veränderungen der intestinalen phosphor-absorption und -exkretion im verlauf der gravidität und...
TRANSCRIPT

W r k u n g der Verfiittcrung ,,Dicnabol" an Mastkiikrn 75
des Kukeils gepriift. Die Ergebnisse deuten auf eine durch die Zulage bewirkte Ver- besserung des Zuwachses hin, die jcdoch von der Dosierung des Wirkstoffes und von der Eiweiflversorgung der Tiere abhangig sein durfie. Rezuglich Futterverwertung und Stickstoff bilanz wurden keine eindeutigen Unterschiede zwischen den Zulage- gruppen und den entsprechenden Kontrollgruppen beobachtet.
Summary
In two experiments with chicks additions to the feed of 1 to 5 nig per kg of a synthetic androgen ,,Dianabol" have been studied. The results suggest a n iniprove- nient of weight increase due to the additive which, however, seem to depend on its level and on the protein level of the diet. Feed cfficiency and nitrogen balance did not show substantial differences between the groups receiving the androgen, and the controls.
Literatur
DESAUI.I.ES, P. A., Ch. KHAHENBUHL, W. SCHULER et H. J. BEIN (1959). Schweiz. Medizinische Vodlcnsdirift 89, 1313-1318. - KOCMAKIAN, C. D., and J. R. MURLIN ((1935). J. Nutrition 10. 437.
Aus den2 Institut fiir Tierphysiologic und Tierernuhrung der Universitut Gottingen Direktor: Prof. Dr. DY. \V. Lenkcit
Veranderungen der intestinalen Phosphor-Absorption und -Exkretion im Verlauf der Graviditat und Laktation des
Schweines bei konstanter Ernahrung
Von J. 0. GUTTE, H. J. LANTZSCH, S . MOLNAR und W. LENKEIT
In fruhereii Untersuchungen an Sauen wurde bei gleichbleibender Ernahrung wah- rend der Graviditat ein Abfall der Ca- und P-Ausscheidungen im K o t von der Mitte bis zum Ende der Graviditat beobachtet (LENKEIT, GUTTE, KIRCHHOFF und SOEHN- GEN 1959 b; FARRIES 1957; SOEHNGEN 1956; KIRCHHOFF 1955).
Wahrend der Laktation .war bei gleichbleibender Ernahrung ebenfalls ein regel- mat3iger Abfall der Ca- und P-Ausscheidungen in1 Kot festzustellen (LENKEIT et al. 1959 b ; LENKEIT 1959; SOEHNGEN 1956; KIRCHHOFF 1955; BEINLICH 1953).
Bei zwei Sauen, die vom Ende der Graviditat bis zum Ende der Laktation gleich- mat3ig gefuttert wurden, lagen die Cn- und P-Ausscheidungen im Kot im Durchschnitt der ersten zehn Tage nach der Geburt leicht iiber dem Niveau der letzten Graviditats-
76 J. 0. Gutte, H. /. Lantzscb, S . MolnLr und W . Lenkeit
tage, um danach bis zum Ende der Laktation abzufallen (LENXEIT et al. 1959 b; KIRCHHOFF 1955).
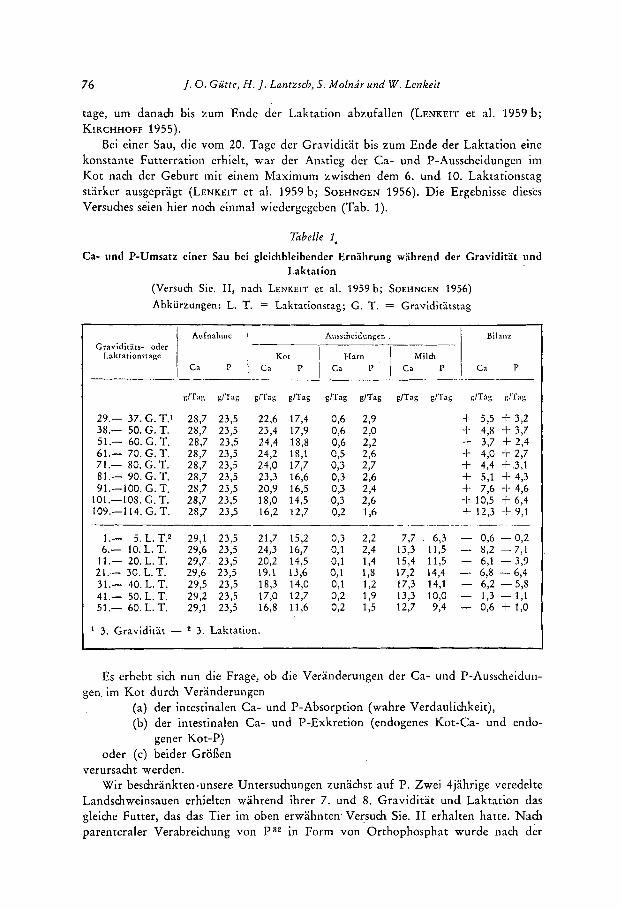
Bei einer Sau, die vom 20. Tage der Graviditat bis zum Ende der Laktation eine konstante Futterration erhielt, war der Anstieg der Ca- und P-Ausscheidungen im Kot nach der Geburt mit eineni Maximum zwischen dem 6. und 10. Laktationstag starker ausgepragt (LENKEIT et al. 1959 b; SOEHNGEN 1956). Die Ergebnisse dieses Versuches seien hier noch einmal wiedergegeben (Tab. 1).
Tabelle 1.
Ca- und P-Umsatz einer Sau bei gleichbleibender Ernlhrung wlhrend der Graviditzt und Laktation
(Versuch Sie. 11, nach LENKEIT et al. 1959 b; SOEHNCEN 1956) Abkiirzungen: L. T. = Laktationstag; G. T . = Graviditatstag
Aufnahme
ca r
Graviditiits- oder Lakrationstage
Aossdkdungen . Bilanz
Kor I Ham cn P I ca P
29.- 37. G. T.' 38.- 50 .G.T. 51.- 60. G. T. 61.- 70.G.T. 71.- 80. G. T. 81.- 90.G.T. 91.-100. G. T.
101.--108. G. T. 109.-114. G.T.
g/Ta::
28,7 28,7 28,7 28,7 28,7 28,7 28,7 28,7 28,7
g/Tag g / T q g/Tag g/Tag
23,s 22,6 17,4 0,6 23,5 23,4 17,9 0,6 23,5 24,4 18,s 0,6 23,5 24,2 18,l 0,5 23,5 24,O 17,7 0,3 23,5 23,3 16,6 0,3 23,5 20,9 16,5 0,3 23,5 18,O 14,5 0,3 23,5 16,2 12,7 0,2
1.- 5. L. T.' 6.- 10. L. T.
11.- 20. L. T. 21:- 30. L. T. 31.- 40. L. T. 41.- 50. L. T. 51.- 60. L. T.
3. Graviditat -
29,l 23,5 21,7 15,2 0,3 29,6 23,5 24,3 16,7 0,l 29,7 23,5 20,2 14,5 0,l 29,6 23,5 19,l 13,6 0,l 29,5 23,5 18,3 14,O 0,l 29,2 23,5 17,O 12,7 0,2 29,l 23,5 16,8 11,6 0,2
2 3. Laktation.
7,7 . 6,3 13,3 11,5 15,4 11,5 17,2 14,4 17,3 14,l 13,3 10,O 12,7 9,4
g/Tag g/Tag
4 5,5 4 3,2 4 4,8 f 3,7 + 3,7 f 2,4 4 4,O 4 2,7 4- 4,4 4- 3,l 4 5,l 4 4,3 + 7,6 f 4,6 4- 10,5 + 6,4 f 12,3 -t 9,l
- 0,6 - 0,2 - 8,2 - 7,l - 6,l - 3,9 - 6,s - 6,4 - 6,2 - 5,s - 1,3 - 1,l - 0,6 f 1,0
Es erhebt sich nun die Frage, ob die Veranderungen der Ca- und P-Ausscheidun- gen in1 Kot durch Veranderungen
(a) der intestinalen Ca- und P-Absorption (wahre Verdaulichkeit), (b) der intestinalen Ca- und P-Exkretion (endogenes Kot-Ca- und endo-
gener Kot-P) oder (c) beider Groflen
verursacht werden. Wir beschrankten-unsere Untersuchungen zunachst auf P. Zwei 4jahrige veredelte
Landschweinsauen erhielten wahrend ihrer 7. und 8. Graviditat und Laktation das gleiche Futter, das das Tier im oben erwahnten Versuch Sie. I1 erhalten hatte. Nach parenteraler Verabreichung von P32 in Form von Orthophosphat wurde nach der

Veranderungen der intestinalen Phosphor-Absorption und -Exkretion 77
Methode von HEVESY, HAHN und REBBE (1939) bestimnit, welche Anteile des Kot-P endogenen und exogenen Ursprungs sind. Daraus lat3t sich die wahre Verdaulichkeit des Futter-P berechnen.
Definition der gebrauchten Begriffe
D a einige Begriffe, wie zum Beispiel ,Resorption", in der Literatur in verschiedener Bedeutung gebraucht werden, seieii die in dieser Arbeit gebraudxen Begriffe folgen- dermat3en definiert: (a) endogener Kot-P derjenige Anteil des Kot-P, der aus dem Organismus des
Tieres stammt (b) exogener Kot-P = derjenige Anteil des Kot-P, der aus dem Futter stammt (c) absorbierter P = Futter-P - exogener Kot-P
=
100 010. absorbierter-P
Futter-P (d) wahre Verdaulichkeit des Futter-P =
Den Begriff ,,Resorption" gebrauchen wir nur im Sinne von ,,Knochen-Resorp- 1:ion". ,,Resorbierter P" bezeichnet also den P, der aus dem Skelett herausgelost wird. Die Aufnahme von P aus dem Verdauungskanal wird dagegen ,,P-Absorption" (,,in- testinale P-Absorption") genannt. Zur Bezeichnung der Einlagerung von P in das Skelett verwendet man den Begriff ,,P-Apposition''
Methoden
Versuchstiere
Im Januar 1960 kauflen wir aus einer Hochzuchtherde veredelter Landschweine zwei 3'/ejihrige Wurfgeschwister. Die Tiere hatten bis dahin 6 Wurfe aufgezogen, immer das gleiche Futter bekommen und standen am Anfang ihrer 7. Gravlditat.
Die Sauen ferkelten am 24. 3. und 25. 3. 1960. Das Tier Farah (F) warf 11 lebende und 2 tote Ferkel und zog 11 Ferkel auf, das. Tier Soraya (S) warf 13 lebende und zog 13 auf. Die Ferkel erhielten bis zum 28. Lebenstage kein Beifutter, das 4-Wochen-Wurfgewicht betrug beim Tier F 74 kg, bei Tier S 82 kg.
- h i Ende der 7. Laktation (19. bis 26. 5 . 1960) wurde ein Probeversuch durch- gefuhrt, uni die Tiere an die Versuchsbedingungen zu gewohnen (Versuchsperioden F 1 und S 1).
Am 26. 5. 1960 wurden die Ferkel abgesetzt. Beide Sauen wurden d a m zweimal, am 31. 5. 1960 und 1. 6. 1960, erfolgreich belegt. Die Versuche zur Ermittlung der wahren Verdaulichkeit wurden in der nun folgenden 8. Gravidi t i t u n d 8. Laktation durchgefuhrt.
'
Futter und Fiitterung
Die Tiere erhielten vom Januar 1960 bis zum Ende der Versuche im November 1960 die in Tab. 2 angegebene Tagesration.

78 1 . 0. Giitte, H . J . Lantzsch, S . Molna'r und W. Lenkeit
Tubelle 2
Futter
Die Tugesrution bestand aus 3050 g Hafcrsclirot 300 g Fischinehl 450 g Waldhof-Futtcrhefc 200 g Luzernc-Gruninclil
I g Roviniix 1/401
4001 g lufitrodrcnes Futter insgesaint 9 bis 18 Liter Trankwasscr
Die Tagesration enthielt Trodrensubsranz . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3473 g
. . . . . . . . . . . . . . . . . . . . . 3229 g
. . . . . . . . . . . . 149 g Rohfaser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N-frcie Exrrakcstoffc . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Gcsaintn~hrsroff . . . P . . . . . . . . . . . . . . . . : . . . . . . . . .
Ca in1 Trinkwasser . . . .
1 Mineralstabiles Vitamin A + D:< Konzentrat der Deutsche Hoffmann-La Roche AG. (Grenzach, Uadcn). Angegebencr Gchalt: 40 000 I. E. Vit. A -I- 8000 I. E. Vit. D3/g.
Diese deckt wahrend der Laktation in ihrem Gehalt an Rohprotein, Energie, Ca und P nur den Bedarf fur mittlere Milchleistung.
Die einzelnen Futtermittel der Ration wurden mit einer Genauigkeit von ? 0,2 O/o
jeweils an einem Tage fur alle Versuchsperioden einschliei3lich der Vorfutterungszeit eingewogen und in Plastikbeuteln aufbewahrt. Dadurch war eine vollig gleiche Futte- rung in allen Versuchsperioden garantiert. Der Zusatz des Vitaminpraparates zum Futter erfolgte taglich. Wahrend des Einwiegens wurden von jedem Futtermittel zwei Proben fur die memische Analyse entnomrnen und von jeder Probe 2 bis 4 Analysen der wesentlichen' Bestandteile durchgefuhrt.
Vor und zwischen den Versuchsperioden erhielten die Tiere das gleiche Futter wie in den Versuchsperioden. Die Einwaage des Futters fur diese Zeit wurde allerdings an anderen Tagen vorgenommen, so dai3 sich Abweichungen im Trockensubstanz- gehalt der einzelnen Komponenten ergeben.
I n der Vorfutterungszeit (5 oder mehr Tage vor Beginn der Samnielperiode) und in den Sammelperioden wurde den Tieren 3mal taglich in genau Sstundigen Inter- vallcn (3.00, 11.00 und 19.00 Uhr) genau '/a der Tagesration gefuttert. Nach Zusatz abgemessener Wassermengen Zuni Futter erhielten die Tiere Trankwasser ad libitum. Die aufgenommenen Wassermengen wurden registriert ; vom Trankwasser wurden Proben zur Mineralstoffanalyse entnommen. In der Zeit zwischen den Versuchs- perioden erhielten die Tiere nur zweimal taglich Futter und Wasser.
Das Tier F nahm die Versuchsration immer vollstandig auf, auch am Tage des Abferkelns. Tier S liefl voni Tage der Geburt bis zum 3 . Laktationstage Futterreste

Verunderungen der intestinalen Phosphor-Absorption und -Exkretion 79
zuriick, die zuruckgewogen und analysiert wurden. Es hatte an diesen 4 Tagcn tag- lich etwa 2/s der verabreichten Tagesration aufgenommen. Vom 4. Laktationstage ab war die Aufnahme der Tagesration bei Tier S wieder vollstandig. Bereits am 5. Lak- tationstage lagen die Kot-P-Ausscheidungen auf der Hohe der folgenden Tage.
Haltung der Tiere, Sammlung von Kot und Harn
Die Tiere wurden wahrend der Versuchsperioden in 2 X 2 m grogen Stallboxen ge- halten, in denen sich keine Streu befand. In der Zeit vor den Versuchen waren sie daran gewohnt worden, die Exkrenlente nach Auslosung eines bedingten Reflexes augerhalb der Stallboxen abzugeben, so daa Kot und Harn sauber getrennt in Sam- melgefal3en aufgefangen werden konnten (LENKEIT und GUTTE 1955). Es gelang in allen Versuchen, die Tiere genau am Ende eines Versuchstages (24.00 Uhr) zur Kot- und Harnabgabe zu bewegen. Die an einem Tag gesammelten Exkremente enthalten daher die Ausscheidungen von genau 24 Stunden, was fur die Auswertung der Resul- tate auflerordentlich gunstig ist.
Die Stalltemperatur wurde durch elektrische Heizung in allen Versuchen zwischen 15 ' und 18 O C gehalten.
Zwischen den Versuchsperioden erhielten die Tiere auch Streu (Stroh) und bei trockener Witterung Auslauf ins Frcie.
Saugungen, Bestimmung der Milchleistung, Probenahme der Milch zur Analyse
Die Ferkel wurden sofort nach der Geburt von der Mutter getrennt und taglich 20- bis 26mal zum Saugen angesetzt. Bei dieser Frequenz blieb bei beiden Tieren taglich bei 1 bis 5 Saugungen die Milchejektion aus.
Die Milchleistung wurde an allen Versuchstagen wie in den fruheren Versuchen bestinimt (GUTTE und LENKEIT 1960; LENKEIT und GUTTE 1955):
Der gesamte Wurf wurde auf einer Tacho-Waage mit einem Fassungsvermogen von 60 kg und einer Ablesegenauigkeit von k 10 g vor und nach jeder Saugung ge- wogen. Die Speichel- und Transpirationsverluste der Ferkel wahrend des Saugens wurden bei den Saugungen ohne Milchejektion festgestellt (taglich 1- bis '5mal in jedeni Wurf) und Lei der Berechnung der Milchmengen berucksichtigt. Kot- und Harnverluste der Ferkel wahrend des Saugens traten nicht auf.
Die Probenahme der Milch zur Analyse wurde ebenfalls wie in den fruheren Ver- suchen durchgefuhrt (GUTTE und LENKEIT 1960; LENKEIT und GUTTE 1955).
Beide Sauen wurden am Anfang der Laktation von 11, am Ende der Laktation von 9 Ferkeln besiiugt.
Bestimmung des P-Gehaltes in Futter, Kot, H a m und Milch
Die Substanzen wurden trocken verascht und der P-Gehalt in den Aschenlosungen nach einer von GERICKE und KURMIES (1952/53) uberarbeiteten Methode (KURMIES 1955; KITSON und MELLON 1944; KOENIG und JOHNSON 1942) bestimmt (als Mo- lybdivanadophosphorsaure in salpetersaurer Losung durch kolorimetrischen Vergleich

80 .J . 0. Giitte, H. /. Lantzsch, S. Molncir irnd W . Lenkeit
Versudx- Versuchs- periode
mit Losungen bekannten P-Gehaltes). Kot und H a r n wurden taglich analysiert, von der Milch kamen Durchschnittsproben von 1 oder 2 Tagen zur Analyse. Einzelheiten wurden von LANTZSCH (1961) beschrieben.
K'rpergewi*x iniizierre Tag der Dauer dcr ZUr der 1 Dosir I Inicktion 1 Versuchsperiode
lnjektion
Injiziertes Isotop
Pa? wurdc als Orthophosphat in isotonischer Kochsalzlosung mit einem pH von 7 vom Radiochemical Centre, Amersham, England, bezogen. Das Volumen der inji- zierten Losungen variierte von 1 bis 8 ml.
Injektion des lsotops und Dauer der Sammelperioden
Das Isotop wurde mit einer 3 cm langen Injektionsnadel in die Keule, etwa 15 cm unterhalb des Sitzbeinhodcers, injiziert. Die Injektion'wurde auflerhalb des Vcrsuchs- stalles durchgefuhrt, um eine Verunreinigung mit Spuren Lon P32 zu vermeidcn. Eine Verunreinigung der Versuchsstalle kann weiterhin dadurch entstehen, daR nach der Injektion einige Tropfchen der injizierten hochaktiven Substanz aus der Injcktions- wunde austreten. Diese wurde daher sofort nach der Injektion mit einer dicken Lage von Verb.andmul1 abgedeckt und mit Heftpflaster verklebt. Der Verband wurde erst
Tabelle 3
Injizierte Dosis von P3* und Ubersicht iiber die Versuchsperioden (Abkurzungen: G. T. = Graviditatstag; L. T. = Laktationstag)
F 2 F 3 F 43 F 4b F 4c F 4d F 5a F 5Ll
s3 S 4a S 4b s 4c S 4d S 5a S 5b
F F F F F F F F
S S S S S S S
kg
201 207 247
191
197 233
176
mc I"?.
1,s 32. G. T. 1,3 49. G. T. 999 98. G. T.
1 3 49. G. T. 999 98. G. T.
3,o 43. L. T.
36.- 42. G. T. 54.- 60. G. T.
103.-108. G. T. 109.--117. G. T.
11.- 15 .L .T . 3.- ~ o . L . T .
53.- 57.L.T.
54.- 60. G. T. 103.-108. G. T.
48.- 52 .L .T.
109.--115. G. T .
11.- 15 .L .T. 5.- ~ o . L . T .
48.- 52.,L. T. 53.- 57. L. T.
nach Beendigung der Sammelperiode abgenommen. E r enthielt stew radioaktive Sub- stanz. Die injizierten Dosen und die Dauer der Perioden, in denen exogener und cndogcner Kot-P bestimmt wurden, sind in Tabelle 3 angegeben.

Veranderungen der intestinalen Phosphor-Absorption und -Exkretion 81
Bestimmung der spezifischen Aktivitat
Die P32-Aktivitat wurde in denselben Aschenlosungen, aus denen der P-Gehalt be- stimmt wurde, mit einem Becher-F1iissigkeits-Geiger-Muller-Zahlrohr der Fa. Siemens & Halske AG. (Typ BFZ-5) gemessen. Die Impulsrate wurde mit dem groi3en Strah- lungsmeflger5t der Fa. Siemens & Halske AG. (Typ GS) registriert.
Bei dieser Mei3anordnung ist die Impulsrate proportional der P32-Konzentration in der gemesscnen Losung.
Durch Messung der Impulsrate von Losungen mit bekanntem Gehalt a n P32 wurde die Meflanlage geeicht und taglich auf ordnungsgemage Funktion iiberpriifk Aus der Impulsrate der analysierten Losungen lafit sich dann durch Multiplikation mit dem Eichfaktor deren P3z-Konzentration errechnen, we!che in p c PSz/l angegeben wird. Diese mui3 um den natiirlichen Zerfall von Psz korrigiert werden.
Durch Division der Konzentration von P3z durch die Konzentration von P er- halt man die spezifische Aktivitat:
" __- ''"* - - - " = spezifische Aktivitat. g PI1 g p
Die Einwaage der zu veraschenden Substanz wurde so groi3 gewahlt, dai3 die P3*- Alctivitft der Aschenlosungen im optimalen MeDbereich des Gerates lag (500 bis 14 000 Impulse pro Minute [I. p. M.] bei einem Null-Effekt von etwa 30 I. p. M.). Die spezifische Aktivitat des Kot-P und des Harn-P wurde taglich in zwei Parallel- proben bestimmt. Von jeder Einzelprobe wurden insgesamt wenigstens 30 000 Tm- pulse gezahlt, um den statistischen Fehler moglichst niedrig zu halten.
Berechnung des endogenen und exogenen Kot-P
Die Methode von HEVESY, HAHN und REBBE (1939) basiert auf der Voraussetzung, dai3 einige Zeit nach parenteraler Verabreichung von P3z die spezifische Aktivitat des P, der vom Organismus in den Verdauungstrakt abgegeben wird, ebenso groi3 ist wie die spezifische Aktivitat des P, der vom Organismus in den H a r n abgegeben wird. I<JERuLF-JENSEN konnte 1941 demonstrieren, d a 8 diese Voraussetzung erfullt ist: Bei 4 Ratren, die eine P-freie Dift erhielren, waren vom 6. bis 13. Tage nach der Injek- tion von die spezifischen Aktivitaten von Kot-P und Harn-P gleich.
Aus dem Verhaltnis der spezifischen Aktivitaten von Kot-P und Harn-P lafit sich folglidi errechnen, uni welchen Prozentsatz der radioaktive endogene Kot-P mit in- aktivem P aus den1 Futter verdiinnt worden ist:
spcz. Akt. des Kot-P s p a . Akt. des Ham-P (1) endogener Kot-P = Gesamt-Kot-P
Dabei mui3 allerdings beriicksichtigt werden, dai3 von der Abgabe des P aus dem Organismus in den Verdauungstrakt bis zur Ausscheidung des P mit dem Kot ein Zeitintervall besteht, das, bei verschiedenen Tieren unterschiedlich sein kann. Wir be- stimmten diese Ausscheidungsverzogerung im Versuch F 2 in der ublichen Weise:
Die spezifische Aktivitat des Harn-P wurde nach der Injektion von P S Z zunachst in allen einzelnen Harnportionen, spater im Sammelharn von 8 Stunden, und vom 4. Tage nach der Injektion an im Sammelharn von 24 Stunden gemessen. Im Kot
82 I . 0. Giitte, H. 1. Lantzsch, S. Molnrir und W. Lenkeit
wurden die Messungen bis zum 3. Tag nach der Injektion in Sammelproben von 8 Stunden, spater von 24 Stunden ausgefiihrt.
220
200
I60
cl 160
$
i 4 I 4 0
120
I00
80
60
40
20
- sprr . Ah1 des H a m - P
w e, .. - d o l - p I I o 2
8 16 24 32 40 4 8 56 64 7 2 5 1 d . 4 6 7 8 9 10 Tagc
ZElT NACH DER I N J E K T I O N
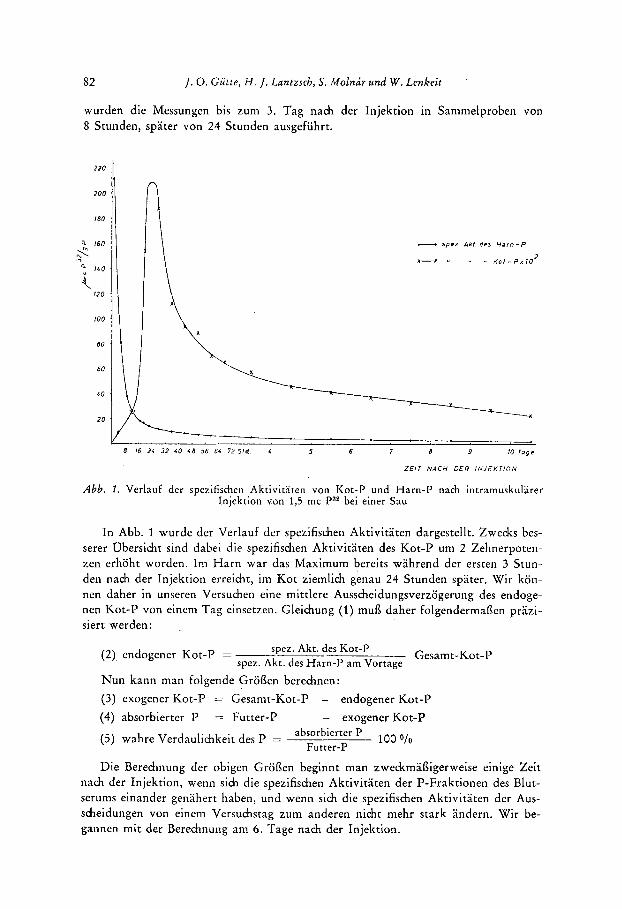
Abb. 1. Verlauf der spezifischen Aktivitaten von Kot-P und Harn-P nach intramuskularer Injektion von 1,5 mc Pa2 bei einer Sau
In Abb. 1 wurde der Verlauf der spezifischen Aktivitaten dargestellt. Zwecks bes- serer Obersicht sind dabei die spezifishen Aktivitaten des Kot-P um 2 Zehnerpoten- Zen erhoht worden. Im H a r n war das Maximum bereits wahrend der ersten 3 Stun- den nach der Injektion erreicht, im Kot ziemlich genau 24 Stunden spater. Wir kon- nen daher in unseren Versuchen eine mittlere Ausscheidungsverzogerung des endoge- nen Kot-P von einem Tag einsetzen. Gleidung (1) mufi daher folgendermafien prazi- siert werden: ,
spez. Akt. des Kot-P spez. Akt . des H a m - P a m Vortage
(2) endogener Kot-P = ~ _ _ _ Gesam t-Kot-P
Nun kann man folgende Grofien berechnen: (3) exogener Kot-P = Gesamt-Kot-P - endogener Kot-P (4) absorbierter P = Futter-P - exogener Kot-P
( 5 ) wahre Verdaulichkeit des P =
Die Berechnung der obigen Grofien beginnt man zweckmafiigerweise einige Zeit nach der Injektion, wenn sich die spezifischen Aktivitaten der P-Fraktionen des Blut- serums einander genahert haben, und wenn sich die spezifischen Aktivitaten der Aus- scheidungen von einem Versuchstag zum anderen nicht mehr stark andern. Wir be- gannen mit der Berechnung am 6 . Tage nach der Injektion.
100 010 absorbierter P
Futter-P

Veranderungen der intestinalen Phosphor-Absorption und -Exkretion 83
Statistische Oberprufung der Resultate
Die statistische Oberprufung der Resultate wurde nach der Varianzanalyse mit dop- pelter Gruppicrung dcr Beobachtungswcrte ausgefuhrr (MUNZNER 1952, SNEDECOR 1957, p. 296). Das Auswertungsschema hat folgendes Aussehen:
Bl
Die A-Gruppen bezeichnen die einzelnen Graviditgts- und Laktationsstadien, die B-Gruppen die beiden Tiere F und S.
Mit diesem Auswertungsschenia kann man prufen, ob die Unterschiede der Resul- tate zwischen den einzelnen Graviditats- und Laktationsperioden und der Unter- schied der l<esultate zwischen den Tieren F und S statistisch gesichert sind.
Die kritischen Punkte der F-Verteilung wurden fur eine Zufallswahrscheinlichkeit P = 0,05, 0,01 und 0,001 einer Tabelle von MUNZNER (1952), fur eine Zufallswahr- scheinlichkeit P = 0,25, 0,10, 0,025 und 0,005 einer Tabelle von SNEDECOR (1957, p. 276-279) entnommen.
Resultate und Diskussion
Verlauf der Versuche
Die Graviditat verlief normal. Beide Tiere begannen am 23. 9. 1960 spat abends zu ferkeln. Bein1 Tier S wurde, wie oft bei alteren Tieren, Wehenschwache beobachtet. Die Geburt wurde daher durch Eingriffe unterstutzt. Kurz nach Mitternacht waren alle 12 Ferkel lebend geboren, die Nachgeburt ging innerhalb der nachsten 12 Stun- den vollstandig ab. Das Tier S hatte die Abendmahlzeit am 23. 9. 1960 ( ' /a der Tagesration) nicht aufgenommen und verzehrte, wie oben erwahnt, an den folgenden drei Tagen cbenfalls nur */a der Tagesration. Die Korperremperatur des Times war immer normal.
Das Tier F gebar bis kurz nach Mitternacht 10 lebende und 1 totes Ferkel ohne Unterstutzung, und die Geburt schien beendet. 36 Stunden spater wurden jedoch noch 2 atrophierte Foeten und 2 vollentwickelte tote Ferkel mit dem Rest der Nachgeburt ausgestofien. Die Milchmengen waren bis zu diesem Zeitpunkt abnorm niedrig, die Milchejektion war nur schwach und blieb bei fast jeder zweiten Saugung ganzlich aus. Nach Ahgabe der toten Ferkel und der Nnchgeburt waren Milchproduktion und Milchejektion normal. Das Tier hat auch an diesen Tagen die Futterration immer
84 J. 0. Giitte, H . J. Lantzsch, S . M o l n h wnd W. Lenkeit
1 Ver- Graviditars- Anfangs- Mittleres P-Gehalt suhs- 1 oder Laktationstag I gewicht I Endgewihr I KGrpergewiht 1 leistung ! der Milch periode
vollstandig aufgenommen, die Korpertemperatur war stets normal. Die Zeit vom Anfang bis zum Ende der Geburt (115. bis 117. Graviditatstag) haben wir beim Tier F zur letzten Graviditatsperiode gerechnet. Die Laktationstage wurden vom Be- ginn der Milchproduktion an gezahlt (116. Graviditatstag). 116. und 117. Gravidi- tatstag entsprechen also dem I. und 2. Laktationstag.
Nach der Geburt wurde sorgfaltig darauf geachtet, daB die Tiere weder Nach- geburt no& Lochialflussigkeit aufnahmen. Die Versudxstalle wurden haufig aus- gewaschen. Denn perorale Aufnahme von Substanzen, die P3* enhalten, wurde die Versuchsergebnisse ganzlich verfalschen.
Da beide Sauen nur 11 leistungsfahige Zitzen hatten (die restlichen waren durch EuBere Verletzungen oder fruhere Entzundungen vollig oder teilweise zerstort, wie dies bei alteren Sauen haufig zu beobachten ist), wurde ein Ferkel vom Tier S dem Tier F angelegt. Am Anfang der Lakiation wurden somit beide Sauen von 11 Fer- keln besaugt, am Ende der Laktation aus versuchstechnischen Grunden nur noch von 9.
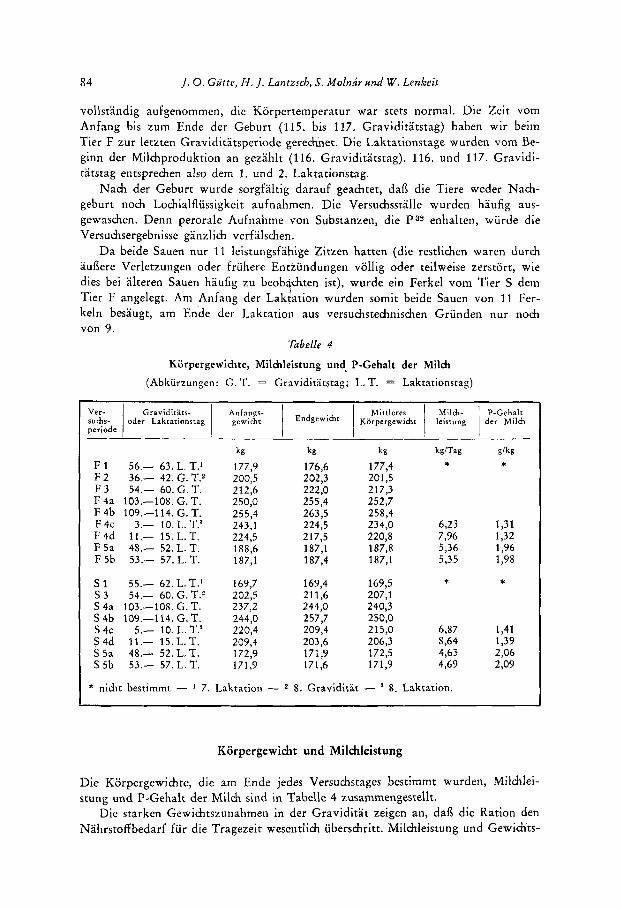
Tabelle 4
Korpergewichte, Milchleistung und. P-Gehalt der Milch (Abkurzungen: G. T. = Graviditatstag; L. T. = Laktationstag)
F 1 56.- 63. L. T.1 F 2 36.- 42.G.T.Z F 3 54.- 60.G.T.
F 4b 109.-114. G. T. F 4 c 3.- 10.L.T.'
F 4a 103.-108. G. T.
F 4 d 11.- 15.L.T. F 5a 48.- 52. L. T. F 5b 53.- 57. L. T.
S 1 55.- 62. L. T.' S 3 54.- 60. G. T.' S 4a 103.-108. G. T. S 4b 109.-114. G. T . S ~ C 5.- 10.L.T.3 S4d 11.- 15.L.T. S 5 a 48.- 52.L.T. S 5b 53.- 57. L. T.
kg
177,9 200,5 212,6 250,O 255,4 243,l 224,5 188,6 187,l
169,7 202,5 237,2 244,O 220,4 209,4 172,9 171,9
kg
176,6 252,3 222,o 255,4 263,s 2#24,5 217,s 187,l 187,4
169,4 21 1,6 244,O 257,7 209,4 203,6 171,9 171,6
kg kglTag g k
177,4 * * 201,5 217,3 252,7 258,4 234,O 6,23 1,31 220,s 7,96 1,32 187,s 5,36 1,96 187,l 5,35 1,98
169,5 * * 207,l 240,3 250,o 215,O 6,87 1,41 206,3 8,64 1,39 172,s 4,63 2,06 171,9 4,69 2,09
I * nicht bestirnrnt - 7. Laktation - 2 8. Graviditat - 8. Laktation.
Korpergewicht und Milchleistung
Die Korpergewichte, die am Ende jedes Versuchstages bestimmt wurden, Milchlei- stung und P-Gehalt der Milch sind in Tabelle 4 zusammengestellt.
Die starken Gewichtszunahmen in der Graviditat zeigen an, daB die Ration den Nahrstoffbedarf fur die Tragezeit wesentlich uberschritt. Milchleistung und Gewichts-
Tabe
lle f
i
Ver
inde
rung
en d
es P
-Um
satz
es i
m V
erla
uf
der
Gra
vidi
tHt
und"
Lak
tati
0n
bei
zwei
Sau
en
(Abk
iirzu
ngen
: G
. T.
=
Gra
vidi
tats
tag;
L. T
. =
Lak
tatio
nsta
g)
F1
F
2
F3
F
4a
F 4b
F
4c
F 4d
F
5a
F 5b
s1
s3
S
4a
S 4b
s 4
c.
s 4d
S 5a
S
5b
56.-
63.L
.T.'
21,9
4 9,
88
* 36
.- 42
. G. T
.2
21,9
4 16
,44
14,8
0 54
.- 60
. G. T
. 21
,94
16,6
7 15
,29
103.
-108
. G
. T.
21,9
4 13
,18
11,7
9 10
9.-1
17.
G. T
. 21
,94
12,6
3 11
,50
3.-
10.L
. T.'
21,9
4 13
,08
11,9
4 11
.- 15
.L.T
. 21
,94
11,l
l 10
,13
48.-
52.L
.T.
21,9
4 9,
63
8,07
53
.- 57
. L. T
. 21
,94
9,02
7,
77
55.-
62. L
. T.'
21,9
4 54
.- 60
. G. T
.e
21,9
4 10
3.-1
08.
G. T
. 21
,94
109.
-115
. G
. T.
21,9
4 5.-
10
.L.T
.3
21,9
4 11
.- 15
.L.T
. 21
,94
48.-
52.L
.T.
21,9
4 53
.- 57
.L.T
. 21
,94
10,3
5 *
16,3
2 15
,Ol
13,0
9 11
,85
12,2
6 11
,28
12,9
1 12
,02
11,4
0 10
,63
10,0
2 8,
71
9,96
8,
93
I * ni
&t
best
imm
t -
7. L
akta
tion
- *
8. G
ravi
dita
t -
8. L
akta
tion
.
* *
1,64
7,
14
1,38
6,
65
1,39
10
,15
1,13
10
,44
1,14
10
,oo
0,98
11
,81
1,56
13
,87
1,25
14
,17
* 1,31
1,
24
0,98
0,
89
0,77
1,
31
1,03
* 6,93
10
,09
10,6
6 9,
92
11,3
1 13
,23
13,O
l
4,22
4,
67
4,03
5,
39
4,91
5,
20
4,18
3,
21
3,32
4,08
4,
45
5,14
4,
46
3,79
2,
54
3,25
3,
06
* *
4- 0,
83
4- 1,
24
+ 3,3
7 0,
55
+ 3,8
5 8,
19
- 4
,53
10,4
9 - 3
,84
10,5
2 - 1,
42
10,5
8 - 0,
98
* *
+ 1,1
7 + 3
,71
+ 53
3
9,70
- 4,
46
12,0
2 - 4,
02
9,54
- 0,
87
9,81
- 0,
90
* 32,5
30
,3
46,3
47
,6
45,6
53
,8
63,2
64
,6
* 31,6
46
,O
48,6
45
,2
51,5
60
,3
59,3

86 J . 0. Giitte, H. J . Lantzscb, S. Molnrir und W . Lenkeit
verluste sind in der Laktation ga in ahnlich wie in dem fruheren Versuch Sie. I1 (GUTTE und LENKEIT 1960; SOEHNGEN 1956), in dem die gleiche Futterration ver- abreicht wurde. Obercinstimmend mit fruheren Untersuchungen ist auch das starke Ansteigen des P-Gehaltes der Milch mit fortschreitender Laktation (LENKEIT, GIMM- LER und SIECK 1959 a ; SOEHNGEN 1956; KIRCHHOFF 1955; PERRIN 1955; BEINLICH 1953).
Die Veranderungen des P-Umsatzes im Verlauf der Graviditat und Laktation
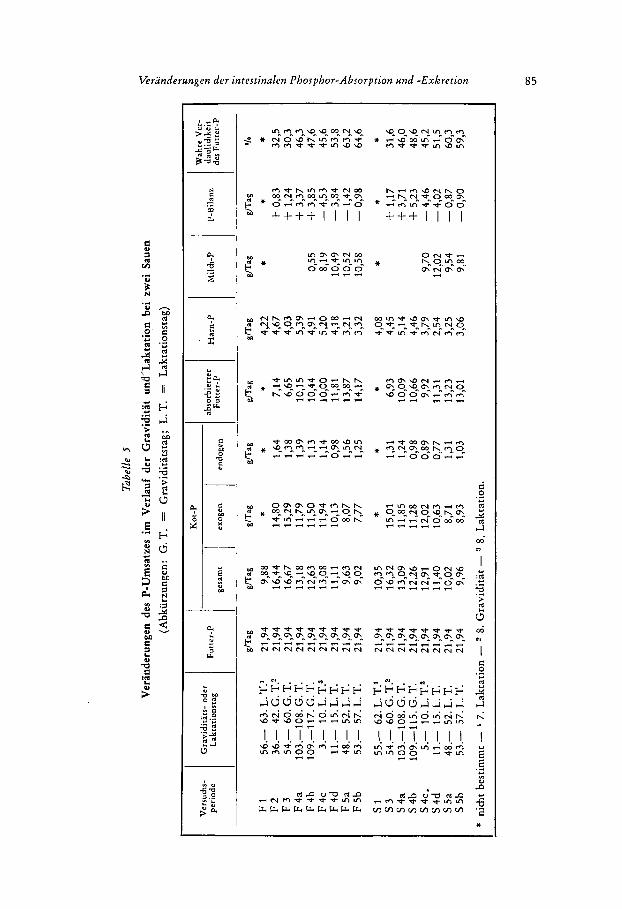
I n Tabelle 5 sind alle erfafiten Daten des P-Umsatzes enthalten. Die Resultate ent- sprechen genau den friiheren Untersuchungen:
P im K o t fallt von der Mitte bis zum Ende der Graviditat, steigt nach der Geburt schwach an und fallt bis zum Ende der Laktation stark ab. Die Bilanz wird am Ende der Graviditat stark positiv. Vermutlich ist auch hier, wie in den fruheren Unter- suchungen (LENKEIT et al. 1959 b; SOEHNGEN 1956; KIRCHHOFF 1955; M!TCHELL, CAR- ROL, HAMILTON und HUNT 1931), die P-Retention wahrend der gesamten Graviditat grofler gewesen als die P-Abgabe durch die Geburt. Am Anfang der Laktation war die P-Bilanz bei bejden Sauen stark negativ, am Ende der Laktation fast ausgeglichen, obwohl die P-Ausscheidungen in der Milch bei beiden Sauen am Ende der Laktation ebenso grofl waren wie in einer der beiden Perioden am Anfang der Laktation (ver- gleiche Pcriode F 4 d mit F 5 a und b, Periode S 4 c mit S 5 a und b).
Aus Tabelle 5 geht bereits hervor, dai3 die Veranderungen der Kot-P-Ausschei- dungen entscheidend durch Veranderungen des exogenen Kot-P beeinfluflt werden. Im folgenden sollen die einzelnen Ausscheidungsposten noch genauer betrachtet wer- den.
Tabelle 6
Veranderungen des endogenen Kot-P wahrend der Graviditat und Laktation von zwei Sauen bei konstanter Ernahrung
(Abkurzungen: G. T. = Graviditatstag; L. T. = Laktationstag)
endogener Kor-P __ Graviditits- oder I
Versudtsperiode I Laktationstag 1 Sau F I S a u S 1 Durchschnicr
F 2 F 3 und S 3 F 4 a und S 4 a F 4 b und S 4 b F4c und S 4c F 4 d und S 4 d F 5a und S 5a F 5b und S 5b
36.- 42. G. T. 54.- 60. G. T.
103.-108. G. T.
3. bzw. 5.-10. L. T. 109. G. T.-Geburt
11.- 1 5 . L . T . 48.- 52. L. T. 53.- 5 7 . L . T .
d T 3 ~
1,64 1,38 1,39 1,13 1,14 0,98 1,56 1,25
dTag
1,31 1,24 0,98 0,89 0,77 1,31 1,03
g/Tng
1,34 1,32 1,06 1,02 0,88 1,44 1,14
Endogener Kot-P
Der endogene Kot-I? ist mit 0,s bis 1,6 g/Tag so gering, dafl dadurch die Gesamt-P- Ausscheidung im Kot (9 bis 17 g/Tag) kaum beeinfluflt wird. Dennocli sind inter- essante Veranderungen dieser P-Fraktion festzustellen (Tab. 6 ) .
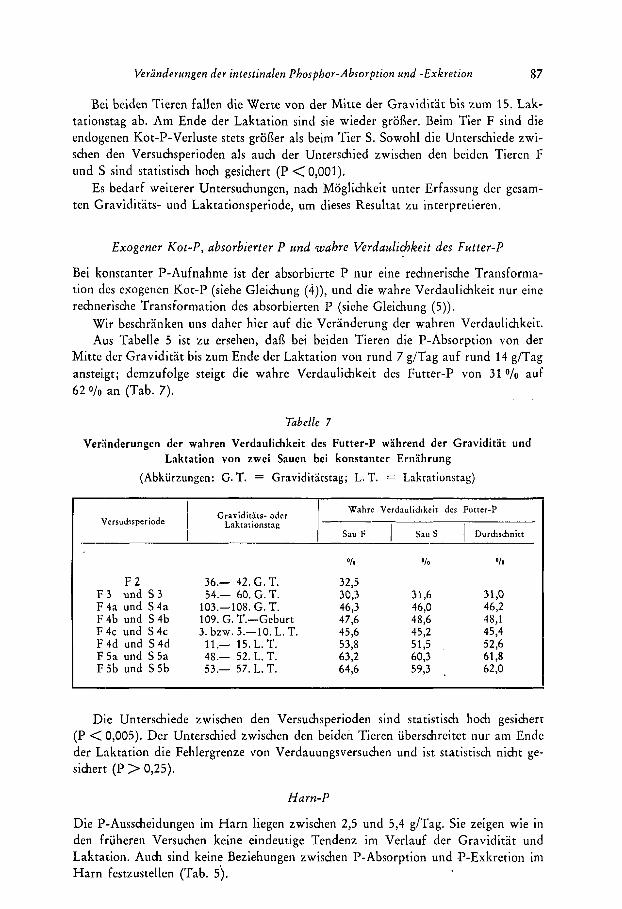
-
Veranderungen der intestinalen Phosphor-Absorption und -Exkretion 87
Bei beiden Tieren fallen die Werte von der Mitte der Graviditat bis zum 15. Lak- tationstag ab. Am Ende der Laktation sind sie wieder grofler. Ueim Tier F sind die
Exogener Kot-P, absorbierter P und wcrhre Verdaulichkeit des Futter-P
Bei konstanter P-Aufnahme ist der absorbierte P nur eine rechnerische Transforma- tion des exogenen Kot-P (siehe Gleichung (4)), und die wahre Verdaulichkeit nur cine rechnerische Transformation des absorbierten P (siehe Gleichung (5) ) .
Wir beschranken uns daher hier auf die Veranderung der wahren Verdaulichkeit. Aus Tabelle 5 ist zu ersehen, dafl bei beiden Tieren die P-Absorption von der
Mitte der Graviditat bis zum Ende der Laktation von rund 7 g/Tag auf rund 14 g/Tag ansteigt; demzufolge steigt die wahre Verdaulichkeit des Futter-P von 31 O/O auf 62 o/o a n (Tab. 7).
Tabelle 7
Verlnderungen der wahren Verdaulimkeit des Futter-P wahrend der Graviditlt und Laktation von zwei Sauen bei konstanter Ernlhrung
(Abkurzungen: G. T. = Gravidititsrag; L. T. = Laktationstag)
I Wahre Verdaulirhkeir des Furrer-P GravidirHts- oder Vcrsurhsperiode Lakrationstag I I
I I Sau F I Sau S I Durchschnitr
F 2 F 3 und S 3 F 4 a und S 4 a F 4 b und S 4 b F 4 c und S 4 c F 4 d und S 4 d F 5a und S 5a F 5 b und S 5 b
36.- 4 2 . G . T . 54.- 60. G. T.
103.-108. G. T.
3. bzw. 5.-10. L. T. 11.- 15. L. T.
, 48.- 52. L. T. 53.- 5 7 . L . T .
109. G. T.-Gcburt
32,5 30,3 31,6 46,3 46,O 47,6 48,6 45,6 45,2 53,s 51,5 63,2 60,3 64,6 59,3 ,
31,O 46,2 48,l 45,4 52,6 61,s 62,O
Die Unterschiede zwischen den Versuchsperioden sind statistisch hoch gesichert (P < 0,005). Der Unterschied zwischen den beiden Tieren uberschreitet nur am Ende der Laktation die Fehlergrenze von Verdauungsversuchen und ist statistisch nicht ge- sichert (P > 0,25).
Ham-P
Die P-Ausscheidungen im H a r n liegen zwischen 2,5 und 5,4 g/Tag. Sie zeigen wie i n den fruheren Versuchen keine eindeutige Tendenz im Verlauf der Graviditat utid Laktarion. Auch sind keine Beziehungen zwischen P-Absorption und P-Exkretion ini H a r n festzustellen (Tab. 5).

88 J . 0. Giitte, H . /. Lantzsch, S. Molnir und W. Lenkeit
Milch-P
Wie in vielen friiheren Versuchen sind die P-Ausscheidungen in der Milch am Ende der Laktation ebenso groa wie am Anfang der Laktation. Die P-Absorption ist trotz- dem am Ende der Laktation groCer als am Anfang der Laktation, so dai3 kein direk- ter Zusammenhang zwischen diesen beiden Groi3en zu bestehen scheint (Tab. 5). Den- noch ist anzunehmen, dai3 unter sonst gleichen Bedingungen (gleiches Laktationssta- dium, gleiche Vergangenheit der Versuchstiere, gleiche Futterung) die P-Absorption bei groi3er P-Ausscheidung in der Milch groaer sein wird als bei geringer P-Aus- scheidung in der Milch.
SchluG folgerungen
Die Ergebnisse lassen sich am besten unter Heranziehung eines Modells verstehen, welches in Anlehnung an BRONNER und HARRIS (1956) in Abbildung 2 dargestellt ist:
In tes t ina le P -Absorplion
4 P-E in lagerung 1. undzer y b P - E i n l a g r r u n g in
extrsskrlellale Gewebe P im B l u l s r r u m
in das Skelcl l
(P-Appos i l ionl
P - Abgabr
a u s dern Skelcll
(P-Resorpl ionl
rx lrare l lu laren
Gewebsllussigkcil
( P - l r a n s p o r l I
er l raske lz l la len Geweben
/ . P - E x k r e l i o n \
Ham Milch Korperoberl lache K O /
,466. 2. Schcmatische Darstellung des P-Umsatzes
Die Futterung war in diesen Versuchen kon- stant. Der Entzug von P aus dem Blut durfte da- gegen im Verlauf der Graviditat durch wach- sende P-Einlagerung i n den Uterus angestiegen sein. Wfhrend der Lak- tation steigt der P-Ent- zug aus dem Blut durch steigende P-Ausscheidun- gen in der Milch bis zur Mitte der Laktation wei- ter an.
Falls ein normaler P- Blutspiegel erhalten wer- den soll, kann der anstei- gende P-Entzug folgen- dermaaen kompensiert werden :
Verminderung der intestinalen P-Exkretion (endogener Kot-P), Verminderung der renalen P-Exkretion (Harn-P), Verminderung der P-Einlagerung in das Skelett (P-Apposition) oder in extra- skelettale Gewebe, Vermehrung der P-Abgabe aus dem Skelett (Knochen-P-Resorption) ,oder aus extraskelettalen Geweben, Vermehrung der intestinalen P-Absorption. Bei den beiden Sauen wird im Verlauf der Graviditat die P-Absorption erhoht.
Mit dem Einsetzen der Laktation werden dann Korperreserven von P mobilisiert, die P-Bilanz wird stark negativ (Tab. 5). Die P-Absorption braucht daher zunachst nicht anzusteigen, um ' den wachsenden P-Entzug aus dem Blut auszugleichen, sie fallt so-

Veranderungen dcr intestinalen Phosphor-Absorption und -Exkretion 89
gar schwach ab. Es mug besonders betont werden, da8 die Mobilisierung von Kor- per-P nach der Geburt ausbleiben kann, wenn ein Tier infolge mangelhafier P-Ver- sorgung vor der Geburt die Laktation mit geringen Korperreserveii an P beginnt (Versuch ROTK. I, LENKEIT et al. 1959 b, Tab. 81 , SOEHNGEN 1956).
Vom 10. Laktationstage ab reichen die mobilisierten Korper-P-Mengen nicht mehr aus. Nun wird die P-Absorption weiter gesteigert. Mit fortschreitender Er- schopf'ung dcr Korperreservcn nahert sich die Bilanz bis zum Ende der Laktation dem Ausgleich, wahrend die intestinale P-Absorption fortgesetzt ansteigt.
Die intestinale und renale P-Exkretion zeigen zwar im Versuchsverlauf Veran- derungen; doch tragen diesc nur in geringem Mai3e zur Regulierung des P-Umsatzes bei. Sie werden offenbar nur zur feineren, eventuell kurzfristigen Regulation einge- setzt. Dies uberrascht insofern, als nian bisher der renalen P-Exkretion, die vom Parathormon gesteuert wird (PULLMAN, LAVENDER, AHO und RASMUSSEN 1960), eine grogere Bedeutung bei der Kegulation zugesclirieben ha t als der intestinal& P-Ab- sorption.
Bei den hier untersuchten Tieren wurde dagegen in erster Linie die intestinale P- Absorption, in zweiter Linie die Speicherung und Mobilisierung von Korper-P (Ske- lect?) und erst in dritter Linie die renale und intestinale P-Exkretion zur lang- fristigen Regulation des P-Umsatzcs eingesetzt.
Zusammenfassung
Zwei 4jahrigen veredelten Landschwein-Sauen wurde wahrend der Graviditat und Laktation eine konstante Tagesration gefuttert, welche 21,94 g P/Tag enthielt. In der Mitte und am Ende der Graviditat, am Anfang und am Ende der Laktation wurden nach parenteraler Verabreichung von' P32 aus dem Verhaltnis der spezi- fischen Aktivitaten von Kot und H a r n endogener Kot-P, exogener Kot-P, absor- bierter Futter-P und wahre Verdaulichkeit des Futter-P berechnet. Daneben wurden die P-Ausscheidungen in H a r n und Milch bestimmt und die P-Bilanz errechnet.
Es wurde ein Ansteigen der wahren Verdaulichkeit des Futter-P von der Mitte der Graviditat bis zum Ende der Laktation von 310/0 auf 620/0 festgestellt. Die Unterschiede der wahren Verdaulihkeit in den einzelnen Graviditats- und Lak- tationsperioden waren statistisch hoch gesichert (P< 0,005). Auch die intestinale P- Exkretion zeigte hoch signifikante Veriinderungen (P< 0,001). Doch waren die Ver- luste von endogenem Kot-P mit 0,8 bis 1,6 g/Tag (7 bis l60/0 des Gesamt-Kot-P) so gering, 'da8 dadurch der gesamte P-Umsatz nur unwesentlich beeinflugt wurde.
Gedankt sei Dr. MANUELA ROLLER-BURESCH, Dr. R. VON BERSWORDT-WALLRABE und Herrn E. PFEFFER fur technische Assistenz, Dr. VON BERSWORDT-WALLRABE fer- ner fur wertvolle Anregungen bei der Diskussion der Resultate.
Auf einen Druckfehler in dieser Tabelle sei hier hingewiesen: Lies im unteren Teil der Tabellc ,,Ansatz in 60 Laktationstagen" anstatc ,Verlust in 60 Laktationstagen".

90 J . 0. Gulte, H . J . Lantzscb, S. Molna'r und W . Lenkeit
Summary
A constant diet, containing 21.94 grams of phosphorus per day, was fed to two 4 years old sows (breed: veredeltes Landschwein) during their 8 t h pregnancy and lactation. A t middle and end of pregnancy, a t beginning and end of lactation, endo- genous fecal P, exogenous fecal P, absorbed P and true digestibility of P were cal- culated from the ratio of specific activities of feces and urine after parenteral adrnini- stration of P3*. Furthermore, excretion of P in urine and milk was determined and P balance calculated.
The true digestibility of P increased from middle of pregnancy up to the end of lactation from 31o/o to 620/0. The differences of true digestibility between diffe- rent stages of pregnancy and lactation were statistically significant (P< 0,005). Also intestinal excretion of P did show significant changes (P< 0,001). But, since losses of endogenous fecal P have been low (0.8 to 1.6 grams per day, 7 to 16 Q / o of total fecal P), total metabolism of phosphorus was not very much affected by changes of this fraction.
Literatur
BEINLICH, W. (1953): Untersuchungen zum Calcium- und Phosphor-Sroff wechsel des saugen- den Schweines. Diss. agr. Gottingen. - BRONNER, F., & R. S. HARRIS (1956): Absorption and Metabolism of Ca in Human Beings, studied with Ca45. Annals New York Acad. Sci. 64, 314. - FARRIES, F. E. (1957): Zum Einflui3 steigender Kaliumgaben auf den Gesamt- stoffwechsel wachsender und gravida Schweine. Diss. sc. agr. Gottingen. - GERICKE, S., & B. KURMlEs (1952/53): Kolorimetrische Bestimniung der Phosphorsaure mit Vanadat- Molybdat. 2. andyt . Chem. 137, 15. - GUTTE, J. O., & W. LENKEIT (1960): Energieumsatz und Nahrstoffbedarf saugender Sauen. Z. Tierphysiol. Tierernah'r. Futtermittelk. 15, 165. - HEVESY, G., L. HAHN & 0. REBBE (1939): Excretion of Phosphorus. Det Kgl. Danske Videnskabernes Selskab. Biologiske Meddelelser, Kebenhavn, 14. Nr . 3. - KIRCHHOFF, W. (1955): Untersuchungen zum Mineralstoffwechsel wahrend der Graviditat und Laktation beim Schwein. Diss. sc. agr. Gottingen. - KITSON, R. E., & M. G. MELLON (1944): Colori- metric Determination of Phosphorus as Molybdivanadophosphoric Acid. Indust. Eng. Chem. 16, 379. - KJERULF-JENSEN, K. (1941/42): Excretion of Phosphorus by the Bowel. Acta Physiol. Scand. 3, 1. - KOENIG, R. A., & C. R. JOHNSON (1942): Colorimetric Deter- mination of I'hosphorus in Biological Materials. Industr. Eng. Chem. 14, 155. - KURMIES, B. (1955): Colorimetrische Bestimmung kleinster Mengen Phosphorsiiure mit Vanadat-Molyb- dat. Phosphorsaare 15, 64. - LANTZSCH, H. J. (1961): Untersuchungen uber die P-Absorp- tion und P-Exkretion an graviden .und laktierenden Sauen unter Verwendung des Radio- isotops P2. Diss. sc. agr. Gottingen. - LENKEIT, W. (1959): Der Tragezeit-Laktationszyklus und die Mineralversorgung. Ziichtungskunde 31, 410. - LENKEIT, W., W. GlhlMLER & K. H. SIECK (1959 a): Beitrag zur Calcium- und Phosphorausschcidung mit der Milch im Ablauf der Laktation ((Kuh, Sau). Archiv Tierernahr. 9, 166. - LENKEIT, W., J. 0. GUTTE, W. KIRCHHOFF & F. K. SOEHNGEN (1959 b): Zur Abhangigkeit des Mineralumsatzes, unter Be- amtung der negativen Ca-, P-Anfangsbilanz wahrend der Laktation, von der Mineralstoff- versorgung wahrend der Graviditat. Z. Tierphysiol. Tierernahmr. Futtermittelk. 14. 3. - LENKEIT, W., & J. 0. GUTTE (1955): Langfristige Untersuchungen zum augeren und innercn Stoffwechsel des graviden und laktierenden Schweines. 1. Mitteilung. Z. Tierernahr. Futter- mittelk. 10, 94. - MITCHELL, H., W. E. CARROL, T. S. HAMILTON & G. E. HUNT (1931): Food Requitrements of Pregnancy in swine. Univ. Illinois, Agr. Exp. Sta. Bull. 375. - MUNZNER (1952): Varianzanalyse. Vorlesung Sommersemester 1952 Gottingen. - PERRIN, D. R. (1955): The Chemical Composition of rhe Colostrum and Milk of the Sow. J. Dairy Res. 22, 103. - PULLMAN, T. N., A. R. LAVENDER, I. AHO & H. RASMUSSEN (1960): Direct Renal Action of a Purified Parathyroid Extract. Endocrinology 67, 570. - SNEDECOR, G . W. (1957): Statistical Methods. 5th ed., Iowa State College Press, Ames, Iowa. - SOEHNGEN, F. K. (1956): Untersuchungen uber dic Speicherung von Mineralstoffen wahrend der Gmvidi- tat und ihre Beziehung zum Mineralst-offwechsel wiihrend der Laktation, nadi Versuchen an Schweinen und Ziegen. Diss. sc. agr. Gottingen.