikels
wurden lichtmikroskopisch vermessen (siehe rote
Doppelpfeile).

15
Materialien und Methodik
In den Serienschnitten der Anagen-late Follikel wurde der Schnitt mit der
maximalen Anzahl der Melaningranula in der keratinogenen Zone gewählt. Die
Melaningranula wurden auf einer Fläche von circa drei Zellen (~60µm) unter
dem Lichtmikroskop gezählt und diese Fläche nach dem Einziehen des
Schnittes mit geeigneter Software vermessen (Mikroskop: Carl Zeiss; Software:
Qwin, Leica Microsystems GmbH, Wetzlar, Deutschland), so dass die Anzahl
der Melaningranula pro Einheit berechnet werden konnte.
3.4 Statistische Analyse
Die Anzahl der Wimpernschafte der Bimatoprost- gegenüber der Vehikelgruppe
bzw. der Bimatoprost- gegenüber der Basiskontrollgruppe, sowie Vehikel-
gegenüber der Basiskontrollgruppe wurde mit dem zweiseitigen Student„s t-Test
geprüft. Für den Vergleich von behandelten und kontralateral unbehandelten
Wimpern des gleichen Tieres wurde der zweiseitig verbundene t-Test
angewandt. Analog dazu wurde der Durchmesser der Wimpernschafte der
Bimatoprost- zur Vehikelgruppe mit dem zweiseitigen Student‟s t-Test, sowie
behandelte Wimpern versus kontralateral unbehandelte Wimpern des gleichen
Tieres mit dem zweiseitigen verbundenen t-Test auf Signifikanz überprüft. Die
Durchschnittslänge in jeder Behandlungsgruppe wurde zuvor mit einfaktorieller
Varianzanalyse (ANOVA) getestet, um zu bestätigen, dass die Dicke nicht
durch die Varianz in der Länge beeinträchtigt wird. Die Anzahl der Follikel von
Bimatoprost- versus Vehikelgruppe, Bimatoprost- versus Basiskontrollgruppe
und Vehikel- versus Basiskontrollgruppe wurde anhand des zweiseitigen
Student‟s t-Test verglichen. Der prozentuale Anteil jeder Phase wurde durch
den zweiseitigen Z-Test überprüft (Bimatoprost versus Vehikel, Bimatoprost
versus Basiskontrollgruppe und Vehikel versus Basiskontrollgruppe). Der
Durchmesser der Follikel und Papillen sowie die Anzahl der Melaningranula
wurden in Bimatoprost- gegen Vehikelgruppe durch den zweiseitigen Student‟s
t-Test auf Signifikanz überprüft. Für alle Analysen galten p-Verhältnisse kleiner
als 0,05 als statistisch signifikant.

16
Ergebnisse
4. Ergebnisse
4.1 Morphologie des Augenlids
Das Augenlid der adulten Maus zeigte die gleichen anatomischen Strukturen,
wie sie für das menschliche Augenlid beschrieben wurden.[53] Zentral wurde es
durch den Tarsus, das Bindegewebsskelett, gestützt, welchem außen die Pars
palpebralis des M. orbicularis oculi anlag, die von subkutanem Fettgewebe und
Epithel bedeckt wurde. Auf der den Augapfel bedeckenden Innenseite des
Tarsus befanden sich die holokrinen Gll. tarsales, die Meibom-Drüsen, und das
Epithel der Konjunktiva. Das Epithel des Augenlids konnte in drei
unterschiedliche Abschnitte unterteilt werden (Fig. 7), die Konjunktiva, das
Epithel, aus welchem die Wimpern austraten, von nun an mit dem Terminus
Wimpernepithel bezeichnet, und das Hautepithel. Das Epithel der Konjunktiva,
welches dicker als das Hautepithel war, bestand aus einem nicht verhornten
Plattenepithel mit reichhaltig Pigmentgranula im Stratum basale und spinosum
(Fig. 7a). Das Epithel der Haut, aus welchem die Hauthaare austraten, war
dünner und bestand aus verhorntem Plattenepithel. Es konnten keine
Pigmentgranula im Epithel beobachtet werden, nur vereinzelt im dermalen
Bindegewebe (Fig. 7c). Zwischen der Haut und der Konjunktiva, begrenzt durch
den Limb. palpebralis ant. und den Limb. palpebralis post., wobei der Limb.
palb. ant. mikroskopisch durch den Übergang in das Hautepithel dargestellt
werden konnte und der Limb. palb. post. durch den Austritt des
Ausführungsganges der Gll. tarsales, befand sich das Epithel, aus dem die
Wimpern austraten. Dieses Epithel besaß ein dickes Stratum corneum, ein gut
entwickeltes Stratum granulosum mit großen Keratohyalingranula und ein
dickes Stratum spinosum und Stratum basale. Es konnten viele Pigmentgranula
im Stratum basale, spinosum und granulosum beobachtet werden (Fig. 7b).
Wimpern konnten aufgrund ihrer Lokalisation zwischen dem M. orbicularis oculi
und den Gll. tarsales und ihrer Wachstumsrichtung von Fellhaaren
unterschieden werden. Sie wuchsen entgegengesetzt zu den Fellhaaren, so
dass die Wimpern in diesen Schnitten longitudinal und die Hauthaare
transversal geschnitten wurden.

17
Ergebnisse
Fig. 7: Morphologie des Augenlids der Maus.
1µm Semidünnschnitt durch das Augenlid einer sechs
Wochen alten Maus (Toluidinblau) Das Wimpernepithel
befand sich zwischen dem Limbus palbebralis anterior
(Limb. palb. ant) und dem Limbus palbebralis post.
(Limb. palb. post.). Die Wimpernfollikel (W) mit den Gll.
sebaceae (Gl. s) befanden sich zwischen dem M.
orbicularis oculi (MO) und den Gll. tarsales (Gll. t). Sie
waren longitudinal angeschnitten und traten aus dem
Wimpernepithel aus. Dadurch konnten sie von den
Fellharen (FH) unterschieden werden, welche
transversal angeschnitten waren und aus dem
Hautepithel austraten. [Skala=100µm]
(a-c) Vergrößerung der markierten Regionen aus dem
Semidünnschnitt des Augenlids. (a) Das Epithel der
Konjunktiva enthielt Pigmentgranula im Stratum basale
und spinosum (weiße Pfeilspitzen) und im dermalen
Bindegewebe (weiße Pfeile). (b) Das Wimpernepithel
war dick und reich an Pigmentgranula im Stratum
basale, spinosum und granulosum (weiße Pfeilspitzen)
69
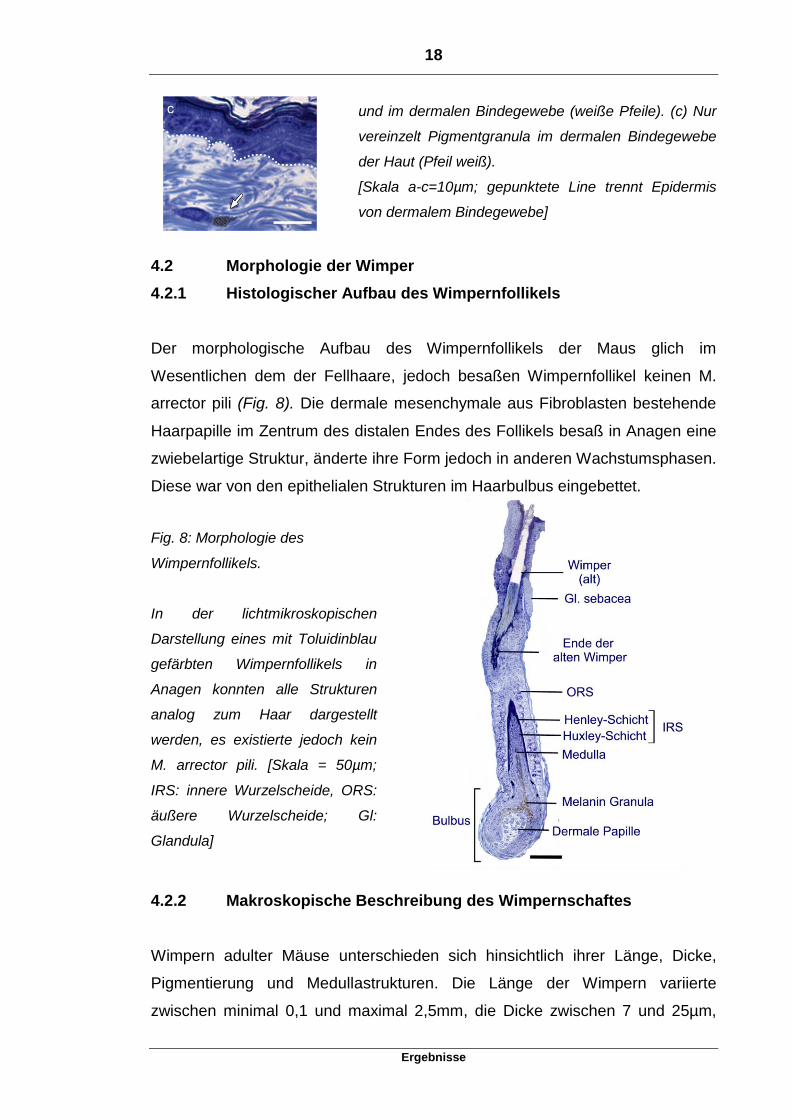
18
Ergebnisse
und im dermalen Bindegewebe (weiße Pfeile). (c) Nur
vereinzelt Pigmentgranula im dermalen Bindegewebe
der Haut (Pfeil weiß).
[Skala a-c=10µm; gepunktete Line trennt Epidermis
von dermalem Bindegewebe]
4.2 Morphologie der Wimper
4.2.1 Histologischer Aufbau des Wimpernfollikels
Der morphologische Aufbau des Wimpernfollikels der Maus glich im
Wesentlichen dem der Fellhaare, jedoch besaßen Wimpernfollikel keinen M.
arrector pili (Fig. 8). Die dermale mesenchymale aus Fibroblasten bestehende
Haarpapille im Zentrum des distalen Endes des Follikels besaß in Anagen eine
zwiebelartige Struktur, änderte ihre Form jedoch in anderen Wachstumsphasen.
Diese war von den epithelialen Strukturen im Haarbulbus eingebettet.
Fig. 8: Morphologie des
Wimpernfollikels.
In der lichtmikroskopischen
Darstellung eines mit Toluidinblau
gefärbten Wimpernfollikels in
Anagen konnten alle Strukturen
analog zum Haar dargestellt
werden, es existierte jedoch kein
M. arrector pili. [Skala = 50µm;
IRS: innere Wurzelscheide, ORS:
äußere Wurzelscheide; Gl:
Glandula]
4.2.2 Makroskopische Beschreibung des Wimpernschaftes
Wimpern adulter Mäuse unterschieden sich hinsichtlich ihrer Länge, Dicke,
Pigmentierung und Medullastrukturen. Die Länge der Wimpern variierte
zwischen minimal 0,1 und maximal 2,5mm, die Dicke zwischen 7 und 25µm,

19
Ergebnisse
wobei die Länge mit der Dicke korrelierte. Alle Wimpernschafte wiesen eine
leichte Biegung auf (Fig. 9a). Sie unterschieden sich nicht nur hinsichtlich ihrer
Länge und Dicke, sondern auch hinsichtlich ihrer Pigmentierung und dem
Aufbau der Medulla. Die langen, dicken Wimpern zeigten die stärkste
Pigmentierung, einen dicken Kortex und es wurden Lufteinschlüsse in der
Medulla beobachtet. Die Medulla war in ein- bis zweireihige Septen gegliedert
und könnte damit eine Ein- bis Zweisäulen Struktur formen. Diese war jedoch
auf die dicken Anteile der Wimpern begrenzt (Fig. 9b,c). Einige Wimpern
besaßen einen fibrösen gut pigmentieren Kortex, jedoch keine Medulla (Fig.
9d). Es existierten ebenso Velluswimpern ohne Pigmentierung (Fig. 9e).
Fig. 9: Länge und Matrixstruktur
der Mäusewimpern. (a) Wimpern
von Mäusen unterschiedlicher
Länge und Dicke. Jeder
Wimpernschaft wies eine Biegung
auf. (b-d) Vergrößerung der in
Figur 9a markierten Regionen. Die
Medulla des dicksten Anteils einer
langen Wimper wies eine
Zweisäulen-Struktur infolge von
Lufteinschlüssen auf (b), während
im dünneren Anteil eine
Einsäulen-Struktur beobachtet
werden konnte (c). Diese Bilder
wurden aufgrund der dichten
Pigmentierung überbelichtet, um die Strukturen zu zeigen. Wimpernschaft mit
Pigmentierung des fibrösen Kortex, doch ohne Medullastruktur (d). Vergrößerte
Ansicht einer kurzen und dünnen Wimper (~100µm) ohne Pigmentierung und
Medullastruktur (e). [Skala b-e: 10µm]
4.3 Unterteilung des Wimpernwachstumszyklus
Mithilfe der in den seriell sagittalen Semidünnschnitten lichtmikroskopisch
sichtbaren morphologischen Merkmale (Form, Struktur, Größe der dermalen
Papille und des Bulbus, Zellphasen, Melaningranula, Form bzw. Vorhandensein
69

20
Ergebnisse
der inneren Wurzelscheide, der äußeren Wurzelscheide und des
Wimpernschaftes) wurde der Wachstumszyklus der Wimper in sechs Stadien
unterteilt. Verglichen mit dorsalen Fellhaaren erforderte die Einteilung teilweise
andere Kriterien, da beispielsweise im Augenlid eine Unterteilung in Epidermis,
Dermis und Subkutis nicht möglich war. Deshalb wurden die Kriterien für die
Phasen des dorsalen Fellhaares der Maus[41] in dieser Studie für die Wimpern
modifiziert und mit einer Gliederung in die sechs Phasen Telogen, Anagen-
early, Anagen-middle, Anagen-late, Katagen-early und Katagen-late
vereinfacht.
4.3.1 Telogen
Wimpernfollikel in Telogen (Fig. 10a), der Ruhephase der Wimpern, drangen
meist nicht tiefer als bis zum oberen Drittel des Lids ein. Morphologisch war ein
Wimpernschaft – die Kolbenwimper – zu sehen. Charakteristisch für dieses
Stadium war die relativ kleine, rundlich geformte dermale Papille mit dicht
kondensierten Fibroblasten, lokalisiert im distalen Ende des Follikels. Weder
Zeichen eines neu wachsenden Haares, noch die innere Wurzelscheide noch
Melaningranula konnten in dieser Wachstumsphase detektiert werden.
Telogen
IRS/WS: WS der Kolben-wimper sichtbar.
DP: Am distalen Ende des Follikels war eine runde, kleine kondensierte DP zu sehen.
MG: --- --- ---
Zellphase: --- --- ---
Äquivalent: Telogen
Durchmesser: DP: 14,6±0,6 (µm) Bulbus: 18,2±0,1 (µm) DP/Bulbus: 83,8±2,4 (%) (Mittelwert±SEM; n=35)
Fig. 10a. Wimpernfollikel in Telogen (Ausschnitt eines 1µm dicken mit Toluidinblau
gefärbten Schnittes durch das Augenlid der adulten Maus, Skala=50µm). IRS/ORS:
innere/äußere Wurzelscheide, WS: Wimpernschaft, DP: dermale Papille, MG:
Melaningranula, Gl.: Glandula
69
21
Ergebnisse
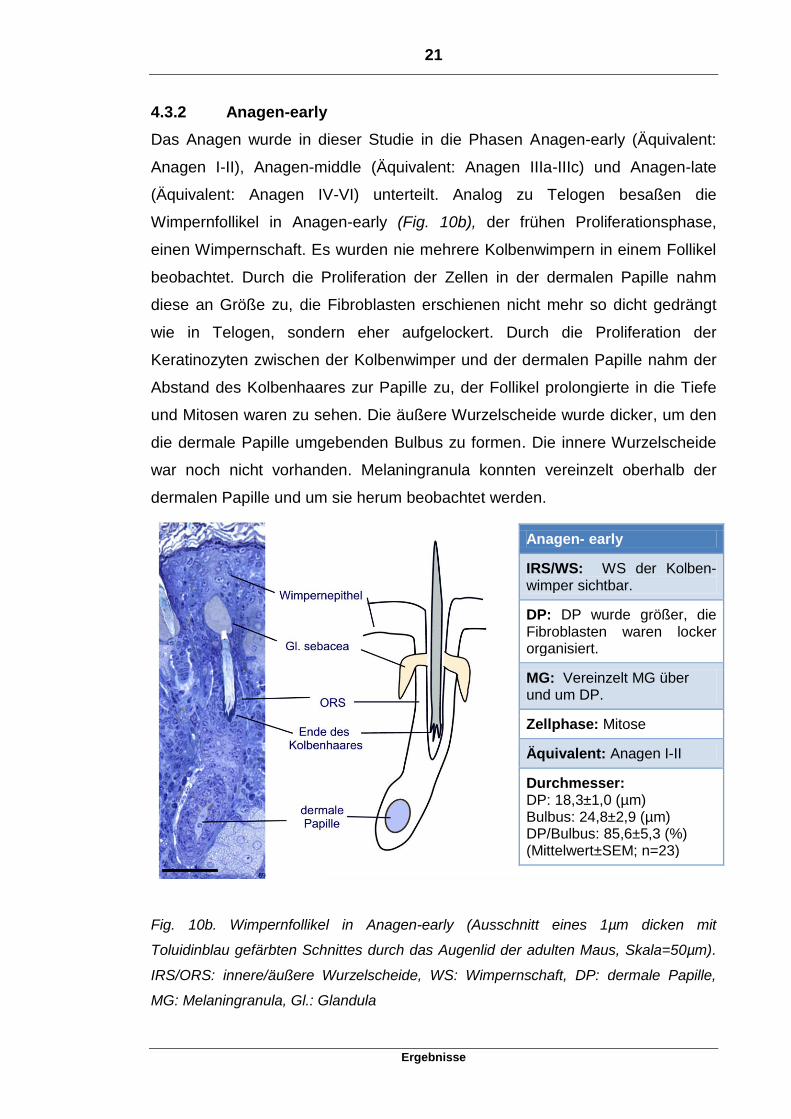
4.3.2 Anagen-early
Das Anagen wurde in dieser Studie in die Phasen Anagen-early (Äquivalent:
Anagen I-II), Anagen-middle (Äquivalent: Anagen IIIa-IIIc) und Anagen-late
(Äquivalent: Anagen IV-VI) unterteilt. Analog zu Telogen besaßen die
Wimpernfollikel in Anagen-early (Fig. 10b), der frühen Proliferationsphase,
einen Wimpernschaft. Es wurden nie mehrere Kolbenwimpern in einem Follikel
beobachtet. Durch die Proliferation der Zellen in der dermalen Papille nahm
diese an Größe zu, die Fibroblasten erschienen nicht mehr so dicht gedrängt
wie in Telogen, sondern eher aufgelockert. Durch die Proliferation der
Keratinozyten zwischen der Kolbenwimper und der dermalen Papille nahm der
Abstand des Kolbenhaares zur Papille zu, der Follikel prolongierte in die Tiefe
und Mitosen waren zu sehen. Die äußere Wurzelscheide wurde dicker, um den
die dermale Papille umgebenden Bulbus zu formen. Die innere Wurzelscheide
war noch nicht vorhanden. Melaningranula konnten vereinzelt oberhalb der
dermalen Papille und um sie herum beobachtet werden.
Anagen- early
IRS/WS: WS der Kolben-wimper sichtbar.
DP: DP wurde größer, die Fibroblasten waren locker organisiert.
MG: Vereinzelt MG über und um DP.
Zellphase: Mitose
Äquivalent: Anagen I-II
Durchmesser: DP: 18,3±1,0 (µm) Bulbus: 24,8±2,9 (µm) DP/Bulbus: 85,6±5,3 (%) (Mittelwert±SEM; n=23)
Fig. 10b. Wimpernfollikel in Anagen-early (Ausschnitt eines 1µm dicken mit
Toluidinblau gefärbten Schnittes durch das Augenlid der adulten Maus, Skala=50µm).
IRS/ORS: innere/äußere Wurzelscheide, WS: Wimpernschaft, DP: dermale Papille,
MG: Melaningranula, Gl.: Glandula
69

22
Ergebnisse
4.3.3 Anagen-middle
Im Follikel von Anagen-middle (Fig. 10c) entstand die innere Wurzelscheide,
welche den neu entstehenden Wimpernschaft umhüllte. Es bestand keine
Verbindung der Spitze der inneren Wurzelscheide mit dem Wimpernkanal. Es
waren deutlich mehr Melaningranula, v. a. in der keratinogenen Zone und der
Matrixzone sichtbar, welche von den Trichozyten aufgenommen und in
Richtung des Haarschafts transportiert wurden. Sowohl die dermale Papille, als
auch der Bulbus erreichten ihre fast maximale Größe, weshalb die Fibroblasten
in der dermalen Papille locker organisiert erschienen. Aufgrund des Wachstums
konnten, wie in allen Phasen des Anagens, Mitosen beobachtet werden.
Fig. 10c. Wimpernfollikel in Anagen-middle.
(Ausschnitt eines 1µm dicken mit Toluidinblau gefärbten Schnittes durch das Augenlid
der adulten Maus, Skala=50µm) IRS/ORS: innere/äußere Wurzelscheide, WS:
Wimpernschaft, DP: dermale Papille, MG: Melaningranula, Gl.: Glandula
4.3.4 Anagen-late
In Anagen-late (Fig. 10d) erreichte die innere Wurzelscheide Anschluss an den
Haarkanal des Kolbenhaares, wodurch der in Anagen entstandene Schaft aus
Anagen-middle
IRS/WS: Trichozyten produ-zierten Trichohyalingranula, um den IRS zu formen (gestrichelte Pfeile). Am Ende der Phase entstand ein neuer WS, jedoch erreichte IRS des WS noch nicht den Wimpernkanal.
DP: DP erreichte fast maximale Größe, die Fibroblasten waren locker organisiert.
MG: Zahlreiche MG von der Matrixzone bis zur Medulla.
Zellphase: Mitosen v.a. im Bulbus (weiße Pfeile).
Äquivalent: Anagen IIIa-IIIc
Durchmesser: DP: 27,2±5,2 (µm) Bulbus: 69,2±11,6 (µm) DP/Bulbus: 39,0±1,2 (%) (Mittelwert±SEM; n=3)
69

23
Ergebnisse
dem Epithel austrat. Der Schaft des Kolbenhaares verblieb im Haarkanal und
war noch nicht epiliert, so dass in Anagen-late immer zwei aus dem Epithel
austretende Schafte beobachtet wurden. Es wurden Melaningranula nicht nur in
der keratinogenen Zone, sondern auch um die dermale Papille aufsteigend in
die Matrixzone und im Wimpernschaft beobachtet. Der Durchmesser der
dermalen Papille und des Bulbus hatte ungefähr dieselbe Größe wie in Anagen-
early. Die Zellen in der dermalen Papille erschienen weiterhin locker organisiert.
Aufgrund der Proliferation konnten Mitosen beobachtet werden.
Anagen-late
IRS/WS: Spitze der IRS des neu geformten WS erreichte Wimpernkanal und trat aus dem Epithel aus. Die Kolbenwimper verblieb im Follikel.
DP: Fibroblasten der DP waren locker organisiert.
MG: MG über und um DP sowie in der Matrixzone (Pfeile weiß).
Zellphase: Mitose
Äquivalent: Anagen IV-VI
Durchmesser: DP: 27,9±2,5 (µm) Bulbus: 69,4±4,7 (µm) DP/Bulbus: 41,6±3,8 (%) (Mittelwert±SEM; n=15)
Fig. 10d. Wimpernfollikel in Anagen-late und selber Follikel 12µm vom Schnitt entfernt.
(Ausschnitte 1µm dicker mit Toluidinblau gefärbter Schnitte durch das Augenlid der
adulten Maus, Skala=50µm) IRS/ORS: innere/äußere Wurzelscheide, WS:
Wimpernschaft, DP: dermale Papille, MG: Melaningranula, Gl.s.: Glandula sebacea
4.3.5 Katagen-early
Die Regressionsphase wurde in dieser Studie in Katagen-early (Äquivalent
Katagen I-VI) und Katagen-late (Äquivalent: Katagen VII-VIII) unterteilt. Im
Katagen-early (Fig. 10e) fing der Abbau mit der Degeneration der inneren
Wurzelscheide an, welcher am distalen Ende beginnend langsam zum
proximalen Ende hin fortschritt, weshalb hier Apoptosen beobachtet werden
konnten. Der Schaft des Kolbenhaares verblieb weiterhin im Haarkanal, so dass
69
69

24
Ergebnisse
in dieser Phase analog zum Anagen-late zwei aus dem Epithel austretende
Schafte pro Follikel beobachtet wurden. Die Medulla des neuen
Wimpernschaftes wies in der Nähe der dermalen Papille eine bürstenförmige
Struktur auf. Melaningranula waren zwar vorhanden, aber die Anzahl nahm
stark ab, so dass sie nur noch oberhalb der dermalen Papille in der
keratinogenen Zone lokalisiert waren, jedoch nicht mehr um die dermale Papille
herum. Der Durchmesser der rundlich, ovalen dermalen Papille mit kugelig
dicht gedrängten Zellen war deutlich reduziert. Auch der Durchmesser des
Bulbus nahm ab. Die Degeneration der äußeren Wurzelscheide war deutlich
sowohl an der Abnahme ihres Durchmessers als auch an dem Auftreten von
Apoptosen zu erkennen. Insgesamt nahm auch die Länge des Follikels ebenso
wie der Durchmesser ab, so dass der Follikel kleiner erschien.
Fig. 10e: Wimpernfollikel in Katagen-early und selber Follikel 24µm vom Schnitt
entfernt. (Ausschnitte 1µm dicker mit Toluidinblau gefärbter Schnitte durch das
Augenlid der adulten Maus, Skala=50µm) IRS/ORS: innere/äußere Wurzelscheide,
WS: Wimpernschaft, DP: dermale Papille, MG: Melaningranula, Gl.s.: Glandula
sebacea
Katagen-early
IRS/WS: Degeneration der IRS begann distal. Die Medulla des WS wies ein fibröses, bürstenförmiges Ende in der Nähe der DP auf. Die Kolbenwimper verblieb im Follikel.
DP: DP war klein mit dicht orga-nisierten Zellen.
MG: Deutlich weniger MG, nur noch über keratinogener Zone sichtbar.
Zellphase: Apoptosen im ORS und am proximalen Ende des IRS.(schwarze Pfeilspitzen)
Äquivalent: Katagen I-VI
Durchmesser DP: 21,1±1,9 (µm) Bulbus: 37,6±5,1 (µm) DP/Bulbus: 59,1±8,3 (%) (Mittelwert±SEM; n=4)
69
69

25
Ergebnisse
4.3.6 Katagen-late
In Katagen-late (Fig. 10f) war die innere Wurzelscheide fast vollständig
degeneriert, nur noch fokale Reste im mittleren Drittel des Wimpernfollikels
waren vorhanden. Infolge des Effluviums der Kolbenwimper (Exogen) war nur
noch der in Anagen neu produzierte Wimpernschaft im Haarkanal zu
beobachten. Man konnte viele Apoptosen finden mit einer lokalen Betonung
über der dermalen Papille und um den Schaft, der Stelle der degenerierten
inneren Wurzelscheide. Die dermale Papille wurde kleiner, nahm fast die Größe
analog zu Telogen an, die Zellen erschienen stark kondensiert. Die dermale
Papille befand sich näher am Ende des neuen Wimpernschaftes.
Fig. 10f: Wimpernfollikel in Katagen-late. (Ausschnitt eines 1µm dicken mit Toluidinblau
gefärbten Schnittes durch das Augenlid der adulten Maus, Skala=50µm) IRS/ORS:
innere/äußere Wurzelscheide, WS: Wimpernschaft, DP: dermale Papille, MG:
Melaningranula, Gl.s.: Glandula sebacea
Während des Wimpernzyklus war damit der Durchmesser der dermalen Papille
und des Bulbus in Telogen am kleinsten. In Anagen nahmen die Durchmesser
zu und erreichten in Anagen-middle und Anagen-late ihre maximale Größe.
Katagen-late
IRS/WS: Zurückbildung des IRS mit sichtbarem Rest. Das alte Kolbenhaar wurde aus-gestoßen (Exogen) und war nicht mehr sichtbar.
DP: DP war klein mit konden-sierten Zellen und befand sich nahe am Ende des WS.
MG: --- --- ---
Zellphase: Apoptosen im ORS (schwarze Pfeilspitzen).
Äquivalent.: Katagen VII-VIII
Durchmesser: DP: 14,9±1,0 (µm) Bulbus: 21,1±1,3 (µm) DP/Bulbus: 73±4,2 (%) (Mittelwert±SEM; n=26)
69

26
Ergebnisse
Dabei war der Anstieg des Durchmessers des Bulbus verhältnismäßig größer,
so dass das Verhältnis von dermaler Papille zu Bulbus in Anagen-middle und
Anagen-late kleiner war. (DP/Bulbus: Telogen 82,8%, Anagen-early 85,6%,
Anagen-middle 39,0%, Anagen-late 41,6%, Katagen-early 59,1%, Katagen-late
73,3%). In Katagen nahm der Durchmesser der dermalen Papille und des
Bulbus wieder ab (Fig. 11).
Fig. 11: Durchmesser des Bulbus und der dermalen Papille.
Sowohl der Durchmesser der dermalen Papille, als auch der Durchmesser des Bulbus
stieg in Anagen-middle und Anagen-late an und sank während Katagen, wobei durch
einen verhältnismäßig größeren Anstieg des Durchmessers des Bulbus das Verhältnis
von dermaler Papille zu Bulbus in Anagen-middle und Anagen-late kleiner war.
Im gesamten Wimpernzyklus wurden keine Wimpernfollikel mit zwei
Kolbenhaaren beobachtet. Follikel in Katagen-early besaßen zwei
Wimpernschafte, den Schaft des Kolbenhaares aus Telogen (Wimper alt) und
den neu in Anagen produzierten Schaft (Wimper neu) und Follikel in Katagen-
late nur einen Schaft (Wimper alt). Diese Beobachtung ließ darauf schließen,
dass Exogen während Katagen in jedem Zyklus stattfinden muss (Fig. 12).

27
Ergebnisse
Fig. 12: Anzahl der aus dem Epithel
austretenden Wimpernschafte pro
Follikel. In Katagen-early waren Follikel
mit zwei austretenden Wimpernschaften
vorhanden, in Katagen-late besaßen die
Follikel nur einen Wimpernschaft.
4.4 Morphogenese und erster Zyklus der Wimper
4.4.1 Makroskopie
Bei der Geburt waren die Augenlider der Mäuse verschlossen und es konnten
mithilfe des Stereomikroskops keine Wimpernschafte an der Oberfläche
beobachtet werden. Ab der ersten Lebenswoche wurden vereinzelt
Wimpernschafte sichtbar, wobei die Anzahl bis zu einem Alter von drei Wochen
post partum rasch anstieg. Danach blieb die Anzahl der Wimpern pro Augenlid
während des Beobachtungszeitraumes konstant bei einem Wert zwischen 130
und 150 (Fig. 13).
Fig. 13: Anzahl der
Wimpern während der
Morphogenese. Die
Anzahl der Wimpern
stieg bis zur dritten
Woche an und blieb
danach konstant.
4.4.2 Histologie
4.4.2.1 Morphogenese in Woche 0 post partum
Die generellen Merkmale der Wimpernmorphogenese entsprachen der
Morphogenese dorsaler Hauthaare.[49] Zwar waren bei Geburt Wimpernschafte
an der Oberfläche des Augenlids noch nicht sichtbar, doch die Anfangsstadien
der Morphogenese waren bereits unter der Haut des noch verschlossenen

28
Ergebnisse
Augenlids in den mikroskopischen Schnitten zu beobachten (Fig. 14a, links).
Epidermale Verdickungen, Wimpernknospen, des relativ dicken und
Keratohyalingranula haltigen Epithels waren infolge einer Kumulation von
Keratinozyten und Fibroblasten sichtbar (St. 1). In die Tiefe elongierte Keime
mit einer Ansammlung von Fibroblasten am distalen Ende wurden beobachtet
(St. 2) und ein beginnender Einschluss dieser Fibroblasten in den Haarkeim (St.
3) deuteten die Entwicklung der dermalen Papille an (Fig. 14a, Mitte).
Insgesamt konnte zu diesem Zeitpunkt kurz nach der Geburt nur eine begrenzte
Anzahl an Follikeln unterschiedlicher Morphogenesestadien beobachtet
werden.
Diese Stadien ähnelten den Morphogenesestadien der dorsalen Fellhaare (Fig.
14a, rechts).
Fig. 14a: Woche 0 post partum.
Skala schwarz=100µm, Skala weiß=20µm, M.o.: Muskulus orbicularis oculi
Fig 14a-g: 1µm dicke mit Toluidinblau angefärbte Schnitte durch das Augenlid (links)
bzw. durch die dorsale Haut (rechts) einer Maus des entsprechenden Alters mit
vergrößerter Ansicht von nummerierten Wimpernfollikeln (Mitte).
Links: Die weiße gepunktete Linie trennt das noch verschlossene Ober- vom
Unterlid. Es waren Follikel in den Morphogenesestadien 1-3 sichtbar
(schwarze Pfeile).
Mitte: Vergrößerte Ansicht von Follikel 1;2;3 im Serienschnitt 9;9;4µm entfernt.
Epidermale Verdickung und Ansammlung dermaler Fibroblasten im St. 1(1).
Kondensation der Fibroblasten im St. 2(2). Entstehung der dermalen Papille
im St. 3 (3).
Graph: Alle im linken Schnitt angeschnittenen Follikel befanden sich in den
Morphogenesestadien 1-3.
Rechts: Die dorsalen Hauthaare desselben Tieres waren in unterschiedlichen
Morphogenesestadien.
69

29
Ergebnisse
4.4.2.2 Morphogenese in Woche 1 post partum
Die Augenlider waren zwar auch in der ersten Woche post partum noch
verschlossen, doch nahm die Anzahl der Follikel (Fig. 14b, links) rasch zu und
es konnten bereits vereinzelt Wimpern an der Oberfläche beobachtet werden.
Die Follikel befanden sich in unterschiedlichen Wachstumsphasen der
Morphogenese (Fig. 14b, Mitte). Es waren Follikel in frühen
Morphogenesestadien zu finden, in welchen die äußere Wurzelscheide begann
die dermale Papille zu umschließen. Aber auch spätere Stadien waren sichtbar,
in welchen sich die innere Wurzelscheide und die Melanogenese entwickelten
und Sebozyten die Talgdrüse formten.
Die meisten Fellhaare befanden sich zu diesem Zeitpunkt im Stadium 8 der
Morphogenese (Fig. 14b, rechts).
Fig. 14b: Woche 1 post partum.
Skala schwarz=100µm, Skala weiß=20µm, M.o.: M. orbicularis oculi, Gll.t. Gll. tarsales
Links: Die weiße gepunktete Linie trennt die noch verschlossenen Lider. Follikel im
St. 3 (schwarzer Pfeil 1); St. 4 (weiße Pfeilspitze), wobei die dermale Papille
fast von der äußeren Wurzelscheide umschlossen war; St. 6 (weißer Pfeil) mit
Entwicklung des IRS und Melanogenese und St. 8 (schwarzer Pfeil 2).
Mitte: Vergrößerung von Follikel 1;2 im Serienschnitt 1;20 µm entfernt. Der
Wimpernkeim war im St. 3 (oben) noch nicht von der äußeren Wurzelscheide
umschlossen. Eine Wimper im St. 8 (unten) mit fast entwickelter Talgdrüse
erreichte fast den Haarkanal.
Graph: Die im linken Schnitt angeschnittenen Follikel befanden sich in
unterschiedlichen Morphogenesestadien. (nicht eindeutig zuordenbare
Stadien im Graph zwischen den Phasen dargestellt)
Rechts: Die dorsalen Hauthaare eines Tieres desselben Wurfes befanden sich am
Ende der Morphogenese.
69

30
Ergebnisse
4.4.2.3 Beginn des ersten Wimpernzyklus in Woche 2 post partum
In der zweiten Woche post partum hatten alle Wimpernfollikel die
Morphogenese abgeschlossen (Fig. 14c, links). Ein großer Anteil der Follikel
befand sich in Katagen und Telogen, vereinzelt wurden aber auch Follikel in
Anagen-early gefunden (Fig. 14c, Mitte). Damit hatte der Eintritt in den ersten
Wachstumszyklus bereits begonnen. Im Gegensatz dazu befanden sich alle
dorsalen Hauthaare in Katagen oder Telogen (Fig. 14 c, rechts).
Fig. 14c: Woche 2 post partum.
Skala schwarz=100µm, Skala weiß=20µm, M.o.: M. orbicularis oculi
Links: Follikel in Katagen-early (weiße Pfeilspitzen), Katagen-late (weiße Pfeile) und
Anagen-early (schwarzer Pfeil).
Mitte: Vergrößerte Ansicht vom Follikel (schwarzer Pfeil) im selben Schnitt (1) bzw.
39µm entfernt (2), aus welcher Ebene das Kolbenhaar und damit der Beginn
von Anagen deutlich wird.
Graph: Die meisten im linken Schnitt angeschnittenen Follikel befanden sich in
Katagen und Telogen, teilweise aber auch in Anagen.
Rechts: Die dorsalen Hauthaare desselben Tieres befanden sich in Katagen und
Telogen.
4.4.2.4 Woche 3 post partum
In der dritten Woche post partum befanden sich alle Wimpernfollikel in Anagen-
middle oder Anagen-late, somit in Anagen und damit synchron in einer Phase
des Wachstumszyklus (Fig. 14d links). Alle dorsalen Hauthaare konnten
Telogen zugeordnet werden (Fig. 14d rechts).
69

31
Ergebnisse
Fig. 14d: Woche 3 post partum.
Skala schwarz=100µm, M.o.: M. orbicularis oculi, Gll. t.: Gll. tarsales
Wimpernfollikel in Anagen (links und Graph), die dorsalen Hauthaare desselben Tieres
in Telogen (rechts). (nicht eindeutig zuordenbare Stadien im Graph zwischen den
Phasen dargestellt)
4.4.2.5 Woche 4 post partum
In der vierten Woche post partum befand sich der Großteil der Wimpern in
Telogen (Fig. 14e, links), nur ein kleiner Teil in Katagen oder Anagen-early
(nicht gezeigt). Die dorsalen Hauthaare hingegen wurden synchron in Anagen-
early vorgefunden (Fig. 14e, rechts).
Fig. 14e: Woche 4 post partum.
Skala schwarz=100µm, M.o.: M. orbicularis oculi, Gll. t.: Gll. tarsales
Wimpernfollikel in Telogen, vereinzelt in Katagen-late (links und Graph), die dorsalen
Hauthaare desselben Tieres in Anagen (rechts).
4.4.2.6 Woche 5 post partum
In der fünften Woche post partum begann der zweite Zyklus der Wimpern,
welche sich teilweise in Telogen, teilweise in Anagen-early (Fig. 14f, links)
befanden. Die dorsalen Hauthaare wurden noch in Anagen-late vorgefunden.
(Fig. 14f, rechts).
69
69

32
Ergebnisse
Fig. 14f: Woche 5 post partum.
Skala schwarz=100µm, M.o.: M. orbicularis oculi, Gll. t.: Gll. tarsales
Ein neuer Zyklus der Wimpernfollikel begann mit Telogen und Anagen (links und
Graph), die Hauthaare desselben Tieres befanden sich in Anagen-late (rechts).
4.4.2.7 Woche 6 post partum
In der sechsten Woche post partum ließen sich Wimpernfollikel in
verschiedenen Wachstumsphasen darstellen (Fig. 14g, links).
Die Synchronität des Wachstumszyklus der dorsalen Hauthaare blieb noch
erhalten. Sie befanden sich in Katagen (Fig. 14g, rechts).
Fig. 14g: Woche 6 post partum.
Skala schwarz=100µm, Skala weiß=20µm M.o.: M. orbicularis oculi
Links: Follikel in unterschiedlichen Phasen, z. B. in Telogen (Pfeil 1), Anagen-early
(Pfeil 2) und Anagen-late (Pfeile 3).
Mitte: Vergrößerte Ansicht von Follikel 1;2;3 im Serienschnitt 18;0;50µm entfernt.
Kompakte dermale Papille in Telogen (1). Aktive Proliferation und große
dermale Papille in Anagen-early (2). Aktive Proliferation und Melanogenese in
Anagen-late (3).
Graph: Die im linken Schnitt angeschnittenen Wimpernfollikel wurden in
verschiedenen Phasen beobachtet.
Rechts: Die dorsalen Hauthaare desselben Tieres befanden sich in Katagen-late.
69
69

33
Ergebnisse
Zusammenfassend endete die Morphogenese der Wimpern analog zum
Fellhaar zwischen der ersten und zweiten Woche. Sofort nach der
Morphogenese begann der erste Wimpern- bzw. Haarzyklus. Dieser erste
synchrone Wimpernzyklus schritt schneller voran, verglichen mit dem dorsalen
Hauthaar (Fig. 15).
Fig. 15: Chronologische Darstellung der Morphogenese und des ersten Zyklus der
Wimper im Vergleich zum Hauthaar.
Sowohl bei den Wimpern als auch bei den dorsalen Fellhaaren begann ein
synchronisierter Zyklus sofort nach der Morphogenese, welcher bei den Wimpern
schneller voranschritt.
4.5 Bimatoprostbehandlung
4.5.1 Makroskopische Untersuchung
Die Analyse der Wimpern unter dem Stereomikroskop ergab keine Anomalitäten
als Folge der Bimatoprostbehandlung im Vergleich zu den kontralateralen
unbehandelten Wimpern (Fig. 16).
Fig. 16: Makroskopischer Vergleich
der Wimpern.
Kein makroskopischer Unterschied
zwischen repräsentativen kontra-
lateralen unbehandelten (Bimato-
prost-kl) und mit Bimatoprost be-
handelten Wimpern (Bimatoprost).
[Skala=100µm]
Die Anzahl der Wimpernschafte pro Augenlid stieg unter
Bimatoprostbehandlung signifikant von ~142 bei den Kontrollen auf ~168 bei
Bimatoprost behandelten Tieren an (Fig. 17). Damit erhöhte sich die Anzahl
der Wimpern pro Augenlid unter Bimatoprostbehandlung statistisch signifikant,
69

34
Ergebnisse
verglichen mit der Basiskontrollgruppe, der Vehikelbehandlung und den
unbehandelten kontralateralen Wimpern. Zwischen den unbehandelten
Kontrollen (Basiskontrollgruppe und kontralateral unbehandelte Wimpern) und
Vehikel-behandelten Wimpern gab es keinen Unterschied.
Fig. 17: Anzahl der Wimpern.
Grafik: Die Anzahl der Wimpern nahm unter Bimatoprostbehandlung signifikant zu im
Vergleich zu allen anderen Gruppen. (*: signifikanter Unterschied (p<0,05), ns: nicht
signifikanter Unterschied)
Tabelle: Anzahl der Wimpern pro Augenlid als Mittelwert±SEM
[n=10 pro Gruppe, Basis: Basiskontrollgruppe; Vehikel/Bimatoprost: rechte behandelte
Augen der Vehikel/Bimatoprost Gruppe; Vehikel-KL/Bimatoprost-KL: linke
unbehandelte Augen der Vehikel/Bimatoprost Gruppe]
Die Wimpern wurden anhand ihrer Länge in drei Gruppen unterteilt
(lang~2570µm, mittel~451µm, kurz~250µm). Die detaillierte Länge,
aufgeschlüsselt nach Behandlungsgruppen, entnehme man Tab. 1. Die
Mittelwerte jeder Behandlungsgruppe wurden mit der einfaktoriellen
behandeltes Auge unbehandeltes Auge (KL)
Basis 144,15±5,34
Vehikel 145,65±5,42 140,35±6,62
Bimatoprost 168,30±3,89 138,75±5,43

35
Ergebnisse
Varianzanalyse ANOVA auf signifikante Unterschiede innerhalb der jeweiligen
Längengruppe überprüft, um eine mögliche Interferenz der Dicke durch zu
große Varianz in der Länge auszuschließen.
Vehikel Vehikel-KL Bimatoprost Bimatoprost-KL Mittelwert
lang 2616,72±42,66 2565,51±32,09 2605,32±45,42 2493,58±44,38 2570,28±20,95
mittel 441,95±10,67 440,54±16,22 451,52±10,32 468,37±11,91 450,60±6,25
kurz 246,46±6,49 244,99±6,91 261,69±5,72 247,19±6,51 250,08±3,23
Tab. 1: Länge der Wimpern, aufgeschlüsselt nach Behandlungsgruppen.
Mittelwert der Länge der Wimpern in µm±SEM [n=3 Wimpern pro Längengruppe bei 10
Augenlidern pro Behandlung] in den Gruppen lang, mittel und kurz aufgeschlüsselt
nach der jeweiligen Behandlung (Vehikel, Vehikel kontralateral (KL), Bimatoprost und
Bimatoprost kontralateral (KL).
Unter Bimatoprostbehandlung nahm der Durchmesser von mittellangen und
kurzen Wimpern signifikant zu (Fig. 18). Es resultierte in keiner Längengruppe
ein signifikanter Unterschied hinsichtlich der Dicke zwischen mit Vehikel
behandelten und kontralateral unbehandelten Augen. Im Gegensatz dazu
waren die mit Bimatoprost behandelten Wimpern signifikant dicker in den
Gruppen „mittel“ und „kurz“, verglichen mit den unbehandelten kontralateralen
Augen, nicht jedoch in Gruppe „lang“. Analog dazu konnte eine Signifikanz
hinsichtlich der Dicke bei den mit Bimatoprost behandelten Wimpern im
Vergleich zu Vehikel behandelten Wimpern in den Gruppen „mittel“ und „kurz“
nachgewiesen werden, nicht jedoch in Gruppe „lang“.

36
Ergebnisse
Vehikel Vehikel-KL Bimatoprost Bimatoprost-KL
lang 22,74±0,82 21,62±0,74 23,19±0,79 21,99±0,70
mittel 12,54±0,60 12,01±0,62 15,75±0,84 12,14±0,65
kurz 7,19±0,42 7,70±0,42 9,79±0,64 7,80±0,55
Fig. 18: Durchmesser der Wimpern.
Grafik: Der Durchmesser der Wimpern nahm unter Bimatoprostbehandlung
(Bimatoprost) signifikant bei mittellangen und kurzen Wimpern zu, nicht jedoch bei den
langen Wimpern, verglichen mit kontralateralen unbehandelten Wimpern (Bimatoprost-
KL/Vehikel-KL) und mit Vehikel behandelten Wimpern (Vehikel). (*: signifikanter
Unterschied (p<0,05), ns: nicht signifikanter Unterschied)
Tabelle: Durchmesser der Wimpern als Mittelwert in µm±SEM
[(n(Wimper)=3 pro Augenlid; n(Augenlid)=10 pro Längengruppe]
4.5.2 Histologische Untersuchung
Die Anzahl der Follikel stieg unter Bimatoprostbehandlung nicht an. Die
Follikelanzahl ergab innerhalb 160µm in der unbehandelten Basiskontrollgruppe
(4 Wochen p.p.) 14,60±0,64 [n=10], unter Vehikelbehandlung (6 Wochen p.p.
am Ende der Behandlung) 15,33±1,01 [n=9] und unter Bimatoprostbehandlung
(6 Wochen p.p. am Ende der Behandlung) 16,00±1,05 [n=9] [Mittelwert±SEM].
Die Anzahl war somit in allen Gruppen ähnlich, es konnte kein signifikanter
Unterschied nachgewiesen werden. Dies deutet darauf hin, dass der
wachstumsstimulierende Effekt von Bimatoprost trotz der höheren
Wimpernanzahl (siehe oben) nicht auf eine de novo Bildung von Wimpernfollikel
zurückgeführt werden konnte (Fig. 19).
Fig. 19: Anzahl der Follikel.
Kein Unterschied hin-
sichtlich der Follikelanzahl
in der Basiskontrollgruppe
(n=10), der Vehikel Gruppe
(n=9) und der Bimatoprost
Gruppe (n=9).

37
Ergebnisse
Der prozentuale Anteil von Anagen stieg und der von Telogen sank unter
Bimatoprostbehandlung. Alle gezählten Follikel innerhalb der 160µm eines
Augenlids wurden einer der sechs Phasen des Wachstumszyklus der Wimper
zugeordnet (Kriterien siehe oben). Aufgrund der mangelnden Anzahl der Follikel
in Anagen-middle und Anagen-late wurden diese beiden Phasen in der
Berechnung zusammengefasst. Zwischen der Basiskontrollgruppe und der mit
Vehikel behandelten Gruppe waren keine signifikanten Unterschiede in der
prozentualen Phasenverteilung feststellbar. Im Gegensatz dazu sank der
prozentuale Anteil von Telogen und Katagen-late unter Bimatoprostbehandlung
signifikant und der Anteil von Anagen-early und Anagen-middle/late stieg
signifikant im Vergleich zur vier Wochen alten Kontrollgruppe an. Katagen als
Gruppe (Katagen-early und Katagen-late) wies keinen signifikanten Unterschied
beim Vergleich von Bimatoprost- zur Kontrollgruppe auf. Unter
Bimatoprostbehandlung ließ sich ein signifikanter Unterschied von Telogen,
Anagen-middle/late und Anagen als Gesamtgruppe, verglichen mit der
Vehikelbehandlung, feststellen (Fig. 20). Diese Ergebnisse deuteten darauf hin,
dass Bimatoprost den prozentualen Anteil der Anagenfollikel, vor allem den in
der Anagen-middle/late Phase, erhöhte.
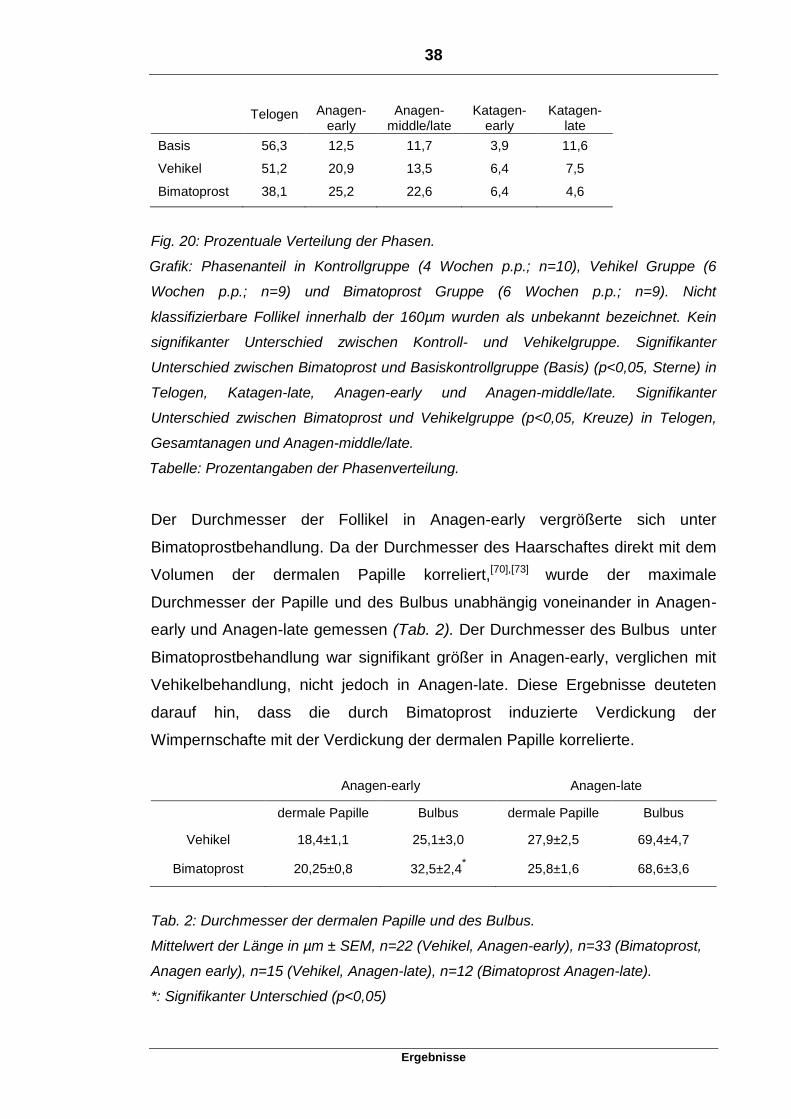
38
Ergebnisse
Telogen Anagen-early
Anagen-middle/late
Katagen-early
Katagen-late
Basis 56,3 12,5 11,7 3,9 11,6
Vehikel 51,2 20,9 13,5 6,4 7,5
Bimatoprost 38,1 25,2 22,6 6,4 4,6
Fig. 20: Prozentuale Verteilung der Phasen.
Grafik: Phasenanteil in Kontrollgruppe (4 Wochen p.p.; n=10), Vehikel Gruppe (6
Wochen p.p.; n=9) und Bimatoprost Gruppe (6 Wochen p.p.; n=9). Nicht
klassifizierbare Follikel innerhalb der 160µm wurden als unbekannt bezeichnet. Kein
signifikanter Unterschied zwischen Kontroll- und Vehikelgruppe. Signifikanter
Unterschied zwischen Bimatoprost und Basiskontrollgruppe (Basis) (p<0,05, Sterne) in
Telogen, Katagen-late, Anagen-early und Anagen-middle/late. Signifikanter
Unterschied zwischen Bimatoprost und Vehikelgruppe (p<0,05, Kreuze) in Telogen,
Gesamtanagen und Anagen-middle/late.
Tabelle: Prozentangaben der Phasenverteilung.
Der Durchmesser der Follikel in Anagen-early vergrößerte sich unter
Bimatoprostbehandlung. Da der Durchmesser des Haarschaftes direkt mit dem
Volumen der dermalen Papille korreliert,[70],[73] wurde der maximale
Durchmesser der Papille und des Bulbus unabhängig voneinander in Anagen-
early und Anagen-late gemessen (Tab. 2). Der Durchmesser des Bulbus unter
Bimatoprostbehandlung war signifikant größer in Anagen-early, verglichen mit
Vehikelbehandlung, nicht jedoch in Anagen-late. Diese Ergebnisse deuteten
darauf hin, dass die durch Bimatoprost induzierte Verdickung der
Wimpernschafte mit der Verdickung der dermalen Papille korrelierte.
Anagen-early Anagen-late
dermale Papille Bulbus dermale Papille Bulbus
Vehikel 18,4±1,1 25,1±3,0 27,9±2,5 69,4±4,7
Bimatoprost 20,25±0,8 32,5±2,4*
25,8±1,6 68,6±3,6
Tab. 2: Durchmesser der dermalen Papille und des Bulbus.
Mittelwert der Länge in µm ± SEM, n=22 (Vehikel, Anagen-early), n=33 (Bimatoprost,
Anagen early), n=15 (Vehikel, Anagen-late), n=12 (Bimatoprost Anagen-late).
*: Signifikanter Unterschied (p<0,05)

39
Ergebnisse
Die Anzahl der Melaningranula unter Bimatoprostbehandlung war nicht
signifikant erhöht. Da die Hyperpigmentierung der Wimpern in Folge von
Therapie mit PGF-2α Analoga beschrieben wurde,[27] wurde die Anzahl der
Melaningranula im Bulbus von Follikeln in Anagen-late analysiert. Die Anzahl
der Melaningranula betrug 1,13±0,06 unter Vehikelbehandlung und 1,44±0,18
unter Bimatoprostbehandlung (Mittelwert pro µm2±SEM). Es konnte kein
signifikanter Unterschied zwischen den Behandlungsgruppen nachgewiesen
werden (Fig. 21).
Fig. 21: Zahl der Melaningranula.
Die Anzahl der Melaningranula pro µm
in Anagen-late schien in der
Bimatoprostgruppe höher, war jedoch
nicht signifikant (n=12 pro Gruppe).

40
Diskussion
5. Diskussion
5.1 Vergleich der Morphologie der Wimpern der Maus mit
menschlichen Wimpern und mit den Fellhaaren
Die Morphologie des Augenlids der Maus glich im Wesentlichen der des
menschlichen Augenlids.[53] Analog zu den menschlichen Wimpern[42]
existierten auch bei der Maus Wimpern unterschiedlicher Länge, Dicke und
Pigmentierung, sowie mit unterschiedlich aufgebauter Medulla. Betrachtete man
den Wimpernschaft, so wiesen die Wimpern der Maus, verglichen mit dem
dorsalen Hauthaar, einen dickeren Kortex, eine stärkere Pigmentierung und
eine gerade Struktur ohne Knicke auf, wie sie in dorsalen Zig-Zag Haaren der
Maus beobachtet wurden.[15] Dies untermauert die Funktion der Wimpern zum
Schutz vor Partikeln oder Sonnenlicht. Der Aufbau der Medulla der Wimper
ähnelte dem Monotrich-Haar.[13] (auch als Guard-Haar bezeichnet[15],[60]). Im
Gegensatz dazu zählte der größte Anteil dorsaler Fellhaare zur Gruppe der Zig-
Zag Haare.[15] Dies könnte auf einen unterschiedlichen Ursprung oder
verschiedene Regulationsmechanismen der Wimpern im Gegensatz zu
Fellhaaren hindeuten. Die Wimpern waren in der Regel gut pigmentiert, wobei
die Pigmentierung bei dicken Wimpern stärker ausgeprägt war.
5.2 Vergleich der Morphogenese und der frühen Entwicklung der
Wimpern mit der der Fellhaare
Analysen hinsichtlich der Morphogenese und der frühen Entwicklung der
Wimpern wurden zur Planung des optimalen zeitlichen Ablaufs der
experimentellen Studie durchgeführt. Im Wesentlichen unterschied sich im
Modell der Maus die Morphogenese der Wimpernfollikel nicht von der
Morphogenese dorsaler Fellhaarfollikel. Die makroskopischen Beobachtungen
hinsichtlich der austretenden Wimpernschafte deuteten auf einen ähnlichen
zeitlichen Ablauf der Morphogenese, verglichen mit dem dorsalen Fellhaar,
hin.[50] Im Anschluss an die Morphogenese durchliefen die Wimpern den ersten
synchronisierten Zyklus analog zum dorsalen Fellhaar, welcher jedoch etwas
früher begann, nämlich schon in der zweiten Woche, und schneller
voranschritt.[49] Die in der Haarwissenschaft oft als Angriffspunkt benutzte

41
Diskussion
synchrone Telogenphase dorsaler Fellhaare, in der siebten Woche p.p., am
Ende des ersten Zyklus vor der Transition in den asynchronen Zyklus,
entspräche bei den Wimpern wohl am ehesten der vierten Woche post partum.
5.3 Vergleich des Wachstumszyklus und des Exogens der Wimper
mit der dorsaler Fellhaare
Gerade in taktilen Haaren und damit auch in Guard-Haaren und Tasthaaren ist
Exogen ein streng regulierter Prozess, vermutlich um mit nur einem Kolbenhaar
pro Follikel die taktile Funktion zu gewährleisten.[13],[24] Im Gegensatz dazu
findet das Effluvium dorsaler Fellhaare nicht während jedes Zyklus statt.[39] Das
Exogen des Hauthaares ereignet sich während Anagen,[39] im Tasthaar läuft es
während des Überganges von Anagen zu Katagen ab,[13],[24] in der Wimper
während des Katagens. Die Daten dieser Studie zeigten, dass das Exogen der
Wimpern analog zu taktilen Haaren während des Durchlaufens jedes Zyklus
stattfand. Fehlregulationen des Exogen könnten hinsichtlich des
Pathomechanismus der Trichomegalie eine wichtige Rolle spielen.
5.4 Effekte von Bimatoprostbehandlung
Die Untersuchungen dieser Studie zeigten, dass durch Bimatoprost bei der
Maus tatsächlich Trichomegalie erzeugt werden konnte, wie es beim Menschen
beschrieben ist.[34],[35],[44],[71] Trotz der Zunahme der Anzahl der Wimpernschafte
nahm die Anzahl der Wimpernfollikel nicht zu. Es gab es in dieser Studie keinen
Hinweis für de-novo Follikelerzeugung. Die Induktion einer de-novo
Follikelgeneration aus adultem Epithel ist beispielsweise durch Überexpression
des Wnt/β-Catenin Pathways möglich.[16],[26] Diese Studie ergab jedoch keine
Hinweise auf morphologisch unreife oder anormale Wimpernfollikel, welche
Follikelneogenese aus dem mit Bimatoprost behandelten Epithel
wiederspiegeln würden. Eine de-novo Generation von Follikeln wurde ebenso
nicht beim Auftragen anderer PGF-2α Analoga auf dorsale Haut beschrieben.[59]
Viele Haarwachstum stimulierende Substanzen, wie Cyclosporin, Minoxidil oder
PGF-2α Analoga induzieren den Anagen-Eintritt von ruhenden
Haarfollikeln.[36],[38],[59] Dennoch konnte ein Anstieg der Wimpernzahl nicht allein

42
Diskussion
durch die Induktion von Anagen und einen daraus resultierenden schnelleren
Progress des Wachstumszyklus erklärt werden, da die Wimpernfollikel Exogen
während jedes Zyklus durchliefen und damit jeder Wimpernfollikel nur eine
Kolbenwimper besaß.
Eine mögliche Erklärung für die steigende Anzahl der Wimpern durch
Bimatoprost Therapie könnte eine steigende Anzahl der Wimpernschafte bei
gleich bleibender Follikelanzahl sein. Das Exogen der Wimpern fand während
jedes Zyklus in jedem Follikel statt, so dass die Follikel während Anagen-late
bis Katagen-early ein zwei-Schaft-pro-Follikel Verhältnis aufwiesen.
Bimatoprost beeinflusste die Regulation von Exogen und das ein-Schaft-pro-
Follikel Verhältnis nicht. Doch Bimatoprost beeinflusste die Anagenphase, was
zu einer erhöhten Anzahl der Follikel in der zwei-Schaft-pro-Follikel Phase
führte, im Vergleich zu den mit Vehikel behandelten Wimpern. Dies würde eine
erhöhte Anzahl an Wimpernschaften ohne steigende Follikelanzahl erklären.
5.5 Zunahme der Wimperndicke und Hyperpigmentierung durch
Bimatoprost
Es ist bekannt, dass PGF-2α Analoga Vellushaar in terminales ausgereiftes
Haar, beispielsweise im Bereich der Wangen- und Kanthalregion,
umwandeln[20],[27] und die Melanogenese in Haarfollikeln aktivieren.[59] In dieser
Studie induzierte die Bimatoprostbehandlung eine Zunahme der Dicke der
mittellangen und kurzen Wimpern, welche durch die histologische Evaluation
der Bulbusdicke weiter verifiziert wurde. Da Veränderungen hinsichtlich der
Form und Dicke nach dem Entstehen der massiven inneren Wurzelscheide
während des Wachstumszyklus nur schwer möglich sind,[28] wird die Dicke und
Form der Wimper in Anagen-early bestimmt. Zwar war die Behandlung in dieser
Studie kurz, doch resultierte eine effektive Verdickung der Anagen-early
Follikel. Dennoch schien eine makroskopische Verdickung der mittellangen und
kurzen Wimpern im Widerspruch zur nicht nachgewiesenen Vergrößerung der
Follikel in Anagen-late zu stehen. Ursache hierfür könnte die kurze
Behandlungsdauer und die geringe Anzahl der Follikel in Anagen-late sein,
welche die Evaluationskriterien erfüllten, so dass eine Größenzunahme der
Follikel nicht signifikant validiert werden konnte.

43
Diskussion
Eine Hyperpigmentierung durch PGF-2α Analoga wurde nicht nur in
Wimpern,[74],[27] sondern auch in der Iris,[1],[10],[61],[67] dem Augenlid,[27]
periokulär[23],[75] und im dorsalen Fellhaarfollikel[59] beschrieben. In der Iris war
die Verdunklung auf eine Größenzunahme der Melaningranula ohne
Einflussnahme auf die Anzahl zurückzuführen.[9],[10] In dieser Studie konnte
ebenfalls keine Zunahme der Melaningranula durch die Bimatoprost Therapie
nachgewiesen werden, eine Veränderung der Melaningranula wäre möglich,
war jedoch lichtmikroskopisch nicht messbar. Die unterschiedliche Dichte der
Melaningranula im selben Follikel, die variierende Größe der Follikel selbst und
die lichtmikroskopisch nicht messbare Größe der Melaningranula, limitierten die
morphometrische Methodik.
5.6 Wimpernverlängerung durch Bimatoprost
Die durch Bimatoprost induzierte Wimpernverlängerung wurde in dieser Studie
indirekt durch die Bestimmung der Wachstumsphase jedes einzelnen Follikels
dargestellt. Da die Dauer der Anagen-Phase die Länge der Wimper
bestimmt,[48] deuteten die Daten dieser Studie auf eine Verlängerung der
Wimpern als Folge einer Beeinflussung des Wachstumszyklus hin. Die
prozentuale Verteilung der Wachstumsphasen unter Bimatoprost Therapie
könnte dahingehend interpretiert werden, dass Telogen Wimpernfollikel früher
in die Anagenphase eintraten. Diese Induktion der Telogen zu Anagen
Konversion stimmt mit beschriebenen Mechanismen durch Applikation von
PGF-2α Analoga auf dorsale Fellhaare überein.[59] Dennoch resultiert nicht
allein aus der Induktion des Anagen eine längere Dauer der Anagenphase. In
dieser Studie wurde gezeigt, dass der Anteil der induzierten Anagen Follikel
nicht in Anagen-early, sondern in Anagen-middle/late größer war. Dies indiziert,
dass Bimatoprost hauptsächlich Anagen-middle/late prolongierte, indem es
möglicherweise den Übergang in Katagen behinderte. Während der Anagen-
middle/late Phase der Wimpern, der äquivalenten Anagen IIIa-c Phase der
dorsalen Fellhaarfollikel, entstand der Haarschaft und es wurde aktive
Keratinisierung beobachtet.

44
Literaturverzeichnis
6. Literaturverzeichnis
1 Albert D. M., Gangnon R. E., Grossniklaus H. E., Green W. R., Darjatmoko S. and Kulkarni A. D. (2008). "A study of histopathological features of latanoprost-treated irides with or without darkening compared with non-latanoprost-treated irides." Arch Ophthalmol 126(5): 626-31.
2 Bayer A., Bagkesen H. and Sobaci G. (2007). "Acquired trichomegaly in uveitis." Can J Ophthalmol 42(1): 101-6.
3 Bergstrom S. and Samuelsson B. (1962). "Isolation of prostaglandin E1 from human seminal plasma. Prostaglandins and related factors. 11." J Biol Chem 237: 3005-6.
4 Carser J. E. and Summers Y. J. (2006). "Trichomegaly of the eyelashes after treatment with erlotinib in non-small cell lung cancer." J Thorac Oncol 1(9): 1040-1.
5 Cedrone C., Mancino R., Cerulli A., Cesareo M. and Nucci C. (2008). "Epidemiology of primary glaucoma: prevalence, incidence, and blinding effects." Prog Brain Res 173: 3-14.
6 Centofanti M., Oddone F., Chimenti S., Tanga L., Citarella L. and Manni G. (2006). "Prevention of dermatologic side effects of bimatoprost 0.03% topical therapy." Am J Ophthalmol 142(6): 1059-60.
7 Chase H. B. (1954). "Growth of the hair." Physiol Rev 34(1): 113-26. 8 Choy I. and Lin S. (2008). "Eyelash enhancement properties of topical
dechloro ethylcloprostenolamide." J Cosmet Laser Ther 10(2): 110-3. 9 Cracknell K. P., Farnell D. J. and Grierson I. (2007). "Monte Carlo
simulation of latanoprost induced iris darkening." Comput Methods Programs Biomed 87(2): 93-103.
10 Cracknell K. P., Grierson I., Hogg P., Appleton P. and Pfeiffer N. (2003). "Latanoprost-induced iris darkening: a morphometric study of human peripheral iridectomies." Exp Eye Res 77(6): 721-30.
11 Cruz A. A., Zenha F., Silva J. T., Jr. and Martinez R. (2004). "Eyelid involvement in paracoccidioidomycosis." Ophthal Plast Reconstr Surg 20(3): 212-6.
12 Deplewski D. and Rosenfield R. L. (2000). "Role of hormones in pilosebaceous unit development." Endocr Rev 21(4): 363-92.
13 Dry F.W. (1926). "The coat of the mouse (Mus Musculus)." Journal of Genetics 16: 287-340.
14 Elgin U., Batman A., Berker N. and Ilhan B. (2006). "The comparison of eyelash lengthening effect of latanoprost therapy in adults and children." Eur J Ophthalmol 16(2): 247-50.
15 Fraser A.S. (1951). "Growth of the mouse coat." J Exp Zool 117: 15-29. 16 Gat U., DasGupta R., Degenstein L. and Fuchs E. (1998). "De Novo hair
follicle morphogenesis and hair tumors in mice expressing a truncated beta-catenin in skin." Cell 95(5): 605-14.
17 Gryglewski R. J. (2008). "Prostacyclin among prostanoids." Pharmacol Rep 60(1): 3-11.
18 Hamada K., Thornton M. J., Laing I., Messenger A. G. and Randall V. A. (1996). "The metabolism of testosterone by dermal papilla cells cultured from human pubic and axillary hair follicles concurs with hair growth in 5 alpha-reductase deficiency." J Invest Dermatol 106(5): 1017-22.

45
Literaturverzeichnis
19 Hamberg M. and Samuelsson B. (1973). "Detection and isolation of an endoperoxide intermediate in prostaglandin biosynthesis." Proc Natl Acad Sci U S A 70(3): 899-903.
20 Hart J. and Shafranov G. (2004). "Hypertrichosis of vellus hairs of the malar region after unilateral treatment with bimatoprost." Am J Ophthalmol 137(4): 756-7.
21 Herane M. I. and Urbina F. (2004). "Acquired trichomegaly of the eyelashes and hypertrichosis induced by bimatoprost." J Eur Acad Dermatol Venereol 18(5): 644-5.
22 Hernandez-Nunez A., Fernandez-Herrera J., Buceta L. R. and Garcia-Diez A. (2002). "Trichomegaly following treatment with interferon alpha-2b." Lancet 359(9312): 1107.
23 Herndon L. W., Robert D. Williams, Wand M. and Asrani S. (2003). "Increased periocular pigmentation with ocular hypotensive lipid use in African Americans." Am J Ophthalmol 135(5): 713-5.
24 Higgins C. A., Westgate G. E. and Jahoda C. A. (2009). "From telogen to exogen: mechanisms underlying formation and subsequent loss of the hair club fiber." J Invest Dermatol 129(9): 2100-8.
25 Ishida N., Odani-Kawabata N., Shimazaki A. and Hara H. (2006). "Prostanoids in the therapy of glaucoma." Cardiovasc Drug Rev 24(1): 1-10.
26 Ito M., Yang Z., Andl T., Cui C., Kim N., Millar S. E. and Cotsarelis G. (2007). "Wnt-dependent de novo hair follicle regeneration in adult mouse skin after wounding." Nature 447(7142): 316-20.
27 Johnstone M. A. (1997). "Hypertrichosis and increased pigmentation of eyelashes and adjacent hair in the region of the ipsilateral eyelids of patients treated with unilateral topical latanoprost." Am J Ophthalmol 124(4): 544-7.
28 Johnstone M. A. and Albert D. M. (2002). "Prostaglandin-induced hair growth." Surv Ophthalmol 47 Suppl 1: S185-202.
29 Jordan D. R., Ahuja N. and Khouri L. (2002). "Eyelash loss associated with hyperthyroidism." Ophthal Plast Reconstr Surg 18(3): 219-22.
30 Kaplan M. H., Sadick N. S. and Talmor M. (1991). "Acquired trichomegaly of the eyelashes: a cutaneous marker of acquired immunodeficiency syndrome." J Am Acad Dermatol 25(5 Pt 1): 801-4.
31 Kerob D., Dupuy A., Reygagne P., Levy A., Morel P., Bernard B. A. and Lebbe C. (2006). "Facial hypertrichosis induced by Cetuximab, an anti-EGFR monoclonal antibody." Arch Dermatol 142(12): 1656-7.
32 Kowing D. (2005). "Madarosis and facial alopecia presumed secondary to botulinum a toxin injections." Optom Vis Sci 82(7): 579-82.
33 Liang Y., Woodward D. F., Guzman V. M., Li C., Scott D. F., Wang J. W., Wheeler L. A., Garst M. E., Landsverk K., Sachs G., Krauss A. H., Cornell C., Martos J., Pettit S. and Fliri H. (2008). "Identification and pharmacological characterization of the prostaglandin FP receptor and FP receptor variant complexes." Br J Pharmacol 154(5): 1079-93.
34 Manni G., Centofanti M., Parravano M., Oddone F. and Bucci M. G. (2004). "A 6-month randomized clinical trial of bimatoprost 0.03% versus the association of timolol 0.5% and latanoprost 0.005% in glaucomatous patients." Graefes Arch Clin Exp Ophthalmol 242(9): 767-70.

46
Literaturverzeichnis
35 Mansberger S. L. and Cioffi G. A. (2000). "Eyelash formation secondary to latanoprost treatment in a patient with alopecia." Arch Ophthalmol 118(5): 718-9.
36 Maurer M., Handjiski B. and Paus R. (1997). "Hair growth modulation by topical immunophilin ligands: induction of anagen, inhibition of massive catagen development, and relative protection from chemotherapy-induced alopecia." Am J Pathol 150(4): 1433-41.
37 Mehta J. S., Raman J., Gupta N. and Thoung D. (2003). "Cutaneous latanoprost in the treatment of alopecia areata." Eye (Lond) 17(3): 444-6.
38 Messenger A. G. and Rundegren J. (2004). "Minoxidil: mechanisms of action on hair growth." Br J Dermatol 150(2): 186-94.
39 Milner Y., Sudnik J., Filippi M., Kizoulis M., Kashgarian M. and Stenn K. (2002). "Exogen, shedding phase of the hair growth cycle: characterization of a mouse model." J Invest Dermatol 119(3): 639-44.
40 Motl S. E. and Fausel C. (2003). "Recurring chemotherapy-associated alopecia areata: case report and literature review." Pharmacotherapy 23(1): 104-8.
41 Muller-Rover S., Handjiski B., van der Veen C., Eichmuller S., Foitzik K., McKay I. A., Stenn K. S. and Paus R. (2001). "A comprehensive guide for the accurate classification of murine hair follicles in distinct hair cycle stages." J Invest Dermatol 117(1): 3-15.
42 Na J. I., Kwon O. S., Kim B. J., Park W. S., Oh J. K., Kim K. H., Cho K. H. and Eun H. C. (2006). "Ethnic characteristics of eyelashes: a comparative analysis in Asian and Caucasian females." Br J Dermatol 155(6): 1170-6.
43 Nakamura M., Sundberg J. P. and Paus R. (2001). "Mutant laboratory mice with abnormalities in hair follicle morphogenesis, cycling, and/or structure: annotated tables." Exp Dermatol 10(6): 369-90.
44 Noecker R. S., Dirks M. S. and Choplin N. (2004). "Comparison of latanoprost, bimatoprost, and travoprost in patients with elevated intraocular pressure: a 12-week, randomized, masked-evaluator multicenter study." Am J Ophthalmol 137(1): 210-1; author reply 211-2.
45 Oh D. J., Martin J. L., Williams A. J., Russell P., Birk D. E. and Rhee D. J. (2006). "Effect of latanoprost on the expression of matrix metalloproteinases and their tissue inhibitors in human trabecular meshwork cells." Invest Ophthalmol Vis Sci 47(9): 3887-95.
46 Oliver G. L. and McFarlane D. C. (1965). "Congenital Trichomegaly: With Associated Pigmentary Degeneration of the Retina, Dwarfism, and Mental Retardation." Arch Ophthalmol 74: 169-71.
47 Pascual J. C., Banuls J., Belinchon I., Blanes M. and Massuti B. (2004). "Trichomegaly following treatment with gefitinib (ZD1839)." Br J Dermatol 151(5): 1111-2.
48 Paus R. and Cotsarelis G. (1999). "The biology of hair follicles." N Engl J Med 341(7): 491-7.
49 Paus R., Muller-Rover S., Van Der Veen C., Maurer M., Eichmuller S., Ling G., Hofmann U., Foitzik K., Mecklenburg L. and Handjiski B. (1999). "A comprehensive guide for the recognition and classification of distinct stages of hair follicle morphogenesis." J Invest Dermatol 113(4): 523-32.
50 Paus R., van der Veen C., Eichmuller S., Kopp T., Hagen E., Muller-Rover S. and Hofmann U. (1998). "Generation and cyclic remodeling of the hair follicle immune system in mice." J Invest Dermatol 111(1): 7-18.

47
Literaturverzeichnis
51 Pucci N., Massai C., Bernardini R., Caputo R., Mori F., De Libero C., Novembre E., De Martino M. and Vierucci A. (2007). "Eyelash length in children with vernal keratoconjunctivitis: effect of treatment with cyclosporine eye drops." Int J Immunopathol Pharmacol 20(3): 595-9.
52 Randall V. A., Hibberts N. A., Thornton M. J., Hamada K., Merrick A. E., Kato S., Jenner T. J., De Oliveira I. and Messenger A. G. (2000). "The hair follicle: a paradoxical androgen target organ." Horm Res 54(5-6): 243-50.
53 Rohen J. W. (1979). "[Functional anatomy of the eyelids]." Ber Zusammenkunft Dtsch Ophthalmol Ges(77): 3-12.
54 Roseborough I., Lee H., Chwalek J., Stamper R. L. and Price V. H. (2009). "Lack of efficacy of topical latanoprost and bimatoprost ophthalmic solutions in promoting eyelash growth in patients with alopecia areata." J Am Acad Dermatol 60(4): 705-6.
55 Ross E. K., Bolduc C., Lui H. and Shapiro J. (2005). "Lack of efficacy of topical latanoprost in the treatment of eyebrow alopecia areata." J Am Acad Dermatol 53(6): 1095-6.
56 Sachdeva S. and Prasher P. (2008). "Madarosis: a dermatological marker." Indian J Dermatol Venereol Leprol 74(1): 74-6.
57 Santiago M., Travassos A. C., Rocha M. C. and Souza S. (2000). "Hypertrichosis in systemic lupus erythematosus (SLE)." Clin Rheumatol 19(3): 245-6.
58 Santmyire-Rosenberger B. R. and Albert M. (2005). "Acquired trichomegaly with topiramate." J Am Acad Dermatol 53(2): 362-3.
59 Sasaki S., Hozumi Y. and Kondo S. (2005). "Influence of prostaglandin F2alpha and its analogues on hair regrowth and follicular melanogenesis in a murine model." Exp Dermatol 14(5): 323-8.
60 Schlake T. (2007). "Determination of hair structure and shape." Semin Cell Dev Biol 18(2): 267-73.
61 Selen G., Stjernschantz J. and Resul B. (1997). "Prostaglandin-induced iridial pigmentation in primates." Surv Ophthalmol 41 Suppl 2: S125-8.
62 Selva D., Chen C. S., James C. L. and Huilgol S. C. (2003). "Discoid lupus erythematosus presenting as madarosis." Am J Ophthalmol 136(3): 545-6.
63 Shaikh M. Y. and Bodla A. A. (2006). "Hypertrichosis of the eyelashes from prostaglandin analog use: a blessing or a bother to the patient?" J Ocul Pharmacol Ther 22(1): 76-7.
64 Sharma R. C., Mahajan V. K., Sharma N. L. and Sharma A. (2002). "Trichomegaly of the eyelashes in dermatomyositis." Dermatology 205(3): 305.
65 Smith W. L., Marnett L. J. and DeWitt D. L. (1991). "Prostaglandin and thromboxane biosynthesis." Pharmacol Ther 49(3): 153-79.
66 Stenn K. S. and Paus R. (2001). "Controls of hair follicle cycling." Physiol Rev 81(1): 449-494.
67 Stjernschantz J. W., Albert D. M., Hu D. N., Drago F. and Wistrand P. J. (2002). "Mechanism and clinical significance of prostaglandin-induced iris pigmentation." Surv Ophthalmol 47 Suppl 1: S162-75.
68 Sundberg J. P., Peters E. M. and Paus R. (2005). "Analysis of hair follicles in mutant laboratory mice." J Investig Dermatol Symp Proc 10(3): 264-70.

48
Literaturverzeichnis
69 Tauchi M., Fuchs T.A., Kellenberger A.J., Woodward D.F., Paus R. Lütjen-Drecoll E. (2010) „Characterization of an in vivo model for the study of eyelash biology and trichomegaly: mouse eyelash morphology, developement, growth cycle, and anagen prolongation by bimatoprost“ ©
2010 Br J Dermatol: 162(6) 1186-97 Mit freundlicher Genehmigung von John Wiley und Söhne
70 Tobin D. J., Gunin A., Magerl M. and Paus R. (2003). "Plasticity and cytokinetic dynamics of the hair follicle mesenchyme during the hair growth cycle: implications for growth control and hair follicle transformations." J Investig Dermatol Symp Proc 8(1): 80-6.
71 Tosti A., Pazzaglia M., Voudouris S. and Tosti G. (2004). "Hypertrichosis of the eyelashes caused by bimatoprost." J Am Acad Dermatol 51(5 Suppl): S149-50.
72 Uno H., Zimbric M. L., Albert D. M. and Stjernschantz J. (2002). "Effect of latanoprost on hair growth in the bald scalp of the stump-tailed macacque: a pilot study." Acta Derm Venereol 82(1): 7-12.
73 Van Scott E. J. and Ekel T. M. (1958). "Geometric relationships between the matrix of the hair bulb and its dermal papilla in normal and alopecic scalp." J Invest Dermatol 31(5): 281-7.
74 Wand M. (1997). "Latanoprost and hyperpigmentation of eyelashes." Arch Ophthalmol 115(9): 1206-8.
75 Wand M., Ritch R., Isbey E. K., Jr. and Zimmerman T. J. (2001). "Latanoprost and periocular skin color changes." Arch Ophthalmol 119(4): 614-5.
76 Ward K. M., Barnett C., Fox L. P. and Grossman M. E. (2006). "Eyelash trichomegaly associated with systemic tacrolimus." Arch Dermatol 142(2): 248.
77 Weaver D. T. and Bartley G. B. (1990). "Cyclosporine-induced trichomegaly." Am J Ophthalmol 109(2): 239.
78 Wolf R., Matz H., Zalish M., Pollack A. and Orion E. (2003). "Prostaglandin analogs for hair growth: great expectations." Dermatol Online J 9(3): 7.
79 Woodward D. F., Liang Y. and Krauss A. H. (2008). "Prostamides (prostaglandin-ethanolamides) and their pharmacology." Br J Pharmacol 153(3): 410-9.

49
Abkürzungsverzeichnis
7. Abkürzungsverzeichnis
A(e) Anagen-early
A(l) Anagen-late
A(m) Anagen-middle
ant. anterior
Äqu. Äquivalenz
bzw. beziehungsweise
C Kohlenstoff
ca. circa
COX Cyclooxygenase
DP dermale Papille
EGRF epidermal growth factor receptor
Fig. Figur
Gl. Glandula
Gl. s. Glandula sebacea
Gll. t. Glandulae tarsales
HIV Human Immunodeficiency Virus
IOP intraocular pressure (Intraokularer Druck)
IRS innere Wurzelscheide (inner root sheath)
K(e) Katagen-early
KL kontralateral
K(l) Katagen-late
Limb. Limbus
M. Musculus
MG Melaningranula
M. o. Musculus orbicularis oculi
N. Nervus
n Anzahl
ns nicht signifikant
ORS äußere Wurzelscheide (outer root sheath)
palp. palpebralis
PBS phosphate buffered saline
PGE Prostaglandin E

50
Abkürzungsverzeichnis
PGF Prostaglandin F
PGF-2α Prostaglandin F-2α
PGG Prostaglandin G
post. posterior
p.p. post partum
SEM standard error of the mean
St. Stadium
T Telogen
Tab. Tabelle
v.a. vor allem
WS Wimpernschaft
µm µMeter

51
Verzeichnis der Vorveröffentlichungen
8. Verzeichnis der Vorveröffentlichungen
M. Tauchi, T.A. Fuchs, D. F. Woodward, E. Lütjen-Drecoll “Characterization of
Eyelash Growth in Mice.” The 13th Annual Meeting of European Hair Research
Society, Genoa, Italy (2008)
M. Tauchi, T.A. Fuchs, D.F. Woodward, E. Lütjen-Drecoll “Bimatoprost-induced
hypertrichosis of eyelashes: murine model.” The Association for Research in
Vision and Ophthalmology, annual meeting, Fort Lauderdale, Florida, U.S.A.
(2009)
Tauchi M., Fuchs T.A., Woodward D.F., Lütjen-Drecoll E. “Murine eyelashes as
a study model for hypertrichosis or alopecia of eyelashes.” The Society of
Investigational Dermatology, annual Meeting, Montreal, Canada (2009)
Tauchi M, Fuchs TA, Woodward DF, Lütjen-Drecoll E “Murine Eyelashes as a
Study Model for Hypertrichosis or Alopecia of Eyelashes.” The 14th annual
Meeting of European Hair Research Society, Graz, Austria (2009)
M. Tauchi, T.A. Fuchs, A.J. Kellenberger, D.F. Woodward, R. Paus, E. Lütjen-
Drecoll “Characterization of an in vivo-model for the study of eyelash biology.”
The 6th World Congress for Hair Research, Cairns, Australia (2010)
M. Tauchi, T.A. Fuchs, A.J. Kellenberger, D.F. Woodward, R. Paus, E. Lütjen-
Drecoll. “Characterization of an in vivo-model for the study of eyelash biology
and trichomegaly: Mouse eyelash morphology, development, growth cycle, and
anagen prolongation by bimatoprost.” British Journal of Dermatology (2010)

52
Lebenslauf
9. Danksagung
Mein größter Dank gilt Frau Prof. Dr. med. Lütjen-Drecoll, Direktorin des
Anatomischen Institutes II der Universität Erlangen-Nürnberg, die mich von
Anfang an für die Anatomie begeistert hat. Ich danke ihr ganz herzlich für die
Themenstellung. Ohne ihre uneingeschränkte Förderung, ihre Unterstützung,
ihren Antrieb und ihr Engagement wäre diese Arbeit nie möglich gewesen.
Frau Dr. med. vet. Dr. rer. nat. Miyuki Tauchi, wissenschaftliche Mitarbeiterin
des Anatomischen Institutes II, an die ich mich jederzeit bei Fragen jeglicher Art
wenden konnte, danke ich sehr für ihre Anleitung und Einführung in die
Thematik, ihre Hilfe und ihre freundliche Art.
Den medizinisch-technischen Assistentinnen Elke Kretzschmar, Gertrud Link
und Anke Fischer danke ich für ihre technische Expertise und Unterstützung.
Meinen Eltern möchte ich an dieser Stelle ganz herzlich danken, dass sie mir
das Studium ermöglicht und mich immer gefördert haben. Lucia möchte ich für
die liebevolle Unterstützung danken.