what does the retrosplenial cortex do?cvcl.mit.edu/sunseminar/vannaggletonmaguirre_2009.pdf · lp,...
TRANSCRIPT

It is 100 years since Brodmann delineated the human RSC (areas 29 and 30) within the posterior cingulate region1. Regrettably, so few advances in our under-standing of the human RSC seemed to have occurred over the next 90 years that Vogt et al. stated ‘nothing is known about its function’2. Early views of RSC function focused on its potential role in brain systems that regu-late emotion. The most famous example of these is the Papez circuit3, in which projections from the anterior thalamic nuclei to the cingulate cortex and from there to other cortical regions emotionally colour our psychic processes. MacLean’s concept of the ‘visceral brain’4 endorsed Papez’ ideas, citing evidence that stimulation of the cingulate cortices can evoke autonomic changes that are linked with emotion. This past focus on emotion and autonomic regulation may, however, have deflected attention away from the potential importance of this region for other cognitive functions.
This Review first considers RSC neuroanatomy, then focuses on evidence that the RSC contributes to memory and navigation, and finally considers how these roles fit with its more recently revealed involvement in other aspects of cognition.
Location, terminology and appearanceFor RSC research, location is everything. The term ‘retro-splenial’ defines the RSC’s position immediately behind the splenium, the most caudal part of the corpus cal-losum. Consequently, the primate RSC is located deep in the midline, much of it hidden from the medial surface of the brain. The RSC forms part of the posterior cingu-late region, the other components of the primate posterior cingulate region being areas 23 and 31 (FIG. 1). Area 31 is
sometimes considered to belong to both the posterior cin-gulate and the precuneate cortices5,6, but it is agreed that area 23 separates the RSC from the precuneate region.
The importance of distinguishing the RSC from other parts of the posterior cingulate region is underlined by the many morphological and connectivity differences that areas 29 and 30 have from area 23 (ReFs 7,8). The RSC is regarded as an ‘intermediate’ cortex as it has a transitional pattern of lamination that reaches a clearly defined pattern of six layers only in the adjacent area 23. Consequently, progressing in a lateral to medial direc-tion around the callosal sulcus above the splenium in the primate brain (both human and monkey), lamination increases from the induseum griseum to Brodmann’s area 26 (not present in the monkey), to lateral area 29 (four layers) to medial area 29 (four or five layers, depending on the authority) to the dysgranular area 30 (six layers but a poorly defined, dysgranular layer IV) and finally to area 23 (six distinct layers)8–11. When refer-ring to the monkey brain in this article, we use the RSC boundaries and terminology of Kobayashi and Amaral8, as their nomenclature accompanies the most detailed analyses of primate RSC connectivity to date (FIG. 1).
The rat posterior cingulate region differs substantially from the primate RSC as it has no direct counterparts to areas 23 and 31, so the entire region is designated the RSC12,13. The term posterior cingulate for the rodent brain is, therefore, potentially misleading. As in the primate, the rat RSC can be separated into dysgranular (area 30) and granular (area 29) regions, and the latter area has been divided into either two13 or three12 sub-regions, depending on the authority. In this Review we refer to the regions and subregions as described by Van
*School of Psychology, Cardiff University, Tower Building, Park Place, Cardiff, CF10 3AT, UK. ‡Wellcome Trust Centre for Neuroimaging, Institute of Neurology, University College London, London, WC1N 3BG, UK.Correspondence to S.D.V.e-mail: [email protected]:10.1038/nrn2733Published online 8 October 2009
What does the retrosplenial cortex do?Seralynne D. Vann*, John P. Aggleton* and Eleanor A. Maguire‡
Abstract | The past decade has seen a transformation in research on the retrosplenial cortex (RSC). This cortical area has emerged as a key member of a core network of brain regions that underpins a range of cognitive functions, including episodic memory, navigation, imagination and planning for the future. It is now also evident that the RSC is consistently compromised in the most common neurological disorders that impair memory. Here we review advances on multiple fronts, most notably in neuroanatomy, animal studies and neuroimaging, that have highlighted the importance of the RSC for cognition, and consider why specifying its precise functions remains problematic.
R E V I E W S
792 | noVEMBER 2009 | VoLuME 10 www.nature.com/reviews/neuro
© 2009 Macmillan Publishers Limited. All rights reserved

e
Nature Reviews | Neuroscience
a
d
b c3, 1, 2
752423
31
2930
Corpuscallosum
Dorsolateralprefrontal cortex
Parietal & occipitalcortex
Anterior thalamicnuclei
Hippocampus
RSC12
11
107a
8, 6
STSd
2423
29 & 30
Clau
PR
HF PH V4Occipital
ATNCaudLP, LD, MPulv
24
818b
172930
b
b
d
d
aAM
ADAV
LPLD
PostSubPreSParaS
A8.6 A5.6 A2.6 A1.6 A0.6
232930 23
46, 9
Groen and Wyss13–15 — that is, dysgranular (Rdg), granu-lar a (Rga) and granular b (Rgb) regions — because some connectivity studies and all studies of selective lesions in areas 29 and 30 have used this terminology. A striking feature of the rat RSC is its size — it extends over half the length of the entire cerebrum, making it one of the largest cortical regions in this species.
Anatomical connectionsIf little else was known about RSC function, its con-nections would immediately point to a role in memory. Axonal tracing studies in monkeys (FIG. 1) have revealed reciprocal connections with three notable regions: the hippocampal formation (subiculum, presubiculum
and parasubiculum), the parahippocampal region (the entorhinal cortex and areas TH and TF) and select tha-lamic nuclei (the anterior and lateral dorsal nuclei). Pathology in all three regions has been repeatedly implicated in amnesic syndromes. Furthermore, the densities of the hippocampal and anterior thalamic con-nections with areas 29 and 30 are much higher than the densities of these connections to the adjacent posterior cingulate area 23 (ReFs 7,16,17), suggesting that within the posterior cingulate region the RSC is most likely to be involved in hippocampus-dependent functions. other notable connections include reciprocal retro-splenial pathways to parts of the prefrontal cortex (areas 46, 9, 10 and 11), which provide an indirect route for
Figure 1 | Retrosplenial neuroanatomy. Location and principal connections of the retrosplenial cortex (RSC). a | Medial surface of a human brain showing the position of the RSC (Brodmann’s areas 29 and 30) (blue) and the remaining posterior cingulate cortex (areas 23 and 31) (green). b | How the RSC can help to integrate information in cortical sensory and prefrontal sites (upper half of the schematic) with subcortical sites required for memory (lower half). c | Principal connections of the macaque (Macaca fascicularis) RSC. The darker arrows indicate denser projections (bicoloured arrows indicate denser projections in the direction indicated by the darker colour)7,16. d | Location and principal connections of the rat RSC; d indicates the dysgranular cortex (area 30); a and b respectively indicate divisions Rga and Rgb within the granular cortex (area 29)12–14. The arrowheads in area 29 do not distinguish between Rga and Rgb. e | A series of coronal sections depicting the location of areas 29 and 30 (the RSC, blue) and area 23 (green) in the posterior cingulate region of M. fascicularis8. Numbers prefixed by ‘A’ indicate the distance (in millimetres) anterior to the auditory meatus8. All other numbers refer to area designations. AD, anterior dorsal thalamic nucleus; AM, anterior medial thalamic nucleus; AV, anterior ventral thalamic nucleus; Caud, caudate nucleus; Clau, claustrum; HF, hippocampal formation; LD, laterodorsal thalamic nucleus; LP, lateroposterior thalamic nucleus; MPulv, medial pulvinar; ParaS, parasubiculum; PH, parahippocampal cortex; Post, postsubiculum; PR, perirhinal cortex; PreS, presubiculum; STSd, dorsal superior temporal sulcus; Sub, subiculum; V4, visual area 4. Part a is modified, with permission, from ReF. 142 © (2001) Elsevier. Part c is modified, with permission, from ReF. 16 © (2007) Wiley. Part d is modified, with permission, from ReF. 12 © (1981) Wiley. Part e is modified, with permission, from ReF. 8 © (2000) Wiley.
R E V I E W S
nATuRE REVIEWS | NeuRoscieNce VoLuME 10 | noVEMBER 2009 | 793
© 2009 Macmillan Publishers Limited. All rights reserved

Head-direction cellsCells that can distinguish, by altering their firing rate, the direction that an animal is facing.
Executive functionThe planning and sequencing of future actions, selected from multiple options.
Immediate-early genesGenes that encode transcription factors that are induced within minutes of raised neuronal activity without requiring a protein signal. Immediate-early gene activation is, therefore, used as an indirect marker of neuronal activation.
Working memoryIn animal research this term refers to information that is required within a trial but may well prove to be misleading across trials or sessions.
Ideothetic informationInternal feedback information that can be used to monitor progress and, hence, position in space.
Path integrationThe continuous integration of self-movement (ideothetic) cues to update representations of current position (and so plot a direct path back to the start of a journey). Also known as ‘dead reckoning’.
hippocampal influences on the dorsolateral prefrontal cortex (areas 46 and 9) and vice versa18. There are also reciprocal links with the dorsal superior temporal sulcus, area V4 and cingulate areas 23 and 24. The dense intra-cingulate connections suggest that interactions between different parts of the cingulate cortex might form addi-tional sets of segregated networks7,19. The pathways for these cortico-cortical and cortico-thalamic connections contribute to the cingulum bundle17.
The rat RSC is dominated by its reciprocal connec-tions with the anterior thalamic nuclei, the lateral dorsal thalamic nucleus and the hippocampal formation13–15. The hippocampal inputs originate in the postsubicu-lum, the subiculum and the presubiculum. The granular regions (area 29) in the RSC (Rga and Rgb) are distin-guished by reciprocal connections with sites (the lateral dorsal and anterior dorsal thalamic nuclei and the post-subiculum) that contain ‘head-direction’ cells, whereas the dysgranular cortex (area 30) in the RSC (Rdg) is more interconnected with visual areas through recipro-cal connections with areas 18b and 17 (ReF. 20). As in the primate, there are reciprocal connections with the anterior cingulate cortex15,19. Subcortical inputs to the rat RSC include projections from the raphe nuclei, the locus coeruleus and the diagonal band21.
It is striking that in both the rodent and the mon-key brain the RSC provides reciprocal, indirect routes between the hippocampal formation and the anterior thalamic nuclei, even though these same regions are also linked by dense direct connections (FIG. 1b). The functional importance of these indirect routes is high-lighted by disconnection studies (see next section) showing that the anterior thalamic nuclei, the RSC and the hippocampus require each other to support spatial learning22,23. These considerations raise the important issue of identifying those RSC connections that are not shared by either the hippocampal formation or the ante-rior thalamic nuclei, so as to obtain clues to the unique mnemonic contributions of the RSC.
Candidate RSC connections not shared with either the hippocampus or the anterior thalamus include recip-rocal connections with areas 46 and 9 in the dorso lateral prefrontal cortex18, which could provide important routes between the hippocampus and cortical regions involved in executive function. In addition, the principal extrinsic cholinergic inputs to the RSC, the hippocam-pus and the anterior thalamic nuclei originate from dif-ferent sources (the basal nucleus of Meynert, the medial septum and vertical limb of the diagonal band, and the laterodorsal tegmental nucleus, respectively). Perhaps most importantly, the RSC receives more early-processed sensory information than either the hippocampus or the anterior thalamic nuclei. The direct inputs from V2 and V4, along with afferents from the claustrum and the lateroposterior thalamic nuclei, could provide visual information21, but additional sensory information might be conveyed through direct parietal cortex (area 7) projections to the RSC. In addition, the parietal cortex (area 7a, area 7b, area LIP (lateral intraparietal cortex) and area DP (dorsal prelunate cortex)) has particularly dense projections to posterior cingulate area 23 (ReF. 7),
which in turn projects heavily to the RSC. Thus, areas 29 and 30 could be vital conduits for sensory (parietal and occipital) information reaching both the hippocampal formation and the anterior thalamic nuclei.
Animal studies of RSC functionThe inaccessibility of the RSC in the human brain has pro-moted research in other animals, which has predominantly used lesion techniques and electrophysiology.
RSC lesions — method and extent. Prompted by the connectivity of the RSC with the hippocampal forma-tion, most RSC lesion studies have focused on cognitive abilities attributed to hippocampal activity. Despite some initial controversy, there is now a general consensus that RSC lesions in rodents impair spatial memory24,25. Many of the earlier disparities can be traced to differ-ences in lesion extent and surgical method26: although the RSC reaches the caudal limit of the rat cerebrum, many lesion studies targeted more rostral parts of the RSC. The importance of the spared caudal retrosplenial tissue for spatial memory has been demonstrated using immediate-early gene mapping27 and electrophysiology28 and by directly comparing the behavioural impact of dif-ferent size RSC lesions29. Lesion size also interacts with lesioning method, as traditional surgical techniques (electrolytic and aspiration) are prone to damaging the underlying cingulum bundle and so potentially discon-necting much of the entire cingulate cortex. A further factor is that the extent of pre-surgical and post-surgical training can alter the impact of RSC lesions30.
Different types of RSC lesion-induced impairments. Most rodent lesion studies have focused on spatial memory and navigation. RSC lesions impair performance on standard spatial memory tasks, including learning the fixed22,31–34 or daily-changing22,32,33,35 location of a platform in a water maze, and performance on working memory tasks in a radial-arm maze31,36. These tasks involve the use of distal visual cues (‘allocentric’ tasks), but lesion-induced impair-ments have also been reported on tasks designed to tax the use of directional information37 (requiring rats to alternate their position around a fixed direction) or the use of ideo-thetic information for path integration34. Inactivation studies (using tetracaine injection) have shown that the RSC is necessary for correct performance in a radial-arm maze task under normal light conditions and for radial-arm maze performance and path integration in the dark38–40. In rats, although the RSC is not required for the detection of novel objects31, it seems to be important for the detec-tion of novel spatial arrangements of objects (‘object-in-place’ detection)31. This wide array of RSC lesion-induced spatial deficits on tasks that rely on different spatial strate-gies (involving both visual and somatosensory feedback) is consistent with the multiple sources of afferent sensory information to the RSC (FIG. 1).
The RSC supports spatial memory in conjunction with at least two other sites, the hippocampus and the anterior thalamic nuclei (FIG. 1). This conclusion is bolstered by findings from spatial working memory tasks in which rats use familiar spatial cues27 or novel
R E V I E W S
794 | noVEMBER 2009 | VoLuME 10 www.nature.com/reviews/neuro
© 2009 Macmillan Publishers Limited. All rights reserved

Nature Reviews | Neuroscience
RSC
ACC
Control MTT lesion
100 µm
Visuospatial conditional tasksTasks in which animals are required to learn fixed associations between visual stimuli or spatial contexts and either spatial locations or objects to gain a reward.
Compound-feature nega-tive-discrimination taskA task in which animals are given two types of trial: one in which a stimulus (for example, a tone) is followed by a reward and a second in which the same stimulus is presented with another stimulus (for example, a light) but no reward. Control animals learn to respond to the tone only but not the tone and light combinations.
Contextual fear conditioningWhen animals learn to associate a specific context or environment with the administration of an aversive stimulus, for example a shock. When re-exposed to the same context or environment control animals will demonstrate a fear response, for example freezing.
Active avoidance in a two-way shuttle boxWhen animals avoid a shock that they have learnt — through prior conditioning — is associated with a stimulus by entering the compartment in which they know they will not be shocked.
configurations of spatial cues41 and show immediate-early gene activation in all three brain structures. The relationships between these three regions were tested more formally in a disconnection study in which unilat-eral lesions were made in the RSC and in either the con-tralateral hippocampus or the anterior thalamic nuclei. Both lesion combinations impaired learning the posi-tion of a fixed platform in a water maze task22 (BOX 1). By contrast, learning in visuospatial conditional tasks is spared in rats with lesions of the RSC36,42. Interestingly, performance on this class of task is sensitive to lesions of the hippocampus and the anterior thalamic nuclei36 but not of other structures known to be necessary for spatial memory — the mammillary bodies43 and fornix44. It is possible that the extensive training required for animals to acquire these conditional discriminations36,44 masks the contribution of some of these brain structures.
The magnitude of the spatial deficits after lesioning of the RSC is typically smaller than the magnitude of the deficits associated with either anterior thalamic or hippocampal damage, which presumably reflects some compensation across structures mediated by their par-allel interconnections (FIG. 1b). The full impact of RSC lesions often emerges only when animals are forced to shift modes of spatial learning — for example, from light to dark38 or from local to distal cues29,37 — indicat-ing that rats with lesions of the RSC cannot switch or integrate parallel modes of task performance. A strik-ing, robust example is when radial-arm maze training
changes so that intra-maze cues are placed in conflict with extra-maze cues29,35,37.
RSC function in rodents is not limited to spatial memory. Recent findings highlight the importance of the RSC for the simultaneous processing of multiple stimuli, with RSC lesions disrupting performance on a compound-feature negative-discrimination task45, contextual fear conditioning46 and the acquisition of active avoidance in a two-way shuttle box47. In addition, RSC lesions severely impair the ability of rabbits to learn how to avoid an elec-tric shock when they hear an acoustic signal48. Although these impairments do not seem to reflect general impairments in the processing of fear-evoking stimuli47, they reveal that RSC function might extend to include learning that is associated with affective stimuli.
Selective subregion lesions. Based on the connectivity of RSC subregions, and recent immediate-early gene studies49, it might be supposed that the dysgranular RSC is more important for visually guided spatial memory and navigation, whereas the granular RSC has a greater involvement in internally directed naviga-tion. To date only two studies in rodents have targeted the individual RSC subregions. one study found that lesions of the Rgb, but not the Rga, disrupted delayed matching to position in a water maze50. The other study51 found that selective Rdg lesions impaired the use of visual allocentric cues in the radial-arm maze, consistent with the predominance of visual inputs to
Box 1 | The retrosplenial cortex as a site of covert pathology?
The retrosplenial cortex (RSC) is unusually sensitive to deafferentation. As noted in BOX 2, RSC hypometabolism is found in patients with Alzheimer’s disease and mild cognitive impairment — conditions that are associated with atrophy in sites that project to the RSC, for example the hippocampal formation and the anterior thalamic nuclei. Studies in rats using immediate-early gene activity as a marker of neuronal activity found that lesions in the anterior thalamic nuclei and hippocampus both produce marked RSC dysfunction119,120. The loss of immediate-early gene (for example, Fos and Egr1) expression in the RSC is striking and seems to be far greater than in comparison sites121. Even indirect disconnections can produce dramatic RSC hypoactivity, for example after mammillothalamic tract (MTT) lesions122 (see the figure: the upper right panel shows loss of Fos expression in the RSC after MTT lesion, whereas the lower right panel shows normal Fos expression in the anterior cingulate cortex (ACC)). Complementary microarray studies revealed a swathe of altered gene activity in the RSC following anterior thalamic lesions, particularly for genes involved in metabolism123.These same limbic lesions have little or no effect on RSC cell numbers120,124.
These findings suggest that the functional impact of some limbic lesions could be exacerbated by distal dysfunctions in the RSC. A direct test of this possibility125 showed that RSC brain slices from rats with anterior thalamic lesions fail to show normal synaptic plasticity. This result is persuasive as the slice preparation ensures that the RSC tissue is stimulated as per normal, but owing to a distal thalamic lesion — made weeks previously — it is not possible to induce long-term depression in the RSC125. This implies that anterior thalamic lesions induce ‘covert pathology’ in the RSC — that is, a seemingly intact cytoarchitecture combined with a persistent functional abnormality. To date, no studies have examined possible effects of RSC lesions on immediate-early gene activation in the hippocampus and anterior thalamic nuclei but, given the apparent interdependence of these structures and the effects of RSC lesions on anterior dorsal thalamic head-direction cells28 and hippocampal place cells40, it would be predicted that other functional measures would also be affected by RSC lesions and potentially contribute to subsequent behavioural impairments. Figure is reproduced, with permission, from ReF. 122 © (2009) Wiley.
R E V I E W S
nATuRE REVIEWS | NeuRoscieNce VoLuME 10 | noVEMBER 2009 | 795
© 2009 Macmillan Publishers Limited. All rights reserved

Place cellsCells that fire differentially depending on the animal’s location in an environment.
Grid cellsNeurons that fire strongly when an animal is at one of several specific locations in an environment and that are organized in a grid-like fashion.
Episodic memoryThe recollection of events with a specific spatial and temporal context, such as personal experiences. Often referred to as autobiographical memory.
the Rdg. Whether the two major RSC subregions can be functionally dissociated on the same task, and in the same study, remains to be determined.
Electrophysiology. Recording studies directly impli-cate the rodent RSC in navigation. Approximately 10% of RSC cells in the rat are ‘head-direction’ cells52. Such cells, along with ‘place cells’ and ‘grid cells’, form part of the triumvirate of the spatial navigation system. Head-direction cells are equally distributed across the granular and dysgranular RSC52. In the granular, but not the dys-granular, RSC the spatial tuning of some of these cells is modulated by aspects of behaviour such as the veloc-ity of locomotion52. In addition, some RSC cells fire in response to particular combinations of location, direc-tion and movement53, although actual place cells have not been detected in the RSC.
Head-direction cells are also found in other brain structures, including the postsubiculum and the anterior dorsal and lateral dorsal thalamic nuclei. Although these regions are reciprocally connected with the RSC, the RSC is not thought to be crucial for the generation of the head-direction signal28. nevertheless, lesions that include the caudal Rdg can disrupt anterior dorsal thalamic head-direction cells such that their preferred firing direction becomes less stable and likely to drift over time28. It is likely that the Rdg contributes visual information to the head-direction circuit, a view that is consistent with its anatomi-cal connections and the impact of selective Rdg lesions51. A similar situation may hold for the place cell circuit, as temporary inactivation of the RSC can transiently change the spatial tuning of hippocampal place cells while leaving other electrophysiological properties intact40.
Rhythmical slow-wave (theta) activity has been recorded in the RSC; it is locally generated and, unlike hippocampal theta activity, is only partially depend-ent on septal inputs54,55. Although RSC theta activity
is independent of hippocampal theta activity55, some aspects of hippocampal rhythmical slow activity (in the 8–12 Hz frequency range) are mediated by the RSC (and/or the underlying cingulum bundle)56. Cells in the RSC also respond differentially to different types of reward, and many RSC cells fire exclusively when animals make a physical response to acquire rewards (not during the pres-entation of conditioned stimuli)57. Finally, electrophysio-logical studies have found that RSC activity increases with the progression of learning an auditory avoidance con-ditioning task48. Parallel studies on the anterior thalamic nuclei led to the proposal that these regions work together to support both training-induced increases in attention to significant cues (for example, the conditioned stimulus) and subsequent cue-based retrieval48.
Together, findings from animal studies highlight the importance of the RSC for various behavioural tasks, in particular those that tax spatial memory or require the integration of different types of stimuli. The importance of the RSC to the latter is most apparent when animals are required to shift spatial strategies. It is also apparent that the RSC is closely interlinked with the hippocampus and the anterior thalamic nuclei.
Human neuropsychological studiesHuman studies also provide evidence for a role for the RSC in navigation and spatial memory. In addition, they have allowed investigations into topics that are less amena-ble to study in animals, such as episodic memory. As early as 1929, reports linked RSC pathology to memory disor-ders and spatial disorientation58, findings that have since been confirmed59–66. unsurprisingly, it has been difficult to exclude the possibility that neighbouring posterior cingulate areas61, the fornix60 or the hippocampus62 also contribute to these symptoms, and this caveat remains. It has also emerged that RSC dysfunction might contribute to Alzheimer’s disease from its earliest stages (BOX 2).
Patients with damage involving the RSC consistently show difficulty acquiring new information presented verbally or visually61. In addition to these learning defi-cits, patients generally have problems retrieving memo-ries of recent but not of very remote autobiographical events59,61,63,65,66. However, the extent of the retrograde amnesia varies from less than 1 year59 to 10 years62. Damage involving the RSC can also cause a selective def-icit in spatial orientation61,64,67–72. In most of these cases, patients can recognize neighbourhood landmarks but fail to navigate effectively in familiar environments, indicat-ing that they are unable to derive directional information from landmark cues. RSC damage can also compromise learning to navigate in new environments61. Patients with hippocampal lesions are also impaired at navigat-ing in familiar and novel environments (for example, see ReF. 73) but, unlike patients with RSC lesions, they are usually able to orient in familiar environments and can have a preserved sense of direction73,74.
In patients with unilateral RSC damage, the spa-tial deficit can resolve after several months61,68, and a neuro imaging study of such a patient suggests that this recovery is supported by the remaining RSC tissue64. Verbal and visual memory deficits are often, but not
Box 2 | Retrosplenial hypoactivity and memory loss
Primary lesions involving the retrosplenial cortex (RSC) can be caused by tumours, arteriovenous malformations, infarction and haemorrhage. Pathological changes in this region can also occur in conditions such as schizophrenia126, bipolar disorder127, post-traumatic stress disorder128, dyslexia129 and fibromyalgia81. More subtle changes, specifically reduced RSC activity, have been found in patients with temporal lobe or diencephalic amnesia130,131. Especially striking is the discovery that the earliest metabolic decline in Alzheimer’s disease, as measured using [18F]-fluorodeoxyglucose positron emission tomography, is centred on the RSC132,133. This finding has been shown in people with mild cognitive impairment (MCI), which is often a prodromal stage of Alzheimer’s disease. Such results are consistent with the link between retrosplenial hypoactivity and memory loss in patients with early Alzheimer’s disease110.
There are currently two explanations for this RSC hypoactivity. The first is that it is a secondary response to atrophy in other brain sites that are known to undergo pathological damage in early Alzheimer’s disease and that have direct RSC inputs133, such as the hippocampus, the entorhinal cortex and the anterior thalamic nuclei (FIG. 1). The second account, which is not mutually exclusive with the first, is that the RSC may itself atrophy in early Alzheimer’s disease134,135. Of particular relevance is an MRI analysis of patients with MCI who subsequently developed Alzheimer’s disease135. These incipient Alzheimer’s cases showed volume decreases in both the RSC and the adjacent posterior cingulate area 23 that were comparable to their hippocampal atrophy135. These findings show that RSC dysfunction is likely to be a major contributor to the cognitive deficits associated with MCI and Alzheimer’s disease133.
R E V I E W S
796 | noVEMBER 2009 | VoLuME 10 www.nature.com/reviews/neuro
© 2009 Macmillan Publishers Limited. All rights reserved

Rotational offsetsubtraction of the allocentric heading direction from the allocentric direction of an environmental landmark.
always, consistent with the side of the lesion; for exam-ple, there is a preponderance of right-sided RSC pathol-ogy in patients with selective spatial disorientation61, but left-sided lesions have also been reported64,70,72,75,76. This indicates that interpreting the cause of such deficits is difficult owing to the rarity of selective RSC lesions and, consequently, the possibility of undetected bilateral pathology.
The type of spatial deficit in patients (and animals) with RSC pathology, along with the anatomical con-nectivity of the RSC, has fostered the idea that the RSC has a ‘translational’ function61,67–69,77,78. It has been pro-posed that the RSC transforms allocentric representa-tions into egocentric representations and vice versa. In other words, the RSC helps to switch between egocen-tric, viewpoint-dependent (mediated by the posterior parietal cortex) and allocentric, viewpoint-independent (mediated by the medial temporal lobe) frames of refer-ence61,67,68,77,78. Based on a computational model, Burgess and colleagues77,78 proposed that the RSC uses head-direction information from the anterior thalamic nuclei to compensate for the rotational offset between egocen-tric and allocentric coordinates. This proposal has been extended to episodic and autobiographical memory77,78, according to which the hippocampus indexes the loca-tion that is embodied in an autobiographical memory, scene or imagined event and the RSC then translates this information into an egocentric representation so that the memory, scene or imagined event can be viewed from a specific viewpoint. For instance, a scene or event is constructed in a person’s hippocampus and medial temporal lobe. This is in an allocentric framework (for example, it is built in relation to a landmark 10 metres north, where north is relative to the environmental cues driving the head-direction system). This representation has to be translated into egocentric directions to enable a person to act, such as left or right from the current (or imagined) direction of view (for example, a landmark 10 metres ahead if the viewpoint is north or 10 metres to the left if the viewpoint is east). In this model77,78 the RSC is involved in the translation process, possibly by acting as a short-term buffer for the representations as they are being translated77,78.
Neuroimaging in humansMemory and spatial navigation. The location of the RSC poses severe challenges to techniques like magneto-encephalography and transcranial magnetic stimulation. By contrast, functional MRI (fMRI) has been so success-ful that activity in the RSC region has been reported to be modulated by a bewildering array of tasks or processes, ranging from basic speech production and comprehen-sion79 to motivation80 and pain81. Most striking, however, is its almost ubiquitous engagement in studies of spatial navigation61,68 and episodic memory82,83, in which it is often the brightest ‘blob’ on the brain activation maps. For example, in a recent meta-analysis82 the RSC emerged as one of a number of brain regions that consistently showed significant activation during autobiographical memory retrieval. This meta-analysis also highlights a recurrent limitation on ascertaining the function of the RSC from
imaging studies, as the authors82 placed studies showing RSC activation and studies showing activation of other posterior cingulate regions into one category. Despite this generalization, plots of activation peaks from the studies they surveyed clearly implicate the right and left RSC in numerous studies82. In some instances, greater RSC activity has been observed during the retrieval of recent versus remote autobiographical experiences84–86. This pat-tern accords with neuropsychological studies in which damage involving the RSC seemed to disproportionately affect the retrieval of recent events59,61,63,65,66. However, in general, the RSC is active during retrieval of any kind of past autobiographical experience, be it recent or remote, emotional87 or neutral82,83.
The RSC is also consistently engaged by spatial navi-gation tasks61,68,74,88–90. These include passive viewing of navigation footage, mental navigation and interactive navigation in virtual-reality environments. The RSC is also active during learning of new environments, navi-gation in recently learned environments and naviga-tion in very familiar environments. Indeed, it is hard to find a navigation or topographical memory task in which the RSC is not activated (for example, see ReF. 91). Importantly, however, some studies have a rather liberal interpretation about what constitutes the RSC2: activa-tion in posterior cingulate areas 23 and 31 (ReFs 92,93), in the parietal-occipital sulcus and in the anterior calcarine region92 has been misattributed to the RSC.
In an effort to determine when during navigation different brain areas are engaged, Spiers and Maguire90 scanned participants navigating through a highly realistic virtual-reality simulation of central London. The analy-sis revealed a complex choreography of neural dynamics: they were both focal and distributed, and both transient and sustained. Activity in the RSC increased specifi-cally when topographical representations needed to be updated, integrated or manipulated for route planning, or when new topographical information was acquired90. Thus, RSC processing is not tonically maintained dur-ing navigation in familiar environments; rather, its activity alters depending on specific circumstances and priorities. This could relate to the proposal that the RSC acts as a short-term buffer for translating between representations77,78, as outlined above.
There has also been considerable interest in the RSC’s role in processing scenes68,94. Bar suggested that rather than supporting spatial navigation and orientation, the RSC (and other ‘scene areas’ like the parahippocam-pal cortex) might process scene-relevant relationships between objects and their contexts94. In support of this hypothesis, one study95 found that the RSC was engaged by viewing objects strongly associated with a specific context more so than by viewing objects that were weakly associated with a context. Recently, Henderson et al.96 directly examined whether the RSC was engaged by three-dimensional geometric structure in scenes (which might underpin its role in navigation) and contrasted this with its purported role in contextual associations. Their findings were not compatible with the contextual-associations hypothesis93 but were consistent with the idea that the RSC supports spatial navigation.
R E V I E W S
nATuRE REVIEWS | NeuRoscieNce VoLuME 10 | noVEMBER 2009 | 797
© 2009 Macmillan Publishers Limited. All rights reserved

Imagination and thinking about the future. In recent years a new theme has emerged in memory research, based on two observations. First, Hassabis et al.97 reported that patients with bilateral hippocampal dam-age not only are unable to recollect past experiences but also are impaired at imagining fictitious and future expe-riences. Second, recalling past experiences was found to activate many of the same brain regions as imagining plausible personal future98–102 and fictitious103 experi-ences. These areas included the hippocampus and the RSC99,101–103. Moreover, these brain areas were also active during spatial navigation (see above), theory of mind tasks104, daydreaming105 and when the brain is in a ‘rest-ing state’ (BOX 3). A meta-analysis106 of studies from a number of these domains confirmed that the RSC is part of a common ‘core network’ that subserves a range of cognitive functions. This implies that the hippocam-pus and the RSC are engaged in a type of processing that may not be purely mnemonic in nature but that is
nevertheless crucial for memory. Several theories have been advanced to account for the functions of this ‘core network’104,107,108. For example, Hassabis and Maguire107 suggested that this network supports ‘scene construction’ — the process of mentally generating and maintaining a complex and coherent scene or event — which they believe underpins functions such as autobiographical memory, navigation and thinking about the future.
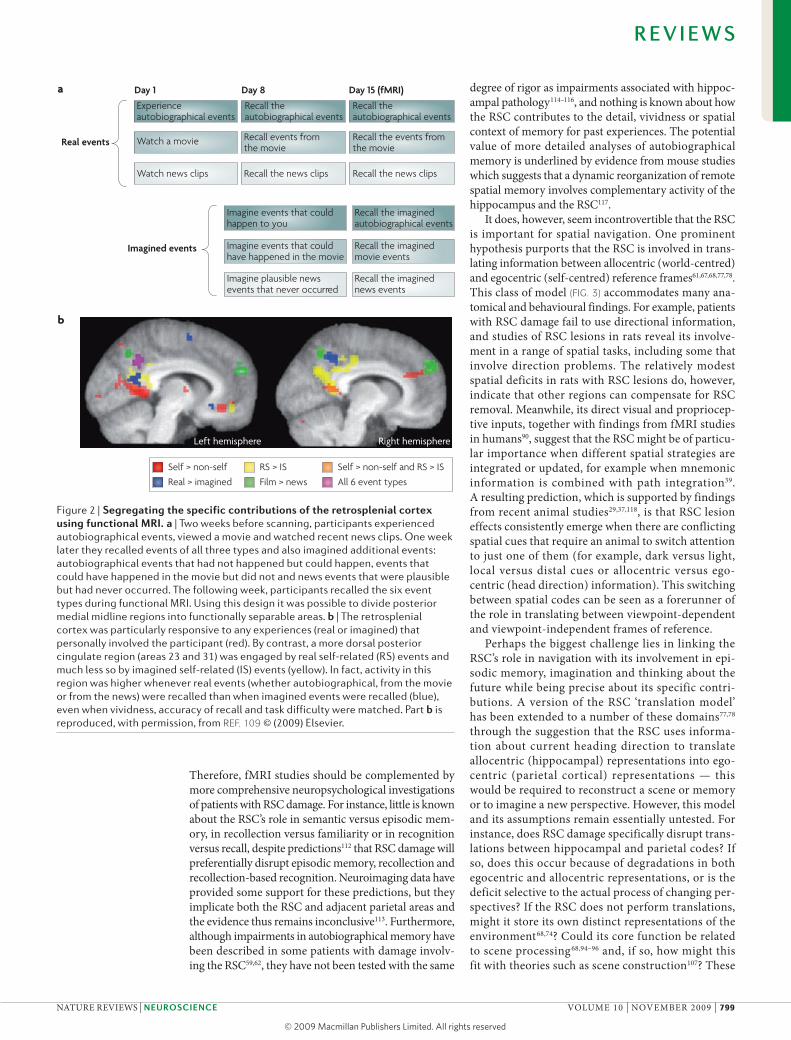
Linking memory and navigation with imagination and thinking about the future, and placing them in the wider context of cognition, opens up new perspectives for understanding the role of the RSC. For example, a recent study used fMRI to examine brain activation during retrieval of autobiographical events, of episodes from a movie and of real news clips, as well as previously imagined events of all three types109 (FIG. 2). Crucially, activation patterns in the RSC differed from those in posterior cingulate areas 23 and 31: parts of areas 23 and 31 were more active during retrieval of real events than during retrieval of imagined events across all categories, which suggests that re-experiencing real events modu-lates activity in this area (FIG. 2). By contrast, the ventral portion of the parietal-occipital sulcus, extending into the left and right RSC, was activated specifically by real and imagined autobiographical events that involved the participant personally. It is possible that the recollection of (real or imagined) autobiographical events requires participants to constantly update their first-person per-spective relative to the changing spatial context of the environment as they progress through the unfolding event. By contrast, the detached, observer perspective from which movie and most news events are recalled requires less self-referencing to specific spatial contexts. This interpretation fits with the proposed function of the RSC in spatial updating and translating between egocen-tric and allocentric frames of reference outlined above.
Conclusions and future directionsEarly studies characterized the memory impairments following RSC damage as disconnection effects resulting from the interruption of pathways from the hippocampus to either the anterior thalamic nuclei59 or the frontal cor-tex18. Although anatomical studies give some credence to this view, they also highlight its shortcomings. As illustrated in FIG. 1, some key mnemonic structures link through the RSC, but they also have direct interconnec-tions. Consequently, the focus has shifted to the contri-butions that the RSC per se makes to memory. This shift is important given the growing evidence of pervasive RSC dysfunction in conditions like Alzheimer’s disease (BOX 2); such dysfunction might contribute to the cogni-tive deficits from the earliest stages of this disease110,111. Determining the relationship between RSC dysfunction and symptoms in Alzheimer’s disease is complicated by the potential for ‘covert’ pathology in the RSC (BOX 1).
Both functional neuroimaging and clinical studies suggest that the RSC is involved in a range of cogni-tive functions, although there is a continued need for greater anatomical precision in these studies. Indeed, the propensity for RSC activation in fMRI studies makes it all the more difficult to determine its precise functions.
Box 3 | The retrosplenial cortex and the default mode network
When participants are not performing tasks during functional neuroimaging, they show a characteristic pattern of brain activity involving the medial frontal and medial temporal lobe regions, lateral and medial parietal areas and the retrosplenial cortex (RSC)136. The areas that are active in this ‘resting brain state’ are together also known as the ‘default mode network’137. Although some default mode network areas show a decrease in activity once a participant engages in a cognitive task136, the RSC response during tests of spatial navigation and autobiographical memory retrieval is significantly above the resting-state baseline, reflecting its true engagement during such tasks61,68,82,83. There are temporal correlations in functional MRI (fMRI) signals between areas in the default mode network138, which might reflect the interconnectivity between regions such as the RSC and the hippocampus. There might also be subsystems in the default mode network that converge on two main hubs — one consisting of posterior cingulate cortex (PCC) areas 23 and 31 and the RSC, and the other lying in the medial prefrontal cortex139. Interestingly, although the default mode network changes across age groups, PCC and RSC involvement is observed consistently, even in 2-week-old infants140, suggesting that the PCC and RSC together are the main hub of the default mode network.
Greicius et al.141 combined resting state fMRI and diffusion tensor imaging (DTI) to assess whether resting-state functional connectivity maps reflect neural connectivity. Using seed regions from functional connectivity maps, DTI revealed robust structural connections (orange fibres in the figure) between the RSC and the medial temporal lobes (green and purple areas), whereas tracts from the medial prefrontal cortex (yellow area) contact the PCC just rostral to the RSC (blue fibres). Fibre tracts connecting the RSC and the medial temporal lobes have previously been identified in macaque tracer studies (see the main text) but have not been shown in humans. Combining fMRI with DTI might, therefore, prove useful for characterizing normal RSC connectivity in humans and for assessing the impact of pathologies such as Alzheimer’s disease (BOX 2). Figure is reproduced, with permission, from ReF. 141 © (2009) Oxford Journals.
R E V I E W S
798 | noVEMBER 2009 | VoLuME 10 www.nature.com/reviews/neuro
© 2009 Macmillan Publishers Limited. All rights reserved
Day 1 Day 8 Day 15 (fMRI)
Experience autobiographical events
Watch a movie
Recall the autobiographical events
Recall events from the movie
Recall the autobiographical events
Recall the events from the movie
Recall the news clipsWatch news clips Recall the news clips
Real events
Imagined events
Recall the imagined autobiographical events
Imagine events that could happen to you
Recall the imagined movie events
Recall the imagined news events
Imagine plausible news events that never occurred
Imagine events that could have happened in the movie
Nature Reviews | Neuroscience
Self > non-self
Real > imagined
RS > IS
Film > news
Self > non-self and RS > IS
All 6 event types
a
b
Left hemisphere Right hemisphere
Therefore, fMRI studies should be complemented by more comprehensive neuropsychological investigations of patients with RSC damage. For instance, little is known about the RSC’s role in semantic versus episodic mem-ory, in recollection versus familiarity or in recognition versus recall, despite predictions112 that RSC damage will preferentially disrupt episodic memory, recollection and recollection-based recognition. neuroimaging data have provided some support for these predictions, but they implicate both the RSC and adjacent parietal areas and the evidence thus remains inconclusive113. Furthermore, although impairments in autobiographical memory have been described in some patients with damage involv-ing the RSC59,62, they have not been tested with the same
degree of rigor as impairments associated with hippoc-ampal pathology114–116, and nothing is known about how the RSC contributes to the detail, vividness or spatial context of memory for past experiences. The potential value of more detailed analyses of autobiographical memory is underlined by evidence from mouse studies which suggests that a dynamic reorganization of remote spatial memory involves complementary activity of the hippocampus and the RSC117.
It does, however, seem incontrovertible that the RSC is important for spatial navigation. one prominent hypothesis purports that the RSC is involved in trans-lating information between allocentric (world-centred) and egocentric (self-centred) reference frames61,67,68,77,78. This class of model (FIG. 3) accommodates many ana-tomical and behavioural findings. For example, patients with RSC damage fail to use directional information, and studies of RSC lesions in rats reveal its involve-ment in a range of spatial tasks, including some that involve direction problems. The relatively modest spatial deficits in rats with RSC lesions do, however, indicate that other regions can compensate for RSC removal. Meanwhile, its direct visual and propriocep-tive inputs, together with findings from fMRI studies in humans90, suggest that the RSC might be of particu-lar importance when different spatial strategies are integrated or updated, for example when mnemonic information is combined with path integration39. A resulting prediction, which is supported by findings from recent animal studies29,37,118, is that RSC lesion effects consistently emerge when there are conflicting spatial cues that require an animal to switch attention to just one of them (for example, dark versus light, local versus distal cues or allocentric versus ego-centric (head direction) information). This switching between spatial codes can be seen as a forerunner of the role in translating between viewpoint-dependent and viewpoint-independent frames of reference.
Perhaps the biggest challenge lies in linking the RSC’s role in navigation with its involvement in epi-sodic memory, imagination and thinking about the future while being precise about its specific contri-butions. A version of the RSC ‘translation model’ has been extended to a number of these domains77,78 through the suggestion that the RSC uses informa-tion about current heading direction to translate allocentric (hippocampal) representations into ego-centric (parietal cortical) representations — this would be required to reconstruct a scene or memory or to imagine a new perspective. However, this model and its assumptions remain essentially untested. For instance, does RSC damage specifically disrupt trans-lations between hippocampal and parietal codes? If so, does this occur because of degradations in both egocentric and allocentric representations, or is the deficit selective to the actual process of changing per-spectives? If the RSC does not perform translations, might it store its own distinct representations of the environment68,74? Could its core function be related to scene processing68,94–96 and, if so, how might this fit with theories such as scene construction107? These
Figure 2 | segregating the specific contributions of the retrosplenial cortex using functional MRi. a | Two weeks before scanning, participants experienced autobiographical events, viewed a movie and watched recent news clips. One week later they recalled events of all three types and also imagined additional events: autobiographical events that had not happened but could happen, events that could have happened in the movie but did not and news events that were plausible but had never occurred. The following week, participants recalled the six event types during functional MRI. Using this design it was possible to divide posterior medial midline regions into functionally separable areas. b | The retrosplenial cortex was particularly responsive to any experiences (real or imagined) that personally involved the participant (red). By contrast, a more dorsal posterior cingulate region (areas 23 and 31) was engaged by real self-related (RS) events and much less so by imagined self-related (IS) events (yellow). In fact, activity in this region was higher whenever real events (whether autobiographical, from the movie or from the news) were recalled than when imagined events were recalled (blue), even when vividness, accuracy of recall and task difficulty were matched. Part b is reproduced, with permission, from ReF. 109 © (2009) Elsevier.
R E V I E W S
nATuRE REVIEWS | NeuRoscieNce VoLuME 10 | noVEMBER 2009 | 799
© 2009 Macmillan Publishers Limited. All rights reserved

Nature Reviews | Neuroscience
Prefrontal cortexExecutive, scene manipulation
Retrosplenial cortexScene translation
Parietal cortexBody-oriented information
Occipital cortexVisual information
Allocentric framework
Egocentricframework
ATNHead direction, theta
Perirhinal cortexObject-based information
ParahippocampalScene-based information
HippocampusEvent within a scene,scene construction
key questions could be addressed by assessing patients with RSC damage on their ability to imagine ficti-tious and future experiences and on how they process scenes, and by using fMRI studies with task designs that specifically target the RSC. The RSC has too long
been the poor relation of cognitive neuroscience. It is our hope that the RSC will now become a major focus of dedicated research, and our belief that defining its role will prove pivotal in understanding a range of crucial cognitive functions.
Figure 3 | The key anatomical and functional relationships of the retrosplenial cortex. Effective episodic memory, navigation and future thinking all require the ability to integrate and manipulate different frameworks of information, for example egocentric (self-centred) and allocentric (world-centred) frameworks. By virtue of its principal connections, the retrosplenial cortex is uniquely placed to enable translation within these domains. ATN, anterior thalamic nuclei.
1. Brodmann, K. Vergleichende Lokalisationslehre der Grosshirnrinde inihren Prinzipien dargestellt auf Grund des Zellenbaues (Barth, Leipzig, 1909).
2. Vogt, B. A., Absher, J. R. & Bush, G. Human retrosplenial cortex: where is it and is it involved in emotion? Trends Neurosci. 23, 195–197 (2000).
3. Papez, J. W. A proposed mechanism of emotion. Arch. Neurol. Psychiatry 38, 725–743 (1937).
4. MacLean, P. Psychosomatic disease and the visceral brain; recent developments bearing on the Papez theory of emotion. Psychosom. Med. 11, 338–353 (1949).
5. Vogt, B. A., Nimchinsky, E. A., Vogt, L. J. & Hof, P. R. Human cingulate cortex: surface features, flat maps, and cytoarchitecture. J. Comp. Neurol. 359, 490–506 (1995).
6. Cavanna, A. E. & Trimble, M. R. The precuneus: a review of its functional anatomy and behavioural correlates. Brain 129, 564–583 (2006).
7. Kobayashi, Y. & Amaral, D. G. Macaque monkey retrosplenial cortex: II. Cortical afferents. J. Comp. Neurol. 466, 48–79 (2003).
8. Kobayashi, Y. & Amaral, D. G. Macaque monkey retrosplenial cortex: I. Three-dimensional and cytoarchitectonic organization. J. Comp. Neurol. 426, 339–365 (2000).
9. Vogt, B. A., Vogt, L., Farber, N. B. & Bush, G. Architecture and neurocytology of monkey cingulate gyrus. J. Comp. Neurol. 485, 218–239 (2005).
10. Morris, R., Petrides, M. & Pandya, D. N. Architecture and connections of retrosplenial area 30 in the rhesus monkey (Macaca mulatta). Eur. J. Neurosci. 11, 2506–2518 (1999).
11. Vogt, B. A. in Cingulate Neurobiology and Disease (ed. Vogt, B. A.) 65–93 (Oxford Univ. Press, London, 2009).A chapter from a recent book devoted to the entire cingulate cortex, of which the RSC is a component area. This is the first book devoted to the neurobiology of this region since 1993.
12. Vogt, B. A. & Peters, A. Form and distribution of neurons in rat cingulate cortex: areas 32, 24, and 29. J. Comp. Neurol. 195, 603–625 (1981).
13. Van Groen, T. & Wyss, J. M. Connections of the retrosplenial granular a cortex in the rat. J. Comp. Neurol. 300, 593–606 (1990).
14. Van Groen, T. & Wyss, J. M. Connections of the retrosplenial granular b cortex in the rat. J. Comp. Neurol. 463, 249–263 (2003).
15. Van Groen, T. & Wyss, J. M. Connections of the retrosplenial dysgranular cortex in the rat. J. Comp. Neurol. 315, 200–216 (1992).
16. Kobayashi, Y. & Amaral, D. G. Macaque monkey retrosplenial cortex: III. Cortical efferents. J. Comp. Neurol. 502, 810–833 (2007).The final paper in a series of three that provide the most detailed descriptions to date of the inputs to the RSC in an Old World monkey brain.
17. Mufson, E. J. & Pandya, D. N. Some observations on the course and composition of the cingulum bundle in the rhesus monkey. J. Comp. Neurol. 225, 31–43 (1984).
18. Morris, R., Pandya, D. N. & Petrides, M. Fiber system linking the mid-dorsolateral frontal cortex with the retrosplenial/presubicular region in the rhesus monkey. J. Comp. Neurol. 407, 183–192 (1999).
19. Jones, B. F., Groenewegen, H. J. & Witter, M. P. Intrinsic connections of the cingulate cortex in the rat suggest the existence of multiple functionally segregated networks. Neuroscience 133, 193–207 (2005).
20. Vogt, B. A. & Miller, M. W. Cortical connections between rat cingulate cortex and visual, motor, and postsubicular cortices. J. Comp. Neurol. 216, 192–210 (1983).
21. Vogt, B. A. in Cerebral Cortex: Association and Auditory Cortices Vol. 4 (eds Peters, A. & Jones, E.) 89–149 (Plenum, New York, 1985).
22. Sutherland, R. J. & Hoesing, J. M. in Neurobiology of Cingulate Cortex and Limbic Thalamus: A
R E V I E W S
800 | noVEMBER 2009 | VoLuME 10 www.nature.com/reviews/neuro
© 2009 Macmillan Publishers Limited. All rights reserved

Comprehensive Treatise (eds Vogt, B. A. & Gabriel, M.) 461–477 (Birkhauser, Boston, 1993).
23. Warburton, E. C., Baird, A., Morgan, A., Muir, J. L. & Aggleton, J. P. The conjoint importance of the hippocampus and anterior thalamic nuclei for allocentric spatial learning: evidence from a disconnection study in the rat. J. Neurosci. 21, 7323–7330 (2001).
24. Aggleton, J. P. & Vann, S. D. Testing the importance of the retrosplenial navigation system: lesion size but not strain matters: a reply to Harker and Whishaw. Neurosci. Biobehav. Rev. 28, 525–531 (2004).
25. Harker, K. T. & Whishaw, I. Q. A reaffirmation of the retrosplenial contribution to rodent navigation: reviewing the influences of lesion, strain, and task. Neurosci. Biobehav. Rev. 28, 485–496 (2004).
26. Meunier, M. & Destrade, C. Effects of radiofrequency versus neurotoxic cingulate lesions on spatial reversal learning in mice. Hippocampus 7, 355–360 (1997).
27. Vann, S. D., Brown, M. W. & Aggleton, J. P. Fos expression in the rostral thalamic nuclei and associated cortical regions in response to different spatial memory tests. Neuroscience 101, 983–991 (2000).
28. Taube, J. S. The head direction signal: origins and sensory-motor integration. Annu. Rev. Neurosci. 30, 181–207 (2007).
29. Vann, S. D. & Aggleton, J. P. Testing the importance of the retrosplenial guidance system: effects of different sized retrosplenial cortex lesions on heading direction and spatial working memory. Behav. Brain Res. 155, 97–108 (2004).Here, rats with RSC lesions that differed in size along the RSC’s rostrocaudal axis were compared for effects on different aspects of spatial memory. Rats with lesions of the whole RSC were consistently more impaired than rats with the more standard lesion that spared the most caudal extent of the structure. This study clarified some of the previous inconsistencies in the literature.
30. Cain, D. P., Humpartzoomian, R. & Boon, F. Retrosplenial cortex lesions impair water maze strategies learning or spatial place learning depending on prior experience of the rat. Behav. Brain Res. 170, 316–325 (2006).
31. Vann, S. D. & Aggleton, J. P. Extensive cytotoxic lesions of the rat retrosplenial cortex reveal consistent deficits on tasks that tax allocentric spatial memory. Behav. Neurosci. 116, 85–94 (2002).
32. Sutherland, R. J., Whishaw, I. Q. & Kolb, B. Contributions of cingulate cortex to two forms of spatial learning and memory. J. Neurosci. 8, 1863–1872 (1988).
33. Harker, K. T. & Whishaw, I. Q. Impaired place navigation in place and matching-to-place swimming pool tasks follows both retrosplenial cortex lesions and cingulum bundle lesions in rats. Hippocampus 14, 224–231 (2004).
34. Whishaw, I. Q., Maaswinkel, H., Gonzalez, C. L. & Kolb, B. Deficits in allothetic and idiothetic spatial behavior in rats with posterior cingulate cortex lesions. Behav. Brain Res. 118, 67–76 (2001).
35. Vann, S. D., Kristina Wilton, L. A., Muir, J. L. & Aggleton, J. P. Testing the importance of the caudal retrosplenial cortex for spatial memory in rats. Behav. Brain Res. 140, 107–118 (2003).
36. St-Laurent, M., Petrides, M. & Sziklas, V. Does the cingulate cortex contribute to spatial conditional associative learning in the rat? Hippocampus 19, 612–622 (2009).
37. Pothuizen, H. H., Aggleton, J. P. & Vann, S. D. Do rats with retrosplenial cortex lesions lack direction? Eur. J. Neurosci. 28, 2486–2498 (2008).
38. Cooper, B. G. & Mizumori, S. J. Retrosplenial cortex inactivation selectively impairs navigation in darkness. Neuroreport 10, 625–630 (1999).
39. Cooper, B. G., Manka, T. F. & Mizumori, S. J. Finding your way in the dark: the retrosplenial cortex contributes to spatial memory and navigation without visual cues. Behav. Neurosci. 115, 1012–1028 (2001).
40. Cooper, B. G. & Mizumori, S. J. Temporary inactivation of the retrosplenial cortex causes a transient reorganization of spatial coding in the hippocampus. J. Neurosci. 21, 3986–4001 (2001).
41. Amin, E., Pearce, J. M., Brown, M. W. & Aggleton, J. P. Novel temporal configurations of stimuli produce discrete changes in immediate-early gene expression in the rat hippocampus. Eur. J. Neurosci. 24, 2611–2621 (2006).
42. Markowska, A. L., Olton, D. S., Murray, E. A. & Gaffan, D. A comparative analysis of the role of fornix and cingulate cortex in memory: rats. Exp. Brain Res. 74, 187–201 (1989).
43. Sziklas, V., Petrides, M. & Leri, F. The effects of lesions to the mammillary region and the hippocampus on conditional associative learning by rats. Eur. J. Neurosci. 8, 106–115 (1996).
44. Dumont, J., Petrides, M. & Sziklas, V. Functional dissociation between fornix and hippocampus in spatial conditional learning. Hippocampus 17, 1170–1179 (2007).
45. Keene, C. S. & Bucci, D. J. Involvement of the retrosplenial cortex in processing multiple conditioned stimuli. Behav. Neurosci. 122, 651–658 (2008).
46. Keene, C. S. & Bucci, D. J. Contributions of the retrosplenial and posterior parietal cortices to cue-specific and contextual fear conditioning. Behav. Neurosci. 122, 89–97 (2008).
47. Lukoyanov, N. V. & Lukoyanova, E. A. Retrosplenial cortex lesions impair acquisition of active avoidance while sparing fear-based emotional memory. Behav. Brain Res. 173, 229–236 (2006).
48. Gabriel, M. & Talk, A. C. in Model Systems and the Neurobiology of Associative Learning (eds Steinmetz, J. A., Gluck, M. & Soloman, R. R.) 149–185 (Erlbaum, Mahwah, New Jersey, 2001).
49. Pothuizen, H. H., Davies, M., Albasser, M. M., Aggleton, J. P. & Vann, S. D. Granular and dysgranular retrosplenial cortices provide qualitatively different contributions to spatial working memory: evidence from immediate-early gene imaging in rats. Eur. J. Neurosci. 30, 877–888 (2009).Rats were trained on spatial working memory tasks or control tasks, in the radial-arm maze, in either the presence or the absence of visual cues (that is, in the light or in the dark). There was increased immediate-early gene activity in the granular RSC in the working memory group both in the light and in the dark conditions, but the dysgranular RSC was involved in spatial memory only in the presence of visual cues (that is, in the light).
50. Van Groen, T., Kadish, I. & Wyss, J. M. Retrosplenial cortex lesions of area Rgb (but not of area Rga) impair spatial learning and memory in the rat. Behav. Brain Res. 154, 483–491 (2004).
51. Vann, S. D. & Aggleton, J. P. Selective dysgranular retrosplenial cortex lesions in rats disrupt allocentric performance of the radial-arm maze task. Behav. Neurosci. 119, 1682–1686 (2005).
52. Chen, L. L., Lin, L. H., Green, E. J., Barnes, C. A. & McNaughton, B. L. Head-direction cells in the rat posterior cortex. I. Anatomical distribution and behavioral modulation. Exp. Brain Res. 101, 8–23 (1994).
53. Cho, J. W. & Sharp, P. E. Head direction, place, and movement correlates for cells in the rat retrosplenial cortex. Behav. Neurosci. 115, 3–25 (2001).
54. Borst, J. G., Leung, L. W. & MacFabe, D. F. Electrical activity of the cingulate cortex. II. Cholinergic modulation. Brain Res. 407, 81–93 (1987).
55. Talk, A., Kang, E. & Gabriel, M. Independent generation of theta rhythm in the hippocampus and posterior cingulate cortex. Brain Res. 1015, 15–24 (2004).A dissociation was found between the theta activity of cue-elicited field potentials and multiple-unit activity recordings in the RSC. Retrosplenial theta activity, as recorded by field potentials, was attenuated by dorsal hippocampal lesions. By contrast, theta-like unit activity in the granular b (29b) RSC was unaffected by the same lesions and, therefore, occurs independently of the hippocampus.
56. Destrade, C. & Ott, T. Is a retrosplenial (cingulate) pathway involved in the mediation of high frequency hippocampal rhythmical slow activity (theta)? Brain Res. 252, 29–37 (1982).
57. Tabuchi, E. et al. Neural correlates to action and rewards in the rat posterior cingulate cortex. Neuroreport 16, 949–953 (2005).
58. Ironside, R. & Guttmacher, M. The corpus callosum and its tumours. Brain 52, 442–483 (1929).
59. Valenstein, E. et al. Retrosplenial amnesia. Brain 110, 1631–1646 (1987).
60. Rudge, P. & Warrington, E. K. Selective impairment of memory and visual perception in splenial tumours. Brain 114, 349–360 (1991).
61. Maguire, E. A. The retrosplenial contribution to human navigation: a review of lesion and neuroimaging findings. Scand. J. Psychol. 42, 225–238 (2001).
A range of single case reports of patients with RSC damage are reviewed. The effect on the patients’ spatial navigation is considered in conjunction with functional neuroimaging studies of navigation that results in RSC activation.
62. Gainotti, G., Almonti, S., Betta, A. M. D. & Silveri, M. C. Retrograde amnesia in a patient with retrosplenial tumour. Neurocase 4, 519–526 (1998).
63. McDonald, C. R., Crosson, B., Valenstein, E. & Bowers, D. Verbal encoding deficits in a patient with a left retrosplenial lesion. Neurocase 7, 407–417 (2001).
64. Ino, T. et al. Directional disorientation following left retrosplenial hemorrhage: a case report with fMRI studies. Cortex 43, 248–254 (2007).
65. Oka, Y. et al. A case of amnesia caused by a subcortical hematoma in the left retrosplenial region. No Shinkei Geka 31, 289–295 (2003) (in Japanese).
66. Osawa, A., Maeshima, S., Kubo, K. & Itakura, T. Neuropsychological deficits associated with a tumour in the posterior corpus callosum: a report of two cases. Brain Inj. 20, 673–676 (2006).
67. Aguirre, G. K. & D’Esposito, M. Topographical disorientation: a synthesis and taxonomy. Brain 122, 1613–1628 (1999).
68. Epstein, R. A. Parahippocampal and retrosplenial contributions to human spatial navigation. Trends Cogn. Sci. 12, 388–396 (2008).
69. Takahashi, N., Kawamura, M., Shiota, J., Kasahata, N. & Hirayama, K. Pure topographic disorientation due to right retrosplenial lesion. Neurology 49, 464–469 (1997).
70. Alsaadi, T., Binder, J. R., Lazar, R. M., Doorani, T. & Mohr, J. P. Pure topographic disorientation: a distinctive syndrome with varied localization. Neurology 54, 1864–1866 (2000).
71. Greene, K. K., Donders, J. & Thoits, T. Topographical heading disorientation: a case study. Appl. Neuropsychol. 13, 269–274 (2006).
72. Osawa, A., Maeshima, S. & Kunishio, K. Topographic disorientation and amnesia due to cerebral hemorrhage in the left retrosplenial region. Eur. Neurol. 59, 79–82 (2008).
73. Maguire, E. A., Nannery, R. & Spiers, H. J. Navigation around London by a taxi driver with bilateral hippocampal lesions. Brain 129, 2894–2907 (2006).
74. Spiers, H. J. & Maguire, E. A. The neuroscience of remote spatial memory: a tale of two cities. Neuroscience 149, 7–27 (2007).
75. Obi, T., Bando, M., Takeda, K. & Sakuta, M. A case of topographical disturbance following a left medial parieto-occipital lobe infarction. Rinsho Shinkeigaku 32, 426–429 (1992) (in Japanese).
76. Maeshima, S. et al. Memory impairment and spatial disorientation following a left retrosplenial lesion. J. Clin. Neurosci. 8, 450–451 (2001).
77. Burgess, N., Becker, S., King, J. A. & O’Keefe, J. Memory for events and their spatial context: models and experiments. Philos. Trans. R. Soc. Lond. B Biol. Sci. 356, 1493–1503 (2001).Presents a neural-level model of the medial temporal and parietal roles in retrieval of the spatial context of an event. The central tenet is that medial temporal allocentric representations are used in long-term storage, whereas parietal egocentric representations are used to imagine, manipulate and re-experience the products of retrieval, with the RSC translating between the two codes.
78. Byrne, P., Becker, S. & Burgess, N. Remembering the past and imagining the future: a neural model of spatial memory and imagery. Psychol. Rev. 114, 340–375 (2007).The authors modelled the neural mechanisms underlying spatial cognition by integrating neuronal systems and behavioural data, and addressed the relationships between long-term memory, short-term memory and imagery, between egocentric and allocentric representations and between visual and ideothetic representations. A key role for the RSC is outlined in translating between reference frames.
79. Awad, M., Warren, J. E., Scott, S. K., Turkheimer, F. E. & Wise, R. J. A common system for the comprehension and production of narrative speech. J. Neurosci. 27, 11455–11464 (2007).
80. Small, D. M., Zatorre, R. J., Dagher, A., Evans, A. C. & Jones-Gotman, M. Changes in brain activity related to eating chocolate: from pleasure to aversion. Brain 124, 1720–1733 (2001).
81. Wik, G. et al. Retrospenial cortical deactivation during painful stimulation of fibromyalgic patients. Int. J. Neurosci. 116, 1–8 (2006).
R E V I E W S
nATuRE REVIEWS | NeuRoscieNce VoLuME 10 | noVEMBER 2009 | 801
© 2009 Macmillan Publishers Limited. All rights reserved

82. Svoboda, E., McKinnon, M. C. & Levine, B. The functional neuroanatomy of autobiographical memory: a meta-analysis. Neuropsychologia 44, 2189–2208 (2006).
83. Maguire, E. A. Neuroimaging studies of autobiographical event memory. Philos. Trans. R. Soc. Lond. B Biol. Sci. 356, 1441–1451 (2001).
84. Gilboa, A., Winocur, G., Grady, C. L., Hevenor, S. J. & Moscovitch, M. Remembering our past: functional neuroanatomy of recollection of recent and very remote personal events. Cereb. Cortex 14, 1214–1225 (2004).
85. Woodard, J. L. et al. Temporally graded activation of neocortical regions in response to memories of different ages. J. Cogn. Neurosci. 19, 1113–1124 (2007).
86. Oddo, S. et al. Specific role of medial prefrontal cortex in retrieving recent autobiographical memories: an fMRI study of young female subjects. Cortex 7 Nov 2008 (doi:10.1016/j.cortex.2008.07.003).
87. Maddock, R. J. The retrosplenial cortex and emotion: new insights from functional neuroimaging of the human brain. Trends Neurosci. 22, 310–316 (1999).
88. Burgess, N., Maguire, E. A. & O’Keefe, J. The human hippocampus and spatial and episodic memory. Neuron 35, 625–641 (2002).
89. Bird, C. M. & Burgess, N. The hippocampus and memory: insights from spatial processing. Nature Rev. Neurosci. 9, 182–194 (2008).
90. Spiers, H. J. & Maguire, E. A. Thoughts, behaviour, and brain dynamics during navigation in the real world. Neuroimage 31, 1826–1840 (2006).
91. Rosenbaum, R. S., Winocur, G., Grady, C. L., Ziegler, M. & Moscovitch, M. Memory for familiar environments learned in the remote past: fMRI studies of healthy people and an amnesic person with extensive bilateral hippocampal lesions. Hippocampus 17, 1241–1251 (2007).
92. Epstein, R. A., Parker, W. E. & Feiler, A. M. Where am I now? Distinct roles for parahippocampal and retrosplenial cortices in place recognition. J. Neurosci. 27, 6141–6149 (2007).
93. Wolbers, T. & Buchel, C. Dissociable retrosplenial and hippocampal contributions to successful formation of survey representations. J. Neurosci. 25, 3333–3340 (2005).
94. Bar, M. Visual objects in context. Nature Rev. Neurosci. 5, 617–629 (2004).
95. Bar, M. & Aminoff, E. Cortical analysis of visual context. Neuron 38, 347–358 (2003).
96. Henderson, J. M., Larson, C. L. & Zhu, D. C. Full scenes produce more activation than close-up scenes and scene-diagnostic objects in parahippocampal and retrosplenial cortex: an fMRI study. Brain Cogn. 66, 40–49 (2008).
97. Hassabis, D., Kumaran, D., Vann, S. D. & Maguire, E. A. Patients with hippocampal amnesia cannot imagine new experiences. Proc. Natl Acad. Sci. USA 104, 1726–1731 (2007).
98. Addis, D. R., Wong, A. T. & Schacter, D. L. Remembering the past and imagining the future: common and distinct neural substrates during event construction and elaboration. Neuropsychologia 45, 1363–1377 (2007).
99. Szpunar, K. K., Watson, J. M. & McDermott, K. B. Neural substrates of envisioning the future. Proc. Natl Acad. Sci. USA 104, 642–647 (2007).
100. Okuda, J. et al. Thinking of the future and past: the roles of the frontal pole and the medial temporal lobes. Neuroimage 19, 1369–1380 (2003).
101. Botzung, A., Denkova, E. & Manning, L. Experiencing past and future personal events: functional neuroimaging evidence on the neural bases of mental time travel. Brain Cogn. 66, 202–212 (2008).
102. D’Argembeau, A., Xue, G., Lu, Z. L., Van der Linden, M. & Bechara, A. Neural correlates of envisioning emotional events in the near and far future. Neuroimage 40, 398–407 (2008).
103. Hassabis, D., Kumaran, D. & Maguire, E. A. Using imagination to understand the neural basis of episodic memory. J. Neurosci. 27, 14365–14374 (2007).
104. Buckner, R. L. & Carroll, D. C. Self-projection and the brain. Trends Cogn. Sci. 11, 49–57 (2007).
105. Mason, M. F. et al. Wandering minds: the default network and stimulus-independent thought. Science 315, 393–395 (2007).
106. Spreng, R. N., Mar, R. A. & Kim, A. S. The common neural basis of autobiographical memory, prospection,
navigation, theory of mind, and the default mode: a quantitative meta-analysis. J. Cogn. Neurosci. 21, 489–510 (2009).
107. Hassabis, D. & Maguire, E. A. Deconstructing episodic memory with construction. Trends Cogn. Sci. 11, 299–306 (2007).
108. Schacter, D. L. & Addis, D. R. The cognitive neuroscience of constructive memory: remembering the past and imagining the future. Philos. Trans. R. Soc. Lond. B Biol. Sci. 362, 773–786 (2007).
109. Summerfield, J. J., Hassabis, D. & Maguire, E. A. Cortical midline involvement in autobiographical memory. Neuroimage 44, 1188–1200 (2009).In this fMRI study of autobiographical memory, the factors of ‘realness’ and ‘self-relevance’ were manipulated and resulted in a relatively clear distinction between activations in the RSC and those in the neighbouring PCC. This finding highlights the importance of designing fMRI experiments to delineate the specific role of the RSC.
110. Desgranges, B. et al. The neural substrates of episodic memory impairment in Alzheimer’s disease as revealed by FDG-PET: relationship to degree of deterioration. Brain 125, 1116–1124 (2002).
111. Nestor, P. J., Fryer, T. D., Ikeda, M. & Hodges, J. R. Retrosplenial cortex (BA 29/30) hypometabolism in mild cognitive impairment (prodromal Alzheimer’s disease). Eur. J. Neurosci. 18, 2663–2667 (2003).
112. Aggleton, J. P. & Brown, M. W. Episodic memory, amnesia, and the hippocampal-anterior thalamic axis. Behav. Brain Sci. 22, 425–444 (1999).
113. Wagner, A. D., Shannon, B. J., Kahn, I. & Buckner, R. L. Parietal lobe contributions to episodic memory retrieval. Trends Cogn. Sci. 9, 445–453 (2005).
114. Moscovitch, M. et al. Functional neuroanatomy of remote episodic, semantic and spatial memory: a unified account based on multiple trace theory. J. Anat. 207, 35–66 (2005).
115. Squire, L. R. & Bayley, P. J. The neuroscience of remote memory. Curr. Opin. Neurobiol. 17, 185–196 (2007).
116. Levine, B., Svoboda, E., Hay, J. F., Winocur, G. & Moscovitch, M. Aging and autobiographical memory: dissociating episodic from semantic retrieval. Psychol. Aging 17, 677–689 (2002).
117. Maviel, T., Durkin, T. P., Menzaghi, F. & Bontempi, B. Sites of neocortical reorganization critical for remote spatial memory. Science 305, 96–99 (2004).
118. Wesierska, M., Adamska, I. & Malinowska, M. Retrosplenial cortex lesion affected segregation of spatial information in place avoidance task in the rat. Neurobiol. Learn. Mem. 91, 41–49 (2009).
119. Albasser, M. M., Poirier, G. L., Warburton, E. C. & Aggleton, J. P. Hippocampal lesions halve immediate-early gene protein counts in retrosplenial cortex: distal dysfunctions in a spatial memory system. Eur. J. Neurosci. 26, 1254–1266 (2007).
120. Jenkins, T. A., Vann, S. D., Amin, E. & Aggleton, J. P. Anterior thalamic lesions stop immediate early gene activation in selective laminae of the retrosplenial cortex: evidence of covert pathology in rats? Eur. J. Neurosci. 19, 3291–3304 (2004).
121. Aggleton, J. P. EPS Mid-Career Award 2006. Understanding anterograde amnesia: disconnections and hidden lesions. Q. J. Exp. Psychol. (Colchester) 61, 1441–1471 (2008).
122. Vann, S. D. & Albasser, M. M. Hippocampal, retrosplenial, and prefrontal hypoactivity in a model of diencephalic amnesia: evidence towards an interdependent subcortical-cortical memory network. Hippocampus 11 Mar 2009 (doi:10.1002/hipo.20574).
123. Poirier, G. L. et al. Anterior thalamic lesions produce chronic and profuse transcriptional de-regulation in retrosplenial cortex: a model of retrosplenial hypoactivity and covert pathology. Thal. Relat. Syst. 4, 59–77 (2008).
124. Poirier, G. L. & Aggleton, J. P. Post-surgical interval and lesion location within the limbic thalamus determine extent of retrosplenial cortex immediate-early gene hypoactivity. Neuroscience 160, 452–469 (2009).
125. Garden, D. L. et al. Anterior thalamic lesions stop synaptic plasticity in retrosplenial cortex slices: expanding the pathology of diencephalic amnesia. Brain 132, 1847–1857 (2009).Slice recording study showing the loss of a form of synaptic plasticity in the RSC following anterior thalamic lesions. The results are striking as the loss
of plasticity is due not to a failure to stimulate the target neurons in the RSC but rather to a local change in the properties of these RSC neurons following deafferentation.
126. Mitelman, S. A., Shihabuddin, L., Brickman, A. M., Hazlett, E. A. & Buchsbaum, M. S. Volume of the cingulate and outcome in schizophrenia. Schizophr. Res. 72, 91–108 (2005).
127. Nugent, A. C. et al. Cortical abnormalities in bipolar disorder investigated with MRI and voxel-based morphometry. Neuroimage 30, 485–497 (2006).
128. Liberzon, I. et al. Brain activation in PTSD in response to trauma-related stimuli. Biol. Psychiatry 45, 817–826 (1999).
129. Temple, E. et al. Neural deficits in children with dyslexia ameliorated by behavioral remediation: evidence from functional MRI. Proc. Natl Acad. Sci. USA 100, 2860–2865 (2003).
130. Fazio, F. et al. Metabolic impairment in human amnesia: a PET study of memory networks. J. Cereb. Blood Flow Metab. 12, 353–358 (1992).
131. Joyce, E. M. et al. Decreased cingulate and precuneate glucose utilization in alcoholic Korsakoff’s syndrome. Psychiatry Res. 54, 225–239 (1994).
132. Minoshima, S. et al. Metabolic reduction in the posterior cingulate cortex in very early Alzheimer’s disease. Ann. Neurol. 42, 85–94 (1997).
133. Villain, N. et al. Relationships between hippocampal atrophy, white matter disruption, and gray matter hypometabolism in Alzheimer’s disease. J. Neurosci. 28, 6174–6181 (2008).
134. Scahill, R. I., Schott, J. M., Stevens, J. M., Rossor, M. N. & Fox, N. C. Mapping the evolution of regional atrophy in Alzheimer’s disease: unbiased analysis of fluid-registered serial MRI. Proc. Natl Acad. Sci. USA 99, 4703–4707 (2002).
135. Pengas, G., Hodges, J. R., Watson, P. & Nestor, P. J. Focal posterior cingulate atrophy in incipient Alzheimer’s disease. Neurobiol. Aging 1 May 2008 (doi:10.1016/j.neurobiolaging.2008.03.014).
136. Buckner, R. L., Andrews-Hanna, J. R. & Schacter, D. L. The brain’s default network: anatomy, function, and relevance to disease. Ann. NY Acad. Sci. 1124, 1–38 (2008).
137. Raichle, M. E. et al. A default mode of brain function. Proc. Natl Acad. Sci. USA 98, 676–682 (2001).
138. Biswal, B., Yetkin, F. Z., Haughton, V. M. & Hyde, J. S. Functional connectivity in the motor cortex of resting human brain using echo-planar MRI. Magn. Reson. Med. 34, 537–541 (1995).
139. Uddin, L. Q., Clare Kelly, A. M., Biswal, B. B., Xavier Castellanos, F. & Milham, M. P. Functional connectivity of default mode network components: correlation, anticorrelation, and causality. Hum. Brain Mapp. 30, 625–637 (2009).
140. Gao, W. et al. Evidence on the emergence of the brain’s default network from 2-week-old to 2-year-old healthy pediatric subjects. Proc. Natl Acad. Sci. USA 106, 6790–6795 (2009).
141. Greicius, M. D., Supekar, K., Menon, V. & Dougherty, R. F. Resting-state functional connectivity reflects structural connectivity in the default mode network. Cereb. Cortex 19, 72–78 (2009).
142. Zilles, K. & Palomero-Gallagher, N. Cyto-, myelo-, and receptor architectonics of the human parietal cortex. Neuroimage 14, S8–S20 (2001).
AcknowledgementsThe authors would like to thank A. Perrotin, N. Burgess, M. Greicius and L. Woods. E.A.M. is supported by the Wellcome Trust. S.D.V. is supported by a Biotechnology and Biological Sciences Research Council (BBSRC) David Phillips Research Fellowship. S.D.V. and J.P.A.’s contributions have also been supported by a BBSRC grant (BB/D002001/1).
DATABASESEntrez Gene: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=geneEgr1 | Fos
FURTHER INFORMATIONSeralynne D. Vann’s homepage: http://www.cardiff.ac.uk/psych/vannJohn P. Aggleton’s homepage: http://www.cardiff.ac.uk/psych/aggletonEleanor A. Maguire’s homepage: http://www.fil.ion.ucl.ac.uk/Maguire
All liNks ARe AcTive iN The oNliNe pdf
R E V I E W S
802 | noVEMBER 2009 | VoLuME 10 www.nature.com/reviews/neuro
© 2009 Macmillan Publishers Limited. All rights reserved

ToC Blurb
000 What does the retrosplenial cortex do?Seralynne D. Vann, John P. Aggleton and Eleanor A. Maguire
Vann and colleagues review anatomical, lesion and imaging studies suggesting that the retrosplenial cortex is involved in navigation and memory. They propose that it achieves this function through a key role in scene construction by translating between different perspectives of the environment.
At a glance•The main connectional properties of the retrosplenial cortex (RSC)
(Brodmann areas 29 and 30) are conserved across mammalian spe-cies. Key features include reciprocal links with the hippocampal formation, the parahippocampal region, the limbic thalamus and the parietal cortex.
• Functional studies of the RSC of rodents consistently point to a role in learning and navigation. These roles are thought to be acted out in concert with the hippocampal formation and the limbic thalamus.
•neuropsychological studies of the human RSC remain problematic owing to difficulties in isolating this area. nevertheless, studies of patients with unilateral retrosplenial damage suggest an important contribution to navigation, whereas bilateral retrosplenial damage is often associated with anterograde and varying degrees of retro-grade amnesia.
•There is consistent evidence that the RSC suffers very early patho-logical changes in the progression of mild cognitive impairment and Alzheimer’s disease. These pathological changes are reflected by a decrease in metabolic activity.
• Functional neuroimaging studies of healthy volunteers suggest broad involvement of the RSC in an array of cognitive abilities, with an almost ubiquitous engagement by tests of spatial naviga-tion and episodic (autobiographical) memory.
•These patterns of retrosplenial activity might relate to a key role in scene construction — the process of mentally generating and manipulating a complex or coherent scene. This process may underpin functions such as autobiographical memory, navigation and thinking about the future.
•one specific hypothesis relating to the retrosplenial cortex is that it translates between different perspectives of the external world, namely viewpoint-dependent and viewpoint-independent frames of reference.
BiosSeralynne Vann obtained her B.Sc. in experimental psychology from the university of Sussex, uK, and was subsequently awarded her Ph.D. in behavioural neuroscience from Cardiff university, uK. Her research focuses on the neural basis of learning and memory and on how the limbic midbrain, medial diencephalon, medial temporal lobe and related cortical structures interact to support memory. Key interests are the mammillary bodies, and her research on this struc-ture is currently supported by a fellowship from the Biotechnology and Biological Sciences Research Council, uK.
John Patrick Aggleton has been Professor of Cognitive neuroscience at Cardiff university since 1994. Prior to that he was a Lecturer and Senior Lecturer at Durham university and before that a Fellow at the Laboratory of neuropsychology at the uS national
Institutes of Health (Bethesda, Maryland). Through a combina-tion of anatomical, behavioural and clinical investigations he has studied the brain systems that support episodic memory in humans and spatial memory in rodents. His particular focus is on the functional interactions between the temporal lobe and the medial diencephalon.
Eleanor A. Maguire is a Wellcome Trust Senior Research Fellow and Professor of Cognitive neuroscience at the Wellcome Trust Centre for neuroimaging, university College London, uK. She heads the Memory and Space research laboratory at the Centre, where her team seeks to understand how spatial and episodic memories are formed, represented and recollected by the human brain.
o n l I n E o n ly
© 2009 Macmillan Publishers Limited. All rights reserved