u~oqs say s~eal luam oupnp %u!pq!oa eas-daap sdma palpq jo...
TRANSCRIPT

-8u!paa3 uop~a leas-daap 01 uo!p~depe Iyssmns 04 pal a~~y sau!g heuopnlona Ou!8ran!p Qap!~ arow 30 om aep!ssaue!sLq ay) u!ql!fi ~~yl snoinqo S! 11 -sapads eas-daap omt pue sapads jjaqs auo uaawaq uospvdwm a Lq pa1saBns S! a3ero~s 103 uo!le~depe leu -!pal@ a~!ssa~%o~d a snua8 auo u~ -saug a~!snpxa d~~enlnu~ pue lua~ajj~p dpla~dwcm OM] %UO@ tnq %uuols pooj afe3s aSq ~oj paldeple S! pnea hluauqle aql sapads Ile q 'saug aualaN!p Ouop A~lua~adde pua walled podfidura puo!l!pva ay1 wog Leme suo!leulrojsuaq ~m!i!o~oydrour gu!y~ea~-~e~ ol pal wy uourea a3q ~J!M %uyeap JOJ
uopmy!pow saIq!puew aql u~ -em$ $lays pue eas-daap paaepr uaawaq wadsax srya ul aana~a~jrp snoyqo d~ale!paurtu! ou S! aIau -spod!ydtue Jaqlo lsow U! ueyl padola~ap rallaq 'uo!lez!pol pooj hrosuasoway~ 203 uoyeldepe p~auaO B se uaas aq ue3 aepu -nalue p!sseule!sAi ay] uo sne8ro hrosuasowaq~ JO ~uawa%uvrra ~spads ayct .wajsAs Ilresuaru!p aqa pue ~uawd~nba fiosuasouray~ aql uy ay~!u 1eqOopa 1e1n39led qaql JOJ suo!leldepe agq!ssod 03 padsal yqh pa!pnls uaaq a~ey 3!3!3ed aql pue 3!lu~pv ay3 wog sp!sseue!sAl gas-daap 3u!paa~-uorr~a~ an5 uo!le%!gsa~u! luasald aql UI .aep!s -seuqslC~ Lp~j ay) 01 Suo~aq sde~l eas-daap q~ns U! pauIlelqo Ll~e1n8al spod!qdwe 1~ 'leyqey ~esdqe ay1 U! s~apaaj-uo!rre:, aueu!wop aql aq 01 spodlydwe pue saqsg u~oqs say s~eaL luam Oupnp %u!pq!oa eas-daap u! sdma palpq jo asn aqsualxx

l. introductory remarks
The introduction in the late 1960's of the baited camera and the re-introduction in the early 1970's of the baited trap as means for studying deep sea epibenthic carni- vores have brought spectacular results. Amphipods have been found to play a leading role in dealing with offered baits, especially at deeper leveIs and are now being treated in a rapidly growing literature (Hessler et al. 1972, Shulenberger and Hessler 1974, Shulenberger and Barnard 1976, Hessler et al. 1978, Thurston 1979).
The carrion feeders, occurring in great numbers, constitute an element of the deep-sea fauna, the quan- titative and metabolic importance of which is only now becoming gradually appreciated. For obvious reasons studies of the ecology of the relevant species are dif- ficult, most of the evidence being inevitably cir- cumstantial. The aim of the present paper is to discuss some aspects of functional amphipod morphology, in- terpreted as adaptations for deep sea carrion feeding.
2. Material and methods
I have had at my disposal deep-sea carrion feeding am- phipods from the Biscayan abyssal plain and the deep Norwegian Sea belonging to the species Eurythenes gryllus (Lichtenstein), Paralicella tenuipes Chevreux, Orchomene (Orchomenopsisj chevreuxi (Stebbing) and a further apparently undescribed species of Orchomene (s.str.). Also Prof. Robert R. HessIer provided me with a number of specimens of Hirondellea gigas from the Philippine Trench. For comparisons with shallow-water carrion feeders specimens of Orchomene (Orchornenopsis) obtusa (G. 0. Sars) and Tmetonyx cicada (Fabricius) from the Swedish and Norwegian west coasts were studied.
Specimens were fixed in ethanol, formaline or al- coholic Bouin, and for EM studies in Karnowski's fluid. For laboratory study various standard histological methods as well as TEM and SEM techniques were applied. Clearing of preserved specimens in clove oil proved useful for studying gut contents and gross gut morphology.
3. Results
3.1. Some general aspects of deep sea carrion feeding in amphipods
Apart from more or less accidental findings reported by Chevreux (1900, 1935) from the extensive and suc- cessful trapping operations carried out during the cruises of prince Albert I of Monaco, all deep-sea am- phipods caught in baited traps belong to the family Lysianassidae. The lysianassids are typical gammarid amphipods conforming 'in all essentials to the basic functional model described by Dahl (1977). Many of
them are scavengers in shelf habitats and a high per- centage of the deep-sea species belong to the family.
Deep-sea carrion feeders occupy a very special ecoIogical niche. To be successful there they need a number of adaptations meeting certain requirements necessary for a feeding strategy based on large meals taken at more or less long time intervals and randomIy dispersed over vast areas of uniform ocean bottom. The most important of these adaptations would seem to be: (1) Ability to localize and to recognize potential food. (2) Ability to feed on large and muscular food items. (3) Ability to consume within a short period large quantities of food and to store that food for gradual utilization during a long time interval. Additional advantages could be gained by using at least temporarily food of lower quality e.g. from the bottom deposit (Hessler et al. 1978) and to maintain a low metabolic rate as shown by Smith and Hessler (1974) for certain deep water fishes.
3.2. Localization of food
Chemosensory stimulation would seem to be the most likely agency by which amphipods are led to carrion on the deep sea bottom. It shodd be noted, however, that the question of long range chemicaI stimulation in water raises theoretical difficulties related i. a. to turbulence (Wilson 1970, Crisp .1974) which have not yet been overcome. The only alternative to chemical stimulation would seem to be mechanical stimulation caused by the rapid sinking of a comparatively -heavy body and its impact on the bottom. Although stimdi of that type are rapidly and widely spread through the water, it is dif- ficult to see how they could produce an attraction of longer duration. As shown by Hessler et al. (1978) and Thurston (1979) amphipods are still attracted to bait after 24 h, and this would seem to point definitely to chernosensory attraction.
Lysianassid amphipods are adapted for efficient chemoreception of stimuli carried by water currents. As shown for Gammarus pulex (L.) (Dah'i 1977) part of the current produced by the beating of the pleopods of the resting animal passes over the proximal part of the an- tennulae, sweeps over the mouth-parts and enters the branchial region behind them. This is the case also in the lysianassids and here one finds a particularly dense accumulation of chemosensorytype setae on the ventral side of the first flagellar article of the antennula (Figs 1 and 2). Also lysianassids in most cases have short and stout antennulae which are normally kept somewhat depressed. This leads to an increased exposure to chemosensory stimuli, and could be seen as a special lysianassid adaptation for chernosensory orientation. Also when swimming the lysianassid sensory apparatus will be strongly exposed to chemical stimuIi in the water through which the amphipod passes.
Electron rnicroscopical investigations of antennular setae of Eurythenes gryllus fixed on board did not give

Fig. 1. Antennula of Orchomene (Orchomenopsis) chevreuxi (Stebbing) in ventral view showing dense tufts of presumably chemosensory setae. x 130.
Fig. 2. Parasagittal section of terminal part of peduncte and proximal part of flagellurn of antennulae in 0. chevreuxi. On upper left base of accessory flagellurn. Ventrally rows of setae bases and above them sensory cell nuclei and dense bundles of nerve fibres. X 150.
technically perfect results. Nevertheless, fixation was good enough to show that the sensory hairs of Euryrhenes are closely similar to those described for Orchesria platerzsis Krayer and Talorchestia deshayeesi Audouin by Dahl (1973), the hairs of Eurythenes also having a terminal sensory pore and 15 to 18 cilia per hair.
3.3. Mandibles
Thurston (1979) pointed out the efficiency of the in- cisor parts of the mandibles in ParaIicella and
- -
12 OIKOS 33:2 (1979)
Euryrhenes and noted that their molars, in contrast to those of the Orchomene species, are nontriturative.
I studied the mandibles of all the five deep sea carrion feeders listed above and can contribute some further observations.
The basic gammarid amphipod mandibte in all prob- ability is of the type found in the genus Gammarus and most other gammarid families (Barnard 1969). It has a strongly and somewhat irregularly serrated incisor part and a well-developed lacinia mobiris on both mandibles. Biting conforms to the standard peracarid pattern (Fryer 1964, Dahl and Hessler unpubl.), with the left

Fig. 3. Ventral view of Iabmm and mandibles in Hirondellea gigas Birstein and Vinogradov. Posterior mouth-parts partly removed. Mandibles nearly closed but shearing mode of biting evident. la labrum, Im left mandible rm right mandible. X 60.
incisor passing anteriad of the right one which in its turn enters the space between it and the left lacinia mobilis, which comes to lie anteriad of the right lacinia. Thus, a double interlocking of four serrate biting elements occurs, two of which, the laciniae, are weaker than the others.
Within the family Lysianassidae most genera have smooth incisor parts with a short edge. There exists a tendency to reduction of the right lacinia. The molar is highly variable, but in some genera, e.g. Hippomedon, Pseudalibrom and Onesimus it is triturative.
Thurston (1979) points out that the incisor edge of the mandible in carrion feeding amphipods tends to be- come long and sharp. Of the genera examined this applies in the first place to Eurythenes, Hirondellea and Paralicella. Biting at least in Eurythenes and Hirondel- lea, probably also in Paralicella, follows a pattern differ- ent from that described above (Dahl and Hessller un- publ.). The right incisor glides along a IOW oblique edge traversing the posterior side of the left incisor (Figs 3,4, 5). The right lacinia has disappeared but the left one, which is usually long and slender and also in a somewhat unusual position in relation to the row of setae which here replace the spine-row, can be presumed to stabilize the right incisor in its effective position. This is the in- ference drawn from the arrangement of the various elements (the opaqueness and general arrangement of mouth-parts preclude all direct observations on Iiving animals).
The corpus mandibulae of all the three genera men- tioned above has become bowl-shaped. The represen- tatives of all three genera cut out large morsels of food
and this bowl-shape will permit them to take larger bites than a more flattened mandible where a piece of meat bit into by the incisor edges might impede their closing and effective shearing (Figs 3,4,6,7). This transforma- tion of the corpus mandibulae is facilitated by the elongation of the cutting incisor edge.
The non-triturative molars of Eurythenes gryllus and Hirondellea gigas appear to have acquired a new func- tion and are of a very distinctive shape, particularly in the latter species. There the molar tends to become transformed into a long, smooth and tapering semitubular process which reaches into the opening of the stomodaeum (Figs 4, 5). When the molars close upon each other the two molar processes with their setose edges form a more or less complete funnel ap- parently guiding larger food particles when they are being sucked up into the stomodaeum. For obvious reasons it has not been possible to study this process in the living animals but the morphology is highly sugges- tive;
In higher magnification the surface of the molar elongations in H. gigas turns out to be composed of various layers of closely packed fibres (Fig. 8).
In Paralicella tenuipes the incisor edge is strikingly Iong and sharp. The molar is elongated to form a long flap-like structure (Shulenberger and Barnard 1976, Fig. 4) the function of which is not clearly understood although it could resemble that suggested for Euryfhenes and Hirondellea.
The mandible of Orchomene differs from those de- scribed above in being more slender with a shorter and blunter incisor edge, a flat instead of a bowl-shape cor-
OIKOS 33:2 (1979)

Figs 4-5. Left and right mandibles of Hiror~dellea gigas. Note in this as in the other species the sharp incisor edge (i) the oblique ledge (01) on the left but not the right mandible, the presence of a lacinia rnobilis ( / c ) only on the left mandible, and the bowl-shapecl corpus mandibulae ( c m ) . Note also the long semitubular molar (tno). X 60. Fig. 6. Right mandible of Elrryrheitrs grjll~cr (Lichtenstein). Note general similarity to H. gigas but less extreme development of molar. X 60. Fig. 7 . Left mandible of PuraliceNa te~rrcipes Chevreux. Note in this comparatively small species t h e very long incisor edge, and the deep bowl of the corpus rnandibttlae. X 130.
12* OIKOS 33:2 (1979) 171
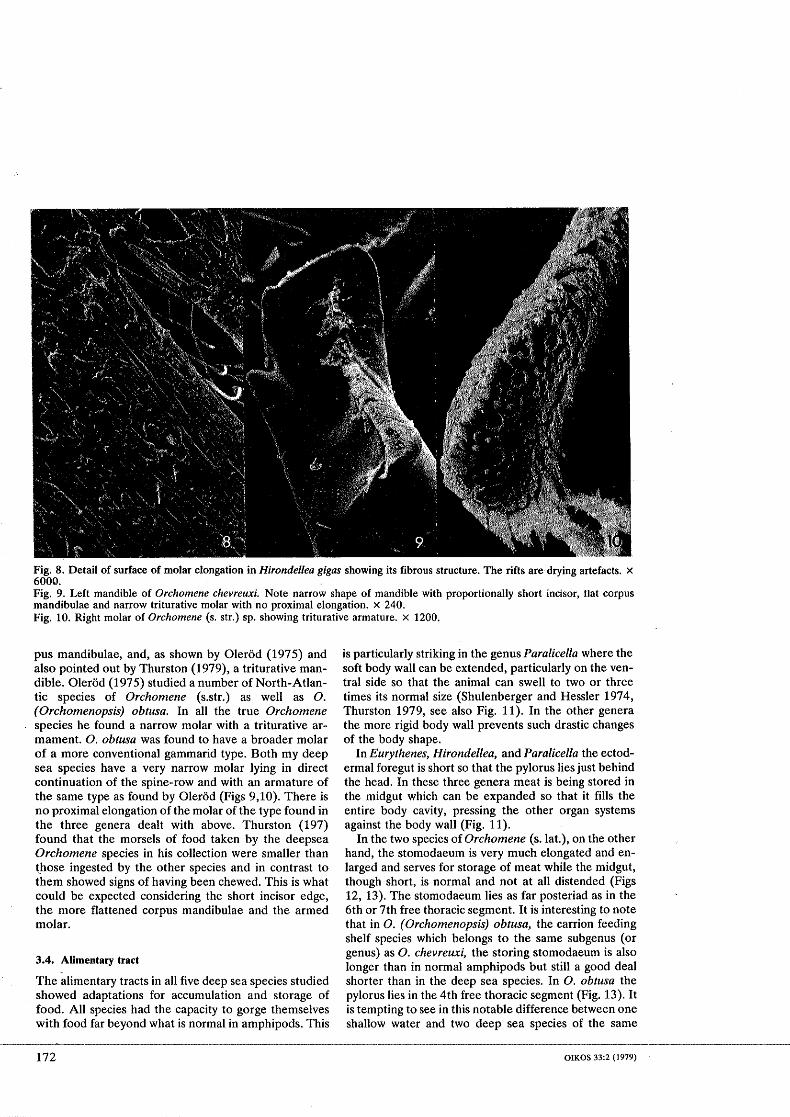
Fig. 8. Detail of surface of molar elongation in Hirondeliea gigas showing its fibrous structure. The rifts are drying artefacts. X 6000. Fig. 9. Left mandible of Orchomene chevreuxi. Note narrow shape of mandible with proportionally short incisor, flat corpus mandibulae and narrow triturative molar with no proximal elongation. X 240. Fig. 10. Right molar of Orchomene (S. str.) sp. showing triturative armature. X 1200.
pus mandibulae, and, as shown by Olerod (1975) and also pointed out by Thurston (19791, a triturative rnan- dible. Olerod (1 975) studied a number of North-Atlan- tic species of Orchomene (S.&.) as well as 0. (Orchornenopsis) obtusa. In all the true Orchomene
. species he found a narrow molar with a triturative ar- mament. 0. obfusa was found to have a broader molar of a more conventional gammarid type. Both my deep sea species have a very narrow molar lying in direct continuation of the spine-row and with an armature of the same type as found by Olerud (Figs 9,lO). There is no proximal elongation of the molar of the type found in the three genera dealt with above. Thurston (197) found that the morsels of food taken by the deepsea Orchomene species in his collection were smaller than [hose ingested by the other species and in contrast to them showed signs of having been chewed. This is what could be expected considering the short incisor edge, the more flattened corpus mandibulae and the armed molar.
3.4. ALimentary tract
is particularly striking in the genus Paralicella where the soft body wall can be extended, particularly on the ven- tral side so that the animal can swell to two or three times its normal size (Shulenberger and Hessler 1974, Thurston 1979, see also Fig. 11). In the other genera the more rigid body wall prevents such drastic changes of the body shape.
In Eurythenes, Hirondellea, and Paralicella the ectod- errnal foregut is short so that the pylorus lies just behind the head. In these three genera meat is being stored in the rnidgut which can be expanded so that it fills the entire body cavity, pressing the other organ systems against the body wall (Fig. l l).
In the two species of Orchomene (S. lat.), on the other hand, the stomodaeum is very much elongated and en- larged and serves for storage of meat while the midgut, though short, is normal and not at all distended (Figs 12, 13). The stomodaeum lies as far posteriad as in the 6th or 7th free thoracic segment. It is interesting to note that in 0. (Orchomenopsi5) obtusa, the carrion feeding shelf species which belongs to the same subgenus (or genus) as 0. chevreuxi, the storing stomodaeum is also longer than in normal amphipods but still a good deal
The alimentary tracts in all five deep sea species studied shorter than in the deep sea species. In 0. obtusa the showed adaptations for accumulation and storage of pylorus lies in the 4th free thoracic segment (Fig. 13). It food. All species had the capacity to gorge themselves is tempting to see in this notable difference between one with food far beyond what is normal in amphipods. This shallow water and two deep sea species of the same
172 OIKOS 33:2 (1979)
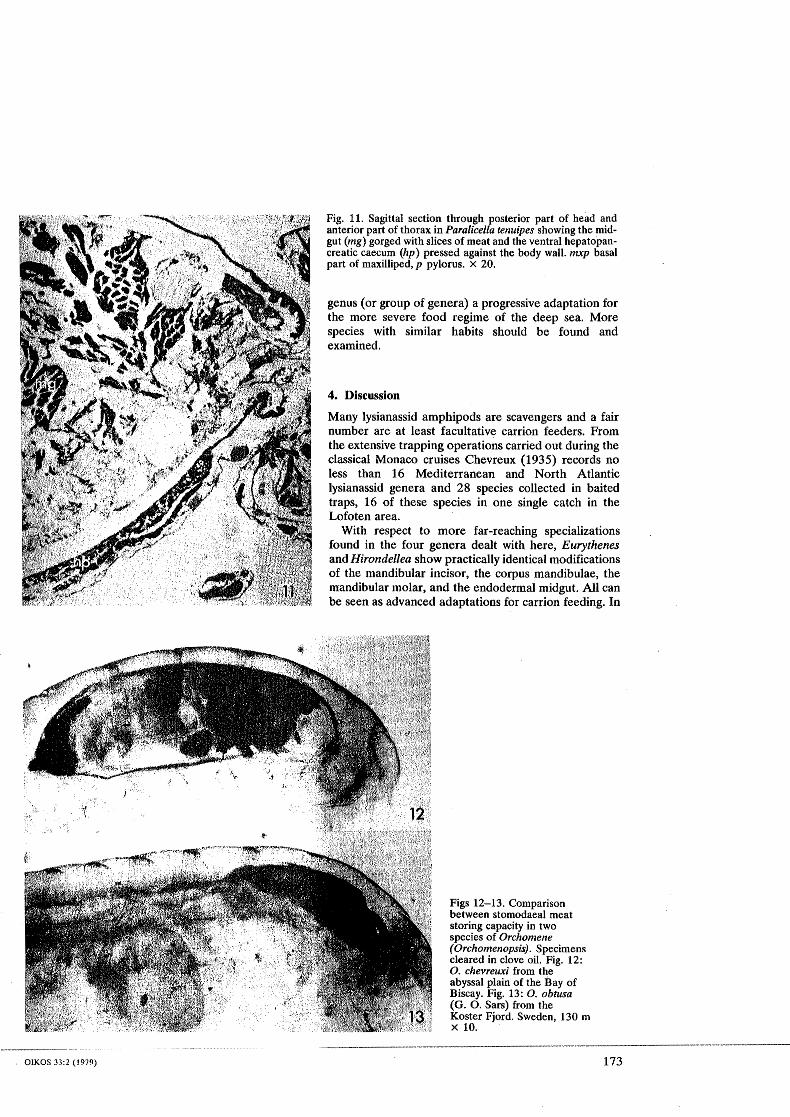
Fig. 11. SagittaI section through posterior part of head and anterior part of thorax in Paralicelfa tenuipes showing the mid- gut (mg) gorged with slices of meat and the ventral hepatopan- creatic caecum (hp) pressed against the body wall. mxp basal part of maxilliped, p pylorus. X 20.
genus (or group of genera) a progressive adaptation for the more severe food regime of the deep sea. More species with similar habits shouId be found and examined.
4. Discussion
Many lysianassid amphipods are scavengers and a fair number are at least facultative carrion feeders. From the extensive trapping operations carried out during the classical Monaco cruises Chevreux (1935) records no Iess than 16 Mediterranean and North Atlantic lysianassid genera and 28 species collected in baited traps, 16 of these species in one single catch in the Lofoten area.
With respect to more far-reaching specializations found in the four genera dealt with here, Eurythenes and Hirondellea show practically identical modifications of the mandibular incisor, the corpus mandibulae, the mandibular molar, and the endodemal midgut. All can be seen as advanced adaptations for carrion feeding. In
Figs 12-1 3. Comparison between stomodaeal meat storing capacity in two species of Orchomene (Orchomenopsis). Specimens cleared in clove oiI. Fig. 12: 0. chevreuxi from the abyssal plain of the Bay of Biscay. Fig. 13: 0. obrusa (G. 0. Sars) from the Koster Fjord. Sweden, 130 m X 10.
OIKOS 33:2 (1979) 173

Paralicella adaptations of the incisor, corpus man- dibulae, and midgut are very similar to those found in these two genera, while the molar is somewhat differ- ent.
The species of Orchomene (S. lat.) differ materially from the others. The mandible is more sIender and much flatter, the incisor edge is neither very long nor very sharp, and the molar, though narrow, is otherwise of a basic type with a triturative armament and no pro- ximal elongation. The meat-storing function is effected by the ectodermal foregut.
Although the adaptations for carrion feeding in the alimentary systems of Eurythenes, Hirondellea, and Paralicella are so similar that the genera could from a purely functional point of view be grouped together, such a grouping would have no taxonomic or evolutio- nary meaning. The family Lysianassidae is the largest of all marine gammarid families, comprising about 20% of all known marine genera (Barnard 1969). The three genera in question fall within the morphologicd frame-work of the family but are otherwise widely separated. It can hardly be doubted that their far-reaching adaptations for carrion feeding represent three separate evolutionary lines and that the remarka- ble similarities, particularly evident in the case of Eurythenes and Hirondellea, are the result of con- vergence.
The mandible of Orchomene is basic enough to per- mit it to be considered as possibly ancestral to more advanced types of carrion feeding mandibles, but the mechanism for meat storing found in Orchomene is so
fundamentally different from that found in the other three genera that they must be considered mutually exclusive, and Orchomene cannot be fitted into the lines of descent of any of the other three carrion feeding types (Fig. 14).
It must be concluded that far-reaching structural and functional morphological adaptations for deep-sea carrion feeding have evolved independently several times within the family Lysianassidae.
The niche occupied by the deep sea carrion feeders must be presumed to be old and has probably existed ever since good-sized cephalopods and fishes began to occur in greater numbers in the pelagid of the open oceans. The groups mentioned are the only ones which are common and widespread enough and simultane- ously composed of members with high enough body weight and density to produce through rapid sinking after death a sufficient and constant though scattered food supply for benthic carrion feeding macro- and megafauna species. It would seem probable that this food source has been available since Paleozoic times and it is unlikeIy that it was left unexploited for very long.
As already mentioned the carrion feeders of greater depths are exclusively or nearly exclusively amphipods. Fossil amphipods are known only since the Eocene and geologically speaking they are generally considered to be a young group. On the other hand, there is an in- creasing amount of evidence showing that the Peracarida were a well-differentiated group in the early Carboniferous and possibly still earlier (Schram 1979).
Fig. 14. Diagrammatic sketch comparing the two mutually exclusive adaptations for meat storage. Above stomodaed storage in Orchomene, below midgut storage in Paralicella (representing also Eurythenes and Hirondellea). mg midgut, hp hepatopancreatic caeca, st stomodaeum.

Within the superorder the Amphipoda assume a very isolated position, and this might indicate an earlier origin for amphipods than generally presumed.
With respect to amphipod occupation of the carrion feeding niche in the deeper parts of the World Oceans four possibilities appear to be open, viz. (1) Amphipods were present in the Paleozoic oceans and evolved carrion feeding types a very long time ago. (2) Carrion feeding amphipods evolved later and suc- ceeded earlier carrion feeders which have disappeared without leaving traces in the present fauna. (3) The food source existed for a long time before it became exploited by macro- and megafauna species. (4) The whole system is of a much later origin than suggested above. It is impossible at present to choose between these four alternatives, although the two last-mentioned ones ap- pear to be less likely. Considering the apparently great importance of carrion feeders to the dispersal of organic matter over the dep-sea bottom (Hessler et al. 1978) the problem is relevant to deep sea ecology and pa1eoecoIogy and merits further attention.
Acknowledgements - It is in the hope that the present con- tribution will interest my dear friend and colleague Per Brinck that it is dedicated to him on the occasion of his 60th birthday. I wish to thank Dr L. Laubier and Dt Robert R. Hessler for the use of vaIuable material and the staff of the Dept of Structural Zoology, Lund University, for efficient help of many different kinds.
References Bamard, J. L. 1969. The families and genera of marine Gam-
rnaridean Amphipoda. - Bdl. U. S. Natn. Mus. 271: 1-535.
Chevreux, E. 1900. Amphipodes provenant des Campagnes de l'Hirondelle (1885-1888). - Res. Camp. Sci. Albert I 16: 1-195, pl. I-XVIII.
- 1935. Amphipodes provenant des Campagnes scientifiques du Prince Albert I"' de Monaco. - Ibid. 90: 1-214, pl. I-XVI.
Crisp, D. J. 1974. Factors influencing the settlement of marine invertebrate larvae. - In: Grant, P. T. and Mackie, A. M. (ed.), Chemoreception in marine organisms. London and New York, pp. 177-265.
DahI, E. 1973. Antenna1 sensory hairs in Talitrid Amphipods. - Acta Zool. 54: 161-171.
- 1977. The amphipod functional model and its bearing upon systematics and phytogeny. - 2001. Scripta 6: 221-228.
Fryer, G. 1964. Studies on the functional morphology and feeding mechanism of MonodeIIa argentarii Stella (Crus- tacea Thermosbaenacea). - Trans. R. Soc. Edinb. 44: 49-90.
Hessler, R. R., Isaacs, J. D., and Mills, E. L. 1972. Giant amphipod from the abyssal Pacific Ocean. - Science 175: 63 6-637.
- , Ingram, C. J., Yayanos, A. A., and Burnett, B. R. 1978. Scavenging amphipods from the floor of the PhiIippine Trench. - Deep-Sea Res, 25: 1029-1047.
Oterod, R. 1975. The mouthparts in some North Atlantic species of the genus Orchomene Boeck (Crustacea Am- phipoda). - 2001. Scripta 4: 206-216.
Schram, F. R. 1979. British Carboniferous Mdacostraca. - Fieldiana Geol. 40: I-VIII, 1-129.
Shulenberger, E. and Barnard, 3. L. 1976. Amphipods from an abyssal trap set in the North Pacific Gyre. - Crustaceans 31: 241-258.
- and HessIer, R. R. 1974. Scavenging abyssal benthic am- phipods trapped under oligotrophic Central North Pacific Gyre waters. - Mar. Biol. 28: 185-187.
Smith, K. L. and Hesder, R. R. 1974. Respiration in bentho- pelagic fishes: in situ measurements at 1230 meters. - Science 184: 72-73.
Thurston, M. H. 1979. Scavenging abyssal amphipods from the North-East Atlantic Ocean. - Mar. Biol. 51: 55-68,
Wlson, E. 0. 1970. Chemical communication within animal species. - In: Sondheimer, E. and Simeone, J. B. (ed.), Chemical Ecology. New York and London, pp. 138-188.
