use of seagrass zostera novazelandica (setchell, 1933) as habitat and food by the crab...
TRANSCRIPT

ELSEVIER Journal of Experimental Marine Biology and Ecology,
214 (1997) 49-65
JOURNAL OF EXPERIMENTAL MARINE BIOLOGY AND ECOLOGY
Use of seagrass Zostera novazelandica (Setchell, 1933) as habitat and food by the crab A4acrophthalmus hirtipes
(Heller, 1862) (Brachyura: Ocypodidae) on rocky intertidal platforms in southern New Zealand
Chris M.C. Woods*, David R. Schiel
Marine Ecology Research Group, Zoology Department, University of Canterbury. Private Bag 4800, Christchurch, New Zealand
Received 2 February 1996; revised 22 October 1996; accepted 13 November 1996
Abstract
Along rocky reefs at Kaikoura, New Zealand, the crab Macroph~hulrnus lzirripes (Heller, 1862) constructs its burrows exclusively in patches of the intertidal seagrass Zustercc novuzelandiu
(Setchell, 1933). The occupation of seagrass patches is widespread, with 63% of patches having
crab burrows. The number of externally visible burrows is highly correlated with the number of crabs inhabiting seagrass patches. Plaster casts of burrows revealed that burrows situated at the edge of seagrass patches differed from those away from the edge in having on average more
entrances and passageways and, therefore, greater total burrow length. There was an average burrow density of 9.4 (rt 1.04 S.E.) per m2 of seagrass, but burrows were several times as
numerous on the edges of patches, especially those that bordered tide pools. The diet of M.
hirtipes was found to consist mostly of living seagrass (50.4t3.40% of foregut contents) and
sediment (39.2+3.11% of foregut contents), with small amounts of isopods, amphipods and conspecifics also consumed. There were no intersexual or site-specific differences in diet.
Ontogenetic differences in diet were found, with smaller crabs consuming more sediment and
larger crabs consuming more seagrass. Crabs contributed significantly to erosion of seagrass patches. There was an average of 22.5% (25.27) loss of patch area where crabs were present compared to I .8% (kO.37) where crabs were absent from patches. Laboratory tests showed that burrow construction took around 250% longer in seagrass patches that had denser blade coverage. In the field, burrows persisted for up to 3 months, even if vacated by crabs. Their abundance at patch edges, combined with the loss of sediment and reduced binding of seagrass roots,
accelerated erosion of seagrass patches. 0 1997 Elsevier Science B.V.
*Corresponding author. Present address: National Institute of Water and Atmospheric Research, P.O. Box
14-901, Wellington, New Zealand. Tel.: ( + 64-4) 386.0300; fax: ( + 64-4) 388-9931; e-mail:
0022.0981/97/$17.00 0 1997 Elsevier Science B.V. All rights reserved
P/f SOO22-098 I (96)02767-O

50 C.M.C. Woods, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 214 (1997) 49-65
Keywords: Burrows; Crab; Macrophthalmus hirtipes; Seagrass; Zostera novazelandica
1. Introduction
Seagrasses are marine angiosperms that have successfully adapted to shallow coastal
and estuarine environments, fulfilling several important ecological roles (Dawes, 1981; Duarte and Sand-Jensen, 1990). Because of their structural complexity, biodiversity and
productivity, they have been described as the marine analog of tropical rainforests (Simenstad, 1994 in Dawes et al., 1995). Seagrasses form extensive continuous meadows, or individually discrete patches (Virnstein, 1995a), providing both a habitat
and a source of food for a wide range of organisms including fishes, gastropods,
crustaceans, and echinoderms (Kikuchi, 1974; Dawes, 198 1; King, 198 1; Den Hartog,
1987; Perez and Bellwood, 1988; Edgar, 1990; Wassenberg, 1990; Bishop, 1992; Rosas et al., 1994; Dawes et al., 1995; Greenway, 1995; Virnstein, 1995b). Seagrasses may also
stabilise bottom sediments, improve water clarity (Ginsberg and Lowenstam, 1958; Dawes, 198 1 ), aid in primary production (Dawes, 1981) and act as substrata for
epiphytes (Humm, 1964; Philippart, 1995a,b).
In New Zealand, seagrasses are represented by the two species Zosteru capricorni and Z. novazelandica (muelleri). Both species are usually found above or at the low tide
mark of sheltered muddy tidal estuaries, river mouths and bay heads (Webb et al., 1990). Zostera cupricorni is found only around the North Island of New Zealand, while Z.
novazelandica is more widespread, occurring in all regions of the mainland (Webb et al.,
1990). Around the rocky exposed coastline of the Kaikoura Peninsula, Z. novazelandica
occurs in isolated areas upon intertidal siltstone platforms. The seagrass patches initially
form in sediment-laden tidal cracks, before expanding along cracks and over the siltstone
to form discrete mats (Ramage, 1995). Although these seagrass patches range up to 10
m’ in size, 75% of them are < 1 m2 (Ramage, 1995), and do not form the extensive
beds often seen in more sheltered habitats such as tidal estuaries. The intertidal platforms
upon which Z. novazelandica occurs are frequently exposed to high energy oceanic swells and storm waves (Rasmussen, 1965), thus exposing the seagrass patches to physical disturbance.
The mud crab Mucrophthalmus hirtipes (Brachyura: Ocypodidae) occurs within the
patches of Z. novazelundica on the Kaikoura Peninsula. This crab is endemic to New
Zealand, most commonly found below the mid-tide level of mud flats of harbours, lagoons and estuaries inhabiting temporary, water-logged burrows. It is in these habitats that such aspects of the biology of M. hirtipes such as burrowing behaviour, feeding,
habitat preferences, population structure and reproduction have been investigated (Nye, 1974; Fielder and Jones, 1978; Simons, 1981; Simons and Jones, 1981; Hawkins and
Jones, 1982; Jones and Simons, 1982; McLay, 1988). However, it is only through the presence of Z. novazelandica that M. hirtipes can populate the Kaikoura Peninsula, as seagrass patches are the only source of permanent sediment on the intertidal platforms.
Due to the isolated nature of individual seagrass patches, the habitat for M. hirtipes is

C.M.C. Woods, D.R. Schiel I J. Exp. Mar. Biol. Ecol. 214 (1997) 49-65 51
more restricted and marginal than that of mudflats in which it usually occurs. Given that
previous studies of M. hirtipes occurring on mudflats have revealed it to be a deposit
feeder (Beer, 1959; Fielder and Jones, 1978), the restriction of crabs to seagrass patches
could affect the diet of M. hirtipes. In this study, we examine the previously uninvestigated use of Z. novazelandica as a resource by M. hirtipes, and the conse- quences of this to seagrass patches on rocky intertidal platforms.
2. Methods
2.1. Study sites
The study was done from May 1994 to July 1995 on the Kaikoura Peninsula on the
central east coast of the South Island of New Zealand (43”25’S, 173”42’E). This area is at the northernmost position of the Subtropical Convergence (Heath, 1985) and is
frequently exposed to high energy oceanic swells and storm waves. The range of annual
sea temperatures is 8.5”C to 19°C (Ottaway, 1976). Two study sites, approximately 3 km
apart on opposite sides of the peninsula, were used (Fig. 1). These two sites are the only locations on the Kaikoura Peninsula possessing significant concentrations of seagrass.
Wairepo Flats has a northeasterly aspect and is composed of an extensive siltstone
platform which gradually slopes down to a subtidal fringe of large brown algae such as Cystophora sp., Corphophyllum maschaloccupum and Macrocystis pyrifera. Intertidally,
extensive mats of turfing coralline algae and the fucalean alga Hormosira banskii are interspersed with seagrass patches and bare siltstone patches dominated by limpets and
trochid gastropods. Mudstone Bay has a southerly aspect and is similarly composed of an extensive, gradually sloping siltstone platform. The algal and animal assemblages are
similar to those at Wairepo Flats. More silt collects at Mudstone Bay, however, due to
the shallower sublittoral bed nearby (Rasmussen, 1965).
Ii5 ii00 IkE ii00
.35’
40’S
.45”
B P
8 P
Al hiudstone Bay Flats
Fig. I. Map of New Zealand showing the location of the Kaikoura Peninsula and the two study sites.

52 C.M.C. Woods, D.R. Schirl I J. Exp. Mar. Biol. Ecol. 214 (1997) 49-6.5
2.2. Sampling methods
Because of the sensitivity of seagrass to disturbance, and the apparent state of decline observed in a number of patches around Kaikoura (Ramage, 199.5) a non-destructive
method of quantifying use of seagrass as a habitat by M. hirtipes was necessary. Warren ( 1990) found that for another ocypodid crab, Heloecius cordiformis, inhabiting
mangrove forests, counts of open burrows provided a quick and reliable estimate of
apparent crab abundance. In May 1994, to determine whether the number of burrows visible on the surface of a patch of seagrass was a reliable indicator of the actual number of crabs inhabiting the seagrass, ten seagrass patches at each site were chosen at random
for sampling. A 0.25 m* quadrat was randomly placed within each patch and the number of visible burrows was counted. The area within the quadrat was then rapidly dug out
down to the bedrock and placed in a high-walled sieve (diameter of sieve = 60 cm, mesh
size = 2 mm). The sediment was washed away from the seagrass and the number of
crabs remaining was counted. The crabs collected were sexed to obtain an estimate of
the sex ratio at each site. For ten of these 0.25 m2 quadrats (five at each site), the vertical
depth of a single burrow selected at random was determined by careful dissection of the seagrass before sieving, measured with a 1 m stainless steel rule and compared with the
vertical depth of the seagrass itself. In June 1994, transects were run perpendicular to the shore at both sites, from the
mean high tide mark to mean low tide, at horizontal intervals of 4 m, along the entire
area of each study site (Wairepo Flats = 900 m wide, Mudstone Bay = 450 m wide). The
number of seagrass patches encountered 2 m either side of the transect and the presence or absence of burrows in these patches were recorded. To obtain an estimate of overall
burrow density, 30 seagrass patches at each site were selected at random and a 1 m’
quadrat placed at random within the patch. The number of burrows within each quadrat
was then recorded.
The spatial distribution of burrows within seagrass patches was determined by randomly selecting ten large-sized ( > 2 m*) patches at each site. For each patch, three
0.0625 m’ quadrats were randomly placed and burrows were counted within each of four zones on a patch: O-30 cm from the patch edge, 30-60 cm from the patch edge, 60-90 cm from the patch edge, and 90- 120 cm from the patch edge. As large seagrass
patches usually border tide pools along some of their perimeter, the sampling procedure was carried out on both tide pool, and non-tide pool regions of the patches. The
percentage cover of seagrass blades was estimated from a grid placed in each quadrat. To examine the morphological characteristics of the burrows constructed by M.
hirtipes, casts were made of 30 burrows from different patches, 15 at each site. Patches were selected at random, then a random allocation was made as to whether a burrow at the edge (O-30 cm from edge) or a non-edge ( > 30 cm from edge) burrow would be
cast. The casts were made when the patches were exposed at low tide using FFFF casting grade Plaster of Paris. Prior to casting, water within the burrows was extracted using a 50 ml syringe with flexible tubing attached which was inserted as far as it would go into the burrow. A liquid slurry of plaster was then poured into the burrows, and the burrows marked with a white upright plastic marker pushed into the seagrass and left for 24 h before the hardened caste was carefully dug out and extracted from the seagrass. Once extracted from the seagrass, the following measurements and observations were

C.M.C. Woods, D.R. Schiel I J. Exp. Mar. Biol. Ecol. 214 (1997) 49-G 53
recorded for each burrow: greatest vertical depth, total burrow length (length of all
entrances and passageways combined), number of crabs per burrow as well as each
crabs’ sex and size, average burrow diameter (mean taken from diameter measured at
burrow entrance, mid-point and end-point), number of entrances, and finally, number of passageways per burrow.
The persistence of crab burrows was determined by placing a 0.25 rn’ quadrat within
each of ten randomly selected patches at each site. The quadrats were marked with plastic corner stakes. A gridded quadrat (100 X 5 cm squares) was laid within the
marked area and the position of all existing burrows was mapped. Every 7 d for a further 1 I weeks, the marked areas were examined and all burrows were mapped.
In August 1994, the effect of blade density on burrowing was determined by selecting I.5 sections (approximately 35 cm X 18 cm X 10 cm deep) which were carefully dug out
intact from large seagrass patches ( > I m’). Five sections had approximately 100% seagrass blade cover, five had 50% blade cover, and five had no blade cover. Once dug
out, these sections were immediately transferred to the laboratory and placed in aquaria
(35 cm X I8 cm X I8 cm), covered with 5 cm of fresh seawater, and provided with a bubbled air supply. One crab was placed into each aquarium and the time from initiation
of burrow construction to time of burrow completion was recorded. To determine the effect of crabs on the rate of patch erosion, each of five patches
possessing crab burrows at each site had a white upright plastic marker placed in the centre in December 1994. Five compass bearings were randomly selected, and the
distance from the plastic marker to the edge of the patch along each of these compass
bearings was measured. The total number of crab burrows present in the patch was recorded, and blade density of the patch was estimated using a 0.25 m’ gridded quadrat. After 6 months, the distance along each compass bearing from the plastic marker in
centre of each patch to the edge of the patch was measured, and number of burrows and blade density were recorded. This treatment included a range of burrow densities from
l-48 crab burrows per patch. For comparison, tive patches that did not possess crab
burrows at each site were treated as above. Sixty crabs were collected from each site for gut content analysis from May to June
1994. Upon capture, crabs were immediately placed in 10% formalin and taken to the laboratory where they were sexed and the carapace width was measured using vernier
callipers (20. I mm). The foregut of each crab was dissected out, opened along the mid-ventral line, and the contents washed out into a gridded (100 X 4 mm” squares) petri
dish. Foregut contents were estimated as the percent volume each different type of food item contributed to the total volume of the foregut contents as estimated by the total
number of 4 mm’ squares covered by the foregut contents.
3. Results
3.1. Occupation qfsseagrass patches by crabs
The number of crab burrows visible at the surface of seagrass patches was positively correlated with actual number of crabs present, both at Wairepo Flats (ux = 0.954, P < O.OOI), and at Mudstone Bay (r8 = 0.853, P < 0.01). For the rest of the study a I: 1
54 C.M.C. Woods, D.R. Schiel I J. Exp. Mar. Biol. Ecol. 214 (1997) 49-65
crab to burrow ratio was assumed. The sex ratio of male to female crabs was 1: 1.1 both
at Wairepo Flats (n = 31) and at Mudstone Bay (n = 19), which do not vary significantly from an even ratio (x2 = 0.04, n.s.). The depth of burrows was related to the depth of
seagrass patches (rx = 0.90, P < 0.001 ), which in large patches can reach 36 cm between the surface of the patch and the hard reef below it.
A total of 2 141 seagrass patches around the Kaikoura Peninsula were examined, 1349
(63%) of which possessed crab burrows. At Wairepo Flats, 65.8% of seagrass patches were occupied by crabs compared to 57.7% at Mudstone Bay. The occupation of
burrows was not dependent on site (x2 = 13.7, P > 0.05). There was no relationship between the number of crab burrows and seagrass patch size (rZx = 0.15, P > 0.05) or
patch depth (Ye = 0.34, P > 0.05). However, there was a negative correlation between
the number of burrows and the density of seagrass blades on patches (r2R = - 0.64, P < 0.01). The overall density of crab burrows per m* was 9.73 (2 1.24) at Wairepo
Flats and 9.1 (2 I .7) at Mudstone Bay.
3.2. Variation within seagrass patches
An ANOVA with the factors Site (Wairepo vs Mudstone), Pool (adjacent or not adjacent to tide pool), Distance (four zones within seagrass patches), Patches within
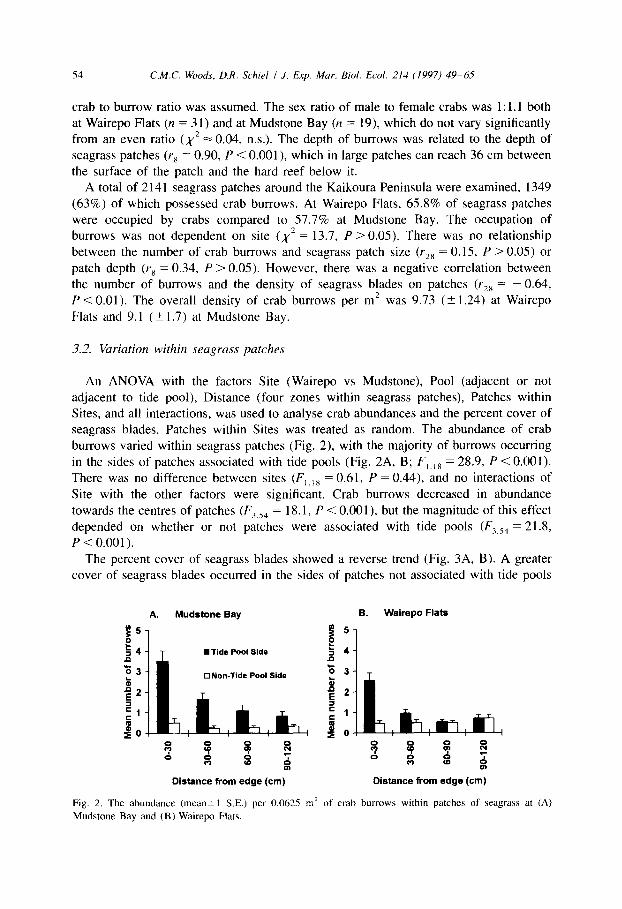
Sites, and all interactions, was used to analyse crab abundances and the percent cover of seagrass blades. Patches within Sites was treated as random. The abundance of crab burrows varied within seagrass patches (Fig. 2) with the majority of burrows occurring
in the sides of patches associated with tide pools (Fig. 2A, B; F,,,, = 28.9, P < 0.001).
There was no difference between sites (F,,,, = 0.61, P = 0.44), and no interactions of
Site with the other factors were significant. Crab burrows decreased in abundance
towards the centres of patches (F,,,, = 18.1, P < O.OOl), but the magnitude of this effect
depended on whether or not patches were associated with tide pools (F,,,, = 21.8,
P < 0.001).
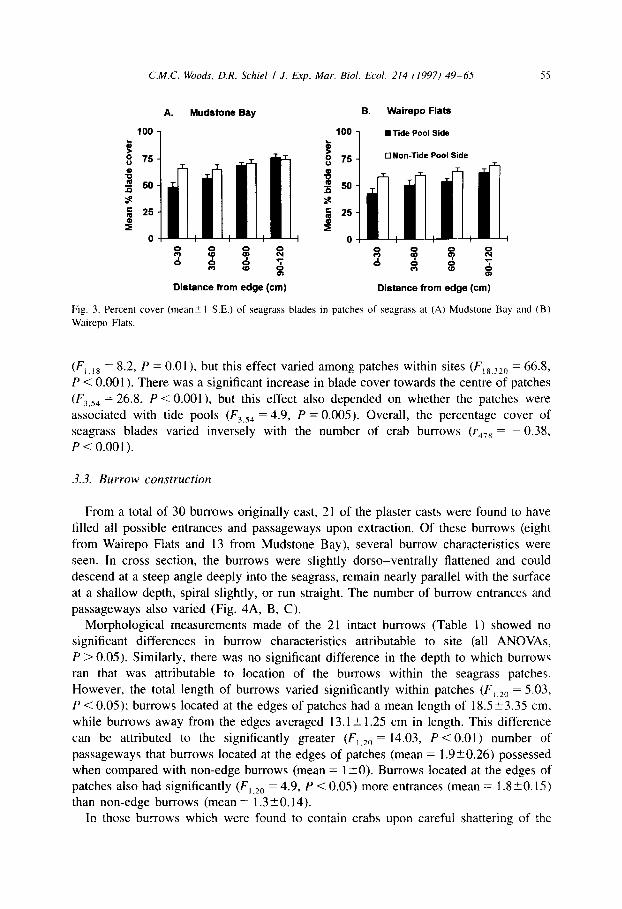
The percent cover of seagrass blades showed a reverse trend (Fig. 3A, B). A greater cover of seagrass blades occurred in the sides of patches not associated with tide pools
A. Mudstone Bay B. Wairepo Flats
Distance from edge (cm) Distance from edge (cm)
Fig. 2. The abundance (mean-+1 S.E.) per 0.06215 m’ of crab burrows within patches of seagrass at (A)
Mudstone Bay and (B) Wairepo Flats.
C.M.C. Woods, D.R. Schiel I J. Exp. Mar. Biol. Ecol. 214 (1997) 49-65 55
A. Mudstone Bay 0. Wairepo Flats
100 , HTide Pool Side t $ 75 0 Non-Tide Pool
8 z5Il
ae 2 25
% 0
Distance from edge (cm) Distance from edge (cm)
Fig. 3. Percent cover (mean21 S.E.) of seagrass blades in patches of seagrass at (A) Mudstone Bay and (B)
Wairepo Flats.
(F ,,,8 = 8.2, P = 0.01) but this effect varied among patches within sites (F,8,32,j = 66.8,
P < 0.001). There was a significant increase in blade cover towards the centre of patches
(Fj.S4 = 26.8, P < O.OOl), but this effect also depended on whether the patches were associated with tide pools (F,,,, = 4.9, P = 0.005). Overall, the percentage cover of seagrass blades varied inversely with the number of crab burrows (Y~,~ = - 0.38,
P<O.OOl).
3.3. Burrow construction
From a total of 30 burrows originally cast, 21 of the plaster casts were found to have
filled all possible entrances and passageways upon extraction. Of these burrows (eight
from Wairepo Flats and 13 from Mudstone Bay), several burrow characteristics were
seen. In cross section, the burrows were slightly dorso-ventrally flattened and could
descend at a steep angle deeply into the seagrass, remain nearly parallel with the surface
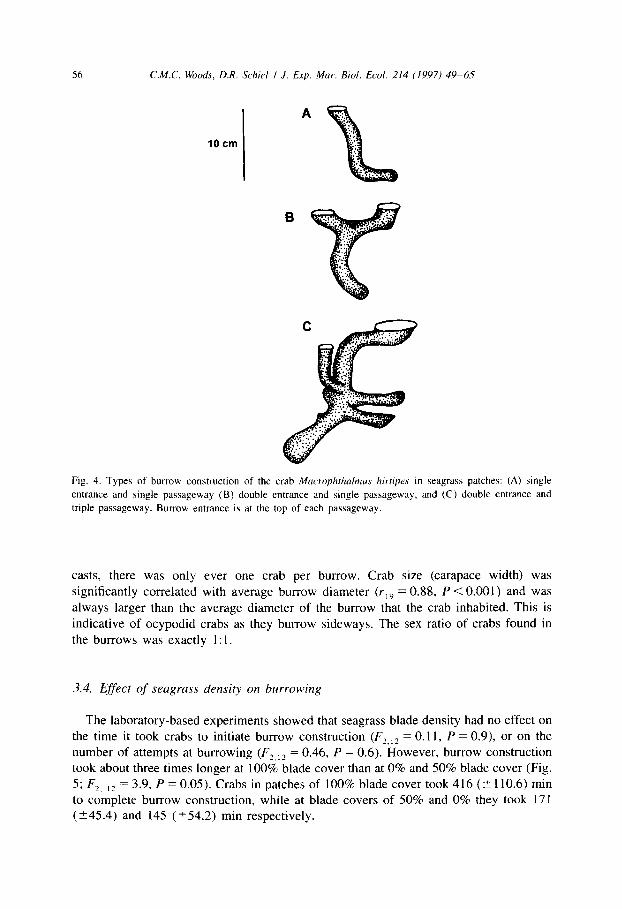
at a shallow depth, spiral slightly, or run straight. The number of burrow entrances and passageways also varied (Fig. 4A, B, C).
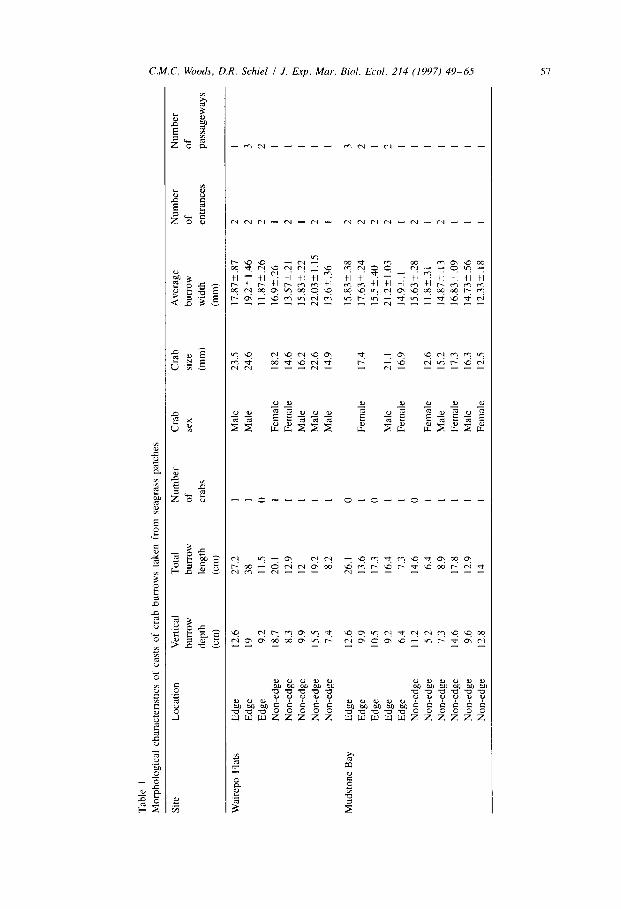
Morphological measurements made of the 21 intact burrows (Table 1) showed no
significant differences in burrow characteristics attributable to site (all ANOVAs,
P > 0.05). Similarly, there was no significant difference in the depth to which burrows ran that was attributable to location of the burrows within the seagrass patches.
However, the total length of burrows varied significantly within patches (F,,,, = 5.03, P < 0.05); burrows located at the edges of patches had a mean length of 18.5k3.35 cm, while burrows away from the edges averaged 13.1 k 1.25 cm in length. This difference
can be attributed to the significantly greater (F,,,, = 14.03, P < 0.01) number of
passageways that burrows located at the edges of patches (mean = 1.9kO.26) possessed when compared with non-edge burrows (mean = 1+-O). Burrows located at the edges of patches also had significantly (F,,,, = 4.9, P < 0.05) more entrances (mean = 1.8’_0.15) than non-edge burrows (mean = 1.3-C0.14).
In those burrows which were found to contain crabs upon careful shattering of the
56 C.M.C. Woods, D.R. Schiel I J. Exp. Mar. Biol. Ecol. 214 (1997) 49-6-r
IO cm
Fig. 4. Types of burrow construction of the crab Macrophrhalmus hirriprs in seagrass patches: (A) single
entrance and single passageway (B) double entrance and single passageway, and (C) double entrance and
triple passageway. Burrow entrance is at the top of each passageway.
casts, there was only ever one crab per burrow. Crab size (carapace width) was
significantly correlated with average burrow diameter (r,9 = 0.88, P < 0.001) and was always larger than the average diameter of the burrow that the crab inhabited. This is indicative of ocypodid crabs as they burrow sideways. The sex ratio of crabs found in
the burrows was exactly 1: 1.
3.4. effect of seagrass density on burrowing
The laboratory-based experiments showed that seagrass blade density had no effect on the time it took crabs to initiate burrow construction (F,,,, = 0.11, P = 0.9), or on the
number of attempts at burrowing (FZ,lS = 0.46, P = 0.6). However, burrow construction took about three times longer at 100% blade cover than at 0% and 50% blade cover (Fig.
5; F*. IZ = 3.9, P = 0.05). Crabs in patches of 100% blade cover took 416 (I? 110.6) min to complete burrow construction, while at blade covers of 50% and 0% they took 171 (k45.4) and 145 (k54.2) min respectively.
Tab
le
I M
orph
olog
ical
ch
arac
teri
stic
s of
ca
sts
of
crab
bu
rrow
s ta
ken
from
se
agra
ss
patc
hes
Site
L
ocat
ion
Ver
tical
T
otal
N
umbe
r C
rab
Cra
b A
vera
ge
Num
ber
Num
ber
burr
ow
burr
ow
of
sex
size
bu
rrow
of
of
+
dept
h le
ngth
cr
abs
(mm
) w
idth
en
tran
ces
pass
agew
ays
(cm
) (c
m)
“2
(mm
)
%
Wai
repo
Fl
ats
Edg
e 12
.6
27.2
I
Mal
e 23
.5
17.8
7+.X
7 2
I
Edg
e I9
38
I
Mal
e 24
.6
19.2
k1.4
6 2
3 i:
Edg
e 9.
2 I
I.5
0 11
.X7+
.26
2 2
P %
Non
-edg
e 18
.7
20.
I 1
Fem
ale
18.2
16
.9k.
26
I I
Non
-edg
e 8.
3 12
.9
I Fe
mal
e 14
.6
13.5
7+.2
1 2
I 5
Non
-edg
e 9.
9 I2
I
Mal
e 16
.2
1583
2.22
I
I P
Non
-edg
e IS
.5
P 19
.2
I M
ale
22.6
22
.03+
I.15
2
I
Non
-edg
e 7.
4 8.
2 I
Mal
e 14
.9
13.6
2.36
I
I $
Mud
ston
e B
ay
Edg
e 12
.6
26.
I 0
1583
%.3
X
2 3
9
Edg
e 9.
9 13
.6
I Fe
mal
e 17
.4
17.6
32.2
4 2
2 e
Edg
e IO
.5
17.3
0
15.s
-t.4
0 2
I !?
Edg
e 9.
2 16
.4
I M
ale
21.1
21
.2+1
.03
2 2
e
Edg
e 6.
4 7.
3 I
Fem
ale
16.9
14
.92.
1 I
I 2
Non
-edg
e I
I.2
14.6
0
15.6
32.2
8 2
I
Non
-edg
e 5.
2 6.
4 I
Fem
ale
12.6
11
.X+.
31
I I
z
Non
-edg
e 7.
3 8.
9 I
Mal
e IS
.2
14.x
7t.1
3 2
I 9
Non
-edg
e 14
.6
17.8
I
Fem
ale
17.3
16
.83-
c.09
I
I ?
Non
-edg
e 9.
6 12
.9
I M
ale
16.3
14
.73%
.56
I I
e N
on-e
dge
12.8
I4
I
Fem
ale
12.5
12
.33-
t.18
I I
58 C.M.C. Woods, D.R. Schiel I J. Exp. Mar. Biol. Ecol. 214 (1997) 49-65
2 600 -
E g 500 - P $ 400 - E
g 300 - 3
: zoo- 2 s loo-
E i= 0,
0 50 100
Percent blade cover (%I
Fig. 5. Time in minutes (mean? I S.E.) taken to complete burrow construction in laboratory-based experiments
using seagrass of 0, SO and 100% blade cover.
3.5. Persistence of burrows and effects qf crabs on patch erosion
The occupation of burrows is not permanent and over time burrows become filled in
with loose sediment except when the edges of patches have been eroded away exposing some portion of the burrow, allowing water movement to flush the burrows clear of
sediment. Following each high tide period, crabs in occupied burrows were observed to
clear away sediment that had begun to fill in their burrows. At both sites, mapped
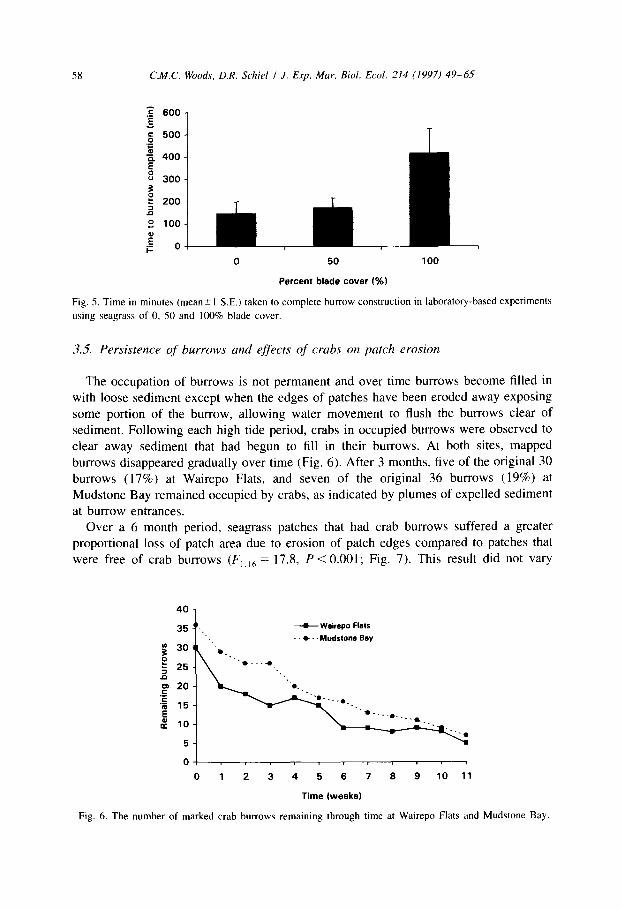
burrows disappeared gradually over time (Fig. 6). After 3 months, five of the original 30
burrows (17%) at Wairepo Flats, and seven of the original 36 burrows (19%) at Mudstone Bay remained occupied by crabs, as indicated by plumes of expelled sediment
at burrow entrances. Over a 6 month period, seagrass patches that had crab burrows suffered a greater
proportional loss of patch area due to erosion of patch edges compared to patches that
were free of crab burrows (F, ,,6 = 17.8, P < 0.001; Fig. 7). This result did not vary
--c Weirspo Flats
. . Mudstone
04 I I I I I I I b I I I
0123456769 10 11
Time (weeks)
Fig. 6. The number of marked crab burrows remaining through time at Wairepo Flats and Mudstone Bay.
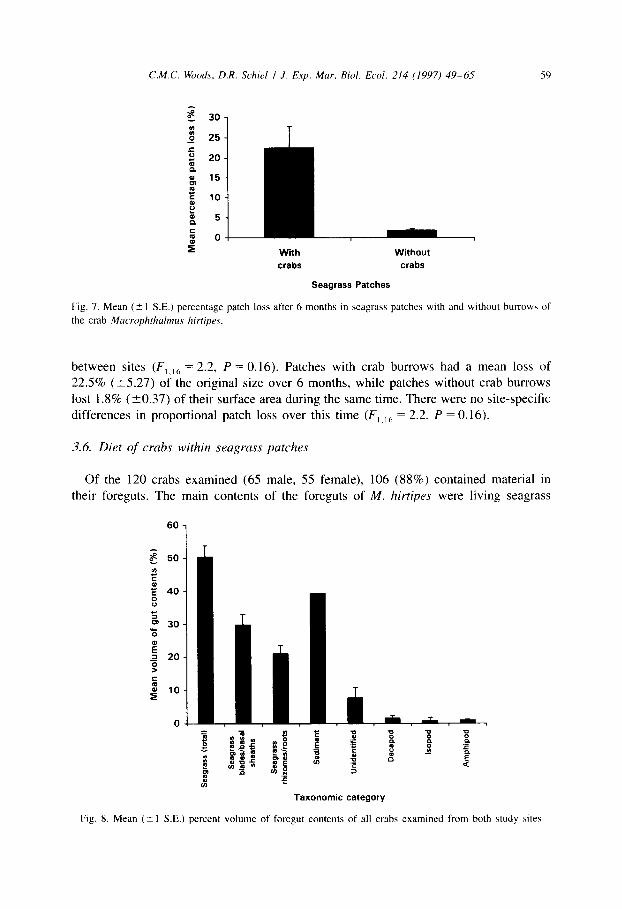
C.M.C. Woods, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 214 (1997) 49-6.~ 59
With Without
crabs crabs
Seagrass Patches
Fig. 7. Mean (2 I S.E.) percentage patch loss after 6 months in seagrass patches with and without burrow of
the crab Macrophthalmus hirriprs.
between sites (F, ,6 = 2.2, P = 0.16). Patches with crab burrows had a mean loss of
22.5% (t5.27) of the original size over 6 months, while patches without crab burrows
lost 1.8% (t0.37) of their surface area during the same time. There were no site-specific differences in proportional patch loss over this time (F, ,,h = 2.2, P = 0.16).
3.6. Diet of crabs within seagrass patches
Of the 120 crabs examined (65 male, 55 female), 106 (88% ) contained material in
their foreguts. The main contents of the foreguts of M. hirtipes were living seagrass
Taxonomic category
Fig. 8. Mean (2 1 SE.) percent volume of foregut contents of all crabs examined from both study bites.
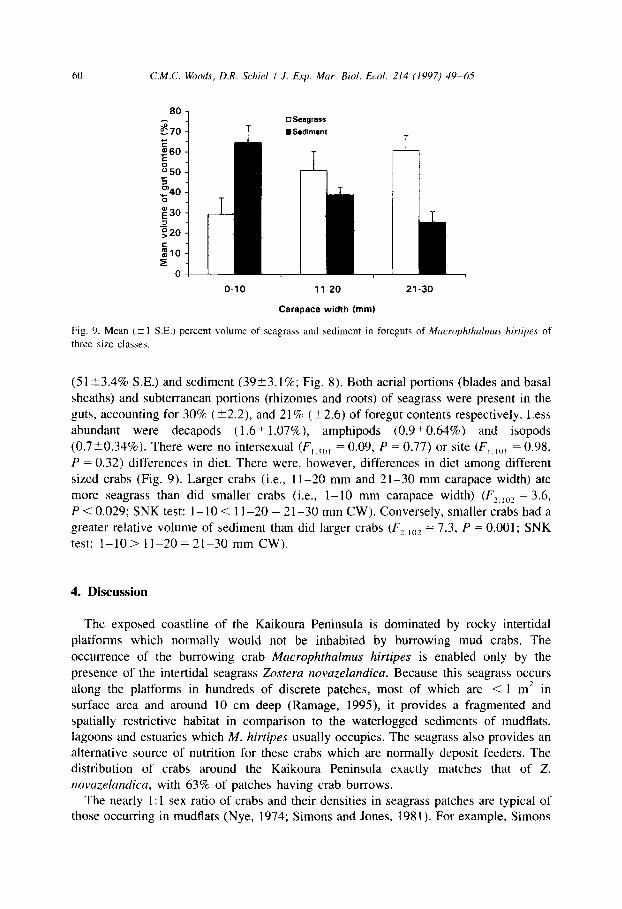
60 C.M.C. Woods, D.R. Schiel I J. Exp. Mar. Biol. Ecol. 214 (1997) 49-13
q seagrsss T @Sediment
i I O-10 11-20
Carapace width (mm)
‘c 21-30
Fig. 9. Mean (t 1 SE.) percent volume of seagraha and sediment m foreguts of Mucrophthulmus hirtipes of
three size classes.
(51?3.4% SE.) and sediment (3923.1%; Fig. 8). Both aerial portions (blades and basal
sheaths) and subterranean portions (rhizomes and roots) of seagrass were present in the
guts, accounting for 30% (-+2.2), and 21% (k2.6) of foregut contents respectively. Less abundant were decapods (1.6-C 1.07%) amphipods (0.9?0.64%) and isopods
(0.7t0.34%). There were no intersexual (F,.,,, = 0.09, P = 0.77) or site (F,,,,,, = 0.98,
P = 0.32) differences in diet. There were, however, differences in diet among different sized crabs (Fig. 9). Larger crabs (i.e., 1 l-20 mm and 21-30 mm carapace width) ate more seagrass than did smaller crabs (i.e., l- 10 mm carapace width) (FZ,,02 = 3.6,
P < 0.029; SNK test: 1- 10 < 1 I-20 = 2 l-30 mm CW). Conversely, smaller crabs had a greater relative volume of sediment than did larger crabs (F~,,,Iz = 7.3, P = 0.001; SNK test: l-10> 1 l-20 = 21-30 mm CW).
4. Discussion
The exposed coastline of the Kaikoura Peninsula is dominated by rocky intertidal platforms which normally would not be inhabited by burrowing mud crabs. The
occurrence of the burrowing crab Macrophthalmus hirtipes is enabled only by the presence of the intertidal seagrass Zostera novazelandica. Because this seagrass occurs
along the platforms in hundreds of discrete patches, most of which are < 1 m2 in surface area and around 10 cm deep (Ramage, 1995), it provides a fragmented and spatially restrictive habitat in comparison to the waterlogged sediments of mudflats, lagoons and estuaries which M. hirtipes usually occupies. The seagrass also provides an alternative source of nutrition for these crabs which are normally deposit feeders. The distribution of crabs around the Kaikoura Peninsula exactly matches that of Z. novazelandica, with 63% of patches having crab burrows.
The nearly 1: 1 sex ratio of crabs and their densities in seagrass patches are typical of those occurring in mudflats (Nye, 1974; Simons and Jones, I98 1). For example, Simons

C.M.C. Woods, D.R. Schiel I J. Exp. Mm. Biol. Ecol. 214 (1997) 49-65 61
and Jones ( 198 1) reported crab densities ranging between 11-41 per m2 in an estuary in
southern New Zealand, and Stephenson (1970) reported crab densities of 11 per m2 in a
central NZ mudflat. At our study sites, the overall densities were around 9 per m2, but
the greatest densities reached > 40 per m2 on the tide pool side of seagrass patches.
According to Beer (1959); Fielder and Jones (1978), M. hirripes is mainly a deposit feeder that uses its hair-lined chelipeds to sieve through the sediment and concentrate organic matter before it is transferred to the mouthparts. It is unable to maintain prolonged recirculation of branchial fluid when exposed to air (Hawkins et al., 1982; Hawkins and Jones, 1982). It seems likely, therefore, that crabs construct their burrows
along patch edges bordering tide pools because it allows them to remain submerged
within tide pools when the seagrass patches and surrounding bedrock are exposed to air during low tide. Indeed, the only crabs observed foraging during low tide at Kaikoura
were found in the tide pools bordering seagrass patches (personal observation). On
mudflats and estuaries, however, M. hirtipes has been observed feeding on the surface
exposed to air at low tide (Fielder and Jones, 1978; Williams et al., 1985). An influencing factor in the abundance of crabs in the tide pool edges of seagrass
patches is the reduced density of seagrass blades along the margins of tide pools. The laboratory-based experiments showed that crabs can burrow into these areas of reduced blade cover 2-3 times faster than into the areas of patches with a dense cover of
seagrass blades. Blade cover and crab abundance clearly interact, however, because
seagrass forms a major component of the diet of crabs. The burrowing behaviour of M. hirtipes in seagrass in the field and laboratory, the
occupation of burrows by single crabs, and the physical characteristics of burrows are
very similar to those described by Nye (1974) in a mudflat habitat. Crab burrows were
usually simple in construction with a single entrance, a passageway with no elaborate
galleries, and were occupied by only one crab. In branching burrows, at least one of the
branches was found to open into tide pools at the edge of the seagrass patch. Except for deep ( > 30 cm deep) seagrass patches, the depth to which crab burrows extended was
restricted to the depth to which the seagrass ran before meeting the underlying bedrock. At our study sites burrow depth averaged 11.5 cm (k4.1 S.D.), compared to 6 cm (5 1 S) in mudflats (Nye, 1974). Once the bedrock was encountered, the subsequent path
the burrow took depended on the contours of the bedrock. The burrows of M. hirtipes in seagrass patches lasted longer than the transient
burrows in mudflats. Nye ( 1974) found that, for burrow entrances on mudflats, only 11%
were in the same position one day later, while none was in the same position one week later. Relatively nomadic in nature, M. hirtipes does not have a strong affinity to a
burrow and will sometimes wander away to construct a new one, even though it may
have just defended its previous burrow (Beer, 1959). In contrast, the seagrass burrows of M. hirtipes were relatively persistent, some being in their original position after 3
months. This implies that crabs have a longer residence time in seagrass, perhaps
because of the lesser amount of suitable habitat available coupled with the energetic cost of burrow construction.
Burrows are constantly being filled in with waterborne sediment or with sediment loosened from the walls of the burrow interior. This requires regular expulsion of sediment to maintain a clear passageway (Nye, 1974; Fielder and Jones, 1978; Jones and

62 C.M.C. Woods, D.R. Schiel I .I. Exp. Mar. Biol. Ecol. 214 (1997) 49-65
Simons, 1982). The difference in burrow persistence between seagrass patches and
mudflats may arise because the interconnecting rhizomes and roots reduce cave-ins of burrow walls. Although the maintenance of burrows by crabs may be reduced, the burrowing activity is to the detriment of seagrass patches. Crabs affect patch stability by reducing the binding effects of the interweaving rhizome/root network. In our study, this was shown to accelerate the erosion of occupied seagrass patches by a factor of 12
compared to patches that were free of crabs. Studies elsewhere have shown similar
effects. In the northern Gulf of Mexico, for example, stone crabs (Menippe spp.) had a
similar detrimental effect on the turtlegrass Thalassia testudinum (Valentine et al., 1994). Over a 7 month period, twice as much recession of seaward edges of turtlegrass
occurred where stone crabs had constructed their burrows.
The burrowing of M. hirtipes along the margins of seagrass patches probably exacerbates patch erosion more so than if burrows were mainly constructed within the
interiors of seagrass patches. The edges of seagrass patches are most prone to erosion through water movement over reef platforms (Ramage, 1995). Once patches begin to decline along their edges, the interiors of burrows are soon exposed. This creates a
greater surface area for further erosion and the area immediately surrounding the burrow is rapidly undermined and torn away by water movement.
Along with a variety of other organisms such as echinoids, limpets, gastropods, parrot
fish, turtles, water fowl, dugongs and manatees that use seagrasses as a source of food
(King, 1981; Greenway, 1995), some crabs have also been identified as grazers of seagrasses. For example, Caine (1980) found that out of 21 specimens of the crab
Pugettia producta collected from San Juan Island, Washington, USA, living seagrass
(Zostera marina) blades accounted for 23% of foregut contents while Perez and
Bellwood (1988) found small quantities of seagrass in the foregut of the crab Matuta
lunaris. In our study, M. hirtipes was both a deposit feeder and a grazer of seagrass, eating aerial and sub-surface portions. Since about 20% of its food is comprised of
seagrass rhizomes and roots, much of the feeding activity of M. hirtipes probably occurs within its burrows. This may be related to the maintenance of burrows, which are free of protruding rhizomes and roots.
In contrast to studies on mudflats, where deposit feeding is the main source of food
for M. hirtipes (Beer, 1959; Fielder and Jones, 1978), the amount of inorganic material
consumed by crabs in our study was second to seagrass consumption. In the field and in
the laboratory, M. hirtipes showed both the deposit-feeding behaviour described by Fielder and Jones (1978) and active grazing of seagrass blades. Crustaceans, such as
amphipods, isopods, and conspecifics formed only minor parts of the diet. Although
necrophagy may contribute to its feeding, M. hirtipes is capable of active predation and has been observed capturing the isopod Isocladus armatus in the field (pers. obs.).
The blade coverage of seagrass patches affects invasion by crabs. Patches with a dense cover of seagrass blades have sediments below them that are firmly consolidated by rhizomes and roots. Laboratory experiments showed that this combination con- siderably lengthens the time it takes crabs to construct burrows. Few of the robust and
healthy-looking patches have crabs inhabiting them. However, patches are temporally variable in size and blade cover, which provides suitable times for invasion by crabs. Harrison (1987) found that when established plots of the seagrass Zostera marina were denuded in summer on Roberts Bank, south-west coast of British Columbia, Canada, the

C.M.C. Woods, D.R. Schiel I J. Exp. Mar. Biol. Ecoi. 214 (1997) 49-65 63
burrowing shrimp Callianasa californiensis was able to burrow into the sediment of the
seagrass plots from which they had previously been excluded because of the blade
coverage. The density of the blade cover of seagrass patches at Kaikoura varies
seasonally, with the lowest density in winter (July-August). This coincides with the period of most severe wave action when patch mortality is greatest and when the majoi erosion of patches occurs (Ramage, 1995). Therefore, the majority of patches have at least a few months per year when they are susceptible to invasion by crabs. This
invasion probably occurs laterally from patches already occupied with crabs, or from larval settlement (newly settled juveniles around 2-3 mm carapace width were occasionally observed hidden under shell fragments and detritus on the surface of
seagrass patches). Invasion probably does not occur from subtidal sources, as snorkelling
and SCUBA dives did not reveal any subtidal populations.
Furthermore, many patches are affected by a wasting disease reportedly caused by the
myxomycete Labyrinthula macrocystis Cienk, as seen across Europe and North America in the 1930s (Armiger, 1964). Labyrinth&a was seen in the seagrass around the
Kaikoura Peninsula (Ramage, 1995) and on another platform in southern New Zealand
(personal observation). Labyrinthula may be a normal secondary composer present in healthy seagrasses (Den Hartog, 1987) and not primarily responsible for the deteriora- tion of seagrass patches. Patches often recover with the growth of new blades, but the
overall effect is that for several months of the year blade cover is drastically reduced and
patches are more vulnerable to possible invasion by crabs and, consequently, to severe erosion.
The physical and biotic factors leading to patch erosion are compounded by
anthropogenic influences, which can be great along accessible intertidal platforms. Foot
and vehicular traffic such as motorbikes and four wheel drive vehicles were seen to
damage patches, which can take years to recover. For example, in another study Zieman (1976) found that seagrass damaged by boat propellers took 2-5 years to recover.
The crab M. hirtipes is an opportunist in rocky intertidal seagrass patches, extending
its range into the only sediment-filled and relatively stable habitat available on rocky platforms. It is probably the most important animal affecting seagrass patch dynamics on rocky platforms in southern New Zealand. Although crabs do not inhabit the smallest
patches forming in shallow tide pools or tidal cracks, they occupy a high proportion of established patches on the rocky platforms. There was no discernible effect of crabs in
the recruitment of new patches, but their major influence in seagrass dynamics is the
destabilisation and partial mortality of established patches. While patches are healthy, with a dense cover of seagrass blades, the crab probably has little effect on them.
However, during times of physical or biotic stress the crabs can contribute significantly
to patch deterioration, primarily through the disruptive effect their burrows have on seagrass patch stability.
Acknowledgments
Thanks to J. vanBerke1 for assistance, to the Zoology Department and Edward Percival Marine Station, University of Canterbury, for facilities. The research was possible through the Public Good Science Fund of New Zealand, grant numbers UOC

64 C.M.C. Woods, D.R. Schiel I .I. Exp. Mar. Biol. Ecol. 214 (1997) 49-6.5
3 18 and UOC 412; we gratefully acknowledge their support. Thanks also to two anonymous reviewers for helpful comments.
References
Armiger, L.C., 1964. An occurrence of Labyrinthula in New Zealand Zostera. N.Z. J. Bot., Vol. 2 (I), pp. 3-9.
Beer, C.G., 1959. Notes on the behaviour of two estuarine crab species. Trans. Royal Sot. N.Z.,Vol. 86 (3/4),
pp. 197-203.
Bishop, N., 1992. Natural history of New Zealand. Hodder and Stoughton, Auckland, New Zealand, 199 pp.
Caine, E.A., 1980. Ecology of littoral species of Caprellid Amphipods (Crustacea) from Washington, USA.
Mar. Biol., Vol. 56, pp. 327-335.
Dawes, C.J., I98 I. Marine Botany. John Wiley, New York, 628 pp.
Dawes, C.J., D. Hanisak and J.W. Kenworthy, 1995. Seagrass biodiversity in the Indian river lagoon. Bull.
Mar. Sci., Vol. 57 (I), pp. 59-66.
Den Hartog, C., 1987. ‘Wasting disease’ and other dynamic phenomena in Zostera beds. Aquat. Bot., Vol. 27,
pp. 3-14.
Duarte, C.M. and K. Sand-Jensen, 1990. Seagrass colonisation: patch formation and patch growth in
Cymodocea nodosa. Mar. Ecol. frog. Ser., Vol. 65, pp. 193-200.
Edgar, G.J., 1990. Predator-prey interactions in seagrass beds. II Distribution and diet of the blue manna crab
Portunus pelagicus Linnaeus at Cliff Head, western Australia. J. Exp. Mar. Biol. Eco/.,Vol. 139, pp. 23-32.
Fielder, D.R. and M.B. Jones, 1978. Observations of feeding behaviour in two New Zealand mud crabs (He/ice
crassa and Macrophthalmus hirtipes). Mauri Ora, Vol. 6, pp. 41-46.
Ginsberg, R.N. and H.A. Lowenstam, 1958. The influence of marine bottom communities on the deposition
environment of sediments. J. Geol., Vol. 66, pp. 310-3 18.
Greenway, M., 1995. Trophic relationships of macrofauna within a Jamaican seagrass meadow and the role of
the echinoid Lytechinus variegatus (Lamarck). Bull. Mar. Sci., Vol. 56 (3) pp. 7 19-736.
Harrison, P.G., 1987. Natural expansion and experimental manipulation of seagrass (Zostera spp.) abundance
and response of infaunal invertebrates. Estuar. Coast. Shelf. Sci., Vol. 24, pp. 7999812.
Hawkins, A.J.S. and M.B. Jones, 1982. Gill area and ventilation in two mud crabs, Helice crassa Dana
(Grapsidae) and Macrophthalmus hirtipes (Jacquinot) (Ocypodidae), in relation to habitat. J. Exp. Mar.
Biol. Ecol., Vol. 60, pp. 103-l 18.
Hawkins, A.J.S., M.B. Jones and I.D. Marsden, 1982. Aerial and aquatic respiration in two mud crabs, He&e
crassa Dana (Grapsidae) and Macrophthalmus hirtipes (Jacquinot) (Ocypodidae). Comp. Biochem. Physiol.,
Vol. 73a, pp. 341-347.
Heath, R.A., 1985. A review of the physical oceanography of the seas around New Zealand- 1982. N.Z. J. Mar.
Freshwater Res.,Vol. 19, pp. 19-124.
Humm, H.J., 1964. Epiphytes of the seagrass Thalassia testudinum, in Florida. Bull. Mar. Sci., of the Gulf and
Caribbea., Vol. 14, pp. 306-341.
Jones, M.B. and M.J. Simon% 1982. Habitat preferences of two estuarine burrowing crabs Helice crassa Dana
(Grapsidae) and Macrophthalmus hirtipes (Jacquinot) (Ocypodidae). J. Exp. Mar. Biol. Ecol., Vol. 56, pp.
49-62.
Kikuchi, T., 1974. Japanese contributions on consumer ecology in eelgrass (Zostera marinu L.) beds, with
special reference to trophic relationships and resources in inshore fisheries. Aquaculture, Vol. 4, pp.
145-160.
King, R.J., 1981. Marine Angiosperms: Seagrasses. In, Marine Botany: an Australasinn Perspective, edited by
M.N. Clayton and R.J. King, Longman Cheshire, Melbourne, pp. 201-210.
McLay, C.L., 1988. Crabs of New Zealand. Leigh Laboratory Bulletin 22, University of Auckland, 463 pp.
Nye, P.A., 1974. Burrowing and burying by the crab Macrophthalmus hirtipes. N.Z. J. Mar. Freshwater Res.,
Vol. 8 (2), pp. 243-254.
Ottaway, J.R., 1976. Inshore sea temperatures at Kaikoura, New Zealand 1973-1975. Mauri Ora, Vol. 4, pp.
69-73.

C.M.C. Woods, D.R. Schiel I J. Exp. Mur. Biol. Em/. 214 (1997) 49-65 65
Perez, OS. and D.R. Bellwood, 1988. Ontogenetic changes in the natural diet of the Sandy Shore Crab, Mtrrura
/unaris (Forskal) (Brachyura: Calappidae). Aust. J. Mar. Fre.shwater Res., Vol. 39, pp. 193- 199.
Philippart, C.J.M., 1995a. Seasonal variation in growth and biomass of an intertidal Zrjsrera n&ii stand in the
Dutch Wadden Sea. Neth. J. Sea. Res., Vol. 33 (2). pp. 205-218.
Philippart. C.J.M., 1995b. Effects of shading on growth, biomass and population maintenance of the intertidal
seagrass Zosterci nolrii Hornem in the Dutch Wadden Sea. J. Exp. Mm. Biol. Ecol., Vol. 188, pp. 199-213.
Ramage, D., 1995. The patch dynatnics and demogruphy of Zosteru novozelundica Serchell on the intertidal
p/a$wm.s qf rhe Kaikouru Peninsula. Unpublished M.Sc. thesis, University of Canterbury, 95 pp.
Rasmussen, R.A.. 1965. The inter&d ecology of the rocky shores of the Koikmm Peninsula. Unpublished
Ph.D. thesis, University of Canterbury, 203 pp.
Rosas, C., E. Lazaro-Chavez and F. Buckle-Ramirez, 1994. Feeding habits and food niche segregation ot
Cullirzectes .wpidu.\, C. rarhhunue, and C. .simi/is in a subtropical coastal lagoon of the Gulf of Mexico. J.
Crust Biol., Vol. I4 (2) pp. 371-382.
Simons, M.J., 1981. Relative growth in the New Zealand mud crab Macrophrhalmus hirtipes (Brachyura:
Ocypodidae). N.Z. J. Mar. Freshwater Res., Vol. 15, pp. 193-200.
Simons, M.J. and M.B. Jones, 1981. Population and reproductive biology of the mud crab, Macrophthalmus
hirtipes (Jacquinot, 1853) (Ocypodidae), from marine and estuarine habitats. J. Nat. Hisf., Vol. IS, pp.
98 I-994.
Stephenson, G., 1970. The distribution of two crab species on the Porirua Harbour mud-flats. Bull. Nut. Sci.
Victoria Univ. Wellington. Vol. I, pp. 21-26.
Valentine, J.F., K.L. Heck Jr., P. Harper and M. Beck, 1994: Effects of bioturbation in controlling turtlegrass
(Thalussicz tesrudinum Banks ex Konig) abundance: evidence from field enclosures and observations in the
Northern Gulf of Mexico. J. Exp. Mar. Biol. Ecol., Vol. 178, pp. 18l- 192.
Virnstein, R.W., 1995a. Seagrass landscape diversity in the Indian River Lagoon, Florida: the importance of
geographic scale and pattern. Bull. Mar. Sci., Vol. 5 I (I). pp. 67-74.
Virnstein, R.W., 1995b. Anomalous diversity of some seagrass-associated fauna in the Indian River Lagoon,
Florida. Bull Mar. Sci., Vol. 57 (I ). pp. 75-78.
Warren, J.H., 1990. The use of open burrows to estimate abundances of intertidal estuarine crabs. Aust. J.
Ewl., Vol. 15, pp. 277-280.
Wassenberg, T.J., 1990. The seasonal feeding on Zosreru cupricnrni seeds by juvenile Penaeu.s esculentus
(Crustacea: Decapoda) in Moreton Bar, Queensland. Ausr. J. Mar. Freshwafer Res., Vol. 41. pp. 301-310.
Webb, C., P. Johnson and B. Sykes, 1990. Flowering planrs of NeM: Zecrland. Caxton Press, Christchurch, 146
PP. Williams, B.G., E. Naylor and T.D. Chatterton, 1985. The activity patterns of New Zealand mud crabs under
lield and laboratory conditions. J. Exp. Mu. Biol. Ecol., Vol. 89, pp. 269-282.
Zieman, J.C., 1976. The ecological effects of physical damage from boat motors on turtlegrass beds in
southern Florida. Aquur. Rot., Vol. 2, pp. 127-139.