a rapid enzyme immunoassay for cholectystokinin octapeptide sulfate
TRANSCRIPT

Neurochem. Int. Vol. 15, No. 1, pp. 55~0, 1989 0197-0186/89 $3.00+0.00 Printed in Great Britain. All rights reserved Copyright © 1989 Pergamon Press pie
A RAPID ENZYME IMMUNOASSAY FOR CHOLECTYSTOKININ OCTAPEPTIDE SULFATE
KATSUMICHI TAKEDA 1, FUMIAKA UCHIUMI l, MASATOSH1 TAKITA l, and TAKESHI KATO 2'*
LDepartment of Life Chemistry, Graduate School at Nagatsuta, Tokyo Institute of Technology, Yokohama 227 and 2Laboratory of Molecular Recognition, Graduate School of Science, Yokohama City
University, Yokohama 236, Japan
(Received 30 June 1988; accepted 5 January 1989)
Abstract--A rapid and sensitive enzyme immunoassay (EIA) for cholecystokinin octapeptide sulfate (CCK-8S) has been developed. The assay is based on a double antibody method using N-terminal specific antibody to CCK-8S, and CCK-8S conjugated with horseradish peroxidase as the enzyme labeled antigen. The time for the first incubation was 2 h. The time for the second incubation, during which the first antibody was precipitated using the second antibody which contained polyethylene glycol 6000 as an accelerator of the immune reaction, was 5 min. The total assay time was less than 3 h. Intra- and interassay coefficients of variation were 4.4-5.8 and 2.7-10.97%, respectively. The minimal detectable dose was 2 pg of CCK-8S in this assay. To demonstrate the utility of this EIA, we applied this assay to evaluate changes in the levels of CCK-8S immunoreactivity in various regions of the brain after kainic acid-induced seizures in the rat. This treatment decreased the contents of CCK-8S immunoreactivity in the frontal cortex, amygdala and hippocampus. These data show that this EIA system is rapid and sensitive enough to measure changes in the tissue levels of CCK-8S.
The peptide cholecystokinin (CCK) was first isolated from gut and identified as a gastrointestinal hormone (Ivy and Oldberg, 1928). Recently, gas t r in /CCK was found in the brain of mammals by radio- immunoassay (RIA) using antibodies directed against gastrin (Vanderhaeghen et al., 1975). The CCKs are widely distributed in most brain regions (Rehfeld, 1978b; Innis et al., 1979). The carboxy-terminal oc- tapeptide sulfate of C C K (CCK-8S) is the predomi- nant form (Rehfeld, 1978b; Dockray, 1980). Since CCK peptides may act as transmitters/ modulators within the limbic system (Beinfeld et al., 1983; Rehfeld, 1985), they may also be involved in the limbic pathologies linked to neuropsychiatric diseases (Ferrier et al., 1983) and may have some important clinical roles (Emson et al., 1980; Verbank et al., 1984). Although many RIA methods for the measurement of CCK have been reported (Rehfeld, 1978a; Himeno et al., 1983; Cantor, 1986; Fukamauchi et al., 1987), the description of non-iso- topic immunoassay methods are fewer (Yamamoto et al., 1986). Here, we report the development and application of a rapid and sensitive EIA for neuronal CCK.
*To whom all correspondence should be addressed.
EXPERIMENTAL PROCEDURES
Materials
CCK-8S, other CCK related peptides and poly L-glutamic acid (poly-Glu) were purchased from protein Research Foundation (Osaka, Japan). Human gastrin I (1-17) sulfate was from UCB Bioproducts (Belgium). Bovine serum albu- min (BSA), rabbit ?-globulin (Cohn Fraction II, III), horseradish peroxidase (HRP), kainic acid (KA) and apro- tinin were obtained from Sigrnan Chemicals (St Louis, Mo., U.S.A). Freund's complete adjuvant was obtained from Difco Labs (Detroit, Mich., U.S.A.). O-Phenylenediamine (OPD) tablets (14 mg of OPD per tablet) and anti-rabbit ),-globulin goat serum were obtained from Dainabot (Tokyo, Japan). Copolymer of D-glutamic acid and D-lysine, MW 6900 (D-GL) were obtained from Miles Labs (Elkhart, Ind., U.S.A.), keyhole limpet hemocyanin (KLH) from Calbiochem (San Diego, Calif., U.S.A.) and 30% hydrogen peroxide and polyethylene glycol 6000 from Wako Pure Chemicals (Osaka, Japan).
Preparation of immunogens CCK-8S conjugated to KLH (CCK-8-KLH) was used as
an immunogen. Cholecystokinin tetrapeptide (CCK-4) con- jugated to D-GL (CCK-4-D-GL) was used as a suppressor to suppress the production of antibody cross-reactive with CCK-4 which shares a common determinant with CCK-8S. Pretreatment of the rabbits with this tolerogenic conjugate leads to the inactivation of cross-reacting antibody forming precursor cells before immunization with CCK-8S (Hashimura et al., 1982). CCK-8-KLH was synthesized by
55

56 KATSUMICHI TAKEDA el al.
the method of Liu et al. (1979) as described in detail in our previous report (Yamamoto and Kato, 1986). Briefly, to prepare acyl-KLH, a solution of KL H (26 mg, 300 #1) was dialyzed against 0.125 M phosphate buffer, pH 7.2, and then mixed with 6.5 pmol (1.13 mg) of S-acetylmercaptosuccinic anhydride in dimethylformamide (DMF) (6 pt). For prep- aration of maleimidobenzoyl CCK-8S (MB-CCK-8), CCK- 8S (0.42 rag) in 0.5 ml of 0.2 M phosphate buffer, pH 8.0, was mixed with m-maleimidobenzoyl-N-hydroxysuccini- mide ester (MBS) (0.6 rag) in 50/al of DMF. After mixing of the acyl-KLH and MV-CCK-8 with nitrogen, 300 #1 of 0.5 M NH4OH was added to the mixture. The reaction mixture was stirred for 2 h at room temperature, and then dialyzed to remove free CCK-8S. CCK-4-D-GL was synthe- sized by the procedure described for the preparation of CCK-8-KLH using CCK-4 (50 mg in 1.2 ml of DMF) and D-GL (200mg in 4.5ml of 0.125 M phosphate buffer, pH 7.2) with the exception that D M F was used to dissolve CCK-4 because of its low solubility in phosphate buffer.
Immunization
Anti-CCK-SS specific antiserum was prepared according to the method of Hashimura et al. (1982). Each of three female white rabbits was injected subcutaneously with CCK-8-KLH (100 ,ug of CCK-8S for each injection) 8 times for 4 months. CCK-4-D-GL (6.25 mg of CCK-4) was also injected intraperitoneally 3 days before the second and third injections with CCK-8-KLH. Complete and incomplete Freund's adjuvants were used for the first 6 and next l0 weeks, respectively. All rabbits were bled 10 days after the final injection. Antibodies directed against CCK-8S were elicited in all of the rabbits.
Preparation o f enzyme conjugate
CCK-8S was conjugated with HRP by a slight modification of the periodate oxidation method of Wilson and Nakane (1978). For the oxidation o f HRP, a 0.5 M carbonate buffer, pH 9.5 instead of distilled water was employed because a higher reaction yield of enzyme conju- gate was obtained. CCK-8S vs HR P in a molar ratio of 1 : I was chosen. The conjugation procedure was performed as follows: 1 ml of HRP (8 mg/ml in 0.5 M carbonate buffer, pH 9.5) was oxidized with 1 ml of freshly prepared 0.I M NaIO4 at room temperature for 30 min, After dialysis of the oxidized product (HRP-aldehyde) against a 50 mM carbon- ate buffer, pH 9.5 overnight at 4°C, 0.5 ml of CCK-8S (0.4 mg/ml in 50 mM carbonate buffer, pH 9.5) was imme- diately added. After a 2 h reaction at room temperature, 0.11 ml of freshly prepared 160 m M sodium borohydride was added. The mixture was stirred for 2 h at 4°C and then dialyzed against 10 mM phosphate-buffered saline, pH 7.2 for 20h at 4°C. The conjugate and excess HRP were separated from unreacted CCK-8S on a column of Sepha- dex G-100 ( l . 6 x 9 3 c m ) in 10raM phosphate-buffered saline, pH 7.2, at 4°C. Aliquots of each column fraction were diluted in EIA buffer and assayed for enzyme conju- gate activity using the C C K EIA reagents. Detailed condi- tions are described below (EIA procedure). The most active fractions were pooled, mixed with BSA (I mg/ml) and then stored in 0.2ml aliquots at - 6 0 ° C . Concentration of the enzyme conjugate was determined by measuring the abs- orbance at 403nm (1%; 403nm; I c m = 2 2 . 5 ) (Tijssen, 1985a).
Preparation o[ rat brain samph, s
KA (10 mg/kg) or saline was injected i.p. into male Wistar rats weighing 30(~350 g as described by Meyer et al., (1986). The rats exhibiting strong limbic convulsions and controls were killed by decapitation 3 h after the injection. The brain tissues were rapidly separated on ice into the following regions: frontal cortex, amygdala, h ippocampus and sub- stantia nigra. These tissues were frozen in liquid nitrogen and stored at -80'~C. CCK peptides were extracted as described by Marley and Rehfeld (1984). Briefly, the brain tissues were weighed, mixed with approx. 20 vol of 90% methanol and sonicated. After centrifugation for 30 min at lO,O00g, the supernatant was lyophilized and frozen at _ 8 0 , ~ C .
EIA procedure
The E1A for CCK-8S was based on the double antibody method (Fig. 1). All assay reactions were performed in a single tube. In this study, we used transparent plastic tubes and a spectrophotometer (Quantum II dual wavelength analyzer, Abbott Labs, Irving, Tex., U.S.A.) which is de- signed to measure the absorbance of the solution directly in the plastic tube. A 10 m M phosphate-buffered saline solu- tion, pH 7.2, containing 0.1% BSA, 200 KIU/ml aprotinin and 0.01% thimerosal was used as the EIA buffer. The second antibody (anti-rabbit ~,-globulin goat serum) was diluted 1:50 with 0.1 M borate buffer, pH 8.6 containing 0.1% BSA, 4% PEG 6000 and 0.01% thimerosal. The total assay volume of 0.3 ml contained 0.1 ml of the diluted ant i -CCK 8S antiserum (1:6000 in EIA buffer containing 100 #g/ml rabbit ),-globulin), 0. l ml of the CCK-8S enzyme
CCK-8S s tandard or sample 100 ~ ]
AntI-CCK antibody 100 ~ 1
CCK-IIRP 100 ~ I
Incubate for 2 b at 4 °C
Add 500 I of a n t l - r a b b l t 7 f; an t ibody s o l u t i o n
Incubate for 5 min a t room tempera ture
C e n t r i f u g e a t 1,500 g for 20 min a t 15 °C
I)Iscard the supe rna t an t
Add 200 lz I of OPD so lu t ion
Incubate for 20 mln a t room tempera ture
Stop thc enzyme r e a c t i o n by adding 1 ml of l N II2SO.
Neasure t.he absorbanee at 492 rim
Fig. 1. CCK-8S EIA double antibody procedure. Both of the immune reactions and the enzyme reaction were per- formed in a single tube. Detailed assay conditions are
described in Experimental Procedures.
57
R E S U L T S
conjugate solution (0.1 #g of HRP per ml) and 0.1 ml of the sample or CCK-8S standard. The reaction mixture was incubated for 2 h at 4°C. To each tube was then added 0.5ml of the second antibody solution. After a 5min incubation at room temperature, free and bound enzyme conjugates were separated by centrifugation at 1500g for 20 min at 15°C. The supernatant was decanted, and to the precipitate was added 0.2 ml of a substrate solution (14 mg of OPD per 5 ml of 0.1 M phosphate-citrate buffer, pH 5.0 containing 0.01% H202). After a 20 min incubation at room temperature, 1 ml of 1 N H2SO 4 was added to stop the enzyme reaction and the absorbance at 492 nm was read in a Quantum II dual wavelength analyzer.
100
Standard curve
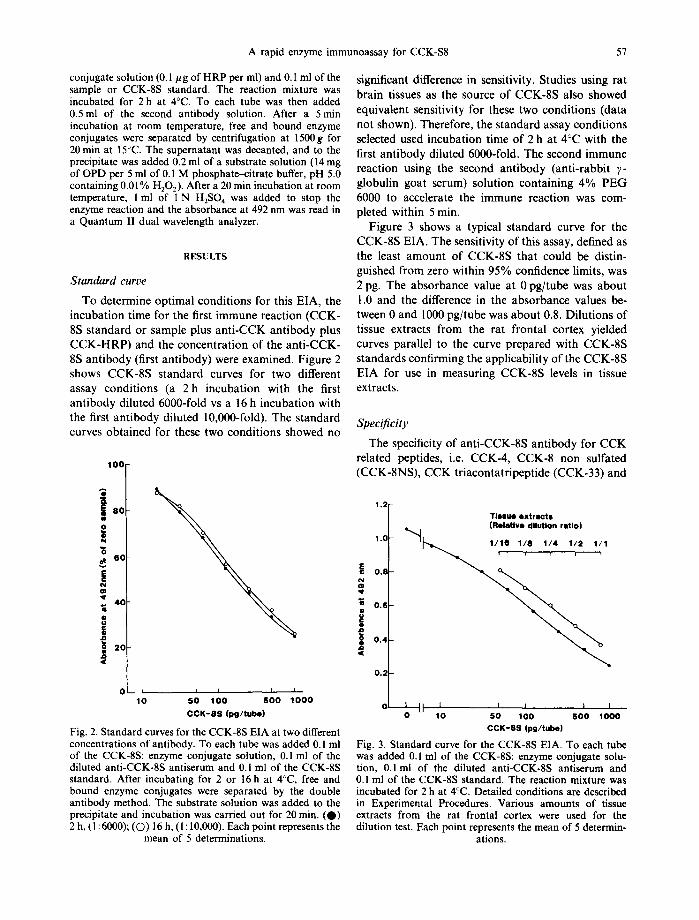
To determine opt imal condi t ions for this EIA, the incuba t ion t ime for the first immune react ion (CCK- 8S s tandard or sample plus an t i -CCK an t ibody plus C C K - H R P ) and the concen t ra t ion of the an t i -CCK- 8S an t ibody (first ant ibody) were examined. Figure 2 shows CCK-8S s tandard curves for two different assay condi t ions (a 2 h incuba t ion with the first an t ibody diluted 6000-fold vs a 16 h incubat ion with the first an t ibody diluted 10,000-fold). The s tandard curves ob ta ined for these two condi t ions showed no
1,2
1.0
o.S
i..0 o z
6 o
i 4 0
o o
i I 2 0
A rapid enzyme immunoassay for CCK-S8
0 i i i , i [
10 50 100 5 0 0 1 0 0 0
CCK-SS (pg/tube)
Fig. 2. Standard curves for the CCK-8S EIA at two different concentrations of antibody. To each tube was added 0.1 ml of the CCK-8S: enzyme conjugate solution, 0.1 ml of the diluted anti-CCK-8S antiserum and 0.1 ml of the CCK-8S standard. After incubating for 2 or 16 h at 4°C, free and bound enzyme conjugates were separated by the double antibody method. The substrate solution was added to the precipitate and incubation was carried out for 20 min. (O) 2 h, (I :6000); (O) 16 h, (1: I0,000). Each point represents the
mean of 5 determinations.
significant difference in sensitivity. Studies using rat bra in tissues as the source of CCK-8S also showed equivalent sensitivity for these two condi t ions (data not shown). Therefore, the s tandard assay condi t ions selected used incuba t ion t ime of 2 h at 4°C with the first an t ibody diluted 6000-fold. The second immune reaction using the second an t ibody (ant i - rabbi t ~- globulin goat serum) solut ion conta in ing 4 % P E G 6000 to accelerate the immune react ion was com- pleted within 5 rain.
Figure 3 shows a typical s t andard curve for the CCK-8S EIA. The sensitivity of this assay, defined as the least a m o u n t of CCK-8S tha t could be distin- guished f rom zero within 95% confidence limits, was 2 pg. The absorbance value at 0 pg/ tube was abou t 1.0 and the difference in the absorbance values be- tween 0 and 1000 pg/ tube was abou t 0.8. Dilut ions of tissue extracts f rom the rat f ronta l cortex yielded curves parallel to the curve prepared with CCK-8S s tandards confi rming the applicabil i ty of the CCK-8S EIA for use in measur ing CCK-8S levels in tissue extracts.
Specificity
The specificity of an t i -CCK-8S an t ibody for C C K related peptides, i.e. CCK-4, CCK-8 non sulfated (CCK-8NS), C C K t r iaconta t r ipept ide (CCK-33) and
Tissue extracts (RelaUve dilution ratio) -.q
0.6
0.4
0.;
I t l 1 I A I I 0 10 50 100 5 0 0 1000
CCK-8S (pg/tube)
Fig. 3. Standard curve for the CCK-8S EIA. To each tube was added 0.1 ml of the CCK-8S: enzyme conjugate solu- tion, 0.1 ml of the diluted anti-CCK-8S antiserum and 0.1 ml of the CCK-8S standard. The reaction mixture was incubated for 2 h at 4°C. Detailed conditions are described in Experimental Procedures. Various amounts of tissue extracts from the rat frontal cortex were used for the dilution test. Each point represents the mean of 5 determin-
ations.
58 KATSUMICHI TAKEDA et al,
Table I. Cross-reactivity of CCK related peptides in the E1A for CCK-8S
CCK related peptide Cross-reactivity* (%)
CCK-SS 100.0 CCK-4 0.31 CCK-SNS 1.26 CCK-33 105.0 Gastrin I 66.0
*As determined by the relative molar amounts needed for 50% displacement of the CCK-8S-HRP conjugate.
gastrin I (human) was studied. Cross-reactivity was expressed as the relative molar amounts needed for 50% displacement of the enzyme conjugated antigen. As shown in Table 1, the cross-reactivity for CCK-4, CCK-8NS, CCK-33 and gastrin I were 0.31, 1.26, 105 and 66%, respectively.
Reproducibi l io '
Intra- and inter-assay precision at three concentra- tions of CCK-8S in tissue extracts (low, medium and high) are shown in Table 2. The intra-assay CV values at each concentration were 4.4 (low), 4.7 (medium) and 5.8% (high), respectively. The inter-assay CV values at each concentration were 4.2 (low), 2.7 (medium) and 10.9% (high), respectively.
K A - i n d u c e d seizures in the rat brain
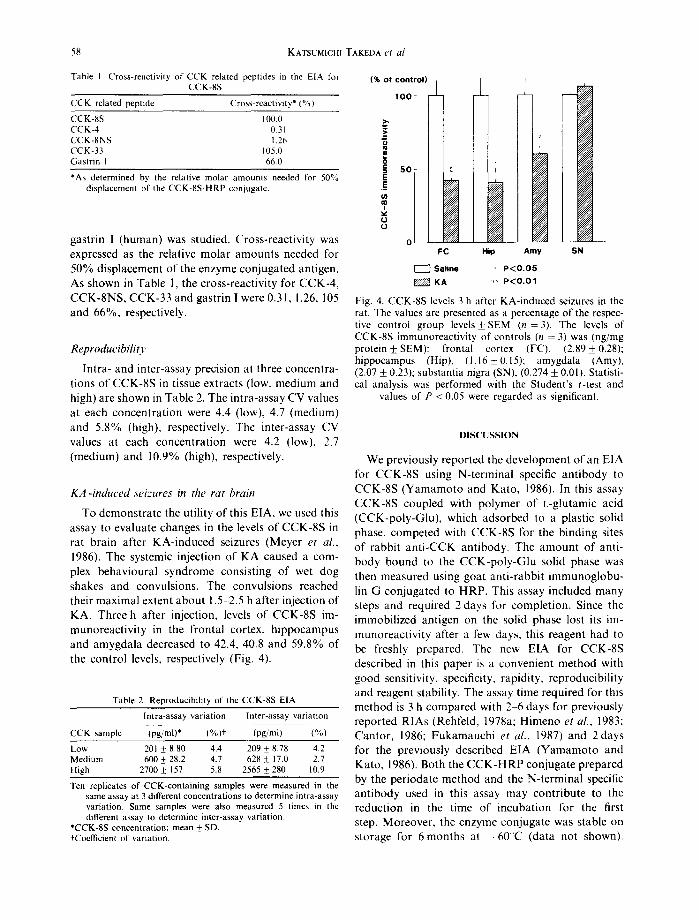
To demonstrate the utility of this E|A, we used this assay to evaluate changes in the levels of CCK-8S in rat brain after KA-induced seizures (Meyer et al., 1986). The systemic injection of KA caused a com- plex behavioural syndrome consisting of wet dog shakes and convulsions. The convulsions reached their maximal extent about 1.5-2.5 h after injection of KA. Three h after injection, levels of CCK-8S im- munoreactivity in the frontal cortex, hippocampus and amygdala decreased to 42.4, 40.8 and 59.8% of the control levels, respectively (Fig. 4).
Table 2. Reproducibility of the CCK-8S EIA
Intra-assay variation Inter-assay variation
CCK sample (pg/ml)* 1%)t (pg/ml) (%)
Low 201 ± 8.80 4.4 209 _+ 8.78 4.2 Medium 600 ± 28.2 4.7 628 ± 17.0 2.7 High 2700 ± 157 5.8 2565 + 280 10.9
Ten replicates of CCK-containing samples were measured in the same assay at 3 different concentrations to determine intra-assay variation. Same samples were also measured 5 times in the different assay to determine inter-assay variation.
*CCK-8S concentration; mean ± SD. ±'Coefficient of variation.
(% of control)
, o o
._~
s o
0
o I FC
Saline
±
Hip Amy SN
' P<O.05 ++ P < O . O 1
Fig. 4. CCK-8S levels 3 h after KA-induced seizures in the rat. The values are presented as a percentage of the respec- tive control group levels_+ SEM (n = 3). The levels of CCK-8S immunoreactivity of controls (n = 3) was (ng/mg protein + SEM): frontal cortex (FC), (2.89_+0.28); hippocampus (Hip), (1.16_+ 0.15); amygdala (Amy), (2.07 _+ 0.23); substantia nigra (SN), (0.274 _+ 0.01). Statisti- cal analysis was performed with the Student's t-test and
values of P < 0.05 were regarded as significant.
DISCUSSION
We previously reported the development of an EIA for CCK-8S using N-terminal specific antibody to CCK-8S (Yamamoto and Kato, 1986). In this assay CCK-8S coupled with polymer of L-glutamic acid (CCK-poly-Glu), which adsorbed to a plastic solid phase, competed with CCK-8S for the binding sites of rabbit anti-CCK antibody. The amount of anti- body bound to the CCK-poly-Glu solid phase was then measured using goat anti-rabbit immunoglobu- lin G conjugated to HRP. This assay included many steps and required 2 days for completion. Since the immobilized antigen on the solid phase lost its im- munoreactivity after a few days, this reagent had to be freshly prepared. The new EIA for CCK-8S described in this paper is a convenient method with good sensitivity, specificity, rapidity, reproducibility and reagent stability. The assay time required for this method is 3 h compared with 2 ~ days for previously reported RIAs (Rehfeld, 1978a; Himeno et al., 1983; Cantor, 1986; Fukamauchi et al., 1987) and 2 days for the previously described EIA (Yamamoto and Kato, 1986). Both the CCK-HRP conjugate prepared by the periodate method and the N-terminal specific antibody used in this assay may contribute to the reduction in the time of incubation for the first step. Moreover, the enzyme conjugate was stable on storage for 6months at - 6 0 C (data not shown).

A rapid enzyme immunoassay for CCK-S8 59
Using the [125I]CCK-8S labeled Bolton-Hunter reagent as a tracer (Rehfeld, 1978a) in this assay, sufficient binding of the tracer to antibody was not observed after short incubation times (data not shown). In our previous EIA system, an overnight incubation was needed for the immune reaction because the rate of antibody-antigen interaction was slower than in solution, due to the necessity for the free immunoreactant to diffuse to the solid phase (Tijssen, 1985b).
It is known that immune reactions can be acceler- ated considerably in the presence of PEG 6000 (Salo- nen and Vaheri, 1981; Konijin et al., 1982). In this new EIA we applied this method to the second immune reaction. Four percent PEG 6000 gave in- creased reaction rates without a significant increase in background levels. Therefore, the second immune reaction could be completed within 5 min, while more than several hours were needed for the conventional method (Himeno et al., 1983). In contrast, a significant acceleration was not observed in the first immune reaction by using the PEG 6000. The per- centage of cross-reactivity for CCK related peptides and human gastrin I in this assay (Table 1) was very similar to our previous data (Yamamoto and Kato, 1986), but higher than the value obtained by Hashimura et al. (1982). Based on the high cross- reactivity for CCK-33 (105%) and human gastrin I (66%), the antibody used in this assay might also recognize the sulfated tyrosine regions (Table 1). Although gastrins are found in the anterior pituitary lobe, they are not detectable in other regions of the central nervous system (Rehfetd, 1978a, b). There- fore, the antibody used in this EIA can measure CCK-8S specifically in brain tissues using selective extraction in cold 90% methanol (Marley and Rehfeld, 1984). The minimal level detectable in this EIA (2pg) is nearly equal to that in the RIA (6.6-1.6 pg) (Himeno et al., 1983; Cantor, 1986). To improve the sensitivity in this EIA we tried to apply the method of delayed addition of labeled antigen. However, a significant increase in sensitivity was not obtained (data not shown).
KA, a rigid analogue of glutamic acid, is a cyto- toxic agent causing strong excitation of neurons of the mammalian central nervous system (Johnston et al., 1974). In the rat, systemic, intraventricular or intra-amygdaloid injection of KA produces marked behavioural changes (Ben-Ari et al., 1979; Pisa et al., 1980; Schwob et al., 1980). After limbic seizures induced by i.p. injection of this toxin, pronounced changes in the levels of CCK-8S immunoreactivity have been reported by Meyer et al. (1986). The results
obtained in the present study (Fig. 4) support their findings. The decrease of CCK-8S immunoreactivity in the frontal cortex, hippocampus and amygdala during the acute seizures may be due to an increase of release and/or to a decrease of the synthesis of the peptide. We are presently investigating the mecha- nisms for this decrease by measuring CCK mRNA levels and the released amounts of the CCK peptides.
The EIA for CCK-8S described here is a useful method for measuring neuronal CCK-8S with signifi- cant advantages when compared to RIA methods. A number of antibodies currently used in RIA pro- cedures for CCK may be applicable to the develop- ment of an EIA method using the principles described in this paper.
Acknowledgement--We wish to thank Dr S. Rose for his generous advice in preparing the manuscript.
REFERENCES
Beinfeld M. C. (1983) Cbolecystokinin in the central ner- vous system--a mini review. Neuropeptides 3, 411-427.
Ben-Ari Y., Lagowska J., Tremblay E. and Le Gal Salle G. (1979) A new model of focal status epilepticus: intra- amygdaloid application of kainic acid elicits repetitive secondarily generalized convulsive seizures. Brain Res. 163, 176-179.
Cantor P. (1986) Evaluation of a radioimmunoassay for cholecystokinin in human plasma. Scand. J. clin. lab. Invest. 46, 213 221.
Dockray G. J. (1980) Cholecystokinin in rat cerebral cortex: identification, purification, and characterization by im- munochemical methods. Brain Res. 188, 155-165.
Emson P. C., Rehfeld J. F., Langevin H. and Possor M. (1980) Reduction in cholecystokinin-like immunoreactiv- ity in the basal ganglia in Huntington's disease. Brain Res. 198, 497-500.
Ferrier I. N., Roberts G. W., Crow T. J., Johnstone E. C., Owens D. G. C., Lee Y. C., O'Shaughnessy D., Adrian T. E., Polak J. M. and Blooms S. R. (1983) Reduced cholecystokinin-like and somatostatin-like immunoreac- tivity in limbic lobe is also associated with negative symptoms in schizophrenia. Life Sci. 33, 475-482.
Fukamauchi F., Yoshikawa T., Kaneno S., Shibuya H. and Takahashi R. (1987) Dopaminergic agents affected neu- ronal transmission of cholecystokinin in the rat brain. Neuropeptides I0, 207-220.
Hashimura E., Shimizu F., Nishino T., Imagawa K., Tateishi K. and Hamaoka T. (1982) Production of rabbit antibody specific for amino-terminal of cholecystokinin octapeptide (CCK-8) by selective suppression of cross-re- active antibody response. J. lmmun. Meth. 55, 375-387.
Himeno S., Tarui S., Kanayama S., Kuroshima T., Shinomura Y., Hayashi C., Tateishi K., Imagawa K., Hashimura E. and Hamaoka T. (1983) Plasma chol- ecystokinin responses after ingestion of liquid meal and intraduodenal infusion of fat, amino acids, or hydro- chloric acid in man: analysis with region specific radio- immunoassay. Am. J. Gastroent. 78, 703-707.

60 KATSUMICHI TAKEDA et al.
lnnis R. B., Corr6a F. M. A., Uhl G. R., Schneider B. and Synder S. H. (1979) Cholecystokinin octapeptide-like immunoreactivity: histochemical localization in rat brain. Pro. natn. Acad. Sci. U.S.A. 76, 521 525.
Ivy A. C. and Oldberg E. (1928) A hormone mechanism for gallbladder contraction and evacuation. Am. J. Physiol. 86, 599 613.
Johnston G. A. R., Curtis D. R., Davies J. and McCulloch R. M. (1974) Spinal interneurone excitation by conforma- tionally restricted analogues of L-glutamic acid. Nature, Lond. 248, 804 805.
Konijin A. A., Levy R., Link G. and Hershko C. (1982) A rapid and sensitive ELISA for serum ferritin employing a fluorogenic substrate. J. lmmun. Meth. 54, 297 307.
Liu F. -T., Zinnecker M., Hamaoka T. and Katz D. H. (1979) New procedure for preparation and isolation of conjugates of proteins and a synthetic copolymer of t~-amino acids and immunochemical characterization of such conjugates. Biochemistry 18, 690-697.
Marley P. D. and Rehfeld J. F. (1984) Extraction techniques for gastrins and cholecystokinins in the rat central ner- vous system. J. Neurochem. 253, 4022-4030.
Meyer D. K., Widmann R. and Sperk G. (1986) Increased brain levels of cholecystokinin octapeptide after kainic acid-induced seizures in the rat. Neurosci. Lett. 69, 208 211.
Pisa M., Sanberg P. C., Corcoran M. E. and Fibiger H. C. (1980) Spontaneously recurrent seizures after intra- cerebral injections of kainic acid in rat: a possible model of human temporal lobe epilepsy. Brain Res. 200, 481 487.
Rehfeld J. F. (1978a) lmmunochemicat studies on chol- ecystokinin. I. Development of sequence-specific radioim- munoassays for porcine triacontatriapeptide cholecysto- kinin. J. biol. Chem. 253, 401(~4021.
Rehfeld J. F. (1978b) lmmunochemical studies on cholecys- tokinin. II. Distribution and molecular heterogeneity in
the central nervous system and small intestine of man and hog. J. biol. Chem. 253, 4022-4030.
Rehfeld J. F. (1985) Neuronal cholecystokinin: one or multiple transmitters? J. Neurochem. 44, I 10.
Salonen E-M. and Vaheri A. (1981) Rapid solid-phase enzyme immunoassay for antibodies to viruses and other microbes: effects of polyethylene glycol. J. Immun. Meth. 41, 95-103.
Schwob J. W., Fuller T., Price J. L. and Olney J. W. (1980) Widespread patterns of neuronal damage following sys- temic or intracerebral injections of kainic acid: a histolog- ical study. Neuroscience 5, 991 1014.
Tijssen P. (1985a) Horseradish peroxidase. In: Practice and Theory o[ Enzyme Immunoassay (Burdon R. H. and Knippenberg P. H., eds), pp. 175 179. Elsevier, Amster- dam.
Tijssen P. (1985b) The immobilization of immunoreactants on solid phases. In: Practice and Theory of Enzyme Immunoassay (Burdon R. H. and Knippenberg P. H., eds), pp. 297 298. Elsevier, Amsterdam.
Vanderhaeghen J-J., Signeau J. C. and Gepts W. (1975) New peptide in the vertebrate CNS reacting with antibodies. Nature 257, 604~05.
Verbank P. M. P., Lotstra F., Gilles C., Linkowski P., Mendlewicz J. and Vanderhaeghen J-J. (1984) Reduced cholecystokinin immunoreactivity in the cerebrospinal fluid of patients with psychiatric disorders. Life Sci. 34, 67 72.
Wilson M. B. and Nakane P. K. (1978) Recent devel- opments in the periodate methods of conjugating horseradish peroxidase to antibodies. In: Immuno- [tuorescence and Related Staining Techniques (Knapp W.. Holuber K. and Wick G., eds), pp. 215-224. Elsevier, Amsterdam.
Yamamoto H. and Kato "1. (1986) Enzyme immunoassay for cholecystokinin octapeptide sulfate and its applica- tion. J. Neurochem. 46, 702 707.