aspects oflife history and of territorial behavior inyoung
TRANSCRIPT

Pacific Science (1975), Vol. 29, No.4, p. 341-348Printed in Great Britain
Aspects of Life History and ofTerritorial Behavior in Young Individuals of Platynereis bicanaliculata
and Nereis vexillosa (Annelida, Polychaeta) I
PAMELA ROE2
ABSTRACT: Plarynereis bicanaliculata (Baird), an annual nereid, spawned in earlyAugust at two areas in Washington state. Spawning was highly synchronous.Young were planktonic for about 1 week. Within 3 weeks they had grown to 4 mmin length, had started building tubes of mucus and diatoms, and showed a period ofrapid growth in size. By the end of September or early October they averaged 10mm in length, at which size they remained until March. In spring they reachedadult length (20-23 mm) and during the summer gametes developed.
Nereis vexillosa Grube egg masses were found from March through August.Nereis has a 2-year life span in both study areas, growing to one-half adult size the1st year and to mature size the 2nd year. In the laboratory, young made tubes within1 week after hatching from egg masses.
Members of both species defend their tubes from intruders. Usually, larger individuals win fighting encounters, especially if they are the occupants of tubes. Smallindividuals successfully defend their tubes from larger individuals in about onehalf of the encounters; and if fights occur between equal sized individuals, occupants are usually not displaced. Fights are real, with jaws used much for biting, andsmaller individuals are sometimes actually eaten by larger ones, especially In Nereisvexillosa. In the laboratory the number of individuals of N. vexillosa kept in fingerbowls decreased in number until only one or two large individuals remained.
MATERIALS AND METHODS
feeding, is discussed, and the effect of nereidterritorial behavior on mud flat communitystructure is suggested.
Nereids were studied from two intertidal mudflats on San Juan Island, Washington: one atMitchell Bay (48°34'9.5" N, 123°9'48" W) andone at Garrison Bay (48°34'57" N, 123°9'17"W). At Mitchell Bay ulvoids were abundantand patches of these algae remained throughoutthe year. At Garrison Bay ulvoids were presentin spring and summer, but were absent throughout winter; other algae were also scarce then.The Mitchell Bay area had some coarse sand andgravel; the Garrison Bay site was soft mud overa hard-packed clay.
Plarynereis bicanaliculata was collected in alStanislaus, DepartmentMonte Vista Avenue, ternate months at the two areas. A primary
transect was laid high in the intertidal; at 20-m
341 22-2
NEREIDS can be abundant and important members of intertidal mud flat communities (Roe1971, Woodin 1974). This paper describes territorial behavior in newly settled young andcompares life history and territorial behaviorin two species, Plarynereis bicanaliculata (Baird)and Nereis vexillosa Grube. Territorial behaviorof several nereid species has been studied in thelaboratory (e.g., Reish and Alosi 1968, Evans1973) using fully grown but usually nonripeanimals. This is the first description of suchbehavior in newly settled nereids. The adaptivesignificance of this territorial behavior, evidenteven before young animals have commenced
I This work was supported in part by the FridayBarbor Laboratories and National Science Foundationgrants GB-6518X and GB-20978 to the Department ofZoology, University of Washington. Manuscript received 16 March 1975.
2 California State College,of Biological Sciences, 800Turlock, California 95380.

342
intervals along this transect, transects perpendicular to it and leading to the lower intertidalwere laid, and every 10 m along these secondarytransects a wooden 1 m2 quadrat was placed.A jar with a surface area at the opening of 20cm2, inverted and pressed into the mud in thecenter of the quadrat, was used to collect mud.During summer when nereids were larger andless abundant, plastic freezer boxes with a surface area of100 cm2 were used. In the laboratory,contents of each container were emptied withsea water into a Pyrex dish and sloshed arounduntil all nereids swam to the water surface.They were counted, and nereids from thesamples with the most individuals were preserved in 70-percent ethyl alcohol and theirlengths were later measured for size frequencyanalysis.
Nereis vexillosa was collected in 1/8 m2 samples of sieved mud from both areas in alternatemonths. Individuals were measured only 1month, but casual observations of the sizes ofother specimens support data for that month.
Egg masses of Nereis vexillosa were broughtfrom both study areas to the Friday HarborLaboratories, placed in fingerbowls in runningsea water (about 100 C), and observed.
Plarynereis trochophores collected from theplankton of Mitchell and Garrison bays werekept in fingerbowls in running sea water(100 C) and observed. All observations of territorial behavior were made in the laboratory,with young nereids in 4.5-inch (11.4 em)diameter fingerbowls being used.
Sizes of newly hatched nereids usually wererecorded as how many segments had grownrather than by actual size.
During fall and winter nereids were held in4- or 16-ounce jars in a refrigerator kept near70 C.
Young nereids were fed diatoms that grew onthe sides of the running seawater tanks; in falland winter they were fed fresh or dried ulvoidsbroken into small pieces. Growth and behaviorwere recorded.
The work by Berkeley and Berkeley (1948)was used to identify Nereis vexillosa; Hartman'swork (1954) was used for Plarynereis bicanaliculata.
PACIFIC SCIENCE, Volume 29, October 1975
OBSERVATIONS AND RESULTS
Life History of Platynereis bicanaliculata (Baird,1863)
Plarynereis is an annual; thousands of smalleggs (132 # in diameter) are spawned by eachfemale into the open water, and, after spawning, both sexes, which have formed epitokalbreeding swarms, die. Spawning is highly synchronous; I observed nearly all individuals ofthe P. bicanaliculata population at Mitchell Bayswarming and spawning gametes near midnighton 3 August 1969, and I observed the beginningof epitokal swarming at 2230 hours on 31 July1968. Much of the Plarynereis population atGarrison Bay spawned the same nights in 1968and 1969 as did the Mitchell Bay population, asevidenced by the very large numbers of gastrulae that I observed in the plankton of bothareas on 4 August 1969. That spawning ishighly synchronous is evidenced by the factthat I saw only nine individuals swimming on29 July 1969 (2300-2330 hours) and only 17 on1-2 August 1969 (2230-0015 hours) at MitchellBay.
At both places there were minor spawningperiods in late June and mid-July: I observed amuch smaller number of nereid gastrulae orthree-segmented larvae in plankton samplesfrom Garrison Bay on 11 July 1969 and on 19July 1969 from Mitchell Bay. I also observed afew P. bicanaliculata males swimming as if inbreeding swarms on 24 June 1969 and 10 July1969 at Mitchell Bay. However, compared tothe enormous numbers of swarming adults seenon 31 July-1 August 1968 and 3 August 1969,and compared to the great number of gastrulaeseen in plankton samples on 4 August 1969, thenumber of adults that spawned earlier constituted only a small portion. Guberlet (1933) reported highly synchronous swarms of Plarynereis at Friday Harbor in June-August 1929~
1932; so this synchronous, massive swarmingbehavior appears to be predictable. That animals die soon after spawning is evidenced bythe fact that I could find very few large individuals of P. bicanaliculata at either study area by23 August; at both places tiny, 4-5 segmentindividuals were then abundant in algae. On 14July 1969, In contrast, there was an average of75 large individuals (it: 20 samples, standard

Plarynereis bicanaliculata and Nereis vexillosa,-RoE 343
error = 27.98) per square meter at MitchellBay. There is nearly a complete turnover of thepopulation every August, although a few adultsspawn (and die) in June and July and a fewhave not spawned by August.
Development and Behavior
Within 1 day eggs cultured in the laboratoryat 10° C are swimming gastrulae and within 2days have become trochophores. The trochophores have lipid droplets and larvae do notfeed until they become benthic. In 7 to 8 daysthey have three segments and more or less reston the bottom, although they do swim up intothe water column at the least disturbance.(Guberlet, 1933, reported the beginning ofthree segments in 80 hours.) In the laboratorythe small nereids started feeding on diatomsbefore 17 days, when they had four segments.By 1 month's time they had eight plus segments, were about 2 mm long, and had builtmucous and diatom houses. In nature (MitchellBay and Garrison Bay) they settled out of theplankton within 3 weeks at four segments orless. Most settled in young ulvoids, seeming toprefer algae to sand.
Counts were not made of the number of eggsproduced per female or the number survivingto trochophore, but there were averages of6021 (in 23 samples, standard error = 1968)and 7000 (in 11 samples, standard error =2969) small individuals of P. bicanaliculata perm2 at Mitchell Bay in September 1968 andOctober 1969, respectively, and averages of2750 (in 10 samples, standard error = 901) and3500 (in four samples, standard error = 2179)per m2 at Garrison Bay in October 1968 and1969 (Roe, unpublished data).
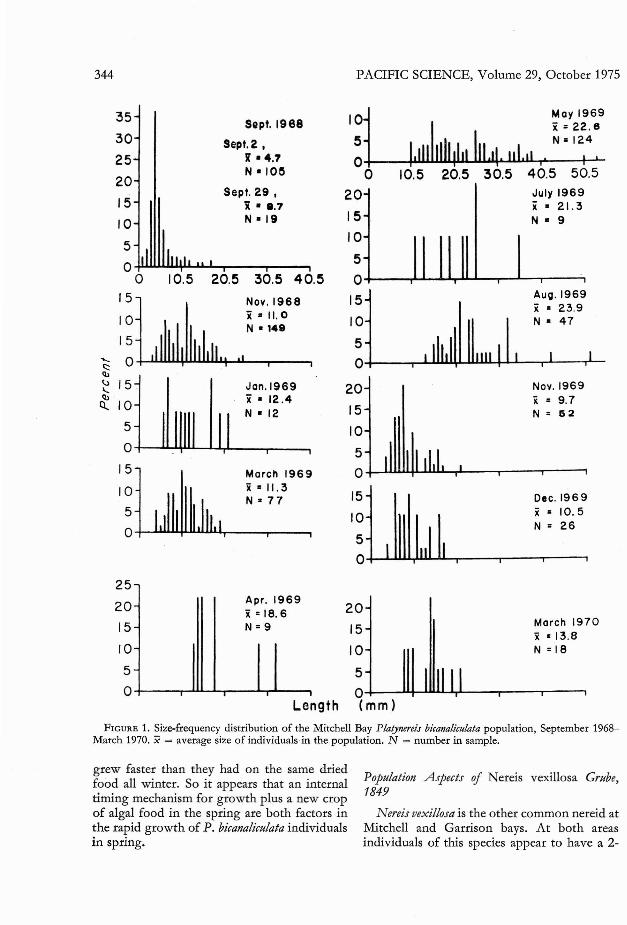
The size-frequency histogram (Figure 1) forMitchell Bay (see Roe 1971 for the similarGarrison Bay population) suggests two periodsof rapid growth by P. bicanaliculata: in AugustSeptember after settlement and in April-May.After settlement, young worms grow to alength of about 10 mm by the end of September. They stay an average of 10-12 mm fromthen until mid-March. In March new ulvoidsstart growing on the mud flats and P. bicanaliculata individuals again grow rapidly (see also thefollowing experiment). During summer the
nereids do not grow in length, but ratherdevelop gametes and become epitokous forspawning. Their diameters increase greatlyduring this time.
In April and May 1969, individuals of P.bicanaliculata were kept in four fingerbowls;five large (about 45 mm long) or 10 small (1520 mm long) worms per bowl. Four pieces ofulvoids, cut into 7.5 x 3 cm rectangles, wereadded as food to each bowl when it becameempty. The 10 larger worms ate the algae fasterthan did the 20 small ones, even though thelatter were twice as many. The five large nereidseating the thinner-bladed ulvoid ate all of it,except for what they used as houses, in anaverage of 4 days; the others ate somewhatslower. The small worms ate so slowly that thealgae grew considerably; so accurate measurements of the amount they ate were not made.
The 10 large worms grew from x 80 mgweight/worm to x130 mg/wormin 12 days (fiveanimals weighed together and total divided by5 each time showed five animals weighing79.3 mg and five weighing 81.3 mg on 2 May;five weighing 128.93 mg and five weighing130.79 mg on 14 May).
The rapid growth rate in nature in springwas first attributed to growth of young algae onthe mud flats. However, this is not the onlyreason for rapid growth by P. bicanaliculatathen. During the fall, winter, and spring of1969 and 1970, I kept some young individualsin the laboratory at 7° C, feeding them onEnteromorpha that I had collected at MitchellBay in October and then dried. The nereidsgrew hardly at all though winter. In April, Idivided these nereids into two groups, feedingone-half on young spring Enteromorpha and onehalf on the dried Enteromorpha. Nereids collected in the field in March that were the same size(average length = 1.5 cm) as those used in theexperiment were used as controls and were notfed. Some nereids fed on the young, fresh algadied after 7 weeks, but they had grown to anaverage length of 5 cm before they died. Theones fed on dried Enteromorpha grew faster thanthey had all winter (to an average length of 2.5cm in 11 weeks). The unfed controls did notgrow during the duration of the experiment.The individuals fed fresh algae grew faster thanthose fed dried food, but those fed dried food
344 PACIFIC SCIENCE, Volume 29, October 1975
Mayl969i =22, eN" 124
Dec. 1969x = 10.5N = 26
AuO·1969i = 23.9N = 47
Nay. 1969x = 9.7N = 6:2
105o+---foIo&.....o 10.5 20.5 30.5 40.5 50.5
20 July 1969It .. 21.3
15 N c 9
10
50+--.,..............1....Lr~---,r--L-.--.....,
1510
5O+--"""'T"""~Wo\JL...W.LLL.J-I-lI..--,.-L----,.L-
201510
5
o15
10
5O+-.L.LLLL,U.lL.LL.,--r---...,..----,
Nay. 1968i .. 11.0N .. 148
March 1969i • 11.3
N • 77
Jon.1969i .. 12.4N .. 12
Sept. 198e
Sept. 2 ,i .. 4.7N" IO~
Sept. 29,x ....7
N" 19
10.5 20.5 30.5 40.5
35
3025201510
5o4-WULLU+LJL..LL.L....,.__~-_,
o15
1015
o""~cu~ 15.
~ 105
o;---........':""t"'U--~L--.,...__-_,15
105
o
Apr. 1969x=18.6N=9
25
2015
1050+---.,.-...LLL-L....-----1~-___,
Length
201510
5O+-----J......-....(mm)
March 1970i =13.8N = 18
FIGURE 1. Size-frequency distribution of the Mitchell Bay Platyllereis bicallaliculata population, September 1968March 1970. x = average size of individuals in the population. N = number in sample.
grew faster than they had on the same driedfood all winter. So it appears that an internaltiming mechanism for growth plus a new cropof algal food in the spring are both factors inthe rapid growth of P. bicanaliculata individualsin spring.
Population Aspects of Nereis vexillosa Grube,1849
Nereis vexillosa is the other common nereid atMitchell and Garrison bays. At both areasindividuals of this species appear to have a 2-

Platynereis bicanaliculata and Nereis vexillosa-RoE
year life span. In May 1969, 36 nereids measuredfell into three major size classes, with eightbeing 3.5-5.5 cm in length, 16 being 6.5-9.5cm, and 13 being 10.0-13.0 cm long. Youngindividuals kept in the laboratory from April toNovember grew to 4.5 and 6 cm, respectively.These data indicate that individuals of thisspecies grow to about one-half adult size their1st year and to adult size their 2nd year, duringwhich spring or summer they become epitokous, spawn, and die. (They can, however,spawn at the end of their 1st year. See Johnson1943.)
Egg masses lying on the surface of the mudwere observed by Roe and Woodin (personalcommunication) between 10 March and 30 Julyat Mitchell Bay; egg masses were observed byRoe at Garrison Bay from late April through 23August. The breeding season is long andasynchronous in both areas.
Eggs are laid in jelly clumps (see Johnson1943 for picture), individual eggs being about250 It in diameter. Small worms hatch from theegg masses at the three-segment stage, about0.322 mm in length, after 3 to 4 days in thelaboratory. They swim rather than crawl untilthe four- to five-segment stage (about 1 weekin the laboratory). Within that week they maketubes from mucus and diatoms and then aremainly benthic. After 3.5 weeks they rangefrom 8 to 25 segments and 0.7 mm to 5.5 mmin length, with some individuals growing morerapidly than others.
Settlement, Tube Building, and Tube Defense
The following refers both to Platynereis bicanaliculata and to Nereis vexillosa, althoughmost reported observations were on Nereissince it is a more benthic species early in its lifeand, therefore, easier to observe; the same observations were made using P. bicanaliculataindividuals (see also Woodin 1974), althoughsuch observations on Platynereis were fewer innumber. All observations of tube building andterritorial behavior were made on individualsin 4.5-inch fingerbowls, usually with a dissecting microscope.
When small nereids settle out of the planktonthey build mucous tubes to live in. They staywithin the tubes, going partway out the front
345
end and biting off algae, grabbing diatoms, etc.,to add to the outside of the tube, anterior endfirst. Nereis vexillosa makes a tube within 1week of hatching from the egg mass, at thefour- to five-segment stage. Platynereis bicanaliCt/lata individuals start making tubes at four tofive segments also, but they do not make tubesthat completely cover them until after theyreach six segments in length. Earlier than sixsegments, they spend much time under anything available (clumps of diatoms, sand grains,etc.). The ratio of tube length to worm lengthin N. vexillosa averaged 1.65:1 (four measurements, range = 4:1 to 1.4:1); tubes of P.bicanaliculata were also longer than the animalsbut were not measured in this study.
Individuals defend their tubes against otherindividuals of the same species. Defense is of asmall territory-of a circle at both openings ofthe tube; i.e., another nereid can build a tubeoverlying the first and there will be no conflictif doorways do not meet. Few measurementswere obtained of the length of worm exposedat the entrance to a tube compared with thetotal length of the same worm, but nereidsmeasured tended to be positioned with aboutone-half of the body out of the tube (9: 19 mmand 13: 25 mm exposed in two measurements),although sometimes most of the worm wasexposed (22: 23 mm exposed, one measutement). The ratio of total tube length to lengthof animal exposed averaged 2.52: 1 in 10measurements (range = 6.4:1 to 1.7:1). As~
suming the exposed length of the nereid equalsthe diameter of the circle most often defendedat each end of the tube (and from observationthis does seem to be the case), then the diameterof this circle is about 2.5 times smaller than thelength of the tube and is about one-half thelength of the occupant.
In laboratory fingerbowls, nereid defense oftube and territory involves fighting (as de~
scribed for adult nereids in Reish and Alosi1968) and is important in keeping them; a wormthat loses a fight is displaced and has to win orbuild another tube. It was observed that largerindividuals almost always win fights and keeptheir tubes when smaller individuals withouttubes fight them (five of five battles won). Infights between worms ofequal sizes, individualsusually win and keep their tubes from intruders

346 PACIFIC SCIENCE, Volume 29, October 1975
.. .. -' ..-,~. ~
.. t •... f \ • "
~ t ;
... -
..,
• .t',I.
a
.. ....: -': "\ ': .") ...,"\ '" ..~ :.........
,,\.,. .. "..... , ~ .. " ..... \
, .. \ ...-.... ., , '" .- til'
~ _ .. " ........ ~ .... - lJ ~ '" or ~t • •__ t"
.'~"" ' ... .,.f • j ..' " ,\~)
d
scale: - = lern
FIGURE 2. Drawings from photographs of distribution of diatom and mucous tubes of young nereids in fingerbowls. Some tubes were built a short way up the sides of the dishes. a, Tubes of Nerds vexillosa, about 2 weeks afterhatching; b, tubes of a second group of Nerds vexillosa, about 3 weeks after hatching; c, tubes of Nereis vexillosa seenin a, about 1 month after hatching; d, tubes of 6-week-old Platynereis bicanaliculata.
(five of seven fights), but occasionally are displaced (two of seven fights). Small individualswith tubes win over larger worms without tubesabout one-half the time (small won five timesand lost five times in 10 episodes). Wins andlosses occur when fights are at the front or therear of a tube, If an intruder tries to displace anoccupant from the rear, the occupant oftenturns around in its tube and fights.
Nereis vexillosa starts making and defendingtubes even before the lipid food reserves of theegg are gone. It does not start to feed until later(at the five- to six-segment stage, about 10 days
after hatching). Before they are five segmentslong, individuals show increased activity butnot aggression when approached by others; butwithin 2 weeks of hatching (seven-eight segments), they have become highly territorial,with much fighting occurring between individuals in fingerbowls in the laboratory. Fromobservations of animals from the same eggmass, I saw that unequal gowth rates occuramong individuals; the fast growers made tubesfirst, usually defended them better, and grew atincreasingly rapid rates compared to slowerindividuals. The smaller worms become pro-

Plarynereis bicanaltculata and Nereis vexillosa-RoE
gressively worse off and, among Nereis vexillosa(but not observed with P. bicanaliculata), I haveseen larger worms actually eat smaller losers offights. This differential growth was also noticedby Johnson (1943) although he did not comment on its significance. High numbers ofNereis hatch from egg masses, but Nereis density is rather low (about 221m2 average atMitchell Bay [Roe 1971]). Spacing and highmortality may be due in part to negative intraspecific interactions. In laboratory fingerbowlsfrom 22 April to 1 June there was a strikinglyeven distribution of nereid tubes (Figure 2 a-c)and the number of tubes decreased from about350 to about 65 (Figure 2 a-c). Tubes alsobecame larger and spaced further apart duringthis time. By November, in three containersonly one (two jars) or two (one jar) large nereidsremained. The smaller ones presumably hadbeen eaten inasmuch as no carcasses were found.
Plarynereis bicanaliculata seems somewhat lessaggressive, with territorial fighting well developed by 20 segments or so. Even spacing oftubes was observed (Figure 2d), but individualswere not seen eating others and many worms ofthis species can be kept together in 4-ounce jarsfor months.
Once a tube is made, a nereid usually comesonly partway out to snatch a clump of algae anddrag it back into the tube to eat it; a worm willleave the tube (in the laboratory) only whenfood is too far away to reach otherwise. If anindividual is thus forced out of its tube, whenit returns it will enter any vacant tube, notnecessarily the one it left.
DISCUSSION AND CONCLUSIONS
The life histories of the common nereids ontwo mud flats in Washington are quite different: Plarynereis bicanaliculata, the smaller of thetwo, is an annual with highly synchronousspawning and planktonic trochophores. Nereisvexillosa has a 2-year life span, and members ofa population shed eggs from 1vfarch throughAugust. Eggs are in benthic jelly masses andthe young are predominately benthic uponhatching. However, behaviors of the two
347
nereids are remarkably similar. In both species,soon after the young become benthic they startbuilding tubes of mucus and any availablematerials (in this study, diatoms), and theydefend these tubes from intruders. This behaviorhas been studied in several species of adultnereids by Reish and Alosi (1968) and by Evans(1973) and is Widespread throughout thefamily.
Territorial behavior and defense of tubes alsooccurs in very young worms soon after theyhave become benthic. In one species (Nereisvexillosa) tube building and defense begins evenbefore the worms begin feeding. A behavioralpattern that begins so early and continuesthroughout the life ot an individual must beadaptive for the individual. Although the adaptive significance of territorial behavior and tubedefense is not known for nereids, several possi~bilities come to mind. Remaining in one's tubeas an antipredator strategy is possibly one majoradaptively significant aspect of this behavior.The nemertean Paranemertes peregrina ate anywhere from 14 to 35 percent of the nereid populations at the areas studied (Roe, unpublisheddata). Paranemertes peregrina appeared to takelonger to locate a nereid if the nereid remainedin its tube.
Maintaining a tube with algae on the outsidesurface also ensures a close food supply for anereid as the attached algae grows (Woodin1974). Woodin (1974) has shown that nereidsthus actually farm algae.
Woodin (1974) has also shown that Plarynereis bicanaliculata is competitively dominant toother polychaetes, such as Armandia brevis andAxiothella rubrocincta, and territorial behaviormay be involved. Further studies in natureshould be carried out to see if P. bicanaliculatauses territorial behavior against A. brevis as ameans of maintaining its competitive dominance for space. I did not study interspecificaggression, but Reish and Alosi (1968) reportedit among some nereid species, and Evans (1973)reported that interspecific aggression is strongerthan intraspecific in some nereid species. It iscertainly important in intraspecific competitionfor space, as this study with Nereis vexillosashows.

348
ACKNOWLEDGMENTS
I wish to thank Dr. Alan J. Kohn for muchhelp throughout this study. Dr. R. L. Fernaldprovided space and facilities at the FridayHarbor Laboratories. Dr. S. A. Woodin provided much discussion. I wish also to thankMr. and Mrs. Jack Pierce and Mr. and Mrs.R. C. Schwarz for letting me use their properties for study areas. Thanks go to C. Argo fortyping the manuscript and to V. Anderson fordrawing Figure 2 from photographs.
LITERATURE CITED
BERKELEY,E., andC. BERKELEY. 1948. Annelida,Polychaeta errantia. Can. Pac. Fauna 9b (1).Fish. Res. Board Can. 100 pp.
EVANS, S. M. 1973. A study of fighting reactions in some nereid polychaetes. Anim.Behav. 21: 138-146.
PACIFIC SCIENCE, Volume 29, October 1975
GUBERLET, J. E. 1933. Observations on thespawning and development of some Pacificannelids. Fifth Pac. Sci. Congr. Proc. 5:4213-4220.
HARTMAN, O. 1954. Australian Nereidae. Trans.R. Soc. South Aust. 77: 1-41.
JOHNSON, M. W. 1943. Studies on the life history of the marine annelid Nereis vexillosa.BioI. Bull. (Woods Hole, Mass.) 84: 106.,-114.
REISH, D. J., and M. C. ALOSI. 1968. Aggressivebehavior in the polychaetous annelid familyNereidae. Bull. South Calif. Acad. Sci. 67:21-28.
ROE, P. 1971. Life history and predator-preyinteractions of the nemertean Paranemertesperegrina Coe. Ph.D. Thesis. University ofWashington, Seattle. 129 pp.
WOODIN, S. A. 1974. Polychaete abundancepatterns in a marine soft-sediment environment: the importance of biological interactions. Ecol. Monogr. 44: 171-187.