a dissertation presented to the faculty of the graduate...
TRANSCRIPT

ENVIRONMENTAL MONITORING OF THE LOWER HUDSON RIVER AND
ASSESSMENT OF FACTORS LIMITING BACTERIAL GROWTH ON IBUPROFEN
ANALOGS
A Dissertation
Presented to the Faculty of the Graduate School
of Cornell University
In Partial Fulfillment of the Requirements for the Degree of
Doctor of Philosophy
by
Amy Joy Risen
January 2012

© 2012 Amy Joy Risen

iii
ENVIRONMENTAL MONITORING OF THE LOWER HUDSON RIVER AND
ASSESSMENT OF FACTORS LIMITING BACTERIAL GROWTH ON IBUPROFEN
ANALOGS
Amy Joy Risen, Ph.D.
Cornell University 2012
Pharmaceuticals and personal care products are increasingly recognized as important
environmental pollutants and have recently gained the attention of both the scientific community
and the public. A base-line study was conducted in the Lower Hudson River of New York State
employing both analytical chemistry and endocrine bioreporters to monitor both traditional and
emerging pollutants. 25 chemicals were detected via gas chromatography-mass spectrometry but
only caffeine (average 186 ng/L) and the polycyclic musk 1,3,4,6,7,8-hexahydro-4,6,6,7,8,8-
hexamethylcyclopenta-γ-2-benzopyran (average 80 ng/L) were found at quantifiable levels.
Using in vitro Chemical Activated LUciferase eXpression (CALUX) bioreporters capable of
measuring endocrine activity, we detected estrogenic (94-701 pg 17b-estradiol Eq/L) activity in
all six samples, both androgenic (1.15 ng DHT Eq/L) and progestrogenic (1.6-4.08 ng/Org2058
Eq/L) activity in one and two samples respectively, but no glucocorticoid activity in any
samples. The potential environmental fates and biological impacts of both these chemicals and
activities are discussed. Since one must look beyond detection of the parent compound in order
to understand the fate of pollutants, this dissertation also reports on the ability of Sphingomonas
sp. strain Ibu2 to metabolize analogs of ibuprofen, a pharmaceutical that has been commonly
detected in wastewater and aquatic environments around the world. While para-substituted

iv
aromatic compounds are typically reported to be more challenging to degrade, strain Ibu2 can
grow on some para-substituted arylacetic acids but not on non-substituted ones. Catabolic rates
for the degradation of both growth and non-growth substrates in the upper pathway and for meta
cleavage (MC) in the lower pathway were measured. Non-growth substrates, 4-tolylacetic acid
(4TAA), 2-propionic acid and phenylacetic acid, were found to cause the inhibition of MC
activity as well as cell death. Spontaneous mutants with the ability to grow on 4TAA were
characterized. The mechanisms behind MC activity inhibition and toxicity were examined and
CoA-thioester intermediates were identified as toxic metabolites during aromatic acid
metabolism.

v
BIOGRAPHICAL SKETCH
Amy Joy Risen was born in Cleveland, OH. As a youth she spent many weekends and
summers hiking, camping, and boating with her family in the whitewater mecca of the eastern
coast of the United States in PA and WV. She did not complete high school and went straight on
to obtain her first degree, an Associates of Applied Science in Recreation and Wildlife
Technology, at Hocking College in Nelsonville, OH in 1997. She received her Bachelor’s of
Science with a major in Biology and minor in Chemistry at Bloomsburg University in
Bloomsburg, PA in 2004. She is passionate about contributing to the world she lives in and has
volunteered as a wildlife rehabilitator, tree planter, trail clearer, public educator, and policy
advisor. Her life is full of a plethora of amazing people and opportunities, which she has
encountered while traversing 16 countries. She loves animals and has a wonderful black lab,
Apollo, whom she hikes with often, and three charming cats. She lives her life as sustainably as
she can to minimize her contribution to the myriad of environmental challenges that the world is
currently facing. She came to Cornell University in Ithaca, NY to pursue her interests in
toxicology and waste management in the Field of Environmental Toxicology. Her graduate
experience has enhanced her appreciation for the complexity of life and this dissertation is
representative of the interdisciplinary approach that she often employs.

vi
ACKNOWLEDGEMENTS
This research was funded by the New York State Department of Environmental
Conservation’s Hudson River Estuary Program and the New York State Water Resources
Institute. My doctoral research was also supported by the Initiative for Future Agriculture and
Food Systems from the U.S. Department of Agriculture’s Cooperative State Research,
Education, and Extension Service (grant # 2001-52104-11484); the National Institute of
Environmental Health Science training grant in Environmental and Molecular Toxicology (grant
# 5T32 ES07052); two teaching assistantships through the Cornell Department of Neurobiology
and Behavior; and a U.S. Geological Survey graduate research assistantship. I would like to
thank the people from the U. S. Geological Survey that I worked with on the Hudson River
project including Patrick J. Phillips, Steven G. Smith, and Daniel D. Edwards. I would also like
to thank my committee members Dr. Eugene L. Madsen and Dr. Larry P. Walker for sharing
their facilities and expertise. Finally, I would like to thank my advisor Dr. Anthony G. Hay for
his guidance and eagerness to pass on a diverse set of skills.

vii
TABLE OF CONTENTS
BIOGRAPHICAL SKETCH .......................................................................................................... v ACKNOWLEDGEMENTS........................................................................................................... vi CHAPTER 1. Introduction ............................................................................................................. 1
1.1. Emerging contaminants ....................................................................................................... 1 1.1.1. Presence of emerging contaminants in aquatic environments ...................................... 3
1.1.1.1. Toxicity and effects in the field ............................................................................. 4 1.1.1.2. WWTPs as a source of emerging contaminants .................................................... 8
1.2. Microbial pathways for the catabolism of aromatic compounds ....................................... 11 1.2.1. Aerobic pathways........................................................................................................ 14
1.2.1.1. Meta cleavage activity, deactivation, and reactivation ........................................ 17 1.2.2. Toxicity during the degradation of aromatic hydrocarbons........................................ 18 1.2.3. Strain Ibu2 and the ipf pathway .................................................................................. 19
1.3. Conclusion ......................................................................................................................... 22 CHAPTER 2. Occurrence of traditional and emerging contaminants and endocrine activity in water samples from HRECOS stations on the Lower Hudson River, New York......................... 23
2.1. Introduction........................................................................................................................ 24 2.2. Materials and Methods....................................................................................................... 27
2.2.1. Site selection ............................................................................................................... 27 2.2.2. Sample collection........................................................................................................ 27 2.2.3. Chemical analysis ....................................................................................................... 29 2.2.4. CALUX analysis......................................................................................................... 29 2.2.5. Chemicals.................................................................................................................... 30 2.2.6. Statistical analysis....................................................................................................... 30
2.3. Results and Discussion ...................................................................................................... 31 2.3.1. Chemical analysis ....................................................................................................... 31
2.3.1.1. Waste indicators................................................................................................... 33 2.3.1.2. Personal care products & detergents.................................................................... 34 2.3.1.3. Flame retardants and plasticizers ......................................................................... 36 2.3.1.4. Pesticides.............................................................................................................. 37 2.3.1.5. Aromatic hydrocarbons from oil and coal ........................................................... 39 2.3.1.6. Non detects & quality control .............................................................................. 40
2.3.2 CALUX........................................................................................................................ 43 CHAPTER 3. Atypical meta cleavage deactivation and growth substrate range in the ibuprofen degradation pathway of Sphingomonas sp. strain Ibu2 ................................................................ 47
3.1. Introduction........................................................................................................................ 48 3.2. Materials and Methods....................................................................................................... 51
3.2.1. Chemicals.................................................................................................................... 51 3.2.2. Culturing of strains ..................................................................................................... 52 3.2.3. Cloning the 4-isobutylcatechol-2,3-dioxygenase (ipfJ) .............................................. 52 3.2.4. Generation of spontaneous 4TAA mutants................................................................. 52 3.2.5. MC activity ................................................................................................................. 53 3.2.6. Deactivation of IpfJ..................................................................................................... 54 3.2.7. Catechol formation by the upper pathway .................................................................. 54

viii
3.3. Results................................................................................................................................ 56 3.3.1. Meta cleavage activity ................................................................................................ 56 3.3.2. Formation of catechol metabolites in the upper pathway ........................................... 60 3.3.3. 4TAA mutants............................................................................................................. 61
3.4. Discussion .......................................................................................................................... 65 CHAPTER 4. Growth inhibition during arylacetic acid metabolism by the ibuprofen degrader Sphingomonas sp. strain Ibu2 ....................................................................................................... 70
4.1. Introduction........................................................................................................................ 72 4.2. Methods.............................................................................................................................. 75
4.2.1. Chemicals.................................................................................................................... 75 4.2.2. Culturing of strains ..................................................................................................... 76 4.2.3. Construction of new strains ........................................................................................ 76 4.2.4. Growth inhibition assays............................................................................................. 76 4.2.5. MC activity ................................................................................................................. 77 4.2.6. Toxicity of CoA-AAA thioesters................................................................................ 77 4.2.7. Toxicity of upper pathway metabolites....................................................................... 78
4.3. Results................................................................................................................................ 79 4.3.1. Toxicity of the IPF analogs......................................................................................... 79 4.3.2. Induction with IPF mitgates analog toxicity............................................................... 81 4.3.3. Ibu2 strains engineered for high MC activity ............................................................. 83 4.3.4. Supplementation with the antioxidant thiourea .......................................................... 85 4.3.5. CoA-AAA thioester toxicity ....................................................................................... 87 4.3.6. Toxicity of AAA metabolism in mutants of the ipf pathway...................................... 88
4.4. Discussion .......................................................................................................................... 90 CHAPTER 5. Conclusion ............................................................................................................. 95
5.1. Monitoring of the Lower Hudson River ............................................................................ 95 5.2. Factors limiting the arylacetic substrate range of strain Ibu2 ............................................ 97
5.2.1. MC activity levels are likely contributing to the lack of growth on 4TAA................ 97 5.2.2. Atypical MC deactivation by 2PPA and PAA............................................................ 98 5.2.3. Expression of the PLFeDox mitigated stress .............................................................. 99 5.2.4. Inhibition of growth by the antioxidant thiourea ...................................................... 100 5.2.5. Conclusion for strain Ibu2 ........................................................................................ 101
APPENDIX................................................................................................................................. 102 A.1. Arylacetic acids that support growth .............................................................................. 102 A.2. Deactivation of MC activity in strain Ibu2 by ibuprofen................................................ 103 A.3. Operon structure.............................................................................................................. 104 A.4. Meta cleavage activity in engineered strains of Ibu2...................................................... 105 A.5. Growth Inhibition by CoA-AAA thioesters.................................................................... 106 A.6. Culture growth conditions have an effect on toxicity ..................................................... 108 A.7. Strain Ibu2 sequence and similarity with sequences in the NCBI databank................... 110
A.7.1. Contig sequences...................................................................................................... 115 A.8. Transcription of ipf genes ............................................................................................... 122

ix
LIST OF FIGURES Figure 1-1. Comparison of typical aerobic, anaerobic and hybrid pathways with the proposed ipf pathway. ........................................................................................................................................ 13 Figure 1-2. The Meta Cleavage Pathway...................................................................................... 16 Figure 1-3. Ibuprofen (IPF) and analogs metabolized by strain Ibu2........................................... 21 Figure 2-1. The Lower Hudson River in NY State with Hudson River Environmental Conditions Observing System (HRECOS) sites indicated with stars and sampling sites indicated with arrows............................................................................................................................................ 28 Figure 2-2. Transactivation of (A) androgen, (B) estrogen, and (C) progesterone response elements in CALUX bioassay by Lower Hudson River samples from winter and spring. .......... 44 Figure 3-1. MC activity of strain Ibu2 metabolizing growth supporting doses (250 mg/L) of analogs for 24 hours...................................................................................................................... 56 deviations, n=3.............................................................................................................................. 56 Figure 3-2. MC substrate range of strains Ibu2 and 3G7 and ipfJ mutants of 3G7 (TnH5 and TnG9)............................................................................................................................................ 58 Figure 3-3. Deactivation of heterologously expressed IpfJ.. ........................................................ 58 Figure 3-4. Induction of MC activity in strain Ibu2 by low doses (50 mg/L) of analogs and a no analog control................................................................................................................................ 59 Figure 3-5. Growth of strain Ibu2 and mutant strains 4TAA1, 4TAA2 and 4TAA3 on (A) 500 mg/L glycerol, (B) 250 mg/L IPF, or (C) 250 mg/L 4TAA. ........................................................ 62 Figure 3-6. MC activity of strain Ibu2 and mutant strains 4TAA1, 4TAA2, and 4TAA3 following 24 hour exposure to 0 (control), 50, or 500 mg/L 4TAA while growing on glycerol. ................. 63 Figure 3-7. Upper pathway activity of strain Ibu2 relative to mutant strains 4TAA1, 4TAA2, and 4TAA3. ......................................................................................................................................... 64 Figure 4-1. Ipf pathway metabolites, enzymes and genes of strain Ibu2 as proposed by Murdoch (2011)............................................................................................................................................ 73 Figure 4-2. Inhibition of growth of strain Ibu2 by IPF analogs.................................................... 80 Figure 4-3. Growth of strain Ibu2 on IPF plus analogs. ............................................................... 81 Figure 4-4. Meta cleavage activity in strain Ibu2 cultures suppressed by 500 mg/L analogs and rescued with 10 mg/L IPF............................................................................................................. 82 Figure 4-5. Growth inhibition in Ibu2 and engineered strains...................................................... 84 Figure 4-6. Combined effect of the antioxidant thiourea (TU) and IPF analog metabolism on strain Ibu2 growth. ........................................................................................................................ 86 Figure 4-7. Growth inhibition by metabolic products of ipf-CoA-ligase (ipfF)........................... 87 Figure 4-8. Toxicity of AAA exposure to E. coli harboring fosmids containing mutations in the ipf pathway.................................................................................................................................... 89 Figure A-1. Growth of strain Ibu2 on 250 mg/L IPF or analogs. ............................................... 102 Figure A-2. Strain Ibu2 MC activity in cultures induced with various concentrations of IPF while growing on glycerol. ................................................................................................................... 103 Figure A-3. Ipf and related MC genes of strain Ibu2.. ................................................................ 104 Figure A-4. MC activity before (A) and after (B) MC deactivation in Ibu2 and engineered strains over expressing ipfJ (Ibu2 pC) or ipfJ and ipfK (Ibu2 pC+P). ........................................ 105 Figure A-5. Growth Inhibition curves for E. coli JM109 in the presence of IPF analogs with and without heterologous expression of the ipf-CoA-ligase (ipfF). .................................................. 107

x
Figure A-6. Inhibition of growth by IPF and analogs in strain Ibu2 under different growth conditions.................................................................................................................................... 109 Figure A-7. Five contigs of strain Ibu2 DNA from fosmid clones 3G7 and 4F6. Upper ipf pathway genes are shown in black.............................................................................................. 111 Figure A-8.1. Transcription of ipf pathway genes. ..................................................................... 123 Figure A-8.2. Gel photos from RT-PCR for the determination of transcript boundaries for strain Ibu2 ipf and related genes, and expression from induced and non-induced cultures. ................ 125 Figure A-8.3. Digest to confirm the identity of band 2, product 7, gel 1 from Figure A-8.2..... 127

xi
LIST OF TABLES
Table 1-1. Organic wastewater contaminants found at mg/L concentrations in U.S. streams in a national reconnaissance (data from (Kolpin et al., 2002). .............................................................. 2 Table 2-1. Geographic coordinates and sampling dates for Lower Hudson River sampling sites........................................................................................................................................................ 28 Table 2-2. Chemicals detected in Lower Hudson River samples. Quantitative results are those detected above the limits of quantification (LOQ). ...................................................................... 32 Table 2-3. Chemicals tested for but not detected in Lower Hudson River samples and limits of quantification (LOQ). ................................................................................................................... 42 Table 3-1. Strains used in this study ............................................................................................. 51 Table 3-2. Formation of catechols by the upper pathway of strain Ibu2. ..................................... 60 Table 4-1. Strains used in this study. ............................................................................................ 75 Table A-7. Sequence similarity between strain Ibu2 and sequences in the NCBI databank. ..... 112 Table A-8.1. Primers used for RNA work .................................................................................. 123 Table A-8.2. Amplicons used to determine transcript boundaries and transcription in the presence and absence of IPF. ..................................................................................................................... 124

1
CHAPTER 1
Introduction
1.1. Emerging contaminants
Emerging pollutants are gaining the attention of both the scientific community and the
public. Pharmaceuticals and personal care products (PPCPs) are a large portion of this new class
of pollutants (Daughton, 2002). Pharmaceuticals originate from both human and agricultural
use, whereas personal care products are ingredients found in items such as soaps and beauty
products including antimicrobials, fragrances, and UV filters. We can expand our view and
consider consumer products including textiles and kitchenware, which contain additives such as
flame-retardants, plasticizers, anticorrosives, and again, antimicrobials. The effects that emerging
pollutants warrant concern (Jobling et al., 1998), and both research and public action are required
to reduce their presence in the environment. This chapter will provide a brief overview of the
toxicity, source, and biodegradability of emerging pollutants, with a focus on aromatic
compounds and ibuprofen in particular.

2
Table 1-1. Organic wastewater contaminants found at µg/L concentrations in U.S. streams in a national reconnaissance (data from (Kolpin et al., 2002). This is a representative list of the contaminants found at the highest concentrations in 139 streams in 30 states in one study. The maximum and median concentrations as well as frequency of detection are shown. Other investigators have found these contaminants at different concentrations and other contaminants have been found at similar concentrations.

3
1.1.1. Presence of emerging contaminants in aquatic environments
Scientists have taken many approaches to investigating emerging pollutants. Several
large projects have been undertaken in both the U.S. and Europe to measure the concentrations
of these chemicals in aquatic environments. Thus for my research, I started by looking at what
has been found via environmental analytical chemistry approaches. In the U.S., the U.S.
Geological Survey has undoubtedly led the way by selecting sample sites across the country and
giving us insight in to the widespread nature of the contamination of our waters (Barnes et al.,
2008; Focazio et al., 2008; Kolpin et al., 2002). Generally speaking, most emerging
contaminants are found at concentrations of ng/L in U.S. surface waters, however, the
concentrations of some chemicals can reach in to the low µg/L level (Table 1-1) (Kolpin et al.,
2002) and similar concentrations have been observed all over the world (Camacho-Munoz et al.,
2010; Sacher et al., 2008; Shala and Foster, 2010; Yoon et al., 2010). For example, in a
groundwater project spanning 23 European countries, investigators found many of the same
compounds listed in Table 1-1, including: benzotriazoles, caffeine, nonylphenol,
sulfamethoxasole, ibuprofen, DEET, and bisphenol A (Loos et al., 2007). They found most
chemicals at low ng/L concentrations on average, however even in groundwater, the maximum
concentrations of 10 analytes were in the µg/L range. In a different study, the River Rhine in
Germany was monitored for 10 years for pharmaceuticals and while statistical analysis indicated
a slight decrease in concentrations over the period of 1997-2006, the authors commented that
concentrations appeared to be fairly constant (Sacher et al., 2008). The pharmaceuticals that
they found at the highest concentrations were the antiepileptic carbamazepine, the lipid lowering
agent bezafibrate, and the non-steroidal anti-inflammatory drug (NSAID) diclofenac; the loading

4
rates at the most downstream site well exceeded one ton per year for each of these drugs.
Emerging contaminants are ubiquitous in our waters, and require our attention to determine what
effects they may be having and how to limit their release into the environment.
1.1.1.1. Toxicity and effects in the field
We have established that emerging contaminants are in our waters, but are the
concentrations that are present biologically relevant? While most emerging contaminants are
present at ng/L concentrations, investigators continue to find many compounds in the µg/L range
(Camacho-Munoz et al., 2010; Dsikowitzky et al., 2004b; Kolpin et al., 2002; Matamoros et al.,
2010; Yoon et al., 2010). Conversely, toxicological studies often find acute effects at mg/L
concentrations, but sublethal effects have been reported in the µg/L to ng/L range. For example,
many of us think of caffeine as a relatively benign substance and it is true that in order to cause
lethality to 50% of the population (LC50) with a simple organism such as the macroinvertebrate
Hydra attenuata it requires concentrations greater than 100 mg/L (Quinn et al., 2008). However,
green frog (Rana pipiens) tadpoles had both reduced activity and an increase in the startle
response during long term (28 days) exposure to environmentally relevant concentrations (600
ng/L) (Fraker and Smith, 2004). The authors suggested that this could decrease their fitness by
increasing their predation rate. We see a similar trend for the NSAID ibuprofen (IPF), with a
high LC50 for Daphnia magna of 133 mg/L (Han et al., 2006). However, a dose dependant
response was observed during chronic exposure of Japanese medaka (Oryzias latipes) to 1 to 100
µg/L IPF, inducing longer days between spawning and more eggs per spawning event (Flippin et
al., 2007). A decrease in cyclooxygenase activity was also observed and the authors suggested

5
that interference with ovulation may be due to the role of prostaglandins in oocyte maturation.
Both of these examples of low dose effects reflect carefully chosen endpoints which mirror the
intended effects of these compounds; ibuprofen inhibits prostaglandin synthesis via
cyclooxygenase inhibition (Smith et al., 2000), and caffeine is well known to be a stimulant. We
must also consider that organisms in the field encounter a myriad of toxicants simultaneously
which can have both additive and synergistic effects (Cleuvers, 2004; Fraker and Smith, 2004).
Diclofenac, another NSAID has also received a lot of attention from the scientific
community and the public. Three Gyps vultures (Gyps bengalensis, G. indicus, and G.
tenuirostris) in Asia were experiencing dramatic population declines and the cause was found to
be diclofenac poisoning (Green et al., 2004). The vultures feed on dead livestock, unfortunately,
one meal of diclofenac-treated livestock was sufficient to cause the kidney failure that had been
observed in the field (Oaks et al., 2004). A questionnaire sent to veterinary personnel revealed
that other NSAIDs including carprofen, flunixin, ibuprofen, and phenylbutazone had had toxic
effects on other wildlife including raptors, storks, cranes, and owls (Cuthbert et al., 2007). The
Indian government was prepared to ban the veterinary use of Diclofenac, so additional testing
was done and the alternative NSAID meloxicam was suggested for safe use (Swan et al., 2006).
The effectiveness of the ban is still to be seen, however the testing of livestock livers in India has
revealed a slight decrease in prevalence of diclofenac from 10.7% to 6.5% in the two years
following the ban (Cuthbert et al., 2011).
Endocrine disruption is another hot topic in environmental toxicology. The endocrine
system is an incredibly complex system that sends out chemical messengers to tissues in distant

6
locations throughout the body to orchestrate metabolism, growth, development, and reproduction
(Casals-Casas and Desvergne, 2011). Endogenous chemicals such as estrogens, androgens,
progesterone, glucocorticoids, mineralocorticoids, thyroid hormone, and vitamin D are all
endocrine signaling molecules and each are recognized by their own receptors to trigger cascades
of gene expression (Casals-Casas and Desvergne, 2011). Bioreporters have been constructed
using these hormone receptor elements, such that exposure to a hormonally active substance will
trigger expression of a luciferase gene which can be easily measured in the laboratory
(Sonneveld et al., 2005). I will demonstrate the use of these reporters in Chapter 2. Some of the
most recognized endocrine disrupting emerging pollutants include: parabens, polycyclic musks,
UV filters, organotins, components of plastics, perflourinated alkanes, brominated flame-
retardants, detergent breakdown products such as alkylphenols, antimicrobials such as triclosan
and triclocarban, bisphenol A, and phthalates (Casals-Casas and Desvergne, 2011; Witorsch and
Thomas, 2010).
Bioreporters alone, however, cannot capture and integrate all aspects of endocrine
disruption. For example, sexual function is a complex process required for reproduction and
therefore is an important factor for maintaining populations of flora and fauna in the wild. A
morphological test was developed for use in the lab in which reed frogs (Hyperolius argus) can
be exposed to exogenous chemicals and observed for sex specific color patterning to indicate
sexual differentiation and endocrine disruption (Hayes, 2005; Hayes and Menendez, 1999).
Other endpoints of sexual morphology are also very valuable for direct enumeration of effects in
the field. One such endpoint is fin anatomy. Female mosquitofish (Gambusia affinis) living
downstream of a pulp paper mill in the Fenholloway River of Florida were found to have

7
masculinized anal fin morphology, an androgen-dependant trait (Parks et al., 2001). In addition,
they had reduced sexual behavior and their pregnancy rate was 55% lower than for mosquitofish
in a nearby uncontaminated river (Toft et al., 2004).
Wastewater treatment plant (WWTP) effluents, on the other hand, have been observed to
have feminizing effects on aquatic species, including skewed sex ratios, elevated vitellogenin (an
egg protein) production, impaired gonadal development, and reduced fertility (Bjerregaard et al.,
2006; Jobling et al., 2009; Jobling et al., 2002; Jobling et al., 1998; Vajda et al., 2008). Certain
species may be more sensitive than others to endocrine effects. For example, smallmouth bass
(Micropertus dolomieu) and carp (Cyprinus carpio) had more highly elevated endocrine
biomarker responses in comparison with largemouth bass (M. salmoides) and bullhead
(Ameiurus nebulosus) at the same Hudson River sites (Baldigo et al., 2006). The contaminants
being investigated were primarily Hg and polychlorinated biphenyls (PCBs). Importantly, at the
sites most contaminated with PCBs there were no smallmouths to be found. In a different U.S.
nationwide study, smallmouth bass were the most common fish found to have intersex
characteristics (Hinck et al., 2009). Sensitivity differences have been observed for frogs as well.
For example, dosing of a Canadian lake with environmentally relevant concentrations of a birth
control ingredient (5-6 ng 17alpha-ethynylestradiol/L) had a greater impact on hatching success
for green frogs (R. pipiens) than mink frogs (R. septentrionalis) (Park and Kidd, 2005).
Many emerging contaminants such as triclosan, polycyclic musks, and halogenated
flame-retardants are bioaccumulative (Coogan and La Point, 2008; Fromme et al., 2001; Letcher
et al., 2009). Nonylphenols, polycyclic musks, and organophosphorus flame-retardants have

8
been found in wild populations of edible fish at concentrations in the high ng/g to low µg/g range
(Fromme et al., 2001; Gatermann et al., 2002; Ramirez et al., 2009; Sundkvist et al., 2010). One
route of human exposure to aquatic pollutants is through the consumption of wild caught fish. In
fact, a study in New York State found that male children whose mothers consumed sport fish
during pregnancy had an elevated rate of birth defects that was significant at 12%, compared to
the background rate of 3-4% (Mendola et al., 2005). A recent biomonitoring project reported on
the presence of a wide variety of emerging contaminants in human blood serum and urine
(Wilding et al., 2011). Polybrominated diphenyl ethers (PBDEs) were found at ng/L
concentrations but others were found in the µg/L range including: triclosan, mercury, bisphenol
A, perflourinated compounds, and phthalates. PBDEs are bioaccumulative neurotoxins and
correlations have been found between human umbilical cord concentrations and the IQs and
mental and psychomotor development of children (Herbstman et al., 2010). Another study
demonstrated that Mexican-American children living in California had higher blood serum levels
of PBDEs than their Mexican counterparts, suggesting that the stringent furniture flammability
standards in place in CA may be having an impact on the chemical body burden of that state’s
inhabitants (Eskenazi et al., 2011). Clearly, emerging contaminants are affecting both wildlife
and humans.
1.1.1.2. WWTPs as a source of emerging contaminants
WWTPs are the major route through which emerging contaminants enter our aquatic
environment (Daughton and Ruhoy, 2009; Metcalfe et al., 2003b; Yoon et al., 2010). Removal
rates for parent compounds in WWTPs vary with near complete removal of natural estrogens and

9
IPF at one end of the spectrum, and essentially no removal of carbamazepine and perflourinated
compounds at the other end (Clara et al., 2005; Heidler and Halden, 2008). Removal rates tend to
be higher in the summer months than the winter months and this may be attributable to the
essential role of microbes in degradation and their higher activity at warmer temperatures
(Castiglioni et al., 2006). During wastewater treatment there are several factors that can
contribute to the effective removal of emerging pollutants. One factor is the amount of time that
these chemicals have for degradation (Quintana et al., 2005; Radjenovic et al., 2008). Studies at
WWTPs have suggested that hydraulic retention times of 12 hours and solids retention times of
10 days often remove 90% of the majority of parent compounds (Clara et al., 2005; Metcalfe et
al., 2003a; Miege et al., 2008). Precipitation events, however, can cause the release of untreated
wastewater through combined sewer overflows, short circuiting the treatment process (Buerge et
al., 2006). This suggests that wastewater treatment plants should be upgraded with higher
capacities to accommodate more flow and longer retention times. While we consider the
upgrading of WWTPs we should consider that new technologies such as chemical oxidation
(mostly ozonation), UV, and membrane bioreactors are being developed that may be more
effective at treating recalcitrant pollutants (Cornel and Krause, 2006; Kim et al., 2009). But do
we have the right set of tools at our disposal to evaluate their effectiveness? Chapter 2 of this
dissertation reports on both analytical chemistry and bioreporter measurements of wastewater
pollutants in the Lower Hudson River in New York State. Its results, demonstrating higher levels
of endocrine disrupting activity than could not be attributed to compounds identified using the
analytical chemistry approach, speak to the need for a multifaceted approach to environmental
monitoring.

10
Current analytical chemistry strategies focus on the detection of parent compounds. The
problem with this is that many metabolites are produced during degradation and some of them
can be more toxic than the parent compounds (Munro et al., 1999; Stratton and Corke, 1982). In
order to know what chemicals to measure we need to know what metabolites might be produced.
These can be metabolites of human metabolism of pharmaceuticals such as glucuronide and
sulfate conjugates as well as hydroxylated and carboxylated products (Knights and Drogemuller,
2000; Reddy et al., 2005; Tan et al., 2002). These can also be biotic and abiotic degradation
products produced during wastewater treatment (Poiger et al., 2001; Quintana et al., 2005). As a
side project (data not presented in this dissertation) I evaluated chemicals that were tentatively
identified in water samples from WWTPs impacted by pharmaceutical manufacturing facilities
(Risen et al., 2009). The majority of these samples were obtained concurrent with those reported
in a USGS study and employed the same methods (Phillips et al., 2010b). However, additional
tentatively identified chemicals (TICs) were found without a priori knowledge of their existence
using gas chromatography-mass spectrometry in the absence of analyte standards; the mass
spectra of all resulting peaks were compared with reference spectra from the mass spectral
library from the 1998 National Institute of Standards and Technology. One of the major findings
of this analysis was that six of the codeine and morphine derivative TICs have previously been
shown to be transformed into one another by the WWTP isolate Pseudomonas putida strain m10
(Lister et al., 1999). This suggests that a compound such as oxycodone could occur in the field
as a metabolite of codeine, despite the fact that both codeine and oxycodone are generally
considered to be parent compounds. Similarly, the NSAID aceclofenac has been reported to be
transformed into the NSAID diclofenac (Perez and Barcelo, 2008). WWTP studies can provide
important information concerning the rate of removal of pollutants from waste streams and these

11
two examples suggest that the metabolic relationship of structurally similar compounds should
be considered when measuring removal rates to improve the accuracy of mass balances. Thus,
metabolic pathway research is valuable as it can provide a potential explanation for the sources
of chemicals found in samples from the field. Basic research examining the biodegradation
pathways used by microbes in WWTPs will also contribute to our knowledge about parent
compounds that might be persistent, as well as metabolites that might accumulate. The study of
unique biodegradation pathways can also provide more general knowledge to serve as a
foundation when studying other degradation pathways.
One of the focuses of my work was to investigate limiting factors in an IPF degradation
pathway (ipf). IPF and many other NSAIDs share an arylacetic acid moiety, as do the
compounds that I will be evaluating. The high prevalence of IPF in our waters and its high
removal rate make it an ideal model compound for degradation studies. Sphingomonas sp. strain
Ibu2 is a WWTP bacterial isolate that can grow on IPF, but it has an unusual substrate range, as I
will discuss in Chapters 3 and 4. The following section will give some background on
biodegradation pathways and the limitations that are often experienced by microorganisms
attempting to grow on aromatic compounds.
1.2. Microbial pathways for the catabolism of aromatic compounds
When microbes grow on a carbon source they must produce the enzymes needed for
metabolism and escape any toxicity incurred. Aromatic compounds are an abundant food source

12
for bacteria and aerobic, anaerobic, and hybrid pathways are used for the catabolism of aromatic
acids (Diaz, 2004; Fuchs, 2008). Each of these pathways must perform two energetically
expensive steps to destabilize the aromatic ring and open it, however the mechanisms employed
to achieve these steps differ. Aerobic pathways destabilize aromatic rings by the addition of
oxygen via monooxygenases or ring hydroxylating dioxygenases and converge on
dihydroxylated intermediates that are cleaved oxygenolytically (Diaz, 2004). This is typified by
the cumate pathway in Figure 1-1 (Eaton, 1996). Anaerobic pathways can employ CoA-
thioesters and they destabilize the ring by reduction, followed by hydrolytic ring opening as
shown for the benzoyl-CoA pathway in Figure 1-1 (Carmona et al., 2009; Fuchs, 2008). In both
cases, so called “lower pathways” further metabolize ring fission products so that they can enter
the Krebs cycle.

13
Figure 1-1. Comparison of typical aerobic, anaerobic and hybrid pathways with the proposed ipf pathway. Aerobic para-cumate (Cmt) pathway as described by (Eaton, 1996). Anaerobic benzoyl-CoA (Bzd) pathway as described by (Carmona et al., 2009). Hybrid phenylacetyl-CoA (Paa) pathway as described by (Teufel et al., 2011). Ipf pathway, including the putative 1,2-cis-diol-1,2-dihydroxy-ibuprofen-CoA as proposed by (Murdoch, 2011). Substrates receiving oxygen addition are indicated. Further catabolism funnels metabolites into the Krebs cycle.

14
Hybrid pathways were initially found in microaerophilic bacteria and may be useful
under fluctuating anoxic/oxic conditions because the ring opening step is non-oxygenolytic
(Fuchs, 2008). These hybrids typically use CoA-ligation and ring epoxidation to destabilize the
aromatic ring and open the ring via hydrolytic ring cleavage, as shown for the phenylacetyl-CoA
pathway in Figure 1-1 (Fuchs, 2008; Rather et al., 2010; Teufel et al., 2011; Teufel et al., 2010).
The recently described ipf pathway combines several steps from anaerobic and aerobic pathways
to produce a unique hybrid (Figure 1-1) (Murdoch, 2011). As with the well-studied anaerobic
and hybrid pathways, the ipf pathway begins with CoA-ligation. The second step is proposed to
be dihydroxylation of the aromatic ring, as is seen in aerobic pathways. The next step is
completely novel for aromatic acid metabolism, however, and results in isobutylcatechol
production in a manner analogous to acetate removal during the metabolism of fatty acids.
Finally, isobutylcatechol enters the lower pathway through oxygenolytic ring cleavage, which is
typical in aerobic pathways but not found in anaerobic or hybrid aromatic acid pathways.
1.2.1. Aerobic pathways
Aerobic microbes use upper pathways to oxidize aromatic rings and form dihydroxylated
intermediates such as catechol, gentisate, and protocatechuate (Diaz, 2004). The process begins
with mono- or di-hydroxylation, which is often performed by a multicomponent enzyme
complex composed of a terminal oxygenase as well as a ferredoxin and a ferredoxin reductase
for the transfer of electrons from NAD(P)H (Harayama et al., 1992). I will limit my discussion
to pathways producing catechol. If the initial substrate is a phenol then a catechol is produced
via monohydroxylation, otherwise non-hydroxylated compounds are dihydroxylated to produce a

15
cis-dihydrodiol which is rearomatized by a dehydrogenase to produce a catechol (Harayama et
al., 1992). Catechols are further catabolized via ortho or meta cleavage (MC) pathways,
beginning with the cleavage of an aromatic C-C bond either between the hydroxyls (ortho) or
adjacent to them (meta), respectively (Diaz, 2004). MC is performed by a ring cleavage
dioxygenase with a Fe2+ active site and it cleaves a variety of substituted and non-substituted
catechols (Harayama et al., 1992). The MC product is channeled down one of two lower
pathway branches, depending on its substitution (Figure 1-2). Cleavage products of meta-
substituted catechols are acted on by the hydrolytic branch, whereas para- and non-substituted
catechols are acted on by the 4-oxalocrotonate branch (Bayly and Dagley, 1969; Sala-Trepat and
Evans, 1971). Both pathways converge on a (substituted)2-hydroxymuconic semialdehyde
before they are funneled into the Krebs cycle (Bayly and Dagley, 1969; Powlowski and Shingler,
1994; Sala-Trepat and Evans, 1971). Ortho cleavage is performed by a dioxygenase that uses a
Fe3+ active site and this pathway is often used to metabolize chlorocatechols because it is not
sensitive to a suicidal oxidative deactivation by the catechol substrate (Harayama et al., 1992).

16
Figure 1-2. The Meta Cleavage Pathway. The hydrolytic branch is used for the metabolism of meta substituted catechols and the 4-oxalocrotonate branch is used for non- and para-substituted catechols. The enzyme names are as follows: catechol 2,3 dioxygenase, C23O; hydroxymuconic semialdehyde hydrolase, HMSH; hydroxymuconic semialdehyde dehydrogenase, HMSD; 4-oxalocrotonate isomerase, 4OI; 4-oxalocrotonate decarboxylase, 4OD; 2-oxopent-4-dienoate hydratase, OEH; 4-hydroxy-2oxalovalerate aldolase, HOA; and plant-like ferredoxin, PLFeDox. Pathways adapted from Bayly and Dagley 1969, Powlowski and Shingler 1994, and Sala-Trepat and Evans 1971.

17
1.2.1.1. Meta cleavage activity, deactivation, and reactivation
MC dioxygenases have an active site consisting of a non-heme ferrous iron (Fe2+) and the
only co-substrate for catechol cleavage is dioxygen (Harayama et al., 1992; Vaillancourt et al.,
2002). The MC dioxygenase is sensitive to oxidative deactivation, which can occur during
catalysis (Figure 1-2) (Vaillancourt et al., 2002). It was originally believed that the mechanism
of deactivation by halogenated catechols was covalent acylation by the reactive acyl halide MC
product (Bartels et al., 1984). However, presently, it is believed that the mechanism for
deactivation by non-, halo-, and alkyl- substituted catechols is through oxidation of the ferrous
iron in the active site (Cerdan et al., 1994; Vaillancourt et al., 2002). The mechanism for catechol
cleavage involves a superoxide intermediate, facilitated by Fe2+, and it can inadvertently oxidize
the iron, deactivating the enzyme and resulting in the release of the unmodified catechol
substrate (Vaillancourt et al., 2002). Halocatechols are the most potent deactivators however
para-substituted catechols are potent enough to pose a challenge for growth on compounds
catabolized via these intermediates. MC activity can be limited sufficiently by 4-methylcatechol
to prevent growth; however larger ethyl or butyl para-substituents are even more challenging
growth substrates (Bird and Cain, 1974; Polissi and Harayama, 1993; Ramos et al., 1987). While
the deactivated enzyme can be reactivated by ferrous iron or reductant (Nozaki et al., 1968),
some microorganisms challenged by deactivation employ a plant-like ferredoxin (PLFeDox)
(Polissi and Harayama, 1993; Powlowski and Shingler, 1994).
It is believed that PLFeDox enzymes transfer an electron to the oxidized iron in the active
site of the MC enzyme (Hugo et al., 1998). PLFeDoxs, like other ferredoxins, possess a [2Fe-2S]

18
redox center. However, the PLFeDox redox center is bound to the polypeptide with four
cysteines and other ferredoxins use two cysteines and two histidines (Park et al., 2002). Other
ferredoxins cannot replace the PLFeDox in reactivating MC enzymes (Hugo et al., 1998). In
addition, activity of the PLFeDox can be specific to the MC enzyme in their pathway. When
using MC enzymes from the chlorobenzene (cbz) and xylene (xyl) pathways, the PLFeDox CbzT
was able to reactivate the MC enzyme XylE however XylT (the xyl PLFeDox) was unable to
reactivate CbzE (Tropel et al., 2002).
1.2.2. Toxicity during the degradation of aromatic hydrocarbons
Research over the past decade has demonstrated that the catabolism of aromatic
hydrocarbons can be toxic to the microorganisms that degrade them. Oxidized aromatic
intermediates in degradation pathways are often more toxic than the parent compound. This is
the case for dihydrodiol, dihydroxy (catechol), and spontaneously produced monohydroxy
products of biphenyl (bph) and PCB degradation by the bph pathway (Camara et al., 2004;
Hiraoka et al., 2002; Yamada et al., 2006). A number of toxic effects have been measured in
response to these metabolites, including: decreased cell viability (Camara et al., 2004), oxidative
stress (Yamada et al., 2006), cell lysis (Camara et al., 2004), and impairment of cell-cell
separation during division (Hiraoka et al., 2002).
Oxidative stress is one mechanism of toxicity in naphthalene (nah) and bph pathways, as
seen by elevations in levels of reactive oxygen species (ROS) and oxidized lipids (Chavez et al.,
2004; Kang et al., 2006; Yamada et al., 2006). The hydroxylated metabolites may induce

19
oxidative stress through interactions with metals. Heavy metals allow catechols to undergo
redox cycling and produce other ROS, including superoxide, which can trigger chain reactions of
lipid peroxidation, inactivate proteins, and alkylate and generate breaks in DNA (Schweigert et
al., 2001). The stress endpoint can be alleviated by supplementing the media with antioxidants
or by using rich media (Kang et al., 2006; Lee et al., 2006; Yamada et al., 2006). During the
metabolism of naphthalene, oxidative stress enzymes can be up-regulated and the engineered
overexpression of antioxidant enzymes such as ferredoxin reductase, alkyl hydroperoxide
reductase, and superoxide dismutase has improved growth (Kang et al., 2006; Kang et al., 2007).
Hydrocarbons also have a high affinity for the cell membrane and will partition into it
and alter its structure and function. Cyclic hydrocarbons induce membrane swelling, leakiness,
and compromise proton gradients, with greater effects seen for chemical species with a high Kow
(Sikkema et al., 1994). Oxidized aromatic species such as phenol and catechol have also been
shown to disrupt the proton gradient across membranes through the mechanism of uncoupling
(Escher et al., 1999). It is unclear to what extent the membrane effects observed during
degradation are due to oxidative stress, however, it is clear that membrane perturbations are
induced by both oxidized intermediates and parent compounds (Camara et al., 2004; Kim et al.,
2001; Rosas et al., 1980; Sikkema et al., 1992).
1.2.3. Strain Ibu2 and the ipf pathway
Sphingomonas sp. strain Ibu2 was isolated from the Ithaca WWTP for its ability to use
IPF as a sole carbon source (Murdoch and Hay, 2005). While characterizing this pathway,

20
several strains were produced that were used for this study (Murdoch, 2011). A fosmid library
was constructed using DNA from strain Ibu2 and a clone, E. coli strain 3G7, was selected for
further evaluation due to its accumulation of catechols, an observation indicating a functional
upper pathway. Loss of function mutants were identified in additional libraries made by random
transposon mutagenesis of strain 3G7. The ipf degradation pathway was proposed by Murdoch
and the upper pathway is described in section 1.2 and pictured in Figure 1-1 (Murdoch, 2011).
Isobutyl catechol and two MC pathway metabolites have been identified although the rest of the
lower pathway has not been characterized.
Strain Ibu2 is able to metabolize IPF, 2-(4-tolyl)propionic acid [2(4T)PA], 4-tolylacetic
acid (4TAA), 3-tolylacetic acid (3TAA), 2-phenylpropionic acid (2PPA), and phenylpropionic
acid (PAA) (see Figure 1-3) to MC products, however further metabolism has not been measured
(Murdoch and Hay, 2005). Strain Ibu2 can only use IPF, 2(4T)PA, and 3TAA as sole carbon
sources, although 4TAA appears to serve as a co-substrate for growth in the presence of IPF
(Murdoch and Hay, 2005). 3TAA will not be discussed further in my dissertation. I chose to
focus on the para- and non-substituted analogs because they are expected to be metabolized via
the same enzymes; given what we know about MC pathways, the non-substituted 2PPA and
PAA and the para-substituted IPF, 2(4T)PA, and 4TAA should all be metabolized by the same
4-oxalocrotonate branch enzymes in the lower MC pathway (Figure 1-2). As will be discussed in
Chapter 3, the range of substrates that support growth of strain Ibu2 does not follow the
paradigm commonly observed for aerobic pathways employing MC. Both Chapters 3 and 4 seek
to shed light on possible reasons for this unexpected behavior.

21
Figure 1-3. Ibuprofen (IPF) and analogs metabolized by strain Ibu2: 2-(4-tolyl)propionic acid [2(4T)PA], 4-tolylacetic acid (4TAA), 3-tolylacetic acid (3TAA), 2-phenylpropionic acid (2PPA), and phenylacetic acid (PAA).

22
1.3. Conclusion
The work presented in this dissertation covers several evolving topics in the field
of emerging contaminants. First, a base-line study of emerging and traditional contaminants in
the Lower Hudson River of New York State is described in Chapter 2. Endocrine bioreporters
are employed to measure the potential of the samples to have a biological effect, while analytical
chemistry is employed for the identification of chemical species. Next, two studies are presented
on the degradation of arylacetic acids by strain Ibu2. The focus is on identifying factors that are
responsible for determining the range of arylacetic substrates that support growth for this
organism. Two approaches are used: 1) the characterization of MC deactivation and 2) toxicity
incurred during degradation.

23
CHAPTER 21
Occurrence of traditional and emerging contaminants and endocrine activity in water
samples from HRECOS stations on the Lower Hudson River, New York
Abstract
We analyzed nine water samples taken from three locations during three seasons from the
Lower Hudson River for 82 common pollutants, and detected 25 of them using gas
chromatography-mass spectrometry. Caffeine, a common indicator of human waste, was
detected at an average of 186 ng/L. 1,3,4,6,7,8-hexahydro-4,6,6,7,8,8-hexamethylcyclopenta-γ-2-
benzopyran (HHCB), a musk that is a bioaccumulative, persistent endocrine disruptor, was
detected at an average of 80 ng/L. Several other compounds including waste indicators, personal
care products, pesticides, flame retardants, detergents, and aromatic hydrocarbons were detected,
but at levels below our limits of quantification (LOQs). Six water samples were also assessed
for endocrine activity using the in vitro Chemical Activated LUciferase eXpression (CALUX)
assay. All samples activated the estrogen receptor (94-701 pg 17b-estradiol Eq/L), two samples
activated the progesterone receptor (1.6-4.08 ng/Org2058 Eq/L), one sample activated the
androgen receptor (1.15 ng DHT Eq/L), but no glucocorticoid receptor activation was observed.
The compounds and endocrine activities detected are discussed with respect to their potential
fates and biological impacts.
1This was a collaborative project in which CALUX testing was coordinated under Peter A. Behnisch at BioDetection Systems, Science Park 406, 1098 XH Amsterdam, The Netherlands. The analytic chemistry was coordinated by Patrick J. Phillips U. S. Geological Survey, Albany, NY, United States and conducted at U. S. Geological Survey National Water Quality Laboratory, Denver, CO, United States.

24
2.1. Introduction
Throughout the world, natural aquatic environments are used as receiving bodies for
treated and untreated waste streams. For several decades we have been aware of the presence
and potential threat of industrial and agricultural pollutants. However, in the past 20 years
additional pollutants such as pharmaceuticals and personal care products (PPCPs), hormones and
steroids, flame retardants, anticorrosives, and plasticizers have become an emerging concern
(Daughton and Ternes, 1999). Some emerging contaminants, including detergent breakdown
products, synthetic musks, pharmaceuticals, and antimicrobials, have been found to accumulate
in edible wild fish and crustacean populations (Ferrara et al., 2008; Ramirez et al., 2009). They
are ubiquitous and have been routinely been found in major waterways (Sacher et al., 2008;
Yoon et al., 2010), coastal regions (Dougherty et al., 2010; Singh et al., 2009), groundwater
(Barnes et al., 2008), and even in national parks (Camacho-Munoz et al., 2010; Smith, 2004).
While wastewater treatment plant (WWTP) effluents are thought to be the major source of these
pollutants (Metcalfe et al., 2003b; Yoon et al., 2010), there are several other recognized sources
including residential septic systems (Dougherty et al., 2010), the land application of biosolids
(Sabourin et al., 2009), agricultural livestock wastes (Fisher and Scott, 2008; Shao et al., 2009),
landfill leachate (Buszka et al., 2009), and waste from pharmaceutical production (Fick et al.,
2009; Phillips et al., 2010b).
Many of these emerging pollutants are biologically active and have been shown to have
endocrine disrupting activity (Kloas et al., 1999; Schreurs et al., 2005; Sonneveld et al., 2005;
Sonneveld et al., 2011). Endocrine functions as diverse as sexual differentiation and energy

25
metabolism can be disrupted through genomic and nongenomic mechanisms; the effects can be
direct or induced indirectly by altering the storage and metabolism of hormones (Casals-Casas
and Desvergne, 2011; Watson et al., 2007). The United States acknowledged the importance of
understanding the endocrine disruption potential of chemicals on a regulatory level more than 15
years ago with the creation of the Endocrine Disruptor Screening Program (Fenner-Crisp et al.,
2000). In vivo assays, such as the Allen-Doisy (estrogens), Hershberger (androgens), and
McPhail (prostaglandins) assays are time consuming and require the use and disposal of animals
(Allen and Doisy, 1923; Gray et al., 2005; McPhail, 1934). Alternatively, in vitro assays are
being increasingly used to measure chemical interference with hormone signaling; however,
progress in developing standardized tests amenable to high throughput analyses has been slow
(Schmidt, 2009). One such in vitro assay is Chemical Activated LUciferase gene eXpression
(CALUX), which uses cell lines engineered to express a luciferase reporter gene when specific
hormone response elements are transactivated (Sonneveld et al., 2005). CALUX assays produce
results that are comparable to other in vitro assays and they are good predictors of in vivo
responses (Allen-Doisy r2=0.87, Hershberger r2=0.46, McPhail r2=0.85) (Sonneveld et al., 2011;
Sonneveld et al., 2006).
In this work we sought to characterize chemical pollutants and endocrine disrupting
activities in the Lower Hudson River of New York State (NYS). While the upstate and Finger
Lakes regions of NYS are primarily rural, the population dramatically increases as one
approaches New York City (NYC). The Lower Hudson River Basin spans this region of
increasing population density and is used as a drinking water source for many adjacent
municipalities including 75,000 people in the greater Poughkeepsie area

26
(http://www.pokwater.com/new/) while also receiving the wastewater discharge from 128
communities (NYSDEC, 2004). The water quality of the Hudson River has been most
extensively studied with respect to polychlorinated biphenyls and heavy metals, but also
surfactants (Baldigo et al., 2006; Butcher and Garvey, 2004; Knutson et al., 1987; Li and
Brownawell, 2010; Sinclair et al., 2006). There are, however, a large number of chemicals from
anthropogenic wastes for which the Hudson River has not been evaluated in the peer-reviewed
literature. The base-line study presented here employed both chemical analysis for 82 chemicals
and in vitro hormonal activity bioassays for androgenic, estrogenic, glucocorticoid, and
progestogenic activities in the Lower Hudson River.

27
2.2. Materials and Methods
2.2.1. Site selection
Sampling for the current study was conducted at field sites established by the Hudson
River Environmental Conditions Observing System (HRECOS), where automatic samplers
monitor general water quality and meteorological data (www.ecostudies.org/hrecos/). The
northern most site was just south of Albany, NY at Schodack Island State Park, the next site was
at the Environmental Center at Norrie Point State Park, and the southern-most site was at
Piermont Pier, just north of NYC (Figure 2-1 and Table 2-1).
2.2.2. Sample collection
Samples were collected during the fall, winter, and spring (Table 2-1), at peak ebb flow
times predicted by tidal current tables on the National Oceanic and Atmospheric Administration
website http://co-ops.nos.noaa.gov/index.shtml. Samples for chemical analysis were collected in
3L Teflon bottles with a weighted cage sampler while samples for CALUX analysis were
collected in 1L organic-free amber glass bottles using a pole sampler. Samples were collected
from depths of approximately 0.5-1.5 meters below the surface at piers or docks then
immediately stored on ice in the dark. One field blank and one replicate were collected for each
sampling season; the field blank was prepared by pouring HPLC grade water into the previously
described collection vessel at the field site. An additional lab blank was prepared for CALUX
using HPLC grade water.

28
Figure 2-1. The Lower Hudson River in NY State with Hudson River Environmental Conditions Observing System (HRECOS) sites indicated with stars and sampling sites indicated with arrows. Figure modified from www.ecostudies.org/hrecos/overview.html Table 2-1. Geographic coordinates and sampling dates for Lower Hudson River sampling sites.

29
2.2.3. Chemical analysis
Water samples were filtered through 0.7µM glass-fiber filters, collected in organic-free
amber glass bottles and shipped on ice to the United States Geological Survey (USGS) National
Water Quality Lab in Denver, CO for analysis of 82 anthropogenic wastewater compounds and
pharmaceuticals as described elsewhere (Phillips et al., 2010a; Zaugg et al., 2007). Briefly,
surrogates (bisphenol-A, caffeine-13C, decafluorobiphenyl, and fluoranthene-d10) were added to
samples that were then loaded on to OASIS-HLB-SPE cartridges (Waters Inc., catalog #
186000115), and eluted with dichloromethane:diethyl ether (4:1). Samples were analyzed by
capillary gas chromatography-mass spectrophotometry (Agilent Technologies model 6890 GC
and model 5973 MS), which was operated under full scan conditions (50-450 atomic mass units
at 1 scan/sec) using electron impact ionization. Sample injection and oven temperature programs
varied for the two methods used (Phillips et al., 2010a; Zaugg et al., 2007).
2.2.4. CALUX analysis
Winter and spring samples were analyzed for endocrine activity with CALUX reporters
for transcriptional activation of HREs using the U2OS human cell line as described elsewhere
(Van der Linden et al., 2008). Briefly, water samples were filtered through 0.7µM glass-fiber
filters, collected in organic-free amber glass bottles, and stored in the dark at 4oC for up to five
days. 500mL of filtrate was extracted with ethyl acetate (polarity index 4.4), resuspended in
dimethyl sulfoxide (DMSO) and shipped to BioDetection Systems in the Netherlands for

30
analysis. This extraction procedure has been shown to have good recovery of many estrogens,
androgens and glucocorticoids (68-120%) and slightly lower recovery of progestagens (40-89%)
(Van der Linden et al., 2008). CALUX cells were grown in 96-well plates and incubated with
DMSO extracts, lysed, and analyzed for luciferase activity with a luminometer (Lucy 2, Anthos,
Austria). Extracts were evaluated for transactivation of androgen (ARE), estrogen (ERE),
glucocorticoid (GRE), and progesterone (PRE) response elements, and results were reported as
equivalents (Eq) of dihydrotestosterone (DHT), 17b-estradiol (E2), dexamethasone (Dex), or
16alpha-ethyl-21-hydroxy-19-nor-4-pregnene-3,20-dione (Org2058) activity respectively. Limits
of detection were 0.46 ng DHT Eq/L, 0.17 ng Org2058 Eq/L, 6.3 ng Dex Eq/L, and 0.022ng E2
Eq/L.
2.2.5. Chemicals
Ethyl acetate, hexanes, methanol, and submicron-filtered HPLC grade water from Fisher
Scientific and DMSO from Sigma-Aldrich were used for sample extraction. All other chemicals
are described in the references for chemical and CALUX analysis.
2.2.6. Statistical analysis
A one-way analysis of variance was conducted using the StatPlus package for Excel to
determine if there were any significant differences in the seasonal or site-specific occurrence of
the chemical classes shown in Table 2-2.

31
2.3. Results and Discussion
2.3.1. Chemical analysis
Caffeine and HHCB were detected at levels above their limits of quantification (LOQs),
while 23 additional compounds were detected below their LOQs (Table 2-2). The detected, but
not quantifiable, compounds included waste indicators, PPCPs, detergents, flame-retardants,
plasticizers, pesticides, and aromatic hydrocarbons (Table 2-2). While twenty-seven of the 60
detects were at Piermont Pier, which is close to the discharge outlets of two municipal WWTPs
(Orangetown Sewer District #2 and Rockland County Sewer District #1), the frequencies of
detection with respect to both location and season were highly variable. Aromatic hydrocarbons
were the only group of compounds whose detection showed a statistically significant seasonal
difference (P=0.016) in the mean number of detects, since they were only detected during the
spring sampling.

32
Table 2-2. Chemicals detected in Lower Hudson River samples. Quantitative results are those detected above the limits of quantification (LOQ). Total detects and shaded totals and subtotals include both quantitative results and qualitative estimates below the LOQ. Abbreviations are as follows: 7-acetyl-1,1,3,4,4,6-hexamethyl-1,2,3,4-tetrahydronaphthalene (AHTN); 1,3,4,6,7,8-hexahydro-4,6,6,7,8,8-hexamethylcyclopenta-γ-2-benzopyran (HHCB); 4-t-octylphenol (4t-OP); 4-t-octylphenol monoethoxylate (4t-OPE1); 4-t-octylphenol diethoxylate (4t-OPE1); tris (2-butoxyethyl) phosphate (TBEP); tris (2-chloroethyl) phosphate (TCEP); and tris (dichloroisopropyl) phosphate (TDCP).

33
2.3.1.1. Waste indicators
Caffeine was detected in every sample, and was found above the LOQ (>60 ng/L) in six
samples; the average detected concentration was 186 ng/L and the maximum was 370 ng/L.
Caffeine is ubiquitous, with concentrations in rivers and streams ranging from 13-5,700 ng/L
(Kolpin et al., 2002; Shala and Foster, 2010; Singh et al., 2009) and in drinking water sources at
up to 122 ng/L (Palmer et al., 2008; Standley et al., 2000). Acute toxicity is low (effective
concentration for Hydra attenuata morphology, EC50 >100 mg/L) (Quinn et al., 2008), however,
long-term exposure (28 days) can reduce activity and elevate the startle response of tadpoles
(Rana pipiens) exposed to environmentally relevant concentrations (600 ng/L) of caffeine
(Fraker and Smith, 2004).
Cotinine, a nicotine metabolite was detected below the LOQ (<600 ng/L) once. Cotinine
has been found at concentrations below 5 and up to 900 ng/L (Hua et al., 2006; Kolpin et al.,
2002; Metcalfe et al., 2003b).
Skatole (3-methyl-1H-indole) was detected below the LOQ (<36 ng/L) three times. It is
a biodegradation product of tryptophan and can be used as an indicator of fecal waste (Schussler
and Nitschke, 1999), however, it is also a component of coal tar and has been used as a
fragrance. The USGS has also detected skatole at levels below their LOQ of 1µg/L in rivers and
streams across the U.S. (Lee et al., 2008; Oblinger et al., 2007; Smith, 2004).

34
2.3.1.2. Personal care products & detergents
The polycyclic musk HHCB was detected in four samples at an average concentration of
80 ng/L and a maximum of 130 ng/L. 7-acetyl-1,1,3,4,4,6-hexamethyl-1,2,3,4-
tetrahydronaphthalene (AHTN) was detected below the LOQ (<28 ng/L) three times. These
musks were only detected during fall and winter sampling. Failure to detect them during the
summer may have been due to their greater susceptibility to photodegradation at the surface of
water bodies during sunny seasons (Buerge et al., 2003). HHCB and AHTN are the most
commonly detected polycyclic fragrances and thus have also been used as waste indicators
(Standley et al., 2000). They have been found at 5-1,590 ng HHCB/L and 2-530 ng AHTN/L in
rivers and lakes worldwide (Buerge et al., 2003; Dsikowitzky et al., 2004b; Fromme et al., 2001;
Sumner et al., 2010; Zhang et al., 2008), and up to 67 ng HHCB/L and 33 ng AHTN/L in
drinking water sources across Canada and the US (Standley et al., 2000). They are of potential
concern as they are persistent, bioaccumulative, and have been reported to have endocrine
effects. While microbes adapted to grow on HHCB and AHTN have been shown to rapidly
degrade these compounds in laboratory experiments (Balk and Ford, 1999a), little to no
degradation was reported for HHCB when examined in oceans and large lakes (Andresen et al.,
2007). They bioaccumulate in fish and eels in the wild, with reported wet weight
bioaccumulation factors (BAFs) of 20-862 (HHCB) and 40-1069 (AHTN) (Fromme et al., 2001;
Gatermann et al., 2002). In a survey of PPCPs in commonly eaten wild-caught fish across the
U.S., Ramirez and collogues found HHCB (100-2,100 ng/g) and AHTN (21-290 ng/g) in fish
fillets at levels which exceeded other detected PPCPs by up to two orders of magnitude (Ramirez

35
et al., 2009). AHTN and HHCB may have a variety of endocrine effects as seen by both agonism
and antagonism of the estrogen receptor, and antagonism of the progesterone receptor in vitro
(Schreurs et al., 2002; Schreurs et al., 2005). However, the lowest observed effect
concentrations for growth, early life stage, and estrogenic effects on aquatic species (ranging
from 35-500 µg AHTN/L and 68-500 µg HHCB/L) are at least two orders of magnitude higher
than concentrations that we found (Balk and Ford, 1999b; Yamauchi et al., 2008).
Benzophenone, a fragrance and UV-filter, was detected below the LOQ (<80 ng/L) four
times. Others have found benzophenone in rivers and streams at 23-130 ng/L (Kawaguchi et al.,
2008; Yoon et al., 2010). Benzophenone may have endocrine effects as it has been shown to
induce cell proliferation and displace 17-beta-estradiol from the rainbow trout ER (Jobling et al.,
1995) and chemically related benzophenones have been shown to have various estrogenic effects
on fathead minnows in vivo at concentrations ranging from 0.4-5,000 µg/L (Kunz and Fent,
2009; Weisbrod et al., 2007).
Camphor, a flavoring agent and component of topical creams and moth repellents, was
detected below the LOQ (<44 ng/L) five times. Methyl salicylate, a component of wintergreen
oil and a plant defense hormone, was detected below the LOQ (<44 ng/L) twice. Neither of
these compounds have received much attention in the peer-reviewed literature, however they
have both been used as indicators of anthropogenic waste by the USGS and have been detected
in several studies, although at levels that were below the LOQs (<0.5-1 µg/L) (Lee et al., 2008;
Oblinger et al., 2007; Smith, 2004). Camphor has also been qualitatively identified in the Lippe
River of Germany (Dsikowitzky et al., 2004a).

36
Three metabolites of alkyl ethoxylate detergents were detected; 4-t-octylphenol (4t-OP)
was detected below the LOQ (<140 ng/L) twice, while both 4-t-octylphenol monoethoxylate (4t-
OPE1) and 4-t-octylphenol diethoxylate (4t-OPE2) were detected below their LOQs (<1,000
ng/L) once. All of the detections for these compounds were at Piermont Pier. These detergent
metabolites are commonly found in rivers and streams at levels ranging from 1-1,585 ng 4t-
OP/L, 0.5-2,000 ng 4t-OPE1/L, and 2-1,574 ng 4t-OPE2/L (Arditsoglou and Voutsa, 2009;
Bennie et al., 1997; Kolpin et al., 2002; Loos et al., 2007; Loyo-Rosales et al., 2010; Matamoros
et al., 2010; Yoon et al., 2010), and have been found in the edible portions of seafood at
concentrations ranging from 0.2-32.3 ng/g (Ferrara et al., 2008). While little investigation has
been done on the toxicity of the ethoxylates, 4t-OP has been shown to have estrogenic effects.
4t-OP is an endocrine disruptor that mimics estrogen (Fu et al., 2007; Jobling and Sumpter,
1993) and has effects such as reduced fecundity, impaired development, and modified sex ratios
in aquatic species at concentrations ranging from 2-20 µg/L (more than 10 times higher than our
LOQ) (Gronen et al., 1999; Isidori et al., 2006; Kloas et al., 1999).
2.3.1.3. Flame retardants and plasticizers
We tested for five organophosphorous compounds, of which the halogenated ones are
primarily used as flame retardants, while the non-halogenated ones are used as plasticizers
(Andresen et al., 2004). Tris (2-butoxyethyl (TBEP)), (2-chloroethyl (TCEP), and
(dichloroisopropyl (TDCP)) phosphates were detected below their LOQs three, one, and two
times respectively, and all but one of the detections were in the fall (LOQs: <800 ng TBEP/L,

37
<100 ng TCEP/L and <160 ng TDCP/L). Other investigators have detected these compounds in
surface waters at concentrations ranging from 10-1,236 ng/L (Andresen et al., 2004; Fries and
Puttmann, 2003; Kolpin et al., 2002; Martinez-Carballo et al., 2007; Yoon et al., 2010). TCEP
and TDCP, which are chlorinated, have been reported to be persistent in environmental waters,
however, TBEP has a half life of <14 days (Andresen et al., 2007). Interestingly, TBEP was
detected more often in this study (three times) than either TCEP (one time) or TDCP (two times).
TCEP and TDCP have also been found in aquatic species and in human breast milk more often
and at higher concentrations than TBEP (Sundkvist et al., 2010). In vitro studies with TCEP
have shown little to no estrogenic, cytotoxic, genotoxic, or mutagenic effects (Follmann and
Wober, 2006) although increases in p21 and p27 cell cycle regulatory proteins have been
observed (Ren et al., 2008). In vivo assays have reported slight fertility decreases and
carcinogenicity, but low bioconcentration factors (TCEP 0.9 & TDCP 3-5) (Sala et al., 1982;
WHO, 1998).
2.3.1.4. Pesticides
Chloroxylenol and triclosan are antimicrobials commonly used in soaps as well as other
personal care and consumer products. Chloroxylenol was detected below the LOQ (<80 ng/L)
once. It has previously been found in rivers at concentrations less than 30 ng/L and as high as
254 ng/L (Kasprzyk-Hordern et al., 2009). There is very little in the peer-reviewed literature
about chloroxylenol; generally speaking it is essentially non-toxic to mammals but
comparatively more toxic to fish, with an acute LC50 for rainbow trout of 760 µg /L (Guess and
Bruch, 1986; USEPA, 1994). Triclosan was detected below the LOQ (<200 ng/L) once. Others

38
have reported triclosan in surface waters at concentrations ranging from 1-2,300 ng/L (Kolpin et
al., 2002; Matamoros et al., 2010; Shala and Foster, 2010; Yoon et al., 2010). Triclosan is
bioaccumulative (BAF=500-4,157 wet weight), and aquatic species have EC50s ranging from
0.23-399 µg/L for endpoints such as growth, survival, activity levels, and endocrine disruption
(Coogan and La Point, 2008; DeLorenzo et al., 2008; Fraker and Smith, 2004; Oliveira et al.,
2009; Orvos et al., 2002; Raut and Angus, 2010). Triclosan is also an inhibitor of p450s and
phase two metabolism enzymes such as sulfo- and glucuronosyl transferases, so it could
negatively impact the detoxification of other pollutants (Hanioka et al., 1996; Wang et al., 2004).
Murray and colleagues have suggested that triclosan is a high priority pollutant and suggest an
acceptable daily intake value of 190 ng/Kg/day (Murray et al., 2010). Since triclosan toxicity
can occur at concentrations close to our LOQ, further analytical sensitivity should be sought after
in future studies.
Metolachlor, a chloroacetamide herbicide, was detected below the LOQ (<80 ng/L) three
times. It has a half life of 12-20 days in surface water (Mazanti et al., 2003), however, it is a
frequently detected pesticide (0.43-8 µg/L) that is found at the highest concentrations after
agricultural application (Lehotay et al., 1998; Vryzas et al., 2011; Whitall et al., 2010).
Metolachlor has low acute toxicity to aquatic species (LC50s 6-80 mg/L) however chronic
exposure has been shown to interfere with responses to chemical stimuli (threats and food) and
reproductive fitness at levels (10-80 µg/L) that are close to environmentally relevant
concentrations (Cook and Moore, 2007; Liu et al., 2006b; Wan et al., 2006; Wolf and Moore,
2002). Our detections were at least three orders of magnitude lower than those for which effects
have been reported.

39
Piperonyl butoxide, a pesticide synergist, was detected below the LOQ (<80 ng/L) once.
Other investigators have detected piperonyl butoxide in surface waters at concentrations ranging
from 0.3-15,000 ng/L, with the highest concentrations occurring after insecticide applications
(Amweg et al., 2006; Schleier et al., 2008; Zulkosky et al., 2005). The toxicity to aquatic species
is low with LC50s ranging from 530-16,000 µg/L (Ankley and Collyard, 1995; Erickson et al.,
1988). Piperonyl butoxide is a cytochrome p450 inhibitor (Weinstein and Garner, 2008). This
activity decreases biotransformation leading to elimination and can increase the bioaccumulation
of co-occurring pollutants, including polycyclic musks and aromatic compounds, both of which
were detected in this study (Artola-Garicano et al., 2003; Weinstein and Garner, 2008).
2.3.1.5. Aromatic hydrocarbons from oil and coal
Seven compounds likely to be of coal and oil origin were detected below their LOQs: 1-
methylnaphthalene (<22 ng/L), 2-methylnaphthalene (<36 ng/L), cumene (<300 ng/L),
fluoranthene (<24 ng/L), naphthalene (<40 ng/L), phenanthrene (<32 ng/L), and pyrene (<42
ng/L). Interestingly, all 13 detections were during the spring sampling (p=0.016). Although
there was no significant correlation between sampling site and the number of detections, the
majority of the detections were at Schodack Island. Other investigators have detected
phenanthrene, pyrene, naphthalene, and flouranthene at concentrations ranging from 7-1,200 ng
/L in waters from Spain and the United States (Kolpin et al., 2002; Matamoros et al., 2010).
PAHs are either petrogenic (coal & oil) or pyrogenic (combustion) in origin and in some cases
sources can be identified based on the proportions of different PAHs present (Pies et al., 2008;

40
Yim et al., 2005). However, since our detections were below the LOQs no source determination
could be made. Bioaccumulation of these compounds can be substantial, with wet weight BAFs
for aquatic species ranging from 50-4,288 for fluoranthene, phenanthrene, pyrene, naphthalene,
and 2-methylnaphthalene (Jonsson et al., 2004; Okay and Karacik, 2008). PAHs have many
toxicological effects, but the one of most concern is cancer; carcinogenesis and mutagenesis can
occur through the action of the parent compounds or by biologically or photo-activated reactive
species (Arfsten et al., 1996; Liu et al., 2008; Skupinska et al., 2004). None of the PAHs that we
detected are recognized carcinogens; however naphthalene, flouranthene, phenanthrene, and
pyrene are listed amongst the 16 PAHs on the USEPA’s priority pollutant list (Collins et al.,
1998).
2.3.1.6. Non detects & quality control
Fifty-seven compounds were not detected in Lower Hudson River samples (Table 2-3).
In addition, detections of nine analytes were censored to avoid false positives and are not
reported in Table 2-2. Decisions on censoring data were based on the results from the field
blanks and laboratory experience. Seven compounds (AHTN, HHCB, tributyl phosphate (TBP),
triphenyl phosphate (TPP), N,N-Diethyl-m-Toluamide (DEET), phenol, and 1,4-
dichlorobenzene) were detected in the field blanks below their LOQs. Phenol was not detected in
any environmental samples. HHCB, DEET and TPP were detected twice each in blanks, and the
rest of the compounds were only detected once in blanks. Two compounds (triethyl citrate and
menthol) were not detected in any blanks however experience in the laboratory with general low
level contamination prompted the censoring of estimates below 50 ng/L and 200 ng/L

41
respectively. Censoring thresholds were set above estimates of detections in the blanks and most
of the data were censored at values below their LOQs; there were exceptions with TBP (LOQ
<160 ng/L) and DEET (LOQ <60 ng/L) for which results were censored for detections below
200 ng/L, and HHCB (LOQ <52 ng/L) for which results were censored for detections below 70
ng/L for the 5/04/10 samples only.

42
Table 2-3. Chemicals tested for but not detected in Lower Hudson River samples and limits of quantification (LOQ).

43
Replicate samples were evaluated for consistency, with 28 compounds being detected at
least once in replicate samples (a total of 40 pairs of detections). All of the paired measurements
were very similar; 33 of the paired values fell within 50% of each other and seven paired values
were of non-detects with detects below their LOQ. 3-beta-coprostanol was detected below the
LOQ in a replicate sample on 11/10/09 at Piermont Pier, but was not found in the primary
sample, therefore it was not included in the results. Results reported in table 2 are for the
primary samples, not the replicates. While we tested for 21 pharmaceuticals, we did not detect
any of them (Table 2-3).
2.3.2 CALUX
Solvent extracts of all samples induced estrogenic responses, ranging from 0.094-0.701
ng E2 Eq/L, with the maximum response coming from the winter Schodack Island sample
(Figure 2-2). Both of the extracts from Piermont Pier induced progestogenic responses ranging
from 1.6-4.08 ng Org2058 Eq/L. The extract from the spring Schodack Island sample induced
an androgenic response of 1.15 ng DHT Eq/L. No glucocorticoid responses were observed in the
environmental samples and no CALUX responses were observed in the blanks.

44
Figure 2-2. Transactivation of (A) androgen, (B) estrogen, and (C) progesterone response elements in CALUX bioassay by Lower Hudson River samples from winter and spring. Activity is reported in hormone equivalents (Eq) for transactivation of the hormone response elements relative to dihydrotestosterone (DHT), 17b-estradiol (E2), or 16alpha-ethyl-21-hydroxy-19-nor-4-pregnene-3,20-dione (Org2058).
Of the compounds detected during the winter and spring sampling, six of them have been
reported to have endocrine activity: HHCB, benzophenone, 4-t-OP, flouranthene, naphthalene,
and triclosan (Fu et al., 2007; Kummer et al., 2008; Passos et al., 2009; Raut and Angus, 2010;
Schreurs et al., 2002; Schreurs et al., 2005). HHCB primarily acts as an antagonist towards the
estrogen (ER), androgen and progesterone receptors, but also has agonistic activity toward the

45
ER; the effects are specific to the cell line and variant of the receptor (Schreurs et al., 2002;
Schreurs et al., 2005). Estrogenic activity factors have been reported in E2 equivalents for
benzophenone (3X10-7) and 4t-OP (10-3) (Fu et al., 2007; Schreurs et al., 2005). Theoretically,
4t-OP could induce an effect equivalent to 0.14 ng E2 Eq/L if it were present in our samples at
our LOQ (<140 ng/L). However, 4t-OP was only detected in one of the six samples (Piermont
Pier, spring) with positive CALUX-ER results and even at the LOQ, which it did not reach,
would still not account for all the estrogenicity detected in most of our samples. Likewise,
benzophenone would only induce 0.024 pg E2 Eq/L if present at its LOQ (<80 ng/L), which is
three to four orders of magnitude below the CALUX-ER responses we measured. We cannot
account for all of the estrogenic activity detected via the CALUX assay (Figure 2-2) based on the
predicted E2 equivalents of compounds detected in our chemical analysis (Table 2-2). This may
be because our chemical analyses did not specifically target steroids and other hormonally active
compounds, as has been done by Schriks and colleagues (Schriks et al., 2010). This may also be
due to the lack of information on the endocrine effects of the compounds that we did detect. Our
CALUX results are, however, consistent with those reported for surface waters in the
Netherlands (0.18-0.5 ng E2 Eq/L, n.d.-4.5 ng Org2058 Eq/L, 0.39-1.3 ng Dex Eq/L, and n.d.-12
ng DHT Eq/L) (Van der Linden et al., 2008).
The androgenic and estrogenic activities at our field sites were lower than, but within one
order of magnitude of activity levels associated with endocrine disruption in the field. For
example, female mosquitofish (Gambusia holbrooki) were found to be masculinized in a Florida
river impacted by a pulp paper mill, where river water androgen activity levels were 6 ng DHT
Eq/L (Jenkins et al., 2001). Feminizing effects, including skewed sex ratios, elevated

46
vitellogenin production in males, impaired gonadal development, and reduced fertility are
commonly reported for aquatic species living in waters heavily impacted by WWTP effluents
(Bjerregaard et al., 2006; Jobling et al., 2009; Jobling et al., 2002; Jobling et al., 1998; Vajda et
al., 2008). For example, streams in the United Kingdom that had greater than 1 ng E2 Eq/L also
had roach (Rutilus rutilus) populations with modified sex ratios (Jobling et al., 2009; Jobling et
al., 2006). In another study, a Canadian lake dosed with 5-6 ng 17alpha-ethynylestradiol/L
caused feminization followed by near extinction of the fathead minnow (Pimenphales promelas),
along with impaired gonadal development and reduced hatching success of anurans (Rana spp.)
(Kidd et al., 2007; Park and Kidd, 2005). Thus, while we do not have specific information on
the chemical species responsible for the endocrine activity measured using the CALUX assay,
our results suggest that current levels of endocrine activity in Lower Hudson River waters may
have biological significance.

47
CHAPTER 3
Atypical meta cleavage deactivation and growth substrate range in the ibuprofen
degradation pathway of Sphingomonas sp. strain Ibu2
Abstract
Sphingomonas sp. strain Ibu2 is capable of growing on ibuprofen (IPF) and 2-(4-
tolyl)propionic acid [2(4T)PA] as sole carbon sources, but not 4-tolylacetic acid (4TAA), 2-
phenylpropionic acid (2PPA), or phenylacetic acid (PAA). Metabolism of all non-growth
substrates (4TAA, 2PPA, and PAA) resulted in inhibition of meta cleavage (MC) activity. A
gene for a MC enzyme, 4-isobutylcatechol-2,3-dioxygenase (ipfJ), was identified in a clone of
strain Ibu2. However, IpfJ was least sensitive to deactivation by non-substituted catechols,
which would be produced during catabolism of 2PPA and PAA. None of the parent compounds
up-regulated the upper pathway during catabolism and all were transformed at approximately the
same rate except for IPF, which was transformed at least 90% slower. Mutants with the ability
to grow on 4TAA were found to have lower rates of catechol formation in the upper pathway and
decreased sensitivity to MC deactivation, however no genetic changes were observed in ipfJ or
its predicted promoter region. This work demonstrates that metabolism of non-growth substrates
2PPA and PAA results in an atypical form of MC deactivation in strain Ibu2 and suggests that,
while managing intracellular catechol concentrations can play an important role in expanding the
range of growth substrates for strain Ibu2, it does not appear to account for all aspects of these
atypical results.

48
3.1. Introduction
While much is known about the biodegradation of traditional pollutants, we are just
beginning to learn about the pathways used for catabolism of emerging pollutants (Kagle et al.,
2009). Pharmaceuticals are an important class of emerging pollutants as they have biological
activity at low concentrations, and their continuous emission into the environment makes them
pseudopersistent even when biodegradable (Daughton, 2002). Wastewater treatment plants
(WWTPs) are one of the major routes through which pharmaceuticals enter the environment
(Daughton and Ruhoy, 2009). While a growing body of literature is available concerning the
removal rates of pharmaceuticals during wastewater treatment, few microbial isolates capable of
degrading pharmaceuticals have been characterized sufficiently to determine the mechanisms of
biotransformation (Kagle et al., 2009). Sphingomonas sp. strain Ibu2, is a bacterial WWTP
isolate that was selected for its ability to use ibuprofen (IPF), an NSAID and substituted
phenylacetic acid, as a sole carbon source (Murdoch and Hay, 2005). Strain Ibu2 can metabolize
a variety of arylacetic acids (AAAs), however, only a subset of these substrates serve as a sole
carbon source for growth (Murdoch and Hay, 2005).
Strain Ibu2 catabolizes IPF via the recently described ipf pathway (Murdoch, 2011). This
pathway appears to be a novel hybrid that combines the use of a CoA-AAA thioester in the upper
pathway with oxygenolytic ring cleavage in the lower pathway (as described in further detail in
section 1.2 of Chapter 1, Figure 1-1). The catecholic metabolite is processed via meta cleavage
(MC) in strain Ibu2. For MC pathways in general, meta-substituted catechols are metabolized
via the hydrolytic branch, whereas non- and para-substituted catechols are metabolized via the 4-

49
oxalocrotonate branch (Figure 1-2) (Sala-Trepat and Evans, 1971). MC enzymes are sensitive to
deactivation that can result from oxidation of the active site iron and which can occur either
during turnover of a catechol substrate or from exposure to either endogenous or exogenous
oxidants (Vaillancourt et al., 2002). Suicide substrate inhibition is most notably reported for
halogenated substrates, but has been routinely reported for alkyl para-substituted substrates as
well (Polissi and Harayama, 1993; Vaillancourt et al., 2002), and more recently for alkene meta-
substituted substrates (George et al., 2011). In some organisms, plant-like ferredoxins
(PLFeDox) are found, which are able to reductively reactivate MC enzymes that have been
oxidatively deactivated as shown in Figure 1-2 (Polissi and Harayama, 1993).
Strain Ibu2 uses the ipf pathway for the aerobic degradation of IPF and related analogs
including 2(4T)PA, 4TAA, 2PPA, and PAA (Figure 1-3) (Murdoch, 2011). Interestingly, strain
Ibu2 can use the para-substituted IPF, and 2(4T)PA as sole carbon sources, but not the para-
substituted 4TAA, or non-substituted 2PPA or PAA, even though all analogs are metabolized by
strain Ibu2 to an MC product (Murdoch and Hay, 2005). It is somewhat surprising that 2PPA and
PAA cannot support growth because compounds metabolized via substituted catechols have
typically been reported to be more difficult to degrade via MC (Polissi and Harayama, 1993;
Ramos et al., 1987).
In order for a compound to support growth, an organism must encode and produce
enzymes that can break it down into high-energy intermediates. In this study, induction of, and
maximum activity rates of the upper and lower ipf pathways were measured in an attempt to
understand strain Ibu2’s unusual growth substrate specificity. In addition, we isolated and

50
characterized mutants with the ability to use 4TAA as a sole carbon source in order to better
understand the factors limiting the wild type’s growth on this substrate.

51
3.2. Materials and Methods
3.2.1. Chemicals
Catechol, 3-methylcatechol, 4-methylcatehcol, 4-tertbutylcatehol, 3-fluorocatechol,
ibuprofen, 2-(4-tolyl)propionic acid, 4-tolylacetic acid, 2-phenylpropionic acid, and phenylacetic
acid were purchased from Acros Organics. HPLC grade methanol was purchased from Fisher
Scientific.
Table 3-1. Strains used in this study
Strain Plasmid and/or characteristics Reference E. coli EPI300 Epicentre E. coli EPI300 3G7 Fosmid pCC1FOS:30-40kb Ibu2 DNA Murdoch 2011 E. coli EPI300 4F6 Fosmid pCC1FOS:30-40kb Ibu2 DNA Murdoch 2011 E. coli EPI300 3G7 TnH5 3G7 with Tn5 transposon in ipfJ Murdoch 2011 E. coli EPI300 3G7 TnG9 3G7 with Tn5 transposon in ipfJ Murdoch 2011 E. coli JM109 pC Plasmid pBBR1MCS-3:ipfJ (forward primer
AATGGTACCGTCCATGCTGAAGTGTGGCGAAAT, reverse primer TCTGGGCCCTGTGTGGGTCAGCTTGTTGTTT)
This study
Sphingomonas sp. Ibu2 Plasmid pBBR1MCS-3:Empty Murdoch & Hay 2005
Sphingomonas sp. Ibu2 4TAA1
Mutant selected on 250mg/L 4TAA This study
Sphingomonas sp. Ibu2 4TAA2
Mutant selected on 250mg/L 4TAA This study
Sphingomonas sp. Ibu2 4TAA3
Mutant selected on 250mg/L 4TAA This study

52
3.2.2. Culturing of strains
Strain Ibu2 and derivatives were grown in liquid mineral salts media (MSM) at room
temperature with aeration by shaking at 150rpm in flasks or conical tubes. Carbon sources were
supplied at concentrations of 250 or 500 mg/L for AAAs, or 0.1% glycerol, and combined when
indicated. Escherichia coli strains were grown at 37oC in Luria Bertani (LB) broth with aeration
by shaking at 150 rpm in flasks or conical tubes, and IPF or analogs were added as described.
Derivatives of the fosmid pCC1FOS (Epicentre) were maintained in E. coli by selection in media
containing 12.5 mg/L chloramphenicol, whereas Tn5 transposon derivatives of the fosmids were
maintained on media containing 5 mg/L tetracycline. Fosmid bearing strains were grown with
0.1% arabinose to increase the copy number of the fosmid. Plasmid pBBR1MCS-3 was
maintained with 6 mg/L tetracycline in strain Ibu2 and 10 mg/L in E. coli.
3.2.3. Cloning the 4-isobutylcatechol-2,3-dioxygenase (ipfJ)
The ipfJ gene was amplified using primers described in Table 3-1, then cloned into the
pBBR1MCS-3 plasmid (Kovach et al., 1995) and transformed into E. coli JM109 to produce
E.coli JM109 pC (Table 3-1).
3.2.4. Generation of spontaneous 4TAA mutants
Strain Ibu2 was grown on MSM and glycerol and cells were washed with 30mM
phosphate buffer to remove the carbon source. Cells were then resuspended at A600=0.02 in

53
MSM with 250 mg/L 4TAA, 2PPA, or PAA as carbon sources. Liquid cultures were monitored
periodically until growth appeared in three different 4TAA cultures after 17 days (Figure A-1).
Aliquots were then spread on LB plates with 10 mg/L tetracycline and one colony was isolated
from each for further evaluation. These mutants are named strains 4TAA1, 4TAA2, and 4TAA3
(Table 3-1).
3.2.5. MC activity
MC activity was assessed by measuring the appearance of MC product (MCP) formation
spectrophotometrically following the addition of the appropriate catecholic substrate to washed
cell suspensions (WCS) or actively growing cultures. Catechol was routinely used as the
substrate to determine relative MC activity levels in actively growing cultures following different
treatments and concentrations used are indicated. Substituted catechols were also used as
substrates when indicated. MC activity rates were reported as µM MCP formed per minute per
unit of optical density of the cell culture or cell suspension at 600 nm. The absorbance
wavelengths and extinction coefficients used were 36/mM/cm for catechol at 375 nm,
13.8/mM/cm for 3-methylcatechol at 388nm, and 28.1/mM/cm for 4-methylcatechol at 382 nm
(Heiss et al., 1995).
In order to determine the ability of analogs to induce MC activity at low doses, early
exponential phase cultures (A600=0.05) of strain Ibu2 that were growing on MSM and glycerol
were provided with 50 mg/L IPF, 2(4T)PA, 4TAA, 2PPA, or PAA for eight hours, and then
assessed for MC activity as described above. In order to determine the effect of metabolizing

54
higher concentrations of the analogs on MC activity, mid exponential phase cultures (A600=0.3)
of strain Ibu2 that were growing on MSM and glycerol were provided with 250 mg/L IPF,
2(4T)PA, 4TAA, 2PPA, or PAA for 24 hours and then assessed for residual MC activity as
described above. In order to evaluate 4-isobutylcatechol-2,3-dioxygenase MC activity, WCS
were prepared from JM109 pC cultures by washing cells two times with 30 mM phosphate
buffer prior to resuspending them in the same to give a final absorbance of A600=2.5 and MC
activity was assessed as described above. To evaluate substrate specificity, strain Ibu2 and E.
coli strains bearing different Ibu2 fosmid library derivatives were induced with 10 mg/L IPF for
7.5 and 6 hours respectively and assessed for MC activity as described above; strain Ibu2 was
grown on MSM and glycerol to early exponential phase (A600=0.05), and overnight cultures of E.
coli strains 3G7, 3G7 TnH5, and 3G7 TnG9 were transferred (1:10) in to fresh LB prior to
induction.
3.2.6. Deactivation of IpfJ
E. coli JM109 pC WCSs were prepared as described above. WCSs were exposed to 250
mg/L catechol or substituted catechol for six hours, then washed again four times and assessed
for MC activity as described above.
3.2.7. Catechol formation by the upper pathway
To test the effect of different analogs on the inducibility of catecholic intermediate
production, strain Ibu2 was grown on MSM and glycerol to mid exponential phase (A600=0.3)

55
then washed and resuspended at A600=0.05 in MSM with glycerol plus equimolar concentrations
of one of the AAAs (309 mg/L IPF, 246 mg/L 2(4T)PA, 225 mg/L 4TAA, 225 mg/L 2PPA, or
204 mg/L PAA). After 12 hours, cells were washed twice and resuspended at A600=4 in 30 mM
phosphate buffer, and 50 mg/L 3-fluorocatechol was added to deactivate MC activity (Murdoch
and Hay, 2005). These WCSs were then provided with the same substrate (225 mg/L PAA) so
that we could distinguish between different levels of induction and different rates of metabolism
of the individual analogs. Catechol formation was measured over three hours by HPLC as
described by Murdoch & Hay (Murdoch and Hay, 2005). In order to determine differences in the
rates of metabolism of the individual analogs, non-induced WCSs were prepared from mid
exponential phase (A600=0.3) cultures of strain Ibu2 that were grown on glycerol. WCSs were
resuspended in phosphate buffer at A600=4 and MC activity was deactivated as described above.
These WCSs were then provided with 500 mg/L IPF, 2(4T)PA, 4TAA, 2PPA, or PAA and
catechol formation was measured over 1.5 hours as described above.
In order to determine the relative differences in the rate of catechol production by strain
Ibu2 and the 4TAA mutants, cultures grown to exponential phase (A600=0.5) on MSM and
glycerol were induced with 250 mg/L 4TAA for eight hours. WCSs were prepared as described
above and then supplied with 500 mg/L 4TAA so that 4-methylcatechol formation could be
measured via HPLC over one hour.

56
3.3. Results
3.3.1. Meta cleavage activity
MC deactivation is known to limit the range of aromatic compounds that can support the
growth of many organisms (Polissi and Harayama, 1993; Ramos et al., 1987). Given the
surprising range of substrates that had previously been reported to support the growth of strain
Ibu2, we sought to quantify the effect that the metabolism of IPF and analogs had on MC
activity. As can be seen in Figure 3-1, MC activity was suppressed below the basal level by non-
growth substrates (4TAA, 2PPA and PAA), but MC activity was elevated above that of the
control for growth substrates (IPF and 2(4T)PA). This was somewhat surprising since
metabolism of both growth substrates resulted in the production of para-substituted catechols,
which have previously been reported to be more potent deactivators of MC activity than their
non-substituted analogs.
Figure 3-1. MC activity of strain Ibu2 metabolizing growth supporting doses (250 mg/L) of analogs for 24 hours. MC activity of growing cultures was assayed with 250 mg/L catechol. All analog treatments were significantly different from the no analog control, and all except for 2(4T)PA were different from IPF (2-tailed t-Tests, p<0.05). Means and standard deviations, n=3.

57
We therefore hypothesized that strain Ibu2’s MC activity might be greater towards para-
substituted catechols or more resistant to their deactivating effects. Conversely, strain Ibu2’s
MC activity was approximately five times greater towards catechol than 4-methylcatechol
(Figure 3-2A). A gene that appeared to encode all of strain Ibu2’s MC activity, ipfJ, was
identified upstream of the ipf upper pathway genes by DNA sequencing of strain 3G7, and a
PLFeDox (ipfK) was identified in a separate operon (Figure A-3). Evidence that IpfJ was the
primary MC enzyme contributing to the ipf pathway comes from the observations that strains
Ibu2 and 3G7 shared similar patterns of IPF inducible MC activity and substrate specificity, and
that there was no MC activity in two ipfJ mutants of 3G7 (Figure 3-2B). Ibu2’s MC activity was
greatest towards 3-methylcatechol however this substrate was merely used as a probe for
comparison of MC activity patterns in the strains. We tested catechol deactivator potency on
heterologously expressed ipfJ encoded MC activity using several commercially available
substituted catechols (isobutylcatechol was not available commercially). Interestingly, however,
para-substituted catechols were the most potent deactivators of MC activity (Figure 3-3), which
is consistent with what has been found for other MC enzymes (Polissi and Harayama, 1993;
Ramos et al., 1987).

58
Figure 3-2. MC substrate range of strains Ibu2 and 3G7 and ipfJ mutants of 3G7 (TnH5 and TnG9). Strains were grown in the presence of 10 mg/L IPF or no inducer (control) and growing cultures of (A) Ibu2 and (B) 3G7 strains were assessed for MC activity towards 500 mg/L of catechol (cat), 3-methylcatechol (3Mcat), or 4-methylcatechol (4Mcat). Statistical significance between induction treatments is indicated by an asterisk (2-tailed t-Tests, p<0.05).
Figure 3-3. Deactivation of heterologously expressed IpfJ. WCSs of E. coli JM109 pC were exposed to 250 mg/L catechol (cat), 4-methylcatechol (4Mcat), 4-tertbutylcatechol (4tBcat), or a methanol control, and then washed and assessed for MC activity towards 500 mg/L catechol. The percent activity remaining after exposure is shown. All treatments were significantly lower than the control and 4tBcat was significantly lower than cat (1-tailed t-Tests, p<0.01). Means and standard deviations, n=3.

59
Non-substituted catechols were less potent deactivators of MC activity than substituted
catechols so we hypothesized that the non-growth substrates might lack or have reduced ability
to induce MC activity in strain Ibu2. Cultures were exposed to a lower dose (50 mg/L rather
than 250 mg/L) and a shorter induction period (8 hours rather than 24 hours) in order to
minimize MC deactivation by metabolites. However, under these conditions, all analogs caused
an increase in MC activity (Figure 3-4).
Figure 3-4. Induction of MC activity in strain Ibu2 by low doses (50 mg/L) of analogs and a no analog control. MC activity of growing cultures was assayed with 500 mg/L catechol. All induction treatments were statistically different from the control, and only the control and PAA treatments were different from IPF (2-tailed t-Tests, p<0.05). Means and standard deviations, n=2.

60
3.3.2. Formation of catechol metabolites in the upper pathway
Table 3-2. Formation of catechols by the upper pathway of strain Ibu2. A) Substrate specificity was determined with uninduced washed cell suspensions (WCSs) and different analogs as substrates. B) Up-regulation was determined with IPF or analog induced WCSs and 2PPA as a substrate. All uninduced WCSs were statistically different from each other, and among the induction data set only 2(4T)PA induction was different from the no inducer control (2-tailed t-Tests, p<0.05). Means and (standard deviations), n=3.
Since strain Ibu2’s MC activity demonstrated a canonical sensitivity to para-substituted
substrates and could not explain its inability to grow on 2PPA or PAA, we decided to look at the
rate of catechol production via the upper pathway. We hypothesized that lower rates of catechol
production would lead to less MC deactivation and be a necessary prerequisite for growth on a
substrate. Because the rates of catechol production could vary due to differential substrate
specificity or differential up-regulation of enzymatic activity, we analyzed both possibilities.
There were differences in substrate specificity, but they were not what we expected (Table 3-2).
The data show that IPF was transformed at less than 1/10th the rate of the other substrates and the
remaining phenylpropionic acids were transformed faster than the phenylacetic acids, although
statistical analysis reveals that results for all treatments were different. When strain Ibu2
cultures were induced with IPF or analogs, the rate of metabolism in the upper ipf pathway was
not greatly affected. Induction with 2(4T)PA resulted in a slightly lower catechol formation rate,

61
however there were no consistent differences between the effects of growth and non-growth
substrates.
3.3.3. 4TAA mutants
Given the lack of a clear explanation for growth substrate range based on either catechol
degradation or production rates, we sought to better understand the phenomenon by
characterizing spontaneous mutants with an expanded growth substrate range. We isolated three
mutants of strain Ibu2 with the ability to use 4TAA as a sole carbon source, however, despite
repeated attempts, no mutants ever arose with the ability to grow on either 2PPA or PAA.
Strains 4TAA1, 4TAA2, and 4TAA3 were tentatively identified as mutants of strain Ibu2 due to
similarity of colony morphology, which was also supported by sequencing of components of the
ipf operon described later in this section. All three mutant strains, but not strain Ibu2, grew on
4TAA within a few days, confirming that they were mutants rather than just wild-type Ibu2 that
had adapted to the experimental conditions (Figure 3-5). Strain 4TAA1 had the slowest growth
rate when growing on any substrate, be it glycerol, IPF or 4TAA. Strain Ibu2 grew slightly faster
on IPF than the mutants.

62
Figure 3-5. Growth of strain Ibu2 and mutant strains 4TAA1, 4TAA2 and 4TAA3 on (A) 500 mg/L glycerol, (B) 250 mg/L IPF, or (C) 250 mg/L 4TAA. Means and standard deviations, n=3.
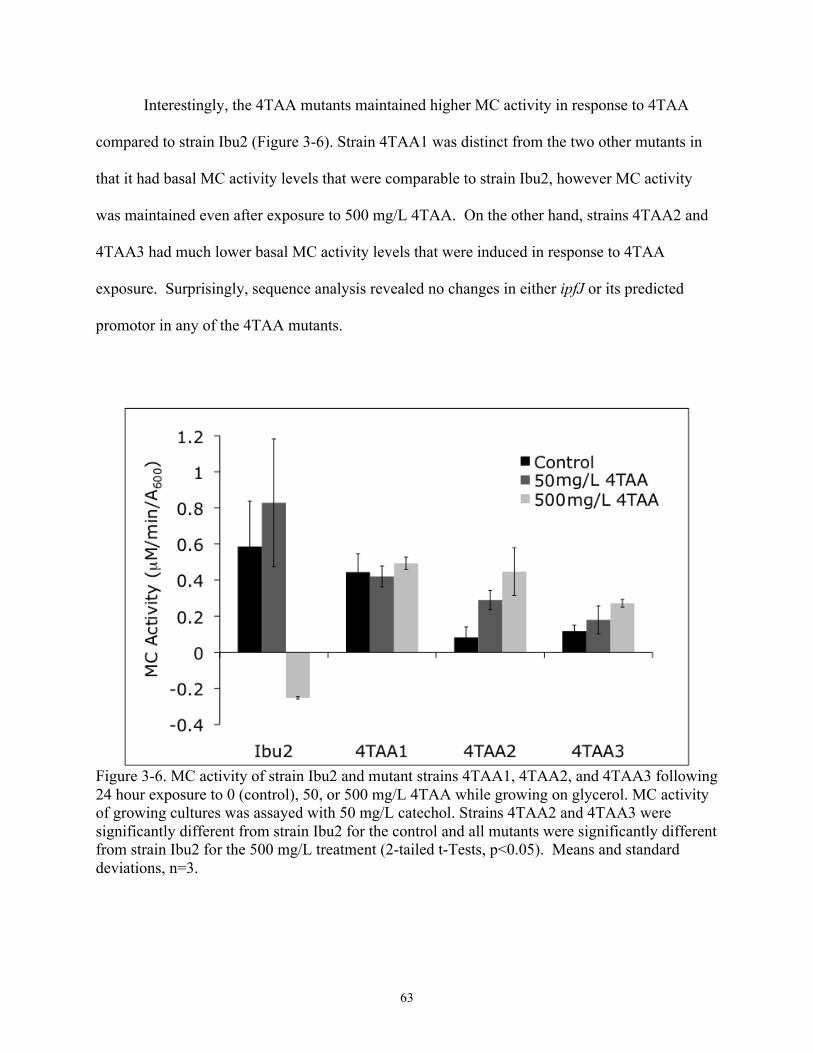
63
Interestingly, the 4TAA mutants maintained higher MC activity in response to 4TAA
compared to strain Ibu2 (Figure 3-6). Strain 4TAA1 was distinct from the two other mutants in
that it had basal MC activity levels that were comparable to strain Ibu2, however MC activity
was maintained even after exposure to 500 mg/L 4TAA. On the other hand, strains 4TAA2 and
4TAA3 had much lower basal MC activity levels that were induced in response to 4TAA
exposure. Surprisingly, sequence analysis revealed no changes in either ipfJ or its predicted
promotor in any of the 4TAA mutants.
Figure 3-6. MC activity of strain Ibu2 and mutant strains 4TAA1, 4TAA2, and 4TAA3 following 24 hour exposure to 0 (control), 50, or 500 mg/L 4TAA while growing on glycerol. MC activity of growing cultures was assayed with 50 mg/L catechol. Strains 4TAA2 and 4TAA3 were significantly different from strain Ibu2 for the control and all mutants were significantly different from strain Ibu2 for the 500 mg/L treatment (2-tailed t-Tests, p<0.05). Means and standard deviations, n=3.

64
Because DNA sequencing did not reveal an explanation for the change in MC activity,
we investigated the rate of catechol production for strain Ibu2 and 4TAA mutants that had been
exposed to 4TAA. 4-Methylcatechol formation was in fact significantly slower for all 4TAA
mutants than strain Ibu2 and catabolic rates for the mutants were not statistically different from
each other (Figure 3-7).
Figure 3-7. Upper pathway activity of strain Ibu2 relative to mutant strains 4TAA1, 4TAA2, and 4TAA3. Relative rates of 4-methylcatechol (4Mcat) production from 4TAA were normalized to that of strain Ibu2. Rates were significantly lower in all mutants than strain Ibu2, but did not differ among mutants (2-tailed t-Test, p<0.05) Means and standard deviations, n=3.

65
3.4. Discussion
We report here on our efforts to understand why strain Ibu2 can easily grow on para-
substituted phenylacetic acid analogs IPF and 2(4T)PA, but not on the para-substituted 4TAA or
non-substituted 2PPA or PAA. Para-substituted catechols, which are produced during
metabolism of IPF, 2(4T)PA, and 4TAA, are known to deactivate the C23Os responsible for
their degradation (Polissi and Harayama, 1993). This has been reported to be an important
obstacle faced by microbes trying to grow on bulky para-substituted compounds (Bird and Cain,
1974; Cerdan et al., 1994; Ramos et al., 1987). With this in mind, we found it intriguing that
strain Ibu2 could easily grow on the para-substituted IPF and 2(4T)PA, but not on the non-
substituted analogs, 2PPA or PAA.
To assess the effect of parent compound metabolism on MC activity, we initially exposed
strain Ibu2 cultures growing on MSM and glycerol to 250 mg/L IPF or its analogs and measured
residual MC activity in growing cultures. MC activity was significantly lower in cultures
exposed to 4TAA and undetectable in cultures exposed to 2PPA, and PAA (Figure 3-1). This
observation correlated with the fact that none of these analogs support growth. Reduction in MC
activity has frequently been reported to be due to suicide substrate inactivation by catechols
(Polissi and Harayama, 1993; Vaillancourt et al., 2002). In addition to suicide inactivation,
differences in the levels of MC activity may also be due to a number of factors including 1)
substrate specificity of the MC enzyme, 2) transcriptional regulation of MC gene expression, 3)
the presence of MC repair enzymes such as a PLFeDox, or 4) differences in the rate at which
catechols are produced (George and Hay, 2011).

66
We systematically assessed each of these possibilities to determine which might play a
role in determining the type of substrates that would support growth. Figure 3-2A demonstrates
that the rate of MC activity was higher for catechol than 4-methylcatechol, suggesting that
substrate specificity was not the factor limiting growth on 2PPA or PAA. Figure 3-3
demonstrates that the MC activity of IpfJ was more sensitive to deactivation by para- than non-
substituted catechols, even though substituted catechols were produced in strain Ibu2 from
substrates that supported growth. This suggests that inactivating potency of the catechol
produced would not limit growth on 2PPA or PAA either. We also found that all of the substrates
induced the expression of MC activity to similar levels, although PAA yielded slightly lower
activity (Figure 3-4). The fact that the level of MC activity in cells exposed to 4TAA and 2PPA
was not different from those exposed to IPF suggests that induction of MC activity could not be
the sole determinant of substrate range. Given that high doses of 4TAA, 2PPA, and PAA inhibit
MC activity (Figure 3-1), it is possible that these non-growth supporting substrates may have the
capacity to up-regulate expression of an MC enzyme but not other lower pathway genes
including those involved in MC enzyme reactivation. Our observations of low substrate
specificity and high deactivation by 4-methylcatechol, suggested that up-regulation of a
PLFeDox may be required for growth. Genetic analysis revealed that strain Ibu2 does have a
PLFeDox (ipfK), but it is not in the same operon as ipfJ (Figure A-3), increasing the possibility
that the two genes are not co-regulated. More work including determination of transcription
levels of these genes via RT-PCR and MC activity assays of induced cells before and after
hydrogen peroxide inactivation (Polissi and Harayama, 1993) should help determine if ipfK is
differentially induced by growth versus non-growth substrates. The high deactivation of MC

67
activity by 2PPA and PAA was unexpected and could not be explained by MC activity up-
regulation, substrate specificity or deactivation by catechol.
The last of the possibilities that we tested was that differences in the rates of catechol
production accounted for growth substrate specificity. This idea was based on the observation
that catechol accumulation can overwhelm cells’ detoxification and degradation capacity,
thereby limiting growth or causing cell death (George and Hay, 2011; Park et al., 2004). To do
this, we assessed the rate at which the parent compounds were converted to catecholic
intermediates. If catechol formation rates were the sole factor controlling MC deactivation
patterns in strain Ibu2, we would have expected to see more rapid transformation rates of 4TAA,
2PPA and PAA but that was not the case (Table 3-2). While strain Ibu2 transformed IPF to its
catechol at least 90% slower than the other substrates, the rate of transformation of 2(4T)PA was
higher than that of both 4TAA and PAA. Thus, no clear differences could be seen in the rates at
which growth and non-growth substrates were transformed to their respective catechols. Nor was
there a clear pattern with respect to the modest changes in upper pathway induction. It therefore
appears that catechol formation rate alone cannot explain the suppression of MC activity levels
in strain Ibu2 by non-growth substrates.
These results do not preclude the possibility, however, that several of these factors in
combination, such as modestly higher catechol production rates, modestly lower catechol
degradation rates, and modestly lower repair enzyme production, contributed to the failure of
these substrates to support growth. For now though, it remains unclear why the non-growth
analogs suppress MC activity, despite the fact that two of these compounds (2PPA & PAA) are

68
metabolized via non-substituted catechols and therefore should not deactivate MC activity to the
same level as the substituted catechols. This work focused on catechols because they are
typically believed to be a source of MC deactivation--an action performed by all non-growth
substrates in strain Ibu2. However, it is possible that another intermediate may be influencing
MC deactivation or limiting growth. No other pathways have been reported that combine MC
with the use of CoA in the upper pathway, so it is possible that some aspect of metabolism via a
CoA-aromatic thioester could be contributing to toxicity (this line of reasoning is addressed
further in Chapter 4).
Our inability to describe a single mechanism that caused non-growth substrates to
suppress MC activity in strain Ibu2 is consistent with our results from the 4TAA mutants in
which we identified at least two possible mechanisms that contributed to the maintenance of MC
activity: altered MC regulation and decreased rates of catechol production (Figures 3-6 and 3-7).
Strains 4TAA2 and 4TAA3 had low basal levels of MC activity that were up-regulated in
response to higher doses of 4TAA. Strain 4TAA1 differed in that basal levels were the same as
those for strain Ibu2, however, unlike Ibu2, 4TAA1 maintained these levels despite being
challenged by 4TAA. Since sequence analysis revealed that ipfJ and its promoter were identical
in all four strains it seems likely that 4TAA1-3 harbor mutations in some, as yet unidentified
regulator. Each of these mutants also produced catechol more slowly than strain Ibu2 (Figure 3-
6). This is consistent with a recently described “less is more” adaptive response in which lowered
activity in the upper pathway was found to expand growth substrate range during the metabolism
of aromatic compounds (George and Hay, 2011).

69
Strain Ibu2 was not able to grow on 2PPA or PAA. While we know that they are both
metabolized to MC products, we have not done any additional work on the lower pathway
beyond ring cleavage. In light of the fact that strain Ibu2 can grow on IPF and 2(4T)PA, and that
the dienoic acid metabolite of the HMSD enzyme was previously detected during IPF
metabolism (Murdoch and Hay, 2005), it is clear that enzyme activities of the 4-oxalocrotonate
branch of the lower pathway are all present in strain Ibu2 (Figure 1-2). MC pathways typically
funnel both para- and non-substituted catechol metabolites through the 4-oxalocrotonate branch,
however it is possible that one or more of the enzymes present in strain Ibu2 do not have activity
towards the non-substituted analogs. It is also possible that 2PPA and PAA lack the ability to
induce transcription of some lower pathway genes, although both clearly induced MC activity
itself.
Strain Ibu2 has not been amenable to genetic manipulation in our lab so our options for
experimental approaches have been limited. In addition, while we have biochemical evidence
that the lower pathway proceeds via MC (Murdoch and Hay, 2005), and preliminary work has
revealed the presence of MC pathway genes in strain Ibu2 (Appendix A.3), we have not
characterized lower pathway metabolism of IPF or its analogs by Ibu2. Additional work is
therefore required to resolve the mechanisms responsible for this unusual pattern of MC
deactivation and to elucidate the role that this phenomenon plays in catabolism of phenylacetic
acids in this novel hybrid pathway.

70
CHAPTER 4
Growth inhibition during arylacetic acid metabolism by the ibuprofen degrader
Sphingomonas sp. strain Ibu2
Abstract
Sphingomonas sp. strain Ibu2 has been previously shown to metabolize ibuprofen (IPF)
and other arylacetic acids (AAAs) using a novel hybrid upper pathway. However the substrates
2-phenylpropionic acid (2PPA) and phenylacetic acid (PAA) could not support growth for strain
Ibu2 and 4-tolylacetic acid (4TAA) could not serve as a sole carbon source. Previous work has
also shown that metabolism of 2PPA and PAA cause unexpected deactivation of meta cleavage
(MC) enzyme activity. The current study demonstrates that non-growth substrates (4TAA,
2PPA, and PAA) are toxic to strain Ibu2 and inhibit growth. In the presence of IPF, MC
deactivation was partially mitigated and there was no growth inhibition by the non-growth
substrates. However, strains engineered to express higher MC activity did not consistently
improve growth in the presence of non-growth substrates; Ibu2 strains with increased expression
of a plant-like ferredoxin (ipfK) had improved growth in the presence of these substrates, but the
4-isobutylcatechol-2,3-dioxygenase (ipfJ) exacerbated growth inhibition both with and without
ipfK. The antioxidant thiourea also exacerbated growth inhibition. Taken together, these results
suggest that the toxicity of these AAAs was not merely due to limited catechol turnover or
oxidative stress. In order to further explore other potential mechanisms of toxicity, the effects of
CoA-AAA thioester formation on growth was assessed in Escherichia coli heterologously
expressing the ipf-CoA-ligase (ipfF). Expression of ipfF was toxic to E. coli in the presence of
IPF or four other AAAs, with effective concentrations for 50% growth inhibition ranging from

71
42 µg/L for 2-(4-tolyl)propionic acid to 9.1 mg/L PAA. Mutants defective in other elements of
the ipf pathway also experienced toxicity, most notably the putative thiolase (ipfD), which
suggests that the putative CoA-AAA diol metabolites were also toxic. This work provides
preliminary data demonstrating that toxicity plays a role in limiting the growth substrate range of
strain Ibu2.

72
4.1. Introduction
For microbes to use an aromatic hydrocarbon for growth they must encode and produce
enzymes that can break it down into high-energy intermediates, without causing lethal toxicity.
Aerobic pathways typically involve dihydroxylated central metabolites such as catechol,
gentisate, and protocatechuate (Diaz, 2004). Many of these hydroxylated intermediates are toxic,
and catechols and their spontaneously oxidized products known to cause oxidative stress and cell
lysis (Camara et al., 2004; Yamada et al., 2006). In order to minimize toxicity while providing
intermediates that support growth via the Krebs cycle, cells must maintain sufficiently high
catechol degradation activity. A widely employed catechol degradation pathway facilitates the
insertion of molecular oxygen adjacent to one of the two ring hydroxyl groups, and is therefore
called meta cleavage (MC). Failure to maintain MC activity can result in the accumulation of
toxic catechol metabolites, incomplete metabolism, and may limit the production of usable
carbon for growth (Munoz et al., 2007; Park et al., 2004; Perez-Pantoja et al., 2003). Levels of
MC enzyme activity are often inducible and therefore dependent upon substrate specificity and
transcription levels, in addition to being affected by suicidal substrate-level inactivation. (Cerdan
et al., 1994; Polissi and Harayama, 1993; Ramos et al., 1987; Vaillancourt et al., 2002). MC
enzymes contain a ferrous iron active site that can be deactivated by oxidation during catechol
cleavage (Vaillancourt et al., 2002). Para-substituted alkylcatechols are generally more potent
deactivators than meta- or non-substituted catechols (Bird and Cain, 1974; Polissi and Harayama,
1993). Plant-like ferredoxins (PLFeDox) can reductively reactivate deactivated MC enzymes and
a PLFeDox is often required for growth on para-substituted compounds (Hugo et al., 2000;
Polissi and Harayama, 1993; Powlowski and Shingler, 1994).

73
Sphingomonas sp. strain Ibu2 can grow on the para-substituted ibuprofen (IPF) and 2-(4-
tolyl)propionic acid [2(4T)PA)] as sole carbon sources but not the para-substituted 4-tolylacetic
acid (4TAA) or the non-substituted 2-phenylpropionic acid (2PPA) or phenylacetic acid (PAA)
(Murdoch and Hay, 2005). Even though strain Ibu2 can metabolize all of these substrates, it is
still not entirely clear why 4TAA, 2PPA and PAA cannot serve as sole carbon sources (Chapter
3). Each of these substrates is metabolized through the upper ipf pathway and the beginning of
the lower pathway to a meta cleavage product (MCP) (metabolite V, Figure 4-1). The central
metabolite catechol is produced at similar rates from 2(4T)PA which supports growth, and
4TAA, 2PPA, and PAA which do not (Chapter 3). The lower pathway has not been fully
characterized but there is clear evidence of MC activity (Murdoch and Hay, 2005). While the
upper pathway is not induced, activity of the MC enzyme in the lower pathway is up-regulated
by IPF and each of the analogs (Chapter 3). However, exposure to higher concentrations (500
mg/L) of non-growth analogs (4TAA, 2PPA, and PAA) suppresses MC activity (Chapter 3).
The MC enzyme, 4-isobutylcatechol-2,3-dioxygenase, was found to be sensitive to deactivation
by catechols with, as expected, para-substituted catechols being the most potent deactivators.
Figure 4-1. Ipf pathway metabolites, enzymes and genes of strain Ibu2 as proposed by Murdoch (2011). Enzymes are ibuprofen-CoA-ligase (CoA Lig), ibuprofen-CoA-dioxygenase (Ring Diox), thiolase/acyltransferase (Thiolase), and 4-isobutylcatechol-2,3-dioxygenase (C23O) and genes are indicated below. Substrates of the pathway include IPF (R1=CH3, R2=CH2CH(CH3)2), 2(4T)PA (R1=CH3, R2=CH3), 4TAA (R1=H, R2=CH3), 2PPA (R1=CH3, R2=H) and PAA(R1=H, R2=H). Metabolites are (I) AAAs, (II) CoA-AAA thioesters, (III) CoA-AAA diols, (IV) catechols, and (V) MCPs.

74
A PLFeDox (ipfK) was identified in strain Ibu2 (Figure A-3). Interestingly, it does not
appear to be co-transcribed with ipfJ, so disregulation of the MC pathway genes is one
hypothesis we have proposed for strain Ibu2’s inability to use the para-substituted compound
4TAA as a sole carbon source (Chapter 3). In this study, we examined the effect of over
expressing ipfJ and ipfK independently and together on the growth inhibition caused by IPF and
its analogs. Given the propensity of catechols to cause oxidative stress, we also examined the
effect of the antioxidant thiourea.
We also considered the possibility that the deleterious effects of catechol might not
account for all of the toxic effects that IPF and its analogs have on bacterial cells. Therefore, we
assessed the toxicity of CoA-AAA thioesters to bacteria since these compounds are known to be
toxic to eukaryotic cells (Knadler and Hall, 1990; Skonberg et al., 2008). We examined the
effects of CoA-AAA thioesters on cell growth by heterologously expressed ipf-CoA-ligase (ipfF)
in E. coli. Toxicities of the other upper pathway metabolites were also examined in E. coli
harboring a fosmid containing a portion of strain Ibu2 DNA that encoded the functional upper ipf
pathway (strain 3G7), along with strains with transposon (Tn) mutants of fosmid 3G7 that were
defective for each of the steps in the upper pathway (Murdoch, 2011).

75
4.2. Methods
4.2.1. Chemicals
Catechol, ibuprofen, 2-(4-tolyl)propionic acid, 4-tolylacetic acid, 2-phenylpropionic acid,
and phenylacetic acid were purchased from Acros Organics. Thiourea was purchased from
Sigma Aldrich. Hydrogen peroxide (3%) was purchased from TopCare. HPLC grade methanol
was purchased from Fisher Scientific.
Table 4-1. Strains used in this study.
Strain Plasmid and/or characteristics Reference E. coli EPI300 Epicentre E. coli EPI300 3G7 Fosmid pCC1FOS:30-40kb Ibu2 DNA Murdoch 2011 E. coli EPI300 3G7 TnF 3G7 with Tn5 transposon in ipfF Murdoch 2011 E. coli EPI300 3G7 TnA 3G7 with Tn5 transposon in ipfA Murdoch 2011 E. coli EPI300 3G7 TnD 3G7 with Tn5 transposon in ipfD Murdoch 2011 E. coli EPI300 3G7 TnJ 3G7 with Tn5 transposon in ipfJ Murdoch 2011 E. coli JM109 E. coli JM109 pC Plasmid pBBR1MCS-3:ipfJ (forward primer
AATGGTACCGTCCATGCTGAAGTGTGGCGAAAT, reverse primer TCTGGGCCCTGTGTGGGTCAGCTTGTTGTTT)
Chapter 3
E. coli JM109 pP Plasmid pBBR1MCS-3:ipfK (forward primer TATGGGCCCGCGAGATCCCGGATGCGAATTT, reverse primer ACCGGATAGGCACGCGTTATTGATGAGCTCGAA)
This study
E. coli JM109 pC+P Plasmid pBBR1MCS-3:ipfJ + ipfK (cloned with both primers above)
This study
E. coli JM109 Plasmid pGEM-T-easy: Empty E. coli JM109 pF Plasmid pGEM-T-easy:ipfF Murdoch 2011 E. coli S17 Sphingomonas sp. Ibu2 WT Plasmid pBBR1MCS-3:Empty Murdoch & Hay 2005 Sphingomonas sp. Ibu2 pC Plasmid pBBR1MCS-3:ipfJ (described above) This study Sphingomonas sp. Ibu2 pP Plasmid pBBR1MCS-3:ipfK (described above) This study Sphingomonas sp. Ibu2 pC+P
Plasmid pBBR1MCS-3:ipfJ + ipfK (described above)
This study

76
4.2.2. Culturing of strains
Ibu2 and engineered strains were grown in liquid culture at room temperature with
aeration by shaking at 150 rpm in flasks or conical tubes. The base medium was a mineral salts
media (MSM) and carbon sources were 250 or 500 mg/L ibuprofen or analog, or 0.1% glycerol,
and sometimes they were combined. Escherichia coli strains were grown on Luria Bertani (LB)
broth. Plasmid pBBR1MCS-3 was maintained with 6 mg/L tetracycline and pGEM-T-easy was
maintained with 150 mg/L ampicillin.
4.2.3. Construction of new strains
The ipfJ and ipfK were PCR amplified using primers described in Table 4-1, then cloned
individually or together (pC, pP and pC+P) into the pBBR1MCS-3 plasmid (Kovach et al., 1995)
and transformed into E. coli JM109 (Table 1). Strains Ibu2 pP, Ibu2 pC, and Ibu2 pC+P were
created by transforming plasmids pP, pC and pC+P into E. coli S17, and were then transferred to
strain Ibu2 by conjugation (Lang and Oesterhelt, 1989).
4.2.4. Growth inhibition assays
Ibu2 strains were grown to exponential phase on MSM and glycerol and diluted in MSM
and glycerol plus 500 mg/L IPF or analog to either a high starting density of A600=0.15 for
experiments with thiourea and those with strains Ibu2 pP, pC, and pC+P, or a low starting

77
density of A600=0.02-0.05 for all other growth inhibition experiments. To test growth on IPF and
analogs in the absence of glycerol, cultures were grown on IPF and diluted to A600=0.05 in MSM
containing 250 mg/L IPF plus 250 mg/L of an analog. For each of the experiments, 15 mL
replicate cultures were grown in 50 mL conicals and shaken on their side to increase aeration.
Growth was measured either spectrophotometrically at 600 nm or by colony forming units
(CFU) on LB plates with 6 mg/L tetracycline.
4.2.5. MC activity
MC activity was assessed by measuring the appearance of MCP formation
spectrophotometrically at 375 nm following the addition of 500 mg/L catechol to whole cultures.
The extinction coefficient used was 36/mM/cm (Heiss et al., 1995). MC activity was reported as
mM MCP formed per minute per unit of optical density of the cell culture or cell suspension at
600 nm.
4.2.6. Toxicity of CoA-AAA thioesters
E. coli JM109 and E. coli JM109 pF cultures were grown at room temperature at volumes
of 200 µL in 96 well plates in 1/20 LB in MSM at room temperature. Expression of ipfF was
induced with 1000 mg/L lactose. Cultures were supplemented with various doses of IPF or
analogs and maximal cell growth was measured spectrophotometrically at 600 nm. The maximal
growth at two days was used to determine the percent growth inhbition in comparison with the
no analog control. Percent growth inhibition values were used to contruct models to predict

78
effective concentrations for growth inhibition using BioDataFit 1.02
(www.changebioscience.com). Models for IPF and 2(4T)PA were four parameter, while models
for 4TAA, 2PPA, PAA were linear.
4.2.7. Toxicity of upper pathway metabolites
E. coli EPI300, 3G7, and 3G7 Tn mutant strains (Table 4-1) were grown at room
temperature in 96 well plates on 1/10 LB in MSM and supplemented with 1000 mg/L IPF or 250
mg/L 2(4T)PA, 4TAA, 2PPA, or PAA. Cell growth was measured spectrophotometrically at 600
nm and maximal growth occurred after one day. For each strain, maximal growth of the IPF and
analog treatments were compared to the no supplement control to determine the percent growth
inhbition.

79
4.3. Results
4.3.1. Toxicity of the IPF analogs
It had previously been shown that strain Ibu2 could grow on IPF and 2(4T)PA as sole
carbon sources, but not 4TAA, 2PPA or PAA (Murdoch and Hay, 2005). We hypothesized that
toxicity might play a role in limiting growth so we grew strain Ibu2 on glycerol as a carbon
source plus 500 mg/L of IPF or analogs. Strain Ibu2 cultures reached higher densities with IPF
and 2(4T)PA than with glycerol alone, however a lag phase was observed in the presence of
4TAA, 2PPA, and PAA (Figure 4-2A). Interestingly, by day six, the 4TAA treatment had a cell
density that surpassed that of the glycerol control. The experiment was repeated and cell
viability was measured via CFU (Figure 4-2B), revealing that at doses of 500 mg/L, 4TAA had a
bacteriostatic effect and that both 2PPA and PAA were bactericidal (Figure 4-2B). Both culture
density and cell viability were compromised by the non-growth substrates only, however
inhibition was brief and growth resumed within one week.

80
Figure 4-2. Inhibition of growth of strain Ibu2 by IPF analogs. Growth on glycerol plus 500 mg/L IPF or analog or no analog control from two different experiments measured as either optical density (A) or cell viability (B). Means and standard deviations, n=3.

81
4.3.2. Induction with IPF mitgates analog toxicity
It had previously been shown that non-growth substrates did not appreciably reduce the
maximal culture density when co-metabolized with IPF (Murdoch and Hay, 2005), so we
wondered if there would be a lag phase under these conditions. That was not the case. When
strain Ibu2 was provided with IPF as as a sole carbon source, maximal growth was greatest for
the 250 mg/L IPF control and slightly lower with 500 mg/L IPF (Figure 4-3). When strain Ibu2
was provided with 250 mg/L of IPF plus an additional 250 mg/L of one of the analogs, growth
proceeded rapidly. No additional lag phase was observed and maximal growth was not
compromised by any of the analogs to a greater degree than with addition of IPF, although a
striking death phase was evident for cells grown in the presence of PAA. By comparing these
results with those in Figure 4-2, it was evident that IPF had a positive effect on growth in the
presence of the other analogs that was not confered by glycerol.
Figure 4-3. Growth of strain Ibu2 on IPF plus analogs. Cell viability is shown for strain Ibu2 growing on 250 mg/L IPF (Control) or 250 mg/L IPF plus an additional 250 mg/L IPF or analog [2(4T)PA, 4TAA, 2PPA, or PAA]. Means and standard deviations, n=3.

82
Previous work had shown that MC activity in strain Ibu2 was reduced during growth in
the presence of 500 mg/L 4TAA, 2PPA or PAA (Chapter 3). Given that IPF imparted an
improvement of growth for strain Ibu2 when stressed by these non-growth substrates (Figure 4-
3), we hypothesized that IPF might also rescued MC deactivation. Cultures that were
supplemented with 10 mg/L IPF did in fact have increased levels of MC activity when the
stressor was a non-growth substrate (Figure 4-4).
Figure 4-4. Meta cleavage activity in strain Ibu2 cultures suppressed by 500 mg/L analogs and rescued with 10 mg/L IPF. Treatment exposure was eight hours. Controls show basal activity (none), and low dose induction (10 mg/L IPF) and high dose inhibition (500 mg/L IPF) by IPF. Statistical significance of adding 10 mg/L IPF is indicated with an asterisk (1-tailed t-Test, p<0.05). Means and standard deviations, n=3.

83
4.3.3. Ibu2 strains engineered for high MC activity
IPF had a positive effect on growth (Figure 4-3) and MC activity (Figure 4-4) in the
presence of other analogs, however it was not clear if increasing MC activity alone would
improve growth. MC activity levels could also be affected by either an MC enzyme or a
PLFeDox. To address this issue we engineered strains of Ibu2 carrying ipfK, ipfJ, or both, and
compared growth inhibition of the engineered and wild type (WT) strains. The growth curves of
the engineered strains were similar to that of the WT in the absence of IPF analogs although their
maximal growth was slightly decreased (Figure 4-5). For all strains the maximal growth was
reduced and a lag phase was present when 2PPA, PAA and 4TAA were added as stressors,
however 4TAA inhibited growth to a slightly greater degree than 2PPA and PAA. This is in
contrast to Figure 4-2 where 4TAA was the least inhibitory of the non-growth substrates.
However, a higher starting culture density was used for the experiment in Figure 4-5 (A600=0.15
vs. 0.02) in order to facilitate faster recovery from growth inhibition and 4TAA has been shown
to be slightly more inhibitory under these conditions in other experiments (data not shown). In
the presence of non-growth analogs ipfK reduced the lag phase these analogs induced.
Interestingly, ipfJ exacerbated growth inhibition, both alone and with ipfK. MC activity was
confirmed to be higher in strains Ibu2 pC and Ibu2 pC+P and with greater MC activity
reactivation in a hydrogen peroxide deactivation and reactivation assay in comparison with strain
Ibu2 (Figure A-4). This suggests that these engineered strains are more resilient to MC
deactivation.

84
Figure 4-5. Growth inhibition in Ibu2 and engineered strains. Engineered strains harbor plasmids with MC genes: ipfJ (Ibu2 pC), ipfK (Ibu2 pP), and ipfJ and ipfK (Ibu2 pC+P). Cultures were grown on glycerol plus no stressor (A) or 500 mg/L of analogs 2PPA (B), 4TAA (C), or PAA (D). Means and standard deviations, n=3.

85
4.3.4. Supplementation with the antioxidant thiourea
Antioxidants have previously been reported to increase growth for organisms stressed by
catechols and related metabolites (Kang et al., 2006; Yamada et al., 2006). Given the growth
improvement imparted by ipfK (Figure 4-5) and since PLFeDoxs function to protect MC
enzymes from oxidative stress, we sought to determine if the more generalized protection from
oxidative stress afforded by an antioxidant could also enable strain Ibu2 to cope with analog
toxicity. However, when strain Ibu2 was supplemented with the antioxidant thiourea, this
treatment did not mitigate growth inhibition by stressors 4TAA, 2PPA or PAA (Figure 4-6). In
fact, thiourea, while not affecting strain Ibu2 in the absence of stressors, dramatically
exacerbated growth inhibition in their presence in a dose dependant manner. Although the
lowest dose of 5 mM thiourea did lengthen the lag phase for the stressor 4TAA, it also increased
the maximal growth in the presence of this analog. This result suggested the possibility that
lower doses might be more beneficial. However, experimentation with lower doses (0.5-10 mM)
also inhibited growth in a dose dependant manner (data not shown). The approaches that we had
used up to this point did not give us a clear answer as to which metabolites might be inducing
growth inhibition. Therefore, we broadened our focus and explored upper pathway metabolites.

86
Figure 4-6. Combined effect of the antioxidant thiourea (TU) and IPF analog metabolism on strain Ibu2 growth. Strain Ibu2 was grown on glycerol and no stressor (A), or 500 mg/L of 2PPA (B), 4TAA (C), or PAA (D). Cultures were supplemented with no TU, 5 mM TU, 20 mM TU, or 75 mM TU. Means and standard deviations, n=3.

87
4.3.5. CoA-AAA thioester toxicity
Since IPF and its analogs are metabolized by strain Ibu2 via CoA-AAA thioesters, which
are known to be toxic to eukaryotic cells (Knadler and Hall, 1990; Skonberg et al., 2008), we
examined the effects of CoA-AAAs on E. coli by heterologously expressing ipf-CoA-ligase
(ipfF). None of the analogs elicited a toxic response at levels below 100 mg/L in the absence of
ipfF, however all analogs generated toxicity at sub-mg/L levels when ipfF was expressed (Figure
A-5). The effective concentrations for which each compound reduced growth by 50% (EC50)
ranged from 42 µg/L for 2(4T)PA to 9.1 mg/L for PAA (Figure 4-7).
Figure 4-7. Growth inhibition by metabolic products of ipf-CoA-ligase (ipfF). EC50s for growth inhibition by IPF analogs for E. coli JM109 heterologously expressing ipfF are graphed and shown above the bars. EC50s were determined using the BioDataFit 1.02 model based on data in Figure A-5.

88
4.3.6. Toxicity of AAA metabolism in mutants of the ipf pathway
The growth effects of the upper pathway metabolites were examined in strain 3G7 (a
clone of strain Ibu2) and Tn mutants of strain 3G7. All strains were inhibited by exposure to
AAAs, however there were no differences between the responses by the E. coli host strain
EPI300 and strain 3G7 (Figure 4-8). Maximal growth was lower in strain TnF (mutation in the
IPF-CoA-ligase) than the strain EPI300 control for all AAAs. If a protein involved in uptake of
the parent compound is present in strain 3G7, then this could suggest that parent compounds
were accumulating in strain TnF, however uptake by the ipf pathway has not been completely
characterized (Murdoch, 2011). There was general improvement in strain TnA (mutation in the
IPF-CoA-dioxygenase) over strain TnF, however maximal growth was lower in strain TnA than
strain EPI300 for non-growth analogs and the differences were significant for 2PPA and 4TAA.
CoA-AAA thioesters would be expected to accumulate in strain TnA (Figure 4-1, metabolite II).
The greatest growth inhibition observed was for strain TnD (mutation in the putative thiolase),
ranging from 41% with 4TAA to 74% with 2(4T)PA in comparison with strain EPI300. In strain
TnD, metabolism is thought to be halted at CoA-AAA diols (Figure 4-1, metabolite III) however
this proposed metabolite has not been identified (Murdoch, 2011). There was general
improvement of growth for strain TnJ (mutation in the gene for the MC enzyme) in comparison
with strain TnD, however growth was less than that of the control. Each of these mutant strains
have been shown to accumulate catechol (Murdoch, 2011), although it would be expected that
strain TnJ would accumulate the most catechol.

89
Figure 4-8. Toxicity of AAA exposure to E. coli harboring fosmids containing mutations in the ipf pathway. E. coli host strain EPI300, strain 3G7 harboring the ipf pathway, and five different derivatives of strain 3G7 with Tn mutations in ipfF (TnF), ipfA (TnA), ipfD (TnD), and ipfJ (TnJ) were grown in the presence of 250 mg/L PAA, 2PPA, 4TAA, 2(4T)PA, or 1000 mg/L IPF and percent growth was determined in comparison with the no analog control. Significant reductions in growth in comparison to the strain EPI300 control are indicated with an asterisk; strain TnA had higher growth than strain TnF for all analogs except PAA; and strain TnJ had higher growth than strain TnD for all analogs (1-tailed t-Tests, p<0.05). There was no difference between strains EPI300 and 3G7 for any analogs (2-tailed t-Tests, p<0.05). Means and standard deviations, n=4.

90
4.4. Discussion
Strain Ibu2 had previously been shown to use IPF and 2(4T)PA as sole carbon sources,
but not 4TAA, 2PPA, or PAA (Murdoch and Hay, 2005). Further work demonstrated that each
of these substrates was metabolized to catechol in the upper pathway at a similar rate, however
MC activity was reduced below basal levels by non-growth substrates only, which likely limited
entry of the latter into the lower pathway (Chapter 3). This scenario is likely to have resulted in
the accumulation of upper pathway metabolites.
In order to determine if toxicity was limiting the growth substrate range of strain Ibu2,
cultures were grown on glycerol as a carbon source plus IPF or analogs as potential stressors.
Non-growth substrates were in fact toxic when they were the only AAA supplied (Figure 4-2). In
contrast, however, non-growth substrates were not appreciably toxic in the presence of IPF
(Figure 4-3) so we suspected that IPF induced protective effects. Supplementation with IPF also
partially mitigated MC deactivation by non-growth analogs (Figure 4-4). This suggested that
increased MC activity might alleviate growth inhibition by non-growth substrates. To test this
hypothesis we constructed Ibu2 strains with increased MC activity and grew them on glycerol in
the presence of IPF and its analogs. Unexpectedly, expression of ipfJ exacerbated growth
inhibition, while ipfK reduced the lag phase for growth on all non-growth substrates (Figure 4-5).
It is unclear what additional growth advantage IpfK might have had besides increasing MC
activity. PLFeDoxs have only been reported to shuttle electrons to deactivated MC enzymes,
reducing the oxidized ferric iron to its active ferrous form (Hugo et al., 1998). Over expression
of a ferredoxin reductase, another enzyme involved in the shuttling of electrons and more general

91
response to oxidative stress, has been shown to reduce oxidative stress and improve growth on
naphthalene (Kang et al., 2007). It is tempting to suggest that PLFeDoxs might act in a similar
manner and shuttle electrons to another substrate(s). While this is possible, it is without
precedent and would require further experimentation to establish. Other PLFeDoxs have only
been shown to act upon MC enzymes, although we could not find any work in the primary
literature that analyzed their interactions with other proteins.
It is also very interesting that expression of ipfJ did not improve growth, but rather
inhibited it further, whether expressed alone or with the ipfK. In similar studies, increasing
expression of meta cleavage enzymes (C23Os) has been beneficial for the growth of organisms
when they were exposed to substrates that caused MC deactivation. For example, Pseudomonas
putida strain F1 is limited in its ability to grow on styrene due to MC deactivation by
vinylcatechol, however strains constructed with increased expression of a somewhat deactivation
resistant C23O or a combination of a sensitive C23O with a PLFeDox were both successful in
allowing strain F1 to grow on styrene (George et al., 2011). In the same work, it was found that
neither increased expression of the PLFeDox nor a sensitive C23O alone was sufficient for
growth, however strains expressing each of these proteins individually did experience less cell
death than the WT when exposed to styrene. We have not identified a single cause of MC
deactivation in strain Ibu2 in previous work (Chapter 3) and the Ibu2 strains engineered with ipfJ
did not behave as would be expected in this work. While the results of both chapters point to a
connection between MC activity, growth inhibition and substrate range, this work suggests that
growth inhibition does not appear to be strictly limited by catechol turnover.

92
Given our results with ipfK and those of Kang et al. (2007) with a ferredoxin reductase,
we wondered if more generalized oxidative stress might contribute to growth inhibition in strain
Ibu2. Surprisingly, in the presence of non-growth substrates, thiourea exacerbated AAA toxicity
to strain Ibu2 in a dose dependant manner (Figure 4-6). Thiourea, a hydroxyl radical scavenger,
has been used to overcome the toxicity associated with oxidative stress in bacteria at
concentrations ranging from 1.5-100 mM (Repine et al., 1981; Wang and Zhao, 2009). Thiourea
can also act as a prooxidant, but at higher concentrations of 263-525 mM (Brennan and Schiestl,
1998). The lack of beneficial effect by thiourea in strain Ibu2 suggests that general cellular
oxidative stress is not the primary mechanism of toxicity exerted during metabolism of 2PPA,
PAA or 4TAA. Additional work is clearly required, however, on the role of antioxidants and
could include an examination of the effect vitamin E, ascorbate, and ferrous iron since they have
been shown to aleviate toxicty in naphthalene and biphenyl degradation (Kang et al., 2006;
Yamada et al., 2006). Because thiourea actually exacerbated the stress effect, our results suggest
that either thiourea is bioactivated into a toxicant in the presence of AAAs, or that it somehow
increases the concentration of toxic AAA metabolites. The latter could occur either through
increased production or decreased removal of metabolites. Thiourea and related thioamides have
been shown to inhibit phenoloxidase (Liu et al., 2006a; Rescigno et al., 2002), although this
enzyme employs a different mechanism than that expected of either the ipf-CoA-dioxygenase
(IpfAB) or 4-isobutylcatechol-2,3-dioxygenase (IpfJ), it is possible that one or both of these was
inhibited in thiourea’s presence.
Given the possibility that thiourea mediated inhibition of the IpfAB could lead to CoA-
AAA thioester accumulation, which has previously been reported to be toxic in eukaryotes, the

93
toxic effects of this metabolite and others in the upper pathway were investigated further. E. coli
heterologously expressing the ipf-CoA-ligase (ipfF) from a high copy plasmid (pF) were
sensitive to AAAs at sub-mg/L concentrations (Figures 4-7 and A-5). However, the CoA-AAA
thioester metabolites of IPF and 2(4T)PA were more toxic than those of the non-growth
substrates, which suggests that the toxicity of the relevant CoA-AAA thioesters was unlikely to
govern substrate toxicity. When the ipf upper pathway was expressed on a low copy number
fosmid, however, the CoA-AAA thioester was only slightly toxic in comparison with the
putative CoA-AAA diol, which was expected to accumulate in strain TnD (Figure 4-8). The
CoA-AAA thioesters generally reduced growth during metabolism of non-growth substrates,
while the putative CoA-AAA diols caused a significant decrease in growth during metabolism of
all AAAs. CoA-AAA thioesters have previously been shown to be toxic during the eukaryotic
metabolism of NSAIDs, because they attack nucleophillic sites on proteins and form covalently
bound protein adducts (Knadler and Hall, 1990; Skonberg et al., 2008). It is possible that this
could contribute to MC enzyme deactivation in strain Ibu2, however additional work would be
required to determine this. To our knowledge, this is the first time that CoA-thioesters have been
reported to be a toxic metabolite of AAA metabolism in bacteria, despite the fact that several
microbial anaerobic and hybrid pathways employ CoA-ligation. However, these pathways do
not use MC for ring cleavage; in anaerobic and hybrid pathways, CoA-thioesters have been
suggested to be less toxic than metabolites of aerobic pathways with meta cleavage (Fuchs,
2008). Despite evident inhibition of E.coli growth, the toxicity pattern of the metabolites
produced during heterologous expression of the ipf pathway did not consistently correlate with
the toxicity of the parent compounds to strain Ibu2. Therefore, more research is required to
determine what role, if any, they have on the growth substrate range in strain Ibu2.

94
This work demonstrates that the non-growth substrates 4TAA, 2PPA, and PAA do
behave as stressors when they are metabolized by strain Ibu2. It shows that growth in the
presence of non-growth substrates is not limited by catechol turnover, despite the fact that they
cause deactivation of MC activity. Toxicity can be mitigated by IPF, however we have not
identified what function is up-regulated. Work with heterologously expressed ipf genes indicated
that CoA-AAA thioesters and their diols can be toxic during AAA metabolism, however
additional work is required to characterize mechanisms of toxicity. Chapters 3 and 4 demonstrate
a clear correlation among MC deactivation, toxicity, and the inability of substrates to support
growth, however additional work is required to fully understand the factors required for growth
on AAAs in strain Ibu2.

95
CHAPTER 5
Conclusion
5.1. Monitoring of the Lower Hudson River
The Lower Hudson River study gave us important base-line information about the
contribution of emerging pollutants in a section downstream of Albany and upstream of New
York City. We identified 25 contaminants but only caffeine (average 186 ng/L) and HHCB
(average 80 ng/L) were detected at quantifiable levels (Table 2-1). We detected nine pollutants
commonly derived from oil, coal, and agricultural pesticides. We also detected many emerging
contaminants, a few are highlighted below. Caffeine, while not typically thought of as a
pollutant, is the one compound that we detected at concentrations (370 ng/L) close to those
shown to effect aquatic life (600 ng/L). We also found ingredients of personal care and
household cleaning products such as antimicrobials, perfumes, and detergents. While biological
effects of alkyl ethoxylate detergent products and the polycyclic musks HHCB and AHTN have
not been measured at the concentrations that we detected in the Lower Hudson River, they are
persistent bioaccumulative endocrine disruptors. Two compounds, triclosan and piperonyl
butoxide are of interest because they can have synergistic effects through their action as
inhibitors of P450 enzymes. Despite the fact that we did not detect concentrations of these
compounds that were biologically relevant when encountered alone, the presence of these
compounds as part of a mixture in our samples leaves open the possibility that they could have as
of yet unidentified effects. Toxicologists have not been able to thoroughly evaluate on an
individual basis, the effects of the tens of thousands of chemicals in our consumer products and

96
this has inspired a national project, Tox21, to employ pathway based toxicological testing
approaches that have enough throughput capacity to begin looking at poorly characterized
compounds as well as combinations of different compounds (Schmidt, 2009).
Our lack of knowledge with respect to the effects of individual compounds can be
addressed to some degree by using endpoint analyses that rely on measures of biologically
relevant responses. This is what prompted our use of endocrine pathway based bioreporters
(CALUX), which allowed us to detect endocrine activity in all samples tested (Figure 2-2).
Fortunately, the activities that we measured were below levels previously shown to cause effects
in the field. However, both estrogen and androgen activity were detected at levels within one
order of magnitude of those found at field sites of other studies where aquatic species had
intersex traits (Jenkins et al., 2001; Jobling et al., 2009; Jobling et al., 2006). Few of the
chemicals that we detected have been assessed for endocrine activity, however, even when we
calculated the known hormone equivalency factors for these compounds we could not account
for the all of our measured CALUX responses. This may in part be due to insufficient
toxicological data, however it is also likely due to the fact that our chemical analysis was focused
on traditional and emerging contaminants rather than hormones. This study is part of an ongoing
project, so in its next phase will schedule chemical analysis of hormones. The remainder of the
work in this dissertation takes a very different approach to studying emerging contaminants. The
focus is on the biodegradation of the pharmaceutical IPF and its analogs by the WWTP isolate
Sphingomonas sp. strain Ibu2.

97
5.2. Factors limiting the arylacetic substrate range of strain Ibu2
Strain Ibu2 can metabolize a variety of AAAs, however only a subset of these can
support growth. The goal of this work was to identify the factors limiting its range of growth
substrates. Non-growth substrates 4TAA, 2PPA, and PAA, but not growth substrates IPF and
2(4T)PA were shown to suppress MC activity below basal levels and inhibit cell growth (Figures
3-1 and 4-2). However, supplementation with a growth substrate, IPF, mitigated the inhibitory
effects of non-growth substrate effects (Figures 4-3 and 4-4). Through the use of heterologously
expressed ipf pathway genes, CoA-AAA thioesters and the putative CoA-AAA diols were
identified as toxic metabolites of the ipf pathway (Figures 4-7 and 4-8) although the relative
toxicity of these compounds did not explain the ability of strain Ibu2 to use them for growth or
not.
5.2.1. MC activity levels are likely contributing to the lack of growth on 4TAA
MC deactivation is commonly observed in microorganisms that are degrading aromatic
compounds, and the catechol substrate has been a focal point of many studies (Bartels et al.,
1984; George et al., 2011; Polissi and Harayama, 1993; Ramos et al., 1987; Vaillancourt et al.,
2002). Our work with 4TAA was consistent with the literature in that the 4-methylcatechol had
lower turnover rates and was a more potent deactivator of MC activity (Figures 3-2 and 3-3).
Mutants were isolated with the ability to grow on 4TAA and they had decreased rates of catechol
production as well as differences in the regulation of the MC activity (Figures 3-6 and 3-7). This
suggests that managing intracellular concentrations of catechols is an important requirement for

98
growth on AAAs in strain Ibu2, as has been previously reported in the literature for nonacidic
aromatic compounds (George and Hay, 2011). Sequencing of ipfJ and its predicted promotor did
not reveal any genetic changes, however work is being continued in the lab to search for genetic
changes in the upper pathway and this may provide a mechanistic explanation for our
observation of decreased upper pathway activity. While mutants with the ability to grow on
4TAA had these changes in catechol management, increasing MC activity alone did not alleviate
the inhibition of cell growth by any of the non-growth substrates (Figure 4-5). This suggests that
additional, as yet unidentified factors are limiting growth.
5.2.2. Atypical MC deactivation by 2PPA and PAA
Our observation of MC deactivation by 2PPA and PAA is not consistent with effects
described in the literature because the non-substituted catechols produced by catabolism are not
typically potent MC deactivators. As would be expected in previously described pathways, IpfJ
was not found to be more sensitive to catechol than 4-methylcatechol (Figure 3-3), and catechol
was not produced more rapidly in the upper pathway (Table 3-2). Because the ipf pathway is
composed of a novel organization of steps with CoA-ligation in the upper pathway and MC in
the lower pathway, additional deactivation mechanisms should be considered. For example,
CoA-thioesters have previously been shown to acylate proteins so future work should explore the
effects of CoA-AAA thioesters on MC activity. It is also possible that inhibitory metabolites are
accumulating in the lower pathway due to incomplete metabolism, although we did not examine
that possibility in this work. The activity of lower pathway enzymes could be measured (Sala-
Trepat and Evans, 1971). If there is low activity of one or more enzymes then a strain could be

99
engineered to combine the upper ipf pathway with a complete and functional lower MC pathway
to determine if MC deactivation can be overcome by improving lower pathway function. It is
also worth noting that while CoA-thioesters were found to be toxic to cells heterologously
expressing ipf genes, their toxicity has not been reported in other pathways employing CoA-
ligation. E. coli strain K12 uses the paa pathway (Figure 1-1) and this strain could be used to
examine the potential toxic effects of CoA-thioesters in other pathways.
5.2.3. Expression of the PLFeDox mitigated stress
While increased expression of the MC enzyme IpfJ exacerbated growth inhibition by
4TAA, 2PPA, and PAA, the PLFeDox IpfK mitigated growth inhibition (Figure 4-5). This
suggests that IpfK may have had an additional benefit besides increasing MC activity.
Ferredoxin reductases have been shown to reduce general oxidative stress (Lee et al., 2006), as
well as promote growth in cells stressed during growth on naphthalene (Kang et al., 2007).
Future work to investigate the possibility that IpfK acts to reduce oxidative stress in the ipf
pathway could employ the superoxide generating agent paraquat and endpoints such as growth or
redox status as measured by the nitroblue tetrazolium assay (Lee et al., 2006). On the other
hand, our experiments with thiourea suggest that oxidative stress may not have been the cause of
toxicity (Figure 4-6). However, future work is required to rule out the possibility that thiourea
did not have other detrimental effects.

100
5.2.4. Inhibition of growth by the antioxidant thiourea
Interestingly, when thiourea was provided at concentrations previously shown to produce
anti-oxidative effects, it exacerbated the toxicity of non-growth substrates; 500 mg/L 4TAA,
2PPA, or PAA plus 75 mM thiourea resulted in the complete inhibition of growth of strain Ibu2
(Figure 4-6). A literature review revealed that thiourea is an inhibitor of phenoloxidase, a
tyrosinase (Liu et al., 2006a). Tyrosinases use a di-copper center to perform both the
hydroxylation of phenols and the oxidation of catechols to quinones, and inhibition by some
thiourea analogs may be due to chelation of copper (Rescigno et al., 2002). The oxygenases of
the ipf pathway (IpfAB and IpfJ) use iron center active sites which employ different mechanisms
(Harayama et al., 1992). However, it is possible that thiourea may have inhibited one of these or
other enzymes in the ipf pathway and caused the accumulation of toxic metabolites in strain
Ibu2. The effects of tyrosinase inhibitors such as Tropolone or phenylthiourea (Rescigno et al.,
2002) should be examined for general toxicity as well as the inhibition of ipf pathway enzymes.
Since thiourea toxicity was not observed in the absence of 4TAA, 2PAA, and PAA, it is also
possible that induction with AAAs lead to the bioactivation of thiourea. Given the toxic
interaction between thiourea and these AAAs, it is likely that toxicity was due to either up-
regulation of the ipf pathway or metabolism of AAAs via the ipf pathway. Future work could
employ AAAs and thiourea to select for growth tolerant isolates, which may harbor mutations in
the ipf pathway and serve as tools for further pathway evaluation. We are not certain why the
antioxidant thiourea exacerbated the toxicity of non-growth substrates. Therefore, additional
testing should be conducted to determine if other antioxidants such as ascorbate or vitamin E can

101
alleviate the toxicity of ibuprofen analogs (Yamada et al., 2006). This would help determine if
oxidative stress might be one of the factors limiting growth on AAAs by strain Ibu2.
5.2.5. Conclusion for strain Ibu2
This investigation suggests that MC deactivation and toxicity are two factors limiting the
range of AAAs that support the growth of strain Ibu2. Toxic metabolites and treatments for
exacerbating and alleviating growth inhibition have also been identified. Additional research
will be required to examine mechanisms underlying these observations.

102
APPENDIX
A.1. Arylacetic acids that support growth
Strain Ibu2 was grown on MSM and glycerol and cells were washed with 30 mM
phosphate buffer to remove the carbon source. Cells were then resuspended at A600=0.02 in
MSM with 250 mg/L IPF, 2(4T)PA, 4TAA, 2PPA, or PAA as carbon sources. Liquid cultures
were monitored spectrophotometrically at 600nm. Strain Ibu2 could readily use IPF and
2(4T)PA as sole carbon sources, but not 4TAA, 2PPA or PAA (Figure A-1). Strain Ibu2 grew on
4TAA after 17 days, but did not grow on 2PPA or PAA. Half maximal growth was achieved on
IPF in 5.7 (+/-0. 6) days, 2(4T)PA in 8.6(+/-0.1)days, and 4TAA in 19 (+/-1) days. Cultures
growing on 4TAA were later confirmed to be mutants.
Figure A-1. Growth of strain Ibu2 on 250 mg/L IPF or analogs. Means and standard deviations, n=3.

103
A.2. Deactivation of MC activity in strain Ibu2 by ibuprofen
The effects of IPF on MC activity were assessed for early exponential phase (A600=0.05)
strain Ibu2 cultures growing in MSM with 0.1% glycerol that were induced with various
concentrations of IPF for 7.5 hours. MC product was measured spectrophotometrically at 375nm
with 500 mg/L catechol as a substrate. MC activity increased following induction with low
concentrations of IPF beginning at 500 µg/L (Figure A-2). Activity continued to increase in a
dose dependant manner up to 5 mg/L, then leveled off at 50 mg/L, and began to drop again at
500 mg/L. At 1000 mg/L, MC activity was suppressed to levels below that of the control.
Figure A-2. Strain Ibu2 MC activity in cultures induced with various concentrations of IPF while growing on glycerol. All treatments were significantly different from the control, with exception to 50 µg/L IPF (2-tailed t-Test, p<0.05). Means and standard deviations, n=3.

104
A.3. Operon structure
In order to investigate the genetic environment surrounding ipf genes, clones from a
strain Ibu2 library (E. coli strains 3G7 and 4F6) and their transposon (Tn) mutants were chosen
for further sequencing (Murdoch, 2011). Strains 3G7 and 4F6 and their Tn mutants were
sequenced using DNA from plasmid preparations or amplicons from PCR using Tn specific
primers (forward: GGATGTGCTGCAAGGCGATTAAGTTGG and reverse:
CTCGTATGTTGTGTGGAATTGTGAGC). Additional primers were used to walk down the
sequence. Sequencing was performed by the Cornell Biotechnology Institute and sequences were
analyzed using DNAstar. Operon mapping revealed that the MC enzyme ipfJ is not in the same
genetic environment as the rest of the MC pathways genes. A PLFeDox (ipfK) was located
within a partial sequence for a MC lower pathway operon on a separate contig.
Figure A-3. Ipf and related MC genes of strain Ibu2. Ipf upper pathway genes are shown in black and pictured in Figure 1-1. Putative MC lower pathway genes are shown in dark gray and pitured in the Figure 1-2. Transposable elements are shown in light gray. The 4-isobutylcatechol-2,3-dioxygenase (ipfJ) and PLFeDox (ipfK) are on separate contigs.

105
A.4. Meta cleavage activity in engineered strains of Ibu2
Cultures of Ibu2 and engineered strains carrying a pBBR1MCS-3 plasmid to over express
a MC enzyme (ipfJ) and a PLFeDox (ipfK) were grown on glycerol to exponential phase (strains
described in Chapter 4). MC activity was assessed by measuring the appearance of MC product
(MCP) formation spectrophotometrically at 375 nm following the addition of 500 mg/L catechol
to whole cultures. The extinction coefficient used was 36/mM/cm (Heiss et al., 1995). To
measure the reactivation of MC activity, MC activity was deactivated with 5mM hydrogen
peroxide (H2O2) and the reappearance of MC activity was measured (Polissi and Harayama,
1993). The engineered strains had higher basal MC activity than strain Ibu2 (Figure A-4A) and
recovered from MC deactivation (Figure A-4B), while strain Ibu2 did not.
Figure A-4. MC activity before (A) and after (B) MC deactivation in Ibu2 and engineered strains over expressing ipfJ (Ibu2 pC) or ipfJ and ipfK (Ibu2 pC+P). Strains Ibu2 pC and pC+P both had higher MC activity than strain Ibu2 before deactivation, as indicated by asterisks (1-tailed t-Test, p<0.005). Means and standard deviations, n=3.

106
A.5. Growth Inhibition by CoA-AAA thioesters
The ipf-CoA-ligase (ipfF) was previously cloned into a pGEM vector and expressed in E.
coli JM109 (Murdoch, 2011). The sequence of the cloned fragment was checked by PCR
amplification and sequenced at the Cornell Biotechnology Institute. Cultures were grown in 1/20
LB in MSM and 1000 mg/L lactose at volumes of 200 µL in 96 well plates and the vector was
maintained with 150 mg/L ampicillin. Cultures were supplemented with various doses of IPF or
analogs and maximal cell growth was measured spectrophotometrically at 600 nm over the
course of two days, at which point the percent growth inhibition was determined (Figure A-5).
Percent growth inhibition values from these curves were used to construct models to predict
effective concentrations for 50% (EC50) growth inhibition using BioDataFit 1.02
(www.changebioscience.com). Models for IPF and 2(4T)PA were four parameter, while models
for 4TAA, 2PPA, PAA were linear.. The analogs were not particularly toxic in the absence of
ipfF so we did not have enough data points to construct models for analog toxicity in the absence
of ipfF.

107
Figure A-5. Growth Inhibition curves for E. coli JM109 in the presence of IPF analogs with and without heterologous expression of the ipf-CoA-ligase (ipfF). Percent of growth inhibition is shown, means and standard deviations, n=3

108
A.6. Culture growth conditions have an effect on toxicity
Work with a high throughput system was initiated to investigate the toxicity of upper
pathway metabolites in E. coli strains, however further work demonstrated that strain Ibu2
responds differently to stress from IPF and its analogs when grown in this manner. We should
not expect to see greater toxicity towards 4TAA, 2PPA, and PAA when cultures are grown in 96-
well plates and this is described below. All previous work with strain Ibu2 had been done using
50mL conicals with the use of rotary shakers for aeration of the growing culture. Under these
aerated conditions, IPF and 2(4T)PA do not inhibit culture growth, however 4TAA, 2PPA, and
PAA do (Figures A-6A and 4-2). We modified our approach to culture 200 µL volumes in 96-
well plates for high throughput; exponential phase cultures of strain Ibu2 grown on glycerol were
resuspended at a starting density of A600=0.08 in MSM with 0.1% glycerol and 250 mg/L IPF or
analog. Under these low aeration conditions, strain Ibu2 grew slower and all analogs inhibited
Ibu2’s growth (Figure A-6B). While 4TAA, 2PPA, and PAA appeared to be slightly less
inhibitory under the low aeration scenario than the high aeration scenario, the concentration was
less in the low aeration scenario (250 mg/L vs. 500 mg/L). Because all analogs are toxic when
strain Ibu2 is grown in 96 well plates, we should not expect that experiments using 96-well
plates will show more drastic growth inhibition by 4TAA, 2PPA, and PAA, however they may
still be useful in identifying toxic metabolites (Figures 4-7 and 4-8). Engineered strains Ibu2 pP,
pC and pC+P from Chapter 4 and as 4TAA mutants from Chapter 3 were also grown under these
conditions and they all yielded comparable results with lower growth rates for all analog
treatments (data not shown). It has been suggested by Fuchs that hybrid pathways may be
beneficial under microaerophillic conditions because both aerobic and anaerobic biodegradation

109
pathways begin with CoA and metabolism could proceed if conditions changed (Fuchs, 2008).
Given that the ipf pathway requires molecular oxygen prior to the release of CoA, this growth
inhibition could be in part due CoA depletion, however this was not investigated. Inhibition
could also be due to the accumulation of other pathway metabolites, as discussed in Chapter 4.
Figure A-6. Inhibition of growth by IPF and analogs in strain Ibu2 under different growth conditions. Cultures were grown on glycerol and (A) supplemented with 500 mg/L analog, grown in 50 ml conicals, and shaken at 150 rpm for aeration, or (B) supplemented with 250 mg/L analog and grown in 96-well plates with no aeration. Cultures were supplemented with no stressor (Control), or analogs IPF, 2(4T)PA, 4TAA, 2PPA or PAA. Means and standard deviations, n=3.

110
A.7. Strain Ibu2 sequence and similarity with sequences in the NCBI databank
Strain Ibu2 genes ipfABDEFG were published in the NCBI data bank by Robert Murdoch
and Anthony Hay. We were curious if we could find lower pathway genes so additional
sequencing was performed. DNA from fosmids 3G7 and 4F6 was sequenced and analyzed with
DNAstar as described in Appendix A.3. The resulting contig sequences were compared to other
known sequences in the NCBI databank using BlastX. This analysis was done simply to gain an
understanding of the genetic environment, and may include genes that are not transcribed into
functional proteins. A visual overview of our current best guess of the genetic environment of ipf
and related genes is shown in Figure A-7. The sequence analysis is shown in Table A-7 and
contig sequences are provided following the table. There are a few caveats that limited additional
clarification of this sequence. For example, there were multiple copies of transposable elements
and ipf genes, which made additional alignments difficult and uncertain. With respect to ipf
genes, one copy of ipfFG is show in contig 5 immediately adjacent to the T7 end of the
pCC1FOS cloning vector and ipfF is truncated by the vector. An additional copy of ipfFG is
located within the ipf operon on contig 1. Transcription of the ipf pathway genes was also
investigated using RT-PCR, as described in Appendix A.8.

111
Figure A-7. Five contigs of strain Ibu2 DNA from fosmid clones 3G7 and 4F6. Upper ipf pathway genes are shown in black. The ipf pathway consists of an ipf-CoA-ligase (ipfF), a ring hydroxylating dioxygenase (ipfAB) with ferredoxin reductase (ipfH) and ferredoxin (ipfI), a putative thiolase/acyltransferase (ipfD) which may act with a protein of unknown function (ipfE), a 4-isobutylcatechol-2,3-dioxygenase (ipfJ), and a plant-like ferredoxin (ipfK).Genes involved with MC activity (ipfJ and ipfK) and putative MC lower pathway genes are shown in dark gray. Transposable elements (Tn) are shown in light gray. Sequence analysis is provided in Table A-7.

112
Table A-7. Sequence similarity between strain Ibu2 and sequences in the NCBI databank.
Name Gene & Species Bp on contig
Aligns with bp in gene
Identities Positives
Contig 1 Tn6 Transposase for insertion sequence IS6100,
Escherichia coli 484-41 1-148 of 266 129/153 133/153
Integrase core domain protein, Escherichia coli MS 57-2
478-41 1-146 of 230 128/151 131/151
IS6100 transposase, Sphingomonas sp. XLDN2-5 478-41 1-146 of 164 128/151 131/151 ipfJ Glyoxalase/bleomycin resistance protein/dioxygenase,
Polaromonas naphthalenivorans CJ2 844-1719 1-292 of 292 282/292 289/292
2,3-dihydroxybiphenyl-1,2-dioxygenase, Burkhoderia xenovorans LB400, Pseudomonas sp., Archromobacter sp.BP3
844-1719 1-292 of 298 262/292 275/292
Dioxygenase, Pseudomonas pseudoalcaligenes, Pseudomonas aeruginosa, Pseudomonas putida
844-1719 1-292 of 298 261/292 274/292
4OI 4-oxalocrotonate tautomerase family enzyme,
Alcycliphilus denitrificans BC, Alcycliphilus denitrificans K601
1852-2019 8-63 of 63 51/56 53/56
4-oxalocrotonate tautomerase, Acidovorox sp. JS42 1852-2019 8-63 of 63 50/56 53/56 4-oxalocrotonate tautomerase, Uncultured bacterium 1852-2019 8-63 of 63 50/56 53/56 ipfA Putative ibuprofen CoA dioxygenase large subunit,
Sphingomonas sp. Ibu2 2091-3362 1-424 of 424 424/424 424/424
ipfB Putative ibuprofen-CoA dioxygenase small subunit,
Sphingomonas sp. Ibu2 3490-4023 1-178 of 178 178/178 178/178
ipfD Putative thiolase/acyl transferase, Sphingomonas sp.
Ibu2 4083-5231 17-399 of 399 383/383 383/383
ipfE Unknown, Sphingomonas sp. Ibu2 5247-5642 1-132 of 132 132/132 132/132 ipfF Ibuprofen CoA Ligase, Sphingomonas sp. Ibu2 5850-7250 1-527 of 527 527/527 527/527 ipG Putative periplasmic binding protein, Sphingomonas
sp. Ibu2 7350-8486 1-379 of 379 379/379 379/379
Contig 2 HOA 4-hydroxy 2-oxovalerate aldolase, Novosphingobium sp.
PP1Y 193-396 1-68 of 343 61/68 62/68
501-1211 103-341 of 343
205/239 219/239
4-hydroxy 2-oxovalerate aldolase, Sphingomonas sp. 14DN-61
193-396 1-68 of 343 61/68 62/68
501-1211 103-341 of 343
205/239 218/239
4-hydroxy-2-ketovalerate aldolase, Novosphingobium aromaticans
193-396 1-68 of 343 59/68 63/68
501-1211 103-341 of 343
203/239 218/239
4OD 4-oxolocrotonate decarboxylase, Sphingomonas
sp.14DN-61 1257-1805 20-202 of 272 143/183 162/183
1805-2014 203-272 of 272
53/70 59/70

113
Name Gene & Species Bp on
contig Aligns with bp in gene
Identities Positives
4-oxolocrotonate decarboxylase, Sphingobium yanoikuyae
1257-1805 4-186 of 256 143/183 162/183
1805-2014 187-256 of 256
53/70 59/70
4-oxolocrotonate decarboxylase, Novosphingobium sp. PP1Y
1257-1811 4-188 of 256 143/185 161/185
1808-2014 188-256 of 256
55/69 59/69
4OI 4-oxalocrotonate tautomerase/isomerase, Sphingobium
yanoikuyae 2018-2200 1-61 of 70 36/61 49/61
4-oxalocrotonate tautomerase/isomerase, Novosphinobium sp. PP1Y
2018-2200 1-61 of 70 35/61 49/61
4-oxalocrotonate isomerase, Sphingobium chungbukense
2018-2200 1-61 of 77 38/61 48/61
ipfK Ferredoxin-like protein, Novosphigobium
aromaticivorans 2245-2505 6-92 of 93 57/87 70/87
Plant-type 2Fe-2S ferredoxin, Sphingobium chungbukense
2245-2505 6-92 of 93 53/87 71/87
Ferredoxin-like protein, Sphigobium yanoikuyae 2245-2499 5-89 of 92 50/85 67/85 Tn Hypothetical protein, alpha proteobacterium BAL199 3621-2671 1-318 of 338 216/318 238/318 Hypothetical protein, Acidophilium multivorum
AIU301 3606-2605 9-342 of 342 218/335 244/335
ISSod11 transposase , Nitrococcus mobilis Nb-231 3618-2656 4-337 of 342 198/334 230/334 ipfH Ferredoxin reductase subunit aromatic oxygenase,
Novosphingobium aromaticivorans
3964-4626 58-278 of 408 142/221 166/221
4629-5006 280-405 of 408
41/126 89/126
3863-3964 24-57 408 24/34 27/34 3801-3848 3-18 of 408 12/16 14/16 FAD-dependent pyridine nucleotide-disulfide
oxidoreductase, Novosphingobium sp. PP1Y 3964-4626 62-283 of 413 136/221 166/221
4629-5006 285-410 of 413
72/126 87/126
3860-3964 28-62 of 413 24/35 28/35 3792-3848 5-23 of 413 12/19 16/19 Naphthalenesulfonate dioxygenase reductase subunit,
Sphingobium xenophagum
3961-4626 57-278 of 409 135/222 168/222
4629-5018 280-409 of 409
73/130 91/130
3863-3949 24-52 of 409 22/29 25/29 3801-3848 3-18 of 409 15/16 15/16 Tn transposase B1, Sphingopyxis macrogoltabida 5544-6365 13-288 of 348 240-276 252/276 transposase B, Sphingomonas sp. KA1 5706-6365 5-224 of 244 200/220 210/220 Transposase, Granulibacter bethesdensis CGDNIH1 5700-6365 5-226 of 286 193/222 209/222 Tn3 Transposase Tn3 family protein, Sphingomonas wittichii
RW1 7074-8141 1-356 of 874 355/356 355/356
Transposase, Mesorhizobium loti MAFF303099 7044-8141 2-367 of 980 276/366 304/366 Transposase Tn3 family protein, Sphingomonas wittichii
RW1 7074-8141 1-356 of 969 273/356 298/356

114
Name Gene & Species Bp on
contig Aligns with bp in gene
Identities Positives
Contig 3 C23O 2,3-dihydroxybiphenyl dioxygenase, Beijerinckia sp. B1
Sphingomonas paucimobilis, Sphingobium yanoikuyae, Sphingomonas sp. P2, Sphingomonas sp. DN1
1000-1641 81-298 of 299 123/219 138/219
Glyoxalase/bleomycin resistance protein/dioxygenase, Novosphingobium sp. PP1Y
81-298 of 299 119/219 136/219
2,3-dihydroxybiphenyl 1,2-dioxygenase , Sphingobium chungbukense
81-299 of 300 116/219 135/219
ipfI Reiske (2Fe-2S) domain-containing protein ,
Novosphingobium sp. PP1Y 1726-2055 1-108 of 108 72/110 87/110
Ferredoxin component of dioxygenase , Sphingomonas yanoikuyae B1, Sphingomonas sp. P2, Sphingomonas sp. DN1
1726-2028 1-101 of 108 67/101 81/101
Reiske-type ferredoxin, Sphingobium chungbukense, Sphingomonas sp. CHY-1, Cloning vector pSKY4-Km24 megaplasmid, Sphingomonas sp. LH128
1726-2055 1-108 of 108 68/110 82/110
ATPase
ATPase Polaromonas sp. JS666 2995-2435 203-389 of 392
110/187 142/187
3392-2970 72-211 of 392 110/187 142/187 Tn6 Transposase, Arcanobacterium pyogenes 3738-4517 1-260 of 264 260/260 260/260 Transposase, Corynebacterium diphtheriae NCTC 13129 1-260 of 264 259/260 260/260 IS6100 transposase, Sphingomonas sp. XLDN2-5 1-260 of 264 259/260 259/260 Res Resolvase, Sphingomonas sp. KA1 4962-5390 1-145 of 189 117/146 124/146 Resolvase domain-containing protein,
Novosphingobium sp. PP1Y 1-145 of 189 116/146 122/146
Fis family transcriptional regulator, Sphingobium chlorophenolicum
1-145 of 189 116/146 123/146
Contig 4 C23O Catechol 2,3-dioxygenase, Sphingomonas sp. A8AN3 51-170 269-308 of
308 34/40 37/40
Glyoxalase/bleomycin resistance protein/dioxygenase, Novosphingobium sp.PP1Y
51-170 268-307 of 307
32/40 35/40
Catechol 2,3-dioxygenase, Sphingomonas sp. B2-7 51-170 268-307 of 307
32/40 35/40
CmpX Hypothetical protein, Novosphingobium
aromaticivorans 320-508 47-109 of 143 37/63 50/63
Unknown, Sphingomonas sp. / phnF Sphingobium chungbukense
320-508 47-109 of 143 37/63 50/63
Hypothetical protein, Sphingomonas sp. P2, Sphingomonas yanoikuyae, Sphingomonas sp. DN1
323-508 48-109 of 143 37/62 48/62
HMSD Aldehyde dehydrogenase, Novosphingobium sp. PP1Y 672-2120 9-494 of 494 378/486 425/486 2-hydroxymuconic semialdehyde dehydrogenase,
Sphingobium yanoikuyae 657-2120 4-494 of 494 378/491 426/491
2-hydroxymuconic semialdehyde dehydrogenase, Sphingomonas sp.
672-2120 9-494 of 494 377/486 424/486
OEH 2-hydroxypent-2,4-dienoate hydratase,
Novosphingobium aromaticivorans 2144-2695 1-184 of 264 142/184 163/184
2-hydroxypent-2,4-dienoate hydratase, Sphingobium chungbukense
2144-2695 1-184 of 264 132/184 157/184
Putative 2-hydroxypent-2,4-dienoate hydratase, Sphingomonas sp. P2, Sphingobium yanoikuyae
2141-2695 4-188 of 270 132/185 158/185

115
A.7.1. Contig sequences
Contig 1 AATTCGGTGAATGCTGCACCAAAGCTTGGCGCTTTTGTCGGGTATTGAGCCGTGGCAGCTTTTTTCCCAAGTGTTTCAGGCTTCGCAAGGGCCTGGCCCAGGAACCGCTTCGTCTTGGCGCTGGGGGTCGGCACAGGTAGAAATCGATCGTGTCGCCCCGCTTGTCGACTGCCCGGTACAGGTAGGTCCACTTGCCCCGCACCTTGACGTAGGTTTCATCCAGGCGCCAGCTCGGATCAAAGCCACGCCGCCAGAACCAGCGCAGCCGCTTCTCCATCTCCGGGGCGTAGCACTGGACCCAGCGATAGATCGTCGTATGGTCGACCGAAATGCCGCGTTCCGCCAGCATTTCCTCAAGGTCGCGATAGCTGATCGGATAGCGACAATACCAGCGCACCGCCCACAGGATCACATCACCCTGGAAATGGCGCCACTTGAAATCCGTCATCGTTCCGTCCGTCCAATCTCCGCCAAGCATGCTCAAGCTTCACGATTTTTGCAACAGAGCCCGGCCTGGGATTCATGAATCTCAGCACGCGTGCGTGGGGTAGTGCGTGCCATCGGATGGACCATACTCATGATGCGTTCTCCTCAGTACCAGAGGCCAGAAGGCCCGCCAAAACAGCACGGGCCCGAAACAACGGCCAGCTGCGACGGGGTACATAATCGCACGGAACTCGACACCTAGGGTTTTTCCCTATCGTAAAAATGTCGAATGTCGATTATATTATCGAGATTATGACGAGACACACGTTGCTGAGCCTTGATTCGGTCCATGCTGAAGTGTGGCGAAATCTTCTGTAATGACCAAGTGAAAAATACAATGAATGCGAGAACTTCAAAATGAGTATCAAAAGCTTGGGTTACATGGGATTCGCGGTACAGGATGTGCCCGCCTGGCGTTCTTTCCTGGCCCAGAAATTGGGCTTGATGGAAGCAGGCGTCACCGATGACGGCGCCTTGTTTCGCATCGATTCGCGCGCCTGGCGGATTGCCGTCCAGCAGGGCGAGGCAGACGATTTGGCCTACGCTGGCTACGAGGTGGCCGATGCAGCGGGTCTGACGCAGATGACAGAAAAACTGCGGCAGGCCGGAATCGAAGTGGTCACGGGCGATGTCGAACTGGCCAAGCGGCGCGGCGTGATGGGCTTGATTTCGTTTGCCGATCCGTTTGGTCTGCCACTGGAAATTTATTATGGCGCCAGCGAGGTGTTCGAAAAACCGTTCATGCCCGGCGCGGCGGTGTCGGGTTTTCTGACCGGCGAGCAGGGGCTGGGTCATTTTGTGCGCTGCGTGCCGGATTCGGACAAGGCGCTGGCGTTTTATACCCAGGTGCTCGGCTTCCAGTTGTCGGACGTCATCGACATGAAAATGGGACCGGACGTGACGATTCCGGCGTATTTTCTGCACTGTAACGAACGCCACCACACCCTCGCGATCGCAGCTTTTCCGCTGCCCAAGCGCATTCACCACTTCATGCTCGAAGTCTCGTCGCTCAATGACGTCGGTTTTGCGTTTGACCGGGTTGACGCTGACGGCTTGATCACCTCCACGCTGGGGTGCCACACCAATGACCATATGGTGTCGTTCTATGCAGCGACCCCGTCCGGAGTCGAGGTCGAGTACGGCTGGGGCGCCCGCACCGTTGACCGCTCCTGGGTGGTGGCACGGCACGATAGCCCGAGCATGTGGGGCCATAAGTCGGTGCGCAACAAAGCATAAACAACAAGCTGACCCACACAGGAGTTTTTGAATGAAGCTTTACTACAGCCCTGGCGCCTGCTCCCTGTCCCCGCACATCGCCTTGCGCGAGGCGGGCCTGAACTTCGAGCTGGTACAAGTCGATCTGACATGATTGAAGGTCGCACCGAAGAGCAAAAACGCGCGGTGATCGAAAAGGTCACGCAGGCGCTGGTTGACGCTGTCGGCGCGCCCAAGGAAAACGTCAGAGTCTGGATCCAGGACGTGCCCAAGGAGAACTGGGGCATTGCCGGGGTGAGCGCCAAGGATCTCGGACGCTAGCTGGCGCGCGGCCCGGGAAGGAAATACTTTCAGCGCCAGTTTTTACACTATTTTTATGGAGAATCAAAATGGCAAGTACACAAACGATCACCATCACACGCAAGGCTCCTGACGTTGATGTCGGTGCCTTGATAGAGCGAGACCGTATTCACGGCAGCTTGTACGCGAACGAGTCGATATTCGAGCTCGAAATGAAGAAAATATTCTACGACGGCTGGGTTTTCGTTGGTCACGACTCCGAAGTGCCAACGGCCGGCGAATACGTGCGCCGCACGCTGGGCCGAGAAGAGGTCTTGATGGTGCGCCAGCGCGATAGCTCCATCGCCGTCATCGCCAACCGTTGCGCGCATCGCGGCAACATGATGTGCATCGCCAACCATGGCAAGGAAAAATACTTCACCTGCACCTACCACGGATGGGTCTATGATTTGGCCGGAAACCTGAAGGACGTTCCCTATCCCGGCGGGTTTGACAAGGACAAGTCGGAGCTGAAGCTGCAGCCCTTGCGCACTGAAGTCTATCGTGGCTTCGTGTTCGCCACCTTCAATGCTTCGGCGCCACCCTTGATGGAACAACTGGGGCGAGGAAAAATCCTGATCGACCGCGCCTGCGACATGTCGCCCACCGGTCGCCTTCAGCTCACGGCGGGCTGGACCAAGCAGCGTTTCGGTGCCAACTGGAAGATGCTGCCCGAGAACGACACGGACGGCTACCACGTCAACGACGTGCATGCTTCCTTTGCCCAAGTGATCGATTCGCACTATGACAGCGCCGCGATTGCTGCAGAAGACAGCCTGCGCTCCCAAGCCAAGGATTGGGGTAACGGGCATACAGAATTGTATCTCTCCCCGACCTACACCGAGTATCTCAAGTGGTTCAACACCACGCCGAATCGTTTTCCGGAGTACATCGCGCAGATGAAGGCCGCCTACGGCGAGGAAAAGGGCGACAACATTCTGCGAGATGGCCCGCCCCACGCCACGATCTTCCCTAACCTGTTCCTCGGCGAGATGAACATCATCATTTTTCTGCCAATCAACGCCCATGAGTGCGTGCAATGGCACACCCCGATGCTGCTTGAAGGCGCTCCTGACGAAGTCAACCAGCGCATTATCCGCAACTCCGAAGCGGCGATGGGCCCCTCGGCCTTCCTGCTTGCCGACGACAGCGTGATTTCGGAGCGCCAGCAAATCGCCTTGCGCGACCGTGCCGACTGGCTGGACGTGTCGCGCGGTCTGAACCGAGAGCATGTCGATGAGATGGGCGTGGTGGTGGGGCACGTGACCGACGAGTGTACCAACCGCGGCTTTTGGCAGCACTACAAAAAGGTGATGACAGCCCCTTCGCCGTCCCCTGTTTGAGCGTATTGATCACCACCTCATCCGCCCCCCTTCGGGGGCGGCGGATTCACTGAAATTTCGTGTTTAAAAAAGCGTCCTGCGAGCAG

116
GACGCGCAAGGAGATACATCATGACAAGCATCACAGCCGCTGTCGGCACCAAGCAAAAGACTGACGTGATCGGGCTGGAAGAGCACCGCGAAGTCTGCGACTTTTTGTACCGAGAGGCGCGCCTGGCTGACGAATCGCGTTACGCCGAATGGGAGGCGTTGGTTGAGGACGACATGACCTATTGGGTTCCGCGCGGCGAGGGTGATTACGACATGAACAAACATGTGTCGATTACCGCGGACAACCGTTCCCGCTTGCGCGTCCGCATCGCGCAGTTGATGACAGGCAAGCGCCATGCCCAACTGCCGGTGTCGTCGATGCGGCGGATCGTCAGCAACATTGAGGTAGAGCATCATGCGCAGGGCGGCTACCGCGTACTGTCGAACTTTGTGCTTTACGAATTGCGGCGCTCATCGACAGGTCAGATAGAAGTTTGGCCGGGGCGGGTGGAGCACCATCTGCGCCGCCGAGCGGATGGCTCGCTGGGCATGTTCTTCAAGAAAGTGGTGCTGATCCATGGGGACGAGGCTGTTCCCAGTCTGGCATTCATCATCTGACGCGCACCATGGTCACGCGAGTGAAGAAAAAGACCAGGGCCAGAGCCGCGATTGCCGGGCTGGGTTTCAGTGCGATGTCACGCCAGCCCGTCGGCACGATCCGCGAGTTGGCGGCCACTGCGGTGGCCGCCGCTGCCGCTGATGCGGGTCTGCGCCTGCAAGACATCGATGGCTTGTTGCTCAACAAAAGCCCGGCAGCAGAGCCTGAAGAGTTGCCCTTGCGTCTGCAAAACGACCTTGGGCTGCGCGACCTGGGGCTGCTGGCTGCGATGGATTCCGAGGGCTCGACAGCGGTGCAGATGGTGCAGTATGCAGCGATGGCGGTGCGTGAAGGACTGGTCAAGTCGGTGGCCTGTGTGTTTGCCGACACCCCGCTCAAAGGATCGGGGGCAGGTGGGGGTGATGCCTTTGCTCTGGCGATGCCGCTGACCGGTGTCGAGGGTTGGGAGGCCCAGCAAGGCTTCCTGGGGGCCACCGCCGCCTATGCGTTGGCCGCGCGCCGTCACATGGCGCTCTATGGCAGCACGGCCGAGCAGCTCGGCGCCTATGCGCTTGCTTGCCGGCAATGGGCGGCGCTGAACCCGCAGGCCTTCCTGCGCAAGCCGATGACGATGGACGACTACCTCGCATCGCCTTTCGTCGTGGAGCCGTTTCGCGTGTTCGACTGCTGCTTCCCGGTCAACGGCGCTGTCGCGCTCATCGTCACCTCCGCCGACCGTGCTGTTGATGGGCCTCAGCCGCCCGTTTTCATCCACGGCATGGGCCAGGGGCACCGCGGCCGCAGTGGCCTGAGCGGCGACGATCCAGAGGTCTTCACGGGTGCCATCCAGGCCGGCCAGACCGCTTACCGTAGTGCCGGGGTGAATGCCAGTGATGTGACTCAGTGCCAGTTTTACGACGCTTTTTCTTACGCCGGCATCCTGGGTCTGGAGGCATATGGGTTGTGTCCCCGGGGTGAGGGCGGGGCCTTCGTCGCGCAGGGCCACACCGCGCCCGGCGGCAAGTTGCCGGTCAATACCGGTGGTGGGCACCTGTCGGGCTTTTACCTGCAGGGCATGACGCCGCTGTCGGAGGCGGTGATCCAGGCACGCGGTGCCGGCGGCGCGCGGCAGGTCGTGCGCAACGACTTGATTCTGGTGACCGGCAATGGTGGCTGCCTTGACTACCACACCTGCGTACTGGTCAGCCCACACCGCACCCTCGCCTGACTGGAGCCATTGATGGATAGCACCTTTTTTCACCGCTACGACGAGGCATTTCTCGCATTCATCGCTGCTGGCGAATTGCGCATTCCCGTGCATACCGAAACCGGCCGGGCGCTGGGTCTGCATCAGCGTGCCTGGTGTGTCGCTGGAGATCACGGCGTGCAATGGAGGCCCGCGTCTGGCCGCGGGGCTGTTCTCTCTTTTACCGTGACGCGTCGGCCCTACACGCCCGAATTTCCCGTGCCTCTGGTGCATGGATTGATCGAGCTGGCCGAGGGGCCGCGACTGATATGCCGGCTTGACGGCGTCACGCCCGAGGCGGTCGCCGTCGGTCAGGCGGTCCAGGCGCATTTCGACCGACAAGGTCTGGTGTTCCGACCGGCCCTTGACGACGGCAATAAGGCCGACAAGTGAAACTTAAAACCAAGGAGACAAGTTATGTTGGCAAGAGACCTGGTTAAGCGCTGTGCGCGTAACTACCCGACCAAGACCGCCTACCTGTGCGGCGAACGCTCGCGTAGCTGGCGCGAGATGGATCAACGCTCCGACCGGTTCGGCGTGGCCTTGCAACAACTCGGCCACCGTCCGGGCGAGGCGGTGGCCATCCTGACGCAAGAAAGCATCGAGGTGTATGAGCACTTCTTTGCCTGCATGAAGATCGCGGCGCCGCGGGTCGGGCTCAATACAGGGTACGTCTGGCCCGAGATGCTGCACGTGCTGAAGGACAGCGAGGTCAAGTTCCTGCTGTTGGATACGCGCTGCCGGCATCTGCTCGCCGAGCGGCTGGGCGAGCTCAAGGCACTGGGCATCACGCTGATTGGCTATGGTGCAGGCCATGGTCTGGAGCGCGACTACGAAAGCTTGCTGGCGACAGCCGAGGGTGAGCCGCACTGGCCAGCGTTGGCGCCCGACGACATCCTTTTTGTCAGCTACACCTCCGGCACGACCGGTGTACCCAAGGGCGTAATGCTCACGCAGGAGGGCGGTGTCAACTGCATCCTCCACTCGCTGATTTCTTTTGGTTTTGGACCTGACGATGTCTGGTACATGCCGGCGGCATCGGCCTGGGTGGTCGTGATACTGAACGCATTCGGGCTCGGCAACGGGATGACGACCGTGATTCCGGACGGCGGATACCAATTACAGGCTTACCTGCGCGACATCGAGCGTTTTCGCGTTACGGTCGGGATGCTCGTCCCGACCATGCTGCAGCGCGCGATTGTTGAAATCCAAACCAATCCAGTTTACGACCTGTCCTCTCTGCGCATGGTGGTGTACGGCTCGTCGCCCGCCACGCCCAAATTGATCCGTGATGCGAGGGCGACCTTCAAGGGGATCAAGCTGCTGCAGGCCTACGCGATGACAGAAGCCACCGGCGGCTGGATCAGCTACCTGACCGATGCCGACCACGAGCATGCGTTGCGCGAGGAGATCGAGCTGCTCAAGTCGGTCGGCCGCATCGGCATTCACTACGACTGTTCGATTCGCGACGAGTCGGGCCAGCCAGTGCCGATCGGTCAATCCGGCGAGATCTGGCTGCGTGGCAATACCATGATGAAGGGTTACCGCAACCTGCCCGAGGCTACGGCGGAAGCGATGCCGGACGGCTGGCTGCGCACCAACGACATCGGGCGGCTCGACGAGCGCGGCTATCTGTATCTGCTGGATCGGCAGAAGTTCCTGATCATTACCGGCGCGGTCAATGTGTTTCCAACCACGGTCGAGGCGATTCTGGTCGAGCACCCGGCGGTGGAGGAGGTGGCCGTGGTCGGCGTGCCGCACCCGGAATGGGGCGAGGCGGTAGTCGCTGTGGTCGTCCGAAAACCTTCGCACCGCGATGTGACGGTGCAGGCGCTTATCGATTTTTGTCACGGCAAGCTCAGTCGCCCCGAAACGCCCAAGCATGTGGTGTTTGTCGATGAACTGCCCAAGACATCCAACGCCAAGCTGAAGAAAGGCGAGTTGAAGAAATGGCTGTCTGGCGGTGCAGTGCCCCTTCCCTGGCAACTCGAAGTTGCTTGACCTTGTAGTGCCCCGTCAAGCAC

117
CTCTAAAAATTCCCCCTGCTTAAAACTTGTAACCCTAGGAGACAAAAATGTTGAAACTACGCAACCCCATGAAATGTTTGATGGCTACCGCCGTCGTGTGTGTGTCCGTCTTGTTGCCGGCGGCAGCTCAAACCAAGCCGCCGTTGAAAATCGGTGCCTATCTGTCCGTCACTGGCCCTGCTTCTTATCTGGGCGCGGTGCCATGAAAACCGTCGAGATGTACGTCGAAGCCATCAACGCCGCTGGCGGCATCGATGGCCGCAAAGTCGAGTTCCAGGGCTACGACGACGAATCGGATGCATCGCGCGCCAATACGCTGGTCAAGCGTCTGATCGAGAACGACAAGGTTCACGTCATCATTGGCGGCTCTACGACAGGCGCGACGATGTCAGCCGTCCCGCTGGTGGAACGGGCCGGTATCGCCATGCTGTCGCTGGCCGGTGGCAGTGTGATCACCGACCCTGTCAAGAAGTTCGTTTTCAGACTGGCCCACAACGATGCGATGGTAGTCAGTCGTCTGTACGACCACATGCTGGGTCGCGGTATCAAGAGCATCGGCATCATTGCCGGTAGCGATGCGTTCGGTCGATCCTGTCTCTCATTCGCGCAGAAATTGGCACCGGGAAAAGGCGTCAAGATTCTGCGCGATGAGTCCTACAACGCCAAGGACACGGACATGACCGTACAGCTGACCAAACTGCGACAGGAACCGGGTTTGCAGGCGATCTTCAATTGCGGCTTCGGCGAGCCGGCGGCGATTGCCACCAAGAACTACAAGCAACTCGGCATCACCGTGCCGCACTACGGAACCCATGCGCTGGCGTCGGACGCTTTCGTCAAATTGGCGGCCGGCGCGGCGGAGGACATGATCATGGCCAACGGCCCCATCCTTGTGTGGGATCAATTGCCGGCCAGCGACCCCCAGCAACCGGTGGTGCAGTCCTATGTGAAGGCCTACCGCGCCAAATTCAACGAAGCCCCTTCCTTCATTGCGGGTATTGCACATGACTCGTTCTTCGCCATCCGGGAGGCGGTGCGGCGCAGTGGCAGTATCGAGCCCGCCAAAATTCGCGATGCCATCGAAAAAGGCAACGCCTTTGTGGGTGTGACAGGGCATTTCCGGATGATGAACGAGACCGACCACATTGGCTTGACCCCCGACTCCTTGCGGATCGTACAGGTCAAGAATGGTCGATGGAAACTCGTTGAATAAGTGTAATGGCT Contig 2 TTCGGATCGAACAACCCGGTCAAGATCCCCGGTTATGGGAGTTCGCGGGGGCTCAAGACGAGCGTCTTCCTGAGGTCGAAGGAGCGGGCGACTATCTCCCCAAATATGCGGGCAATCTGACATCATGACCGCGGCCGCCAAGGCCGCCGGCGAGATGCTGGCGGCCCGCATGCTCAATCGGAAGGCTGCAGCATGACGCTCTATCCCACCCAGACCAAGCTCTACATCCAGGACGTGACCTTGCGTGACGGCATGCACGCGATCCTGCACAATTATGGCACCGAAAGTGTCCGCACGATCGCCAAGGCGCTCGACGATGCCGGTGTCGACGCGATCGAAGTGTCGCATGGCGACGGGCTCAACGGCAGCTCGTTCAACTATGGCTTCGGCGCGCACTTAAATTGCACTGAAATCTAGAAATATTTTATCTGATTAATAAGATGATCCCCGGGTACCGAGCTCGAATTCATCGATGATGGTTGAGATGTGTATAAGAGACAGCTTATGCGCTCGGCGTGCGCTCGGTGCGCGTCGCGACGCACTGCACCGAAGCCGACGTCGGCAAGCAGCATATCGGCATCGCGCGCGATCTCGGCATGGACGTGTCGGGCTTCCTGATGATGAGCCACATGCTCGAGCCCGAGGCGCTCGCCCAGCAGGCGTTGCTGATGGAAAGCTATGGCGCGCATTGCGTCTATGTGACCGACAGCGGCGGTGCGCTCGACATGGATGGCGTCCGCGCCCGCCTGCAGGCCTATGACCGGGTGCTCAAGCCCGAGACGCAGCGCGGCATGCACGCCCACCACAATCTCTCGCTCGGCGTCGCCAACTCGATCGTTGCAGCGCAGGAAGGTGCGATCCGCATCGACGCCAGCCTCGCTGGCATGGGCGCGGGCGCCGGCAATGCGCCGCTCGAGGTGTTCATCGCCGCGATCGACCGCAAGGGCTGGAAGCACGGCTGCGATGTGATGGCGCTGATGGACGCCGCCGAGGATCTGGTCCGGCCGCTTCAGGATCGCCCAGTTCCGCGTCGACCGCGAGACCCTGAGCCTGGGTTATGCGGGCGTATCGCGTCTGCCCACGCCGAAAAGGCCGCCGAACAATACGGACTCGACACACGCGAAATCCTGGTCGAACTTGGCCGGCGCCGGATGGTCGGCGGACAGGAAGACATGATCGTCGACGTCGCTCTCGATATCCTGAGCGCGCGCAAGGGGTGAGACCGAGCCCAAGGGGGAAATGGCCGCGTGACTTCGCTTGCTGAATATGCCGAGATTCTCGACCGTGCCGCGCACGAAGCGCATGCCACGCCTCAGATCACCCACAGCAATGACAAGCTGACCGTCGCCGACGCTTATGCGATCCAAAAGCTGTCGGTCGAGCGCCGCCTGGCGCGCGGCGAGAAGCGGATCGGCGTCAAGATGGGCCTCACCAGCCGCGCCAAGATGCAGCAGGTCGGGGTCGACGAGGTCGCCTGGGGGCGGCTCACCGACGCGATGCTGCTCGAGGAGGGTGCGGCGCTGTCGCTGTCGCGCTTCGTCCATCCGCGGATCGAGCCGGAGATCGCCTTTCTGATGAAGGCGCCGCTCGCGGGCAAGGTCACGGCGGCGCAGGCGCTCGCCTGCGTCGAGGCCGTCGCCCCAGCGATGGAAGTGATCGACAGCCGCTTCGAGAATTTCAAGTTCGCGTTGGTCGACGTGGTCGCCGACAACACCTCGTCTTCGGGCCTGGTGGTCGGCGGCTGGGGCGATCCGATGCAGGATCTCTCGAACCTCGGCGTGATCCTGGAAATCAACGGCGAGGTCGTGAGGTCGGCTCGACTGCCGCGATCCTCGGCCATCCGTTGCGCTCGCTGGTCGCCGCCGCGCGGTTGATCGGCGAGGCGGGCGAGACGATCAACGCCGGCGACATCGTCATGGCCGGCGGCATCACGGCGGCGCCGACCCTCAAGGCCGGCCAGACCATCCGCAACACCGTCCAGAACCTGGGATCGGTCAGCATCACGGTGGAAGCATAAATGCCGATCATGGAAGTCACTTTGGTCGAGGGCCGCACGACCGAGAACAAAAAGGCGTTCATCCTCGCGCTGACCGATGCCGCGGAGCAGTCGCTCGGCGTGCCTCGCGAGTCGATCCGCGTGATCCTGCGCGAGATCCCGGATGCGAATTTCGCGGTGGCGGGCGTGACTTTCGCCGCGCGCAAAACCAATACTTTGAAAGCGGCAAGCTGACCACGATGATCAAGCATCAAGTCCAGATCGTCGATGGCAGCCGCTTCGAATGCCCCAAGGAGGAACGGGTCCTGATCGCGATGGAGCGCTTGGGGCTGAATGACATTTTCGTCGGCTGCCGCGGCGGCGGGTGCGGCGTGTGCAAGGTGAAGGTGACCAGCGGAAATTATCGCACCGGTAAAATGAGCAGGCTACAAGTGAGCGAGGCG

118
GAAGAGGCGGGGGGTTATGCGCTCGCCTGCAAGCTGTTCCCGCTCGACAATCTCGTCATCGAATTGCCCAACTAAAAACTTTTTGTCAGAAGACGGAAAATGCAGCCTAAGCAATCTCGACGACCGCAAGATCGGCATGTTCGATCAATAACGCGTGCCTATCCGGTTAACGCAGACACCCACTACATGTGGGGTGCGCCCAGCCACCGGAATATAGCTCGGCGTAGGTCGAGCTTTCACCTCGCCCGCAATTCCGAGGAGCGAGCGGAAGGCGTTGAACGGGTAGAACCGTCTGTTGAAGCGGAACGTGAACTCGTTGAGGTAGGCCTGCAAATGCTTTGCGCTCACGCCGTGGTGGATGCCGTTGAGCCAAGTCTTCAGGTTGCTGAAGACGAGGTGGATGATCGGCATGAACTCCTCGGCCACCTCGGGGTCGCCACATTCGGCGATTGCGTGGTGGTCATAGCCGCGTGTCCGCAGGTTGCCATAGCCGCTCCAATCGTCGGTGACGATCAGCGACCCCGGCATGACGGCACTCCCTACGAAGCCGCACAGCGAATCGGCGCTGCGGTCGGCGGCGATAGACAGCCGCACCCGACCGGCAAAGCGGCCATCCTTACGCTTGTCCTGGGCTGTACCCGGCTCCCGGTGGCGGACTTCGACGGCGGCAGCCACCAGCGTCTTATGATGAACACCCCGGCCTTCGCCGCGTGTCTTTCCACCGACATAGGTTTTCGTCCACCTCGACATGACCGTCCGTCTTGCCGCCGATCCGGTCCTGCTCGGGCCGTACCATGCCCGCGCGGAGCTTGTGCAGGATTCCGAAGGCGGTTTCGTACCGTGTCAGCCCAAGCTGCTGGAACTGAACCGCCGACATGCCTGCGTCTGGCTGGCGACCAGATAGGCTGCCCAGAACCATGTGCTCAAGGGCGTGTGGCTGCGTTCCATGACCGTGCCGACGGTCAATCCCGTCTGGCGGCGGCAAGCGCGACAGGTCAGGACGCCGGGCCGCGTAGCGATGCGGAAAGGCTCACCCACAATGCCACAATGCGGGCAGGCGAACCCGTCACCCCAGCGAATCCTTTCCAAGTAGGCGGCGCAGGCCCCATCGTCGGGAAAAAGCCGCTGAAACTCGGGGAGCGATTGCGGGAAGGGCAGATCGGTGCGGCCGAGAACGTCCATAACCCACTACCCGCTTCACGCACACAATCTATCCCTAGTGGGTGTATGCGTCAACCGGATAGGCACGCAATAACGATCGTCACGAAAAGCAGAGGAAAGTTAGGCCAATCCCGGTTCAATCAAACCGGGATTGAGTCTAAGTTCGGTTCGAAGGGTACGGAGGCTAATTTATGATCGCTTCGGTTGTGATTGTCGGCGCCAACCTTGCAGGGGGACGCGCCGCCGAACGCTCCGGCTCAACGGCTATGAAGGACGGATCGTCCTGATCGGGGAGGAGCGGTGGCTGCCCTATGAGCGGCCGCCGCTCTCCAAGGAATGCCTGTGGGACAGGGACCAGCTTCCGAGAATTTCTTCCTCCACGACCAGCAATGGTATGAAGACAATAAGATAGAGCTGGAGCTCGGCGTGCGCGCCGAAGCACTGGAGCTTTCCGGCCGAGGGGTTCGGCTGGCCTCCGGCAAGGAAATCCCGGCCGATCGCATCCTGCTCGCGACCGGCGGCAAGGCGCGCTTGTTGCCGCTCGATGGTGCGACCGCCGCCAATGTCCATCATTTGCGGACCAAGGATGACGCCGACCGGCTCGCGGCAGATCTCAAGCCGGGTGCGCGTATCGTCGTCATCGGCATGGGCGTGATCGGCGCTGAGGTCGCCGCGAGCGCGCGCAAGAGCGGCTGTGAGGTCACCGCGATCGAGCCGGCGCCGGTGCCGATGATCCGCACGCTCGGTGCGCATTTTGGCGCCTGGCTCGGCCGCGAGCACGACATGCGGGGGGTGAAGGCGCGTTACGGCATCGGCGTGACCAGGCTGTATCTTGACGGCGGCCTGGTCCGCACCGTCGAGCTCGACGATGGCACGCGCATCGATTGCGACGCGGTGGTGGTCGGGATCGGCATCGTGCCCTCGACCGAGCTTGCCGCCAACGCCGGGCTAGCGATCGGCAACGGCATCGTCGTCGATCGCCAGGGGCGCACCAGCCATGAGGCGGTGTTCGCGGCGGGCGACGTCCCGACCAGCCCAATTTCTTCGGCGGCCGGGTGCGGCTCGAAACCTATCAGAATGCCGCCGATCAGGGCATGGCCGCGGCGCAGGCGATGATCGGCCGCGAGGTCGATTATCTGAAGCCCTGCTGGTTCTGGTCCGACCAATATGACATCAACATCCAGGTCTCCGGCCGGATCGACGACAGCCTGCCGGTGGTGATGCGCGGCGAGCTCGACAGCAGCCAGTTCACCGCTTTCTTCCTCGACGGCAACGTCGTCGCCGGCGTGCTGACCGCAAATCGCGCGGTCGACATGGGCTCGGTGAAGAGGATGGTCGAGCGTCGGCTCGAGGTCGACCCGGGTCAGCTCGGCGACGCCTCGATTCCGCTCCGCGAGTTCTTGAAGCCCAAGGCGCGGGCGGCATAGGCCTGCGGTGAAACCCTCTCCCGTACGGAAGAGGGTGGGCGCCTTGGACGCCCCCTCAGATCAGCCTATCCCGCCGCCTGCCACGTACAGCGTCTGGCCGGTGATGTAGGACGCCTCGGGCGAGGCGAAAAAGCAGATCGCGGCGGCCAATTCTTCCGGCGCCGCGAAGCGCTTCATCAAGGTGTCGCGCAACGTCTGCTCGCCGACTTCGCCGAAACCCTTGATGTCGGCCTCGGTCGGCGGCTTGGGATTGCGCGGCGTGACCCGCTCGACATTGACTCCGCCGGGAGCGACGCAATTGACCCGGATCGGCCGGTCGTCGAGTCGAACGACAGCGCCTTGGGTAAGCGCCGACACTCCGCCTTTGGCCGCGGCATAGGGGACCCGGTTGATGCCGCGCGTTGCGACCGGGCCGATGTTTACGATCGCGCCGCTGTTCGCTTCGAGCATCAGCGGCAGCACGGCATGGCAGCACCACATTGTCGGCCATAAGGAGCGCTGGATCTCGGCCTCTCAGGCGCGCCGTATGAAGGATCTGGAGAAGGAGAATCTGCGGCTTGGCCGGGCGATATCCAACCTGACGCTGGATAAGCTCATTCTCCAGGAGGCCGCCCGGGGAAACTTCTGAGCCCCGCGCGGCGACGGCGCTGTATCGACCAGCTGTGGCGAGAGCTACCGGTGTCCGAACGACGGATTTGCCAGGTGCTCGGACAGCATCGGTCGACCCATCGCAAGGTAGCGCGCGGGGCGGACGACAAGCAGGCGCTGACCGAGGACATCATCGCTTTGGCCAAGCAATATGGTCGCTATGGCTATCGACGTGTCACCGCCCTGCTGCGCGACGCGGGGTGGACAGTGAACCGCAAGCGGGTCGAGCGGATCTGGCGACGCGAGGGGCTCAAGGTGCCGCAGCGGCAGCCGAAGCGCGGGCGGCTGTGGCTCAACGATGGCTCCTGTATCAGGCTCCGGCCGGAATATCCGGGCCATGTGTGGGCCTATGACTTCGTTCAGGGCCGCACTCACGACGGGCGCAAGTTCCGGATCCTGACCATTATCGATGGGGCCAGCCGGGAATGCCTGGCGCTCATCGTCGCCAGGAAGCTACGTCACGAGGACGTGCTGGCCGCGTTGGCCGAGTTGTTCGTCATGCGCGGGCCTCCGGCACACATCAGGTCCGCTAATGGTCCTGAATTTATCGCTACGGCCGTCC

119
AGAAGTGGCTCGGCAAGATCGGCGTGAAGACGCTCTACATCGCGCCGGGATCACCTTGGGAGAATGGCTACAATGAGAGCTTCAACGGGTCGCTGCGCGACGAGCTGCTCAACGGCGAGATCTTCTATCCTCCGAGCGCTCCCAGCTGATATAACCTTCATGCGAGCCGGGAATCAGCGCCAACCACTCATCCCGGGCCTTGCGACGACTCCTCGTCCGCAGGGCGGCGCCATCATATCCAGCTACAACGTGGGTTTTACCATAGGCATACAGGATCGCCTTGCGCTCAAGATGCTGGGGACCGATCTTCTCACAGATCATCGCTCTTCTCCTCCCGTGCCGCGCAGGGGGCGCCGGTGGCGTGGTCGAGCAGTAGACGCACGATCAGGGGCATCAACTTCAGCCCGCGAAGGCCGTTCTTCGCCCCTACGTCGGCGAACAGCTTGTCGAGTTCCCGCGCGATGTAGGGCACGTGCGCGAAACGCGCACCGCCCTTGGCGGTGTCGACGGTGCGGTCCTGGCCAGCCCATTCGTAGAGGTCTTGGAAGAGATGCTTATGCAGGGCACGGTAGCCGTCGGCCGTTGCCGGAAAGGTCAGGCGCGCGGCTTCCGCGCCGCGCGCCTGAGTAAGTCGCCGCTCGGCCTCTGTGAGCGTCTTATCATCGGTGATCCCAAGCCGGTTCCGAAGCGTGTCGGTGCCGGGATAGGTATAAGGCCCAGTCGGCCCCAAGCTGCGATTTTGGCCAAGTCGGAGAACGTTTTGGGAACGTGGCATTCGGTATTGACGATGGAAATCATAGAAGAGGACAGTTCGCTCCATCATAGGGAGTGGGTTGATGGATCGGAGTATTCCGGGCGGGGATTTGATCGGACGATGGAGCCTGAGCTTTGCCGATATTGATTTTGTAAATTCGAAGCCGGCCCTGACGCGCCTTGGCCTCGCCGCGCAGCTGAAATTCTTCGCTTCCCTGGGGTTTTTCGCGATCGATCCCGGCTCAATCCCTACCGATGGCCTCTCGTATCTGGCCGAGCAACTCGGTGTCGAGGCTGGCGAGATAGCCGGTTATGACTTTTCCAGTCGGACAGCACGACGGCATTGTGCGGAGATCCTGATCCATCTTGGATATCACCGCATGAAGCGGGTGGATCGCGCGCAATTGACGGAATGGATTGCTGGCGAGCTGTGCCCGGGCGGCCAGTCGATCAATGCCATGCTTGAGCATGTTTTCCTGTGGTGCCGGGACCGGCGTATTTATGGGCCGTCGCGCAAGGAGCTTGAACGTGTCGTTCGCTCACAACGGCAAGATTATCTGGACACCTGGCTGATCGGAGCCAGTGATCGGCTTTCGTCAGATGCGGTGGCGTTATTGGAAGCCTCGCTTGCCGATCCGGACAGCTCGACCGGATTCAACAGGATGAAGGGTGACGCCGGACAGGCAACGCTCGACAACATTCTCGACGTGACCGAGAAACTCGCCTTTATCCAGAGACTTGATCTTCCCCATGATCTCCTGACGGCTACGGGCAAGCCATGGGTCGATCAGATTGTTCGCCGCGTTGCCGGTGAAAAGGCCTCGGAGATGCGCCGGCATGCGCCGGCGCGACAGCTCGGCCTTTATGCGATTTATCTAATGTCGCGGGAGGCGCAACTCACTGACGCGATGATCGACCTGCTGATCGAAACCGTTCACAAGATCGGAACGCGCTCGAAACGCAAGGTGGTGGGCGATATCGCGAAAGACATCGAGCGGGTCTATGGAAAGGAGCGCCTGCTGGTCGAGATCGCCAGCGCCTCGATCAATGAACCATCGGGGCGCATCTGCGATGTCATTTTCCCGATCGCCGGTAAGGCCAAGCTGGCGGCGATCGTCAAGGAGAGCCATGCGAAGGGCGCTCTGGACCGCGCATCTACAAGGTGATGTGTCAACGGCGGAGCAAAAGTCGGCCATTCGGCGGCGTAAAACCAGGCCATCGT Contig 3 ATAGCCAGCGCGCCGCCGAAGCTGTTGCCGACCAGCGACGCCTTCTCGATGCCCATCGCATCGAGGAAAGCCGGCGAGATGGTCGACCCAGATCTGCCGGTCCTTGATGCCGCCTGCCGGGGCGTCGGTGTAGCCGAAGCCGATCATGTCGGGTGCGTAGACGTGGAAGTCCTTGGCCAACTCGGGCAGGTTGGCGCGCCAGTTCGCCCACGCCGTCACACCGGGACCCGAGCCGTGGATCAGGAGCAGCGGCGGCCCCTCGCCCTGCTCATGATAATTGGTACGAACGCCGTTCACCGAGACGGTCTTGCCGATCTCGGGGATCGTAGCTTGTTTTTCCATTACTGCGTCCGACATGGAGATGACAGGAATTGACCTACAGGGCGCGCCACCAGGAGGTCGTCTCACGGAATTCGCCGGAAAGGACCGGCTGTGCGCTCTTGTTTTAGTTGCACATAGAGGAACTCCTTACAAGGCGAATCTCGCCGTTCAGGCCCTCGAACACGTCGATAATCAGCGCTTGCGTGCTTCGCAAGCGCCGAGCCTGAAGCCGCGTTTCGACATTTTTCCCGATCTGGCTGATCTACGCTGTCGTTCAACCACTCCAAAAGCGGTTCGGGCCGGGGCGTTATCAGGCGAATCCCGTGCAAACCGTTGTCCTGATGCGGTTGCGGAGGTGGTCGAGTTGGACTAAGGCGCGGGGGGCGGGACCGGGCTGGGGTGCCGGGCTTGAAGTAGAGACAAGAGAGAGGATGAGCGCGATGTCGCAAGTGACCGAGCTGGGTTATATCGGTCTGTCCATATCGACGTTGAGGCGTGGAAGGATTATGCGGCGTCGATCGTCGGCATGGAGATCGTCGACGAGGGTGAGGGCGACCGCATTTATCTGCGGATGGACAAGTGGCATCACCGCCTCACGCTGCACATCGATGGCGGCGACGACCTGGCCTATATCGGCTGGCGGTTGCGGGTCCGTCCGAGTTCAACGAGCTGGTCGAGAAGCTGCGGCTCAACAACCTCGACATTCGCGTGGCGTCGTAGGAGGAATGCGCCGAGCGGCGTGTTCTGGGCCTGGCCAAGACGGTCGATCCGGGCGGCAACCCGACCGAGATCTTCTATGGTCCGCTGGTCGACAACTGGCGTGCGTTCCACCCGGGGCGGCCGATGTTCGGCAAGTTCATGACCGCCAAGGAAGGCATCGGCCACTGAATCTTGCGCCAGGACGACATTCCTGCCGCGGTGCGCTTCTACGAGATGCTCGGCCTCACCGGCTCGGTCGAGTACAAGCTTCCGCTTCCCGGCGGCATGGTGGCGCAGCCTTATTTCATGCACGTCAACGGGCGCCAACATTCGGTGGCGTTCGGCGCTTGGGCCGATGAAGAAGCGGATCAAGATGCTCGAATATACCGATCTCGACGATCTGGGGGTGGCGCACGACATCATCCGCCAGCGCAAGATCGACGTTGCGCTGCAGCTGGGCAAGCATTCGACCGACGAGGCTTTGACCTTCTACCTGTCCTATACACTCGGCCTTGGAGCTGGCTGGGTGCCAAGAAGGCGGACAACCAGCAGGAATATTACAGCCGCGACATTTTCGGCCACGGCAACGAGGCCGCCGGCTACGGGATG

120
GACATCGAGCTCTAACCGAGCGTTTGAAGTCTTGGGCGATGGACGCCGGGGTCTTGATGATCCCGGCGTCCCAGCGACGATTGCGAGAAGGATGATATGAGCGAGCTGATTCGCCTGTGCCGAGTGGATGAGGTGAAGGAGGGCGAGCCGGTGGCGGCGCATGTCGCCGGGCTGCCGCCGTTTGCGGTCTATGATGTCGGGGGCACTTATTATGTGACCGACAACATCTGCACGCACGGCAATGCGATGCTGACCGACGGCTATCAGGACGGCGGCACGATCGAATGCCCGTTCCATGGCGGCGCGTTCGACATTGCGAGCGGGGCGGCGACGGTGTTCCCGTGCCAGATCCCGCTCAAGACCTATAGTGTCGAGGTCGACGACGGCTGGATCGCGATCCGGCTGGCATCGCCCGAAGCGGGCGCCTGAAATGGGAGCTGAGATGGGGGCACGGACGGGGTTCGAGGAAGCTGGGGCGCGCGATCTTCACCAGGAGGTTGAGGACTGGTGGCGCGCCATGCCGAGCTGATCACGAGGACCGTCTGGAGGAATGGCCCGAGCTGTTCACCCAGGAGTGCGTCTACAAGATCATCGCGCGCGAGAATGTCGACCGCGGCCTGCCGATCGCGGCGATCTTCTGCGACAGCCGCAAGATGCTGGTGTGAACCGACAATTTAAGCTGCAATCTGCTTTGAACGCGGAATGCTGCCAACCGACAGATTATGCTGCAGCGGACGAGACTTCACACGCACCGTGATTGCCAGCCCGGGCGCAATTCTCATGATGGTTTTGAAGTCCCGGCCACCAGCTGCTCCAGCGCCTGCACCAGAGGCATTGCCGTGATGCCGTGCGGCTTCGGGTAGGCCATTTCTCCGGGGTAAATCAGCCAGCGCTTCTCGATCTCGAGATCGTCACAGGCAAGGTGAAACCCGCGCTCAACGCTGGGCGCCGTCGACCGCTTCACTTCGATGGCCACTTCGGGAACGCCGCCGGACACGAGGATGAGGTCTATCTCCGCACCATTCGCCGTCCGGTAGAAATGGGCATCGTAGCGATCGCCGACCGCGCCGATGATGTTCTCGATAACCATTCCTTCCCAGCTGGTGCCGGCGACAGGATGGGAAAGAATGTCGTCGAGCTTTTCGAGGCGCAGCAGGGCATGCAACAGCCCGCTGTCGCGAACATAGAGCCTCGGTGCCTTCACCAGTCGTTTGCCGACATTGGCAAACCAGGGATTGAGCCTGCGGACGAGGCCGAGATCACACAGGATGTCGATATAGTTGGCAACGGTCGGTGCCGAGATGGAGAGACTCCCCGCAAGGCCAGAGGCATTGAACAGACCCCCCTGCCCGTGGGCGAGCATCGTCCCAGAGCCGGTTGATGGTCGCACTCGGCATGCGAGGAACCATGAAAGGCAGATCGCGTTCAACATAGGTGCGGATCAGGTCGAGCCTGCGGCGGAAGCTTTCGGTGTCGTTGGGCGCGGTCAGGCTCTCGGGAAAGCCGCCGCGCACCCACAACTGGTTGAGATCGTCCAGTCCGGCTGACCGCGCTTCGTCTGCGTCGATCGGACCCATGTCCATATAGGCGACGCGTCCGGCGAGCGACTCCGAACTCTGATTGAGCAGAATTCGGTTTGCCGAGCCGAGCAGCAGGAAATGACCCGCCCCCTCCCCTGCACGGCGGCGCTCGTCGATAACGCCGCGCAATGTCGAAAACAGATCCGGTGCCCGCTGGACCTCGTCAATGACGACAAGCTTGCCAACGCCTATCGTACGTAAATGTCACTTTCGTGCAGTGACCCCCGCTTCTCGCAAGGCTCGAGCGGCACCGTCGGCAACCGGCATATTTTCAGCATTGTCTCGCGGCCTATAAGCCCGAATATACCTGTACGCCGGTTCCCGGTCCGATCACGCTGCCTCCACGCAGCGGATGAGCCGGTCAGGGTGATGCGCCGGCACGGCTCGGCCGTCGTTCCGCTTGCGCCAACACGATCTCAACTTCACGCAGCTTGCCGATAATCTCTTCCGGCTTGTGCTTCTTGCTCGGCATTCGTCGTCCCTTTCGTGATCCAGACGGCTCTGTTGCAAAAATCGTGAAGCTTGAGCATGCTTGGCGGAGATTGGACGGACGGAACGATGACGGATTTCAAGTGGCGCCATTTCCAGGGTGATGTGATCCTGTGGGCGGTGCGCTGGTATTGTCGCTATCCGATCAGCTATCGCGACCTTGAGGAAATGCTGGCGGAACGCGGCATTTCGGTCGACCATACGACGATCTATCGCTGGGTCCAGTGCTACGCCCCGGAGATGGAGAAGCGGCTGCGCTGGTTCTGGCGGCGTGGCTTTGATCCGAGCTGGCGCCTGGATGAAACCTACGTCAAGGTGCGGGGCAAGTGGACCTACCTGTACCGGGCAGTCGACAAGCGGGGCGACACGATCGATTTCTACCTGTCGCCGACCCGCAGCGCCAAGGCAGCGAAGCGGTTCCTGGGCAAGGCCCTGCGAGGCCTGAAGCACTGGGAAAAGCCTGCCACGCTCAATACCGACAAAGCGCCGAGCTATGGTGCAGCGATCACCGAATTGAAGCGCGAAGGAAAGCTGGACCGGGAGACGGCCCACCGGCAGGTGAAGTATCTCAATAACGTGATCGAGGCCGATCACGGAAAGCTCAAGATACTGATCAAGCCGGTGCGCGGTTTCAAATCGATCCCCACGGCCTATGCCACGATCAAGGGATTCGAAGTCATGCGAGCCCTGCGCAAAGGACAGGCTCGCCCCTGGTGCCTGCAGCCCGGCATCAGGGGCGAGGTGCGCCTTGTGGAGAGAGCTTTTGGCATTGGGCCCTCGGCGCTGACGGAGGCCATGGGCATGCTCAACCACCATTTCGCAGCAGCCGCCTGATCGGCGCAGAGCGACAGCCTACCTCTGACTGCCGCCAATCTTTGCAACAGAGCCGGCATCGCCTCCCCTGTTCTGTATCAACTGCGCCAGTGTGATGCCCGGGTTGGCGCAAGAGCGCGAGTTGCAGCGCGACGCCCAACTGGTTCCGTCGTTCCCGTCGCGCGCCGATGATCTCGATGTCCGCAGGCTCGAAGGTGTAGAGCCGAGCCAAGTGGTCGCGATCGGTCGGGATGGCAAGGATCTGATTGCGCTCGCTCTCGGTCAGGAGTTGATGCTTGCGCTTCGTCATGCCGATTCCCGTCCACAAAAGGTCAACTGCGCCTATGGACAGCCATGAGTAAAGAGACATAGTATGTGGACGGCTATGGGCGGGGTGAAGCGGTCGCCCTTCAGAGCGTCCACAGAACGACCGTTTGGGAGACACGATAGATGGGCGGCATTCTGGGCTACGCCCGCGTCAGCACCGGCGATCAGGATGTGGCGGGCCAGACCATGCGACTGGAGAATGCCGGCGCCATCAAGGTGTTCACCGATGTAATATCGGGAAAAAGCATGGAGCGGCCTGGACTGGCCGAACTGATCGCCTATGCCCGCAAGGGTGACACGCTCGCCGTGGTCCGCCTCGATCGGCTTGGGCGTTCGCTGACTGAACTGCTCGCCACAGTCGAAACTTTGCGCAGCCAAGGCATCGCCCTCCTGAGTCTTGAGGAAAAAATCGACACCTCGTCGGCCGCCGGCGAACTCATCTTCCATGTGTTCGGAGCTATCGCCATTTCGAGCGGCGGCTATCTCTGAGCGGACCCGCGATGGGATCGCCGCTGCACCGCCAAGGAACAACCTGGCCGTCAGCCGCTCGAATGTCCAAGGTGATGCGGCCATCAAA

121
Contig 4 AGCAGAGCGACAGCCTACATCTGACTGCCGCCAATCTTTAAACAGAGCCCTCCGGCGGCTATGATTATTATCCGGACAACCCCCGCCGCGTGTGGAGCGCGGACGAGGCCGGCAAGGCGATCTTCTACTATGAGCGGCAGCTCAACGATCGCTTCATGGCCGTCAATACATGAGTGAACCGATCGTGCGTATCAGGCTGGTCAAGACGATCAGTCCTGAGGCCGCTCATGCCGCCGTCGGCGCCGCCCTCCGGGCCGGCGCCGACGAAGGTTGTGCGGTGGTGGCGGCCGTGGTCGACGCAGCAGGCGAGCTTGTCGCCTTCGTGCGGGCCACGGGCGCGCCGTTTCATTCGAGCGACATCGCTCGGGACAAGGCGTATACGGCCGCGAGCTTCAAGATGCCGACCGCGGACATCCCGGGCATCGTTTCGGGCAGCGACGCACTTCGCGACGGCATCACGCGGCGCAGGAACATGGTGATGTTCGGCGGAGGACTCCCGATCGTCGTCGACGGCGAGATCGTCGGCGGCATCGGCGTATCGGGCGGCAGCGAGGCGATGGACACGGCTTATGCCAATGCCGGCCTTGCCGCGATCGGCGCCAAACGGTTCGACTGAAGTTTCGAATTAACGGAGAAAGAAGATGGCGCGAGGCGACGAGCCCTTGCCGACGGTGACGGCGGAAGTTTTGAATTATATCGACGGCGCCTATCGCAAGGGCAGCGAAGGCAAGAGCTTCGTCAACATCGACCCGGCGACCGGCAAGGAGATCGGCCGCGTCTATGAGGCGAGCAGGGAGGACGTCGACGCTGCCGTTCAGGCCGCCAAGCGCGCCTTGAAGGGCCCGTGGGGCAAGATGACGATTCCCGAGCGAGTCAAGCTGATCATCAAGGTCGCCGACGAGATCGAGCGCCGCTTCGACGATTTCCTCGCGGCCGAGGTCGCCGACACCGGCAAGCCGATCTCGATCGCCTCGCACATCGACATTCCGCGCGGCGCCGCCAATTTCCGCATGTTCGCGGACGTCGTCTCGACGACCCCCACCGAATCGTTCATGACCCCGACTCCCGACGGGTCGAAGGCGCTGAACTATGCGGTGCGCAAGCCCAAGGGCGTGGTTGCGGTGGTCTGCCCGTGGAACTTCCCGTTGCTGCTGATGACCTGGAAGGTCGGCCCCGCGCTCGCGTGCGGCAACACCGTCGTCGTCAAGCCGTCCGAGGAAACGCCGCGCACCGCGGCGCTGCTCGGCGAGGTGATGGACGCGGTCGGCATGCCAAAGGGCGTGTTCAACGTCGTCCACGGCTTCGGCCCGGATTCGGCCGGTGAGTTCCTGACCAGCCACAAGGACGTCGATGCGATCACCTTCACCGGCGAGACCGGCACCGGCACGGCGATCATGAAGCAGGCCGCGGTTGGCCTGCGCGACATTTCGTTCGAGCTCGGCGGCAAGAACCCGGCGATCGTGTTCGCCGATTGCGACATCGACAAGGCGATCGAGGGTGTTGCTCGCGCCACCTTCCTCAACACCGGCCAGGTGTGCCTCGGGACCGAGCGCATCTTCGTCGAGCGGTCGATCTTCGACAAGTTCGTCGAGCGGCTTGCCGCGGCGGCGCAGGCGTTCGAGCCCGGCGATCCGAAGCATCCCCAATATATGGGCCCGCTGATCAGCGACGAGCATCGCTCGAAGGTGCTCGGCTATTACCAGCGCGCGGTCGACGAAGGCGCGACCGTCGTGACCGGCGGCGGCGTTCCGAAGAACGTTGACGGCGGTTTCTACGTCGAGCCGACGATCTGGACCGGCCTTGCTCCCGATTCCACGGTTATGCGCGAGGAAGTATTCGGCCCGTGCTGCGGCGTCATTCCATTCGACACCGAGGAAGAAGCCATTGCGATGGCCAACGACACGCCCTACGGGCTCAGCGCGACGATCTGGACGCAGGATCTTTCCCGCGCCCATCGCGTGTCGGCCGAGATGGAGGTCGGCATCTGCTGGGTGAACTGCTGGTTCCTGCGTGACCTGCGCACCGCCTTCGGCGGCGCCGGGCAATCCGGCATCGGCCGTGAAGGCGGGGTGCACAGCCTCGAATTCTACACCGAGATCGAAAACATCTGCGTGAAGTTGTGAGCGCCATGGCTGATCAGGAAGTCACATTCGATAGAAGCAACATCGACAAGGCCGCGGCCCAGTTGAGGCACGCCGCCGAGACGATGAATCCTGGCCCGCCGATCCGGGATCTGATCTCGGCCGGCGGCGTCGAGGCCGCTTATGCGGTGCAGGACGCCAATACGACGCACTATCTTGCCAACGGCCGCCGGCTGGTGGGCCGCAAGACCGGGCTCACGTCGAAGTCGGTGCAGAAGCAGCTCGGCGTCAACCAGCCCGACTACGGCATGCTCTATGCGGACATGGACGTGCCCGAAGGCCAGCCGATCCTGCTGACGCGCGTGGTGCAGCCCAAGGTCGAGGCGGAGATCGCTTTGGTGATCGGGCGCGACCTCGACGATCCCGACGTGACCACCGCCGAAATGCTGCGCGCGATCGAATATGTCGTGCCGGCGATCGAGATCGTCGACAGCCGCATCGCCAATTGGGATATCAAGATCTGGGACACGATCGCCGACAATGCGTCGAGCGGGGTGTACACTCTGGGGGNCGTTCCCAAGAAGCTTGACGGGCTCGACCTGCGCACCTGCGGATGGTGATGGA

122
A.8. Transcription of ipf genes
RNA was isolated from strain Ibu2 cultures using a RNeasy Mini Kit (Qiagen). DNA was
digested with DNase (Promega) and RNases were inhibited with RNasin (Promega). RNA was
subjected to reverse transcription PCR (RT-PCR) using the superscript one-step RT-PCR
(Invitrogen) and intergenic regions were targeted to identify the boundaries of the transcripts.
RNA and primers were held at 94oC for two minutes prior to synthesis of cDNA in order to
denature secondary structures. RNA from both induced and non-induced cultures were compared
to assess induction by IPF and basal transcription, as indicated in Figure A-8.2. Primers are
shown in Table A-8.1.
Three transcriptional units were identified on contig 1 due to the absence of RT-PCR
amplicons spanning intergenic regions (Table A-8.2, Figure A-2.2) and pictured in Figure A-8.1.
Due to the presence of a light band in gel 4 for product 8, additional work would be required to
determine if transcripts 2a and 2b are distinct, as previously thought, or one combined transcript
(Table A-8.2, Figure A-8.2). However, the absence of products 9 and 12 provide support that
transcripts 2 and 3 are distinct. The end of transcript 3 has not been identified. A minimum of
four transcripts (transcripts 2-5, Figure A-8.1) are thought to be required for expression of the
upper ipf pathway. Transcripts 1, 2, 3, and 5 were evaluated for transcription in cultures induced
with 50 mg/L IPF. Each of these transcripts was transcribed in both the presence and absence of
IPF (Table A-8.2, Figure A-8.2). While many pathways for aromatic hydrocarbon catabolism are
organized in to operons, it is common in Sphingomonads for these pathway genes to be
distributed among several transcriptional units and spread out in distinct genetic environments
(Pinyakong 2003, Romine 1999, Suar 2004).

123
Figure A-8.1. Transcription of ipf pathway genes. Genes are described in Appendix A.2. Additional work is required to determine if transcripts 2a and 2b are separate. Transcripts 1,2,3 and 5 are transcribed in both the presence and absence of IPF induction. Table A-8.1. Primers used for RNA work. When a base is indicated as a “M” it consists of either A or C bases, and “Y” consists of either C or T bases.

124
Table A-8.2. Amplicons used to determine transcript boundaries and transcription in the presence and absence of IPF. Primer sequences are provided in Table A-8.1 and bands from RT-PCR can are shown in Figure A-8.2.

125
Figure A-8.2. Gel photos from RT-PCR for the determination of transcript boundaries for strain Ibu2 ipf and related genes, and expression from induced and non-induced cultures. Amplicons are described in Table A-8.2 and primer sequences are provided in Table A-8.1

126
FigureA‐8.2continued.

127
Figure A-8.3. Digest to confirm the identity of band 2, product 7, gel 1 from Figure A-8.2. The band was excised and digested with Cla I, Not I, and Sma I and the product is shown in the gel. The predicted restriction enzyme digest performed by NEBcutter for the correct amplicon matched that shown in the gel: Cla I, 500 and 1100 bp; Not I, 1100 and 500 bp; Sma I 300 and 1200 bp.

128
LITERATURE CITED
Allen, E., Doisy, E.A., 1923. An Ovarian Hormone. Journal of the American Medical Association 81, 819-821. Amweg, E.L., Weston, D.P., Johnson, C.S., You, J., Lydy, M.J., 2006. Effect of Piperonyl Butoxide on Permethrin Toxicity in the Amphipod Hyallela Azteca. Environmental Toxicology and Chemistry 25, 1817-1817. Andresen, J.A., Grundmann, A., Bester, K., 2004. Organophosphorus flame retardants and plasticisers in surface waters. The Science of the Total Environment 332, 155-166. Andresen, J.A., Muir, D., Ueno, D., Darling, C., Theobald, N., Bester, K., 2007. Emerging pollutants in the North Sea in comparison to Lake Ontario, Canada, data. Environmental Toxicology and Chemistry / SETAC 26, 1081-1089. Ankley, G.T., Collyard, S.A., 1995. Influence of piperonyl butoxide on the toxicity of organophosphate insecticides to three species of freshwater benthic invertebrates. Comparative Biochemistry and Physiology Part C: Pharmacology, Toxicology and Endocrinology 110, 149-155. Arditsoglou, A., Voutsa, D., 2009. Partitioning of endocrine disrupting compounds in inland waters and wastewaters discharged into the coastal area of Thessaloniki, Northern Greece. Environmental Science and Pollution Research 17, 529-538. Arfsten, D.P., Schaeffer, D.J., Mulveny, D.C., 1996. The Effects of Near Ultraviolet Radiation on the Toxic Effects of Polycyclic Aromatic Hydrocarbons in Animals and Plants: A Review. Ecotoxicology and Environmental Safety 33, 1-24. Artola-Garicano, E., Sinnige, T.L., van Holsteijn, I., Vaes, W.H.J., Hermens, J.L.M., 2003. Bioconcentration and acute toxicity of polycyclic musks in two benthic organisms (Chironomus riparius and Lumbriculus variegatus). Environmental Toxicology and Chemistry 22, 1086-1092. Baldigo, B.P., Sloan, R.J., Smith, S.B., Denslow, N.D., Blazer, V.S., Gross, T.S., 2006. Polychlorinated biphenyls, mercury, and potential endocrine disruption in fish from the Hudson River, New York, USA. Aquatic Sciences 68, 206-228. Balk, F., Ford, R.A., 1999a. Environmental risk assessment for the polycyclic musks AHTN and HHCB in the EU: I. Fate and exposure assessment. Toxicology Letters 111, 57-79. Balk, F., Ford, R.A., 1999b. Environmental risk assessment for the polycyclic musks, AHTN and HHCB: II. Effect assessment and risk characterisation. Toxicology Letters 111, 81-94. Barnes, K.K., Kolpin, D.W., Furlong, E.T., Zaugg, S.D., Meyer, M.T., Barber, L.B., 2008. A national reconnaissance of pharmaceuticals and other organic wastewater contaminants in the United States -- I) Groundwater. Science of The Total Environment 402, 192-200.

129
Bartels, I., Knackmuss, H.J., Reineke, W., 1984. Suicide Inactivation of Catechol 2,3-Dioxygenase from Pseudomonas putida mt-2 by 3-Halocatechols. Appl Environ Microbiol 47, 500-505. Bayly, R.C., Dagley, S., 1969. Oxoenoic acids as metabolites in the bacterial degradation of catechols. Biochem J 111, 303-307. Bennie, D.T., Sullivan, C.A., Lee, H.B., Peart, T.E., Maguire, R.J., 1997. Occurrence of alkylphenols and alkylphenol mono- and diethoxylates in natural waters of the Laurentian Great Lakes basin and the upper St. Lawrence River. Science of The Total Environment 193, 263-275. Bird, J.A., Cain, R.B., 1974. Microbial degradation of alkylbenzenesulphonates. Metabolism of homologues of short alkyl-chain length by an Alcaligenes sp. Biochem J 140, 121-134. Bjerregaard, L.B., Korsgaard, B., Bjerregaard, P., 2006. Intersex in wild roach (Rutilus rutilus) from Danish sewage effluent-receiving streams. Ecotoxicology and Environmental Safety 64, 321-328. Brennan, R.J., Schiestl, R.H., 1998. Free radicals generated in yeast by the Salmonella test-negative carcinogens benzene, urethane, thiourea and auramine O. Mutat Res 403, 65-73. Buerge, I.J., Buser, H.-R., Muller, M.D., Poiger, T., 2003. Behavior of the Polycyclic Musks HHCB and AHTN in Lakes, Two Potential Anthropogenic Markers for Domestic Wastewater in Surface Waters. Environmental Science & Technology 37, 5636-5644. Buerge, I.J., Poiger, T., Muller, M.D., Buser, H.R., 2006. Combined sewer overflows to surface waters detected by the anthropogenic marker caffeine. Environ Sci Technol 40, 4096-4102. Buszka, P.M., Yeskis, D.J., Kolpin, D.W., Furlong, E.T., Zaugg, S.D., Meyer, M.T., 2009. Waste-Indicator and Pharmaceutical Compounds in Landfill-Leachate-Affected Ground Water near Elkhart, Indiana, 2000–2002. Bulletin of Environmental Contamination and Toxicology 82, 653-659. Butcher, J.B., Garvey, E.A., 2004. PCB Loading from Sediment in the Hudson River:  Congener Signature Analysis of Pathways. Environmental Science & Technology 38, 3232-3238. Camacho-Munoz, M.D., Santos, J.L., Aparicio, I., Alonso, E., 2010. Presence of pharmaceutically active compounds in Doñana Park (Spain) main watersheds. Journal of Hazardous Materials 177, 1159-1162. Camara, B., Herrera, C., Gonzalez, M., Couve, E., Hofer, B., Seeger, M., 2004. From PCBs to highly toxic metabolites by the biphenyl pathway. Environ Microbiol 6, 842-850.

130
Carmona, M., Zamarro, M.T., Blazquez, B., Durante-Rodriguez, G., Juarez, J.F., Valderrama, J.A., Barragan, M.J., Garcia, J.L., Diaz, E., 2009. Anaerobic catabolism of aromatic compounds: a genetic and genomic view. Microbiol Mol Biol Rev 73, 71-133. Casals-Casas, C., Desvergne, B., 2011. Endocrine disruptors: from endocrine to metabolic disruption. Annual Review of Physiology 73, 135-162. Castiglioni, S., Bagnati, R., Fanelli, R., Pomati, F., Calamari, D., Zuccato, E., 2006. Removal of pharmaceuticals in sewage treatment plants in Italy. Environ Sci Technol 40, 357-363. Cerdan, P., Wasserfallen, A., Rekik, M., Timmis, K.N., Harayama, S., 1994. Substrate specificity of catechol 2,3-dioxygenase encoded by TOL plasmid pWW0 of Pseudomonas putida and its relationship to cell growth. J Bacteriol 176, 6074-6081. Chavez, F.P., Lunsdorf, H., Jerez, C.A., 2004. Growth of polychlorinated-biphenyl-degrading bacteria in the presence of biphenyl and chlorobiphenyls generates oxidative stress and massive accumulation of inorganic polyphosphate. Appl Environ Microbiol 70, 3064-3072. Clara, M., Kreuzinger, N., Strenn, B., Gans, O., Kroiss, H., 2005. The solids retention time-a suitable design parameter to evaluate the capacity of wastewater treatment plants to remove micropollutants. Water Res 39, 97-106. Cleuvers, M., 2004. Mixture toxicity of the anti-inflammatory drugs diclofenac, ibuprofen, naproxen, and acetylsalicylic acid. Ecotoxicol Environ Saf 59, 309-315. Collins, J.F., Brown, J.P., Alexeeff, G.V., Salmon, A.G., 1998. Potency Equivalency Factors for Some Polycyclic Aromatic Hydrocarbons and Polycyclic Aromatic Hydrocarbon Derivatives. Regulatory Toxicology and Pharmacology 28, 45-54. Coogan, M.A., La Point, T.W., 2008. Snail Bioaccumulation of Triclocarban, Triclosan, and Methyltriclosan in a North Texas, USA, Stream Affected by Wastewater Treatment Plant Runoff. Environmental Toxicology and Chemistry 27, 1788-1788. Cook, M.E., Moore, P.A., 2007. The Effects of the Herbicide Metolachlor on Agonistic Behavior in the Crayfish, Orconectes rusticus. Archives of Environmental Contamination and Toxicology 55, 94-102. Cornel, P., Krause, S., 2006. Membrane bioreactors in industrial wastewater treatment--European experiences, examples and trends. Water Sci Technol 53, 37-44. Cuthbert, R., Parry-Jones, J., Green, R.E., Pain, D.J., 2007. NSAIDs and scavenging birds: potential impacts beyond Asia's critically endangered vultures. Biol Lett 3, 90-93. Cuthbert, R., Taggart, M.A., Prakash, V., Saini, M., Swarup, D., Upreti, S., Mateo, R., Chakraborty, S.S., Deori, P., Green, R.E., 2011. Effectiveness of action in India to reduce exposure of gyps vultures to the toxic veterinary drug diclofenac. PLoS One 6, e19069.

131
Daughton, C.G., 2002. Environmental stewardship and drugs as pollutants. Lancet 360, 1035-1036. Daughton, C.G., Ruhoy, I.S., 2009. Environmental footprint of pharmaceuticals: the significance of factors beyond direct excretion to sewers. Environ Toxicol Chem 28, 2495-2521. Daughton, C.G., Ternes, T.A., 1999. Pharmaceuticals and personal care products in the environment: agents of subtle change? Environmental Health Perspectives 107, 907-938. DeLorenzo, M.E., Keller, J.M., Arthur, C.D., Finnegan, M.C., Harper, H.E., Winder, V.L., Zdankiewicz, D.L., 2008. Toxicity of the antimicrobial compound triclosan and formation of the metabolite methyl-triclosan in estuarine systems. Environmental Toxicology 23, 224-232. Diaz, E., 2004. Bacterial degradation of aromatic pollutants: a paradigm of metabolic versatility. Int Microbiol 7, 173-180. Dougherty, J.A., Swarzenski, P.W., Dinicola, R.S., Reinhard, M., 2010. Occurrence of herbicides and pharmaceutical and personal care products in surface water and groundwater around Liberty Bay, Puget Sound, Washington. Journal of Environmental Quality 39, 1173-1180. Dsikowitzky, L., Schwarzbauer, J., Kronimus, A., Littke, R., 2004a. The anthropogenic contribution to the organic load of the Lippe River (Germany). Part I: qualitative characterisation of low-molecular weight organic compounds. Chemosphere 57, 1275-1288. Dsikowitzky, L., Schwarzbauer, J., Littke, R., 2004b. The anthropogenic contribution to the organic load of the Lippe River (Germany). Part II: quantification of specific organic contaminants. Chemosphere 57, 1289-1300. Eaton, R.W., 1996. p-Cumate catabolic pathway in Pseudomonas putida Fl: cloning and characterization of DNA carrying the cmt operon. J Bacteriol 178, 1351-1362. Erickson, D.A., Goodrich, M.S., Lech, J.J., 1988. The effect of piperonyl butoxide on hepatic cytochrome P-450-dependent monooxygenase activities in rainbow trout (Salmo gairdneri). Toxicology and Applied Pharmacology 94, 1-10. Escher, B.I., Hunzinker, R., Schwarzenbach, R.P., Westall, J.C., 1999. Kinetic Model To Describe the Intrinsic Uncoupling Activity of Substituted Phenols in Energy Transducing Membranes. Environ Sci Technol 33, 560-570. Eskenazi, B., Fenster, L., Castorina, R., Marks, A.R., Sjodin, A., Rosas, L.G., Holland, N., Guerra, A.G., Lopez-Carrillo, L., Bradman, A., 2011. A Comparison of PBDE Serum Concentrations in Mexican and Mexican-American Children Living in California. Environ Health Perspect Apr 15. [Epub ahead of print]

132
http://ehp03.niehs.nih.gov/article/fetchArticle.action?articleURI=info%3Adoi%2F10.1289%2Fehp.1002874. Fenner-Crisp, P.A., Maciorowski, A.F., Timm, G.E., 2000. The Endocrine Disruptor Screening Program Developed by the U.S. Environmental Protection Agency. Ecotoxicology 9, 85-91. Ferrara, F., Ademollo, N., Delise, M., Fabietti, F., Funari, E., 2008. Alkylphenols and their ethoxylates in seafood from the Tyrrhenian Sea. Chemosphere 72, 1279-1285. Fick, J., Soderstrom, H., Lindberg, R.H., Phan, C., Tysklind, M., Larsson, D.G.J., 2009. Contamination of surface, ground, and drinking water from pharmaceutical production. Environmental Toxicology and Chemistry 28, 2522-2522. Fisher, P.M.J., Scott, R., 2008. Evaluating and controlling pharmaceutical emissions from dairy farms: a critical first step in developing a preventative management approach. Journal of Cleaner Production 16, 1437-1446. Flippin, J.L., Huggett, D., Foran, C.M., 2007. Changes in the timing of reproduction following chronic exposure to ibuprofen in Japanese medaka, Oryzias latipes. Aquat Toxicol 81, 73-78. Focazio, M.J., Kolpin, D.W., Barnes, K.K., Furlong, E.T., Meyer, M.T., Zaugg, S.D., Barber, L.B., Thurman, M.E., 2008. A national reconnaissance for pharmaceuticals and other organic wastewater contaminants in the United States--II) untreated drinking water sources. Sci Total Environ 402, 201-216. Follmann, W., Wober, J., 2006. Investigation of cytotoxic, genotoxic, mutagenic, and estrogenic effects of the flame retardants tris-(2-chloroethyl)-phosphate (TCEP) and tris-(2-chloropropyl)-phosphate (TCPP) in vitro. Toxicology Letters 161, 124-134. Fraker, S.L., Smith, G.R., 2004. Direct and interactive effects of ecologically relevant concentrations of organic wastewater contaminants onRana pipiens tadpoles. Environmental Toxicology 19, 250-256. Fries, E., Puttmann, W., 2003. Monitoring of the three organophosphate esters TBP, TCEP and TBEP in river water and ground water (Oder, Germany). Journal of Environmental Monitoring 5, 346-352. Fromme, H., Otto, T., Pilz, K., 2001. Polycyclic musk fragrances in different environmental compartments in Berlin (Germany). Water Research 35, 121-128. Fu, K.-Y., Chen, C.-Y., Chang, W.-m., 2007. Application of a yeast estrogen screen in non-biomarker species Varicorhinus barbatulus fish with two estrogen receptor subtypes to assess xenoestrogens. Toxicology in Vitro 21, 604-612. Fuchs, G., 2008. Anaerobic metabolism of aromatic compounds. Ann N Y Acad Sci 1125, 82-99.

133
Gatermann, R., Biselli, S., Huhnerfuss, H., Rimkus, G.G., Hecker, M., Karbe, L., 2002. Synthetic Musks in the Environment. Part 1: Species-Dependent Bioaccumulation of Polycyclic and Nitro Musk Fragrances in Freshwater Fish and Mussels. Archives of Environmental Contamination and Toxicology 42, 437-446. George, K.W., Hay, A.G., 2011. Bacterial strategies for growth on aromatic compounds. Adv Appl Microbiol 74, 1-33. George, K.W., Kagle, J., Junker, L., Risen, A., Hay, A.G., 2011. Growth of Pseudomonas putida F1 on styrene requires increased catechol-2,3-dioxygenase activity, not a new hydrolase. Microbiology 157, 89-98. Gray, L.E., Furr, J., Ostby, J.S., 2005. Hershberger Assay to Investigate the Effects of Endocrine-Disrupting Compounds with Androgenic or Antiandrogenic Activity in Castrate-Immature Male Rats. Current Protocols in Toxicology. John Wiley & Sons, Inc., Hoboken, NJ, USA. Green, R.E., Newton, I., Shultz, S., Cunningham, A.A., Gilbert, M., Pain, D.J., Prakash, V., 2004. Diclofenac poisoning as a cause of vulture population declines across the Indian subcontinent. J Appl Ecol 41, 793-800. Gronen, S., Denslow, N., Manning, S., Barnes, S., Barnes, D., Brouwer, M., 1999. Serum Vitellogenin Levels and Reproductive Impairment of Male Japanese Medaka (Oryzias latipes) Exposed to 4-tert-Octylphenol. Environmental Health Perspectives 107, 385-390. Guess, W.L., Bruch, M., 1986. A Review of Available Toxicity Data on the Topical Antimicrobial, Chloroxylenol. Cutaneous and Ocular Toxicology 5, 233-262. Han, G.H., Hur, H.G., Kim, S.D., 2006. Ecotoxicological risk of pharmaceuticals from wastewater treatment plants in Korea: occurrence and toxicity to Daphnia magna. Environ Toxicol Chem 25, 265-271. Hanioka, N., Omae, E., Nishimura, T., Jinno, H., Onodera, S., Yoda, R., Ando, M., 1996. Interaction of 2,4,4'-trichloro-2'-hydroxydiphenyl ether with microsomal cytochrome p450-dependent monooxygenases in rat liver. Chemosphere 33, 265-276. Harayama, S., Kok, M., Neidle, E.L., 1992. Functional and evolutionary relationships among diverse oxygenases. Annu Rev Microbiol 46, 565-601. Hayes, T.B., 2005. Welcome to the revolution: integrative biology and assessing the impact of endocrine disruptors on environmental and public health. Integr Comp Biol 45, 321-329. Hayes, T.B., Menendez, K.P., 1999. The effect of sex steroids on primary and secondary sex differentiation in the sexually dichromatic reedfrog (Hyperolius argus: Hyperolidae) from the Arabuko Sokoke Forest of Kenya. Gen Comp Endocrinol 115, 188-199.

134
Heidler, J., Halden, R.U., 2008. Meta-analysis of mass balances examining chemical fate during wastewater treatment. Environ Sci Technol 42, 6324-6332. Heiss, G., Stolz, A., Kuhm, A.E., Muller, C., Klein, J., Altenbuchner, J., Knackmuss, H.J., 1995. Characterization of a 2,3-dihydroxybiphenyl dioxygenase from the naphthalenesulfonate-degrading bacterium strain BN6. J Bacteriol 177, 5865-5871. Herbstman, J.B., Sjodin, A., Kurzon, M., Lederman, S.A., Jones, R.S., Rauh, V., Needham, L.L., Tang, D., Niedzwiecki, M., Wang, R.Y., Perera, F., 2010. Prenatal exposure to PBDEs and neurodevelopment. Environ Health Perspect 118, 712-719. Hinck, J.E., Blazer, V.S., Schmitt, C.J., Papoulias, D.M., Tillitt, D.E., 2009. Widespread occurrence of intersex in black basses (Micropterus spp.) from U.S. rivers, 1995-2004. Aquat Toxicol 95, 60-70. Hiraoka, Y., Yamada, T., Tone, K., Futaesaku, Y., Kimbara, K., 2002. Flow cytometry analysis of changes in the DNA content of the polychlorinated biphenyl degrader Comamonas testosteroni TK102: effect of metabolites on cell-cell separation. Appl Environ Microbiol 68, 5104-5112. Hua, W.Y., Bennett, E.R., Maio, X.-S., Metcalfe, C.D., Letcher, R.J., 2006. Seasonality effects on pharmaceuticals and s-triazine herbicides in wastewater effluent and surface water from the Canadian side of the upper Detroit River. Environmental Toxicology and Chemistry / SETAC 25, 2356-2365. Hugo, N., Armengaud, J., Gaillard, J., Timmis, K.N., Jouanneau, Y., 1998. A novel -2Fe-2S- ferredoxin from Pseudomonas putida mt2 promotes the reductive reactivation of catechol 2,3-dioxygenase. J Biol Chem 273, 9622-9629. Hugo, N., Meyer, C., Armengaud, J., Gaillard, J., Timmis, K.N., Jouanneau, Y., 2000. Characterization of three XylT-like [2Fe-2S] ferredoxins associated with catabolism of cresols or naphthalene: evidence for their involvement in catechol dioxygenase reactivation. J Bacteriol 182, 5580-5585. Isidori, M., Lavorgna, M., Nardelli, A., Parrella, A., 2006. Toxicity on crustaceans and endocrine disrupting activity on Saccharomyces cerevisiae of eight alkylphenols. Chemosphere 64, 135-143. Jenkins, R., Angus, R.A., McNatt, H., Howell, W.M., Kemppainen, J.A., Kirk, M., Wilson, E.M., 2001. Identification of androstenedione in a river containing paper mill effluent. Environmental Toxicology and Chemistry / SETAC 20, 1325-1331. Jobling, S., Burn, R.W., Thorpe, K., Williams, R., Tyler, C., 2009. Statistical modeling suggests that antiandrogens in effluents from wastewater treatment works contribute to widespread sexual disruption in fish living in English rivers. Environmental Health Perspectives 117, 797-802.

135
Jobling, S., Coey, S., Whitmore, J.G., Kime, D.E., Van Look, K.J.W., McAllister, B.G., Beresford, N., Henshaw, A.C., Brighty, G., Tyler, C.R., Sumpter, J.P., 2002. Wild Intersex Roach (Rutilus rutilus) Have Reduced Fertility. Biology of Reproduction 67, 515-524. Jobling, S., Nolan, M., Tyler, C.R., Brighty, G., Sumpter, J.P., 1998. Widespread Sexual Disruption in Wild Fish. Environmental Science & Technology 32, 2498-2506. Jobling, S., Reynolds, T., White, R., Parker, M.G., Sumpter, J.P., 1995. A variety of environmentally persistent chemicals, including some phthalate plasticizers, are weakly estrogenic. Environmental Health Perspectives 103, 582-587. Jobling, S., Sumpter, J.P., 1993. Detergent components in sewage effluent are weakly oestrogenic to fish: An in vitro study using rainbow trout (Oncorhynchus mykiss) hepatocytes. Aquatic Toxicology 27, 361-372. Jobling, S., Williams, R., Johnson, A., Taylor, A., Gross-Sorokin, M., Nolan, M., Tyler, C.R., van Aerle, R., Santos, E., Brighty, G., 2006. Predicted Exposures to Steroid Estrogens in U.K. Rivers Correlate with Widespread Sexual Disruption in Wild Fish Populations. Environmental Health Perspectives 114, 32-39. Jonsson, G., Bechmann, R.K., Bamber, S.D., Baussant, T., 2004. Bioconcentration, biotransformation, and elimination of polycyclic aromatic hydrocarbons in sheepshead minnows (Cyprinodon variegatus) exposed to contaminated seawater. Environmental Toxicology and Chemistry / SETAC 23, 1538-1548. Kagle, J., Porter, A.W., Murdoch, R.W., Rivera-Cancel, G., Hay, A.G., 2009. Biodegradation of pharmaceutical and personal care products. Adv Appl Microbiol 67, 65-108. Kang, Y.S., Kim, Y.J., Jeon, C.O., Park, W., 2006. Characterization of Naphthalene-Degrading Psedomonas Species Isolated from Pollutant-Contaminated Sites: Oxidative Stress During their Growth on Naphthalene. J. Microbiol. Biotechnol. 16, 1819-1825. Kang, Y.S., Lee, Y., Jung, H., Jeon, C.O., Madsen, E.L., Park, W., 2007. Overexpressing antioxidant enzymes enhances naphthalene biodegradation in Pseudomonas sp. strain As1. Microbiology 153, 3246-3254. Kasprzyk-Hordern, B., Dinsdale, R.M., Guwy, A.J., 2009. The removal of pharmaceuticals, personal care products, endocrine disruptors and illicit drugs during wastewater treatment and its impact on the quality of receiving waters. Water Research 43, 363-380. Kawaguchi, M., Ito, R., Honda, H., Endo, N., Okanouchi, N., Saito, K., Seto, Y., Nakazawa, H., 2008. Simultaneous analysis of benzophenone sunscreen compounds in water sample by stir bar sorptive extraction with in situ derivatization and thermal desorption-gas chromatography-mass spectrometry. Journal of Chromatography. A 1200, 260-263.

136
Kidd, K.A., Blanchfield, P.J., Mills, K.H., Palace, V.P., Evans, R.E., Lazorchak, J.M., Flick, R.W., 2007. Collapse of a fish population after exposure to a synthetic estrogen. Proceedings of the National Academy of Sciences 104, 8897-8901. Kim, I., Yamashita, N., Tanaka, H., 2009. Performance of UV and UV/H2O2 processes for the removal of pharmaceuticals detected in secondary effluent of a sewage treatment plant in Japan. J Hazard Mater 166, 1134-1140. Kim, I.S., Lee, H., Trevors, J.T., 2001. Effects of 2,2',5,5'-tetrachlorobiphenyl and biphenyl on cell membranes of Ralstonia eutropha H850. FEMS Microbiol Lett 200, 17-24. Kloas, W., Lutz, I., Einspanier, R., 1999. Amphibians as a model to study endocrine disruptors: II. Estrogenic activity of environmental chemicals in vitro and in vivo. The Science of the Total Environment 225, 59-68. Knadler, M.P., Hall, S.D., 1990. Stereoselective arylpropionyl-CoA thioester formation in vitro. Chirality 2, 67-73. Knights, K.M., Drogemuller, C.J., 2000. Xenobiotic-CoA ligases: kinetic and molecular characterization. Curr Drug Metab 1, 49-66. Knutson, A.B., Klerks, P.L., Levinton, J.S., 1987. The fate of metal contaminated sediments in Foundry Cove, New York. Environmental Pollution 45, 291-304. Kolpin, D.W., Furlong, E.T., Meyer, M.T., Thurmam, E.M., Zaugg, S.D., Barber, L.B., Buxton, H.T., 2002. Pharmaceuticals, hormones, and other organic wastewater contaminants in U.S. streams, 1999-2000: a national reconnaissance. Environmental Science & Technology 36, 1202-1211. Kovach, M.E., Elzer, P.H., Hill, D.S., Robertson, G.T., Farris, M.A., Roop, R.M., 2nd, Peterson, K.M., 1995. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 166, 175-176. Kummer, V., Maskova, J., Zraly, Z., Neca, J., Simeckova, P., Vondracek, J., Machala, M., 2008. Estrogenic activity of environmental polycyclic aromatic hydrocarbons in uterus of immature Wistar rats. Toxicology Letters 180, 212-221. Kunz, P.Y., Fent, K., 2009. Estrogenic activity of ternary UV filter mixtures in fish (Pimephales promelas) -- An analysis with nonlinear isobolograms. Toxicology and Applied Pharmacology 234, 77-88. Lang, F.S., Oesterhelt, D., 1989. Gene transfer system for Rhodopseudomonas viridis. J Bacteriol 171, 4425-4435.

137
Lee, K.E., Yaeger, C.S., Jahns, N.D., Schoenfuss, H.L., 2008. Occurrence of Endocrine Active Compounds and Biological Responses in the Mississippi River‚ Study Design and Data, June through August 2006. U.S. Geological Survey Data Series 368, 27p. with Appendix. Lee, Y., Pena-Llopis, S., Kang, Y.S., Shin, H.D., Demple, B., Madsen, E.L., Jeon, C.O., Park, W., 2006. Expression analysis of the fpr (ferredoxin-NADP+ reductase) gene in Pseudomonas putida KT2440. Biochem Biophys Res Commun 339, 1246-1254. Lehotay, S.J., Harman-Fetcho, J.A., McConnell, L.L., 1998. Agricultural pesticide residues in oysters and water from two chesapeake bay tributaries. Marine Pollution Bulletin 37, 32-44. Letcher, R.J., Gebbink, W.A., Sonne, C., Born, E.W., McKinney, M.A., Dietz, R., 2009. Bioaccumulation and biotransformation of brominated and chlorinated contaminants and their metabolites in ringed seals (Pusa hispida) and polar bears (Ursus maritimus) from East Greenland. Environ Int 35, 1118-1124. Li, X., Brownawell, B.J., 2010. Quaternary Ammonium Compounds in Urban Estuarine Sediment Environments - A Class of Contaminants in Need of Increased Attention? Environmental Science & Technology 44, 7561-7568. Lister, D.L., Kanungo, G., Rathbone, D.A., Bruce, N.C., 1999. Transformations of codeine to important semisynthetic opiate derivatives by Pseudomonas putida m10. FEMS Microbiol Lett 181, 137-144. Liu, G., Niu, Z., Van Niekerk, D., Xue, J., Zheng, L., 2008. Polycyclic aromatic hydrocarbons (PAHs) from coal combustion: emissions, analysis, and toxicology. Reviews of Environmental Contamination and Toxicology 192, 1-28. Liu, G., Yang, L., Fan, T., Cong, R., Tang, Z., Sun, W., Meng, X., Zhu, L., 2006a. Purification and characterization of phenoloxidase from crab Charybdis japonica. Fish Shellfish Immunol 20, 47-57. Liu, H., Ye, W., Zhan, X., Liu, W., 2006b. A comparative study of rac- and S-metolachlor toxicity to Daphnia magna. Ecotoxicology and Environmental Safety 63, 451-455. Loos, R., Hanke, G., Umlauf, G., Eisenreich, S.J., 2007. LC-MS-MS analysis and occurrence of octyl- and nonylphenol, their ethoxylates and their carboxylates in Belgian and Italian textile industry, waste water treatment plant effluents and surface waters. Chemosphere 66, 690-699. Loyo-Rosales, J.E., Rice, C.P., Torrents, A., 2010. Fate and distribution of the octyl- and nonylphenol ethoxylates and some carboxylated transformation products in the Back River, Maryland. Journal of Environmental Monitoring 12, 614-614. Martinez-Carballo, E., Gonzalez-Barreiro, C., Sitka, A., Scharf, S., Gans, O., 2007. Determination of selected organophosphate esters in the aquatic environment of Austria. The Science of the Total Environment 388, 290-299.

138
Matamoros, V., Jover, E., Bayona, J.M., 2010. Part-per-Trillion Determination of Pharmaceuticals, Pesticides, and Related Organic Contaminants in River Water by Solid-Phase Extraction Followed by Comprehensive Two-Dimensional Gas Chromatography Time-of-Flight Mass Spectrometry. Analytical Chemistry 82, 699-706. Mazanti, L., Rice, C., Bialek, K., Sparling, D., Stevenson, C., Johnson, W.E., Kangas, P., Rheinstein, J., 2003. Aqueous-Phase Disappearance of Atrazine, Metolachlor, and Chlorpyrifos in Laboratory Aquaria and Outdoor Macrocosms. Archives of Environmental Contamination and Toxicology 44, 67-76. McPhail, M.K., 1934. The assay of progestin. The Journal of Physiology 83, 145-156. Mendola, P., Robinson, L.K., Buck, G.M., Druschel, C.M., Fitzgerald, E.F., Sever, L.E., Vena, J.E., 2005. Birth defects risk associated with maternal sport fish consumption: potential effect modification by sex of offspring. Environ Res 97, 134-141. Metcalfe, C.D., Koenig, B.G., Bennie, D.T., Servos, M., Ternes, T.A., Hirsch, R., 2003a. Occurrence of neutral and acidic drugs in the effluents of Canadian sewage treatment plants. Environ Toxicol Chem 22, 2872-2880. Metcalfe, C.D., Miao, X.-S., Koenig, B.G., Struger, J., 2003b. Distribution of acidic and neutral drugs in surface waters near sewage treatment plants in the lower Great Lakes, Canada. Environmental Toxicology and Chemistry / SETAC 22, 2881-2889. Miege, C., Choubert, J.M., Ribeiro, L., Eusebe, M., Coquery, M., 2008. Removal efficiency of pharmaceuticals and personal care products with varying wastewater treatment processes and operating conditions - conception of a database and first results. Water Sci Technol 57, 49-56. Munoz, R., Diaz, L.F., Bordel, S., Villaverde, S., 2007. Inhibitory effects of catechol accumulation on benzene biodegradation in Pseudomonas putida F1 cultures. Chemosphere 68, 244-252. Munro, N.B., Talmage, S.S., Griffin, G.D., Waters, L.C., Watson, A.P., King, J.F., Hauschild, V., 1999. The sources, fate, and toxicity of chemical warfare agent degradation products. Environ Health Perspect 107, 933-974. Murdoch, R.W., 2011. Dissertation - The Metabolism of Ibuprofen and Phenylacetic Acid by Sphingomonas Ibu-2. Cornell University. Murdoch, R.W., Hay, A.G., 2005. Formation of catechols via removal of acid side chains from ibuprofen and related aromatic acids. Appl Environ Microbiol 71, 6121-6125. Murray, K.E., Thomas, S.M., Bodour, A.A., 2010. Prioritizing research for trace pollutants and emerging contaminants in the freshwater environment. Environmental Pollution (Barking, Essex: 1987) 158, 3462-3471.

139
Nozaki, M., Ono, K., Nakazawa, T., Kotani, S., Hayaishi, O., 1968. Metapyrocatechase. II. The role of iron and sulfhydryl groups. J Biol Chem 243, 2682-2690. NYSDEC, 2004. Descriptive Data of Municipal Wastewater Treatment Plants - NYS Dept. of Environmental Conservation. NYS Dept. of Environmental Conservation, Division of Water Bureau of Water Compliance. Oaks, J.L., Gilbert, M., Virani, M.Z., Watson, R.T., Meteyer, C.U., Rideout, B.A., Shivaprasad, H.L., Ahmed, S., Chaudhry, M.J., Arshad, M., Mahmood, S., Ali, A., Khan, A.A., 2004. Diclofenac residues as the cause of vulture population decline in Pakistan. Nature 427, 630-633. Oblinger, C.J., Gill, A.C., McPherson, A.K., Meyer, M.T., Furlong, E.T., 2007. Occurrence of Selected Pharmaceuticals, Personal-Care Products, Organic Wastewater Compounds, and Pesticides in the Lower Tallapoosa River Watershed near Montgomery, Alabama, 2005. U.S. Geological Survey Scientific Investigations Report 2007-5266, 23 p. Okay, O.S., Karacik, B., 2008. Bioconcentration and phototoxicity of selected PAHs to marine mussel Mytilus galloprovincialis. Journal of Environmental Science and Health. Part A, Toxic/Hazardous Substances & Environmental Engineering 43, 1234-1242. Oliveira, R., Domingues, I., Koppe Grisolia, C., Soares, A.M.V.M., 2009. Effects of triclosan on zebrafish early-life stages and adults. Environmental Science and Pollution Research International 16, 679-688. Orvos, D.R., Versteeg, D.J., Inauen, J., Capdevielle, M., Rothenstein, A., Cunningham, V., 2002. Aquatic toxicity of triclosan. Environmental Toxicology and Chemistry / SETAC 21, 1338-1349. Palmer, P.M., Wilson, L.R., O'Keefe, P., Sheridan, R., King, T., Chen, C.-Y., 2008. Sources of pharmaceutical pollution in the New York City Watershed. Science of The Total Environment 394, 90-102. Park, B.J., Kidd, K.A., 2005. Effects of the synthetic estrogen ethinylestradiol on early life stages of mink frogs and green frogs in the wild and in situ. Environmental Toxicology and Chemistry / SETAC 24, 2027-2036. Park, D.W., Chae, J.C., Kim, Y., Iida, T., Kudo, T., Kim, C.K., 2002. Chloroplast-type ferredoxin involved in reactivation of catechol 2,3-dioxygenase from Pseudomonas sp. S 47. J Biochem Mol Biol 35, 432-436. Park, W., Jeon, C.O., Cadillo, H., DeRito, C., Madsen, E.L., 2004. Survival of naphthalene-degrading Pseudomonas putida NCIB 9816-4 in naphthalene-amended soils: toxicity of naphthalene and its metabolites. Appl Microbiol Biotechnol 64, 429-435. Parks, L.G., Lambright, C.S., Orlando, E.F., Guillette, L.J., Ankley, G.T., Gray, L.E., 2001. Masculinization of Female Mosquitofish in Kraft Mill Effluent-Contaminated Fenholloway

140
River Water Is Associated with Androgen Receptor Agonist Activity. Toxicological Sciences 62, 257-267. Passos, A.L.S., Pinto, P.I.S., Power, D.M., Canario, A.V.M., 2009. A yeast assay based on the gilthead sea bream (teleost fish) estrogen receptor beta for monitoring estrogen mimics. Ecotoxicology and Environmental Safety 72, 1529-1537. Perez, S., Barcelo, D., 2008. First evidence for occurrence of hydroxylated human metabolites of diclofenac and aceclofenac in wastewater using QqLIT-MS and QqTOF-MS. Anal Chem 80, 8135-8145. Perez-Pantoja, D., Ledger, T., Pieper, D.H., Gonzalez, B., 2003. Efficient turnover of chlorocatechols is essential for growth of Ralstonia eutropha JMP134(pJP4) in 3-chlorobenzoic acid. J Bacteriol 185, 1534-1542. Phillips, P.J., Smith, S.G., Kolpin, D.W., Zaugg, S.D., Buxton, H.T., Furlong, E.T., 2010a. Method Description, Quality Assurance, Environmental Data, and other Information for Analysis of Pharmaceuticals in Wastewater-Treatment-Plant Effluents, Streamwater, and Reservoirs, 2004-2009. U.S. Geological Survey Open-File Report 2010-1102, 36 p. Phillips, P.J., Smith, S.G., Kolpin, D.W., Zaugg, S.D., Buxton, H.T., Furlong, E.T., Esposito, K., Stinson, B., 2010b. Pharmaceutical Formulation Facilities as Sources of Opioids and Other Pharmaceuticals to Wastewater Treatment Plant Effluents. Environmental Science & Technology 44, 4910-4916. Pies, C., Ternes, T.A., Hofmann, T., 2008. Identifying sources of polycyclic aromatic hydrocarbons (PAHs) in soils: distinguishing point and non-point sources using an extended PAH spectrum and n-alkanes. Journal of Soils and Sediments 8, 312-322. Poiger, T., Buser, H.R., Muller, M.D., 2001. Photodegradation of the pharmaceutical drug diclofenac in a lake: pathway, field measurements, and mathematical modeling. Environ Toxicol Chem 20, 256-263. Polissi, A., Harayama, S., 1993. In vivo reactivation of catechol 2,3-dioxygenase mediated by a chloroplast-type ferredoxin: a bacterial strategy to expand the substrate specificity of aromatic degradative pathways. EMBO J 12, 3339-3347. Powlowski, J., Shingler, V., 1994. Genetics and biochemistry of phenol degradation by Pseudomonas sp. CF600. Biodegradation 5, 219-236. Quinn, B., Gagne, F., Blaise, C., 2008. An investigation into the acute and chronic toxicity of eleven pharmaceuticals (and their solvents) found in wastewater effluent on the cnidarian, Hydra attenuata. The Science of the Total Environment 389, 306-314.

141
Quintana, J.B., Weiss, S., Reemtsma, T., 2005. Pathways and metabolites of microbial degradation of selected acidic pharmaceutical and their occurrence in municipal wastewater treated by a membrane bioreactor. Water Res 39, 2654-2664. Radjenovic, J., Perez, S., Petrovic, M., Barcelo, D., 2008. Identification and structural characterization of biodegradation products of atenolol and glibenclamide by liquid chromatography coupled to hybrid quadrupole time-of-flight and quadrupole ion trap mass spectrometry. J Chromatogr A 1210, 142-153. Ramirez, A.J., Brain, R.A., Usenko, S., Mottaleb, M.A., O'Donnell, J.G., Stahl, L.L., Wathen, J.B., Snyder, B.D., Pitt, J.L., Perez-Hurtado, P., Dobbins, L.L., Brooks, B.W., Chambliss, C.K., 2009. Occurrence of pharmaceuticals and personal care products in fish: results of a national pilot study in the United States. Environmental Toxicology and Chemistry / SETAC 28, 2587-2597. Ramos, J.L., Wasserfallen, A., Rose, K., Timmis, K.N., 1987. Redesigning metabolic routes: manipulation of TOL plasmid pathway for catabolism of alkylbenzoates. Science 235, 593-596. Rather, L.J., Knapp, B., Haehnel, W., Fuchs, G., 2010. Coenzyme A-dependent aerobic metabolism of benzoate via epoxide formation. J Biol Chem 285, 20615-20624. Raut, S.A., Angus, R.A., 2010. Triclosan has endocrine-disrupting effects in male western mosquitofish, Gambusia affinis. Environmental Toxicology and Chemistry / SETAC 29, 1287-1291. Reddy, S., Iden, C.R., Brownawell, B.J., 2005. Analysis of steroid conjugates in sewage influent and effluent by liquid chromatography-tandem mass spectrometry. Anal Chem 77, 7032-7038. Ren, X., Lee, Y.J., Han, H.J., Kim, I.S., 2008. Effect of tris-(2-chloroethyl)-phosphate (TCEP) at environmental concentration on the levels of cell cycle regulatory protein expression in primary cultured rabbit renal proximal tubule cells. Chemosphere 74, 84-88. Repine, J.E., Fox, R.B., Berger, E.M., 1981. Hydrogen peroxide kills Staphylococcus aureus by reacting with staphylococcal iron to form hydroxyl radical. J Biol Chem 256, 7094-7096. Rescigno, A., Sollai, F., Pisu, B., Rinaldi, A., Sanjust, E., 2002. Tyrosinase inhibition: general and applied aspects. J Enzyme Inhib Med Chem 17, 207-218. Risen, A.J., Hay, A.G., Smith, S.G., Zaugg, S.D., Phillips, P.J., 2009. Pharmaceuticals, precursors and metabolites in wastewater impacted by pharmaceutical manufacturing facilities. EmCon, Colorado State University. Fort Collins, CO, USA. Rosas, S.B., Secco, M., Ghittoni, N.E., 1980. Effects of pesticides on the fatty acid and phospholipid composition of Escherichia coli. Appl Environ Microbiol 40, 231-234.

142
Sabourin, L., Beck, A., Duenk, P.W., Kleywegt, S., Lapen, D.R., Li, H., Metcalfe, C.D., Payne, M., Topp, E., 2009. Runoff of pharmaceuticals and personal care products following application of dewatered municipal biosolids to an agricultural field. The Science of the Total Environment 407, 4596-4604. Sacher, F., Ehmann, M., Gabriel, S., Graf, C., Brauch, H.-J., 2008. Pharmaceutical residues in the river Rhine--results of a one-decade monitoring programme. Journal of Environmental Monitoring 10, 664-664. Sala, M., Gu, Z.G., Moens, G., Chouroulinkov, I., 1982. In vivo and in vitro biological effects of the flame retardants tris(2,3-dibromopropyl) phosphate and tris(2-chlorethyl)orthophosphate. European Journal of Cancer & Clinical Oncology 18, 1337-1344. Sala-Trepat, J.M., Evans, W.C., 1971. The meta cleavage of catechol by Azotobacter species. 4-Oxalocrotonate pathway. Eur J Biochem 20, 400-413. Schleier, J.J., 3rd, Peterson, R.K.D., Macedo, P.A., Brown, D.A., 2008. Environmental concentrations, fate, and risk assessment of pyrethrins and piperonyl butoxide after aerial ultralow-volume applications for adult mosquito management. Environmental Toxicology and Chemistry / SETAC 27, 1063-1068. Schmidt, C.W., 2009. TOX 21: New Dimensions of Toxicity Testing. Environmental Health Perspectives 117, A348-A353-A348-A353. Schreurs, R.H.M.M., Quaedackers, M.E., Seinen, W., van der Burg, B., 2002. Transcriptional Activation of Estrogen Receptor ER[alpha] and ER[beta] by Polycyclic Musks Is Cell Type Dependent. Toxicology and Applied Pharmacology 183, 1-9. Schreurs, R.H.M.M., Sonneveld, E., Jansen, J.H.J., Seinen, W., van der Burg, B., 2005. Interaction of polycyclic musks and UV filters with the estrogen receptor (ER), androgen receptor (AR), and progesterone receptor (PR) in reporter gene bioassays. Toxicological Sciences: An Official Journal of the Society of Toxicology 83, 264-272. Schriks, M., van Leerdam, J.A., van der Linden, S.C., van der Burg, B., van Wezel, A.P., de Voogt, P., 2010. High-Resolution Mass Spectrometric Identification and Quantification of Glucocorticoid Compounds in Various Wastewaters in The Netherlands. Environmental Science & Technology 44, 4766-4774. Schussler, W., Nitschke, L., 1999. Death of fish due to surface water pollution by liquid manure or untreated wastewater: analytical preservation of evidence by HPLC. Water Research 33, 2884-2887. Schweigert, N., Zehnder, A.J., Eggen, R.I., 2001. Chemical properties of catechols and their molecular modes of toxic action in cells, from microorganisms to mammals. Environ Microbiol 3, 81-91.

143
Shala, L., Foster, G.D., 2010. Surface water concentrations and loading budgets of pharmaceuticals and other domestic-use chemicals in an urban watershed (Washington, DC, USA). Archives of Environmental Contamination and Toxicology 58, 551-561. Shao, B., Chen, D., Zhang, J., Wu, Y., Sun, C., 2009. Determination of 76 pharmaceutical drugs by liquid chromatography-tandem mass spectrometry in slaughterhouse wastewater. Journal of Chromatography A 1216, 8312-8318. Sikkema, J., de Bont, J.A., Poolman, B., 1994. Interactions of cyclic hydrocarbons with biological membranes. J Biol Chem 269, 8022-8028. Sikkema, J., Poolman, B., Konings, W.N., de Bont, J.A., 1992. Effects of the membrane action of tetralin on the functional and structural properties of artificial and bacterial membranes. J Bacteriol 174, 2986-2992. Sinclair, E., Mayack, D.T., Roblee, K., Yamashita, N., Kannan, K., 2006. Occurrence of Perfluoroalkyl Surfactants in Water, Fish, and Birds from New York State. Archives of Environmental Contamination and Toxicology 50, 398-410. Singh, S.P., Azua, A., Chaudhary, A., Khan, S., Willett, K.L., Gardinali, P.R., 2009. Occurrence and distribution of steroids, hormones and selected pharmaceuticals in South Florida coastal environments. Ecotoxicology 19, 338-350. Skonberg, C., Olsen, J., Madsen, K.G., Hansen, S.H., Grillo, M.P., 2008. Metabolic activation of carboxylic acids. Expert Opin Drug Metab Toxicol 4, 425-438. Skupinska, K., Misiewicz, I., Kasprzycka-Guttman, T., 2004. Polycyclic aromatic hydrocarbons: physicochemical properties, environmental appearance and impact on living organisms. Acta Poloniae Pharmaceutica 61, 233-240. Smith, D.G., 2004. Surface- and ground-water monitoring and mapping of selected features at the Mt. Pisgah Campground, Haywood County, North Carolina, 2002. U.S. Geological Survey Open-File Report 2004-1073, 33p. Smith, W.L., DeWitt, D.L., Garavito, R.M., 2000. Cyclooxygenases: structural, cellular, and molecular biology. Annu Rev Biochem 69, 145-182. Sonneveld, E., Jansen, H.J., Riteco, J.A.C., Brouwer, A., van der Burg, B., 2005. Development of Androgen- and Estrogen-Responsive Bioassays, Members of a Panel of Human Cell Line-Based Highly Selective Steroid-Responsive Bioassays. Toxicological Sciences 83, 136-148. Sonneveld, E., Pieterse, B., Schoonen, W.G., van der Burg, B., 2011. Validation of in vitro screening models for progestagenic activities: Inter-assay comparison and correlation with in vivo activity in rabbits. Toxicology in Vitro 25, 545-554.

144
Sonneveld, E., Riteco, J.A.C., Jansen, H.J., Pieterse, B., Brouwer, A., Schoonen, W.G., van der Burg, B., 2006. Comparison of In Vitro and In Vivo Screening Models for Androgenic and Estrogenic Activities. Toxicological Sciences 89, 173-187. Standley, L.J., Kaplan, L.A., Smith, D., 2000. Molecular Tracers of Organic Matter Sources to Surface Water Resources. Environmental Science & Technology 34, 3124-3130. Stratton, G.W., Corke, C.T., 1982. Comparative Fungitoxicity of the Insecticide Permethrin and Ten Degradation Products. Pestic Sci 13, 679-685. Sumner, N.R., Guitart, C., Fuentes, G., Readman, J.W., 2010. Inputs and distributions of synthetic musk fragrances in an estuarine and coastal environment; a case study. Environmental Pollution 158, 215-222. Sundkvist, A.M., Olofsson, U., Haglund, P., 2010. Organophosphorus flame retardants and plasticizers in marine and fresh water biota and in human milk. Journal of Environmental Monitoring 12, 943-943. Swan, G., Naidoo, V., Cuthbert, R., Green, R.E., Pain, D.J., Swarup, D., Prakash, V., Taggart, M., Bekker, L., Das, D., Diekmann, J., Diekmann, M., Killian, E., Meharg, A., Patra, R.C., Saini, M., Wolter, K., 2006. Removing the threat of diclofenac to critically endangered Asian vultures. PLoS Biol 4, e66. Tan, S.C., Patel, B.K., Jackson, S.H., Swift, C.G., Hutt, A.J., 2002. Stereoselectivity of ibuprofen metabolism and pharmacokinetics following the administration of the racemate to healthy volunteers. Xenobiotica 32, 683-697. Teufel, R., Gantert, C., Voss, M., Eisenreich, W., Haehnel, W., Fuchs, G., 2011. Studies on the mechanism of ring hydrolysis in phenylacetate degradation: a metabolic branching point. J Biol Chem 286, 11021-11034. Teufel, R., Mascaraque, V., Ismail, W., Voss, M., Perera, J., Eisenreich, W., Haehnel, W., Fuchs, G., 2010. Bacterial phenylalanine and phenylacetate catabolic pathway revealed. Proc Natl Acad Sci U S A 107, 14390-14395. Toft, G., Baatrup, E., Guillette, L.J., Jr., 2004. Altered social behavior and sexual characteristics in mosquitofish (Gambusia holbrooki) living downstream of a paper mill. Aquat Toxicol 70, 213-222. Tropel, D., Meyer, C., Armengaud, J., Jouanneau, Y., 2002. Ferredoxin-mediated reactivation of the chlorocatechol 2,3-dioxygenase from Pseudomonas putida GJ31. Arch Microbiol 177, 345-351. USEPA, 1994. Reregistration Eligibility Decision Chloroxylenol List C Case 3045. United States Environmental Protection Agency.

145
Vaillancourt, F.H., Labbe, G., Drouin, N.M., Fortin, P.D., Eltis, L.D., 2002. The mechanism-based inactivation of 2,3-dihydroxybiphenyl 1,2-dioxygenase by catecholic substrates. J Biol Chem 277, 2019-2027. Vajda, A.M., Barber, L.B., Gray, J.L., Lopez, E.M., Woodling, J.D., Norris, D.O., 2008. Reproductive Disruption in Fish Downstream from an Estrogenic Wastewater Effluent. Environmental Science & Technology 42, 3407-3414. Van der Linden, S.C., Heringa, M.B., Man, H.-Y., Sonneveld, E., Puijker, L.M., Brouwer, A., Van der Burg, B., 2008. Detection of multiple hormonal activities in wastewater effluents and surface water, using a panel of steroid receptor CALUX bioassays. Environmental Science & Technology 42, 5814-5820. Vryzas, Z., Alexoudis, C., Vassiliou, G., Galanis, K., Papadopoulou-Mourkidou, E., 2011. Determination and aquatic risk assessment of pesticide residues in riparian drainage canals in northeastern Greece. Ecotoxicology and Environmental Safety 74, 174-181. Wan, M.T., Buday, C., Schroeder, G., Kuo, J., Pasternak, J., 2006. Toxicity to Daphnia magna, Hyalella azteca, Oncorhynchus kisutch, Oncorhynchus mykiss, Oncorhynchus tshawytscha, and Rana catesbeiana of Atrazine, Metolachlor, Simazine, and Their Formulated Products. Bulletin of Environmental Contamination and Toxicology 76, 52-58. Wang, L.-Q., Falany, C.N., James, M.O., 2004. Triclosan as a substrate and inhibitor of 3'-phosphoadenosine 5'-phosphosulfate-sulfotransferase and UDP-glucuronosyl transferase in human liver fractions. Drug Metabolism and Disposition: The Biological Fate of Chemicals 32, 1162-1169. Wang, X., Zhao, X., 2009. Contribution of oxidative damage to antimicrobial lethality. Antimicrob Agents Chemother 53, 1395-1402. Watson, C.S., Bulayeva, N.N., Wozniak, A.L., Alyea, R.A., 2007. Xenoestrogens are potent activators of nongenomic estrogenic responses. Steroids 72, 124-134. Weinstein, J.E., Garner, T.R., 2008. Piperonyl butoxide enhances the bioconcentration and photoinduced toxicity of fluoranthene and benzo[a]pyrene to larvae of the grass shrimp (Palaemonetes pugio). Aquatic Toxicology (Amsterdam, Netherlands) 87, 28-36. Weisbrod, C.J., Kunz, P.Y., Zenker, A.K., Fent, K., 2007. Effects of the UV filter benzophenone-2 on reproduction in fish. Toxicology and Applied Pharmacology 225, 255-266. Whitall, D., Hively, W.D., Leight, A.K., Hapeman, C.J., McConnell, L.L., Fisher, T., Rice, C.P., Codling, E., McCarty, G.W., Sadeghi, A.M., Gustafson, A., Bialek, K., 2010. Pollutant fate and spatio-temporal variability in the choptank river estuary: factors influencing water quality. The Science of the Total Environment 408, 2096-2108.

146
WHO, 1998. Environment health criteria No. 209: Flame retardants : tris(chloropropyl) phosphate and tris(2-chloroethyl) phosphate. World Health Organization, Geneva. Wilding, B.C., Curtis, K., Welker-Hood, K., 2011. Hazardous Chemicals in Health Care- A Snapshot of Chemicals in Doctors and Nurses www.psr.org/assets/pdfs/hazardous-chemicals-in-health-care.pdf. Witorsch, R.J., Thomas, J.A., 2010. Personal care products and endocrine disruption: A critical review of the literature. Crit Rev Toxicol 40 Suppl 3, 1-30. Wolf, M.C., Moore, P.A., 2002. Effects of the Herbicide Metolachlor on the Perception of Chemical Stimuli by Orconectes rusticus. Journal of the North American Benthological Society 21, 457-467. Yamada, T., Shimomura, Y., Hiraoka, Y., Kimbara, K., 2006. Oxidative stress by biphenyl metabolites induces inhibition of bacterial cell separation. Appl Microbiol Biotechnol 73, 452-457. Yamauchi, R., Ishibashi, H., Hirano, M., Mori, T., Kim, J.-W., Arizono, K., 2008. Effects of synthetic polycyclic musks on estrogen receptor, vitellogenin, pregnane X receptor, and cytochrome P450 3A gene expression in the livers of male medaka (Oryzias latipes). Aquatic Toxicology (Amsterdam, Netherlands) 90, 261-268. Yim, U.H., Hong, S.H., Shim, W.J., Oh, J.R., Chang, M., 2005. Spatio-temporal distribution and characteristics of PAHs in sediments from Masan Bay, Korea. Marine Pollution Bulletin 50, 319-326. Yoon, Y., Ryu, J., Oh, J., Choi, B.-G., Snyder, S.A., 2010. Occurrence of endocrine disrupting compounds, pharmaceuticals, and personal care products in the Han River (Seoul, South Korea). Science of The Total Environment 408, 636-643. Zaugg, S.D., Smith, S.G., Schroeder, Barber, L.B., Burkhardt, M.R., 2007. Determination of Wastewater Compounds in Whole Water by Continuous Liquid‚ Liquid Extraction and Capillary-Column Gas Chromatography/Mass Spectrometry. U.S. Geological Survey Water-Resources Investigations Report 01-4186, 37 p. Zhang, X., Yao, Y., Zeng, X., Qian, G., Guo, Y., Wu, M., Sheng, G., Fu, J., 2008. Synthetic musks in the aquatic environment and personal care products in Shanghai, China. Chemosphere 72, 1553-1558. Zulkosky, A.M., Ruggieri, J.P., Terracciano, S.A., Brownawell, B.J., 2005. Acute toxicity of resmethrin, malathion and methoprene to larval and juvenile American lobsters and analysis of pesticide levels in surface waters after Scourge, Anvil and Altosid application.