applications of phenological models to predict the future carbon sequestration potential of boreal...
TRANSCRIPT

APPLICATIONS OF PHENOLOGICAL MODELS TO PREDICTTHE FUTURE CARBON SEQUESTRATION POTENTIAL
OF BOREAL FORESTS
ILKKA LEINONEN 1 and KOEN KRAMER 2
1Department of Botany & Microbiology, University of Oklahoma, Norman, OK 73019, U.S.A.E-mail: [email protected]
2Alterra, Department of Ecology and Environment, P.O. Box 47, 6700 AA Wageningen,The Netherlands
E-mail: [email protected]
Abstract. Changes in the duration of the photosynthetically active period strongly influence thechanges in the carbon sequestration potential of boreal forests under climatic warming. In this paper,current theories on the effects of environmental variables such as spring air and soil temperature, pho-toperiod and chilling temperatures on the timing and initiation of photosynthesis in boreal deciduousand coniferous trees are discussed. Different dynamic phenological modeling approaches are re-viewed, and model simulations are utilized to demonstrate model predictions under changing climaticconditions. A process-based forest ecosystem model is applied to estimate the relative importance ofthe duration of the photosynthetically active period on the amount of annual gross primary productionand net primary production of boreal coniferous forests. All applied modeling approaches predict anincreasing duration of the photosynthetically active period as a result of climatic warming. However,the magnitude of the response to increasing temperature varies between models and therefore affectsthe predictions of the changes in production.
1. Introduction
Phenology studies the seasonal and annual rhythms of biological phenomena inrelation to climatic factors (e.g., Häkkinen, 1999). In the case of forest trees, suchphenomena include the timing of bud burst, leaf senescence and leaf fall, or moregenerally the timing of the beginning and the end of the photosynthetically activeperiod. The timing of phenological events and the duration of the growing seasonstrongly influence the amount of annual photosynthetic production of boreal forests(e.g., Black et al., 2000; Myneni et al., 1997). A change in phenology may be oneof the most important factors affecting the future carbon sequestration potentialof boreal forests with rising global temperatures. The most significant changes inforest carbon sequestration are likely to occur in spring, when temperature stronglyregulates events such as the timing of bud burst and the onset of photosynthesis,and when a high level of incoming radiation is already available for photosyntheticproduction.
It may not be possible to make quantitative predictions on the effects of cli-matic change on the growth and carbon sequestration of forests without applying
Climatic Change 5: 99–113, 2002.© 2002 Kluwer Academic Publishers. Printed in the Netherlands.

100 ILKKA LEINONEN AND KOEN KRAMER
a modeling approach. Recently, process-based forest ecosystem models have beenutilized for this purpose (Mohren et al., 2000). However, since the model predic-tions are dependent on the original assumptions built into the models, it is necessaryto carefully evaluate the reliability of the model behavior. This holds for all differ-ent sub-processes included in the models, and one of the most important of these,at least in the case of modeling boreal forest ecosystems, is tree phenology.
The aim of this paper is to summarize the current knowledge on the environ-mental regulation of the phenological events in boreal forest trees, concentratingespecially on the phenomena determining the timing of the beginning of the photo-synthetically active period in spring. An overview of existing phenological modelsand their applications in predicting changes in the carbon sequestration potentialof boreal forests under changing climatic conditions is presented and demonstratedthrough model simulations.
2. Modeling Approaches
2.1. TIMING OF BUD BURST
Spring temperature is the most important single factor affecting the timing ofbud burst of trees. The simplest possible approach to describe the relationshipbetween these two factors is to apply empirical relationships between observedbud burst dates and standard meteorological variables such as monthly mean tem-peratures (e.g., Maak and von Storch, 1997). However, in cases where more precisemeteorological information is available, such as daily mean, minimum and max-imum temperatures, it is likely that utilization of more detailed, usually dynamicmodeling approaches gives more accurate predictions.
Dynamic modeling of the timing of bud burst is based on the fact that the rateof spring bud development is dependent on air temperature. Usually the modelsinclude a certain threshold temperature, below which the rate of development iszero, while above this threshold a quantitative temperature response (either linearor non-linear) is assumed. The state of development at each instant is determinedby integrating the rate of development over time, and the bud burst occurs when acertain critical value of the developmental state is attained.
The oldest and probably most widely used approach for modeling the timingof phenological events, the temperature sum or thermal time method, is analogousto the dynamic modeling approach described above. In this method, accumulateddaily or hourly mean temperatures above a certain threshold are used as an in-dependent variable to predict, for example, the occurrence of bud burst of trees.In this case, the momentary temperature can be seen to be directly related to therate of bud development, whilst the accumulated temperature sum represents thedevelopmental state of the bud.
According to the traditional temperature sum approach, the rate of bud devel-opment is assumed to be linearly dependent on temperature. However, different

PHENOLOGY AND CARBON SEQUESTRATION OF BOREAL FORESTS 101
response functions have been presented. Sarvas (1972) measured the developmen-tal rate at different constant temperatures and found a logistic temperature responsefunction, which was applicable for several tree species. In later dynamic bud burstmodels, this response has been sometimes used to replace the linear temperaturesum function (Hänninen, 1990; Häkkinen et al., 1998).
In addition to the important role of spring temperatures, other driving variableshave also been included in the phenological models. The occurrence of dormancybreaking chilling temperatures has been advocated as having an important rolein the bud development of several tree species. It has been proven that a suffi-ciently long exposure to low temperatures is required before buds can respond tohigh spring temperatures which induce the development leading to bud burst. Indynamic phenological modeling, there are several alternatives used to take thisphenomenon into account. One approach is based on conceptual modeling princi-ples presented by Sarvas (1974). According to his theory, the annual cycle of treesis divided into subsequent dormant and active periods. The dormant period consistsof two separate parts, called autumn dormancy and winter dormancy. During theautumn dormancy (biochemical) changes in the bud are driven by low chillingtemperatures, the efficiency of which is modeled by using a bell-shaped responsefunction. During the winter dormancy, the temperature response is assumed tofollow a logistic function, where high temperatures are the most effective. Afterthe completion of winter dormancy, the active period is assumed to begin. Duringthis period, the ontogenetic development, leading to bud burst, occurs at the ratedetermined by a logistic temperature response.
The Sarvas approach has been widely utilized in recent phenological modeling(e.g., Hänninen, 1990; Kramer, 1994; Chuine et al., 1998). However, the later mod-eling approaches have not taken into account the dissimilarity between the winterdormancy and the active period. Instead, the ontogenetic development towardsbud burst has been assumed to begin as soon as the autumn dormancy has beencompleted, i.e., the chilling requirement of the bud has been attained.
An alternative approach to model the chilling effect was presented by Cannelland Smith (1983). In their model, the accumulated chilling, i.e., number of hourswhen temperature is below a certain threshold, was assumed to reduce the temper-ature sum required for bud burst according to a decreasing exponential function.This approach has been found to be applicable especially for some temperate treespecies that have a high chilling requirement for dormancy release (e.g., Murrayet al., 1989). The original model by Cannell and Smith (1983) was modified byHänninen (1990) according to the dynamic modeling approach. He assumed thatthe quantitative relationship between chilling and high spring temperatures can beinterpreted in a way that increased chilling gradually increases the competence ofbud to respond to temperatures which drive the ontogenetic development towardsthe bud burst.
The photoperiod controls the timing of phenological events of various borealtree species. The role of photoperiod is pronounced in events such as growth ces-

102 ILKKA LEINONEN AND KOEN KRAMER
sation and bud set in autumn (e.g., Heide, 1974; Koski and Sievänen, 1985). In thecase of the timing of bud burst in spring, the effect of photoperiod is, however,less understood, although some evidence indicate that photoperidic response isinvolved also in this event (e.g., Heide, 1993). The photoperiodic effect can beinterpreted to be included indirectly in some models predicting bud burst. Sometemperature-based models include a fixed calendar date (i.e., threshold day length)as a starting point of the accumulation of spring temperature sum (Häkkinen etal., 1998; Wielgolaski, 1999). A good performance of such models may be due tothe physiological effect of photoperiod on spring bud development, and thereforesimple phenological models including a calendar day as a parameter may havequite realistic bases.
2.2. SEASONAL PHOTOSYNTHETIC CAPACITY IN CONIFERS
The photosynthetic capacity of boreal evergreen conifers is strongly limited duringwinter, mainly due to the reduced photochemical efficiency (e.g., Öquist, 1983;Ottander and Öquist, 1991; Bauer et al., 1994). The photochemical down regula-tion of photosynthesis has been seen at least partly to be caused by acclimationagainst unfavorable winter conditions by protecting the photosynthetic machineryfrom damage caused by the combination of low temperatures and light energy(e.g., Gillies and Vidaver, 1990; Huner et al., 1993). The timing and rate ofspring recovery of photosynthesis strongly influences the amount of annual carbonsequestration of trees. It has been generally observed that the recovery of photo-synhetic capacity from a negligible winter level to its summertime maximum is agradual process which may take several weeks (Pharis et al., 1970; Öquist, 1983;Ottander and Öquist, 1991). Furthermore, this recovery is known to be stronglydependent on air temperature (e.g., Lundmark et al., 1988).
Dynamic modeling approaches have been utilized to predict the winter recoveryand autumn cessation of photosynthetic capacity of boreal conifers. Pelkonen andHari (1980) developed a model for the seasonal photosynthetic activity of Scotspine. In their model, the rate of change (increase or decrease) of the photosyntheticcapacity depended on air temperature and on the prevailing stage of development.According to the model, warming spring temperatures caused a gradual recovery ofphotosynthesis from winter depression, while the occurrence of low temperaturescaused a drawback of this development. Later comparisons with empirical datahave proven the model to be successful in predicting the timing of the beginning ofthe carbon uptake period of Scots pine stands in spring (e.g., Mohren et al., 2000).
Bergh et al. (1998) modeled the seasonality of photosynthetic capacity of bo-real Norway spruce. The main principles of their model were basically similar tothe model by Pelkonen and Hari (1980). The spring recovery of photosynthesiswas modeled according to the temperature sum approach, and frost temperatureswere assumed to reduce or reverse this development. In addition to the air tem-perature, soil temperature was also included in their model, and the phosynthetic

PHENOLOGY AND CARBON SEQUESTRATION OF BOREAL FORESTS 103
recovery was assumed to be hastened after soil thawing in spring. Bergh et al.(1998) also used their model to demonstrate the importance of the seasonal changesof photosynthetic capacity for the annual photosynthetic production of Norwayspruce.
3. Simulation Examples
3.1. METHOD
In the following, the application of phenological modeling in predicting the effectsof climatic warming on the carbon sequestration of boreal forests is demonstrated.In the simulations, climatic scenario data, based on the output of HadCM2 climatemodel (Mitchell et al., 1995) and generated for southern boreal conditions (Erhardet al., 2001), were used. The data covered the time period from 1950 to 2100and predicted an increase in annual average temperature by approximately 2 ◦Cduring this period. Four phenological models were utilized to predict the annualduration of the photosynthetically active period in deciduous trees and conifers.The effect of phenology on annual gross primary production (GPP) and net primaryproduction (NPP) of a coniferous stand under the conditions of climatic warmingwas quantified with the aid of a process based forest growth model. Only the tem-perature effect was taken into account in the simulations, i.e., the direct effect ofincreased atmospheric CO2 concentration was excluded.
Three models, developed and parameterized for predicting the timing of budburst in birch, were used to predict the beginning of the photosynthetically ac-tive period in deciduous trees. First, in the model developed and parametrized byHäkkinen et al. (1998), the rate of spring bud development was dependent on airtemperature according to a logistic function (Sarvas, 1972). The model includestwo parameters, namely the critical developmental stage when the bud burst occurs,and the calendar day when buds begin to respond to warm temperatures, both ofwhich were estimated from long term phenological time series in Southern Fin-land (Häkkinen et al., 1998). Second, the model by Hänninen (1990) included theSarvas’ (1972, 1974) principles of sequential periods of dormancy and ontogeneticdevelopment. The parameters for the critical developmental stages for the com-pletion of dormancy and the bud burst were estimated by Häkkinen et al., (1998).Third, the model by Cannell and Smith (1983) assumed the simultaneous effectof chilling and high temperatures. The parameters for the exponential function forthe critical temperature sum were estimated by Murray et al. (1989) for birch inScotland.
Seasonal changes in the photosynthetic capacity of conifers were predicted by atemperature-based model, developed for Scots pine (Pelkonen and Hari, 1980, seeabove). The model predicts the alternation of the photosynthetic capacity betweenwintertime zero-level and maximum summertime level, and has been applied forboth the spring recovery and the autumn cessation.

104 ILKKA LEINONEN AND KOEN KRAMER
In order to quantify the effects of annual variation in weather conditions andclimatic warming in both deciduous trees and conifers, the amount of annual radi-ation available for photosynthetic production was calculated. In connection of theresults of this study, the concept of ‘available radiation’ indicates the total incomingshort-wave solar radiation on the top of the canopy during the period when treesare photosynthetically active. The active photosynthetic period was determined,in the case of deciduous trees, to begin at the time of bud burst, predicted by themodel by Häkkinen et al. (1998) and end at a fixed calendar day, i.e., only thebeginning of this period was affected by climatic conditions. In the case of conifers,the photosynthetic period included all days when the photosynthetic capacity wasabove the level of 50% from the maximum summertime level.
Variations in annual GPP and NPP in a Scots pine stand were predicted bya process-based forest growth model FORGRO (Mohren, 1987, 1994; Kramer,1995). The FORGRO model contains detailed descriptions for light intercep-tion, photosynthesis, respiration and allocation. To describe light interception ina canopy with a mixture of species, the leaf areas, weighted by the extinctioncoefficients, are summed over the species (Kropff and Van Laar, 1993). Canopyphotosynthesis is calculated by integration over five shaded and sunlit leaf layers,assuming a uniform distribution of the leaf area over the canopy height. Moredetails of this approach can be found in Goudriaan and Unsworth (1990), andGoudriaan and Van Laar (1994). Leaf photosynthesis depends on light, CO2 andtemperature according to the biochemical model developed by Farquhar and VonCaemmerer (1982). Respiration of living biomass depends on its biochemical com-position (Penning de Vries et al., 1974), and is also dependent on temperatureaccording to an exponential function. Allocation of assimilates to the differentorgans is done daily, based on empirical allocation keys with the exception ofallocation to the foliage and the reserve pool for which saturation curves are used(Kramer, 1995).
The simulations consisted of three subsequent model runs, each covering a 50-year time period and applying the same initial stand conditions (20-year-old Scotspine stand). The seasonal changes in photosynthetic capacity were included in thesimulations by using the model by Pelkonen and Hari (1980) as a sub-model.
3.2. RESULTS
The timing of bud burst in birch, as predicted by three models under the scenarioclimate, is shown in Figure 1. There is a clear trend of hastened bud burst as a resultof climatic warming. Two of the models (Häkkinen et al., 1998; Cannell and Smith,1983) show a very similar response of the timing of bud burst to climate. The thirdmodel (Hänninen, 1990) shows a response similar to the other two models at thebeginning of the simulation period when no climatic warming has occurred, butpredicts systematically earlier bud burst in a warmer climate.

PHENOLOGY AND CARBON SEQUESTRATION OF BOREAL FORESTS 105
Figure 1. Predictions of the timing of bud burst of birch by three phenological models under thescenario climate.
Since the beginning of the photosynthetically active period in conifers is agradual process, no single event such as the timing of bud burst in the case ofdeciduous trees can be determined. Instead, in Figure 2, the seasonal course ofphotosynthesis, as predicted by the FORGRO model, is shown for two yearsdiffering strongly in temperature conditions (Figure 2a). The results show anapproximate one month difference in the timing of the maximum level of photo-synthesis in spring (Figure 2b). Despite the differences in summertime temperature,the maximum photosynthesis remains at a similar level during both years.
The annual amount of radiation available for photosynthetic production, aspredicted by the two phenological models in the scenario climate, is presentedin Figure 3. For comparison, the results are also shown for the case where a fixedcalendar day is used for both the beginning and end of the photosynthetically activeperiod. In this case, the date for bud burst is determined to be the mean date of thepredictions of the birch model from years without climatic warming (1951–2000).In cases where phenological models were applied, both strong annual variation anda clear increasing long-term trend in the amount of available radiation were evident.The observed trend was solely determined by the effect of rising temperature, sinceno trend in the amount of total annual radiation is included in the scenario.
The results of two models shown in Figure 3 are not directly comparable, sincedifferent mechanisms determine the photosynthetically active period in evergreenconifers and deciduous trees. Despite that, the results demonstrate the relative dif-ference in the temperature effect in models developed for different tree species. Thepine model predicts a stronger increase in radiation under changing climatic con-ditions compared to the birch model. This is caused by differences in the assumedenvironmental responses included in the models. In the pine model, the photo-synthetic capacity responds relatively quickly to rising temperatures in spring,while in the birch model, a longer accumulation of temperature sum is required
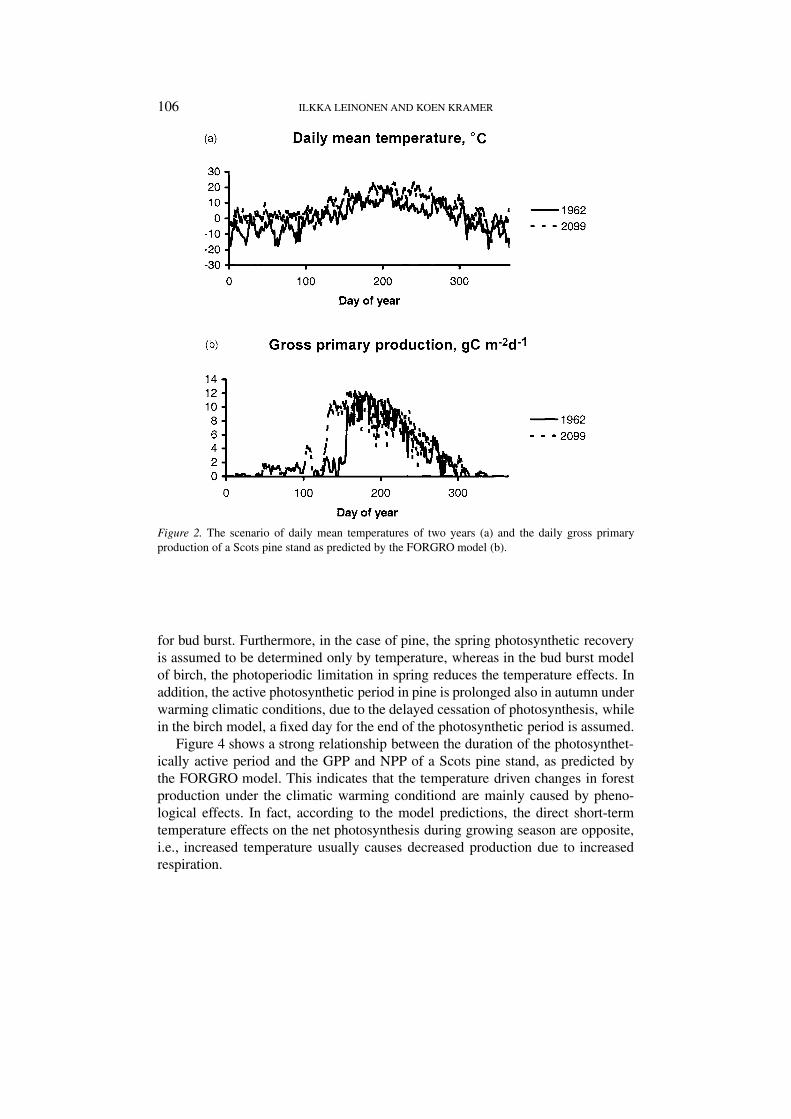
106 ILKKA LEINONEN AND KOEN KRAMER
Figure 2. The scenario of daily mean temperatures of two years (a) and the daily gross primaryproduction of a Scots pine stand as predicted by the FORGRO model (b).
for bud burst. Furthermore, in the case of pine, the spring photosynthetic recoveryis assumed to be determined only by temperature, whereas in the bud burst modelof birch, the photoperiodic limitation in spring reduces the temperature effects. Inaddition, the active photosynthetic period in pine is prolonged also in autumn underwarming climatic conditions, due to the delayed cessation of photosynthesis, whilein the birch model, a fixed day for the end of the photosynthetic period is assumed.
Figure 4 shows a strong relationship between the duration of the photosynthet-ically active period and the GPP and NPP of a Scots pine stand, as predicted bythe FORGRO model. This indicates that the temperature driven changes in forestproduction under the climatic warming conditiond are mainly caused by pheno-logical effects. In fact, according to the model predictions, the direct short-termtemperature effects on the net photosynthesis during growing season are opposite,i.e., increased temperature usually causes decreased production due to increasedrespiration.

PHENOLOGY AND CARBON SEQUESTRATION OF BOREAL FORESTS 107
Figure 3. Model-predicted annual available radiation for photosynthetic production under the sce-nario climate as affected by the timing of bud burst in deciduous trees (Birch model), beginning andcessation of the photosynthetically active period in conifers (Pine model) and constant duration ofthe photosynthetically active period (fixed days).
Figure 4. The relationship between the annual GPP and NPP of a Scots pine stand, and the avail-able radiation during the photosynthetically active period. Results from three simulation runs of theFORGRO model.

108 ILKKA LEINONEN AND KOEN KRAMER
4. Discussion
The results of this study show that when changes in the photosynthetic productionof boreal forests under future conditions are predicted, the results are strongly de-pendent on the predictions of the duration of the photosynthetically active period,which is consecutively determined by the timing of phenological events. Despitethe different mechanisms behind seasonal changes in the photosynthetic capacityin evergreen conifers and deciduous trees, the timing of these changes is equallyimportant. Therefore, it can be concluded that the correct behavior of the appliedphenological model is necessary in order to obtain reliable estimates of the overallchanges in carbon sequestration of boreal forests under climatic change.
The first step in applying ecological models for predictive purposes is to ensuretheir realism. This means that models must include the actual driving variablesaffecting the modeled phenomenon and the response to these variables must becorrectly formulated (e.g., Hänninen, 1995a). As described above, several model-ing approaches are available to predict the spring phenology of boreal trees and alsoto estimate the possible changes, caused by the climatic warming, in the duration ofgrowing season and carbon sequestration potential of boreal forests. Unfortunately,different models are based on partly conflicting theories of the environmental reg-ulation of tree development, and therefore their predictions may also be partlycontrasting. In this study, the results of three different models, predicting the timingof bud burst under changing climatic conditions, are presented. Although the long-term trend was similar in all models, they differed in the magnitude of the responseof model output to climatic change. Due to such variation, in order to ensure thereliability of the model predictions, it is essential to carry out tests where the resultsof the models are compared with observed data. In the case of the timing of budburst, such tests have been carried out for both boreal deciduous and coniferoustree species. Data for the tests have been obtained from both long phenologicaltime series and from experiments carried out under natural or semi-controlled con-ditions. One of the most important questions in the model testing concerned therole of chilling to determine the timing of bud burst of trees. Although differenttree species were included in these studies, the results seem to be surprisinglyconsistent.
Hänninen (1995b) compared different phenological models in predicting thetiming of bud burst in Scots pine at an open-top chamber experiment in easternFinland, where the winter and spring air temperature was highly manipulated. Theoutcome of his study was that the models including chilling as an independentvariable were less precise compared to models that included the effect of highspring temperatures only. Hannerz (1999) used Norway spruce clones to assess thetiming of spring bud burst at different sites in Sweden over several years. He foundthat the best model predicting the timing of bud burst was a simple thermal timemodel based on spring temperatures. Also, the model performance could not beimproved by including the effect of chilling. Häkkinen et al. (1998) used long-term

PHENOLOGY AND CARBON SEQUESTRATION OF BOREAL FORESTS 109
phenological time series from central Finland to test different models to predictthe timing of bud burst in birch. Also their results showed that more accurateand realistic predictions could be achieved by using a model including only thespring temperatures occurring after a fixed starting day, compared to a model whichincludes the chilling effect.
The results of earlier studies described above show generally that the use ofthe occurrence of chilling temperatures as an additional model variable will notimprove the behavior of models predicting the timing of spring bud burst underboreal conditions. These results are opposite to those obtained for some tree speciesgrowing in the temperate climate zone (for review, see Linkosalo, 2000). Theremay be two main explanations for this disagreement. First, with a boreal climate,winter temperatures are so low that the chilling requirement will always be fulfilled.This situation will probably remain the same under conditions of climatic warming(Myking and Heide, 1995). Second, with northern tree species and provenances,the chilling requirement has been observed to be lower compared to the southernones (e.g., Leinonen, 1996), i.e., relatively short exposure to low temperature issufficient to release bud dormancy.
In the present study, the model by Hänninen (1990), assuming the sequential pe-riods of dormancy and ontogenetic development, predicted the strongest responseof the timing of bud burst to climatic warming. This is due to the fact that in thismodel, in addition to the increasing temperature in spring, warm temperatures inautumn, after sufficient chilling exposure and the completion of dormancy, havean effect on ontogenetic bud development. On the contrary, in two other models(Häkkinen et al., 1998; Cannell and Smith, 1983), no ontogenetic developmentoccurs in autumn, but only after a certain calendar day (or day length) has beenattained. Despite some principal differences between these two models and thefact that they have been parametrized for birch origins representing different re-gions, their predictions are very consistent. These models differ in the assumptionconcerning the effect of chilling on the timing of bud burst, which is not taken intoaccount in the model by Häkkinen et al. (1998). However, at the boreal climate, theimportance of the chilling effect mostly disappears also in the case of the modelby Cannell and Smith (1983). Sufficient amount of chilling days occurs every year,and therefore in both models the main factor determining the timing of bud burstis the occurrence of warm temperatures in spring.
Compared to the bud burst phenology, fewer results are available concerningthe modeling of the seasonal changes of the photosynthetic capacity in borealconifers. The springtime recovery of photosynthesis has been modeled for bothScots pine (Pelkonen and Hari, 1980) and Norway spruce (Bergh et al., 1998), andthe main principles of these models are relatively consistent and are also supportedby empirical data. In the case of both tree species, the dynamics of the springtimephotosynthetic capacity has been found to be a gradual process, the rate of whichis dependent on air temperature. Furthermore, the occurrence of low temperaturesduring the recovery period may cause a drawback in this development. Due to

110 ILKKA LEINONEN AND KOEN KRAMER
between-year variation in weather conditions, the timing and rate of photosyntheticrecovery in conifers may vary strongly. Such variation has been observed in thecarbon exchange measurement both at the shoot level (Pelkonen and Hari, 1980;Wallin et al., 2000) and at the ecosystem level (Lindroth et al., 1998).
In addition to the effect of low air temperatures on reduction of the photosyn-thetic capacity of conifers, the freezing of soil may limit photosynthesis through thereduced availability of water. This effect can also be included in models predictingthe photosynthetic recovery (Bergh et al., 1998), but alone it may not be a sufficientvariable to explain completely the recovery process. The effects of both soil andair temperature on photosynthetic recovery in Norway spruce were studied exper-imentally by Bergh and Linder (1999). It was found that that the timing of springsoil warming had some effect on the timing of recovery, but mainly this processwas controlled by air temperature.
In conclusion, current knowledge of the principles of environmental regula-tion of the seasonalitity of photosynthetic production in boreal coniferous anddeciduous forest trees makes it possible to develop realistic models to predictthe changes of the carbon sequestration of boreal forests under changing climaticconditions. However, quantifying these processes requires further research sincetheir environmental responses are likely to be highly variable between tree species.Furthermore, the environmental regulation of phenological events appears to varystrongly within a single species (e.g., Beuker, 1994; Hannerz, 1994). For exam-ple, in tree species with a broad range of distribution, adaptation to local climaticconditions has caused differentiation between geographical origins. Therefore, itis likely that within-species genetic differences should be taken into account inorder to make reliable predictions on the effects climate change on boreal forests(Oleksyn et al., 1998).
Acknowledgements
This study was funded by the Academy of Finland (contracts 69806 and 2770)and by the EU Environment and Climate Research Programme (ENV4-CT97-0577). The climate scenario data were provided by M. Erhard, Potsdam Institutfür Klimafolgenforschung, Potsdam, Germany.
References
Bauer, H., Nagele, M., Comploj, M., Galler, V., Mair, M., and Unterpertinger, E.: 1994, ‘Photosyn-thesis in Cold Acclimated Leaves of Plants with Various Degrees of Freezing Tolerance’, Physiol.Plant. 91, 403–412.
Bergh, J. and Linder, S.: 1999, ‘Effects of Soil Warming during Spring on Photosynthetic Recoveryin Boreal Norway Spruce Stands’, Global Change Biol. 5, 245–253.
Bergh, J., McMurtrie, R. E., and Linder, S.: 1998, ‘Climatic Factors Controlling the Productivity ofNorway Spruce: A Model-Based Analysis’, Forest Ecol. Manage. 110, 127–139.

PHENOLOGY AND CARBON SEQUESTRATION OF BOREAL FORESTS 111
Beuker, E.: 1994, ‘Adaptation to Climatic Changes of the Timing of Bud Burst in Populations ofPinus Sylvestris L. and Picea Abies (L.) Karst’, Tree Physiol. 14, 961–970.
Black, T. A., Chen, W. J., Barr, A. G., Arain, M. A., Chen, Z., Hogg, E. H., and Nesic, Z.: 2000,‘Impact of Spring Temperature on Carbon Sequestration by a Boreal Aspen Forest’, in TheRole of Boreal Forests and Forestry in the Global Carbon Budget, May 8–12, 2000, Edmonton,Alberta, Canada, Abstracts, p. 99.
Cannell, M. G. R. and Smith, R. I.: 1983, ‘Thermal Time, Chill Days and Prediction of Budburst inPicea Sitchensis’, J. Appl. Ecol. 20, 951–963.
Chuine, I., Cour, P., and Rousseau, D. D.: 1998, ‘Fitting Models Predicting Dates of Flowering ofTemperate-Zone Trees Using Simulated Annealing’, Plant Cell Environ. 21, 455–466.
Erhard, M., Lindner, M., and Cramer, W.: 2001, ‘Climate Data’, in Mohren, G. M. J. and Kramer, K.(eds.), Long-Term Effects of Climate Change on Carbon Budgets of Forests in Europe, Alterra-Report 194, Alterra, Wageningen, The Netherlands, pp. 151–164.
Farquhar, G. D. and von Caemmerer, S.: 1982, ‘Modelling of Photosynthetic Response to Envi-ronmental Conditions’, in Lange, O. L., Nobel, P. S., Osmond, C. B., and Ziegler, H. (eds.),Physiological Plant Ecology II: Water Relations and Carbon Assimilation, 12B, Springer-Verlag,Germany, pp. 549–587.
Gillies, S. L. and Vidaver, W.: 1990, ‘Resistance to Photodamage in Evergreen Conifers’, Physiol.Plant. 80, 148–153.
Goudriaan, J. and Unsworth, M. H.: 1990, ‘Implications of Increasing Carbon Dioxide and ClimateChange for Agricultural Productivity and Water Resources’, in Impact of Carbon Dioxide, TraceGases, and Climate Change on Global Agriculture, ASA Special Publication 53, pp. 111–130.
Goudriaan, J. and Van Laar, H. H.: 1994, Modelling Potential Crop Growth Processes, KluwerAcademic Publishers, The Netherlands, p. 238.
Häkkinen, R.: 1999, Analysis of Bud-Development Theories Based on Long-Term Phenological andAir Temperature Time Series: Application to Betula sp. Leaves, Finnish Forest Research Institute,Research Papers 754.
Häkkinen, R., Linkosalo, T., and Hari, P.: 1998, ‘Effects of Dormancy and Environmental Factors onTiming of Bud Burst in Betula Pendula’, Tree Physiol. 18, 707–712.
Hannerz, M.: 1994, ‘Predicting the Risk of Frost Occurrence after Budburst of Norway Spruce inSweden’, Silva Fenn. 28, 243–249.
Hannerz, M.: 1999, ‘Evaluation of Temperature Models for Predicting Bud Burst in Norway Spruce’,Can. J. Forest. Res. 29, 9–19.
Hänninen, H.: 1990, ‘Modelling Bud Dormancy Release in Trees from Cool and Temperate Regions’,Acta For. Fenn. 213, 1–47.
Hänninen, H.: 1995a, ‘Assessing Ecological Implications of Climatic Change: Can We Rely on OurSimulation Models?’, Clim. Change 31, 1–4.
Hänninen, H.: 1995b, ‘Effects of Climatic Change on Trees from Cool and Temperate Regions: AnEcophysiological Approach to Modelling of Bud Burst Phenology’, Can. J. Bot. 73, 183–199.
Heide, O. M.: 1974, ‘Growth and Dormancy in Norway Spruce (Picea Abies). I. Interaction ofPhotoperiod and Temperature’, Physiol. Plant. 30, 1–12.
Heide, O. M.: 1993, ‘Daylength and Thermal Time Responses of Budburst during Dormancy Releasein Some Northern Deciduous Trees’, Physiol. Plant. 88, 531–540.
Huner, N. P. A., Öquist, G., Hurry, V. M., Krol, M., Falk, S., and Griffith, M.: 1993, ‘Photosynthesis,Photoinhibition and Low Temperature Acclimation in Cold Tolerant Plants’, Photosynth. Res.37, 19–39.
Koski, V. and Sievänen, R.: 1985, ‘Timing of Growth Cessation in Relation to the Variations in theGrowing Season’, in Tigerstedt, P. M. A., Puttonen, P., and Koski, V. (eds.), Crop Physiology ofForest Trees, Helsinki University Press, Helsinki, pp. 167–193.
Kramer, K.: 1994, ‘Selecting a Model to Predict the Onset of Growth of Fagus Sylvatica’, J. Appl.Ecol. 31, 172–181.

112 ILKKA LEINONEN AND KOEN KRAMER
Kramer, K.: 1995, ‘Modelling Comparison to Evaluate the Importance of Phenology for the Effectsof Climate Change on Growth of Temperate-Zone Deciduous Trees’, Clim. Res. 5, 119–130.
Kropf, M. J. and Van Laar, H. H.: 1993, Modelling Crop-Weed Interactions’, Cab International,Oxford, U.K., p. 274.
Leinonen, I.: 1996, ‘Dependence of Dormancy Release on Temperature in Different Origins of PinusSylvestris and Betula Pendula Seedlings’, Scand. J. For. Res. 11, 122–128.
Lindroth, A., Grelle, A., and Morén, A-S.: 1998, ‘Long-Term Measurements of Boreal Forest CarbonBalance Reveal Large Temperature Sensitivity’, Global Change Biol. 4, 443–450.
Linkosalo, T.: 2000, Analyses of the Spring Phenology of Boreal Trees and its Response to ClimateChange, University of Helsinki Department of Forest Ecology Publications 22.
Lundmark, T., Hällgren, J-E., and Hedén, J.: 1988, ‘Recovery from Winter Depression of Photosyn-thesis in Pine and Spruce’, Trees 2, 110–114.
Maak, K. and von Storch, H.: 1997, ‘Statistical Downscaling of Monthly Mean Air Temperature tothe Beginning of Flowering of Galanthus Nivalis L. in Northern Germany’, Int. J. Biometeorol.41, 5–12.
Mitchell, J. F. B., Johns, T. C., Gregory, J. M., and Tett, S. F. B.: 1995, ‘Climate Response toIncreasing Levels of Greenhouse Gases and Sulphate Aerosols’, Nature 376, 501–504.
Mohren, G. M. J.: 1987, Simulation of Forest Growth, Applied to Douglas Fir Stands in TheNetherlands, Thesis, Agricultural University Wageningen, The Netherlands, p. 184.
Mohren, G. M. J.: 1994, ‘Modelling Norway Spruce Growth in Relation to Site Conditions andAtmospheric CO2’, in Veroustraete, F. and Ceulemans, R. (eds.), Vegetation, Modelling andClimate Change Effects, SPB Academic Publishing, The Hague, The Netherlands, pp. 7–22.
Mohren, G. M. J., Bartelink, H. H., Kramer, K., Magnani, F., Sabaté, S., and Loustau, D.: 2000,‘Modelling Long-Term Effects of CO2 Increase and Climate Change on European Forests, withEmphasis on Ecosystem Carbon Budgets’, in Ceulemans, R. J. M., Veroustraete, F., Gond, V., andVan Rensbergen, J. B. H. F. (eds.), Forest Ecosystem Modelling, Upscaling and Remote Sensing,SPB Academic Publishing bv, The Hague, The Netherlands. pp. 179–192.
Murray, M. B., Cannell, M. G. R., and Smith, R. I.: 1989, ‘Date of Budburst of Fifteen Tree Speciesin Britain Following Climatic Warming’, J. Appl. Ecol. 26, 693–700.
Myking, T. and Heide, O. M.: 1995, ‘Dormancy Release and Chilling Requirement of Buds ofLatitudinal Ecotypes of Betula Pendula and B. Pubescens’, Tree Physiol. 15, 697–704.
Myneni, R. B., Keeling, C. D., Tucker, C. J., Asrar, G., and Nemani, R. R.: 1997, ‘Increased PlantGrowth in the Northern High Latitudes from 1981 to 1991’, Nature 386, 698–702.
Oleksyn, J., Tjoelker, M. G., and Reich, P. B.: 1998, ‘Adaptation to Changing Environment in ScotsPine Populations across a Latitudinal Gradient’, Silva Fennica 32, 129–140.
Öquist, G.: 1983, ‘Effects of Low Temperature on Photosynthesis’, Plant Cell Environ. 6, 281–300.Ottander, C. and Öquist, G.: 1991, ‘Recovery of Photosynthesis in Winter-Stressed Scots Pine’, Plant
Cell Environ. 14, 345–349.Pelkonen, P. and Hari, P.: 1980, ‘The Dependence of the Springtime Recovery of CO2 Uptake in
Scots Pine on Temperature and Internal Factors’, Flora 169, 398–404.Penning de Vries, F. W. T., Brunsting, A., and Van Laar, H. H.: 1974, ‘Products, Requirements and
Efficiency of Biosynthesis; a Quantitative Approach’, J. Theor. Biol. 45, 339–377Pharis, R. P., Hellmers, H., and Schuurmans, E.: 1970, ‘Effects of Subfreezing Temperatures on Pho-
tosynthesis of Evergreen Conifers under Controlled Environment Conditions’, Photosynthetica4, 273–279.
Sarvas, R.: 1972, ‘Investigations on the Annual Cycle of Development of Forest Trees. ActivePeriod’, Commun. Inst. For. Fenn. 76, 1–110.
Sarvas, R.: 1974, ‘Investigations on the Annual Cycle of Development of Forest Trees. II. AutumnDormancy and Winter Dormancy’, Commun. Inst. For. Fenn. 84, 1–101.

PHENOLOGY AND CARBON SEQUESTRATION OF BOREAL FORESTS 113
Wallin, G., Bergh, J., Räntfors, M., and Linder, S.: 2000, ‘Measurement and Modeling of Post-Winter Recovery of Photosynthetic Capacity in Boreal Norway Spruce’, in The Role of BorealForests and Forestry in the Global Carbon Budget, May 8–12, 2000, Edmonton, Alberta, Canada,Abstracts, p. 135.
Wielgolaski, F-E.: 1999, ‘Starting Dates and Basic Temperatures in Phenological Observations ofPlants’, Int. J. Biometeorol. 42, 158–168.
(Received 23 January 2001; in revised form 14 March 2002)