16.11.14 (???) bkj · ! 4! tak til… jeg vil gerne takke min vejleder, mikael kristian sejr,...
TRANSCRIPT


2
Tak til… ......................................................................................................................................................... 4
Forord ............................................................................................................................................................ 5
Introduktion ................................................................................................................................................ 6
Tungmetaller i det arktiske Grønland .......................................................................................................... 7 Den arktiske tidevandszone ............................................................................................................................. 9 Blåmuslingen Mytilus edulis .......................................................................................................................... 12 Blåmuslingens økologi og biogeografi ...................................................................................................................... 12 Blåmuslingen som miljøindikator ............................................................................................................................... 13 Blåmuslingens kuldetolerance ..................................................................................................................................... 14 Blykontaminerede blåmuslinger i Arktis ................................................................................................................. 15
Cellemembraner hos ektotermer stresses af kulde og bly ................................................................. 16 Cellemembranen ................................................................................................................................................................. 16 Cellemembranens temperatursensitivitet .............................................................................................................. 18 Kulde stresser membranen hos ektoterme organismer ................................................................................... 18 Peroxidering af cellemembranen ................................................................................................................................ 19 Bly øger peroxidering og stresser membranen .................................................................................................... 20
Synergetisk stressinteraktion mellem kulde og tungmetal ............................................................... 21 Aktuelt studie ..................................................................................................................................................... 23
Manuscript ................................................................................................................................................ 27
Abstract ................................................................................................................................................................ 29 1. Introduction .................................................................................................................................................... 30 2. Materials and Methods ................................................................................................................................ 33 2.1 Animals and holding conditions ........................................................................................................................... 33 2.2 Experimental design .................................................................................................................................................. 33 2.3 Pb analysis ..................................................................................................................................................................... 34 2.4 PLFA analysis ................................................................................................................................................................ 34 2.5 Statistical analysis ...................................................................................................................................................... 35
3. Results .............................................................................................................................................................. 35 3.1 Pb content in mussels ............................................................................................................................... 35 3.2 Effects of Pb on PLFAs .............................................................................................................................................. 36 3.3 Effects of sub-‐zero temperatures and Pb on survival ................................................................................. 37
4. Discussion ....................................................................................................................................................... 39 4.1 Internal lead .................................................................................................................................................................. 39

3
4.2 Pb effect on PLFA composition and survival .................................................................................................. 40 4.3 Implications for risk-‐assessments ....................................................................................................................... 41
Acknowledgements .......................................................................................................................................... 42 References ........................................................................................................................................................... 43

4
Tak til… Jeg vil gerne takke min vejleder, Mikael Kristian Sejr, Seniorforsker ved Arktisk
Forskningscenter, Institut for Bioscience, Aarhus Universitet, for god og brugbar vejledning og en
stor villighed til at give mig de bedste rammer for mit speciale. Jeg vil også takke Jakob Thyrring,
Ph.d. studerende ved Arktisk Forskningscenter, Institut for Bioscience, Aarhus Universitet, som har
været medvejleder og en stabil, hjælpsom og inspirerende støtte og samarbejdspartner igennem hele
projektet. Endvidere vil jeg takke Martin Holmstrup, Professor ved Jordfaunaøkologi og
Økotoksikologi, Institut for Bioscience, Aarhus Universitet, for vejledning og hjælp til
gennemførelse af og forståelse for laboratoriearbejdet. I denne forbindelse vil jeg også takke Lise
Lauridsen, Stine Slotsbo og Ana Silva Patricio, som ligeledes har været meget behjælpelige og
imødekommende i forbindelse med mit laboratoriearbejde. Jeg vil gerne takke Matilde Rasmussen
og Siri Elmegaard for inspirerende feedback og mine kontormakkere Ane Middelbo og Sofie
Mathiessen for mange gode timer på kontoret. Afslutningsvis takker jeg Bo Michelsen samt familie
og venner for støtte og opmuntring igennem hele processen.

5
Forord Hypotesen i dette specialestudie er, at der er synergi imellem effekterne af bly i økologisk
realistiske koncentrationer og realistisk temperatur på blåmuslingens kuldetolerance i Arktis. Som
mulig mekanistisk forklaring herpå undersøges det, hvorvidt den potentielle synergetiske
interaktion på kuldetolerancen skyldes, at bly resulterer i membranskader via ændringer i
cellemembranens fedtsyrekomposition. Specialet består af en introduktion til emnet, hvor den
teoretiske baggrund for hypotesen gennemgås. Efterfølgende forefindes manuskript til en artikel
omhandlende selve studiet med henblik på udgivelse i det videnskabelige tidsskrift Aquatic
Toxicology.
Introduktionen giver først et overblik over den grønlandske minehistorie og hvordan
tungmetalforurening, såsom bly, i visse marine områder udgør en risiko for den arktiske flora og
fauna. Herefter følger en kort beskrivelse af den arktiske tidevandszone, hvor arktiske organismer
udsættes for ekstrem kulde. Det tredje afsnit omhandler specialets forsøgsorganisme, blåmuslingen,
som er en dominerende art i den arktiske tidevandszone og en hyppigt anvendt marin miljøindikator
for tungmetaller. Fjerde afsnit er et teoretisk overblik over cellemembranens fysiske og fysiologiske
egenskaber og vil belyse hvorledes denne stresses af både kulde og tungmetal. Dette afsnit danner
således baggrund for vores hypotese om den mulige synergetiske stresseffekt af disse to faktorer på
blåmuslingens kuldetolerance. Afslutningsvis opsummeres det teoretiske ræsonnement for det
aktuelle studie, som manuskriptet omhandler.

6
Introduktion
Test af mulige synergetiske
interaktioner mellem frysegrader og bly (Pb) kontaminering i grønlandske blåmuslinger
(Mytilus edulis)

7
Tungmetaller i det arktiske Grønland Tidligere blev havet opfattet som et enormt reservoir for sikker bortskaffelse af forurenende
stoffer såsom tungmetaller (Ansari et al. 2004, Torres et al. 2008). I dag er marin forurening med
tungmetaller af antropogen oprindelse en global bekymring (AMAP 2005, Reichelt-Brushett 2012).
Dette skyldes tungmetallers bioakkumulering i, og mulige toksiske effekter på, marine organismer
(Josefson et al. 2008, Torres et al. 2008, Søndergaard 2013). Den primære tungmetalforurening af
Arktis sker via atmosfæren. Dette, samt minedriften i Arktis, har i de seneste årtier medført en
tiltagende bekymring for forurening af det arktiske marine økosystem (AMAP 2010). I Arktis har
cadmium (Cd) og kviksølv (Hg) størst økotoksikologisk betydning. Disse metaller findes i så høje
koncentrationer i arktisk flora og fauna, at det har sundhedsmæssige konsekvenser for organismerne
selv, men også for de mennesker der indtager dem (AMAP 2005). Endvidere er også bly fortsat en
kilde til bekymring i Arktis på trods af, at det atmosfæriske indhold af bly er faldet signifikant efter
forbuddet imod blyholdigt benzin i den vestlige verden (AMAP 2005). Særligt i visse marine
områder nær minedrift findes der fortsat høje blykoncentrationer, hvilket uddybes nedenfor.
Minedrift har fundet sted i Grønland siden 1840’erne og landet er fortsat særdeles rigt på mange
ressourcer såsom petroleum, gas, olie og forskellige mineraler, eksempelvis bly (Pb), zink (Zn), jern
(Fe), kobber (Cu) og guld (Au) (Figur 1) (Rosing et al. 2014). Vedtagelsen af Grønlands
Selvstyrelov i 2009 betød, at Grønland fik rettighederne over landets ressourcerigdom, inklusive
underjordiske mineralressourcer. Dette har medført en politisk og økonomisk vilje til i langt højere
grad at udnytte ressourcerne ved blandt andet minedrift, så landet kan opnå sin målsætning om at
blive en selvstændig økonomi (Rosing et al. 2014). Eksempelvis har Grønland i de seneste år solgt
et stigende antal minerettigheder til udenlandske minedriftskompagnier (Nuttall 2014) og minedrift
forventes at bidrage betydeligt til den grønlandske økonomi allerede indenfor de næste 5-10 år
(Rosing et al. 2014).
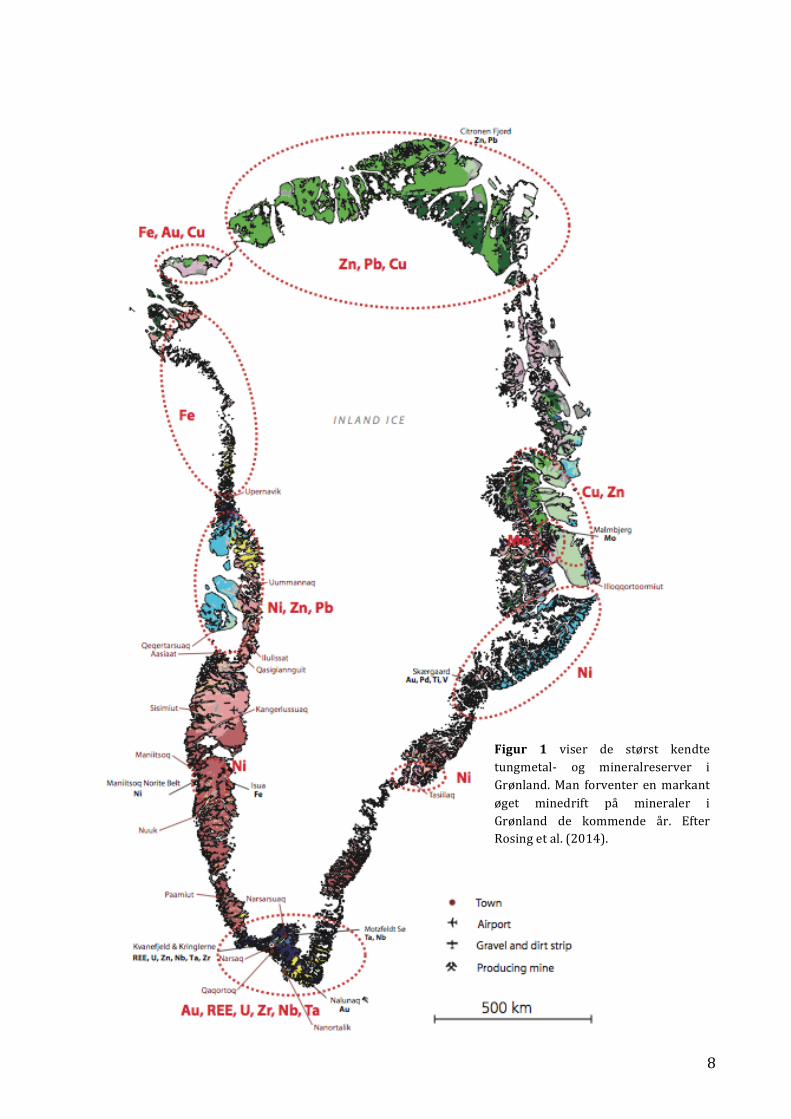
8
Figur 1 viser de størst kendte tungmetal-‐ og mineralreserver i Grønland. Man forventer en markant øget minedrift på mineraler i Grønland de kommende år. Efter Rosing et al. (2014).

9
Udledning af mineaffald har betydet en markant tungmetalforurening af nogle grønlandske
fjorde. Den forhenværende bly- og zinkmine Black Angel i Maarmorilik, Vestgrønland, lukkede
årligt 500.000 tons mineaffald direkte ud i det omkringliggende fjordsystem indtil den lukkede i
1990 efter 17 års drift (Loring & Asmund 1989b). Da minedriften var på sit højeste, målte man
blyindhold i blåmuslinger (Mytilus edulis) på op til 800 µg/g (baggrundsindholdet var 3 µg/g) og
man observerede bly- og zinkkontaminerede organismer op til 50 km fra minen (Loring & Asmund
1989b). Regelmæssig monitering fra minens åbning og frem til 2009 har vist, at koncentrationen af
opløst bly og zink i havvandet faldt kraftigt ved minens lukning (0,1% i 1995 i forhold til 1988
(Schiedek et al. 2009)). Dog er indholdet af bly og zink i fjordsystemets organismer og sediment
stadig kraftigt forhøjet 20 år efter minens lukning og dette forhøjede indhold ses op til 12 km fra
minen (Søndergaard et al. 2011). Man har således for nylig fundet blåmuslinger ved Maarmorilik
med blyindhold på op imod 3000 µg/g (Bach, 2014, pers. komm.) og man forventer at se forhøjede
niveauer af zink og bly i fjordsystemets planter og dyr de næste 50 år (Søndergaard et al. 2011).
Rettighederne til Black Angel minen blev solgt i 2008 og bly-, zink- og sølvudvinding forventes
genoptaget inden for de næste par år (Råstofstyrelsen 2013).
Lignende eksempler på tungmetalforurening af grønlandske fjorde fra minedrift er fra en
olivinmine i Fiskefjorden i Sydvestgrønland (Søndergaard 2013) samt tungmetaludledning fra en
kryolitmine i Ivittuut i Sydgrønland. I sidstnævnte eksempel måltes i 2007, 20 år efter
kryolitminens lukning, et blyindhold i blåmuslinger på op til 200-500 gange baggrundsniveau (0,7-
1,7 µg/g) (Johansen et al. 2008b). Tungmetalforurening har således langvarige konsekvenser for
den arktiske flora og fauna i Grønland og dette kan forblive et økologisk problem mange år endnu -
både på grund af bioakkumulering i arktiske organismer og på grund af den forventede øgede
mineaktivitet i Grønland.
Den arktiske tidevandszone Arktis kan defineres som området nord for polarcirklen (66°32′ N), men i denne introduktion
defineres Arktis ud fra en isoterm med en maksimal gennemsnitlig lufttemperatur på 10°C i årets
varmeste måned, hvilket økologisk set svarer til de 4,8% af jordens overflade nord for trægrænsen
(Figur 2) (AMAP 1998, CAFF 2013).

10
Arktis rummer nogle af de mest ekstreme
habitater på jorden kendetegnet ved
permanent eller midlertidigt isdække samt
op til 6 måneders kontinuerligt vintermørke,
hvor primærproduktionen stort set ophører
(Sejr, pers. komm.). Temperaturen kan
komme ned på -25°C i vintermånederne, og
der er store temperatursvingninger både
dagligt og over året (Blicher et al. 2013).
Der er endvidere store stedslige variationer i
temperaturen imellem de enkelte
mikrohabitater. Dette fremgår eksempelvis
af figur 3, som viser, at alger har en
markant isolerende effekt relativt til
eksponerede klipper i både den øvre og
midterste tidevandszone (Sejr et al.,
upubliceret).
Figur 2 viser forskellige definitioner på Arktis. Den stiplede blå linie angiver polarcirklen, imens den røde linie angiver en isoterm med en maksimal gennemsnitlig lufttemperatur på 10°C i årets varmeste måned. Når der i denne introduktion tales om Arktis menes der området nord for denne isoterm.
Figur 3 viser temperaturen i tidevandszonen over året 2011-‐2012 i Nuuk, Grønland. Temperaturen vises i (A) den øvre tidevandszone, (B) den midterste tidevandszone og (C) den nedre tidevandszone. Temperaturen vises både på eksponerede klipper (rød graf) og under alger (sort graf). Efter Sejr et al. upubliceret.

11
Tidevandszonen er den del af kysten, der skiftevis dækkes og blotlægges af havvand som følge
af tidevandets cyklus (Figur 4). I økotoksikologisk sammenhæng er tidevandszonen interessant, da
denne zone ofte er særligt udsat for antropogen forurening (eksempelvis olieudslip og minedrift).
Desuden giver zonen let adgang til indsamling af marine organismer til økotoksikologiske studier.
Blotlægningen af havbunden i tidevandszonen eksponerer (semi)-sessile organismer for luft. I
Arktis betyder det, at zonens organismer skal kunne klare lufttemperaturer ned til -25°C (Blicher et
al. 2013), hvorfor lufttemperatur er vigtigt for hyppigheden og distributionen af organismer i den
arktiske tidevandszone. Endvidere er andre abiotiske stresspåvirkninger som eksempelvis bølger og
is også af betydning herfor (Aarset 1982, Aunaas et al. 1988). Scrosati og Eckersley (2007)
undersøgte kanten af landisens, den såkaldte ”isfods”, effekt i tidevandszonen i St. Lawrence
Bugten i det østlige Canada, der har lufttemperaturer tilsvarende i Arktis. Studiet viste, at isfoden
har en isolerende effekt, da lufttemperaturer ned til -20°C aldrig gav en lavere temperatur under
isfoden end -7°C. Studiet indikerer, at isfoden yder langsigtet beskyttelse til zonens organismer
imod meget lave frysegrader (Scrosati & Eckersley 2007).
Figur 4 viser en skematisk oversigt over tidevandszonen (”intertidal”) og de zoner, der grænser op her til.

12
Blåmuslingen Mytilus edulis Organismer i tidevandszonen er som
nævnt interessante i økotoksikologiske
studier, da tidevandszonen ofte er særligt
udsat for forurening. I sådanne studier
anvendes blåmuslingen (Mytilus edulis)
ofte, da den er en ideel miljøindikator
(Figur 5), hvilket blandt andet vil blive
uddybet i dette afsnit.
Blåmuslingen M. edulis er en del af et
artskompleks med Mytilus trossulus og
Mytilus galloprovincialis. Arterne er
morfologisk set ens, men de adskiller sig
genetisk (Mcdonald & Koehn 1988, Westfall & Gardner 2010). I Grønland er der kun identificeret
M. edulis (Wanamaker et al. 2007, Versteegh et al. 2012). Når ”blåmuslingen” i denne introduktion
benævnes, refereres der til M. edulis, med mindre andet er angivet.
Blåmuslingens økologi og biogeografi
Blåmuslingen er en bentisk, semisessil filtrator (Bayne et al. 1976, Seed 1976, Gosling 2003).
Arten kan tilpasse sig relativt store fluktuationer i abiotiske forhold (som temperatur, salinitet,
udtørring og ilt) og kan derfor leve i en bred vifte af habitater (Seed & Suchanek 1992).
Eksempelvis findes M. edulis fra tidevandszonen og ned til 40 m, i både brakvand og havvand samt
under både milde og ekstreme temperaturforhold (Seed & Suchanek 1992)(FAO 2014a). Den er
fremherskende på stejle, bølgeudsatte kyster og dominerer både i tidevandszonen og kystzonen
(”subtidal” på figur 4). I den arktiske tidevandszone er lufttemperatur samt lokal fysisk stress fra
skurrende isflager afgørende for blåmuslingens øvre udbredelsesgrænse (Gosling 2003, Blicher et
al. 2013). Arten danner store banker ved hjælp af fasthæftning med byssustråde til substrat og til
hinanden. Blåmuslingen er økologisk set en vigtig art, da den danner fødegrundlag for mange
organismer såsom fugle, fisk og invertebrater. De tætte muslingebanker fungerer endvidere som
habitat for et artsrigt alge- og dyresamfund (Seed 1976, Seed & Suchanek 1992). Blåmuslingen er
desuden af stor kommerciel betydning (FAO 2014b).
Figur 5. Blåmuslinger vokser i tætte banker og filtrerer vandet for fødepartikler med gællerne.

13
Temperatur er afgørende for blåmuslingens biogeografi og artens nordlige grænse afgøres af
minimum lufttemperatur, hvilket eventuelt kan modificeres af en isfod (Seed 1976, Scrosati &
Eckersley 2007, Jones et al. 2010). Blåmuslingen er særligt udbredt på den nordlige halvkugle
(Seed & Suchanek 1992) og den findes i arktiske områder i Norge, Rusland, Canada og Grønland i
både tidevandszonen og kystzonen (Figur 4) (Blicher et al. 2013). I de seneste år er blåmuslingen
endvidere blevet observeret på både Svalbard og i Qaanaaq i det nordvestlige Grønland. Hvorvidt
muslingerne gyder i disse områder eller fragtes hertil i larvestadiet med havstrømme vides endnu
ikke (AMAP 2012, Thyrring, pers. komm.).
Blåmuslingen som miljøindikator
Organismers optag af tungmetaller er oftest proportionelt med mængden af metallet i
omgivelserne (Livingstone & Pipe 1992). En miljøindikator for tungmetal kan defineres som en art,
der akkumulerer tungmetaller i sit væv. Herved fås et mål for biotilgængelighed af tungmetal i det
pågældende miljø, det vil sige hvor stor en andel af den totale tungmetalmængde, der er tilgængelig
for dyr og planter og således økotoksikologisk relevant (Rainbow 1995). Dette mål afhænger af
miljøindikatorens optagelses- og udskillelsesrate af et pågældende metal (Søndergaard et al. 2014).
Miljøindikatorer kan således anvendes til at analysere geografiske og temporale variationer i
biotilgængeligheden af tungmetal i marine økosystemer (Rainbow 1995).
Udover at opfylde kravet om akkumulering af tungmetaller i vævet, så er ideelle miljøindikatorer
sessile, hyppigt forekommende, nemme at identificere, robuste overfor skiftende abiotiske forhold
og de har en relativt lang levetid (Søndergaard et al. 2014). Blåmuslingen lever oftest op til de fleste
af disse kriterier. Som filtrator optager blåmuslingen både tungmetal opløst i vandet og bundet til
partikulært materiale (Rainbow 1995), hvilket gør blåmuslingen særligt attraktiv til risikovurdering
af tungmetaller i akvatiske miljøer, også af bly (Schulz-Baldes 1974, Phillips 1976, Simpson 1979,
Bourgoin 1990, Widdows & Donkin 1992, Mubiana et al. 2005). Blåmuslingen er særdeles velegnet
til risikovurdering af kystnære områder nær minedrift, da organismen er en dominerende art langs
klippekyster (Rainbow 1995), og den har da også hyppigt været anvendt til dette formål i Grønland
(Loring & Asmund 1989b, Riget et al. 1997, Johansen et al. 2008b, Søndergaard et al. 2011,
Zimmer et al. 2011, Søndergaard 2013, Søndergaard et al. 2014).

14
Blåmuslingens kuldetolerance
Blåmuslingen er en af de mest kuldetolerante organismer i den arktiske tidevandszone (Aarset
1982). Kuldestress og -mortalitet er normalt i den arktiske region, og kuldetolerance er således af
afgørende betydning for blåmuslingens fitness og distribution (Aarset 1982, Bourget 1983). Når
blåmuslingen udsættes for kold luft i tidevandszonen lukkes skallen i, hjerteraten og cilieaktiviteten
nedsættes og anaerob metabolisme indtræffer (Aarset 1982). Blåmuslingens kuldetolerance består i,
at den tåler frysning af ekstracellulærvæsken, ligesom mange andre kuldetolerante organismer
(Aarset 1982). Supercooling, en proces hvor en væske holdes flydende under dets frysepunkt, er
ifølge ældre studier ikke observeret hos blåmuslingen (Williams 1970, Aarset 1982), men det er for
nylig observeret, at blåmuslinger først fryser ved -5 til -8°C afhængig af størrelse (Sejr, upub. data).
Så længe cellerne ikke fryser intracellulært, kan de potentielt overleve perioder med lave
frysegrader (Mazur 1977, Odintsova & Boroda 2012). Når ekstracellulærvæsken fryser, mindskes
molfraktionen af frit vand, og intracellulært vand bevæger sig da ud i det ekstracellulære rum ved
osmose, hvilket fører til cellulær dehydrering (Aarset 1982). Cellernes kuldetolerance afgøres af en
maksimal tolerance overfor dehydrering og/eller et minimum tolereret cellevolumen. Williams
(1970) har påvist, at blåmuslingen kan overleve frysning af op til 64% af dens kropsvæske.
Kuldeskade indtræffer således, når en vis mængde af kropsvæsken fryser (Williams 1970, Aarset
1982). En del af forklaringen herpå kan være, at intracellulære proteiner ofte er hydrerede, hvilket
stabiliserer proteinstrukturen. Ved cellulær dehydrering kan det bundne vand ekstraheres ved
osmose, hvormed proteinerne kan denaturere med kronisk celleskade som følge. Dehydrering af
cellen kan desuden medføre membranskader (Karow & Webb 1965, Meryman 1971). Irreversible
kuldeskader ved kuldestress kan resultere i en øget mortalitet hos blåmuslingen, der først indtræffer
flere uger efter stresspåvirkningen (Aarset 1982).
En afgørende del af blåmuslingens kuldetolerance menes at bestå i, at arten kan regulere den
intracellulære koncentration af frie aminosyrer. Ved at øge koncentrationen kan cellen sænke
intracellulærvæskens frysepunkt samt opretholde et vist osmotisk potentiale, der muliggør
regulering af cellevolumen. Cellen kan således undgå en letal grad af dehydrering ved kuldestress
(Williams 1970). I overensstemmelse med denne teori er det observeret, at blåmuslinger
akklimatiseret til høj salinitet har en højere kuldetolerance (Williams 1970). Det kan skyldes, at det
hyperosmotiske saline miljø får cellerne til at dehydrere. Det resulterer i et cellulært respons, der
øger mængden af frie aminosyrer. Herved er cellerne bedre rustet til at modstå efterfølgende

15
kuldestress. Desuden medfører den høje salinitet en mindre isdannelse ekstracellulært og deraf en
lavere grad af cellulær dehydrering (Lange 1963, Williams 1970).
Udover fysiologisk baseret kuldetolerance kan blåmuslingen også beskyttes via artens tendens til
at vokse i banker, under alger og i klippesprækker, hvilket kan beskytte dem imod kulde, is og
bølger (Williams 1970). Det er endvidere muligt, at blåmuslingens kappehule fungerer som et
internt varme-reservoir, når blåmuslingen ved lavvande fryser i den arktiske tidevandszone.
Kappehulen rummer cirka 10 gange så meget vand som vævet. Havvand i kappehulen fryser før
vævet. Isdannelsen er en ektoterm proces, der frigiver latent varme til vævet således, at
vævsfrysning forsinkes. Frysning af kappehulen kan dermed forkorte den periode, hvor vævet er
frosset ved lavvande. Dette bidrager til blåmuslingens kuldetolerance, eftersom blåmuslingens
mortalitet ved kulde i høj grad afgøres af, hvor længe vævet eksponeres for frysning (Williams
1970, Aarset 1982). Store individer er mere kuldetolerante end små individer (Bourget 1983),
sandsynligvis fordi deres kappehule indeholder mere vand og fordi de har et mindre
overflade/volumen forhold, hvilket også giver en langsommere afkøling.
Blåmuslingen adapteres til kulde ved naturlig selektion. Bourget (1983) fandt, at LT50
(temperatur, hvor 50% dør) for blåmuslinger i tidevandszonen i den canadiske St. Lawrence Bugt
var -17,5°C for store muslinger (> 3cm) ved 24 timers stresseksponering. Ved sammenligning med
et tidligere studie, der viste en lavere kuldetolerance hos blåmuslinger fra et mildere klima (-10°C i
Massachusetts, New England, (Williams 1970)), konkluderede Bourget (1983), at der hos
blåmuslingen selekteres for kuldetolerance i arktisk klima.
Det enkelte individ kan endvidere akklimatisere sig til kulde ved at øge sin kuldetolerance over
tid. Gællevæv fra blåmuslingen øger kuldetolerancen efter 2-3 uger ved overførsel fra 15 til 5°C,
hvorimod kuldetolerancen hos kulde-akklimatiserede dyr allerede mindskes igen efter 1 uge ved
overførsel til højere temperatur (Theede 1972).
Blykontaminerede blåmuslinger i Arktis
Blåmuslingen bioakkumulerer tungmetaller opløst i havvandet samt via føden. Artens optag af
tungmetal opløst i havvand er ikke en temperaturafhængig proces (Baines et al. 2006), hvorimod
artens tungmetaloptag fra føden er langt mere effektiv ved lav temperatur (2°C) end ved høj
temperatur (12°C) (Baines et al. 2005). Forskellen i assimileringseffektiviteten var 122-945%
(afhængig af metallet) højere ved lav temperatur, hvilket skyldes blåmuslingens øgede

16
fødeassimilationsrate ved lav temperatur (Baines et al. 2005). Endvidere var udskillelsesraten lavere
ved 2°C (50-80%) end ved 12°C på grund af nedsat respiration (Baines et al. 2005). Disse
temperatureffekter kan have stor indflydelse på organismers bioakkumulering af tungmetal fra føde
og bør inddrages som faktor, når man vurderer udledte tungmetallers toksicitet for flora og fauna i
forskellige miljøer (Baines et al. 2005).
Nogle organismer er aktivt i stand til at udskille tungmetaller fra vævet, men det er muslinger
generelt ikke (Depledge & Rainbow 1990). Fordøjelseskirtlen (hepatopankreas) er hos M.
galloprovincialis vist at optage og binde tungmetaller, hvormed stofferne uskadeliggøres. Organet
spiller derved en vigtig rolle i tungmetalmetabolismen (Viarengo et al. 1981), hvilket sandsynligvis
også gør sig gældende hos M. edulis. Flere transplantationsstudier i Grønland har vist, at
blåmuslingen frigiver bly langsomt efter kontaminering (Riget et al. 1997, Søndergaard et al. 2011,
Zimmer et al. 2011), hvilket kan skyldes langvarig akkumulering i fordøjelseskirtlen. Riget et al.
(1997) viste eksempelvis, at blykontaminerede blåmuslinger fra Maarmorilik overført til et
ukontamineret område i løbet af det første år nedsatte blyindholdet fra 744 til 400 µg/g tørvægt,
hvorefter koncentrationen forblev konstant de næste fire år. Blyindholdet i det enkelte individ kan
således kun mindskes ved fortynding og/eller fysiologisk udskiftning af væv, efterhånden som
muslingen vokser (Zimmer et al. 2011). Dette kan øge de langsigtede økologiske konsekvenser af
tungmetalforurening for blåmuslingen.
Cellemembraner hos ektotermer stresses af kulde og bly Dette afsnit er et teoretisk overblik over cellemembranens fysiske og fysiologiske egenskaber og
hvorledes denne stresses af både tungmetal og kulde, hvilket lægger til grund for studiets hypotese
om, at den mulige synergi imellem effekterne af kulde og bly på blåmuslingens kuldetolerance
skyldes ændringer i membranens fedtsyrekomposition.
Cellemembranen
Alle celler er omgivet af en semipermeabel membran, der virker som barriere til det omgivende
miljø. Membranens funktionalitet er afgørende for cellens (og organismens) overlevelse, da
cellemembranen gør det muligt for cellen at kontrollere sit indre miljø (Singer & Nicolson 1972,
Hazel & Williams 1990). En cellemembran er en dynamisk todimensionel lagdelt struktur
bestående af orienterede lipider med indlejrede transportproteiner, der tillader effektiv transport af
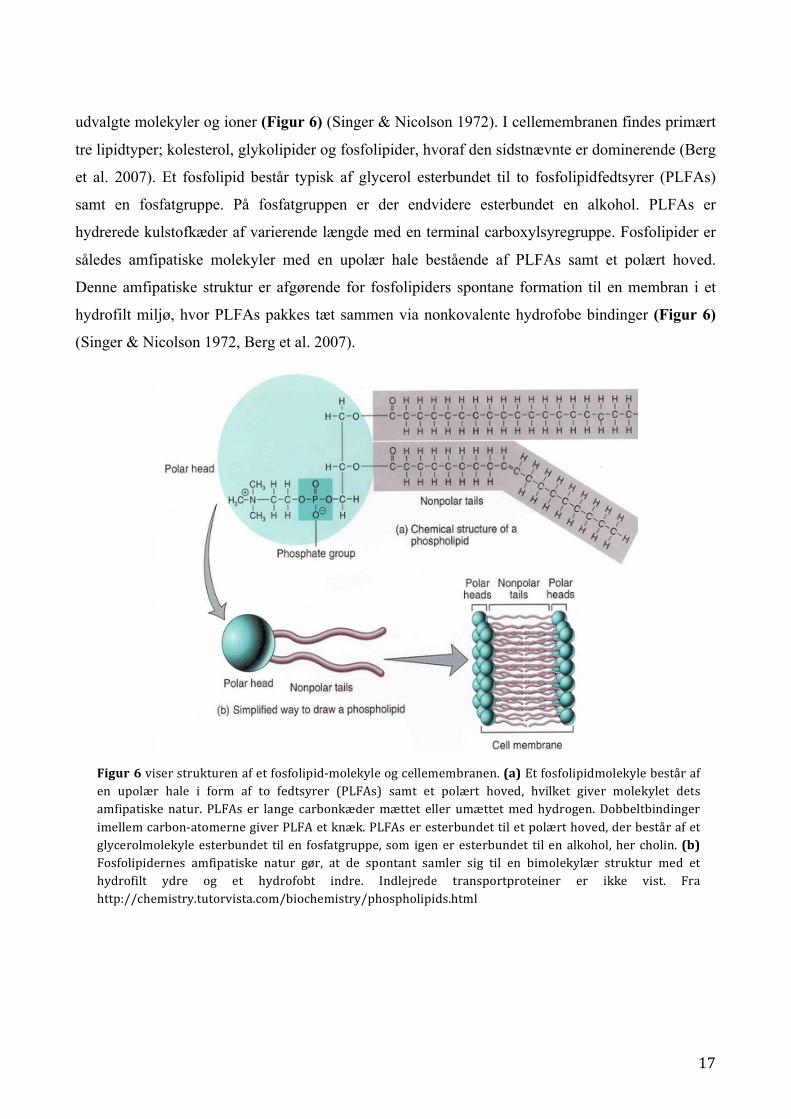
17
udvalgte molekyler og ioner (Figur 6) (Singer & Nicolson 1972). I cellemembranen findes primært
tre lipidtyper; kolesterol, glykolipider og fosfolipider, hvoraf den sidstnævnte er dominerende (Berg
et al. 2007). Et fosfolipid består typisk af glycerol esterbundet til to fosfolipidfedtsyrer (PLFAs)
samt en fosfatgruppe. På fosfatgruppen er der endvidere esterbundet en alkohol. PLFAs er
hydrerede kulstofkæder af varierende længde med en terminal carboxylsyregruppe. Fosfolipider er
således amfipatiske molekyler med en upolær hale bestående af PLFAs samt et polært hoved.
Denne amfipatiske struktur er afgørende for fosfolipiders spontane formation til en membran i et
hydrofilt miljø, hvor PLFAs pakkes tæt sammen via nonkovalente hydrofobe bindinger (Figur 6)
(Singer & Nicolson 1972, Berg et al. 2007).
Figur 6 viser strukturen af et fosfolipid-‐molekyle og cellemembranen. (a) Et fosfolipidmolekyle består af en upolær hale i form af to fedtsyrer (PLFAs) samt et polært hoved, hvilket giver molekylet dets amfipatiske natur. PLFAs er lange carbonkæder mættet eller umættet med hydrogen. Dobbeltbindinger imellem carbon-‐atomerne giver PLFA et knæk. PLFAs er esterbundet til et polært hoved, der består af et glycerolmolekyle esterbundet til en fosfatgruppe, som igen er esterbundet til en alkohol, her cholin. (b) Fosfolipidernes amfipatiske natur gør, at de spontant samler sig til en bimolekylær struktur med et hydrofilt ydre og et hydrofobt indre. Indlejrede transportproteiner er ikke vist. Fra http://chemistry.tutorvista.com/biochemistry/phospholipids.html

18
Cellemembranens temperatursensitivitet
Cellemembranens funktionalitet, herunder membranens fluiditet og fasetilstand, er sensitiv
overfor temperaturændringer (Hazel & Williams 1990). Temperaturen, hvor membranen abrupt går
fra en funktionel flydende-krystallin fase til en dysfunktionel stiv gelfase kaldes
fasetransitionstemperaturen (Tm). Fasetransitionen ændrer i høj grad membranens fysiske
organisering og således dens fysiologiske egenskaber (Hazel & Williams 1990).
Det mest observerede cellulære respons til temperaturændringer hos ektotermer er regulering af
cellemembranen (Hazel & Williams 1990). Dette sker på flere måder. Kolesterol regulerer
fluiditeten i animalske membraner og herved også Tm (Crockett 1998, Berg et al. 2007). Et øget
kolesterolindhold giver rigiditet til membranen og mindsker endvidere risikoen for transition til
gelfase, der ellers kan medføre en række forstyrrelser af membranen (Seelig 1978, Quinn 1985,
Hazel 1995). Sammenligninger af kolesterolindhold i akklimatiserede (og akklimerede) ektotermer
viser et stigende kolesterolindhold ved stigende temperatur (Crockett 1998). Endvidere afhænger
Tm af egenskaber ved PLFAs. PLFAs er en divers gruppe af molekyler med meget forskellige
kemiske og fysiske egenskaber (Hazel & Williams 1990, Berg et al. 2007). Både længde samt antal
og placering af cis carbon-carbon dobbeltbindinger er afgørende for de enkelte PLFA’s smeltepunkt
og således for membranens overordnede Tm (Figur 6). Lange PLFAs interagerer kraftigere med
hinanden end korte, hvorfor lange PLFAs relativt til korte øger Tm. Ligeledes øger mættede PLFAs
Tm, da disse er lige molekyler, der pakkes helt tæt og interagerer effektivt med nabomolekyler.
Modsat sænker umættede PLFAs Tm, særligt flerumættede fedtsyrer (PUFAs), da dobbeltbindinger
giver PLFAs et knæk, hvormed pakningen af PLFAs bliver mere løs og dermed relativt svagere
(Wang et al. 1999, Berg et al. 2007). Således korresponderer højere Tm værdier med højere grad af
orden i pakningen af PLFAs (Separovic & Gawrisch 1996).
Kulde stresser membranen hos ektoterme organismer
Ektoterme organismer har samme kropstemperatur som omgivelserne og har derfor varierende
kropstemperatur over året og døgnet. Omgivelsernes temperatur er således af afgørende fysiologisk
og økologisk betydning for disse organismer (Cairns et al. 1975, Hazel & Williams 1990, Pernet et
al. 2007). I den arktiske tidevandszone indtræffer daglige temperaturfluktuationer som følge af
tidevandets cyklus. Her når ektoterme organismers kropstemperatur ned på lave frysegrader. Kulde
stresser biologiske membraner, som er det primære sted for kuldeskade hos invertebrater (Aarset
1982, Hazel & Williams 1990). Kuldechok, hvor organismer kortvarigt udsættes for lav temperatur,

19
kan enten have umiddelbar død eller kroniske skader på muskler og nervesystem som følge. Det
menes at være resultatet af, at membranen overgår til den dysfunktionelle gelfase, hvor membranen
eksempelvis ikke kan opretholde vitale iongradienter (Quinn 1985, Kelty et al. 1996).
Inden for de genetiske rammer er det muligt for et individ at akklimatisere sig til foranderlige
temperaturforhold (Aarset 1982, Heugens et al. 2001). Mange ektoterme dyr er endvidere adapteret
til temperaturvariationer, således at fysiologiske processer kan forløbe effektivt over et relativt bredt
temperaturspektrum (Crockett 1998). Den mest almindelige tilpasning til temperaturændringer er
regulering af cellemembranen. Dette fænomen kaldes homeoviskøs adaption. Ektoterme dyr
regulerer primært membranen ved at regulere membranens PLFA komposition og dermed
membranens grad af orden (Hazel & Williams 1990, Hazel 1995). Denne membranregulering som
respons på temperaturændringer er observeret hos flere ektoterme tidevandsarter (Hall et al. 2002,
Pernet et al. 2006, Pernet et al. 2007), endda indenfor få timer hos M. californianus samlet øverst i
tidevandszonen (Williams & Somero 1996).
Når ektoterme organismer udsættes for kulde øges graden af umættethed (primært PUFAs) i
membranen for at sænke Tm og dermed opretholde en funktionel fluiditet ved lav temperatur (Hazel
1995). Dette er eksempelvis påvist hos gællevæv fra henholdsvis blåmusling og østers (Crassostrea
virginica). Begge arter viste en signifikant relation imellem den overordnede grad af umættethed i
membranen og akklimatiseringstemperatur (Pernet et al. 2007). En øget grad af umættethed øger
overlevelsen ved kolde temperaturer (Hazel & Williams 1990, Hazel 1995) og særligt et forøget
PUFA indhold ses i de mest kuldetolerante organismer (Hazel 1988). I henhold til dette observerede
Pernet (2007), at blåmuslingen havde en højere grad af umættethed og PUFA i sine cellemembraner
end østers, der er en mindre kuldetolerant art.
Peroxidering af cellemembranen
Som ovenfor beskrevet stresses cellemembranen hos ektoterme dyr af kulde. Alle organismer er
endvidere kontinuerligt udsat for oxidativt stress forårsaget af reaktive oxygenarter (ROS). ROS
skabes ved en ufuldstændig reduktion af ilt (O2) (Livingstone et al. 1990). Som forsvar har alle
organismer et antioxidantsystem, der nedbryder ROS (Sukhotin et al. 2002).
Peroxidering betyder oxidering af lipider og er en skadelig proces, hvor ROS oxiderer og
degraderer membranlipiderne, hvilket resulterer i effekter varierende fra lokale ændringer i
membranfluiditeten til celledød (Marcelo 2004, Valko et al. 2005). Den mest almindelige ROS til

20
peroxidering anses for at være OH- (Nyska & Kohen 2002, Marcelo 2004). Peroxidering initieres
ved, at OH- abstraherer H+ fra en methylengruppe ved siden af en dobbeltbinding i en umættet
PLFA. Herved dannes en fedtsyreradikal (Figur 7.1). PUFAs er meget følsomme overfor
peroxidering, da C=C
dobbeltbindingen gør C-H
bindingen i methylengruppen
svag og derfor mere følsom
overfor oxidering (Nyska &
Kohen 2002, Valko et al. 2005).
Den dannede fedtsyreradikal
reagerer med omgivende O2 og
danner en lipoperoxyl radikal
(Figur 7.2). Denne er ligeledes
meget reaktiv og kan reagere
med et nabo-PUFA molekyle,
hvormed der dannes en
lipidhydroperoxid. Samtidig
omdannes nabo-PUFA til en
fedtsyreradikal, der kan indgå i
samme reaktionskæde (Figur
7.3). Lipidhydroperoxid reagerer
eventuelt med Fe2+, men
undergår under alle
omstændigheder degradering
(Figur 7.4 og 7.5). Peroxidering af én PUFA via ROS igangsætter altså en autokatalytisk proces i
membranen, hvor PUFAs nedbrydes, indtil der ikke er flere eller indtil en antioxidant bremser
kædereaktionen (Nyska & Kohen 2002, Marcelo 2004, Valko et al. 2005).
Bly øger peroxidering og stresser membranen
Det er velkendt, at mange tungmetaller kan inducere produktionen af ROS og øge peroxidering
(Christie & Costa 1984, Viarengo et al. 1990, Valko et al. 2005). Flere studier har vist, at bly øger
peroxidering i både planter og dyr (Gerber et al. 1978, Shafiq Ur 1984, Lawton & Donaldson 1991,
Figur 7 viser peroxidering af flerumættede fedtsyrer (PUFAs) ved eksponering til reaktive oxygenarter. Se tilhørende tekst for detaljer. Efter Marcelo (2004)

21
Stohs & Bagchi 1995, Verma & Dubey 2003). Blykontaminering er generelt forbundet med en
dosisafhængig stigning i peroxidering og mange af blys toksiske effekter synes at være forbundet
med ændringer i membranens PLFAs (Donaldson & Knowles 1993).
Geret et al. (2002) har vist, at flere tungmetaller (Ag, Cd og Hg) signifikant øger peroxidering i
blåmuslingen. I henhold til dette viste studiet endvidere, at tungmetaleksponeringen inducerede
produktionen af proteinet metallothionein (MT) (Geret et al. 2002). MT binder overskydende
metalioner i vævet (sekvestrering) og regulerer således indirekte peroxidering (Geret et al. 2002).
Desuden har MT en direkte antioxidant virkning, da proteinet binder og uskadeliggør OH-
(Thornalley & Vasak 1985). Af disse grunde kan MT anvendes som indikator på biologiske effekter
af tungmetalkontaminering (Schiedek et al. 2009). Blåmuslinger fra Maarmorilik kontamineret med
høje blykoncentrationer (>100 x baggrundsniveau) er ligeledes vist at have et øget MT indhold
(Schiedek et al. 2009).
Viarengo et al. (1991) har vist, at blåmuslingen har et lavere antioxidantniveau om vinteren.
Dette kan skyldes den ektoterme organismes nedsatte metabolisme eller at fysiologisk
antioxidantproduktion kræver energi. Blåmuslingen i Arktis er ofte begrænset af
fødetilgængelighed, særligt i de mørke vintermåneder, hvor produktionen af fytoplankton stort set
går i stå på grund af lysmangel (Sejr, pers. komm.). Den observerede reduktion i
antioxidantniveauet blev tilsvaret af en øget peroxidering af cellemembranerne. Længerevarende
lave vintertemperaturer øger ifølge dette studie således indirekte peroxidering og mindsker dermed
potentielt mængden af PUFA. (Viarengo et al. 1991).
Synergetisk stressinteraktion mellem kulde og tungmetal I 1984 fremsatte Environmental Protection Agency (EPA) risikovurderingsprocedurer til at
vurdere og minimere risikoen af forskellige giftige stoffer i miljøet (EPA 1984). Disse danner
baggrund for undersøgelser af forskellige stoffers toksicitet og letale koncentration ud fra
standardiserede økotoksikologiske laboratorietests, hvor alle andre parametre end
stofkoncentrationen holdes konstante (og typisk favorable) for testorganismen (Løkke et al. 2013).
Det kan dog være problematisk at ekstrapolere resultaterne fra laboratoriet til det naturlige miljø,
hvor organismer sjældent oplever optimale forhold og oftest er udsat for flere stressfaktorer ad
gangen (Holmstrup et al. 2010).

22
Når flere former for stress interagerer og
simultant påvirker en organisme, kan det resultere
i synergetisk stress (figur 8). Det betyder, at den
kombinerede stresseffekt er større end de
adderede effekter af de enkelte stressfaktorer. Der
tages ikke højde for synergetiske interaktioner
med andre stressfaktorer eller for subletale
effekter, hvis man vurderer toksiciteten af et stof i
et som ovenfor beskrevet standard-
laboratorieforsøg (Holmstrup et al. 2010).
Eftersom den omgivende temperatur hos
ektoterme organismer påvirker visse fysiologiske
processer, forventes en synergetisk interaktion
med et toksisk stof, der påvirker de samme
fysiologiske processer (Heugens et al. 2001). I
henhold til dette kan metalkontaminering mindske
organismens tolerance overfor ekstreme
temperaturer og omvendt (Ansari et al. 2004),
eftersom både tungmetal og kulde stresser
cellemembraner. Synergetisk stresspåvirkning af
kulde og tungmetal er ikke et velstuderet område,
men fokus på interaktioner imellem kemisk og
fysisk stress er dog øget i det seneste årti, primært på grund af klimaforandringer (EPA 2003, Noyes
et al. 2009, Hooper et al. 2013, Løkke et al. 2013). Synergi imellem kulde og tungmetal er blevet
undersøgt hos nogle få terrestriske arter, men ikke marine arter. Bindesbøl et al. (2009a) viste
eksempelvis, at både nikkel (Ni) og kviksølv (Hg) interagerede synergetisk med kuldestress (-6°C)
på frysetolerancen hos regnorm (Dendrobaena octaedra). Bly havde derimod ingen interaktion
med kuldestress i dette studie, sandsynligvis på grund af for lave anvendte blykoncentrationer
(ingen mortalitet af bly i kontroller uden fryseeksponering) (Bindesbøl et al. 2009a). Holmstrup et
al. (2008) har som et andet eksempel vist en signifikant synergetisk interaktion imellem kviksølv og
kuldechok på springhaler (Folsomia candida). Studiet viste, at LT50 under kontrolforhold (ingen
Hg) var -6°C. Med stigende subletale kviksølvkoncentrationer øgedes LT50 gradvist til -2°C.
Figur 8 viser et skematisk eksempel på en synergetisk interaktion imellem en kemisk og en fysisk (”natural”) stressfaktor. (A) viser, at en organismes performance falder, når et kritisk stressniveau nås, altså hvor organismen ikke længere kan kompensere for de negative effekter af en naturlig stressfaktor. (B) viser, at ved tilstedeværelsen af en kemisk stressfaktor indtræffer dette kritiske stressniveau ved en lavere grad af fysisk stress. Efter Løkke (2013).

23
Omvendt øgede kuldechok toksiciteten af kviksølv, således at kviksølvkoncentrationer, der under
kontrolforsøg med optimale temperaturforhold ikke havde haft letale effekter, fik det. På baggrund
af deres resultater påpegede Holmstrup et al. (2008) risikoen for at undervurdere tungmetallers
toksicitet med laboratorieforsøg, hvor organismer kun udsættes for én stresspåvirkning ad gangen
(Holmstrup et al. 2008). Også kobber (Cu) er vist at interagere synergetisk med kuldestress på
frysetolerance hos flere arter (Holmstrup et al. 1998, Bossen 2001). Bindesbøl et al. (2005) viste
eksempelvis en signifikant synergetisk interaktion imellem subletale kobberkoncentrationer og
kuldestress hos regnorm (D. octaedra) fra Grønland og konkluderede derfor, at kobberforurening er
af betydning for regnormens biogeografi (Bindesbøl et al. 2005). Et senere studie af samme gruppe
har vist, at kobbers skadelige effekt på regnormens kuldetolerance skyldes tungmetallets toksiske
effekt på cellemembranerne, da gruppen fandt et signifikant fald i PUFAs samt et signifikant øget
peroxideringsniveau ved kobbereksponering (Bindesbøl et al. 2009b). Flere andre studier viser
ligeledes, at den synergetiske interaktion imellem tungmetal og kuldestress på kuldetolerance
(delvist) skyldes membranskader (se Holmstrup 2010 for review).
Aktuelt studie Dette afsnit opsummerer, hvorledes den ovenstående teoretiske introduktion danner baggrund
for det aktuelle videnskabelige studie, som manuskriptet omhandler.
I de seneste årtier er der kommet øget fokus på tungmetalforurening af marine miljøer. Store
mængder af tungmetal er udledt til havet, og dette forventes at fortsætte i fremtiden. Udledte
tungmetaller vækker stor bekymring grundet deres akkumulering i sediment og i marine organismer
samt stoffernes potentielle toksiske (sub-)letale effekter (Ansari et al. 2004). Særligt marine
områder nær minedrift er kraftigt kontaminerede med tungmetaller såsom bly.
Blåmuslinger i den arktiske tidevandszone udsættes for ekstremt lave lufttemperaturer i
vintermånederne (Scrosati & Eckersley 2007, Blicher et al. 2013). I kontaminerede grønlandske
fjordsystemer er de samtidig udsat for høje blykoncentrationer (Johansen et al. 2008, Bach, pers.
komm.). Lave temperaturer udøver stress på cellemembranerne hos ektoterme organismer, da
membranen kræver en vis fluiditet for at være fuldt funktionel. Kommer temperaturen under
fasetransitionstemperaturen Tm går membranen i en dysfunktionel stiv gelfase (Hazel & Williams
1990). Bly stresser cellemembranen ved at øge peroxidering, som kan mindske mængden af

24
PUFAs. Færre PUFAs øger generelt Tm, da membranen bliver mere rigid og derfor kræver en
højere temperatur for at holdes flydende.
Eftersom både kulde og bly stresser biologiske membraner, kan bly potentielt gøre organismer
mere sårbare overfor kulde og omvendt (Heugens et al. 2001). En sådan synergetisk interaktion af
tungmetal og kulde er tidligere påvist hos flere terrestriske organismer, men ikke hos marine arter
(Bindesbøl et al. 2005, Holmstrup et al. 2008, Bindesbøl et al. 2009a, b). Disse synergetiske
stressinteraktioner er generelt vigtige at forstå, før man ekstrapolerer resultater fra laboratoriet til
naturen. En subletal blykoncentration i laboratoriet kan have letale konsekvenser i det naturlige
økosystem, hvor flere stressfaktorer simultant interagerer på organismens fitness (Heugens et al.
2001).
Det er således hypotesen i dette studie, at der er en synergetisk interaktion mellem effekterne af
henholdsvis bly i økologisk realistiske koncentrationer og økologisk realistisk kulde på
blåmuslingens kuldetolerance i Arktis. Dette undersøges i et fuld-faktorielt forsøg, hvor
blåmuslinger indsamlet i tidevandszonen i Nuuk, Grønland (maj 2014), udsættes for 5
blykoncentrationer og 6 temperaturer (figur 9-11). Som mulig forklaring herpå undersøges det,
hvorvidt den potentielle synergetiske interaktion skyldes, at bly skader membranen via ændringer i
PLFA kompositionen grundet øget peroxidering.
Figur 9. Blåmuslinger samlet i Nuuk, Grønland, kontamineres med forskellige blykoncentrationer i en uge.
Figur 10. 100 blåmuslinger tørres enkeltvist inden blyanalyse af vævet.

25
Blåmuslingen anvendes som testorganisme, da den er en
velstuderet og hyppigt anvendt miljøindikator. Der findes
således en baggrundsviden om denne art, også fra arktiske
lokaliteter. Bly undersøges af flere grunde: (1) bly er en
kilde til bekymring i arktiske marine miljøer, eftersom bly
fortsat findes i høje koncentrationer i visse arktiske
fjordsystemer; (2) bly har toksiske effekter ved både letale
og subletale koncentrationer (AMAP 2005, Liu et al. 2011,
Ouyang et al. 2012, Liu et al. 2014) og (3) bly
bioakkumuleres og øger cellulær peroxidering. Desuden
forventer man flere blyminer i Grønland i fremtiden (Rosing
et al. 2014), som kan forårsage yderligere blyforurening. Bly
og kuldes mulige synergetiske interaktion på blåmuslingens
frysetolerance er yderst relevant at undersøge, da et positivt resultat kan indebære, at man ikke kan
vurdere blys toksicitet alene ud fra standardiserede økotoksikologiske laboratorietests. Yderligere
bør man i så fald overveje, om man kan overføre europæiske grænseværdier1 for bly i marine
miljøer til Arktis, da arktiske organismer udsættes for ekstrem kulde. Synergetiske interaktioner
angående kuldetolerance gør sig potentielt gældende for en række andre arter og tungmetaller. Det
er kritisk at forstå disse toksiske stoffers interaktion med andre former for stress (i Arktis særligt
kuldestress) for at kunne vurdere risikoen for det arktiske marine økosystem i forbindelse med de
antropogene tungmetaludledninger, som vi forventer vil fortsætte i fremtiden.
1 Grænseværdien for bly i marine vande er i EU er fastsat til 0,34 μL/L (Miljøministeriet 2010).
Figur 11. PLFA komposition af 100 blåmuslinger analyseres.

26

27
Manuscript
A test of possible synergetic
interactions between sub-‐zero temperatures and lead (Pb) contamination in Greenland blue mussel
(Mytilus edulis).

28
Authors and addresses
Bodil Klein Juhl1, Jakob Thyrring1*, Martin Holmstrup1,2, Mikael K. Sejr1,3
1Department of Bioscience, Aarhus University, Arctic Research Centre, C. F. Møllers Allé 8,
building 1110, DK-8000 Aarhus C, Denmark 2Department of Bioscience, Aarhus University, Soil Fauna Ecology and Ecotoxicology, Vejlsøvej
25, DK-8600 Silkeborg, Denmark 3Greenland Climate Research Centre, Greenland Institute of Natural Resources, Nuuk, Greenland
*Corresponding author: [email protected]; phone: +45 87156697
Keywords
Arctic, Freeze tolerance, Freezing, Mining, membrane lipids; PLFA, Risk assessment

29
Abstract In their natural habitat, organisms are exposed to multiple stressors. Previous studies have
shown, that interactions between chemical and physical stressors can lead to increased mortality.
Heavy metal contamination stresses the cell membrane due to increased peroxidation. Likewise,
sub-zero temperatures also potentially reduce membrane functionality. We conducted a full
factorial experiment with 5 Pb concentrations between 0 and 6.50 mg Pb/L and 6 temperatures from
0 to -17ºC to test the hypothesis that environmentally realistic levels of Pb may interact
synergetically with realistic sub-zero temperatures on the freezing mortality of the blue mussel
(Mytilus edulis) collected in Nuuk, Greenland. We further investigated whether these interactions
are due to Pb-induced alterations of PLFA composition and membrane damage. We found a highly
significant interaction between temperature and mortality, however no effect of lead was seen. Nor
a significant interaction between temperature and Pb on mortality of blue mussels was found, but a
trend was observed at intermediate sub-zero temperatures with increased freezing mortality with
increasing Pb concentrations. At low temperature, ectothermic organisms will regulate the
composition of phospholipid fatty acids (PLFAs) typically by increasing the amount of
polyunsaturated fatty acids (PUFAs) of the cell membrane to sustain a proper fluidity and
functionality. Pb had a significant effect on the molar percentage of PUFAs, which was reduced
from 72.3% (control) to 70.8% in mussels exposed to the highest Pb concentration. Also a
significant increase in monounsaturated fatty acids (MUFAs) was observed, in particular due to an
increase in 18:1. Further, blue mussels exposed to the highest Pb concentrations (4.50 and 6.50 mg
Pb/L) had a significantly higher !"#$!"#$
ratio compared to controls and mussels exposed to the lowest
Pb concentration (0.50 mg Pb/L). The overall unsaturation index was not significantly related to Pb
concentration. Thus, this study revealed significant effects of sub-lethal concentrations of the
hazardous heavy metal Pb on a cellular level in blue mussels. A biochemical background for a
decreased freezing tolerance over time is possibly observed as Pb significantly decreases PUFA
content. This and other studies indicate that synergetic interactions between physical and chemical
stressors should be considered in future risk assessments of heavy metal contamination of natural
habitats.

30
1. Introduction Declining sea ice cover and political and economical interests in the vast potential of oil and
hard mineral reserves in the Greenlandic underground are all drivers for Greenland’s aspirations
concerning the future exploitation of mineral recourses, including iron (Fe), gold (Au), zinc (Zn),
lead (Pb), gas and oil (Nuttall 2014, Rosing et al. 2014).
In Greenland, mining activities have been conducted for over a century with well-documented
long-term environmental impacts, including heavy metal contamination of fjords and organisms
(Søndergaard et al. 2011, Søndergaard et al. 2014). For example, at an abandoned mining site in
South-Greenland, Johansen et al. (2008) found Pb concentrations in blue mussels (Mytilus edulis)
elevated to 200-500 times above background concentration, and Loring & Asmund (1989) reported
Pb tissue concentrations exceeding 800 µg/g (background 3 µg/g) in mussels collected close to the
Maarmorilik mine in West Greenland. Because of previous environmental contamination, recent oil
and mineral exploration in the Arctic region have lead to growing concerns about uncontainable
pollution of the Arctic marine ecosystems (AMAP 2010). Heavy metal waste from increasing
mining activities is an environmental threat due to potential pollution and contamination of the
sensitive marine flora and fauna (Reichelt-Brushett 2012, Søndergaard 2013).
In the Arctic, contaminated organisms simultaneously get exposed to multiple stressors, both
chemical and physical. Chemical contamination may under some conditions reduce the cold
tolerance of invertebrates, leading to increased mortality when low temperature is combined with
contamination (Holmstrup et al. 2008, Bindesbøl et al. 2009a, b). Still, in traditional
ecotoxicological laboratory studies organisms are usually exposed to a single chemical, while held
at otherwise optimal conditions. Risk-assessment and environmental monitoring methods often
consist of point sampling and transplant studies with the focus of estimating uptake/release rates of
contaminants, mortality and metal tissue content (Søndergaard et al. 2011, Zimmer et al. 2011).
However, such studies have limitations, as they do not consider synergetic stress interactions of
multiple stressors acting simultaneously on organisms in natural environments. Both physical and
chemical stressors can interact synergetically and in the Arctic, extreme temperatures, increasingly
acidic oceans (AMAP 2013), additive effects of multiple contaminants, food scarcity during winter
and other relevant stressors are factors worth considering when conducting risk assessment in this
region. Also, such traditional studies often do not include sub-lethal effects. Sub-lethal
concentrations of metals may affect growth, and enzyme activities of plants and animals (Naimo
1995, Ouyang et al. 2012). Consequently, as both chemical and physical induced stress is known to

31
have chronic and (sub)-lethal effects, multi-factorial studies investigating synergetic interactions
between multiple stressors are crucial to incorporate in future risk-assessments of chemicals in the
Arctic (Heugens et al. 2001).
In marine environments a number of species are frequently included when conducting risk-
assessments (Søndergaard et al. 2014). One key indicator organism is the blue mussel (M. edulis)
because it is relatively long-lived, sessile and abundant with potential for concentrating and
assimilating metals directly from seawater and suspended particles (Zimmer et al. 2011,
Søndergaard et al. 2014). In subarctic Greenland, intertidal blue mussels are widely distributed
(Blicher et al. 2013). Here they create unique habitats with a rich associated fauna and they are an
important food source for higher trophic levels such as fish, sea birds and mammals (Gosling 2003).
In the intertidal zone, sub-zero air temperatures are a primary factor influencing blue mussels
distributions-patterns (Blicher et al. 2013). With minimum winter temperatures down to -25ºC
(Blicher et al. 2013), cold stress and freezing mortality is common among intertidal blue mussels in
the Arctic region (Bourget 1983). When acclimatizing to sub-zero temperatures, ectotherms modify
the lipid composition of their cell membrane. The phase behaviour and physical properties of
membrane phospholipid fatty acids (PLFAs) are extremely sensitive to temperature changes (Hazel
& Williams 1990). Fully functional membranes are in a liquid-crystalline phase, but if membranes
are cooled to a threshold temperature [the phase-transition-temperature (Tm)], the membrane enters
a dysfunctional rigid gel phase (Hazel 1995). To lower Tm and thereby ensure the proper fluidity at
low temperatures, ectotherms regulate cholesterol content and the proportion of unsaturated fatty
acids, mainly they up-regulate the content of polyunsaturated fatty acids (PUFAs) (Cossins &
Raynard 1987, Hazel 1995). Such changes in cell membrane PLFA composition are probably
important for freeze tolerance in cold adapted ectothermic animals (Hazel 1995, Kostal et al. 2003).
When blue mussels are exposed to sub-zero temperatures (approximately -7°C depending on body
size (Sejr, unpublished data)), ice forms in extracellular fluids. Since only water molecules form the
ice, the concentration of extracellular solutes increases. This causes cells to dehydrate as
intracellular water moves down the osmotic gradient into the extracellular space. Excessive cell
dehydration is lethal as it disrupts membrane function and protein structures (Meryman 1971). To
increase freezing tolerance, blue mussels change the intracellular free amino acid concentration,
altering the freezing point of intracellular fluids and preventing an intolerable degree of dehydration
(Williams 1970, Aarset 1982).

32
When the hazardous heavy metal lead (Pb) is released into marine environments, it may be
assimilated into marine organisms. Filter-feeding bivalves are exposed to large quantities of water
during feeding and respiration, and in this process they accumulate Pb in both soft tissues and the
shell (Sericano 2000). On a cellular level, Pb contamination has been shown to cause mitochondrial
swelling, deformation and decreased membrane permeability (Goyer & Krall 1969, Raghavan et al.
1981). Heavy metal contamination with various metals has been demonstrated to increase reactive
oxygen species (ROS) production, which accelerates lipid peroxidation, destabilises cell
membranes and alternate PLFA composition (Christie & Costa 1984, Gallego et al. 1996, Valko et
al. 2005). Increased ROS production has also been demonstrated with Pb contamination (Lawton &
Donaldson 1991, Verma & Dubey 2003). ROS affects both monounsaturated (MUFAs) and
polyunsaturated fatty acids, but PUFAs are particularly susceptible to oxidation by ROS (Nyska &
Kohen 2002). PUFAs are vulnerable to peroxidation since double bonds weaken the adjacent
monoallylic C-H bonds and facilitate the reaction of –H with an oxidizing agent, resulting in lipid
radicals and eventually lipid chain termination (Marcelo 2004). Lipid peroxidation will hereby alter
the overall degree of unsaturation from a highly polyunsaturated cell membrane to a more ordered
and rigid membrane structure. This would in theory increase the phase-transition-temperature (Tm)
of the membrane, which leads to a local reduction in membrane fluidity (Hazel & Williams 1990).
If ambient temperature decreases to Tm it can lead to a loss of vital functions and selective
properties (Hazel & Williams 1990). Since both sub-zero temperatures and Pb possibly stress cell
membranes by making it inadequately fluid for full functioning, a synergetic effect of these
stressors can possibly exist.
In the present study we test the hypothesis that synergetic interactions between Pb at ecological
relevant tissue concentrations and realistic sub-zero air temperatures will increase mortality due to
freezing in the arctic blue mussel. Further, to understand the importance of altering cell membrane
fluidity in the freezing tolerance strategy of blue mussels, we test the possibility that Pb
contamination destabilizes the cell membrane by modifying the PLFA composition, and hereby
cause blue mussels to become more vulnerable to the sub-zero temperatures of the arctic intertidal
zone.

33
2. Materials and Methods
2.1 Animals and holding conditions
We used blue mussels (Mytilus edulis) of similar size (~3 cm) collected in the intertidal zone of
Kobbefjord near Nuuk, subarctic Greenland (64°N) in May 2014. Collected animals were
immediately transferred to aerated tanks before transported to holding facilities at Aarhus
University, Denmark. Blue mussels were kept in 40 L aquariums of constant salinity (27 PSU) and
temperature (1ºC). Animals were allowed to acclimate to laboratory conditions for seven days
before experiments were conducted. Mussels were fed with commercially available phytoplankton
mix (Reed Mariculture, Shellfish Diet – 1800 Formula, Campbell, CA, USA). Half the water
volume was daily exchanged with fresh filtrated seawater to minimise build-up of pseudofeces and
excretion products.
2.2 Experimental design
A full factorial experimental design was used to determine survival after exposure to six sub-
zero air temperatures (0, -7, -8, -11, -14, -17ºC) and five Pb concentrations (0, 0.50, 1.80, 4.50 and
6.50 mg Pb/L seawater), giving a total of 30 treatments including control groups.
Pb contaminated seawater was made using a stock solution (10 g Pb/L) made from the analytical
salt lead(II)nitrate [Pb(NO3)2] and deionised water. Mussels were exposed to water concentrations
of 0, 0.50, 1.80, 4.50 and 6.50 mg Pb/L seawater for seven days at 5ºC prior to the freezing
experiment. Water was aerated by air pumps and changed every 24 hours. After seven days of
exposure, 15 mussels were randomly allocated to every experimental combination of Pb and
temperature (a total of 450 mussels). To simulate aerial exposure in the arctic intertidal zone,
mussels from each Pb treatment were transferred to a freezing cabinet (WTB Binder Labortechnik,
Tuttlingen, Germany) programmed to gradually lower the temperature from 5ºC to experimental
temperature over 45 minutes. Mussels were kept at the target temperature (±0.5ºC) for 60 minutes
before a gradual increase of temperature to 5ºC over 45 minutes was initiated. All mussels were
then returned to holding aquariums (5ºC) to recover.
Survival rates were assessed after 1 and 24 hours, respectively, using tactile stimuli and cardio
activity as indicators of survival (using infrared sensors) (Aarset 1982, Burnett et al. 2013).
Mussels were considered alive if they reacted to tactile stimuli and had cardio activity. The mussels

34
were finally frozen and stored at -80ºC until further analysis of tissue Pb content and phospholipid
fatty acid (PLFA) composition.
2.3 Pb analysis
We randomly sampled 20 mussels from each Pb treatment for analysis of Pb tissue content and
PLFA composition. Approximately 10 mg (fresh weight) of gill tissue was dissected out and stored
at -80ºC for PLFA analysis. Remaining tissue was dried for 24 h (60ºC) and acid-digested in 65%
nitric acid at increasing temperatures (80-110ºC) until all fluid had evaporated. Tissue was then re-
dissolved in 6 mL 2% nitric acid and whirly mixed until clear. Pb content was analysed using flame
atomic absorption spectrometry (AAS) (Perkin-Elmer 4100, Ueberlingen, Germany). Certified
reference material (lobster hepatopancreas from National Research Council Canada) was used to
verify the analytical quality, resulting in a measured concentration of 139% (± 7.83% SE, N = 5) of
the certified value.
2.4 PLFA analysis
All gill tissue (see above) was transferred to centrifuge glass tubes and extracted with a mixture
of 1.5 mL chloroform, 1.25 mL buffer (50 mM K2HPO4, pH 7.4) and 3 mL methanol (Waagner et
al. 2013). The chloroform phase containing lipids was transferred to a test tube and evaporated
under nitrogen flow. Lipid extract was dissolved in 300 µL chloroform and fractionated on
prepacked columns with 100 mg of silic acid (Bond Elut Extraction Cartridges, Varian US). PLFAs
were extracted from the column using 1.5 mL methanol and dried under nitrogen flow. Collected
PLFAs were trans esterified by mild alkaline methanolysis (Dowling et al. 1986). Heptane, ELGA
water, and acetic acid (1 M) (1:1:0.15 v/v/v) were added. The biphasic mixture was whirly mixed
and centrifuged for 5 min at 1500 g. The heptane phase containing the fatty acid methyl esters
(FAMEs) was collected. Extraction with heptane was repeated. Combined heptane fractions were
dried under nitrogen flow. Resulting FAMEs were re-dissolved in heptane for gas chromatographic
analysis coupled with mass spectrometry on a Shimadzu GCMS-QP2010 Plus. Identification of
individual FAMEs was based on mass spectra of known FAME standards (Nu-check Prep, Elysian,
MN, USA). Identified fatty acids were designated as X:Y, where X is the number of carbon atoms
and Y is the number of double bonds. Areas of identified peaks were quantified using external
standards, and mol% distributions were calculated (Waagner et al. 2013).

35
2.5 Statistical analysis
We used one-way analysis of variance (ANOVA) to test for differences in the relative amounts
of polyunsaturated fatty acids (PUFAs), monounsaturated fatty acids (MUFAs) and the !"#$!"#$
relationship among Pb treatments. Post-hoc pair-wise (Tukey HSD) tests were used to compare
significant treatment effects (p < 0.05). Finally, generalized linear models (GLM) were used to
model the effects of temperature and Pb and any interactions between the two on survival.
Prior to any analysis, data exploration was carried out following the protocol of Zuur et al. (2010).
Relationships between covariates were assessed using Pearson correlation coefficients (Zuur et al.
2010). Cook’s plot and boxplots were used to identify outliers and to investigate relationships
between variables. PUFA, MUFA and !"#$!"#$
data showed homogeneity and normality of
distribution. For survival we found no indication of zero-inflation or over-dispersion, and we
decided to use GLM with a binomial distribution for the survival model. Once we had identified
valid models we re-examined the residuals to ensure model assumptions were acceptable. Sample
size is denoted n.
3. Results
3.1 Pb content in mussels We found a positive relationship between internal Pb concentration and Pb concentration in
seawater. Mussel tissue Pb content increased with increasing Pb concentration of the seawater,
except at the highest concentration (Fig. 1). The highest tissue concentration was found in mussels
exposed to Pb concentrations of 4.50 mg/L with a mean tissue concentration of 3516 µg Pb/g dry
weight (dw). Control mussels had an average Pb content of 39.9 µg Pb/g dw.

36
Figure 1. Internal lead (Pb) concentrations (µg/g dw) in blue mussels (Mytilus edulis) exposed to five different lead
concentrations (mg Pb/L seawater) for seven days at 5ºC. Error bars indicate standard error. Sample size, n = 100.
3.2 Effects of Pb on PLFAs
Nine different phospholipid fatty acids (PLFAs) were identified (Table 1). The overall
unsaturation index was not significantly related to Pb concentration.
Polyunsaturated fatty acids (PUFAs) were the most abundant fatty acids found in the cellular
membranes of gill tissue (Table 1). Three predominant PUFAs; 20:2, 20:4 and 20:5 each accounted
for at least 14% of the total PLFAs. Pb had a significant effect on the molar percentage of PUFAs,
which was reduced from 72.3% (control) to 70.8% in mussels exposed to 6.50 mg Pb/L (Fig. 2a;
ANOVA, p = 0.0115). Of the individual fatty acids, 18:1 responded significantly to Pb exposure
(data not shown) (ANOVA, p < 0.0001). The monounsaturated fatty acids (MUFAs); 18:1 and 20:1,
accounted for approximately 13% of the total PLFAs. MUFA mol% increased significantly with Pb
concentrations (Fig. 2b; ANOVA, p = 0.0016). Finally, we found a highly significant increase in
the ratio between MUFAs and PUFAs (Fig. 2c; ANOVA, p < 0.0001), indicating Pb contamination
induced a change in the PLFA composition of the cell membrane, with relatively more MUFAs
compared to PUFAs. Tukey HSD tests revealed that mussels exposed to Pb concentrations of 4.50
and 6.50 mg Pb/L had a significantly higher !"#$!"#$
ratio compared to control and mussels exposed to
the lowest Pb concentration (0.50 mg Pb/L) (Fig. 2c; Tukey HSD, all p < 0.05).
0 1 2 3 4 5 6 7
0
500
1000
1500
2000
2500
3000
3500
4000
Water Pb concentration (mg/L)
Inte
rnal
Pb
conc
entr
atio
n (µ
g/g
dw)

37
3.3 Effects of sub-‐zero temperatures and Pb on survival
Temperature had a highly significant effect on the survival of blue mussels (Table 2; p <
0.0001). However, we found no effects of lead (Table 2; p = 0.5322) nor a synergetic interaction
between temperature and Pb on survival of blue mussels was found (Table 2; p = 0.8852). The
survival rate declined significantly from 100% at 0ºC to 0% in mussels exposed to -17ºC, with the
exception of mussels exposed to 1.80 mg Pb/L, where 20% survived (Table 2). Although not
significant, Pb exposed mussels showed a trend of being less tolerant to freezing at intermediate
sub-zero temperatures (-8 and -11ºC) (Table 2).
PLFA 0 0.50 1.80 4.50 6.50
16:0 7.9 ± 0.2 8.1 ± 0.4 7.9 ± 0.2 8.3 ± 0.3 8.4 ± 0.3
18:0 7.4 ± 0.2 6.8 ± 0.2 7.1 ± 0.2 7.3 ± 0.2 7.3 ± 0.2
18:1 2.5 ± 0.1 2.6 ± 0.2 3.4 ± 0.2 3.5 ± 0.2 3.3 ± 0.2
20:1 9.9 ± 0.3 9.8 ± 0.2 9.9 ± 0.2 10.0 ± 0.2 10.1 ± 0.2
20:2 16.5 ± 0.6 15.1 ± 0.7 15.0 ± 0.6 15.3 ± 0.6 14.6 ± 0.6
20:3 3.2 ± 0.1 3.0 ± 0.1 2.8 ± 0.1 3.0 ± 0.1 2.9 ± 0.1
20:4 16.0 ± 0.7 15.5 ± 0.8 15.4 ± 0.8 15.1 ± 0.7 15.4 ± 0.9
20:5 29.6 ± 0.8 32.2 ± 0.8 31.8 ± 0.6 30.7 ± 0.9 31.3 ± 0.8
22:2 7.0 ± 0.3 7.1 ± 0.3 6.7 ± 0.2 6.9 ± 0.4 6.6 ± 0.2
MUFAs 12.4 ± 0.3 12.4 ± 0.2 13.3 ± 0.3 13.5 ± 0.3 13.5 ± 0.2
PUFAs 72.3 ± 0.4 72.8 ± 0.4 71.7 ± 0.5 70.9 ± 0.5 70.8 ± 0.5
MUFA/PUFA 0.2 ± 0.0 0.2 ± 0.0 0.2 ± 0.0 0.2 ± 0.0 0.2 ± 0.0
Table 1. Molar percentage distribution (mean ± standard error) of phospholipid fatty acids (PLFAs)
among five different lead (Pb) contamination levels (mg Pb/L). Total percentage of monounsaturated
fatty acids (MUFAs), polyunsaturated fatty acids (PUFAs) and ratio of MUFAs versus PUFAs are
presented in the bottom of the table. Sample size, n = 87.

38
0 1 2 3 4 5 6 7
70
71
72
73
74
PU
FA (M
olar
pro
cent
age) ab
a
ab
b b
0 1 2 3 4 5 6 7
12.0
12.5
13.0
13.5
14.0
MU
FA (M
olar
pro
cent
age)
aababc
c
bc
0 1 2 3 4 5 6 7
0.16
0.17
0.18
0.19
0.20
Water Pb concentration (mg/L)
MU
FA/P
UFA
a
b
a
b
ab
a)
b)
c)
Figure 2. Molar percentage of phospholipid fatty acids (PLFA) in blue mussels (Mytilus edulis)
exposed to five different lead (Pb) concentrations (mg Pb/L) for seven days at 5ºC. a) The molar
percentage of monounsaturated fatty acids (MUFA). b) The molar percentage of polyunsaturated fatty
acids (PUFA). c) The ratio of MUFA versus PUFA. Different letters indicate a significant difference (p
< 0.05) among lead concentrations. Error bars indicate standard error. Sample size, n = 87.

39
Lead concentration (mg/L)
Temperature 0 0.50 1.80 4.50 6.50
0 100 100 100 100 100
-7 93 87 100 93 93
-8 33 67 47 27 20
-11 20 20 13 7 0
-14 0 7 0 7 7
-17 0 0 20 0 0
4. Discussion Investigating sub-lethal effects and potential interactions among multiple stressors is important
in order to understand and evaluate environmental impacts of heavy metal contamination in natural
environments. This study reveals significant effects of sub-lethal concentrations of the hazardous
heavy metal lead (Pb) on a cellular level in blue mussels (Mytilus edulis), a commonly used
biological indicator species for marine risk-assessment.
4.1 Internal lead
Blue mussels were exposed to multiple Pb concentrations, which resulted in a net increase of the
Pb concentration in whole-body extracts. With the exception of mussels exposed to 6.50 mg Pb/L,
tissue concentration increased correspondingly to water concentration. In general, mussels cannot
actively excrete heavy metals (Depledge & Rainbow 1990), but when exposed to high Pb
concentrations (i.e. 6.50 mg Pb/L) mussels may close valves or reduce filtration rates to reduce
heavy metal uptake (Kramer et al. 1989, Liu et al. 2014). This is consistent with our observation
that mussels of the 6.50 mg Pb/L treatment often had valves closed (data not shown). Internal Pb
content ranged from approximately 40 - 3500 µg/g dw, which is within tissue concentrations found
in blue mussels collected at contaminated fjords near mining operations, where it can exceed 3000
µg/g dw (Ansari et al. 2004, Bach unpublished data).
Table 2. Percentage survival of Mytilus edulis after 7 days of Pb exposure followed by various
sub-zero temperatures. Survival was evaluated after 24 hours. Sample size, n = 450.

40
4.2 Pb effect on PLFA composition and survival
We found that Pb tissue content induced a significant increase in molar percentage of
monounsaturated fatty acids (MUFAs), while the molar percentage of polyunsaturated fatty acids
(PUFAs) overall decreased. This caused a net shift to a relatively higher proportion of MUFAs
compared to PUFAs. This is consistent with the more general observation that contaminants
influence fatty acid molar composition of cell membranes (Valko et al. 2005, Bindesbøl et al.
2009b). However, this is the first study to show that heavy metal exposure alternate phospholipid
fatty acid (PLFA) composition in a marine invertebrate, and only few studies have investigated
heavy metal effects on fatty acids in terrestrial animals. For example, Donaldson & Knowles (1993)
concluded that Pb toxicosis can result from altered fatty acid composition, and Bindesbøl et al.
(2009b) showed how copper can change PLFA composition to a lesser degree of unsaturation and
increase freezing mortality in earthworms (Dendrobaena octaedra). These results correspond with
the observed reduction in PUFA content in blue mussels due to Pb contamination. PUFAs are
particular susceptible to oxidation by ROS, which previously has been shown accelerated by Pb
(Lawton & Donaldson 1991, Nyska & Kohen 2002, Verma & Dubey 2003). That the relative
amount of MUFAs will increase as PUFAs are terminated by lipid peroxidation corresponds with
our present findings. The membrane hereby changes from a highly polyunsaturated cell membrane
to a more ordered and rigid membrane structure. Theoretically, this increases the phase-transition-
temperature (Tm) of the membrane and leads to a local reduction in membrane fluidity (Hazel &
Williams 1990). If ambient temperature decreases to Tm and the gel phase og the membrane is
entered, it can lead to a diminished membrane function (Hazel 1995).
As functional membranes with a high degree of unsaturation (especially PUFAs) are important
for ectothermic animals to survive sub-zero temperatures, we hypothesised a synergetic interaction
between effects of Pb and sub-zero temperatures on freezing tolerance of the blue mussel, since Pb
increases peroxidation. However, we did not find any significant interactions on survival among
such stressors, although a trend was seen at -8ºC and -11ºC with higher mortalities in Pb exposed
groups. A biochemical background for a decreased freezing tolerance over time is possibly
observed as Pb significantly decreases PUFA content. We therefore envision a long-term study to
fully understand if any interactions of temperature and Pb are present in naturally exposed
populations as a supplement to this short-term experiment, where mussels were exposed to Pb for
only seven days. Long-term contamination (e.g. months or years) could result in more
comprehensive changes of the cellular PLFA composition and thereby enhance the observed effects

41
of Pb. In this regard, replicating our study on resident blue mussel from a contaminated site (e.g.
near mining operations) would improve basic understanding of chronic Pb toxicosis and
vulnerability to freezing. Furthermore, this study was conducted in spring acclimated mussels
(collected late May), which were well-fed and in good energetic condition. Viarengo et al. (1991
showed a seasonal variation in antioxidant enzyme systems in blue mussels. The study showed a
lower antioxidant content during winter, presumably due to an altered metabolic state and scarce
food availability. The enzyme systems (e.g. superoxide dismutase, catalase) protect cells against
lipid peroxidation. The reduction in antioxidant system during cold and food scarcity is of special
concern in the Arctic where blue mussels are exposed to low temperatures most of the year and
food scarcity during winter. In the cold Arctic climate, lipid peroxidation could therefore have a
significant effect on survival as antioxidant defence systems are reduced, making oxidative stress
and cellular damage more likely. Thus, it must be well described how the energetic status of an
individual influences its vulnerability to contaminants. We therefore argue that future studies, in
general, should regard seasonal variation in the use of bio-monitoring species (see also Nahrgang et
al. 2013).
4.3 Implications for risk-‐assessments
Besides altered PLFA composition and potentially reduced freezing tolerance, heavy metals like
Pb may impair cellular structures and functions. Heavy metals and oil spills are known to cause
multiple sub-lethal effects, e.g. mitochondria swelling, membrane leakage, gill degradation and
reduced fecundity (Johnson 1977, Donaldson & Knowles 1993, Liu et al. 2014). Considering both
sub-lethal effects of toxicants and possible synergy between multiple stressors is important to
develop thorough ecotoxicological risk assessments of pollutants dispersed from mining and oil
activities and hereby assess the environmental impact of bioavailable elements and, if needed,
address measures to reduce it. Today, most risk assessments are based on studies where only
bioaccumulation, lethal concentrations or single factor experiments are being assessed. The results
of this study and previous work (Bindesbøl et al. 2005, Holmstrup et al. 2008, Bindesbøl et al.
2009a, Bindesbøl et al. 2009b) indicate that assessment of pollutants by such traditional methods
may be inadequate to estimate the impact of pollutant on the resident biota under natural conditions,
where organisms are exposed to multiple stressors. Especially in the Arctic, where future large-
scale mining and oil explorations are expected, it is crucial that future risk measures and
environmental assessments also consider local abiotic (e.g. low temperature, increasingly acidic

42
oceans, ice cover, decreasing salinity) and biotic (food scarcity) stressors and their interactions with
various contaminants in this region.
Acknowledgements The authors thank Lise Lauridsen for technical support and Christian Damgaard for statistical
advice. We also thank Kattegatcentret, Denmark, for providing filtrated seawater. JT was supported
by a grant from ‘Selskabet for Arktisk Forskning og Teknologi (SAFT)’.

43
References • AMAP (1998) AMAP Assessment 1998. Arctic Monitoring and Assessment Programme
(AMAP)
• AMAP (2005) AMAP Assessment 2002: Heavy metals in the Arctic. Arctic Monitoring and
Assessment Programme (AMAP)
• AMAP (2010) AMAP Assessment 2007: Oil and gas activities in the Arctic - effects and
potential effects. Arctic Monitoring and Assessment Programme (AMAP)
• AMAP (2013) AMAP Assessment 2013: Arctic ocean acidification. Arctic Monitoring and
Assessment Programme (AMAP)
• Ansari TM, Marr IL, Tariq N (2004) Heavy metals in marine pollution perspective - a mini
review. Journal of Applied Sciences 4:1-20
• Aunaas T, Denstad JP, Zachariassen KE (1988) Ecophysiological importance of the
isolation response of hibernating blue mussels (Mytilus Edulis). Marine Biology 98:415-419
• Baines SB, Fisher NS, Kinney EL (2005) Influence of temperature on dietary metal uptake
in Arctic and temperate mussels. Marine Ecology Progress Series 289:201-213
• Baines SB, Fisher NS, Kinney EL (2006) Effects of temperature on uptake of aqueous
metals by blue mussels Mytilus edulis from arctic and temperate waters. Marine Ecology
Progress Series 308:117-128
• Bayne BL, Thompson RJ, Widdows J (1976) Physiology: I. In: Bayne BL (ed) Marine
mussels: their ecology and physiology. Cambridge University Press, New York
• Berg JM, Tymoczko JL, Stryer L (2007) Biochemistry. W.H Freeman and Company, New
York, USA
• Bindesbøl AM, Bayley M, Damgaard C, Holmstrup M (2009a) Impacts of heavy metals,
polyaromatic hydrocarbons, and pesticides on freeze tolerance of the earthworm
Dendrobaena octaedra. Environmental Toxicology and Chemistry 28:2341-2347
• Bindesbøl AM, Bayley M, Damgaard C, Hedlund K, Holmstrup M (2009b) Changes in
membrane phospholipids as a mechanistic explanation for decreased freeze tolerance in
earthworms exposed to sublethal copper concentrations. Environmental Science and
Technology 43:5495-5500

44
• Bindesbøl AM, Holmstrup M, Damgaard C, Bayley M (2005) Stress synergy between
environmentally realistic levels of copper and frost in the earthworm Dendrobaena
octaedra. Environmental Toxicology and Chemistry 24:1462-1467
• Blicher ME, Sejr MK, Hogslund S (2013) Population structure of Mytilus edulis in the
intertidal zone in a sub-Arctic fjord, SW Greenland. Marine Ecology Progress Series
487:89-100
• Bourget E (1983) Seasonal variations of cold tolerance in intertidal mollusks and relation to
environmental conditions in the St. Lawrence Estuary. Canadian Journal of Zoology
61:1193-1201
• Bourgoin BP (1990) Mytilus edulis shell as a bioindicator of lead pollution - considerations
on bioavailability and variability. Marine Ecology Progress Series 61:253-262
• Burnett NP, Seabra R, de Pirro M, Wethey DS, Woodin SA, Helmuth B, Zippay ML, Sara
G, Monaco C, Lima FP (2013) An improved noninvasive method for measuring heartbeat of
intertidal animals. Limnology and Oceanography-Methods 11:91-100
• CAFF (2013) Arctic Biodiversity Assessment. Status and trends in Arctic Biodiversity.
Synthesis. Conservation of Arctic Flora and Fauna, Akureyri
• Cairns J, Jr., Heath AG, Parker BC (1975) Temperature influence on chemical toxicity to
aquatic organisms. Journal of the Water Pollution Control Fed 47:267-280
• Christie NT, Costa M (1984) In vitro assessment of the toxicity of metal compounds : IV.
Disposition of metals in cells: interactions with membranes, glutathione, metallothionein,
and DNA. Biological Trace Element Research 6:139-158
• Cossins AR, Raynard RS (1987) Adaptive responses of animal cell membranes to
temperature. In: Bowler K, J. FB (eds) Temperature and Animal Cells. The Company of
Biologists, Limited, Cambridge
• Crockett EL (1998) Cholesterol function in plasma membranes from ectotherms:
membrane-specific roles in adaptation to temperature. American Zoologist 38:291-304
• Depledge MH, Rainbow PS (1990) Models of regulation and accumulation of trace metals
in marine invertebrates. Comparative Biochemistry and Physiology C - Pharmacology
Toxicology & Endocrinology 97:1-7
• Donaldson WE, Knowles SO (1993) Is lead toxicosis a reflection of altered fatty acid
composition of membranes? Comparative Biochemistry and Physiology Part C:
Comparative Pharmacology 104:377-379

45
• Dowling NJE, Widdel F, White DC (1986) Phospholipid ester linked fatty acid biomarkers
of acetate-oxidizing sulfate-reducers and other sulfide-forming bacteria. Journal of General
Microbiology 132:1815-1825
• EPA (1984) Estimating concern levels for concentrations of chemical substances in the
environment. US. Environmental Protection Agency, Washington D. C., USA
• EPA (2003) Framework for cumulative risk assessment. US. Environmental Protection
Agency, Washington D.C., USA
• FAO (2014a) Species fact sheets: Mytilus edulis. Accessed 13.10.14.
• FAO (2014b) Cultured aquatic species information programme: Mytilus edulis. Accessed
13.10.14.
• Gallego SM, Benavides MP, Tomaro ML (1996) Effect of heavy metal ion excess on
sunflower leaves: evidence for involvement of oxidative stress. Plant Science 121:151-159
• Gerber GB, Maes J, Gilliavod N, Casale G (1978) Brain biochemistry of infant mice and
rats exposed to lead. Toxicology Letters 2:51-63
• Geret F, Jouan A, Turpin V, Bebianno MJ, Cosson RP (2002) Influence of metal exposure
on metallothionein synthesis and lipid peroxidation in two bivalve mollusks: the oyster
(Crassostrea gigas) and the mussel (Mytilus edulis). Aquatic Living Resources 15:61-66
• Gosling E (2003) Bivalve molluscs: biology, ecology and culture. Fishing News Books,
Oxford, England
• Goyer RA, Krall R (1969) Ultrastructural transformation in mitochondria isolated from
kidneys of normal and lead-intoxicated rats. Journal of Cell Biology 41:393-400
• Hall JM, Parrish CC, Thompson RJ (2002) Eicosapentaenoic acid regulates scallop
(Placopecten magellanicus) membrane fluidity in response to cold. Biological Bulletin
202:201-203
• Hazel JR (1988) Homeoviscous adaption in animal cell membranes. Advances in
Membrane Fluidity - Physiological Regulation of Membrane Fluidity 6:149-188
• Hazel JR (1995) Thermal adaptation in biological membranes: is homeoviscous adaptation
the explanation? Annual Review of Physiology 57:19-42
• Hazel JR, Williams EE (1990) The role of alterations in membrane lipid-composition in
enabling physiological adaptation of organisms to their physical environment. Progress in
Lipid Research 29:167-227

46
• Heugens EHW, Hendriks AJ, Dekker T, van Straalen NM, Admiraal W (2001) A review of
the effects of multiple stressors on aquatic organisms and analysis of uncertainty factors for
use in risk assessment. Critical Reviews in Toxicology 31:247-284
• Holmstrup M, Aubail A, Damgaard C (2008) Exposure to mercury reduces cold tolerance
in the springtail Folsomia candida. Comparative Biochemistry and Physiology C -
Toxicology & Pharmacology 148:172-177
• Holmstrup M, Bindesbøl AM, Oostingh GJ, Duschl A, Scheil V, Kohler HR, Loureiro S,
Soares AMVM, Ferreira ALG, Kienle C, Gerhardt A, Laskowski R, Kramarz PE, Bayley M,
Svendsen C, Spurgeon DJ (2010) Interactions between effects of environmental chemicals
and natural stressors: a review. Science of the Total Environment 408:3746-3762
• Hooper MJ, Ankley GT, Cristol DA, Maryoung LA, Noyes PD, Pinkerton KE (2013)
Interactions between chemical and climate stressors: a role for mechanistic toxicology in
assessing climate change risks. Environmental Toxicology and Chemistry 32:32-48
• Johansen P, Asmund G, Riget F, Johansen K (2008a) Environmental monitoring at the
cryolite mine in Ivittuut, South Greenland 2007. National Environmental Research Institute,
Aarhus Universitet, Danmark
• Johnson FG (1977) Sublethal biological effects of petroleum hydrocarbon exposures:
bacteria, algae, and invertebrates In: Malins DC (ed) Effects of petroleum on Arctic and
Subarctic marine environments and organisms. Academic press, Inc., New York, USA
• Jones SJ, Lima FP, S. WD (2010) Rising environmental temperatures and biogeography:
poleward range contraction of the blue mussel, Mytilus edulis L., in the western Atlantic.
Journal of Biogeography 37:2243-2259
• Josefson AB, Hansen JLS, Asmund G, Johansen P (2008) Threshold response of benthic
macrofauna integrity to metal contamination in West Greenland. Marine Pollution Bulletin
56:1265-1274
• Karow AM, Webb WR (1965) Tissue freezing: a theory for injury and survival.
Cryomology 2:101-108
• Kelty JD, Killian KA, Lee RE (1996) Cold shock and rapid cold-hardening of pharate adult
flesh flies (Sarcophaga crassipalpis): effects on behaviour and neuromuscular function
following eclosion. Physiological Entomology 21:283-288
• Kostal V, Berkova P, Simek P (2003) Remodelling of membrane phospholipids during
transition to diapause and cold-acclimation in the larvae of Chymomyza costata

47
(Drosophilidae). Comparative Biochemistry and Physiology B - Biochemistry & Molecular
Biology 135:407-419
• Kramer KJM, Jenner HA, Dezwart D (1989) The valve movement response of mussels - a
tool in biological monitoring. Hydrobiologia 188:433-443
• Lange R (1963) Osmotic function of amino acids and taurine in mussel, Mytilus Edulis.
Comparative Biochemistry and Physiology 10:173-179
• Lawton LJ, Donaldson WE (1991) Lead-induced tissue fatty-acid alterations and lipid-
peroxidation. Biological Trace Element Research 28:83-97
• Liu GX, Chai XL, Shao YQ, Hu LH, Xie QL, Wu HX (2011) Toxicity of copper, lead, and
cadmium on the motility of two marine microalgae Isochrysis galbana and Tetraselmis chui.
Journal of Environmental Sciences - China 23:330-335
• Liu GX, Shu MA, Chai XL, Shao YQ, Wu HX, Sun CS, Yang SB (2014) Effect of chronic
sublethal exposure of major heavy metals on filtration rate, sex ratio, and gonad
development of a bivalve species. Bulletin of Environmental Contamination and Toxicology
92:71-74
• Livingstone DR, Martinez PG, Michel X, Narbonne JF, Ohara S, Ribera D, Winston GW
(1990) Oxyradical production as a pollution-mediated mechanism of toxicity in the common
mussel, Mytilus edulis L., and other mollusks. Functional Ecology 4:415-424
• Livingstone DR, Pipe KR (1992) Mussels and environmental contaminants: molecular and
cellular aspects. In: Gosling E (ed) The Mussel Mytilus: ecology, physiology, genetics and
culture. Elsevier, Amsterdam
• Loring DH, Asmund G (1989) Heavy metal contamination of a Greenland fjord system by
mine wastes. Environmental Geology and Water Sciences 14:61-71
• Løkke H, Ragas AMJ, Holmstrup M (2013) Tools and perspectives for assessing chemical
mixtures and multiple stressors. Toxicology 313:73-82
• Marcelo HL (2004) Oxygen in biology and biochemistry: role of free radicals. In: Storey
KB (ed) Functional metabolism: regulation and adaption. John Wiley & Sons, New Jersey,
USA
• Mazur P (1977) The role of intracellular freezing in the death of cells cooled at
supraoptimal rates. Cryobiology 14:251-272
• Mcdonald JH, Koehn RK (1988) The mussels Mytilus galloprovincialis and Mytilus
trossulus on the pacific coast of North-America. Marine Biology 99:111-118

48
• Meryman HT (1971) Osmotic stress as a mechanism of freezing injury. Cryobiology 8:489-
500
• Miljøministeriet (2010) Bekendtgørelse om miljøkvalitetskrav for vandområder og krav til
udledning af forurenende stoffer til vandløb, søer eller havet., Book 1022. Miljøministeriet
• Mubiana VK, Qadah D, Meys J, Blust R (2005) Temporal and spatial trends in heavy metal
concentrations in the marine mussel Mytilus edulis from the Western Scheldt Estuary (The
Netherlands). Hydrobiologia 540:169-180
• Nahrgang J, Brooks SJ, Evenset A, Camus L, Jonsson M, Smith TJ, Lukina J, Frantzen M,
Giarratano E, Renaud PE (2013) Seasonal variation in biomarkers in blue mussel (Mytilus
edulis), Icelandic scallop (Chlamys islandica) and Atlantic cod (Gadus morhua)—
Implications for environmental monitoring in the Barents Sea. Aquatic Toxicology 127:21-
35
• Naimo T (1995) A review of the effects of heavy metals on freshwater mussels.
Ecotoxicology 4:341-362
• Noyes PD, McElwee MK, Miller HD, Clark BW, Van Tiem LA, Walcott KC, Erwin KN,
Levin ED (2009) The toxicology of climate change: environmental contaminants in a
warming world. Environment International 35:971-986
• Nuttall M (2014) Zero-tolerance, uranium and Greenland's mining future. The Polar Journal
3:368-383
• Nyska A, Kohen R (2002) Oxidation of biological systems: oxidative stress phenomena,
antioxidants, redox reactions, and methods for their quantification. Toxicologic Pathology
30:620-650
• Odintsova NA, Boroda AV (2012) Cryopreservation of the cells and larvae of marine
organisms. Russian Journal of Marine Biology 38:101-111
• Ouyang HL, Kong XZ, He W, Qin N, He QS, Wang Y, Wang R, Xu FL (2012) Effects of
five heavy metals at sub-lethal concentrations on the growth and photosynthesis of
Chlorella vulgaris. Chinese Science Bulletin 57:3363-3370
• Pernet F, Tremblay R, Comeau L, Guderley H (2007) Temperature adaptation in two
bivalve species from different thermal habitats: energetics and remodelling of membrane
lipids. Journal of Experimental Biology 210:2999-3014

49
• Pernet F, Tremblay R, Gionet C, Landry T (2006) Lipid remodeling in wild and selectively
bred hard clams at low temperatures in relation to genetic and physiological parameters.
Journal of Experimental Biology 209:4663-4675
• Phillips DJH (1976) The common mussel Mytilus edulis as an indicator of pollution by
zinc, cadmium, lead and copper. Effects of environmental variables on uptake of metals.
Marine Biology 38:59-69
• Quinn PJ (1985) A lipid-phase separation model of low temperature damage to biological
membranes. Cryobiology 22:128-146
• Raghavan SRV, Culver BD, Gonick HC (1981) Erythrocyte lead-binding protein after
occupational exposure. 2. Influence on lead inhibition of membrane Na+-K+-
adenosinetriphosphatase. Journal of Toxicology and environmental Health 7:561-568
• Rainbow PS (1995) Biomonitoring of heavy metal availability in the marine environment.
Marine Pollution Bulletin 31:183-192
• Reichelt-Brushett A (2012) Risk assessment and ecotoxicology limitations and
recommendations for ocean disposal of mine waste in the coral triangle. Oceanography
4:40-51
• Riget F, Johansen P, Asmund G (1997) Uptake and release of lead and zinc by blue
mussels. Experience from transplantation experiments in Greenland. Marine Pollution
Bulletin 34:805-815
• Rosing M, Mosbech A, Hansen AM, Mortensen BOG, Ulfbeck VG, Alfredsson G, Sejersen
F, Helgesen G, Hansen KG, Rasmussen MV, Borch OJ, Rasmussen RO, Nielsen SB (2014)
For the benefit of Greenland - the committee for greenlandic mineral resources to the benefit
of society
• Råstofstyrelsen GS (2013) Mineralefterforskning i Grønland 2013 - beskrivelse af
aktivitetet. Råstofstyrelsen, Grønlands Selvstyre, Nuuk, Grønland
• Schiedek D, Asmund G, Johansen P, Rigét F, Johansen K, Strand J, Mølvig S (2009)
Environmental monitoring at the former lead-zink mine in Maarmorilik, Northwest
Greenland, in 2008. National Environmental Research Institute, University of Aarhus,
Denmark
• Schulz-Baldes M (1974) Lead uptake from seawater and food, and lead loss in the common
mussel Mytilus edulis. Marine Biology 25:177-193

50
• Scrosati R, Eckersley LK (2007) Thermal insulation of the intertidal zone by the ice foot.
Journal of Sea Research 58:331-334
• Seed R (1976) Ecology. In: Bayne BL (ed) Marine mussels: their ecology and physiology.
Cambridge University Press, New York
• Seed R, Suchanek T (1992) Population and community ecology of Mytilus. In: Gosling E
(ed) The mussel Mytilus: ecology, physiology, genetics and culture. Elsevier, Amsterdam
• Seelig J (1978) P-31 nuclear magnetic-resonance and head group structure of phospholipids
in membranes. Biochimica Et Biophysica Acta 515:105-140
• Separovic F, Gawrisch K (1996) Effect of unsaturation on the chain order of
phosphatidylcholines in a dioleoylphosphatidylethanolamine matrix. Biophysical Journal
71:274-282
• Sericano JL (2000) The Mussel Watch approach and its applicability to global chemical
contamination monitoring programmes. International Journal of Environment and Pollution
13:340-350
• Shafiq Ur R (1984) Lead-induced regional lipid peroxidation in brain. Toxicology Letters
21:333-337
• Simpson RD (1979) Uptake and loss of zinc and lead by mussels (Mytilus edulis) and
relationships with body-weight and reproductive cycle. Marine Pollution Bulletin 10:74-78
• Singer SJ, Nicolson GL (1972) The fluid mosaic model of the structure of cell membranes.
Science 175:720-731
• Stohs SJ, Bagchi D (1995) Oxidative mechanisms in the toxicity of metal ions. Free Radical
Biology and Medicine 18:321-336
• Sukhotin AA, Abele D, Portner HO (2002) Growth, metabolism and lipid peroxidation in
Mytilus edulis: age and size effects. Marine Ecology Progress Series 226:223-234
• Søndergaard J (2013) Dispersion and bioaccumulation of elements from an open-pit
olivine mine in Southwest Greenland assessed using lichens, seaweeds, mussels and fish.
Environmental Monitoring and Assessment 185:7025-7035
• Søndergaard J, Asmund G, Johansen P, Riget F (2011) Long-term response of an arctic
fiord system to lead-zinc mining and submarine disposal of mine waste (Maarmorilik, West
Greenland). Marine Environmental Research 71:331-341
• Søndergaard J, Bach L, Gustavson K (2014) Measuring bioavailable metals using diffusive
gradients in thin films (DGT) and transplanted seaweed (Fucus vesiculosus), blue mussels

51
(Mytilus edulis) and sea snails (Littorina saxatilis) suspended from monitoring buoys near a
former lead-zinc mine in West Greenland. Marine Pollution Bulletin 78:102-109
• Theede H (1972) Vergleichende ökologisch-physiologische Untersuchungen zur zellulären
Kälteresistenz mariner Evertebraten. Marine Biology 15:160-191
• Thornalley PJ, Vasak M (1985) Possible role for metallothionein in protection against
radiation-induced oxidative stress - kinetics and mechanism of its reaction with superoxide
and hydroxyl radicals. Biochimica Et Biophysica Acta 827:36-44
• Torres MA, Barros MP, Campos SCG, Pinto E, Rajamani S, Sayre RT, Colepicolo P (2008)
Biochemical biomarkers in algae and marine pollution: a review. Ecotoxicology and
Environmental Safety 71:1-15
• Valko M, Morris H, Cronin MTD (2005) Metals, toxicity and oxidative stress. Current
Medicinal Chemistry 12:1161-1208
• Verma S, Dubey RS (2003) Lead toxicity induces lipid peroxidation and alters the activities
of antioxidant enzymes in growing rice plants. Plant Science 164:645-655
• Versteegh EAA, Blicher ME, Mortensen J, Rysgaard S, Als TD, Wanamaker AD (2012)
Oxygen isotope ratios in the shell of Mytilus edulis: archives of glacier meltwater in
Greenland? Biogeosciences 9:5231-5241
• Viarengo A, Canesi L, Pertica M, Livingstone DR (1991) Seasonal-variations in the
antioxidant defense systems and lipid-peroxidation of the digestive gland of mussels.
Comparative Biochemistry and Physiology C - Pharmacology Toxicology & Endocrinology
100:187-190
• Viarengo A, Canesi L, Pertica M, Poli G, Moore MN, Orunesu M (1990) Heavy metal
effects on lipid-peroxidation in the tissues of Mytilus galloprovincialis lam. Comparative
Biochemistry and Physiology C - Pharmacology Toxicology & Endocrinology 97:37-42
• Viarengo A, Zanicchi G, Moore MN, Orunesu M (1981) Accumulation and detoxication of
copper by the mussel Mytilus Galloprovincialis lam - a study of the sub-cellular distribution
in the digestive gland-cells. Aquatic Toxicology 1:147-157
• Wanamaker AD, Kreutz KJ, Borns HW, Introne DS, Feindel S, Funder S, Rawson PD,
Barber BJ (2007) Experimental determination of salinity, temperature, growth, and
metabolic effects on shell isotope chemistry of Mytilus edulis collected from Maine and
Greenland. Paleoceanography 22

52
• Wang GQ, Li SS, Lin HN, Brumbaugh EE, Huang CH (1999) Effects of various numbers
and positions of cis double bonds in the sn-2 acyl chain of phosphatidylethanolamine on the
chain-melting temperature. Journal of Biological Chemistry 274:12289-12299
• Westfall KM, Gardner JPA (2010) Genetic diversity of southern hemisphere blue mussels
(Bivalvia: Mytilidae) and the identification of non-indigenous taxa. Biological Journal of the
Linnean Society 101:898-909
• Widdows J, Donkin P (1992) Mussels and environmental contaminents: bioaccumulation
and physiological apects. In: Gosling E (ed) The mussel Mytilus: ecology, physiology,
genetics and culture. Elsevier, Amsterdam
• Williams EE, Somero GN (1996) Seasonal-, tidal-cycle- and microhabitat-related variation
in membrane order of phospholipid vesicles from gills of the intertidal mussel Mytilus
californianus. Journal of Experimental Biology 199:1587-1596
• Williams RJ (1970) Freezing tolerance in Mytilus edulis. Comparative Biochemistry and
Physiology 35:145-161
• Waagner D, Holmstrup M, Bayley M, Sorensen JG (2013) Induced cold-tolerance
mechanisms depend on duration of acclimation in the chill-sensitive Folsomia candida
(Collembola). Journal of Experimental Biology 216:1991-2000
• Zimmer LA, Asmund G, Johansen P, Mortensen J, Hansen BW (2011) Pollution from
mining in South Greenland: uptake and release of Pb by blue mussels (Mytilus edulis L.)
documented by transplantation experiments. Polar Biology 34:431-439
• Zuur AF, Ieno EN, Elphick CS (2010) A protocol for data exploration to avoid common
statistical problems. Methods in Ecology and Evolution 1:3-14
• Aarset AV (1982) Freezing tolerance in intertidal invertebrates - a review. Comparative
Biochemistry and Physiology A - Physiology 73:571-580