vasoactive intestinal polypeptide containing neurones in monkey medial prefrontal cortex (mpfc):...
TRANSCRIPT

Ž .Brain Research 744 1997 179–184
Short communication
Vasoactive intestinal polypeptide containing neurones in monkey medialŽ .prefrontal cortex mPFC : colocalisation with calretinin
P.L.A. Gabbott ), S.J. BaconUniÕersity Department of Pharmacology, Mansfield Road, Oxford OX1 3QT, UK
Accepted 15 October 1996
Abstract
Ž .Neurones immunoreactive for vasoactive intestinal polypeptide VIP were studied in monkey medial prefrontal cortex. The majorityŽ . Ž .78.0% of VIPq neurones were bipolar cells located mainly in layers 2r3. Calretinin CR immunoreactivity was colocalised in 80.5%of VIPq neurones. Furthermore, VIPq puncta formed pericellular baskets around GABA immunonegative somata in layers 2r3. Theseresults indicate that VIPrCR are colocalised in some bipolar cells and superficial pyramidal somata are likely targets of VIPq neurones.
Keywords: Interneurone; Pericellular basket; Inhibition; Vertical organisation; Limbic cortex
Ž .Vasoactive intestinal polypeptide VIP is present in adefined subpopulation of local circuit GABA-containingneurones in the m am m alian cerebral cortexw x3,11,14,16,20,21,24–27,31,33,34 . This group of cells ismainly composed of vertically oriented bipolar corticalinterneurones with narrow axonal arbors that traverse sev-
w xeral cortical layers 21,23–26,33,34 . Morphological evi-dence suggests that some VIP-containing cells have thepotential to strongly influence the activity of not only other
w xinterneurones 10,13,24–26,31 but also pyramidal cellswsituated in a confined radial column of the cortex 13,24–
x26,31 .w xIn a recent immunocytochemical study 7 we have
described several types of interneurone in the monkeyŽ .medial prefrontal cortex mPFC containing the calciumŽ .binding protein calretinin CR that are morphologically
w xsimilar to VIP-containing cells 24–27,33 . Indeed, previ-ous investigations have demonstrated the colocalisation of
w xVIP and CR in some cortical neurones 11,28 . The pur-pose of this short study was two-fold: firstly, to define themorphology and distribution of VIP containing local cir-cuit neurones in monkey mPFC, and secondly, to deter-mine the extent, if any, of VIP and CR colocalisation.
The material came from two adult female cynomolgusŽ w x.monkeys Macaca fascicularis 7 . Each animal had been
) Ž .Corresponding author. Fax: q44 865 27-1853; E-mail: [email protected]
deeply anaesthetised with sodium pentobarbitone and sub-sequently perfused transcardially with a mixture of alde-hyde fixatives containing 3% paraformaldehyde and0.5%–0.7% glutaraldehyde in 0.1 M phosphate buffer pH
w x7.4 7 .Ž .Vibratome tissue sections 100 mm thick were cut
w xfrom areas 24a,b,c, 25 and 32 of the mPFC 7 . TheseŽ .sections were washed in Tris-HCl pH 7.4 Tris buffer and
either bathed in 0.3% Triton X-100 for 30 min or freeze-w xthawed 5 . The tissue was then processed for dual
Ž .immunocytochemistry: i incubation in 20% normal serum,Ž .ii incubation in a primary antibody, VIP rabbit polyclonalŽeither from Penninsula Labs., USA, diluted 1:2000–1:5000, or from Sigma Chemicals, Code: V3508, diluted
. Ž .1:500–1:2000 , iii visualisation of immunolabelling us-w Žing an anti-rabbit Vectastain ABC kit Vector Laborato-
. Xries and developed either with 3,3 -diaminobenzidineŽ .DAB and 0.1% H O for 3–8 min or with the Vector2 2
w Ž . Ž .SG peroxidase substrate kit Vector Laboratories , ivŽ . Ž .avidinrbiotin blocking kit Vector Laboratories , v incu-
Žbation in either an anti-CR serum goat polyclonal, Code.CG1, S. Want, Switzerland, diluted 1:1000–1:8000 or an
Žanti-GABA serum mouse monoclonal, Code GB69, Sigma. Ž .Chemicals, diluted 1:200–1:500 , vi either anti-goat or
anti-mouse antibodies conjugated with fluorescein isothio-Ž .cyanate FITC, diluted 1:50–1:200 . All of the sera were
diluted in Tris buffer. Between each of the above steps theŽ .sections were washed in Tris buffer 3=10 min and
incubation in the primary sera lasted for 48 h at 48C or
0006-8993r97r$17.00 Copyright q 1997 Elsevier Science B.V. All rights reserved.Ž .PII S0006-8993 96 01232-2

( )P.L.A. Gabbott, S.J. BaconrBrain Research 744 1997 179–184180
overnight at room temperature. In control incubations,primary andror secondary antibodies were omitted. Therewas a lack of specific immunolabelling in control sections.
After the immunocytochemical reactions the tissue sec-tions were thoroughly washed in Tris buffer and mountedin 100% glycerol on glass slides. Some additional sectionswere Nissl stained, dehydrated and mounted in DPX –these sections helped to identify the cytoarchitectonic areas
w xand cellular lamination of mPFC 7,8 . The tissue wassubsequently examined in a Leitz photomicroscopeequipped with both bright-field and fluorescence optics.
Under bright-field illumination, vertical strips of cortexfrom pia to white matter were sampled and individualVIPq neurones examined for their CR immunoreactivity
Ž .by switching to fluorescence optics Fig. 3D . Usingbright-field and fluorescence optics together it was possi-ble to simultaneously visualise both VIP and CR or GABA
Ž .immunoreactivities Fig. 3D . Quantitative estimates of thenumber of cells that contained both VIP and CR im-
Ž .munoreactivities VIPqrCRq or that were only im-munolabelled for VIPq or CRq were made directly fromthe tissue. Five sections from area 24b were analysed fordual labelling in each animal. Care was taken only tosample regions of the tissue penetrated by both immunore-
Ž X X .agents Fig. 3E,E and Fig. 3F,F . Only somatic profilescontaining a nucleus were included in the morphometricanalysis. In addition, the somatodendritic morphologies ofwell labelled VIPq neurones in given layers were noted.
Neurones immunoreactive for VIP were present in alllayers and in all areas of monkey mPFC. VIP immunoreac-tive neurones bore the characteristic morphological hall-
w xmarks of cortical interneurones 6,12,17,22,23,33 . Pyrami-dal and glial cells were not immunolabelled. The distribu-tion and morphology of VIPq neurones were consistent
Ž .across cortical areas Fig. 1 . The peak cortical depthdistribution of VIPq somata occurred at the layer 2r3border with the VIPq cell population being distributed asfollows: layer 1, 1.9%, layer 2, 11.4%, layer 3, 62.8%, and
Ž .layers 5r6, 18.7% Fig. 1 . Some VIPq cells were foundŽ .in the white matter Fig. 1 .
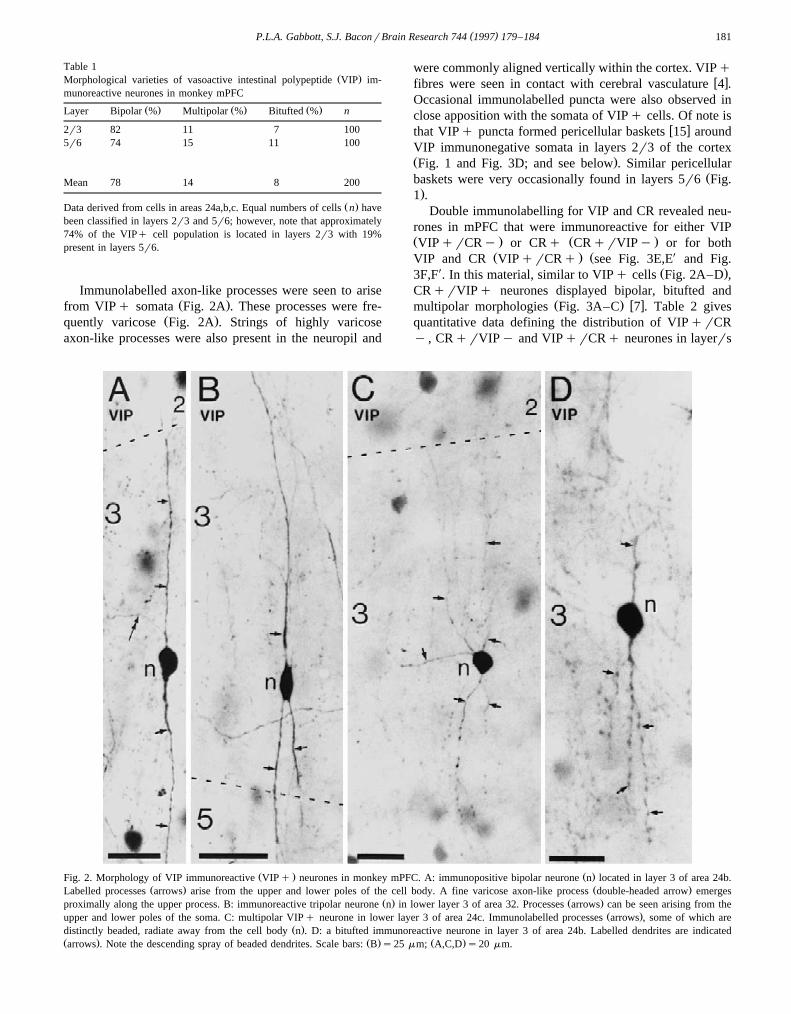
Three basic morphological types of VIPq neuronesŽ .were present in monkey mPFC Fig. 2A–D . The predomi-
nant type of VIPq neurone was bipolar in appearanceŽ .Table 1 . These cells commonly possessed small-sizedovoidrfusiform cell bodies with their major somatic axis
Ž .oriented vertically Fig. 2A . The mean minor and majorsomatic profile diameters of these cells were 7.5 mm=11.8mm. Bipolar neurones had narrow vertically oriented as-piny dendritic arbors consisting of an ascending and adescending dendrite. The primary processes would com-monly bifurcate into two or more daughter branches that
Ž .tapered gradually Fig. 2A,B . On occasions, these narrowdendritic fields would span several cortical layers. SomeVIPq neurones belonging to this class had a third den-drite arising proximally to one of the main processesthereby giving the parent somata a tripolar appearance
Ž .Fig. 1. Cortical depth distribution of VIP immunoreactive VIPq somatain area 24b of monkey mPFC. Black dots represent labelled VIPqneuronal cell bodies. Laminar boundaries are indicated. Open trianglesindicate the positions of somatic profiles encircled by VIPq pericellularbaskets. Note the peak distribution of VIPq somata at the level of thelayer 2r3 border and the superficial distribution of profiles surrounded bypericellular baskets. The distribution was derived from several sections.WM, white matter. Scale bars500 mm.
Ž .Fig. 2B . Birtripolar VIPq neurones were predomi-Ž .nantly found in layers 2r3 Table 1 .
Two other types of VIPq neurone were present inŽ .mPFC multipolar and bitufted cells Table 1 . VIP im-
munoreactive multipolar neurones commonly possessedŽ .circular somata with radiate dendritic arbors Fig. 2C .
Two subtypes of multipolar cells were present, the firsthad aspiny dendrites whereas the higher order dendrites of
Žthe second subtype were very sparsely spinous not illus-.trated . Bitufted VIPq neurones had clusters of narrow
vertically oriented dendrites arising either directly fromopposite somatic poles or from short dendritic processesŽ .Fig. 2D . The somata of bitufted cells were either circularor ovoid in outline and had highly beaded and aspiny
Ž .dendrites Fig. 2D .The proportions of birtripolar, multipolar and bitufted
cell types varied between the superficial and deep layers ofŽ .the cortex Table 1 . VIPq neurones were also found in
Ž .layer 1 Fig. 1 , such cells were commonly either smallmultipolar cells near the layer 1r2 border or cells withhorizontally elongated dendritic arbors.
( )P.L.A. Gabbott, S.J. BaconrBrain Research 744 1997 179–184 181
Table 1Ž .Morphological varieties of vasoactive intestinal polypeptide VIP im-
munoreactive neurones in monkey mPFC
Ž . Ž . Ž .Layer Bipolar % Multipolar % Bitufted % n
2r3 82 11 7 1005r6 74 15 11 100
Mean 78 14 8 200
Ž .Data derived from cells in areas 24a,b,c. Equal numbers of cells n havebeen classified in layers 2r3 and 5r6; however, note that approximately74% of the VIPq cell population is located in layers 2r3 with 19%present in layers 5r6.
Immunolabelled axon-like processes were seen to ariseŽ .from VIPq somata Fig. 2A . These processes were fre-
Ž .quently varicose Fig. 2A . Strings of highly varicoseaxon-like processes were also present in the neuropil and
were commonly aligned vertically within the cortex. VIPqw xfibres were seen in contact with cerebral vasculature 4 .
Occasional immunolabelled puncta were also observed inclose apposition with the somata of VIPq cells. Of note is
w xthat VIPq puncta formed pericellular baskets 15 aroundVIP immunonegative somata in layers 2r3 of the cortexŽ .Fig. 1 and Fig. 3D; and see below . Similar pericellular
Žbaskets were very occasionally found in layers 5r6 Fig..1 .
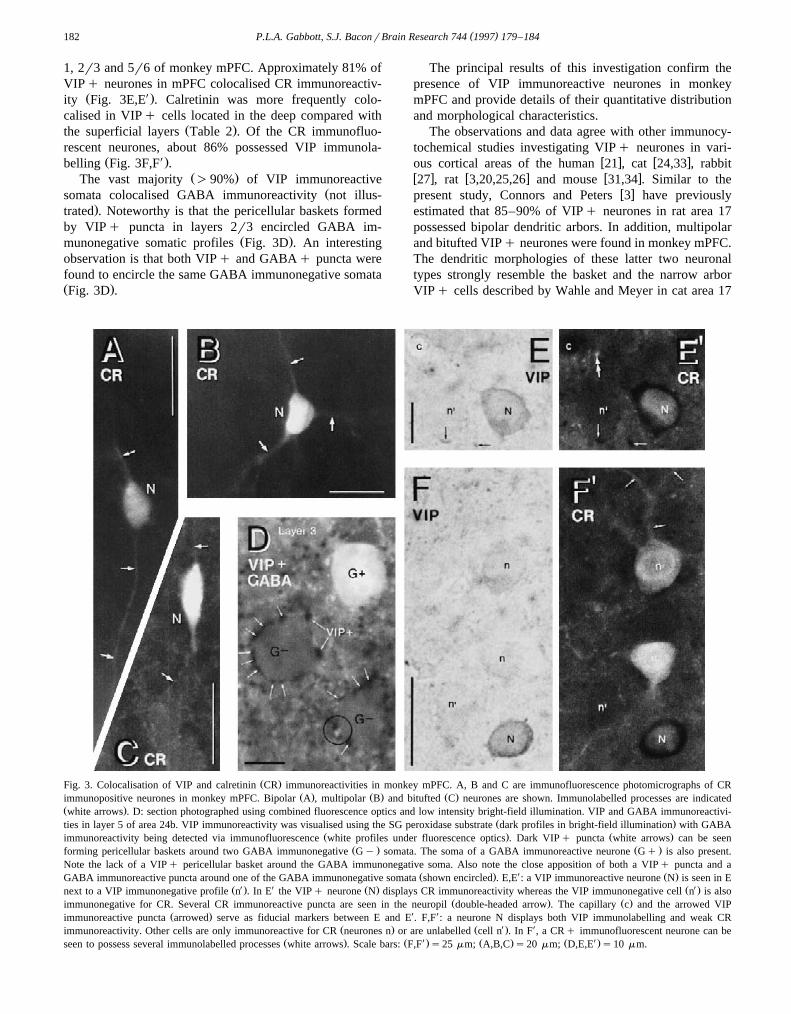
Double immunolabelling for VIP and CR revealed neu-rones in mPFC that were immunoreactive for either VIPŽ . Ž .VIPqrCRy or CRq CRqrVIPy or for both
Ž . Ž XVIP and CR VIPqrCRq see Fig. 3E,E and Fig.X Ž .3F,F . In this material, similar to VIPq cells Fig. 2A–D ,
CRqrVIPq neurones displayed bipolar, bitufted andŽ . w xmultipolar morphologies Fig. 3A–C 7 . Table 2 gives
quantitative data defining the distribution of VIPqrCRy , CRqrVIPy and VIPqrCRq neurones in layerrs
Ž . Ž .Fig. 2. Morphology of VIP immunoreactive VIPq neurones in monkey mPFC. A: immunopositive bipolar neurone n located in layer 3 of area 24b.Ž . Ž .Labelled processes arrows arise from the upper and lower poles of the cell body. A fine varicose axon-like process double-headed arrow emerges
Ž . Ž .proximally along the upper process. B: immunoreactive tripolar neurone n in lower layer 3 of area 32. Processes arrows can be seen arising from theŽ .upper and lower poles of the soma. C: multipolar VIPq neurone in lower layer 3 of area 24c. Immunolabelled processes arrows , some of which are
Ž .distinctly beaded, radiate away from the cell body n . D: a bitufted immunoreactive neurone in layer 3 of area 24b. Labelled dendrites are indicatedŽ . Ž . Ž .arrows . Note the descending spray of beaded dendrites. Scale bars: B s25 mm; A,C,D s20 mm.
( )P.L.A. Gabbott, S.J. BaconrBrain Research 744 1997 179–184182
1, 2r3 and 5r6 of monkey mPFC. Approximately 81% ofVIPq neurones in mPFC colocalised CR immunoreactiv-
Ž X.ity Fig. 3E,E . Calretinin was more frequently colo-calised in VIPq cells located in the deep compared with
Ž .the superficial layers Table 2 . Of the CR immunofluo-rescent neurones, about 86% possessed VIP immunola-
Ž X.belling Fig. 3F,F .Ž .The vast majority )90% of VIP immunoreactive
Žsomata colocalised GABA immunoreactivity not illus-.trated . Noteworthy is that the pericellular baskets formed
by VIPq puncta in layers 2r3 encircled GABA im-Ž .munonegative somatic profiles Fig. 3D . An interesting
observation is that both VIPq and GABAq puncta werefound to encircle the same GABA immunonegative somataŽ .Fig. 3D .
The principal results of this investigation confirm thepresence of VIP immunoreactive neurones in monkeymPFC and provide details of their quantitative distributionand morphological characteristics.
The observations and data agree with other immunocy-tochemical studies investigating VIPq neurones in vari-
w x w xous cortical areas of the human 21 , cat 24,33 , rabbitw x w x w x27 , rat 3,20,25,26 and mouse 31,34 . Similar to the
w xpresent study, Connors and Peters 3 have previouslyestimated that 85–90% of VIPq neurones in rat area 17possessed bipolar dendritic arbors. In addition, multipolarand bitufted VIPq neurones were found in monkey mPFC.The dendritic morphologies of these latter two neuronaltypes strongly resemble the basket and the narrow arborVIPq cells described by Wahle and Meyer in cat area 17
Ž .Fig. 3. Colocalisation of VIP and calretinin CR immunoreactivities in monkey mPFC. A, B and C are immunofluorescence photomicrographs of CRŽ . Ž . Ž .immunopositive neurones in monkey mPFC. Bipolar A , multipolar B and bitufted C neurones are shown. Immunolabelled processes are indicated
Ž .white arrows . D: section photographed using combined fluorescence optics and low intensity bright-field illumination. VIP and GABA immunoreactivi-Ž .ties in layer 5 of area 24b. VIP immunoreactivity was visualised using the SG peroxidase substrate dark profiles in bright-field illumination with GABA
Ž . Ž .immunoreactivity being detected via immunofluorescence white profiles under fluorescence optics . Dark VIPq puncta white arrows can be seenŽ . Ž .forming pericellular baskets around two GABA immunonegative Gy somata. The soma of a GABA immunoreactive neurone Gq is also present.
Note the lack of a VIPq pericellular basket around the GABA immunonegative soma. Also note the close apposition of both a VIPq puncta and aŽ . X Ž .GABA immunoreactive puncta around one of the GABA immunonegative somata shown encircled . E,E : a VIP immunoreactive neurone N is seen in E
Ž X . X Ž . Ž X .next to a VIP immunonegative profile n . In E the VIPq neurone N displays CR immunoreactivity whereas the VIP immunonegative cell n is alsoŽ . Ž .immunonegative for CR. Several CR immunoreactive puncta are seen in the neuropil double-headed arrow . The capillary c and the arrowed VIP
Ž . X Ximmunoreactive puncta arrowed serve as fiducial markers between E and E . F,F : a neurone N displays both VIP immunolabelling and weak CRŽ . Ž X . Ximmunoreactivity. Other cells are only immunoreactive for CR neurones n or are unlabelled cell n . In F , a CRq immunofluorescent neurone can be
Ž . Ž X . Ž . Ž X .seen to possess several immunolabelled processes white arrows . Scale bars: F,F s25 mm; A,B,C s20 mm; D,E,E s10 mm.

( )P.L.A. Gabbott, S.J. BaconrBrain Research 744 1997 179–184 183
Table 2Ž .Percentage colocalisation of vasoactive intestinal polypeptide VIP and
Ž .calretinin CR immunoreactivities in layers 1–6 of area 24b in themonkey
Layer VIPqrCRy VIPqrCRq CRqrVIPy nŽ . Ž . Ž .% % %
Ž . Ž . Ž .1 4.5 1 86.4 19 9 2 22Ž . Ž . Ž .2r3 21.8 42 66.8 129 11.4 22 193Ž . Ž . Ž .5r6 12.5 15 75.8 91 11.7 14 120
VIPqrCRy VIPqrCRq CRqrVIPy nŽ . Ž . Ž .% % %
a ab bŽ . Ž . Ž .Value 17.3 58 71.3 239 11.3 38 335for cortex
Ž .Data expressed as the percentage of sampled neurones in a given layer sŽ .that contained either a single VIPqrCRy or CRqrVIPy or double
Ž . Ž .VIPqrCRq immunolabel. The total number of profiles examined nŽ . Žin each layer s are given. The actual number of cells in each class
Ž . .layer s are shown in parentheses. The overall value for the cortex is alsogiven.a Of the total VIPq cell population: VIPqrCRy neurones constitute19.5%; VIPqrCRq constitute 80.5%.b Of the total CRq cell population: CRqrVIPy neurones constitute13.7%; CRqrVIPq constitute 86.3%.
w x33 . They are also similar to the VIPq basket and doublebouquet cells reported by Kawaguchi and Kubota in the
w xfrontal cortex of the rat 14 .The double-labelling results presented here indicate that
Ž .the majority 80.5% of VIPq neurones contain CR. Thishigh degree of colocalisation is in accord with otherstudies where 69–86% of cortical VIP immunoreactive
w xneurones colocalised CR immunolabelling 16,28,29 . Con-Ž .sequently on the basis of cell frequencies Table 1 , it is
likely that more than 73% of the bipolar neurones inmonkey mPFC are CRqrVIPq . Recent evidence indi-cates that calretinin is present in bipolar cells, double-bouquet neurones, multipolar cells and a class of small
w xbasket neurone 2,7,9,19 — cell types represented withinthe VIPq neurone population. Furthermore, since most, if
w xnot all, VIPq neurones contain GABA 16,28 and thatVIP is not colocalised in the specific neuronal classes
Ž .containing the calcium binding proteins, calbindin CBŽ . Žw x w x.and parvalbumin PV 15,28–30 ; see also ref. 7 , it is
therefore likely that the VIPqrCRq neurones describedhere form a neurochemically distinct subset of local circuitGABA-containing neurones in monkey mPFC.
w xWahle and Meyer 33 have previously identified theaxonal projection patterns of VIPq neurones during thedevelopment of cat area 17. Superficially placed multipolarand bitufted VIPq cells in the adult cat commonly pos-sessed single long vertically oriented axonal fibres thatdescended to layers 5r6 before ramifying into varicoseterminal fields. In contrast, bipolar VIPq neurones haveaxonal fields ramifying close to their parent somata. Wahleand Meyer also described multipolar VIPq basket neu-
w xrones that were predominantly located in layers 2r3 33 .
Similar to monkey mPFC, these basket cells formed VIPqŽ .pericellular baskets in the superficial layers Fig. 1 .
The synaptic connectivity of identified VIPq bipolarneurones in the visual cortices of the adult cat and rat has
w xbeen investigated 3,24–26 . These cells make symmetricŽ w x.synaptic junctions Gray type II 24,26 and principally
target the somata and processes of pyramidal neurones aswell as the smooth dendritic shafts of other interneuronesw x3,24–26 . Although electron microscopic evidence of thestructures innervated by identified bitufted and multipolarVIPq cortical neurones is presently unavailable, Martinand Meskenaite have described the post-synaptic targets ofsimilar cells in CR immunolabelled material from monkey
w xstriate cortex 18 . Of interest is that bitufted and multipo-lar CRq neurones mainly innervated deep layer pyramidswhereas in the superficial layers GABAq cells were thepredominant targets – thus indicating a double functional
Ž .role direct inhibition vs. disinhibition of the CRq inner-vation to lower and upper layers respectively.
Several aspects of the organisation of VIPq neuronesin monkey mPFC suggest that this subpopulation of localcircuit inhibitory neurones participates in the functional
w xcolumnar architecture of monkey mPFC 1 . The narrowlyconfined vertical orientation of immunoreactive dendritesand axon-like processes indicates that specific members ofthis category of interneurone receive and distribute infor-
w xmation in a columnar fashion 1,13,15 . The distribution ofVIPq neurones in the superficial layers further suggeststhat the majority of VIPq neurones receive synaptic inputfrom corticalrsubcortical regions or local intracorticalsources which preferentially innervate the superficial com-
Žpared with the deep layers of the cortex e.g., amygdalaw x.and thalamic input 32 . Such innervation could differen-
tially influence specific output pathways from the upperw xand lower layers of the cortex 18 . Indeed, the predomi-
nant location of VIPq pericellular baskets around GABAimmunonegative profiles in layers 2r3 supports this ideaand indicates a close involvement of VIPq interneurones
w xin cortico-cortical and cortico-subcortical interactions 20a .These structural features suggest that the inhibition medi-
w xated by VIPq neurones 13 could significantly affectintracortical processing mechanisms as well as influencethe capacity of the mPFC to distribute information to other
w xparts of the limbic system in the monkey 32 .
Acknowledgements
We are grateful to Paul Jays and Tracy-Ann Warner fortheir technical assistance. We also thank Professor A.Cowey, Department of Experimental Psychology, Oxford,for providing us with the monkey material. This work wassupported by the MRC, the Beit Memorial Research Fundand The Royal Society. P.L.A.G. is a Beit MemorialResearch Fellow.

( )P.L.A. Gabbott, S.J. BaconrBrain Research 744 1997 179–184184
References
w x1 Arikuni, T., Sakai, M. and Kubota, K., Columnar aggregation ofprefrontal and anterior cingulate cortical cells projecting to thethalamic mediodorsal nucleus in the monkey, J. Comp. Neurol.,
Ž .220 1983 116–125.w x2 Conde, F., Lund, J.S., Jacobowitz, D.M., Baimbridge, K.G. and´
Lewis, D.A., Local circuit neurons immunoreactive for calretinin,calbindin D-28k or parvalbumin in monkey prefrontal cortex:
Ž .distribution and morphology, J. Comp. Neurol., 341 1994 95–116.w x3 Connors, J.R. and Peters, A., Vasoactive intestinal polypeptide-im-
Ž .munoreactive neurons in rat visual cortex, Neuroscience, 12 19841027–1044.
w x4 Chedotal, A., Cozzari, C., Faure, M.P., Hartman, B.K. and Hamel,´Ž .E., Distinct choline acetyltransferase ChAT and vasoactive in-
Ž .testinal polypeptide VIP bipolar neurons project to local bloodŽ .vessels in the rat cerebral cortex, Brain Res., 646 1994 181–193.
w x Ž .5 Cuello, A.C. Ed. , Immunohistochemistry II. IBRO HandbookSeries: Methods in Neurosciences, Vol. 14, Wiley, Chichester,1993.
w x6 Feldman, M.L. and Peters, A., The forms of non-pyramidal neuronsŽ .in the visual cortex of the rat, J. Comp. Neurol., 179 1978
761–794.w x7 Gabbott, P.L.A. and Bacon, S.J., Local-circuit neurons in the
Ž .medial prefrontal cortex areas 24a,b,c, 25 and 32 in the monkey.1. Cell morphology and morphometrics, J. Comp. Neurol., 364Ž .1996 567–608.
w x8 Gabbott, P.L.A. and Bacon, S.J., Local-circuit neurons in theŽ .medial prefrontal cortex areas 24a,b,c, 25 and 32 in the monkey.
2. Quantitative areal and laminar distributions, J. Comp. Neurol.,Ž .364 1996 609–636.
w x9 Glezer, I.I., Hof, P.R. and Morgane, P.J., Calretinin-immunoreac-tive neurons in the primary visual cortex of dolphin and human
Ž .brains, Brain Res., 595 1992 181–188.w x10 Gulyas, A.I., Hajos, N. and Freund, T., Interneurons containing´ ´
calretinin are specialised to control other interneurons in the ratŽ .hippocampus, J. Neurosci., 16 1996 3397–3411.
w x11 Hendry, S.H.C., Jones, E.G., DeFelipe, J., Schmechel, D., Bran-don, C. and Emson, P.C., Neuropeptide containing neurons of thecerebral cortex are also GABAergic, Proc. Natl. Acad. Sci. USA,
Ž .81 1984 6526–6530.w x12 Jones, E.G., Varieties and distribution of non-pyramidal cells in the
somatic sensory cortex of the squirrel monkey, J. Comp. Neurol.,Ž .160 1975 205–268.
w x13 Jones, E.G., Hendry, S.H.C., DeFelipe, J. and Benson, D.L.,GABA neurons and their role in activity-dependent plasticity of
Ž .adult primate visual cortex. In A. Peters and K.S. Rockland Eds. ,Cerebral Cortex. Vol. 10. Primary Visual Cortex in Primates,Plenum Press, New York, 1994, pp. 61–140.
w x14 Kawaguchi, Y. and Kubota, Y., Physiological and morphologicalidentification of somatostatin- or vasoactive intestinal polypeptide-containing cells among GABAergic cell subtypes in rat frontal
Ž .cortex, J. Neurosci., 16 1996 2701–2715.w x15 Kisvarday, Z.F., GABAergic networks of basket cells in the visual´
Ž .cortex. In R.R. Mize, R.E. Marc and A.M. Sillito Eds. , GABA inthe Retina and Central Visual System. Prog. Brain Research, Vol.90, Elsevier, Amsterdam, 1992.
w x16 Kubota, Y., Hattori, R. And Yui, Y., Three distinct subpopulationsof GABAergic neurons in rat frontal agranular cortex, Brain Res.,
Ž .639 1994 159–173.w x17 Lund, J.S. and Lewis, D.A., Local circuit neurons of developing
and mature prefrontal cortex: Golgi and immunocytochemical char-
Ž .acteristics, J. Comp. Neurol., 328 1993 282–312.w x18 Martin, K.A.C. and Meskenaite, V., Calretinin-immunoreactive
local circuit neurons in the area 17 of cynomolgus monkey, MacacaŽ .fascicularis, J. Comp. Neurol., 1996 in press.
w x19 McDonald, A.J., Calretinin immunoreactive neurons in the basolat-Ž .eral amygdala of the rat and monkey, Brain Res., 667 1994
238–242.w x20 McDonald, J.K., Parnavelas, J.G., Karamanlidis, A. and Brecha,
N., The morphology and distribution of peptide-containing neuronsin the adult and developing visual cortex of the rat. II. Vasoactive
Ž .intestinal polypeptide, J. Neurocytol., 11 1982 825–837.w x20a Nimchinsky, E.A., Hof, P.R., Young, W.G. and Morrison, J.H.,
Neurochemical, morphologic, and laminar characterisation of corti-cal projection neurons in the cingulate motor areas of the Macaque
Ž .monkey, J. Comp. Neurol., 374 1996 136–160.w x21 Ong, W.Y. and Garey, L.J., Distribution of GABA and neuropep-
tides in the human cerebral cortex: a light and electron microscopeŽ .study, Anat. Embryol., 183 1991 397–414.
w x22 Peters, A. and Regidor, J., A reassessment of the forms of non-pyramidal neurons in area 17 of the cat visual cortex, J. Comp.
Ž .Neurol., 203 1981 685–716.w x Ž .23 Peters, A., Bipolar cells. In A. Peters and E.G. Jones Eds. ,
Cerebral Cortex. Vol. 1. Cellular Components of the CerebralCortex, Plenum Press, New York, 1984, pp. 381–407.
w x24 Peters, A., Meinecke, D.L. and Karamanlidis, A.N., Vasoactiveintestinal polypeptide immunoreactive neurons in cat primary vi-
Ž .sual cortex, J. Neurocytol., 16 1987 23–38.w x25 Peters, A. and Harriman, K.M., Enigmatic bipolar cell of rat visual
Ž .cortex, J. Comp. Neurol., 267 1988 409–432.w x26 Peters, A., The axon terminals of vasoactive intestinal polypeptide
Ž .VIP -containing bipolar cells in rat visual cortex, J. Neurocytol.,Ž .19 1990 672–685.
w x27 Ramon-y-Cajal-Agueras, S., Contamina, P., Parra, P. and´Martinez-Millan, L., The distribution of vasoactive intestinalpolypeptide immunoreactive neurons in the visual cortex of adult
Ž .rabbits and during postnatal development, Brain Res., 370 1986333–337.
w x28 Rogers, J.H., Immunohistochemical markers in rat cortex: co-localisation of calretinin and calbindin-D28k with neuropeptides
Ž .and GABA, Brain Res., 587 1992 147–157.w x29 Rogers, J.H. and Resbois, A., Calretinin and calbindin-D28 in rat
Ž .brain. Patterns of partial colocalization, Neuroscience, 51 1992843–865.
w x30 Somogyi, P. and Cowey, A., Double bouquet cells. In A. PetersŽ .and E.G. Jones Eds. , Cerebral Cortex. Vol. 1. Cellular Compo-
nents of the Cerebral Cortex, Plenum Press, New York, 1984, pp.337–360.
w x31 Staiger, J.F., Zilles, K. and Freund, T., Innervation of VIP-im-munoreactive neurons by the ventroposteromedial thalamic nucleus
Ž .in the barrel cortex of the rat, J. Comp Neurol., 367 1996194–204.
w x Ž .32 Vogt, B.A. and Gabriel, M. Eds. , Neurobiology of CingulateCortex and Limbic Thalamus. A ComprehensiÕe Handbook,Birkhauser, Basel, 1993.¨
w x33 Wahle, P. and Meyer, G., Early postnatal development of vasoac-tive intestinal polypeptide- and peptide histidine isoleucine-im-munoreactive structures in the cat visual cortex, J. Comp. Neurol.,
Ž .282 1989 215–248.w x34 Zilles, K., Hajos, F., Csillag, A., Zilles, K., Kalan, M., Sotonyi, P.´ ´ ´ ´
and Schleicher, A., Vasoactive intestinal polypeptide immunoreac-Ž .tive structures in the mouse barrel field, Brain Res., 618 1993
149–154.