universidad mayor de san...
TRANSCRIPT

UNIVERSIDAD MAYOR DE SAN ANDRÉS
FACULTAD DE CIENCIAS FARMACÉUTICAS Y BIOQUÍMICAS INSTITUTO DE INVESTIGACIONES FÁRMACO BIOQUÍMICAS
““OOPPTTIIMMIIZZAACCIIOONN DDEE MMEEDDIIOOSS DDEE CCUULLTTIIVVOO,, PPAARRAA LLAA PPRROODDUUCCCCIIOONN DDEE EENNZZIIMMAASS LLIIGGNNIINNOOLLIITTIICCAASS,, PPOORR CCEEPPAASS
FFUUNNGGIICCAASS AAIISSLLAADDAASS DDEELL AALLTTIIPPLLAANNOO BBOOLLIIVVIIAANNOO””
TTEESSIISS DDEE GGRRAADDOO PPAARRAA OOPPTTAARR EELL TTIITTUULLOO DDEE LLIICCEENNCCIIAATTUURRAA EENN BBIIOOQQUUIIMMIICCAA
POSTULANTE: PAMELA LAURA USNAYO GONZALES
LA PAZ – BOLIVIA
2007

UNIVERSIDAD MAYOR DE SAN ANDRÉS
FACULTAD DE CIENCIAS FARMACÉUTICAS Y BIOQUÍMICAS INSTITUTO DE INVESTIGACIONES FÁRMACO BIOQUÍMICAS
““OOPPTTIIMMIIZZAACCIIOONN DDEE MMEEDDIIOOSS DDEE CCUULLTTIIVVOO,, PPAARRAA LLAA PPRROODDUUCCCCIIOONN DDEE EENNZZIIMMAASS LLIIGGNNIINNOOLLIITTIICCAASS,, PPOORR CCEEPPAASS
FFUUNNGGIICCAASS AAIISSLLAADDAASS DDEELL AALLTTIIPPLLAANNOO BBOOLLIIVVIIAANNOO””
TTEESSIISS DDEE GGRRAADDOO PPAARRAA OOPPTTAARR EELL TTIITTUULLOO DDEE LLIICCEENNCCIIAATTUURRAA EENN BBIIOOQQUUIIMMIICCAA
POSTULANTE: PAMELA LAURA USNAYO GONZALES
ASESORES: ENRIQUE TERRAZAS SILES Ph.D. MARIA TERESA ALVAREZ Ph.D. ALBERTO GIMENEZ TURBA Ph.D.
LA PAZ – BOLIVIA 2007

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
"De todo éxito, por pequeño
que sea, surgirá un día un
esfuerzo más grande
que lo completará."
“Whitman”

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
2
Dedicatoria
"A mis padres, Heriberto y Susana a quienes amo
y admiro por la fortaleza que tienen, y les agradezco todo,
porque a ellos debo lo que soy hoy en día y por estar
siempre a mi lado apoyándome y confiando en mi"

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
3
Agradecimientos
A Dios por haberme dado la vida y lo mejor de este mundo mi familia.
A la Universidad mayor de San Andrés, en especial a la facultad de Cs. farmacéuticas y Bioquímicas,
por darme la oportunidad de convertirme en profesional.
A la cooperación del proyecto “Biodiversidad microbiana” del programa UMSA, ASDI-SAREC de Suecia,
ya que sin su colaboración este trabajo no se habría realizado.
A la cooperación del IFS (Internacional Foundation for Science) por la colaboración brindada.
Al Instituto de Investigaciones Fármaco Bioquímicas (IIFB) por abrirme las puertas y darme la plena
libertad de trabajar en este proyecto, por incentivarme a salir adelante cada día.
Al Instituto de Investigaciones de Productos Químicos (IIDEPROQ) y todo su personal, por acogerme en
sus instalaciones y brindarme la ayuda necesaria para realizar el presente trabajo.
Muy especialmente al Dr. Enrique Terrazas por ser un gran maestro en mi formación, por transmitirme
sus conocimientos, por la paciencia, sobre todo por haberme dado la oportunidad de realizar este trabajo.
A la Dra. Teresa Alvarez por su paciencia, apoyo, toda la ayuda que me brindo para hacer realidad este
trabajo, a pesar de ser una gran docente es una gran amiga, la cual agradezco haber encontrado.
Al Dr.Alberto Gimenez por abrirme las puertas del Instituto, por su constante apoyo, y por sacarse tiempo
para brindarme su ayuda.
Al Ing. Cristhian Carrasco quien siempre me brindo su colaboración, por toda la confianza puesta en mi,
apoyo y su sincera amistad brindada.
A la Ing. Daysy Torrico por su constante apoyo y confianza, mas aun por su amistad.
A Karen Cabero por su amistad incondicional, su ayuda permanente, y por tus palabras de aliento.
A Laura Mendoza por su ayuda desinteresada, apoyo y consejos.
A todo el personal del IIFB Dra. Greis, Dr. Avila, Dra. Magui, Dra.Flores,Crispin, a mis compañeros
Kenny, Neida, Efraín, Romina, Silvia, Luis, Juan, Lencho, Hugo, Marina, Carla, Guido, Nelson, Rosita,
gracias por todos lo momentos vividos, por la paciencia que me tuvieron, y por la amistad que nos une.
Gracias a todos mis amigos y amigas de la facultad de Bioquímica y Farmacia, de Ingenieria, a mis
amigos del CBA, y otros, que si bien no escribo cada uno de sus nombres los llevo en mi corazon, gracias
por todo el apoyo brindado y su amistad que alegra mi vida.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
4
Abreviaturas
ASDI/SAREC The Swedish International Development Agency / Swedish Agency for Research Cooperation
ABTS 2, 2’-Azinobis (3-ethylbenzthiazoline-6-sulfonic acid).2NH4-salt
BFP Bifenilos policlorados
ºC Grados centigrados
CMC Carboxi – Metil – Celulosa
DMF Dimethoxyphenol
DNS Acido 3,5 Dinitrosalicilico
g Gramos
IB Ingeniería y Bioquímica
IIDEPROQ Instituto de Investigación y Desarrollo de Procesos Químicos
IIFB Instituto de Investigaciones Fármaco Bioquímicas
IUPAC International Union of Pure and Applied Chemistry
IUB International Union of Biochemistry
KDa Kilo Daltons
L Litros
Lac Lacasa
LiP Lignina peroxidasa
M Concentración molar
MnP Manganeso peroxidasa
mM Milimolar
mg Miligramos
ml Mililitros
mm Milímetro
PB Podredumbre blanca
PCF Pentaclorofenol
PDA Papa dextrosa agar
PV Peroxidasa versátil
PHA Hidrocarburos policíclicos aromáticos
μl Microlitro
μmol Micromol
nm Nanómetro
UI Unidad Internacional

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
Índice de Contenido
1. INTRODUCCION .................................................................................................. 12
1.1 OBJETIVOS ............................................................................................................................................... 13 1.2.1 Objetivo General................................................................................................................................... 13 1.2.2 Objetivos Específicos............................................................................................................................ 13
1.3 JUSTIFICACION DE LA INVESTIGACION.- ...................................................................................... 14
2. MARCO TEORICO CONCEPTUAL.- ................................................................... 17
2.1 LA LIGNINA.-............................................................................................................................................ 17 2.1.1 Estructura y biodegradación de la molécula de lignina.- ..................................................................... 17
a. Propiedades físicas.................................................................................................................................. 19 b. Ligninas comercializadas ...................................................................................................................... 20
2.1.2 Funciones de la lignina.- ....................................................................................................................... 20
2.2 DEGRADACION DE LA LIGNINA POR HONGOS.- ............................................................................ 20 2.2.1 Hongos.- ............................................................................................................................................. 21
a. Características generales.-..................................................................................................................... 21 b. Estructura.- ............................................................................................................................................ 22 c. Hábitat.-.................................................................................................................................................. 24 d. Reproducción.-......................................................................................................................................... 24 e. Clasificación.- ........................................................................................................................................... 25 f. Fisiología de los Hongos.-........................................................................................................................ 25 g. Hongos ligninolíticos.-............................................................................................................................. 27
2.3 ENZIMAS.-.................................................................................................................................................. 27 2.3.1 Estructura y características generales.-................................................................................................ 28
a. Especificidad.-........................................................................................................................................... 28 b. Actividad Enzimática.-.............................................................................................................................. 28 c. Cinética de reacción (Michaelis-Menten).-.............................................................................................. 28 d. Temperatura.- ........................................................................................................................................... 29 e. pH.- ............................................................................................................................................................ 29 f. Clasificación Internacional de las enzimas.-............................................................................................ 29
2.3.2 Enzimas ligninolíticas.-........................................................................................................................... 30 a. Peroxidasas ligninolíticas.-....................................................................................................................... 30 b. Lacasas.-.................................................................................................................................................... 32
2.3.3 Aspectos biotecnológicos.-...................................................................................................................... 34
2.4 RECURSOS LIGNOCELULOSICOS.- ......................................................................................................... 36 2.4.1 Paja Brava.-.............................................................................................................................................. 37

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
6
2.4.2 Bagazo.- .................................................................................................................................................... 38 2.4.3 Aserrín.-.................................................................................................................................................... 38 2.4.4 Residuos de quinua real.-......................................................................................................................... 39 2.4.5 Cáscara de arroz.- .................................................................................................................................... 40
3. METODOS.- .......................................................................................................... 41
3.1 RECOLECCION DE MUESTRAS.- ............................................................................................................. 43
3.2 METODOS DE AISLAMIENTO DE CEPAS FUNGICAS.- .................................................................... 45 3.2.1. Codificación de cepas fúngicas aisladas.- .............................................................................................. 48
3.3 METODOS DE IDENTIFICACION DE CEPAS FUNGICAS.- .............................................................. 49 3.3.1 Identificación Macroscópica.- ................................................................................................................. 49 3.3.2 Identificación Microscópica.- .................................................................................................................. 50 3.3.3 Elaboración de Historial de cepas fúngicas.-....................................................................................... 52
3.4 METODOS DE DETERMINACION DE LA ACTIVIDAD ENZIMATICA.-.............................................. 52 3.4.1 Determinación Cualitativa.-..................................................................................................................... 52 3.4.2 Determinación Cuantitativa.- ................................................................................................................. 55
a. Manganeso Peroxidasa.- ........................................................................................................................ 60 b. Lignina Peroxidasa.- .............................................................................................................................. 61 c. Lacasa.- ................................................................................................................................................... 62
3.4.3 Cuantificación de las Enzimas.- .......................................................................................................... 63
3.5 OPTIMIZACION DE MEDIOS DE CULTIVO PARA LA PRDUCCION DE ENZIMAS LIGNINOLITICAS.- ............................................................................................................................................. 63
3.5.1 Medios de cultivo con variación de concentración de Cobre.- .............................................................. 64 3.5.2 Medios de cultivo con diferentes sustratos lignocelulosicos.-............................................................... 65
3.6 Determinación de Actividad Celulolítica.- .................................................................................................. 66
3.7 Cuantificación de proteínas totales.-........................................................................................................... 67
3.8 Comparación de la Actividad Enzimática Ligninolítica de las enzimas producidas frente a enzimas comerciales.- .......................................................................................................................................................... 68
3.9 Análisis estadístico.- ..................................................................................................................................... 69
4. RESULTADOS Y DISCUSION ............................................................................. 70
4.1 DETERMINACION CUALITATIVA DE LA ACTIVIDAD ENZIMATICA LIGNINOLITICA.-.............. 70
4.2. DETERMINACION CUANTITATIVA DE LA ACTIVIDAD ENZIMATICA LIGNINOLITICA.- .......... 71 4.2.1. Manganeso Peroxidasa.-........................................................................................................................ 71 4.2.2. Lignina Peroxidasa.- .............................................................................................................................. 76
4.2.3. Lacasa.- ........................................................................................................................................................81

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
4.3. OPTIMIZACION DE MEDIOS DE CULTIVO PARA LA PRDUCCION DE ENZIMAS LIGNINOLITICAS.- ............................................................................................................................................. 87
4.3.1 Medios de cultivo con variación de concentración de Cobre.- ............................................................. 87 a. Manganeso Peroxidasa.- ........................................................................................................................ 88 b. Lignina Peroxidasa.- ................................................................................................................................ 92 c. Lacasa.- ..................................................................................................................................................... 95
4.3.2. Medios de cultivo con diferentes sustratos lignocelulosicos.-.............................................................. 100 I. Paja Brava.- ............................................................................................................................................ 101
a. Manganeso Peroxidasa.- .................................................................................................................... 101 b. Lignina Peroxidasa.- .......................................................................................................................... 102 c. Lacasa.- ............................................................................................................................................... 103
II. Bagazo.- ................................................................................................................................................. 105 a. Manganeso Peroxidasa.- .................................................................................................................... 105 b. Lignina Peroxidasa.- .......................................................................................................................... 107 c. Lacasa.- ............................................................................................................................................... 108
III. Aserrín.- ............................................................................................................................................... 110 a. Manganeso Peroxidasa.- .................................................................................................................... 110 b. Lignina Peroxidasa.- .......................................................................................................................... 111 c. Lacasa.- ............................................................................................................................................... 112
IV. Residuos de Quinua Real.-.................................................................................................................. 114 a. Manganeso Peroxidasa.- .................................................................................................................... 114 b. Lignina Peroxidasa.- .......................................................................................................................... 115 c. Lacasa.- ............................................................................................................................................... 116
V. Cáscara de Arroz.- ................................................................................................................................ 118 a. Manganeso Peroxidasa.- .................................................................................................................... 118 b. Lignina Peroxidasa.- .......................................................................................................................... 119 c. Lacasa.- ............................................................................................................................................... 121
4.4. Determinación de Actividad Celulolítica.- ................................................................................................... 122
4.5. Determinación de proteínas totales.-.......................................................................................................... 123
4.6. Comparación de la Actividad Enzimática Ligninolítica de las enzimas producidas frente a enzimas comerciales.- ........................................................................................................................................................ 124
5. CONCLUSIONES ............................................................................................... 127
6. RECOMENDACIONES ....................................................................................... 131
7. BIBLIOGRAFIA .................................................................................................. 132

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
8
Índice de figuras
Figura 1. Estructura parcial de la lignina............................................................................... 19
Figura 2. Corte esquemático de un hongo de la clase basidiomiceto y orden de los agaricales. ................................................................................................................................. 23
Figura 3. Procedimiento general “Producción de enzimas ligninolíticas, a partir de cepas fúngicas del Altiplano boliviano”. ............................................................................. 42
Figura 4. Orillas del Lago Titicaca en la localidad de Llamacachi. ................................ 43
Figura 5. Orillas del Río Desaguadero en el departamento de Oruro. ........................... 43
Figura 6. Paja Brava a orillas del río Desaguadero. ............................................................ 44
Figura 7. Suelo donde crece la planta Paja Brava a orillas del lago Titicaca. ............ 44
Figura 8. Métodos de aislamiento de las cepas fúngicas................................................. 46
Figura 9. Tronco de madera que contiene cepas fúngicas, recolectada de orillas del lago Titicaca. ............................................................................................................................. 47
Figura 10. Preparación de medio Papa Dextrosa Agar (PDA). ........................................ 47
Figura 11. Cepas fúngicas en crecimiento a temperatura ambiente............................ 48
Figura 12. Morfología macroscópica de la cepa fúngica ................................................ 50
Figura 13. Identificación microscópica ................................................................................ 51
Figura 14. Morfología microscópica...................................................................................... 52
Figura 15. Preparación de medio ABTS y Poli – R. .............................................................. 53
Figura 16. Decolorización del medio sólido Poly R............................................................ 54
Figura 17. Decolorización del medio sólido ABTS. .............................................................. 54
Figura 18. Estructura generaliza de ABTS (2,2'-Azinobis (3-ethylbenzthiazoline-6-sulfonic acid).2NH4-salt)........................................................................................................... 55
Figura 19. Determinación cualitativa de la actividad enzimática ligninolítica............. 56
Figura 20. Fotografía del equipo Espectrofotómetro de absorción................................ 58

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
9
Figura 21. Medio de cultivo Mn (II) para determinación cuantitativa............................ 58
Figura 22. Determinación actividad enzimática ligninolítica por métodos espectrofotométricos. .............................................................................................................. 59
Figura 23. Reacción enzimática para MnP por oxidación de 2,6 dimetoxifenol.......... 60
Figura 24. Reacción enzimática para LiP por oxidación de ABTS. .................................. 61
Figura 25. Reacción enzimática para Lac por oxidación de ABTS.................................. 62
Figura 26. Diagrama determinación enzimática en medio de cultivo Cu (II)............... 64
Figura 27. Determinación enzimática en medios de cultivo con diferentes sustratos. 66
Figura 28. Determinación de la actividad Celulolítica. ..................................................... 67
Figura 29. Determinación de proteínas totales. .................................................................. 68
Figura 30. Determinación de la actividad enzimática de enzimas comerciales. ........ 69
Figura 31. Actividad enzimática Manganeso peroxidasa para la cepa fúngica IB-26C (Trinacrium) de los 8 medios evaluados en función de glucosa, extracto de levadura, Mn.............................................................................................................................. 73
Figura 32. Efecto de la glucosa y extracto de levadura sobre la actividad MnP para la cepa fúngica IB-26C (Trinacrium).......................................................................................... 74
Figura 33. Efecto de la glucosa y Mn sobre la actividad MnP para la cepa fúngica IB-26C (Trinacrium) ........................................................................................................................ 75
Figura 34. Actividad enzimática Lignina peroxidasa para la cepa fúngica IB-26C (Trinacrium) de los 8 medios evaluados en función de glucosa, extracto de levadura, Mn.............................................................................................................................. 77
Figura 35. Efecto de la glucosa y extracto de levadura sobre la actividad LiP para la cepa fúngica IB-26C (Trinacrium).......................................................................................... 79
Figura 36. Efecto de la glucosa y Mn sobre la actividad LiP para la cepa fúngica IB-26C (Trinacrium) ........................................................................................................................ 80
Figura 37. Actividad enzimática Lacasa para la cepa fúngica IB-29B (Blasthomyces) de los 8 medios evaluados en función de glucosa, extracto de levadura, Mn......... 83
Figura 38. Efecto de la glucosa y extracto de levadura sobre la actividad Lacasa

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
0
para la cepa fúngica IB-29B (Blasthomyces)...................................................................... 85
Figura 39. Efecto de la glucosa y Mn sobre la actividad Lacasa para la cepa fúngica IB-29B (Blasthomyces). ............................................................................................................ 85
Figura 40. Actividad enzimática Manganeso peroxidasa para la cepa fúngica IB-29B (Blasthomyces) de 8 medios evaluados en función de glucosa, extracto de levadura, Cu.............................................................................................................................. 89
Figura 41. Efecto de la glucosa y extracto de levadura sobre la actividad MnP para la cepa fúngica IB-29B (Blasthomyces). ................................................................................... 90
Figura 42. Efecto de la glucosa y Cu sobre la actividad MnP para la cepa fúngica IB-29B (Blasthomyces). ................................................................................................................. 91
Índice de tablas Tabla 1. Clasificación Internacional de las enzimas..............................................................................30
Tabla 2. Características principales de enzimas ligninolíticas ............................................................... 34
RESUMEN
Las enzimas ligninolíticas junto a las enzimas celulolíticas son importantes en los procesos de
fermentación in situ. En este trabajo se realizó la producción de enzimas ligninolíticas
Manganeso Peroxidasa (MnP), Lignino Peroxidasa (LiP) y Lacasa a partir de hongos aislados
de muestras recolectadas del Altiplano Boliviano. De un total de 31 hongos, 4 cepas fúngicas
presentaron actividad ligninolítica cualitativa determinada por Poly R-478 y ABTS. Las cepas
que presentaron mayor actividad fueron: IB-29B (Blasthomyces), IB-18Cb (Bdellospora), IB-
26C (Trinacrium), IB-15Da (Cephalosporium).
Para la optimización de la producción de estas enzimas se desarrollo un diseño factorial 23
con dos diferentes concentraciones glucosa o sustratos lignocelulósicos paja brava, bagazo,

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
aserrín, residuo de quinua, cascarilla de arroz, extracto de levadura, manganeso o cobre. La
cuantificación de enzimas ligninolíticas fue determinado espectrofotométricamente por
oxidación de ABTS y 2,6 DMF. La máxima actividad enzimática fue de 245,56 UI/L para
MnP, de 487,82 UI/L para LiP y de 287, 30 UI/L Lacasa.
Palabras Claves: Enzimas ligninolíticas, manganeso peroxidasa, lignino peroxidasa, lacasa.
ABSTRACT
The ligninolític and cellulolític enzymes are important in fermentation processes in situ. In this
thesis three kind of ligninolítics enzyme has been produced, Manganese peroxidase (MnP),
Lignin Peroxidase (LiP) and Laccase from fungi isolated using samples collected of the
Bolivian Andean High Plateau. Thirty one fungal strains were isolated, four fungal strains
showed a qualitative ligninolítica activity by Poly R-478 and ABTS. The strains that presented
greater activity were: IB-29B (Blasthomyces), IB-18Cb (Bdellospora), IB-26C (Trinacrium),
IB-15Da (Cephalosporium).
In order to optimize the production of these enzymes a factorial design 23 was developed with
two different concentrations of lignocellulósic material such as brave straw, bagasse, sawdust,
remainder of quinua, husk of rice or glucose, yeast extract, manganese or cupper.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
2
The ligninolíticas enzyme quantification was determined spectrophotométrically by oxidation
of ABTS and 2.6 DMP. The greater enzymatic activity was of 245.56 UI/L for MnP, 487.82
UI/L for LiP and 287, 30 UI/L for Lacasa.
Key words: Ligninolíticas enzymes, manganese peroxidasa, lignino peroxidasa, laccasa.
1. INTRODUCCION
Las enzimas son catalizadores biológicos, es decir, proteínas que tienen la capacidad de
acelerar ciertas reacciones químicas. En los últimos años su uso en gran cantidad de industrias
ha adquirido gran relevancia. El desarrollo de técnicas y métodos para el desarrollo de estos
procesos han sido un importante requisito para el avance en la enzimología en las últimas tres
décadas y por muchos es considerada un arte [17, 11].
Actualmente en Bolivia, la producción de enzimas viene desarrollándose por los
métodos de la biotecnología clásica que incluye dos etapas principales: fermentación, en la
que se multiplica el microorganismo productor de la enzima, y la de recuperación y
purificación, en la que se aísla la enzima y se lleva al grado de pureza adecuado para su uso,

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
3
éste último en plena proyección, cuyo pionero es el Instituto de Investigaciones Fármaco-
Bioquímicas desde el año 1999.
Diversas investigaciónes sobre producción de enzimas ligninolíticas; Lignina
peroxidasa, Manganeso peroxidasa y Lacasa, obtenidos a partir de cultivos fúngicos, han
ayudado a lograr un mayor entendimiento sobre las cualidades que éstas brindan, cuyas
aplicaciones industriales son variadas, basta mencionar algunas: relacionadas con procesos de
deslignificación, detoxificación de suelos y efluentes industriales, versátil y atractivo para
fines ambientales porque puede servir para eliminar diversos contaminantes difíciles de
degradar, como los hidrocarburos del petróleo, plaguicidas, explosivos, colorantes sintéticos
que se utilizan en la industria textil y otros compuestos que son tóxicos para la flora, fauna y el
hombre.
La finalidad del presente estudio fue investigar la capacidad de producción de enzimas
ligninolíticas por los hongos del Altiplano boliviano y encontrar mayores rendimientos de
actividad enzimática ligninolítica, mediante la optimización de condiciones de cultivo a través
del establecimiento de diseños factoriales de los diferentes componentes del medio
1.1 OBJETIVOS
1.2.1 Objetivo General
- Optimizar medios de cultivo para la producción de enzimas ligninolíticas, por
cepas fúngicas aisladas del Altiplano boliviano.
1.2.2 Objetivos Específicos
- Aislar cepas fúngicas a partir de muestras recolectadas de las regiones cercanas al
lago Titicaca y río Desaguadero a través de técnicas de aislamiento axenico.
- Identificar y caracterizar taxonómicamente las cepas fúngicas aisladas.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
4
- Seleccionar cepas fúngicas capaces de producir enzimas con actividad
ligninolítica (Manganeso peroxidasa, Lignina peroxidasa, Lacasa) a través de un
método cualitativo (Poly- R y ABTS).
- Optimizar la producción de enzimas ligninolíticas a través de un diseño factorial
23, considerando la variación de glucosa, manganeso, extracto de levadura como
nutrientes que influyen en la síntesis de enzimas ligninolíticas.
- Optimizar la producción de enzimas ligninolíticas a través de un diseño factorial
23, considerando la variación de glucosa, cobre, extracto de levadura como
nutrientes que influyen en la síntesis de enzimas ligninolíticas.
- Evaluar medios de cultivo complejos con diferentes sustratos lignocelulósicos
como paja, bagazo, aserrín, residuos de quinua, cáscara de arroz en base las
condiciones optimizadas de extracto de levadura y manganeso, para la
determinación de enzimas ligninolíticas.
-
1.3 JUSTIFICACION DE LA INVESTIGACION
La lignina en la madera confiere una protección física a la celulosa y hemicelulosa
contra el ataque enzimático. Existe una gran cantidad de organismos (bacterias y hongos) con
enzimas hidrolíticas necesarias para degradar celulosa y hemicelulosa, pero en lo referente al
ataque y mineralización de la lignina el número de organismos es mucho mas limitado. Los
únicos organismos descritos con la capacidad de degradar y mineralizar la lignina son un
grupo de basidiomicetes causantes de pudrición blanca [4]. Estos hongos, poseen un complejo
de enzimas oxidasas y peroxidasas que catalizan las primeras reacciones que rompen uniones
dentro de la compleja molécula de la lignina, generando moléculas más pequeñas [5], y luego
hay una incorporación de estos productos de degradación a los ciclos metabólicos del

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
organismo que como producto final dan CO2 [3].
Después de los polisacáridos, la lignina es el polímero orgánico más abundante en el
mundo vegetal. Es importante destacar que la lignina es la única fibra no polisacárido que se
conoce [16]. El conocimiento del proceso de biodegradación de la lignina es importante para
comprender aspectos claves del ciclo bioquímico del carbono en la Tierra, dado que la mayor
parte del carbono y la energía fijados por fotosíntesis se acumulan en la molécula de celulosa,
un polímero de glucosa que es el constituyente mayoritario de la biomasa vegetal y que está
íntimamente ligado a la lignina en las plantas [9].
Sin duda, uno de los grandes retos de la humanidad en este inicio del siglo XXI es el
convertir los procesos productivos en procesos limpios y eficientes energéticamente. Por otro
lado, se quiere tener la capacidad tecnológica para restaurar los sitios dañados
ambientalmente, La biotecnología tiene un papel importante que desempeñar en esta
transformación tecnológica. La investigación deberá estar enfocada en al utilización de nuevas
herramientas biotecnológicas para la prevención, control y remediación de las
contaminaciones ambientales [12].
Este sistema ligninolítico, ha demostrado ser muy versátil y atractivo para fines
ambientales, porque puede servir para eliminar diversos contaminantes difíciles de degradar,
como los hidrocarburos del petróleo, plaguicidas, explosivos, colorantes sintéticos que se
utilizan en la industria textil y otros compuestos que son tóxicos para la flora, fauna y el
hombre [8].
Todos los mencionados anteriormente son compuestos muy contaminantes y los
sistemas actualmente implantados para su depuración se basan en la aplicación de tratamientos
físicos o químicos, la mayoría de los cuales son caros [12]. El tratamiento biológico
representa una alternativa atractiva. En este sentido, los hongos ligninolíticos se han estudiado
como posibles agentes para tales procesos debido a que sus sistemas enzimáticos
extracelulares inespecíficos les permite degradar diversos tipos de sustancias recalcitrantes. La
capacidad de estos microorganismos para degradar distintos tipos de compuestos

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
6
contaminantes ha sido demostrada [10].
Adicionalmente, las enzimas ligninolíticas pueden ser aplicadas como indicadores del
deterioro de jugos y vinos y para la síntesis químico-enzimática de nuevos antibióticos y
alcaloides [8]. Además tienen amplias aplicaciones industriales; actualmente, las enzimas son
empleadas principalmente para la obtención de lácteos, edulcolorantes, fármacos, alimentos,
licores, detergentes, etc. [11].
Por otro lado, uno de los problemas que encara el mundo es el agotamiento progresivo
de sus recursos energéticos basados mayoritariamente en combustibles no renovables. Al
mismo tiempo, el consumo de energía aumenta a ritmos cada vez más crecientes. De otro lado,
el consumo global de combustibles genera enormes cantidades de gases contaminantes que
son liberados a la atmósfera. Este tipo de contaminación ha causado cambios en el clima del
planeta, por lo que se ha convertido en una de las problemáticas que mas preocupan a los
gobiernos. La única forma de encarar esta problemática es mediante de recursos energéticos
renovables. Para ello la biotecnología ofrece múltiples alternativas tecnológicas. Una solución
renovable es el uso de energía solar en forma de biomasa, la cual esta representada en los
materiales lignocelulósicos y en cultivos de plantas ricas en energía [13].
Los hongos causantes de la pudrición blanca están adaptados a una amplia variedad
desechos lignocelulósicos, tales como paja brava, bagazo de caña de azúcar, aserrín, residuos
de quinua, cáscara de arroz, entre otros. La producción de estas enzimas es importante en el
proceso de biodegradación de los mencionados sustratos lignocelulósicos, ya que son muy
poco aprovechados, pero no seria posible la utilización de estos como materia prima para la
producción de alcohol, sin la intervención de enzimas como las ligninolíticas y de enzimas
celulolíticas [15].
En este sentido en el Instituto de Investigaciones Fármaco Bioquímicas (IIFB), con el
apoyo de la cooperación Sueca ASDI/SAREC, en el proyecto “Biodiversidad Microbiana”

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
conjuntamente con el Instituto de Investigaciones y Desarrollo de Procesos Químicos
(IIDEPROQ), con la colaboración del IFS (Internacional Foundation for Science) debido al
alto costo de este tipo de enzimas comerciales se toma la iniciativa de producir enzimas
ligninolíticas, requiriendo identificar al hongo capaz de producir dichas enzimas.
El presente estudio de investigación se enfoca íntegramente en la producción de
enzimas ligninolíticas, a partir de hongos aislados de muestras recolectadas del Altiplano
boliviano, en el cual se verifica cualititavamente a la cepa fúngica capaz de producir este tipo
de enzimas redox, utilizando medios sólidos que tiene como indicadores Poly-R y ABTS,
posteriormente se desarrollara diseño factorial 23 con dos diferentes concentraciones de los
nutrientes: glucosa o sustratos lignocelulósicos (paja brava, bagazo, aserrín, residuo quinua,
cáscara arroz), extracto de levadura, manganeso o cobre, se tomen en cuenta las unidades de
color y el consumo de sustrato sintéticos como ABTS y 2,6 DMF por oxidación, los cuales
fueron determinados por métodos espectrofotométricos, la posterior purificación y aplicación
de dicha enzima en diferentes procesos ya descritos, se realizara en trabajos posteriores.
2. MARCO TEORICO CONCEPTUAL
2.1 LA LIGNINA
La palabra lignina proviene del término latino lignum, que significa madera; así, a las
plantas que contienen gran cantidad de lignina se las denomina “leñosas” [16]. La lignina está
en las plantas, íntimamente ligada a la celulosa, constituyente mayoritario a su vez de la
biomasa vegetal. Actúa como pegamento entre las fibras vegetales, recubre y protege a las
moléculas de celulosa en la pared de las fibras [9].
2.1.1 Estructura y biodegradación de la molécula de lignina

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
8
La lignina, después de la celulosa, es el mayor componente de la materia vegetal y
forma más abundante de material aromático en la biosfera. La composición química de la
madera y otros tejidos vasculares es muy variable. Se compone principalmente de celulosa,
lignina, hemicelulosa, y de un 5% a un 10% de otros materiales. La lignina representa entre un
16 % hasta un 33% del peso según el tipo de madera u otro tejido lignocelulósico [19]. La
mayor parte de esta se encuentra dentro de las paredes celulares, entremezclada con las
hemicelulosas y formando una matriz que rodea las ordenadas microfibrillas de celulosa. Este
compuesto provee de rigidez a las plantas superiores ya que actúa como pegamento entre las
fibras de celulosa formando la lámina media [5]. Además, protege a los carbohidratos
fácilmente degradables (celulosa, hemicelulosa) de la hidrólisis enzimática microbiana [12].
Una estructura química representativa de la lignina se muestra en la figura 1. Este
biopolímero contiene alrededor de 10-20% de grupos hidroxilo fenolitos que le confieren
rigidez a la pared celular de las plantas y además las protege del ataque de organismos
patógenos [12, 19].
Biosintéticamente, la lignina proviene de tres alcoholes precursores: el alcohol
p-hidroxicinamilico (cumarilico), el alcohol 4-hidroxi-3-metoxicinamilico (coniferilico) y el
alcohol 3,5-dimetoxi-4-hidroxicinamilico. La copolimerización por radicales libres de estos
alcoholes, iniciada por peroxidasa vegetales, da lugar al polímero de lignina [5].
Químicamente este polímero es heterogéneo, amorfo, ópticamente inactivo, y altamente
ramificado [12].
La lignina es un caso especial en el que la biodegradabilidad depende de la
disponibilidad de oxigeno. A menudo no existe una degradación sustancial por que la mayoría
de los hongos filamentoso que degradan lignina, pueden actuar solo en presencia de oxigeno
[22].

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
9
Figura 1. Estructura parcial de la lignina
Fuente: Rev Temuco Universidad de la Frontera [20].
Las características estructurales de este heteropolímero vegetal imponen ciertas
restricciones para su biodegradación. Definiendo a una molécula de lignina con un peso
molecular de entre 600-1000 kDa, es evidente que su tamaño le impide poder ser degradada
intracelularmente [7, 9]. Además por el tipo de enlaces covalentes que presenta (aril-eter, aril-
aril y carbono-carbono) y su hetereogeneidad, no puede ser degradado por mecanismos típicos
de hidrólisis [21]. Por lo tanto, cualquier enzima o grupo de enzimas capaces de atacar
inicialmente la lignina, deben ser además de extracelulares, no hidrolíticas y bastante
inespecíficas.
a. Propiedades físicas

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
20
Las ligninas son polímeros insolubles en ácidos y en álcalis fuertes, que no se digieren
ni se absorben y tampoco son atacados por la microflora del colon. Pueden ligarse a los ácidos
biliares y otros compuestos orgánicos (por ejemplo, colesterol), retrasando o disminuyendo la
absorción en el intestino delgado de dichos componentes. El grado de lignificación afecta
notablemente a la digestibilidad de la fibra [19]. La lignina, que aumenta de manera ostensible
en la pared celular de la planta con el curso de la maduración, es resistente a la degradación
bacteriana, y su contenido en fibra reduce la digestibilidad de los polisacáridos fibrosos.
b. Ligninas comercializadas Sólo existen dos tipos de lignina comercialmente disponibles: las ligninas sulfonadas y
las kraft ligninas. La capacidad de elaboración de productos de lignina en el mundo oriental es
aproximadamente de 1,4106 toneladas/año. Sólo una compañía produce kraft ligninas; las
restantes producen ligninas sulfonadas. Los productos de lignina han empezado a tener una
importancia creciente en distintas aplicaciones industriales [19].
2.1.2 Funciones de la lignina.- La lignina realiza múltiples funciones que son esenciales para la vida de las plantas.
Por ejemplo, posee un importante papel en el transporte interno de agua, nutrientes y
metabolitos. Proporciona rigidez a la pared celular y actúa como puente de unión entre las
células de la madera, creando un material que es notablemente resistente a los impactos,
compresiones y flexiones. Realmente, los tejidos lignificados resisten el ataque de los
microorganismos, impidiendo la penetración de las enzimas destructivas en la pared celular.
2.2 DEGRADACION DE LA LIGNINA POR HONGOS
Los biocatalizadores o enzimas pueden ser obtenidos de animales, plantas, sin embargo
la fuente mas importante constituyen los microorganismos como las bacterias, hongos y
levaduras. El 90% de las enzimas empleadas industrialmente son de origen microbiano. Los
microorganismos son fácilmente cultivables industrialmente bajo condiciones controladas y
por lo tanto las enzimas son obtenidas con un buen rendimiento y de manera

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
2
constante. Además los microorganismos son productores inducibles y en algunos casos
constitutivos de enzimas [29].
Existe un limitado número de microorganismos que producen esta clase de enzimas,
principalmente hongos y bacterias, las enzimas ligninolíticas que se encuentran en el mercado
principalmente provienen de cepas fúngicas, de esta forma las cepas fúngicas son un grupo de
interés en la producción de enzimas. Hasta hoy día se conoce una sola clase de seres vivos
capaces de degradar eficientemente lignina. Se trata de los hongos Basidiomicetes,
denominados "pudrición blanca", ya que producen un blanqueamiento de la madera al
degradar la lignina [20].
En este sentido debido a sus propiedades ya descritas, nos interesa estudiar las
diferentes propiedades de los hongos.
2.2.1 Hongos
a. Características generales
Los hongos constituyen un grupo muy numeroso de organismos (se han descrito
aproximadamente 500.000, pero se estima que pueden existir entre 1 y 1,5 millones de
especies) que presentan una amplia distribución en la naturaleza, contribuyendo ala
descomposición de la materia orgánica y participando en ciclos biológicos. Un pequeño
número son patógenos de animales y plantas [24].
Son un grupo diverso de organismos unicelulares o pluricelulares que se alimentan
mediante la absorción directa de nutrientes. Los alimentos se disuelven mediante enzimas que
secretan los hongos; después se absorben a través de la fina pared de la célula y se distribuyen
por difusión simple en el protoplasma. Junto con las bacterias, los hongos son los causantes de
la putrefacción y descomposición de toda la materia orgánica. Hay hongos en cualquier parte
en que existan otras formas de vida. Algunos son parásitos de organismos vivos y producen
graves enfermedades en plantas y animales. La disciplina científica que estudia los hongos se

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
22
llama micología.
Los hongos figuraban en las antiguas clasificaciones como una división del reino
Plantas (Plantae). Se pensaba que eran plantas carentes de tallos y de hojas que, en el
trascurso de su transformación en organismos capaces de absorber su alimento, habían perdido
la clorofila, y con ello, su capacidad para realizar la fotosíntesis. Sin embargo, en la actualidad
los científicos los consideran un grupo completamente separado, que evolucionó a partir de
flagelados sin pigmentos. Ambos grupos se incluyen dentro del reino Protistas, o bien se
coloca a los hongos como un reino aparte, debido a la complejidad de su organización (ver
clasificación más adelante). Hay unas cien mil especies conocidas de hongos. Se cree que los
grupos más complejos derivan de los tipos más primitivos, los cuales tienen células flageladas
en alguna etapa de su ciclo vital [25].
b. Estructura.-
Los hongos se diferencian en inferiores y superiores. Los inferiores son los que no se
pueden ver a simple vista, teniendo que utilizar un microscopio para su observación, de aquí
que se llamen también hongos microscópicos. Los superiores o macroscópicos, son las
populares setas que se recolectan al bosques, y que en realidad se trata de la parte fructífera del
hongo, figura 2-3 [24].
La pared celular de los hongos posee diferentes constituyentes químicos como ser
polisacáridos, proteínas, lípidos y otras sustancias. La constitución varía entre las diferentes
especies. También varia con la edad del hongo, ya que sustancias que pueden estar presentes
en las hifas jóvenes, desaparecen en las más viejas o depositar otros materiales y enmascarar la
presencia de constituyentes iniciales, también la composición del medio, el pH y la
temperatura, influyen en la composición de las paredes de los hongos [25].
La mayoría de los hongos están constituidos por finas fibras que contienen
protoplasma, llamadas hifas. Éstas a menudo están divididas por tabiques llamados septos. En
cada hifa hay uno o dos núcleos y el protoplasma se mueve a través de un diminuto poro que

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
23
ostenta el centro de cada septo.
Figura 2. Corte esquemático de un hongo de la clase basidiomiceto y orden de los agaricales.
Fuente: El reino de los hongos [24].
No obstante, hay un filo de hongos, que se asemejan a algas, cuyas hifas generalmente
no tienen septos y los numerosos núcleos están esparcidos por todo el protoplasma. Las hifas
crecen por alargamiento de las puntas y también por ramificación. La proliferación de hifas,
resultante de este crecimiento, se llama micelio. Cuando el micelio se desarrolla puede llegar a
formar grandes cuerpos fructíferos ya anteriormente mencionados, tales como las setas y los
pedos o cuescos de lobo. Otros tipos de enormes estructuras de hifas permiten a algunos
hongos sobrevivir en condiciones difíciles o ampliar sus fuentes nutricionales. Las fibras, a
modo de cuerdas, del micelio de la armilaria color de miel (Armillaria mellea), facilitan la
propagación de esta especie de un árbol a otro [27].

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
24
c. Hábitat.-
Los hongos están muy distribuidos en todos los ecosistemas, desde 0 a 25ºC, aunque
pocos a 0ºC y con un máximo de 50ºC. para los hongos del suelo el óptimo se ubica entre 20-
30ºC, en pH muy variados: el óptimo estaría de Ph 6 y dominan en ambientes ácidos [25]. La
luz esencial para la esporulación de muchas especies [25].
d. Reproducción.-
La mayoría de los hongos se reproducen por esporas, diminutas partículas de
protoplasma rodeado de pared celular. El champiñón silvestre puede formar doce mil millones
de esporas en su cuerpo fructífero; así mismo, el pedo o cuesco de lobo gigante puede producir
varios billones.
Las esporas se forman de dos maneras. En el primer proceso, las esporas se originan
después de la unión de dos o más núcleos, lo que ocurre dentro de una o de varias células
especializadas. Estas esporas, que tienen características diferentes, heredadas de las distintas
combinaciones de genes de sus progenitores, suelen germinar en el interior de las hifas. Los
cuatro tipos de esporas que se producen de esta manera (oosporas, zigosporas, ascosporas y
basidiosporas) definen los cuatro grupos principales de hongos. Las oosporas se forman por la
unión de una célula macho y otra hembra; las zigosporas se forman al combinarse dos células
sexuales similares entre sí. Las ascosporas, que suelen disponerse en grupos de ocho unidades,
están contenidas en unas bolsas llamadas ascas. Las basidiosporas, por su parte, se reúnen en
conjuntos de cuatro unidades, dentro de unas estructuras con forma de maza llamadas basidios
[27].
El otro proceso más común de producción de esporas implica la transformación de las
hifas en numerosos segmentos cortos o en estructuras más complicadas de varios tipos. Este
proceso sucede sin la unión previa de dos núcleos. Los principales tipos de esporas

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
2
reproductivas formadas así son: oídios, conidios y esporangiosporas. Estas últimas se originan
en el interior de unos receptáculos, parecidos a vesículas, llamados esporangios. La mayoría
de los hongos producen esporas sexuales y asexuales.
e. Clasificación.-
Existen dos tipos de clasificaciones:
Según la forma de los órganos productores de esporas
Ascomicetos: Son aquellos hongos que producen sus esporas en el interior de unas
pequeñas vainas a modo de saco microscópico llamadas ascas. Las esporas maduran en el
interior de este pequeño saco, que se abrirá para expulsarlas al exterior. Cada asca contiene
generalmente 8 esporas.
Basidiomicetos: Producen sus esporas sobre unos pequeños órganos a modo de mazas,
formando una especie de racimo microscópico de, generalmente, 4 esporas [26].
Según sus características microscópicas y morfológicas
Existen en realidad más categorías, como el subgénero, sección, tribu, variedad, forma,
etc; pero las mencionadas antes son las principales, y la mayoría de las veces simplemente
utilizaremos las dos últimas para referirnos a un hongo: el Género y la Especie [26].
Por ejemplo si observamos al microscopio una muestra podremos observar que, las
esporas se producen en basidios, con lo cual pertenecerán a la subdivisión Basidiomicotina,
clase Basidiomicetes, etc.
f. Fisiología de los Hongos.-
En la mayoría de los hongos las paredes de las hifas están compuestas principalmente

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
26
por quitina y algunas hemicelulosas. La celulosa, que está presente sólo en unos pocos grupos
de hongos, es característica de los oomicetes. La proporción de agua de los hongos
mucilaginosos generalmente es mayor del 90%. Las esporas pueden tener menos del 50% de
agua; otras estructuras de resistencia, tales como los esclerocios, contienen aún menos. Los
hongos requieren oxígeno para su crecimiento, así como grandes cantidades de agua y de
hidratos de carbono u otras fuentes de carbono.
La mayoría de los hongos utilizan azúcares como la glucosa y la levulosa (D-
fructosa), pero algunos usan otros compuestos orgánicos como alimento, según su capacidad
para sintetizar las enzimas adecuadas. Ciertas micorrizas toman directamente el nitrógeno de
la atmósfera; sin embargo, todos los demás hongos lo obtienen de nitratos, sales de amonio u
otros compuestos orgánicos o inorgánicos de nitrógeno. Los hongos, además, precisan otros
elementos como potasio, fósforo, magnesio y azufre. También son necesarios, aunque en muy
pequeñas cantidades, hierro, manganeso, cobre, molibdeno, zinc y galio; así como factores de
crecimiento [24]. Determinados hongos son deficitarios, al menos en parte, en uno o más
factores de crecimiento.
Las enzimas de los hongos pueden actuar sobre una gran variedad de sustancias. Un
grupo de enzimas, llamado el complejo zimasa, permite a las levaduras llevar a cabo la
fermentación alcohólica [27].Otras enzimas, como la protopectinasa, la pectasa y la pectinasa,
hidrolizan los compuestos pectídicos que hay en las capas medias de las paredes celulares de
las plantas. La amilasa, celobiasa, citasa, dextrinasa, invertasa, lactasa, maltasa, proteasa y la
tanasa son también enzimas producidas por los hongos.
El glucógeno, sustancia relacionada con el almidón y con la dextrina, es la reserva de
hidratos de carbono más común en los hongos. Además, algunos hongos forman polisacáridos
y alcoholes polihidroxílicos, como el manitol y la glicerina. Otros producen proteínas y grasas
en abundancia. Muchos hongos sintetizan ácido oxálico y otros ácidos orgánicos, como
cítrico, fórmico, pirúvico, succínico, málico y acético; la producción de ácido láctico sólo la
realiza una familia de hongos [26]. Otros productos del metabolismo fúngico son compuestos
de azufre, sustancias que contienen cloro y numerosos pigmentos. Unos cuantos hongos tienen

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
2
la facultad de formar compuestos volátiles de arsénico cuando crecen sobre sustratos que lo
contienen.
g. Hongos ligninolíticos.-
Estos hongos ligninolíticos denominados hongos de la pudrición blanca de la madera,
comprenden un grupo de organismos cuya característica es su capacidad para mineralizar
eficientemente la lignina. Presumiblemente esta degradación selectiva les permite tener acceso
a la celulosa y/o hemicelulosa, las cuales finalmente representan su fuente de energía y
carbono [12].
La mayoría de los hongos ligninolíticos pertenecen al grupo Basidiomycetes y son los
microorganismos más eficientes en degradar totalmente la lignina. Estos organismos secretan
varias enzimas extracelulares que son esenciales para la transformación inicial de la lignina y
que en conjunto logran su mineralización [28].
Los hongos ligninolíticos has desarrollado un sistema enzimático único y no especifico
que funciona en el ambiente extracelular. El mecanismo del sistema degradador de lignina esta
basado en la producción de radicales libres. Este mecanismo permite que estas enzimas sean
cataliticamente activas sobre una gran diversidad e sustratos orgánicos. La enorme diversidad
estructural de los contaminantes que son degradados por estos hongos, les confiere un uso
potencial en biorremediación.
Las enzimas ligninolíticas son expresadas por los hongos como un proceso idiofasico,
el cual esta a la limitación de carbono y/o nitrógeno o sulfato en el medio y su síntesis es
particularmente activa con altas tensiones de oxigeno [23].
2.3 ENZIMAS.-
Las enzimas o biocatalizadores son proteínas cuya función es catalizar las reacciones
que realizan los seres vivos. Las enzimas son altamente selectivas para la reacción que

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
28
catalizan. Estas se unen a un sustrato y lo biotransforman.
2.3.1 Estructura y características generales.-
Las enzimas, como proteínas globulares de diversos tamaños, tiene su estructura
definida por la estructura primaria, segundaria, terciaria y cuaternaria.
Existe también una serie de características que diferencian alas enzimas de otras
sustancias. Algunas de ellas son: especificidad, actividad enzimática, cinética de reacción,
temperatura, pH, activación enzimática e inactivación [31].
a. Especificidad.-
Las enzimas son específicas, es decir, hidrolizan y sintetizan un compuesto en
particular. En algunos casos, su acción esta limitada a ligaciones especificas dentro de los
compuestos con los cuales ejercen reacción.
b. Actividad Enzimática.-
La actividad de una enzima o preparación enzimáticas se expresa por peso, en bases de
volumen y/o también en unidades, que es lo más común. Las condiciones propicias para una
óptima actividad enzimática, tales como pH óptimo, concentración de sustrato, cofactores e
inhibidores, temperatura, tiempo de duración del análisis y actividad de agua, son utilizadas
para determinar la unidad de actividad de la enzima.
c. Cinética de reacción (Michaelis-Menten).-
La acción enzimática se caracteriza por la formación de un complejo que representa el
estado de transición.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
29
El sustrato se une al enzima a través de numerosas interacciones débiles como son:
puentes de hidrógeno, electrostáticos, hidrófobos, etc, en un lugar específico, el centro activo.
Este centro es una pequeña porción del enzima, constituido por una serie de aminoácidos que
interaccionan con el sustrato.
Algunas enzimas actúan con la ayuda de estructuras no proteícas. En función de su
naturaleza se denominan:
Cofactor es cuando se trata de iones o moléculas inorgánicas.
Coenzima es cuando es una molécula orgánica. Aquí se puede señalar, que muchas
vitaminas funcionan como coenzimas; y realmente las deficiencias producidas por la falta de
vitaminas responden más bien a que no se puede sintetizar un determinado enzima en el que la
vitamina es la coenzima [30,31].
d. Temperatura.-
La velocidad de reacciones enzimáticas aumenta con el incremento de la temperatura,
de modo semejante a las reacciones químicas.
e. pH.-
La acción catalítica de una reacción enzimática es alcanzada dentro de límites muy
estrechos de pH. Cada reacción tiene un pH óptimo, que para la mayoría de las enzimas se
encuentra entre 4,5 y 8,0, y allí la enzima presenta su actividad máxima.
f. Clasificación Internacional de las enzimas.-
Según la Internacional Comision on Enzyme, han formado con el consentimiento de la

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
30
IUPAC (Internacional Union of Pure and Applied Chemistry) y IUB (internacional Union of
Biochemistry) un sistema para la nomenclatura de las enzimas [30,41] (ver tabla 2-1).
Tabla 1. Clasificación Internacional de las enzimas.
2.3.2 Enzimas ligninolíticas.-
Se han identificado dos familias de enzimas extracelulares, las peroxidasas y las
lacasas. Estas enzimas son capaces de oxidar una gran variedad de compuestos aromáticos y
se diferencian en sus requerimientos catalíticos, potencial de oxido-reducción, y en su modo
de acción que puede ser directo o indirecto [46].
a. Peroxidasas ligninolíticas.-
Lignina peroxidasa (LiP) diaril propano O2 H2O2 oxidoreductasa EC 1.11.1.14
Esta enzima fue descrita por primera vez en P. chrysosporium [5, 47, 48]. Es una
glucoproteína monomerica que contiene un grupo hemo.
El ciclo catalítico de esta peroxidasa comienza con la oxidación de la enzima por
peróxidos, dando lugar al compuesto I, que contiene un complejo oxo-Fe+4 radical
Nº Clase Tipo de reacción catalizada
1 Oxidoreductasas Transferencia de electrones (iones hidruro o átomos de H)
2 Transferasas Reacciones de transferencia de grupos
3 Hidrolasas Reacciones de hidrólisis ( Transferencia de grupos funcionales al agua)
4 Liasas Adición de grupos a dobles enlaces o formación de dobles enlaces por eliminación de grupos
5 Isomerasas Transferencia de grupos dentro de moléculas dando formas isomericas
6 Ligasas Formación de enlaces C-C, C-S, C-O, C-H mediante reacciones de condensación acopladas a la rotura de ATP

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
3
cationico de la porfirina [49] (1). El compuesto I es reducido en dos pasos consecutivos por
compuestos aromáticos, para formar el Compuesto II y la enzima nativa (2) (3). El exceso de
peroxido da lugar a una reacción irreversible en la que se forma en Compuesto III que es
inactivo (4).
LiP Fe 3+ + H2O2 -------- LiP C-I Fe 4+ = O (P)+ + H2O (1)
LiP C-I Fe 4+ = O (P) + + RH ------- LiP C-II Fe 4+ = O (P) + R * + H + (2)
LiP C-II Fe 4+ = O (P) + RH + H ------- LiP Fe 3+ + R * + H2O (3)
LiP C-II Fe 4+ = O + H2O ------- LiP C-III Fe 3 + O2 + (4)
La LiP se diferencia de otras peroxidasas por su alto potencial de oxido-reducción, lo
que permite oxidar directamente compuestos aromáticos no fenólicos, y por tanto, la mayor
parte de las unidades de la lignina [48].
Peroxidasas que oxidan el manganeso (MnP) Mn (II): H2O2 oxidoreductasa EC
1.11.1.13
Mas frecuente que la producción de peroxidasas que oxidan el Mn 2+ a Mn 3+ en
presencia de peróxidos. Estas peroxidasas manganeso dependientes se han descrito en al
mayoría de los hongos ligninolíticos de la podredumbre blanca de la madera [50].
El Mn 3+ formado en la reacción es un fuerte oxidante, que estabilizado con ácidos
dicarboxilicos secretados por el hongo, actúa como intermediario de la oxidación de
compuestos fenólicos y no fenólicos [51].
El ciclo catalítico de la MnP es semejante al de otras peroxidasas incluidas la LiP y
transcurre a través de la formación de intermediarios de la enzima MnP C-I y MnP C-II.
Mn Fe 3+ + H2O2 -------- MnP C-I Fe 4+ = O (P)+ + H2O (1)

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
32
MnP C-I Fe 4+ = O (P) + + Mn +2 ------- MnP C-II Fe 4+ = O (P) + Mn 3+ (2a)
MnP C-I Fe 4+ = O (P) + RH ------- MnP C-II Fe 4+ + O (P) + R * (2b)
MnP C-II Fe 4+ = O (P) + Mn +2 ------- MnP Fe 3 + + Mn 3+ + H2O (3)
MnP C-II Fe 4+ = O + H2O2 ------- MnP C-III Fe 3 + O2 + (4)
El primer producto de la reacción de la MnP nativa con el peroxido de hidrogeno es el
compuesto I (1). La reducción monoelectrónica del compuesto I por Mn 2+ conduce al
compuesto II que de nuevo se reduce por Mn 2+ para generar el enzima nativo. Mientras que el
compuesto I puede ser reducido además de por el Mn 2+ (2a), por otros donadores como los
fenoles (2b), el compuesto II de la MnP solo puede ser reducido por Mn 2+ por lo que depende
de este catión para cerrar su ciclo (3).
Al igual que la LiP la MnP es sensible a latas concentraciones de H2O2 que provocan la
inactivación reversible del enzima al compuesto III que es un estado de oxidación inactivo
cataliticamente (4).
La mayoría de estas peroxidasas se caracterizan por depender del Mn 2+ para cerrar su
ciclo catalítico [49]. Son enzimas que solo actúan de manera indirecta sobre la lignina.
b. Lacasas.-
Lacasas EC.1.10.3.2. Benzodiol oxigeno: oxigeno reductasa
Las lacasas son fenoloxidasas ampliamente distribuidas en plantas, hongos de
diferentes clases [52] y algunas bacterias [53].
Las lacasas se descubrieron en el látex de la planta Rhus vernicifera [54], como la
sustancia termolábil responsable del endurecimiento de la resina. En plantas la lacasa tiene una
función protectora frente a lesiones sufridas por el vegetal, pues la oxidación de fenoles del
látex por la lacasa provoca radicales libres que tienden a polimerizar lo que origina una

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
33
cubierta protectora en la lesión.
La función de la lacasa en los hongos no esta clara, se ha relacionado con la
morfogénesis, con la pigmentación de los conidios, con la formación de rizoformos, con el
desarrollo de cuerpos fructificantes [54], con la patogénesis [55] y en la protección frente a
compuestos fenólicos tóxicos liberados durante la degradación de la lignina [56].
La lacasa mejor estudiada es la de Trametes versicolor [56]. Es una O2 p-difenol
oxidoreductasa que se diferencia de la mayoría de fenoloxidasas en que se produce agua en
lugar de peroxido en la reducción del oxigeno. Son glucoproteínas con 4 átomos de cobre y de
peso molecular variable [57]. Los átomos de cobre están clasificados en tres tipos (1, 2, 3) con
diferentes propiedades [58].
Cobre tipo 1 (T1): es responsable del color azul de la proteína. Tiene una alta
absorbancia en la región del visible (605nm) provocada por la union covalente cobre-cisteina.
Debido al alto potencial redox de este cobre es el sitio donde ocurre la oxidación del sustrato.
Cobre tipo 2 (T2): se caracteriza por no presentar absorbancia detectable en al región
visible y por tener una alta afinidad por aniones (F, CN...) que actúan como inhibidores de la
actividad del enzima.
Cobre tipo3 (T3): es un complejo binario formado por un par de iones Cu2+-Cu 2+
unidos por puente hidróxido con un máximo de absorbancia a 330 nm y un espectro de
fluorescencia característico. La reacción que cataliza la lacasa es la siguiente:
4 AH + O2+ ---------- 4A + H2O [AH: Dador de electrones]
Los cobres T2 y T3 forman un cluster trinuclear que es el lugar donde se reduce el
oxigeno. El cobre del centro T1 es el primer aceptor de electrones del sustrato. Después los
electrones se trasfieren secuencialmente al centro T2-T3 que tras recibir cuatro electrones
reducen una molécula de oxigeno a agua. Así la oxidación monoelectrónica del sustrato va
acoplada a la reducción por cuatro electrones del oxigeno molecular.
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
34
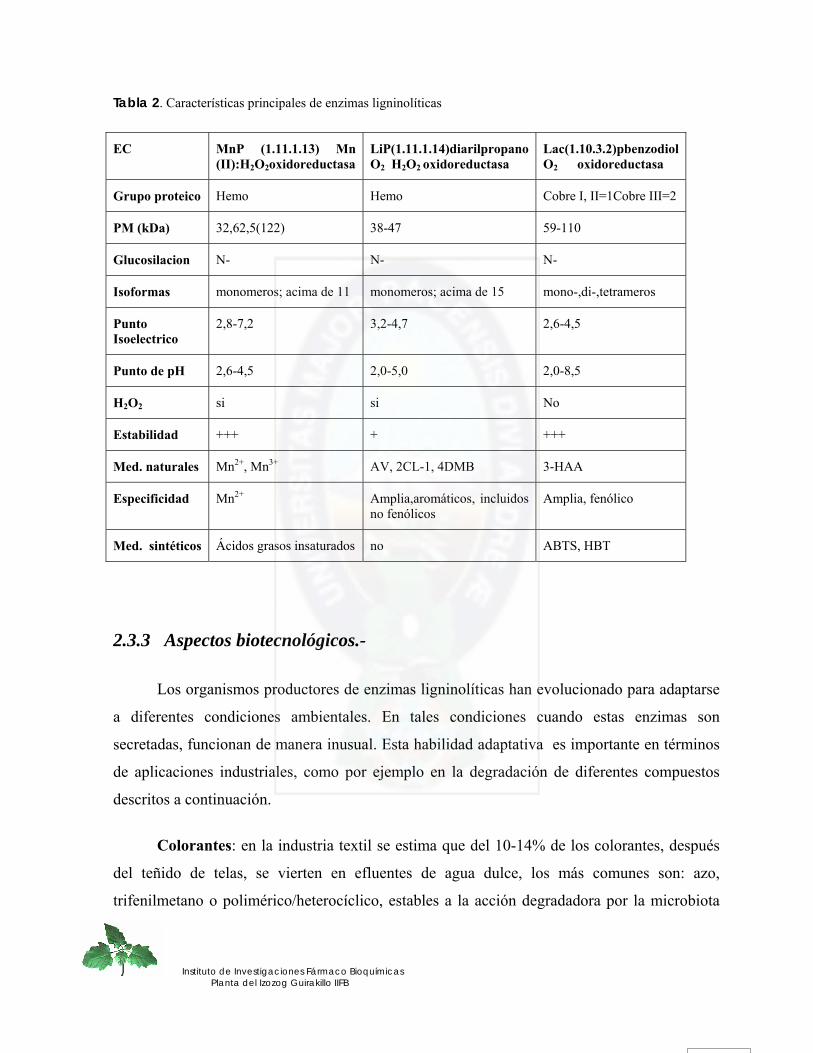
Tabla 2. Características principales de enzimas ligninolíticas
2.3.3 Aspectos biotecnológicos.-
Los organismos productores de enzimas ligninolíticas han evolucionado para adaptarse
a diferentes condiciones ambientales. En tales condiciones cuando estas enzimas son
secretadas, funcionan de manera inusual. Esta habilidad adaptativa es importante en términos
de aplicaciones industriales, como por ejemplo en la degradación de diferentes compuestos
descritos a continuación.
Colorantes: en la industria textil se estima que del 10-14% de los colorantes, después
del teñido de telas, se vierten en efluentes de agua dulce, los más comunes son: azo,
trifenilmetano o polimérico/heterocíclico, estables a la acción degradadora por la microbiota
EC MnP (1.11.1.13) Mn (II):H2O2oxidoreductasa
LiP(1.11.1.14)diarilpropano O2 H2O2 oxidoreductasa
Lac(1.10.3.2)pbenzodiol O2 oxidoreductasa
Grupo proteico Hemo Hemo Cobre I, II=1Cobre III=2
PM (kDa) 32,62,5(122) 38-47 59-110
Glucosilacion N- N- N-
Isoformas monomeros; acima de 11 monomeros; acima de 15 mono-,di-,tetrameros
Punto Isoelectrico
2,8-7,2 3,2-4,7 2,6-4,5
Punto de pH 2,6-4,5 2,0-5,0 2,0-8,5
H2O2 si si No
Estabilidad +++ + +++
Med. naturales Mn2+, Mn3+ AV, 2CL-1, 4DMB 3-HAA
Especificidad Mn2+ Amplia,aromáticos, incluidos no fenólicos
Amplia, fenólico
Med. sintéticos Ácidos grasos insaturados no ABTS, HBT

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
3
natural, dada la similitud química de estos con los compuestos de los procesos vitales
celulares. Para disminuir ese impacto negativo, existe la alternativa de usar hongos de la
podredumbre blanca y/o sus enzimas extracelulares inespecíficas que oxidan la lignina [59].
Aromáticos: en la biorremediación de ambientes contaminados con xenobioticos
aromáticos, se recomienda aplicar hongos de la PB. [60]
Hidrocarburos aromáticos policíclicos (HAP): los HAP son compuestos homólogos
de benceno formados por 2 o más anillos fusionados, que se liberan en emisiones industriales,
durante la combustión incompleta de la gasolina en automotores, en plantas de generación de
energía, naturalmente en depósitos de carbón, en el proceso de transformación de derivados
aromáticos de terpenos, esteroles, quinonas etc., sin embargo estos compuestos son
recalcitrantes y potenciales carcinogénicos. La literatura reporta que los hongos de la PB, así
como sus extractos extracelulares, tienen la capacidad potencial para la degradación de HAP,
por ello son una alternativa en la biorremediación de ambientes contaminados con estos
poliaromáticos [61,62].
Bifenilos policlorados (BFP): los BFP se utilizan en la industria como fluidos
dieléctricos e hidráulicos, como diluyentes orgánicos, plastificantes, etc., por su estabilidad
química, son recalcitrantes a la biodegradación, por ello, causan serios problemas de
contaminación, por esa razón existen estrategias de biorremediación para su eliminación.
Diversos reportes en la literatura muestran la capacidad degradativa de los hongos de la PB
sobre los BFP [63, 64].
Pesticidas: son los compuestos que contaminan aguas superficiales y subterráneas,
causan un grave problema ambiental, pues algunos de estos químicos son carcinogénicos, más
por su bioaumentación a través de la cadena alimenticia, son de alto riesgo para la vida, por
eso es necesario su tratamiento mediante biorremediación para minimizar o reducir su
negativo impacto ambiental. Los hongos de la PB y sus enzimas ligninolíticas son un recurso
viable en la eliminación de los pesticidas o de algunos de sus componentes [65, 66].

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
36
Nitroaromáticos: estos son compuestos como: el nitrobenceno, el nitrotolueno, los
nitrofenoles y los nitrobenzoatos, se utilizan en síntesis de pesticidas, explosivos, colorantes, y
farmacéuticos, pero son un problema ambiental por su recalcitrancia. Ciertos nitroaromáticos
son tóxicos para humanos y animales. Investigación reporta que los hongos de la PB oxidan
nitroaromáticos, como el trinitrotolueno (TNT) son una alternativa en la recuperación de
ambientes contaminados con estos compuestos.
Otros aromáticos: existen otros compuestos sintéticos que por su estructura y su
composición química compleja no se investigan, pero evidencia sugiere que pueden ser
degradados y/o mineralizados por el sistema enzimático de los hongos de la PB como: C.
versicolor, Funalia trogii, P. chrysosporium y P. pulmonaris en la decoloración de vinasas en
cultivo estático. Los reportes al respecto muestran que no solo decoloran las vinasas sino
también reducen su demanda química de oxígeno [67]. Otro estudio demostró la capacidad
oxidativa de P. crhysosporium, P. ostreatus, Lentinus edodes, T. versicolor en un licor
residual del molido de pulpa y papel en un medio plástico poroso, donde los hongos de la PB
oxidaron en un 71% la lignina para disminuir en un 48% la demanda química de oxígeno, lo
que baja el costo de producción del proceso [68].
Además de estar implicadas en la degradación de otros compuestos aromáticos que
producen problemas de contaminación ambiental, anteriormente ya descritos tiene amplias
aplicaciones industriales; actualmente, las enzimas son empleadas principalmente para la
obtención de lácteos, edulcolorantes, fármacos, alimentos, licores, detergentes, el blanqueo de
textiles y de pulpa de papel, síntesis orgánica, biorremediación ,detergentes de lavado, etc.
2.4 RECURSOS LIGNOCELULOSICOS.-
Bolivia es uno de los países mas ricos del mundo en diversidad biológica, ya que en el
país se encuentran entre el 35 a 45% de toda la diversidad biológica, aun que el territorio
boliviano representa apenas el 0,2% de la superficie mundial, y sus bosques alcanzan el 3,5%
de los bosques del mundo [27].

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
3
Dada la disminución de las reservas de materias orgánicas fósiles, en los últimos años
en todo el mundo se dedica mucha atención al tratamiento químico y biotecnológico de la
biomasa vegetal de madera y plantas agrícolas. La biomasa vegetal (fitomasa) se forma como
resultado de la actividad fotosintética de las plantas. A diferencia de las fuentes fósiles, las
reservas de fitomasa se renuevan como resultado de la actividad de las plantas superiores.
Cada año en el planeta se forman cerca de 200 millones de toneladas de biomasa vegetal
celulósica. La biosíntesis de la celulosa es la mayor de las síntesis en el pasado, presente y
futuro.
En este siglo los principales tipos de materiales orgánicos serán la madera, las plantas
agrícolas y el carbón de piedra que a su vez es de origen vegetal. Por una valoración
aproximada, las reservas conocidas de petróleo son prácticamente iguales a las de la madera
en nuestro planeta, sin embargo, los recursos petroleros se agotan, al tiempo que los de
maderas y otros desechos agrícolas, como resultado del crecimiento natural, se elevan [72].
La generación de Biomasa a partir de materiales lignocelulósicos tiene gran
importancia para todos los países. Los residuos agrícolas y forestales constituyen una fuente
importante de sustratos orgánicos para organismos que son capaces de degradar en la
naturaleza la lignina y la celulosa.
2.4.1 Paja Brava.-
En Bolivia se encuentran especies de plantas y animales de casi todas las zonas climáticas
debido a la variada elevación de su territorio. La paja brava es una planta que crece en la zona
más elevada del extenso Altiplano, se utiliza para confeccionar techos, denominados techos de
quincha, es empleada como cama para animales, dígase en caballerizas, también se utiliza para
mantener la humedad después de haber sembrado pasto, puede utilizarse como material de
construcción
Tabla 3: Composición química de la paja brava. Parámetro Unidad Valor Humedad % 8,17
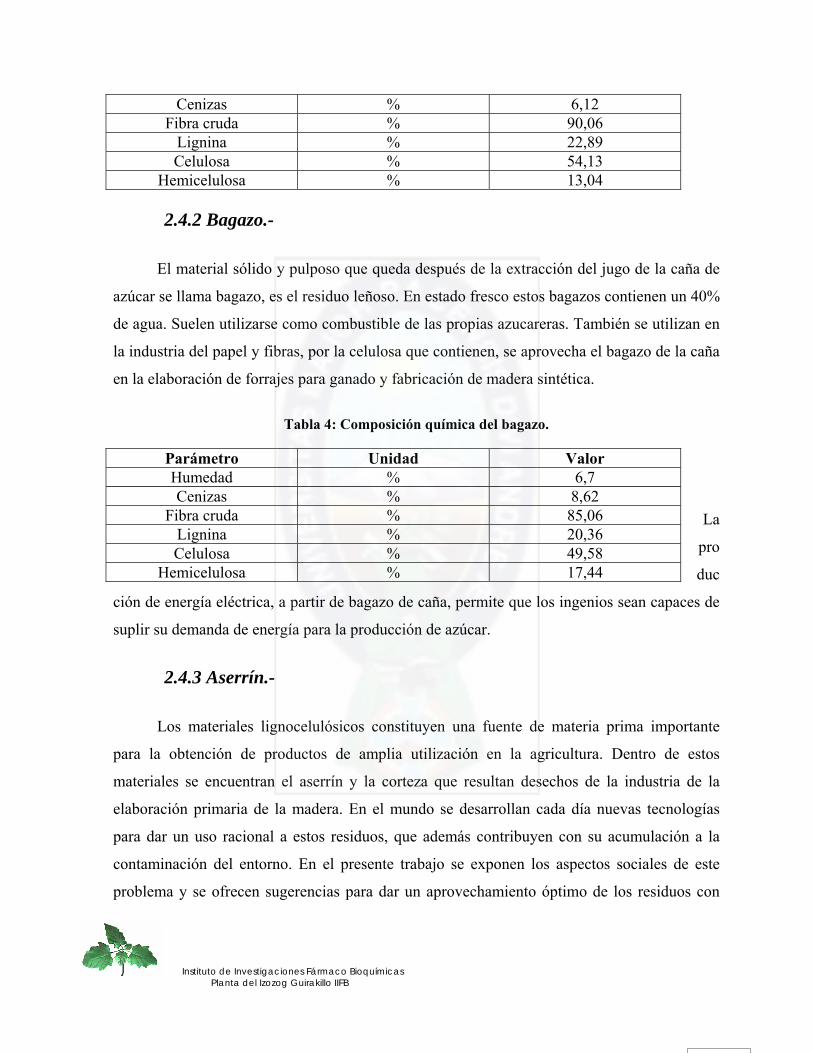
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
38
Cenizas % 6,12 Fibra cruda % 90,06
Lignina % 22,89 Celulosa % 54,13
Hemicelulosa % 13,04
2.4.2 Bagazo.-
El material sólido y pulposo que queda después de la extracción del jugo de la caña de
azúcar se llama bagazo, es el residuo leñoso. En estado fresco estos bagazos contienen un 40%
de agua. Suelen utilizarse como combustible de las propias azucareras. También se utilizan en
la industria del papel y fibras, por la celulosa que contienen, se aprovecha el bagazo de la caña
en la elaboración de forrajes para ganado y fabricación de madera sintética.
Tabla 4: Composición química del bagazo.
La
pro
duc
ción de energía eléctrica, a partir de bagazo de caña, permite que los ingenios sean capaces de
suplir su demanda de energía para la producción de azúcar.
2.4.3 Aserrín.-
Los materiales lignocelulósicos constituyen una fuente de materia prima importante
para la obtención de productos de amplia utilización en la agricultura. Dentro de estos
materiales se encuentran el aserrín y la corteza que resultan desechos de la industria de la
elaboración primaria de la madera. En el mundo se desarrollan cada día nuevas tecnologías
para dar un uso racional a estos residuos, que además contribuyen con su acumulación a la
contaminación del entorno. En el presente trabajo se exponen los aspectos sociales de este
problema y se ofrecen sugerencias para dar un aprovechamiento óptimo de los residuos con
Parámetro Unidad Valor Humedad % 6,7 Cenizas % 8,62
Fibra cruda % 85,06 Lignina % 20,36 Celulosa % 49,58
Hemicelulosa % 17,44

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
39
enfoque ambientalista [71].
Tabla 5 Composición química del aserrín.
El
aser
rín
solo, como fertilizante, es poco efectivo, ya que contiene bajo contenido de elementos
nutritivos: aproximadamente 0,1 % de N, 0,02 % de P2O5 y 0,12 % de K2O. Si se suministra
fresco, sin compostar, puede provocar carencia de Nitrógeno en la planta en el primer y a
veces en el segundo años de haberlo suministrado al suelo. Por otra parte, el complejo
lignocelulósico del aserrín puede ser utilizado para aumentar el nivel de humus del suelo.
Efecto positivo y continuado del aserrín se observó con su introducción en forma de capas
para la reforestación en suelos pobres, arenosos. Ejemplo de esto se observó que durante un
experimento en el cual al cabo de 14 años de plantado un bosque, se conservó sólo el 7 % sin
enmienda, y para el suelo que tuvo aserrín, 43 - 50 %, con un diámetro y altura de los árboles
2 - 3 veces superior [73].
2.4.4 Residuos de quinua real.-
La Quinua Real, es uno de los granos más importantes de los Andes, cuyo origen se
remonta a más de 5000 años, fue el principal alimento que corresponde a las necesidades y
requerimientos del mundo moderno. El ciclo vegetativo de la planta tiene una duración de 8
meses. La siembra generalmente se la realiza en el mes de septiembre, y la planta llega a su
fase de maduración en el mes de abril, para efectuar la cosecha y trilla en los meses de Mayo y
Junio.
Tabla 6: Composición química de residuos de quinua.
Parámetro Unidad Valor Humedad % 5,9259 Cenizas % 0,9884
Fibra cruda % 81,8280 Lignina % 17,2935 Celulosa % 56,6415
Hemicelulosa % 7,8929
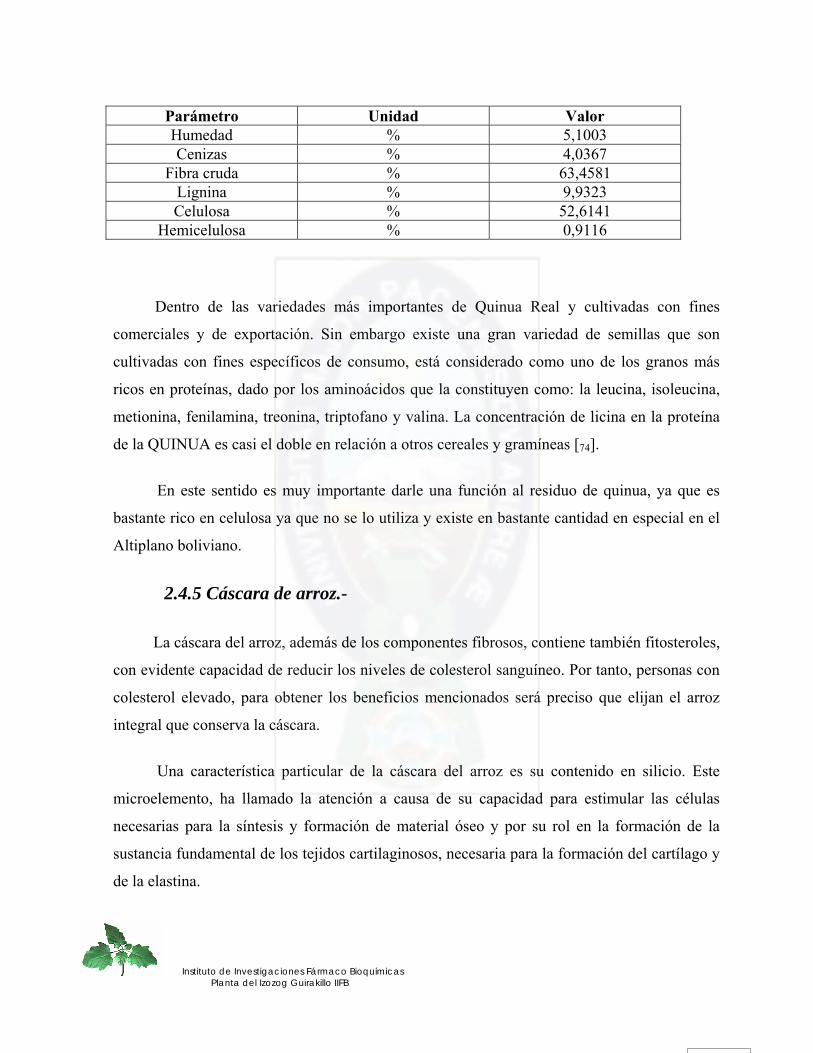
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
40
Dentro de las variedades más importantes de Quinua Real y cultivadas con fines
comerciales y de exportación. Sin embargo existe una gran variedad de semillas que son
cultivadas con fines específicos de consumo, está considerado como uno de los granos más
ricos en proteínas, dado por los aminoácidos que la constituyen como: la leucina, isoleucina,
metionina, fenilamina, treonina, triptofano y valina. La concentración de licina en la proteína
de la QUINUA es casi el doble en relación a otros cereales y gramíneas [74].
En este sentido es muy importante darle una función al residuo de quinua, ya que es
bastante rico en celulosa ya que no se lo utiliza y existe en bastante cantidad en especial en el
Altiplano boliviano.
2.4.5 Cáscara de arroz.-
La cáscara del arroz, además de los componentes fibrosos, contiene también fitosteroles,
con evidente capacidad de reducir los niveles de colesterol sanguíneo. Por tanto, personas con
colesterol elevado, para obtener los beneficios mencionados será preciso que elijan el arroz
integral que conserva la cáscara.
Una característica particular de la cáscara del arroz es su contenido en silicio. Este
microelemento, ha llamado la atención a causa de su capacidad para estimular las células
necesarias para la síntesis y formación de material óseo y por su rol en la formación de la
sustancia fundamental de los tejidos cartilaginosos, necesaria para la formación del cartílago y
de la elastina.
Parámetro Unidad Valor Humedad % 5,1003 Cenizas % 4,0367
Fibra cruda % 63,4581 Lignina % 9,9323 Celulosa % 52,6141
Hemicelulosa % 0,9116
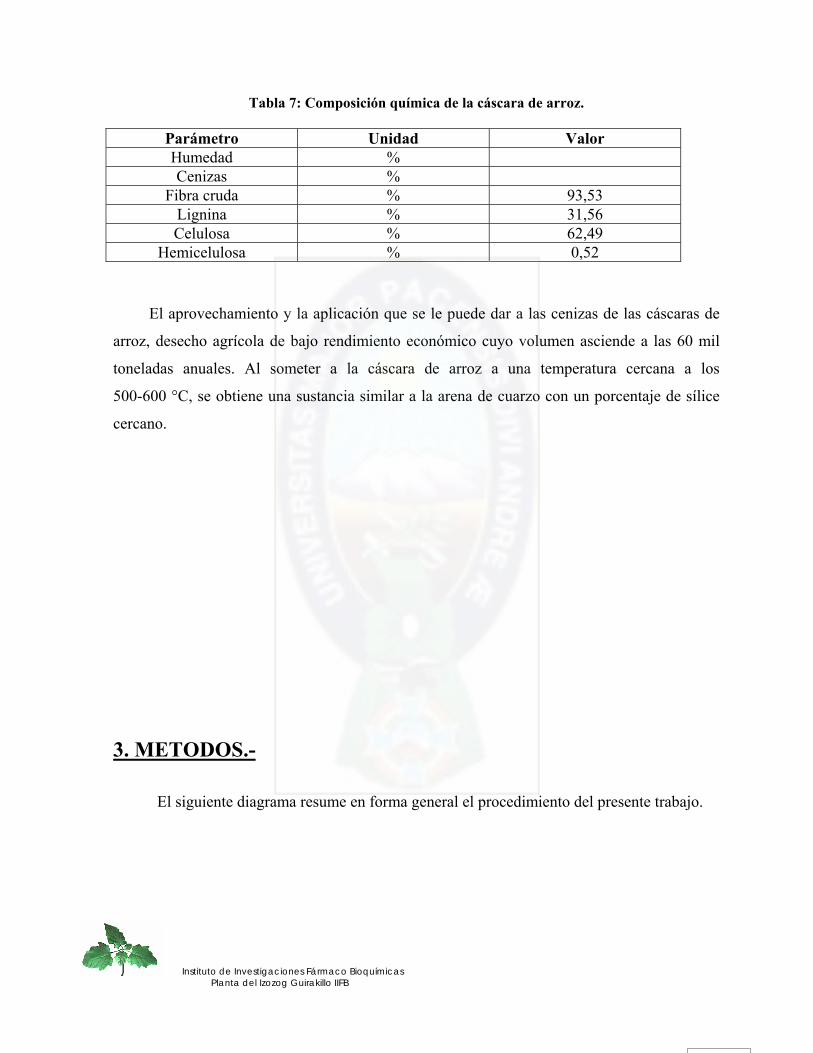
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
4
Tabla 7: Composición química de la cáscara de arroz.
El aprovechamiento y la aplicación que se le puede dar a las cenizas de las cáscaras de
arroz, desecho agrícola de bajo rendimiento económico cuyo volumen asciende a las 60 mil
toneladas anuales. Al someter a la cáscara de arroz a una temperatura cercana a los
500-600 °C, se obtiene una sustancia similar a la arena de cuarzo con un porcentaje de sílice
cercano.
3. METODOS.-
El siguiente diagrama resume en forma general el procedimiento del presente trabajo.
Parámetro Unidad Valor Humedad % Cenizas %
Fibra cruda % 93,53 Lignina % 31,56 Celulosa % 62,49
Hemicelulosa % 0,52
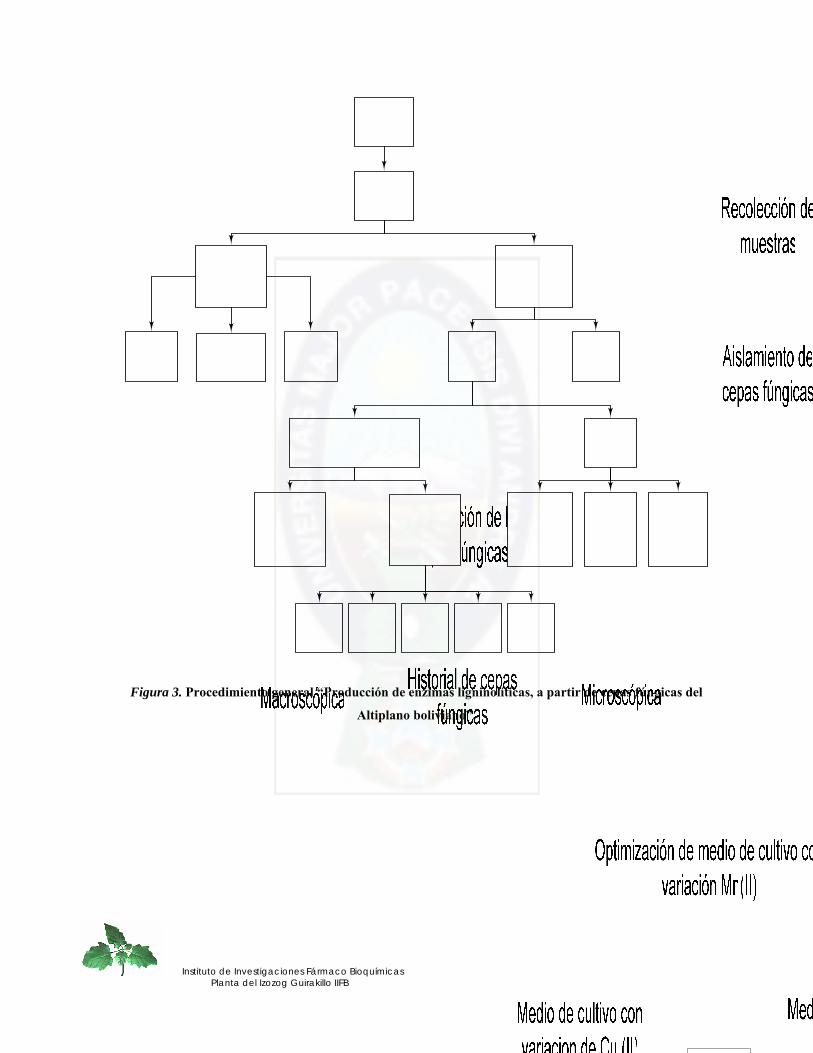
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
42
Figura 3. Procedimiento general “Producción de enzimas ligninolíticas, a partir de cepas fúngicas del
Altiplano boliviano”.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
43
3.1 RECOLECCION DE MUESTRAS.-
Se realizo un viaje a lugares cercanos al lago Titicaca mas específicamente a la
localidad de Llamacachi, cantón Compi (L.S. 17º 51, 966´ NO 67º 20,603´ 3737 m.s.n.m.) y
el rió Desaguadero (L.S. 17º 38,061´ NO 67º 30,500´ 3750 m.s.n.m.) ubicados en los
departamentos de La Paz y Oruro respectivamente.
Figura 4. Orillas del Lago Titicaca en la localidad de Llamacachi.
Figura 5. Orillas del Río Desaguadero en el departamento de Oruro.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
44
En los lugares mencionados se procedió a recolectar muestras de cepas fúngicas
presentes en la parte baja de la paja brava y quinua real, las muestras se tomaron de la raíz, del
tallo, del nudo vital, e incluso del suelo que rodeaba las mencionadas plantas, las muestras
fueron tomadas utilizando pinzas estériles, preparadas a los métodos tradicionales. El
muestreo se realizo utilizando frascos de vidrio estériles, frascos de plástico irradiados por luz
U.V., y bolsas de polietileno.
Figura 6. Paja Brava a orillas del río Desaguadero.
Figura 7. Suelo donde crece la planta Paja Brava a orillas del lago Titicaca.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
4
Se tomaron los datos de muestreo de cada espécimen recolectado con los siguientes
parámetros: ubicación geográfica, la altura, descripción del lugar y apariencia general de la
planta (ANEXO B-2).
En este viaje realizado se aprovecho para la recolección de algunos sustratos utilizados
más adelante, tales como la paja brava, residuos de quinua real.
De la toma de muestra realizada se recolectaron 30 muestras diferentes, entre ellas 19
pertenecían a la paja brava, 5 a quinua real, 4 a tierra que rodeaba mencionadas plantas, y 2 a
las setas.
3.2 METODOS DE AISLAMIENTO DE CEPAS FUNGICAS.-
La inoculación se realizo de dos formas, una indirecta y otra directa.
INDIRECTO: Se preparo diluciones de hipoclorito de sodio al 10% (v/v), alcohol al
70% (v/v). La muestra fue sumergida por 5 segundos en cada dilución y finalmente en agua
destilada y fue trasladada a medio Papa Dextrosa Agar (PDA).
DIRECTO: Se traslado la muestra directamente en medio Papa Dextrosa Agar (PDA).
En ambos casos se incubo a temperatura de 20ºC durante 5 días. Se comprobaron la
pureza de las cepas aisladas mediante resiembras en medio Papa Dextrosa Agar (PDA).
De las 30 muestras recolectadas, se logro aislar 102 cepas fúngicas, de las cuales
muchas eran repetidas, obteniéndose 31 cepas fúngicas diferentes (anexo B-1).
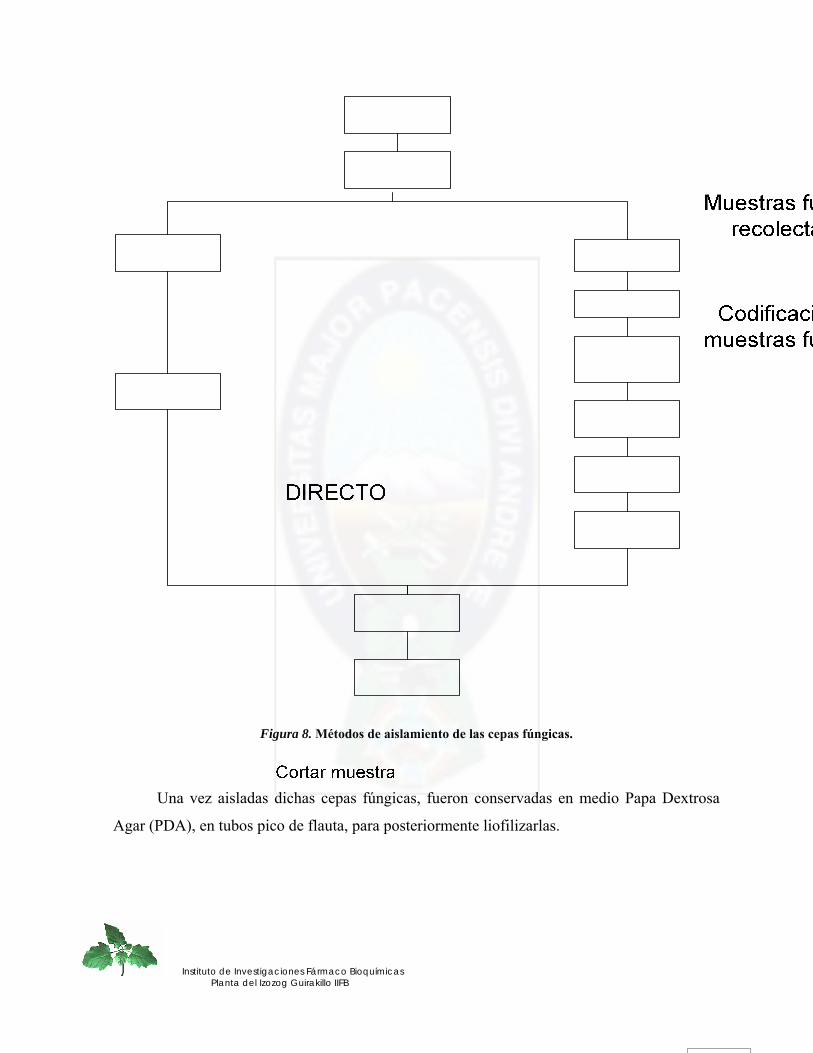
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
46
Figura 8. Métodos de aislamiento de las cepas fúngicas.
Una vez aisladas dichas cepas fúngicas, fueron conservadas en medio Papa Dextrosa
Agar (PDA), en tubos pico de flauta, para posteriormente liofilizarlas.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
4
Figura 9. Tronco de madera que contiene cepas fúngicas, recolectada de orillas del lago Titicaca.
Las muestras recolectadas fueron manipuladas con los debidos cuidados, y a las 24 hrs.
de ser recolectadas.
Figura 10. Preparación de medio Papa Dextrosa Agar (PDA).
La preparación de los medios de cultivo se realizó a condiciones estériles, usando los
materiales adecuados y las normas de Bioseguridad.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
48
Figura 11. Cepas fúngicas en crecimiento a temperatura ambiente.
El crecimiento de las cepas fúngicas aisladas fue a unos 20ºC, teniendo cuidado de no
cruzar las cepas, esto por la esporulación de los hongos.
3.2.1. Codificación de cepas fúngicas aisladas.-
El presente trabajo se realizo en el Instituto de Investigaciones fármaco Bioquímicas
(IIFB) de la Facultad de ciencias Farmacéuticas y Bioquímicas, en conjunto con el Instituto de
Investigación y Desarrollo de Procesos Químicos (IIDEPROQ) de la Facultad de Ingeniería,
motivo el cual se codifico a las cepas fúngicas aisladas como IB.
I = Ingeniería B = Bioquímica
Para la codificación de las cepas fúngicas aisladas se les asigno un numero según el
orden de muestras recolectadas, seguidas de una letra mayúscula el cual representa el primer
aislamiento de la muestra , seguido de una letra minúscula el cual significa el segundo
aislamiento de la misma muestra.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
49
3.3 METODOS DE IDENTIFICACION DE CEPAS FUNGICAS.-
Una vez aisladas las cepas fúngicas se procedió a identificarlas mediante la
identificación de los hongos filamentosos se basa en el examen macroscópico de la colonia y
en sus características microscópicas.
3.3.1 Identificación Macroscópica.-
Se observo cuidadosamente todas las características que pudieran presentar dichas
cepas, tales como: tiempo de crecimiento, forma de crecimiento, color característico de la cepa
fúngica, para así poder compararla con la bibliografía e identificarla según mencionadas
características.
Semejanzas macroscópicas como la forma de la colonia, el color de la superficie, la
textura y la producción de pigmentos son muy útiles para la identificación.
En general, la morfología macroscópica de los hongos es estable y presenta pocas
variaciones. La identificación definitiva se basa en la forma característica, método de
producción y ordenamiento de las esporas, siendo también importante conocer el tamaño y la
disposición de las hifas [18].

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
0
A: Vista anterior de la cepa fúngica B: Vista posterior (observamos el (se observa micelios algodonados) (pigmento que produce el hongo)
Figura 12. Morfología macroscópica de la cepa fúngica
Los estudios macroscópicos consistieron en evaluar el tipo de micelio aéreo, tipo de micelio
vegetativo, color del micelio vegetativo y del micelio aéreo, producción de pigmentos
difusibles en el medio de cultivo y consistencia de las colonias. Todas estas observaciones se
realizaron en medio Papa Dextrosa Agar (PDA).
3.3.2 Identificación Microscópica.-
Se basa en la observación al microscopio con la lente (40x) de la muestra de la cepa
fúngica en un portaobjetos, evaluar el tipo y disposición de hifas, micelios y esporas de cada
una de las cepas, guiándonos en la bibliografía, y así poder identificarlas.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
A: Selección de colonia fúngica. B: Preparación de muestra
Figura 13. Identificación microscópica
La observación al microscopio permite distinguir la forma y disposición de las esporas. Este
método permite la observación microscópica del hongo mientras se desarrolla directamente
debajo del cubreobjetos, de forma que las características microscópicas se distinguen con
facilidad, las estructuras se mantienen intactas y puede observarse un gran número de áreas
representativas [18]. Se comparo información morfológica y taxonómica con bibliografía.
Muchas especies de hongos se pueden diferenciar, identificar y clasificar según su
morfología, estructura, mecanismo de formación y elementos formadores de las esporas. El
conocimiento de estas características puede ser suficiente para identificar especies que
pertenecen al grupo de los hongos filamentosos; sin embargo, cuando se trata de identificar
otras especies es imprescindible la aplicación de pruebas de biología molecular.
Se identificaron microorganismos de los géneros Circinutrichum, Stephanoma,
Cephalosporium, Bdellospora, Blatomyces, Trinacrium, Verticium, Dendrodochium,
Choanephora, Botritys, Paecielomyces, Mycotypha.
De las 31 cepas fúngicas asiladas (100%), solo 25 cepas fúngicas (80,64%) fueron
identificadas, el resto 6 (19, 35%) no pudieron ser identificadas, ya muchas de ellas no
coincidían con la taxonómica de ninguna bibliografía utilizada.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
2
A: Morfología microscópica Botrytis B: Morfología microscópica Cephalosporium
Figura 14. Morfología microscópica
3.3.3 Elaboración de Historial de cepas fúngicas.-
Una vez identificadas las cepas fúngicas, se procedió a elaborar un historial de las cepas
fúngicas recolectadas (ver anexo B).
Este contiene información sobre: numero de muestra, lugar de origen, coordenadas de
la ubicación geográfica, la altura, nombre común, la descripción general del hábitat, apariencia
general, nombre científico de las cepas fúngicas identificadas.(ver anexo A-2). La
identificación microscópica se realizo de acuerdo al procedimiento descrito (3.3.1). Se
comparo información morfológica y taxonómica con bibliografía.
3.4 METODOS DE DETERMINACION DE LA ACTIVIDAD
ENZIMATICA.-
3.4.1 Determinación Cualitativa.-
En este estudio, se determino la actividad enzimática ligninolítica que sean capaces de
producir cepas fúngicas.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
3
Figura 15. Preparación de medio ABTS y Poli – R.
Ver composición en ANEXO A-1
Las cepas identificadas por sus características morfológicas en medio PDA de esos
cultivos se tomo una pieza de micelio aproximadamente de 10mm de diámetro, esos fueron
transferidos a un medio sólido Poly R-478, y otra pieza de micelio del mismo diámetro se
traslado a un medio sólido ABTS, el diámetro de decolorización fue examinado después de 2
días, hasta los 5 días, a 20ºC, se tuvo cuidado de incubarlas a oscuridad, ya que luz puede
afectar al respuesta, y podríamos obtener resultados verdaderos positivos, por ultimo, fue
comparado con las zonas formadas del medio de referencia de Trametes versicolor, esto por
bibliografía [56].
Las figuras 4-5 muestra claramente el resultado de esta prueba, ya que se puede notar
la decolorización del medio alrededor de la cepa fúngica en crecimiento, este resultado se vio
entre el tercer y quinto día. La decolorización se debe a la oxidación del colorante Poly R-478
por acción de las enzimas ligninoliticas, lo que significa que el microorganismo es capaz de
degradarlo y por ende producir dichas enzimas [34].

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
4
Figura 16. Decolorización del medio sólido Poly R
Fuente: Elaboración propia
La figura 4-6 muestra el resultado de decoloración del medio solidó ABTS, este
resultado se presento a los 3 días y nos da a entender que el cambio de color es causada por la
oxidación del colorante ABTS, lo cual indica que la cepa fúngica en el medio tiene la
capacidad de producir enzimas ligninolíticas con actividad ligninolitica.
Figura 17. Decolorización del medio sólido ABTS.
Fuente: Elaboración propia
El ABTS (2,2'-Azinobis (3-ethylbenzthiazoline-6-sulfonic acid).2NH4-salt)
(figura 4-6) es un compuesto el cual tiene una estructura compleja, en este sentido podemos
decir que cuando hay una degradación de ABTS lo cual esta representado por la
decolorización, hay presencia de enzimas ligninolíticas [41].

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
Figura 18. Estructura generaliza de ABTS (2,2'-Azinobis (3-ethylbenzthiazoline-6-sulfonic acid).2NH4-salt)
3.4.2 Determinación Cuantitativa.-
De la cepa fúngicas que presentaron elevadas actividades según los criterios
establecidos anteriormente, se seleccionaron las que como resultado dieron positiva,
para determinar esta actividad enzimática se realizo un diseño factorial 23, siendo los factores:
glucosa, manganeso, extracto de levadura.
Obteniendo un diseño factorial 23 con 8 combinaciones posibles, se utilizaron frascos
de vidrio de 100 ml de capacidad, cada uno con 10 ml de medio de cultivo Mn (II). A partir
del cultivo agarizado, se tomo discos de agar de 10mm de diámetro que fueron transferidos a
los frascos con medio de cultivo, y se incubo en oscuridad, a 20ºC, durante 4 semanas, se tomo
una muestra de cada medio de cultivo cada 7 días para determinaciones de las actividades
enzimáticas, proteínas totales y celulasas, cabe recalcar que las pruebas fueron realizadas por
triplicado, esto para confirmar resultados.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
6
Figura 19. Determinación cualitativa de la actividad enzimática ligninolítica.
Tabla 8: Composición de medios de cultivo Mn (II), resultado del diseño factorial 23
Por otro lado se determinó la influencia de glucosa, extracto de levadura, manganeso,
sobre la producción de las enzimas ligninolíticas por parte de los hongos seleccionados.
Glucosa: Fuente de carbono
Medio Glucosa (g/L) Manganeso(mg/L) Ext. de lev. (g/L) 1 1 40 0.5 2 1 40 2 3 1 80 0.5 4 5 40 0.5 5 1 80 2 6 5 40 2 7 5 80 0.5 8 5 80 2

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
El medio de cultivo utilizado tiene como única fuente carbono a la glucosa, por tanto
podremos verificar que tan importante es el carbono para la producción de este tipo de
enzimas. Considerando a al glucosa como fuente de carbono e inductor de la actividad
enzimática, en estudios en los cuales se utilizo fuente de carbono para producción de este tipo
de enzimas fue observado que las fuentes utilizadas fueron eficientes y rápidamente utilizadas,
resultando al altos niveles de actividades [34].
Extracto de levadura: Fuente de nitrógeno y vitaminas
El extracto de levadura contiene aminoácidos, péptidos, vitaminas solubles en agua y
carbohidratos. La producción de enzimas ligninolíticas esta en dependencia tanto de fuente de
carbono como de nitrógeno, en este trabajo fue utilizado como fuente de nitrógeno y fuente de
vitaminas el extracto de levadura, lo cual pueda explicar varias concentraciones encontradas, a
pesar de la importancia de la fuente de nitrógeno para la producción de estas enzimas ha sido
recientemente apuntada por algunos autores que su actividad es mayor en condiciones
limitantes de nitrógeno [38].
Manganeso: Inductor de enzimas
La producción de enzimas por hongos, por el medio modificado por el diseño factorial,
requieren glucosa y manganeso según bibliografía. Las actividades de MnP, LiP aumentan
simultáneamente por la adición del mencionado en el medio el cual puede estimular la
producción de mas MnP que LiP [1].

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
8
Figura 20. Fotografía del equipo Espectrofotómetro de absorción.
Figura 21. Medio de cultivo Mn (II) para determinación cuantitativa.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
9
Figura 22. Determinación actividad enzimática ligninolítica por métodos espectrofotométricos.
De los medios de cultivo preparados se tomo muestra del sobrenadante de cada frasco,
se tomo 0,5 mL de muestra, cuidando de no extraer micelio, se procedió a determinar tres
tipos de enzimas ligninolíticas: Manganeso peroxidasa, Lignina peroxidasa y Lacasa.
A continuación detallamos el procedimiento de cada enzima.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
60
a. Manganeso Peroxidasa.-
Para determinar la actividad de esta enzima se leyó la variación de absorbancia del
cultivo (entre 120 segundos) en espectrofotómetro (470nm), en cubetas de plástico con: 2,6
DMF 1mM (100μ), MnSO4 3mM (100μl), buffer succinato pH 5 100mM (400μl),
sobrenadante de medio de cultivo (100μl) y peroxido de hidrogeno 0,5mM (100μl).
La actividad para MnP fue estimada por oxidación de 2,6 dimetoxifenol, a coerulignona
(ε470 = 24500 M-1cm-1) monitoreada a 470nm a pH 5.
Figura 23. Reacción enzimática para MnP por oxidación de 2,6 dimetoxifenol
La figura 4-8 muestra el cambio de coloración cuando se lleva acabo la reacción
producida por MnP, la diferencia es clara, para la cepa IB-26C (Trinacrium) que fue la que
presento mayor actividad enzimática que las demás.
La MnP, esta enzima es una peroxidasa que oxida componentes fenólicos de la lignina,
mediante la reacción de oxidación del Mn2+ a Mn3+, la cual es dependiente del H2O2, ya que la
reacción es iniciada por este compuesto[80]. Una unidad de actividad enzimática de MnP es
definida como la formación de coerulignona por minuto.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
6
b. Lignina Peroxidasa.-
La actividad de esta enzima se determino mediante la variación de absorbancia del
cultivo (entre 120 segundos) en espectrofotómetro (420nm) en cubetas de plástico con: ABTS
10mM (100μl), buffer acetato pH 3 100mM (400μl), sobrenadante de medio de cultivo (100μl)
y peroxido de hidrogeno 0,5mM (100μl).
LiP fue medida a pH 3 por oxidación de ABTS en presencia de agua oxigenada a un
catión radical (ε420 = 36000 m-1 cm-1) por el monitoreo en el cambio de absorbancia a 420 nm
[34] (figura 4-12).
La LiP que su actividad depende del peroxido de hidrogeno, además posee algunas
características con las hemoproteínas transportadoras de oxigeno.
La degradación que causa sobre de lignina se da por la ruptura de anillos aromáticos,
por síntesis endógena oxida compuestos aromáticos no fenólicos, en la reacción se generan
radicales arilo y alquilo que se anabolizan intracelularmente.
Figura 24. Reacción enzimática para LiP por oxidación de ABTS.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
62
c. Lacasa.-
La actividad de la lacasa se determino mediante la variación de absorbancia del cultivo
(entre 120 segundos) en espectrofotómetro (420nm) en cubetas de plástico con: ABTS 10mM
(100μl), buffer acetato pH 3 100mM (400μl), sobrenadante de medio de cultivo (100μl).
La actividad de la lacasa se determino mediante la oxidación del sustrato ABTS la
oxidación se midió mediante el incremento de absorbancia (ε420 = 36000 m-1 cm-1) a 420nm (ver
figura 4-16). Lacasa una fenol oxidasa con cobre, enzima que reduce el oxigeno molecular a
agua y a través la utilización de ciertos compuestos redox puedes ser capaz de ampliar su
espectro de sustratos, logrando así la oxidación de porciones no fenólicas de la lignina [12].
Figura 25. Reacción enzimática para Lac por oxidación de ABTS.
Esta enzima con cuatro átomos de cobre como cofactor y cuyo aceptor de electrones es
el O2 dando agua como producto [86]. La enzima oxidada ataca principalmente sustratos
fenólicos, pero en este caso en presencia de ABTS [87,88]. Estos actúan como cooxidantes y
capacitan ala enzima para atacar sustratos más difícilmente oxidables como los de tipo no
fenólico.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
63
Para todas las medidas de actividad enzimática, una unidad de actividad enzimática fue
definida como la cantidad de enzima, que catalizó la formación de 1 mol de producto por
minuto bajo condiciones del análisis.
3.4.3 Cuantificación de las Enzimas.-
Determinado por la ecuación (1) según bibliografía.
[ ] ⎟⎠⎞
⎜⎝⎛ ×== −1
m
t min L
U Y V x ε
xCV x 1000 x AbsiónConcentrac
Abs: variación de la absorbancia en los dos minutos (min.-1)
Vt: volumen total de la solución (mL)
Vm: volumen de la muestra
ε: coeficiente de extinción molar, donde:
ε470 = 24500 M-1cm-1 (Manganeso Peroxidasa)
ε420 = 36000 m-1 cm-1 (Lignina Peroxidasa – Lacasa).
1000: factor de conversión de moles a moles.
C: distancia de la cubeta por donde pasa el haz luminoso (1cm)
3.5 OPTIMIZACION DE MEDIOS DE CULTIVO PARA LA PRDUCCION
DE ENZIMAS LIGNINOLITICAS.-
Una vez cuantificada la actividad enzimática ligninolítica, se procedió a realizar una
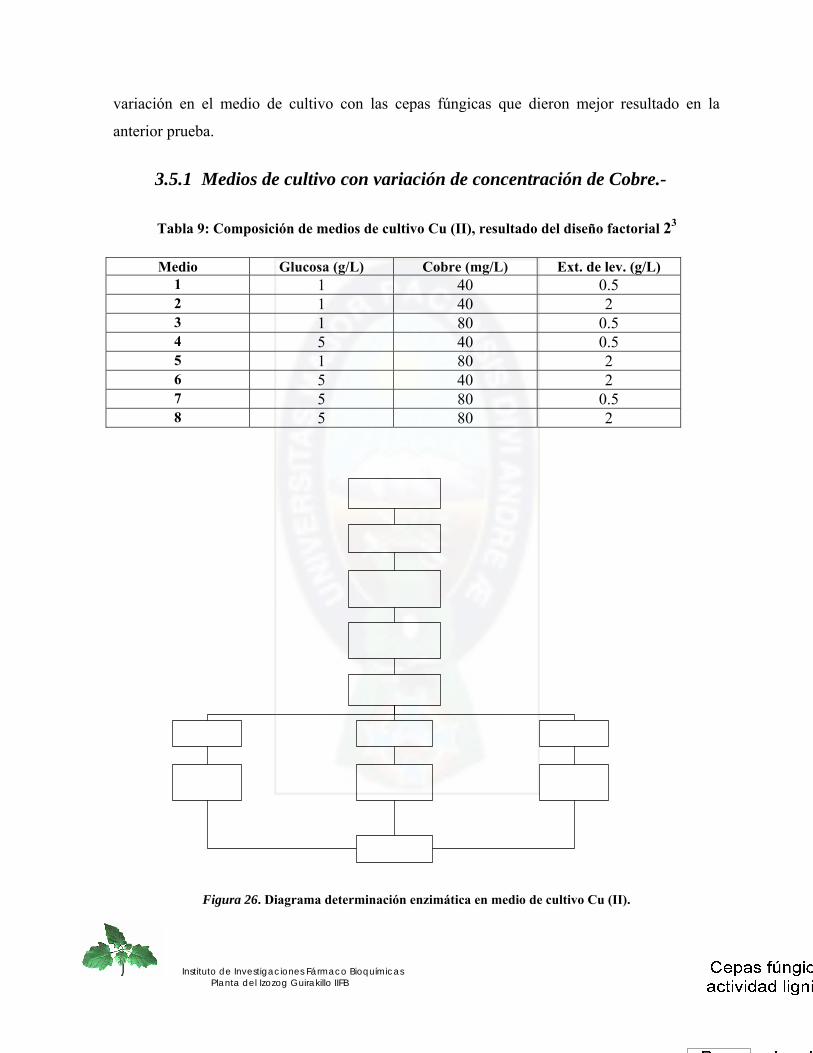
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
64
variación en el medio de cultivo con las cepas fúngicas que dieron mejor resultado en la
anterior prueba.
3.5.1 Medios de cultivo con variación de concentración de Cobre.-
Tabla 9: Composición de medios de cultivo Cu (II), resultado del diseño factorial 23
Figura 26. Diagrama determinación enzimática en medio de cultivo Cu (II).
Medio Glucosa (g/L) Cobre (mg/L) Ext. de lev. (g/L) 1 1 40 0.5 2 1 40 2 3 1 80 0.5 4 5 40 0.5 5 1 80 2 6 5 40 2 7 5 80 0.5 8 5 80 2

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
6
Se preparo un medio de cultivo Cu (II), en el cual se hizo un diseño factorial 23 con 8
combinaciones posibles, siendo los factores: glucosa, manganeso, extracto de levadura. Se
inoculo las cepas fúngicas, se incubo a temperatura ambiente en oscuridad, a 20 ºC, durante 4
semanas, se tomo una muestra de cada medio de cultivo cada 7 días, las pruebas fueron
realizadas por triplicado, de igual forma que la anterior se procedió a determinar la actividad
enzimática de MnP, LiP y Lacasa.
3.5.2 Medios de cultivo con diferentes sustratos lignocelulosicos.-

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
66
Figura 27. Determinación enzimática en medios de cultivo con diferentes sustratos.
Se preparo un medio de cultivo igual al anterior, con la diferencia de que en este medio
de cultivo se suprimió la glucosa, y en su lugar se trabajo con diferentes sustratos
lignocelulosicos, como ser: paja brava, bagazo, aserrín, residuos de quinua real, y cáscara de
arroz., se inocularon las cepas fúngicas en este medio, se incubo a temperatura ambiente en
oscuridad, a 20 ºC, durante 4 semanas, se tomo una muestra de cada medio de cultivo cada 7
días, las pruebas fueron realizadas por triplicado, de igual forma que la anterior, se procedió a
determinar la actividad enzimática de MnP, LiP y Lacasa.
3.6 Determinación de Actividad Celulolítica.-

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
6
Figura 28. Determinación de la actividad Celulolítica.
Se realizo la determinación cuantitativa de la actividad enzimática celulolítica
combinada, se utilizo el método del “Acido 3,5 Dinitrosalicilico” (DNS), donde se agrego a la
muestra (500μl) buffer citrato (1000μl) y se adiciono 100g de papel filtro, se dejo incubar por
60min., posteriormente se adiciono reactivo DNS (3000μl), ebullir la mezcla por 5min. y se
procede a leer la absorbancia de la muestra en espectrofotómetro (550nm).
3.7 Cuantificación de proteínas totales.-
La concentración de proteínas de los sobrenadantes de los cultivos se midió por el

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
68
método de Lowry, utilizando el reactivo y el procedimiento de Bio-Rad reactivo folin
Ciocalteo en un espectrofotómetro, teniendo en cuenta la capacidad de union del colorante del
reactivo a dichas moléculas, posteriormente se leyó de absorbancia de esta muestra en
espectrofotómetro (660nm) en cubetas de plástico.
Muestra (sobrenadante de medio de cultivo) 0,1 ml
Añadir agua (0,1ml) + reactivo A (1ml)
Incubar a 35ºC por 35 min.
Adicionar reactivo B ( 0,1ml)
Incubar por 20 min. a 37 ºC
Leer abs. (660nm)
Figura 29. Determinación de proteínas totales.
3.8 Comparación de la Actividad Enzimática Ligninolítica de las enzimas
producidas frente a enzimas comerciales.-
Para poder verificar la efectividad de las enzimas ligninolíticas producidas en el
presente trabajo, se las comparo frente a enzimas comerciales, esto como una prueba no
especifica, ya que las enzimas producidas no están previamente purificadas.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
69
Enzima comercial purificada
Dilución de la enzima
Lignina peroxidasa LacasaManganeso peroxidasa
Variación abs. (470 nm) 2,6 DMF, H2O2
Variación abs. (420 nm) ABTS,
H2O2
Variacion abs. (420 nm) ABTS
Cuantificación de enzimas
Comparacion de la actividad enzimatica frente a las enzimas
producidas
Figura 30. Determinación de la actividad enzimática de enzimas comerciales.
Las enzimas comerciales utilizadas para esta prueba fueron Lacase de “Rhus
vernificera” y Peroxidase de “Horseradish”.
3.9 Análisis estadístico.-
El análisis de varianza (ANOVA), fue realizado en el software estadístico Statistica
Release 6.0 Stat Soft con un nivel de confianza de 95%. Las superficies de respuesta fueron
realizadas en el programa Microsoft Office Excel 2003.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
0
4. RESULTADOS Y DISCUSION
4.1 DETERMINACION CUALITATIVA DE LA ACTIVIDAD ENZIMATICA
LIGNINOLITICA.-
A continuación se presenta la tabla 4-1 en la que se resume los resultados positivos,
obtenidos en la determinación cualitativa de actividad ligninolítica.
Tabla 10: Determinación de la actividad ligninolítica en medio solidó.
Fuente: Elaboración propia
(-) No hubo decoloración (+) decoloración poco visible
(++) decoloración media (+++) buena decoloración
Cuatro cepas dieron positivo en los dos medios de cultivo, fueron: IB-15B (Stephanoma),
IB-26C (Trinacrium), IB-15A (Dactylella), IB-29B (Blasthomyces).
Cinco cepas fúngicas mostraron un resultado positivo para el medio sólido ABTS,
fueron: IB-15Da (Cephalosporium), IB-18Cb (Bdellospora), IB-18Cb (Bdellospora), IB-20Ca
(Mycotypha), IB-14A (Triposporium), IB-6 (Circinutrychum).
Cepas fúngicas ABTS Poli-R
IB-15B (Stephanoma) +++ +
IB-15Da (Cephalosporium) ++ -
IB-26C (Trinacrium) +++ +
IB-18Cb (Bdellospora) ++ -
IB-15A (Dactylella) + ++
IB-30B (Sepedonium) - ++
IB-29B (Blasthomyces) + +++
IB-5 (Spiromyces) - +
IB-20Ca (Mycotypha) ++ -
IB-14A (Triposporium) + -
IB-18B (Fusoma) - ++
IB-6 (Circinutrychum) + -

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
Las tres cepas restantes presentaron un resultado positivo respecto al medio sólido Poly
R-478, fueron: IB-30B (Sepedonium), IB-5 (Spiromyces), IB-18B (Fusoma).
De las 31 (100%) cepas fúngicas aisladas que fueron inoculadas en medios ABTS y
Poly-R, 12 (38,7%) cepas fúngicas presentaron cambio de coloración en el medio, es decir
presentaron actividad ligninolítica, el restos de los hongos 19 (61,3%) no presentaron
actividad ligninolítica, por que no se los tomo en cuenta para otras pruebas en adelante.
4.2. DETERMINACION CUANTITATIVA DE LA ACTIVIDAD ENZIMATICA
LIGNINOLITICA.-
4.2.1. Manganeso Peroxidasa.-
De las doce cepas fúngicas con actividad enzimática, los resultados fueron los
siguientes, para MnP.
Como se puede observar en la tabla 11 los resultados son muy variados para cada cepa
fúngica, ya que varían en el medio de cultivo, ya que cada una tiene diferentes requerimientos,
pero el tiempo necesario para alcanzar el máximo de producción (o actividad) de manganeso
peroxidasa es el mismo. La concentración de enzima detectada es influida por el medio de
cultivo que se utilice (ver anexo C-1).
Las doce cepas fúngicas alcanzo su actividad máxima a un tiempo de 28 días.
Tabla 11: Actividad enzimática máxima para MnP de diferentes cepas fúngicas con variación
Mn
Cepa fúngica Medio Glucosa (g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
MnP (UI/L)
IB-26C (Trinacrium) 6 5 40 2 28 76,65
IB-15B (Stephanoma) 3 1 80 0.5 28 57,89
IB-29B (Blasthomyces) 5 1 80 2 28 40,24

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
2
IB-18Cb (Bdellospora) 8 5 80 2 28 17,68
IB-15Da (Cephalosporiu) 2 1 40 2 28 13,60
IB-6 (Circinutrychum) 3 1 80 0,5 28 6,80
IB-20Ca (Mycotypha) 5 1 80 20 28 3,26
IB-5 (Spiromyces) 4 5 40 0,5 28 2,72
IB-18B (Fusoma) 4 5 40 0,5 28 2,31
IB-14A (Triposporium) 4 5 40 0,5 28 2,31
IB-30B (Sepedonium) 7 5 80 0,5 28 2,17
IB-15A (Dactylella) 6 5 40 2 28 1,90
La cepa fúngica IB-26C (Trinacrium) (figura 31) fue la que presento mayor actividad
enzimática para Manganeso peroxidasa (MnP) a los 28 días, en el medio 6, llegando a 76,65
(UI/L), en este medio existe mayor fuente de carbono, de vitaminas, de nitrógeno, pero la
concentración de manganeso es la mínima, por lo podemos afirmar que esta cepa no necesita
mayor concentración de manganeso para producir enzima.
La actividad de MnP es sensible a la presencia de algunos metales (manganeso, cobre,
hierro, molibdeno, zinc, níquel) son esenciales para el crecimiento fúngico y también actúan
como co-factores enzimáticos induciendo la actividad ligninolítica. Entretanto, estos metales
son tóxicos para el hongo cuando existen en exceso, ya que pueden inhibir el crecimiento de la
cepa fúngica y la actividad enzimática [83].
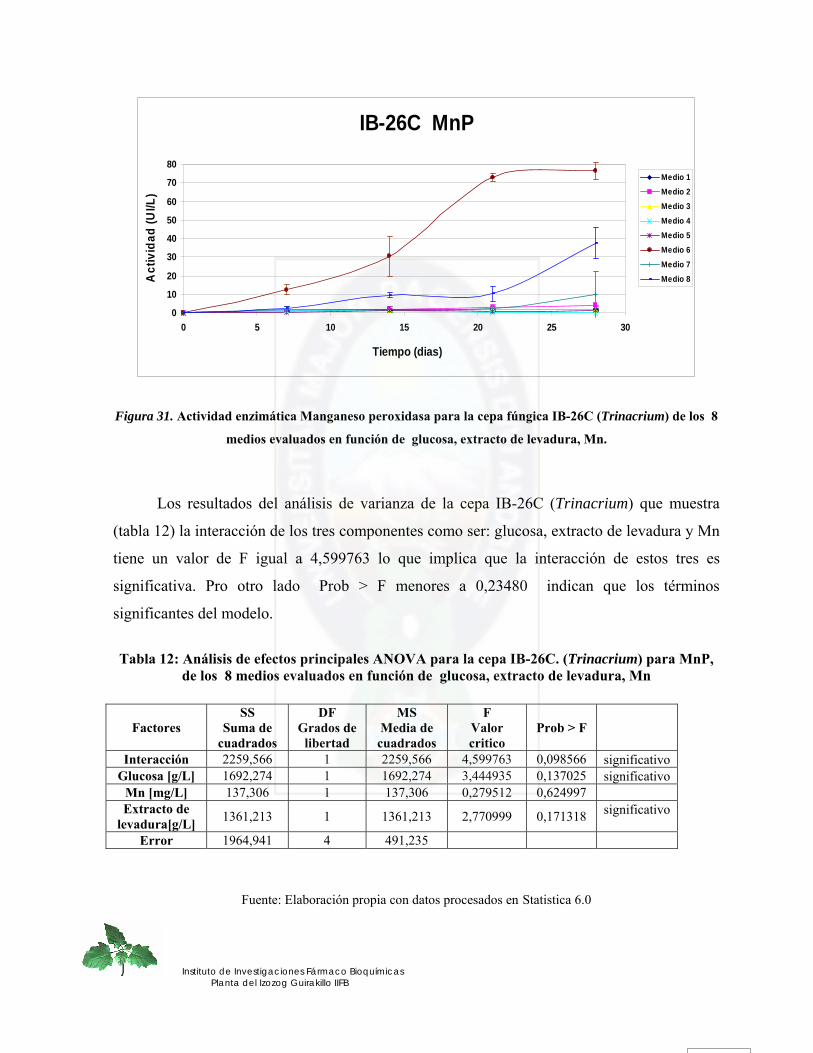
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
3
IB-26C MnP
0
10
20
30
40
50
60
70
80
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L)
Medio 1Medio 2Medio 3
Medio 4Medio 5Medio 6
Medio 7Medio 8
Figura 31. Actividad enzimática Manganeso peroxidasa para la cepa fúngica IB-26C (Trinacrium) de los 8
medios evaluados en función de glucosa, extracto de levadura, Mn.
Los resultados del análisis de varianza de la cepa IB-26C (Trinacrium) que muestra
(tabla 12) la interacción de los tres componentes como ser: glucosa, extracto de levadura y Mn
tiene un valor de F igual a 4,599763 lo que implica que la interacción de estos tres es
significativa. Pro otro lado Prob > F menores a 0,23480 indican que los términos
significantes del modelo.
Tabla 12: Análisis de efectos principales ANOVA para la cepa IB-26C. (Trinacrium) para MnP, de los 8 medios evaluados en función de glucosa, extracto de levadura, Mn
Fuente: Elaboración propia con datos procesados en Statistica 6.0
Factores SS
Suma de cuadrados
DF Grados de
libertad
MS Media de cuadrados
F Valor critico
Prob > F
Interacción 2259,566 1 2259,566 4,599763 0,098566 significativo Glucosa [g/L] 1692,274 1 1692,274 3,444935 0,137025 significativo
Mn [mg/L] 137,306 1 137,306 0,279512 0,624997 Extracto de
levadura[g/L] 1361,213 1 1361,213 2,770999 0,171318 significativo
Error 1964,941 4 491,235
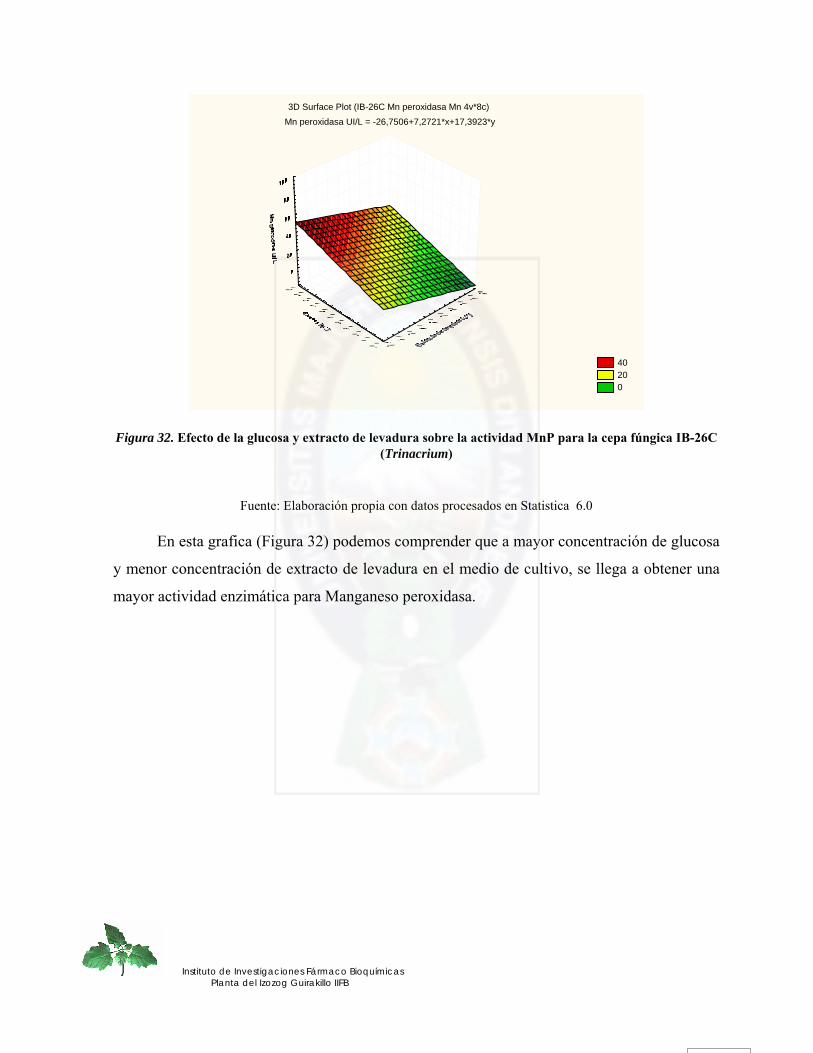
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
4
3D Surface Plot (IB-26C Mn peroxidasa Mn 4v*8c)Mn peroxidasa UI/L = -26,7506+7,2721*x+17,3923*y
40 20 0
Figura 32. Efecto de la glucosa y extracto de levadura sobre la actividad MnP para la cepa fúngica IB-26C (Trinacrium)
Fuente: Elaboración propia con datos procesados en Statistica 6.0
En esta grafica (Figura 32) podemos comprender que a mayor concentración de glucosa
y menor concentración de extracto de levadura en el medio de cultivo, se llega a obtener una
mayor actividad enzimática para Manganeso peroxidasa.
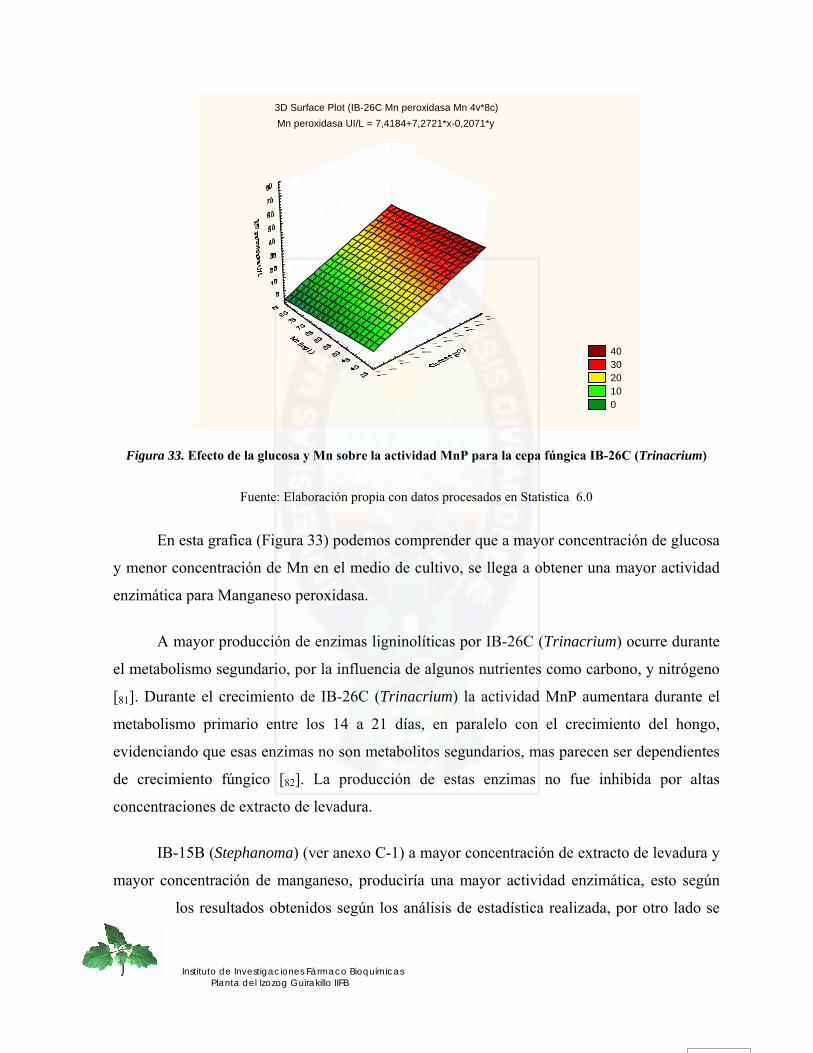
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
3D Surface Plot (IB-26C Mn peroxidasa Mn 4v*8c)Mn peroxidasa UI/L = 7,4184+7,2721*x-0,2071*y
40 30 20 10 0
Figura 33. Efecto de la glucosa y Mn sobre la actividad MnP para la cepa fúngica IB-26C (Trinacrium)
Fuente: Elaboración propia con datos procesados en Statistica 6.0
En esta grafica (Figura 33) podemos comprender que a mayor concentración de glucosa
y menor concentración de Mn en el medio de cultivo, se llega a obtener una mayor actividad
enzimática para Manganeso peroxidasa.
A mayor producción de enzimas ligninolíticas por IB-26C (Trinacrium) ocurre durante
el metabolismo segundario, por la influencia de algunos nutrientes como carbono, y nitrógeno
[81]. Durante el crecimiento de IB-26C (Trinacrium) la actividad MnP aumentara durante el
metabolismo primario entre los 14 a 21 días, en paralelo con el crecimiento del hongo,
evidenciando que esas enzimas no son metabolitos segundarios, mas parecen ser dependientes
de crecimiento fúngico [82]. La producción de estas enzimas no fue inhibida por altas
concentraciones de extracto de levadura.
IB-15B (Stephanoma) (ver anexo C-1) a mayor concentración de extracto de levadura y
mayor concentración de manganeso, produciría una mayor actividad enzimática, esto según
los resultados obtenidos según los análisis de estadística realizada, por otro lado se
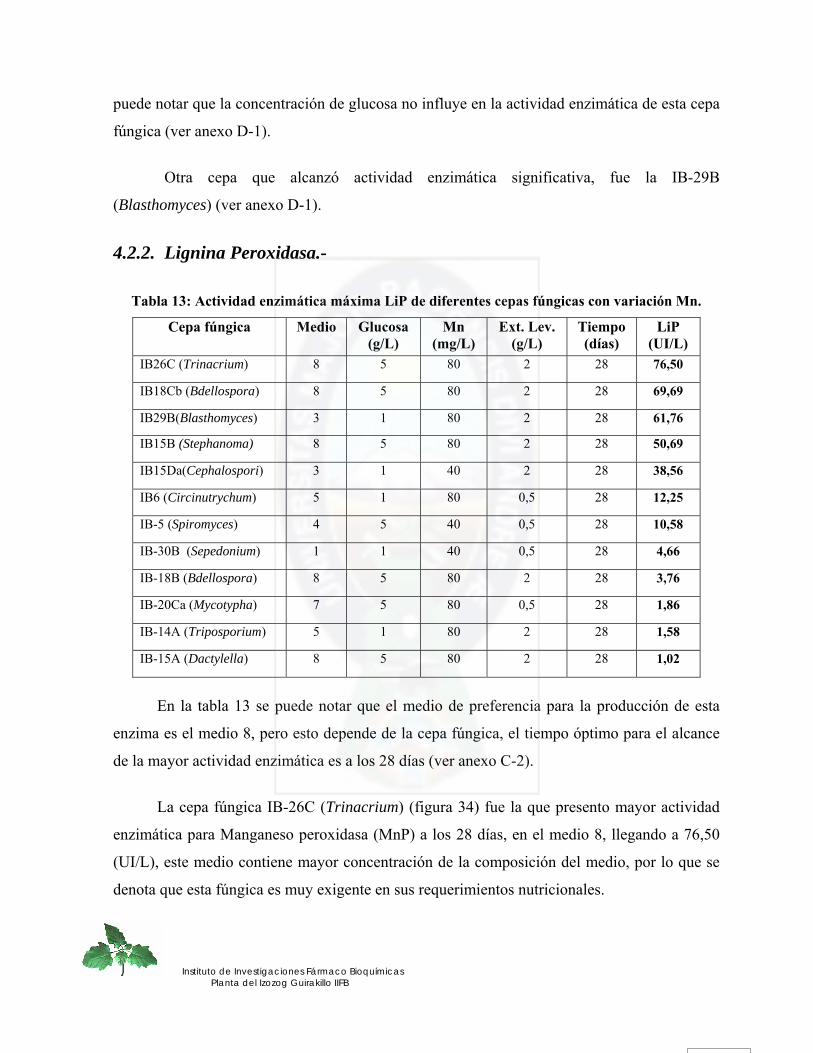
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
6
puede notar que la concentración de glucosa no influye en la actividad enzimática de esta cepa
fúngica (ver anexo D-1).
Otra cepa que alcanzó actividad enzimática significativa, fue la IB-29B
(Blasthomyces) (ver anexo D-1).
4.2.2. Lignina Peroxidasa.-
Tabla 13: Actividad enzimática máxima LiP de diferentes cepas fúngicas con variación Mn.
Cepa fúngica Medio Glucosa (g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
LiP (UI/L)
IB26C (Trinacrium) 8 5 80 2 28 76,50
IB18Cb (Bdellospora) 8 5 80 2 28 69,69
IB29B(Blasthomyces) 3 1 80 2 28 61,76
IB15B (Stephanoma) 8 5 80 2 28 50,69
IB15Da(Cephalospori) 3 1 40 2 28 38,56
IB6 (Circinutrychum) 5 1 80 0,5 28 12,25
IB-5 (Spiromyces) 4 5 40 0,5 28 10,58
IB-30B (Sepedonium) 1 1 40 0,5 28 4,66
IB-18B (Bdellospora) 8 5 80 2 28 3,76
IB-20Ca (Mycotypha) 7 5 80 0,5 28 1,86
IB-14A (Triposporium) 5 1 80 2 28 1,58
IB-15A (Dactylella) 8 5 80 2 28 1,02
En la tabla 13 se puede notar que el medio de preferencia para la producción de esta
enzima es el medio 8, pero esto depende de la cepa fúngica, el tiempo óptimo para el alcance
de la mayor actividad enzimática es a los 28 días (ver anexo C-2).
La cepa fúngica IB-26C (Trinacrium) (figura 34) fue la que presento mayor actividad
enzimática para Manganeso peroxidasa (MnP) a los 28 días, en el medio 8, llegando a 76,50
(UI/L), este medio contiene mayor concentración de la composición del medio, por lo que se
denota que esta fúngica es muy exigente en sus requerimientos nutricionales.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
La producción de LiP aparentemente es limitada por ciertos hongos basidiomicetos,
pero no se descarta la hipótesis de que otros hongos sean capaces de producir dicha enzima, su
excepcional potencial degradativo no esta limitado a la lignina, ya que la ultima década fueron
descritos como posibles sustratos para esta enzima derivados geo-bioquímicos de lignina, y
contaminantes orgánicos entre los cuales están los xenobioticos [76].
IB-26C LiP
0
10
20
30
40
50
60
70
80
0 5 10 15 20 25 30
Tiempo ( dias)
Act
ivid
ad (U
I/L)
Medio 1Medio 2Medio 3
Medio 4Medio 5Medio 6
Medio 7Medio 8
Figura 34. Actividad enzimática Lignina peroxidasa para la cepa fúngica IB-26C (Trinacrium) de los 8
medios evaluados en función de glucosa, extracto de levadura, Mn.
Fuente: Elaboración propia
La secreción de esta enzima probablemente resulta de modificaciones fisiológicas
características del metabolismo primario a metabolismo segundario, son diferentes
condiciones y tipo de sustrato [76].
La actividad enzimática alcanzada se puede deber a la interacción de los componentes
del medio y la concentración, lo cual favorezca al metabolismo del microorganismo y se
pueda hacer posible la producción de enzima tipo LiP.
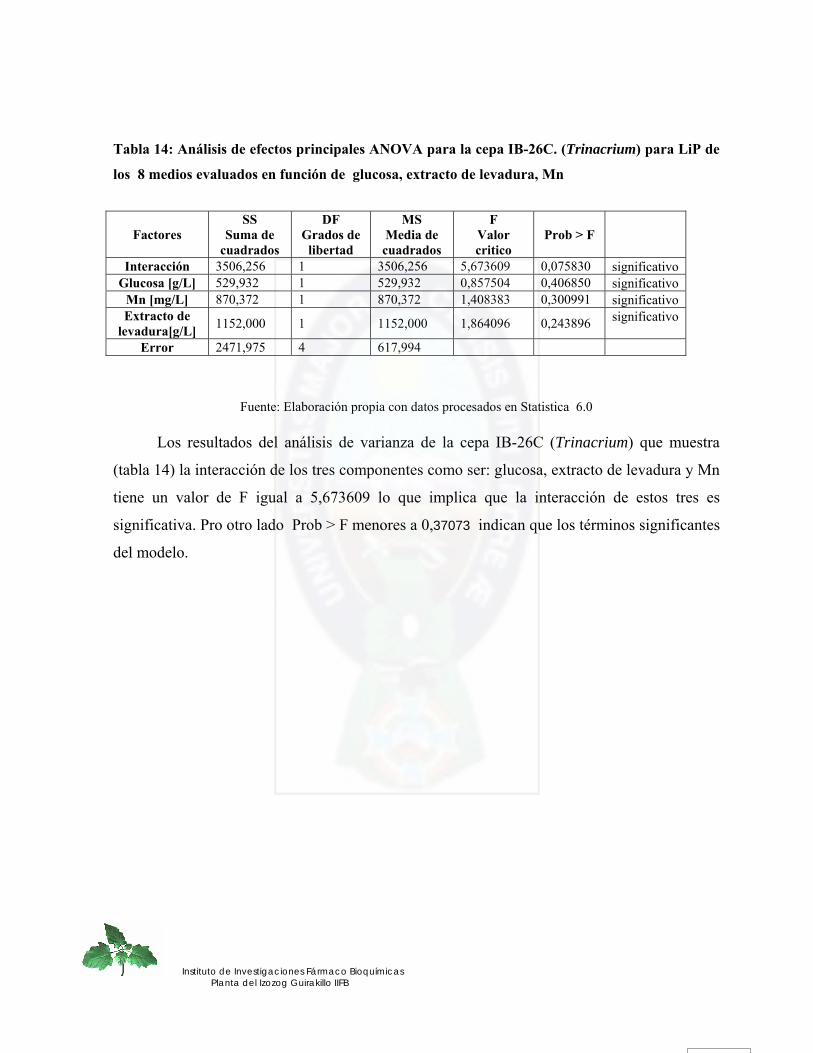
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
8
Tabla 14: Análisis de efectos principales ANOVA para la cepa IB-26C. (Trinacrium) para LiP de
los 8 medios evaluados en función de glucosa, extracto de levadura, Mn
Fuente: Elaboración propia con datos procesados en Statistica 6.0
Los resultados del análisis de varianza de la cepa IB-26C (Trinacrium) que muestra
(tabla 14) la interacción de los tres componentes como ser: glucosa, extracto de levadura y Mn
tiene un valor de F igual a 5,673609 lo que implica que la interacción de estos tres es
significativa. Pro otro lado Prob > F menores a 0,37073 indican que los términos significantes
del modelo.
Factores SS
Suma de cuadrados
DF Grados de
libertad
MS Media de cuadrados
F Valor critico
Prob > F
Interacción 3506,256 1 3506,256 5,673609 0,075830 significativo Glucosa [g/L] 529,932 1 529,932 0,857504 0,406850 significativo
Mn [mg/L] 870,372 1 870,372 1,408383 0,300991 significativo Extracto de
levadura[g/L] 1152,000 1 1152,000 1,864096 0,243896 significativo
Error 2471,975 4 617,994

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
9
3D Surface Plot (IB-26C Li peroxidasa Mn 4v*8c)Li peroxidasa UI/L = -11,2731+4,0694*x+16*y
40 30 20 10 0
Figura 35. Efecto de la glucosa y extracto de levadura sobre la actividad LiP para la cepa fúngica IB-26C (Trinacrium)
Fuente: Elaboración propia con datos procesados en Statistica 6.0
En esta grafica (Figura 35) podemos comprender que a mayor concentración de glucosa
y menor concentración de extracto de levadura en el medio de cultivo, se llega a obtener una
mayor actividad enzimática para Lignina peroxidasa, es posible que este hongo requiera
mayor fuente de carbono para aumentar su actividad enzimática, ya que la glucosa, como bien
sabemos puede ser un co-factor para la producción de estas enzimas, de todas maneras el
extracto de levadura no influye en la actividad de la enzima.
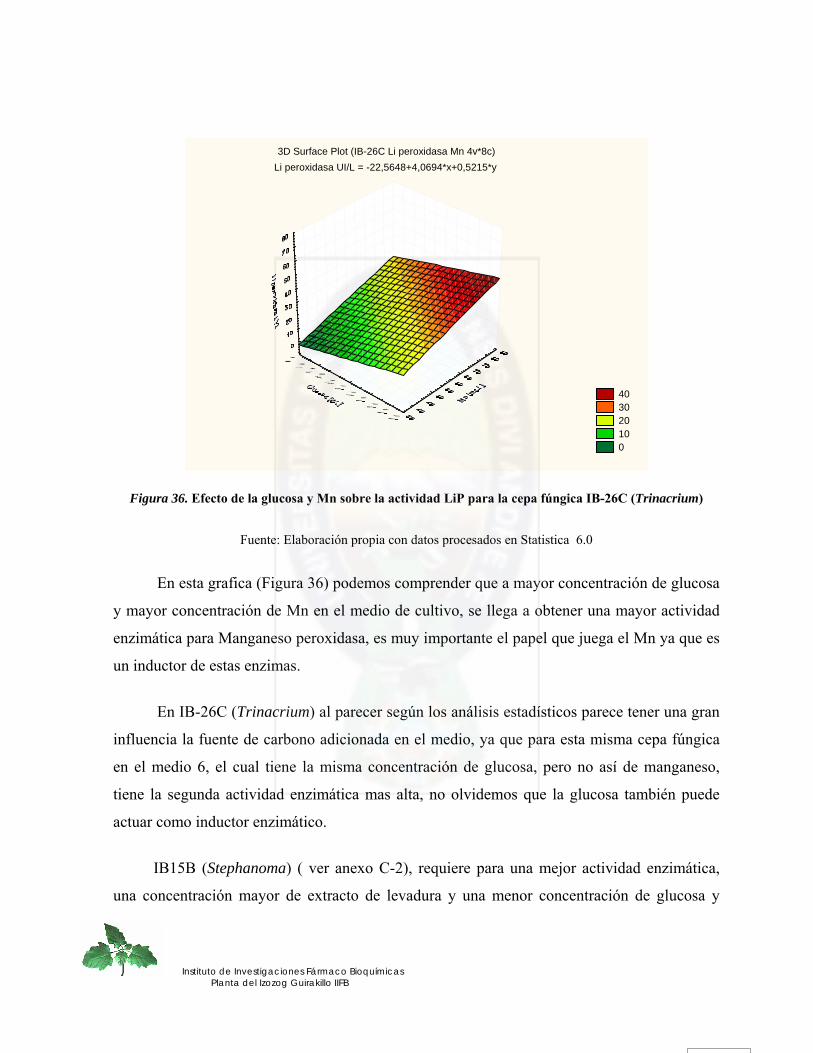
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
80
3D Surface Plot (IB-26C Li peroxidasa Mn 4v*8c)Li peroxidasa UI/L = -22,5648+4,0694*x+0,5215*y
40 30 20 10 0
Figura 36. Efecto de la glucosa y Mn sobre la actividad LiP para la cepa fúngica IB-26C (Trinacrium)
Fuente: Elaboración propia con datos procesados en Statistica 6.0
En esta grafica (Figura 36) podemos comprender que a mayor concentración de glucosa
y mayor concentración de Mn en el medio de cultivo, se llega a obtener una mayor actividad
enzimática para Manganeso peroxidasa, es muy importante el papel que juega el Mn ya que es
un inductor de estas enzimas.
En IB-26C (Trinacrium) al parecer según los análisis estadísticos parece tener una gran
influencia la fuente de carbono adicionada en el medio, ya que para esta misma cepa fúngica
en el medio 6, el cual tiene la misma concentración de glucosa, pero no así de manganeso,
tiene la segunda actividad enzimática mas alta, no olvidemos que la glucosa también puede
actuar como inductor enzimático.
IB15B (Stephanoma) ( ver anexo C-2), requiere para una mejor actividad enzimática,
una concentración mayor de extracto de levadura y una menor concentración de glucosa y

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
8
extracto de levadura, la influencia de la fuente de nitrógeno es muy importante en la
producción de estas enzimas, por otro lado, el extracto de levadura contiene algunas vitaminas
que pueden favorecer a la permeabilidad de la membrana plasmática, esto también puede
influir en el proceso de expresión de enzimas (ver anexo C-2).
Los requerimientos nutricionales son muy distintos para cada una de las cepas fúngicas,
es por eso que el medio de cultivo no se repite.
Algunos hongos productores de LiP no producen enzima en compuestos fenólicos, ya
que acusan inactivación de la actividad de LiP e interfieren en la oxidación de ABTS,
causando inhibición competitiva por el sustrato enzimático, En estas condiciones de cultivo se
torna la necesidad la utilización de otros sustratos para la determinación de actividad LiP,
como por ejemplo veratryl alcohol [83].Talvez la baja actividad enzimática de algunas cepas se
deba a que no se utilizo un sustrato adecuado.
4.2.3. Lacasa.-
Notamos claramente en la tabla 16 que solo cinco cepas fúngicas alcanzaron una
actividad enzimática significativamente buena, ya que las restantes lograron producir enzimas
de actividad lacasa, pero no con éxito, esto puede deber se a diferentes factores los cuales
discutiremos mas adelante.
IB-18Cb (Bdellospora) (ver anexo C-3) a mayor concentración de extracto de levadura
y manganeso, llegaría a una mejor actividad enzimática, podemos notar que la concentración
de glucosa no afecta en la producción de enzima (ver anexo D-3).
La cepa IB-26C (Trinacrium) tuvo una actividad enzimática que fue significativa (ver
anexo C-3), es por esta razón que se la considero entre las cepas fúngicas seleccionadas, para
las pruebas que se realizaran mas adelante.
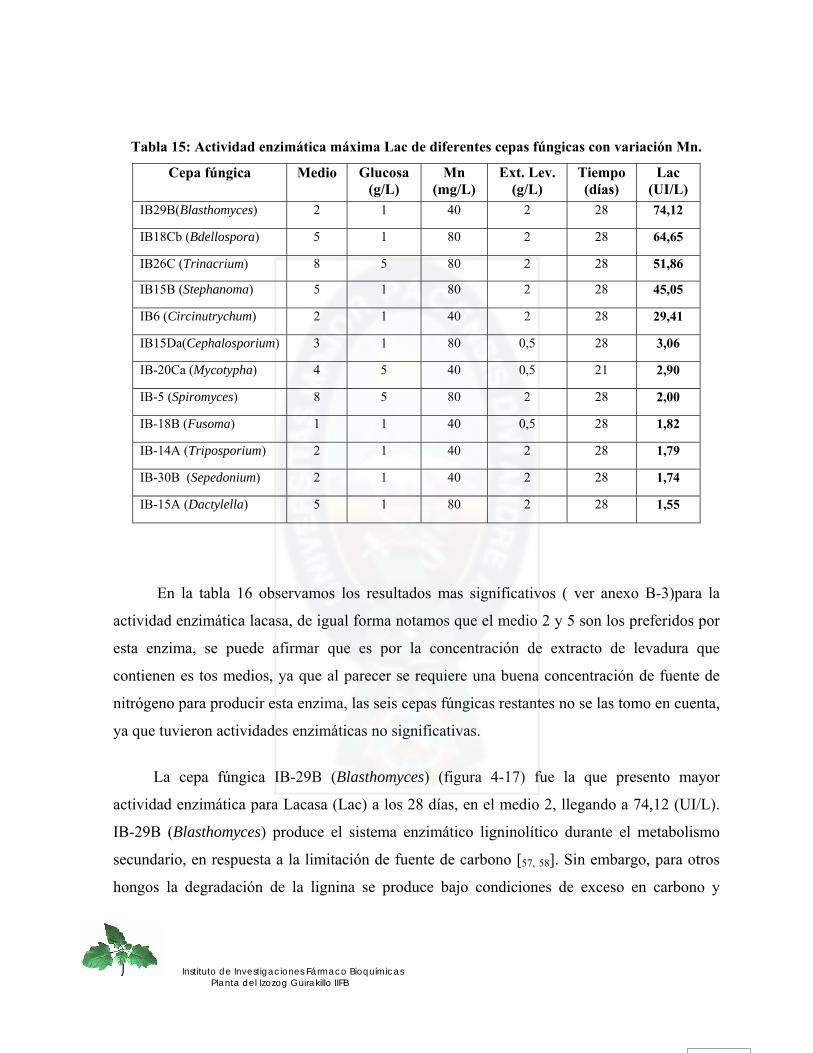
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
82
Tabla 15: Actividad enzimática máxima Lac de diferentes cepas fúngicas con variación Mn. Cepa fúngica Medio Glucosa
(g/L) Mn
(mg/L) Ext. Lev.
(g/L) Tiempo (días)
Lac (UI/L)
IB29B(Blasthomyces) 2 1 40 2 28 74,12
IB18Cb (Bdellospora) 5 1 80 2 28 64,65
IB26C (Trinacrium) 8 5 80 2 28 51,86
IB15B (Stephanoma) 5 1 80 2 28 45,05
IB6 (Circinutrychum) 2 1 40 2 28 29,41
IB15Da(Cephalosporium) 3 1 80 0,5 28 3,06
IB-20Ca (Mycotypha) 4 5 40 0,5 21 2,90
IB-5 (Spiromyces) 8 5 80 2 28 2,00
IB-18B (Fusoma) 1 1 40 0,5 28 1,82
IB-14A (Triposporium) 2 1 40 2 28 1,79
IB-30B (Sepedonium) 2 1 40 2 28 1,74
IB-15A (Dactylella) 5 1 80 2 28 1,55
En la tabla 16 observamos los resultados mas significativos ( ver anexo B-3)para la
actividad enzimática lacasa, de igual forma notamos que el medio 2 y 5 son los preferidos por
esta enzima, se puede afirmar que es por la concentración de extracto de levadura que
contienen es tos medios, ya que al parecer se requiere una buena concentración de fuente de
nitrógeno para producir esta enzima, las seis cepas fúngicas restantes no se las tomo en cuenta,
ya que tuvieron actividades enzimáticas no significativas.
La cepa fúngica IB-29B (Blasthomyces) (figura 4-17) fue la que presento mayor
actividad enzimática para Lacasa (Lac) a los 28 días, en el medio 2, llegando a 74,12 (UI/L). IB-29B (Blasthomyces) produce el sistema enzimático ligninolítico durante el metabolismo
secundario, en respuesta a la limitación de fuente de carbono [57, 58]. Sin embargo, para otros
hongos la degradación de la lignina se produce bajo condiciones de exceso en carbono y
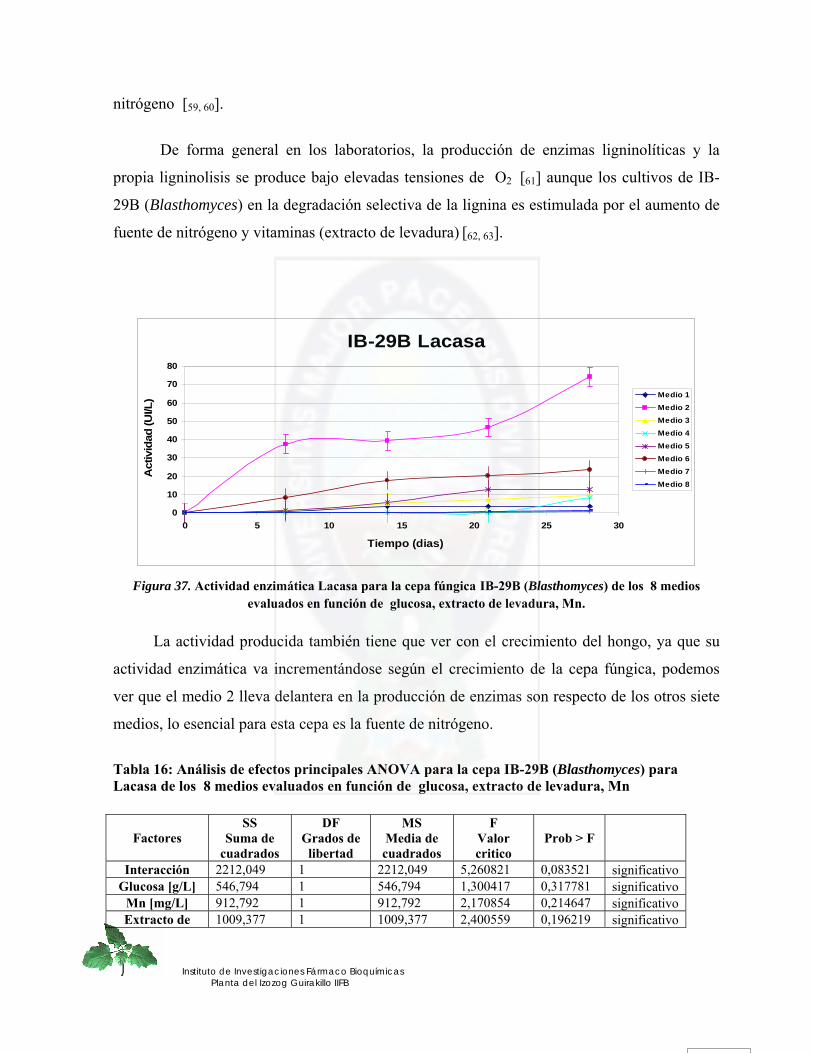
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
83
nitrógeno [59, 60].
De forma general en los laboratorios, la producción de enzimas ligninolíticas y la
propia ligninolisis se produce bajo elevadas tensiones de O2 [61] aunque los cultivos de IB-
29B (Blasthomyces) en la degradación selectiva de la lignina es estimulada por el aumento de
fuente de nitrógeno y vitaminas (extracto de levadura) [62, 63].
IB-29B Lacasa
0
10
20
30
40
50
60
70
80
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L) Medio 1
Medio 2Medio 3Medio 4
Medio 5Medio 6Medio 7
Medio 8
Figura 37. Actividad enzimática Lacasa para la cepa fúngica IB-29B (Blasthomyces) de los 8 medios
evaluados en función de glucosa, extracto de levadura, Mn.
La actividad producida también tiene que ver con el crecimiento del hongo, ya que su
actividad enzimática va incrementándose según el crecimiento de la cepa fúngica, podemos
ver que el medio 2 lleva delantera en la producción de enzimas son respecto de los otros siete
medios, lo esencial para esta cepa es la fuente de nitrógeno.
Tabla 16: Análisis de efectos principales ANOVA para la cepa IB-29B (Blasthomyces) para Lacasa de los 8 medios evaluados en función de glucosa, extracto de levadura, Mn
Factores SS
Suma de cuadrados
DF Grados de
libertad
MS Media de cuadrados
F Valor critico
Prob > F
Interacción 2212,049 1 2212,049 5,260821 0,083521 significativo Glucosa [g/L] 546,794 1 546,794 1,300417 0,317781 significativo
Mn [mg/L] 912,792 1 912,792 2,170854 0,214647 significativo Extracto de 1009,377 1 1009,377 2,400559 0,196219 significativo

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
84
Los resultados del análisis de varianza de la cepa IB-29B (Blasthomyces) que muestra
(tabla 15) la interacción de los tres componentes como ser: glucosa, extracto de levadura y Mn
tiene un valor de F igual a 5,260821 lo que implica que la interacción de estos tres es
significativa. Pro otro lado Prob > F menores a 0,26245 indican que los términos significantes
del modelo.
En esta grafica (Figura 38) podemos comprender que a menor concentración de glucosa
y mayor concentración de extracto de levadura en el medio de cultivo, se llega a obtener una
mayor actividad enzimática para Lacasa.
3D Surface Plot (IB-29B Lacasa Mn 4v*8c)lacasa UI/L = 10,3084-4,1337*x+14,9769*y
40 30 20 10 0
levadura[g/L] Error 1681,904 4 420,476

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
8
Figura 38. Efecto de la glucosa y extracto de levadura sobre la actividad Lacasa para la cepa fúngica IB-
29B (Blasthomyces).
Fuente: Elaboración propia con datos procesados en Statistica 6.0
En esta grafica (Figura 39) podemos comprender que a menor concentración de glucosa
y mayor concentración de Mn en el medio de cultivo, se llega a obtener una mayor actividad
enzimática para Manganeso peroxidasa.
3D Surface Plot (IB-29B Lacasa Mn 4v*8c)Lacasa UI/L = 61,0747-4,1337*x-0,5341*y
40 30 20 10 0
Figura 39. Efecto de la glucosa y Mn sobre la actividad Lacasa para la cepa fúngica IB-29B
(Blasthomyces).
Fuente: Elaboración propia con datos procesados en Statistica 6.0
También se dice que este tipo de enzimas pueden ser reguladas por metales como el
manganeso [35]. La influencia del pH para oxidar el ABTS tanto como el DMF es importante,
este fue conducido en el caso del ABTS por tampón acetato a pH 3 100mM y para DMF por

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
86
tampón succinato pH 5 100 mM, ya que según la bibliografía revisada [81], estos niveles de pH
son óptimos para cada tipo de actividad enzimática que se requiere determinar.
Como las enzimas son proteínas, la variación de pH afecta profundamente al carácter
iónico dos grupos carboxilo y amino de la superficie de la molécula, influenciando bastante en
la naturaleza catalítica. Estas enzimas como ya lo mencionamos tiene un pH optimo
característico exactamente igual al medio en el que se encuentran [84].
Son muchos los factores que afectan la producción de enzimas ligninolíticas en hongos.
Además, las diferentes cepas corresponden de un modo particular a cada un de estos factores.
Debido a esto, el estudio combinado de dos o más factores es acertado para estudiar no solo la
producción de enzimas sino la posible interacción entre estos compuestos.
La entrada de aire u oxigeno tiene gran relevancia debido a que la enzimas
ligninolíticas son activadas a diferentes tensiones de oxigeno en el medio.
Se ha demostrado que la lignina no es una fuente aprovechable para el crecimiento de
los hongos sino que es necesaria la presencia de un co-sustrato [85]. Por lo tanto la adquisición
de la habilidad para mineralizar lignina seria ventajoso par el aprovechamiento de otras
moléculas más fácilmente metabolizables, teniendo en cuenta las relaciones intermoleculares
presentes en la estructura de las paredes celulares.
El patrón de actividad de estas enzimas es específico del género y especie de hongo de
la PB involucrado, algunos excretan la LiP, la MnP y no la Lac ó la MnP, la Lac y no la LiP;
La síntesis fúngica de la LiP y la MnP se realiza bajo una alta tensión de oxígeno, se reprime
en agitación o cuando los hongos de la PB, se cultivan en medio líquido, sin embargo la Lac se
sintetiza sólo en agitación, estas tres enzimas son inespecíficas y se inducen en condición
limitante de nutrientes, como la fuente de nitrógeno [39].
Según los resultados obtenidos, podemos comprobar las referencias de que una misma
cepa fúngica es capaz de producir las tres enzimas ligninolíticas, ya que en este caso, cuatro
cepas son capaces de producir al mismo tiempo MnP, LiP, Lac, unas con mas actividad

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
8
enzimática en LiP y otras con mayor actividad enzimática en Lac.
Las cepas que fueron capaces de producir los tres tipos de enzimas fueron: IB-29B, IB-
26C, IB-15B, IB-18Cb, (ver anexos C-1, C-2, C-3) según la caracterización realizada se cree
que son hongos basidiomicetos, pero esto aun esta por confirmarse, en posteriores pruebas
moleculares.
Es interesante notar que las condiciones óptimas para la producción de estas enzimas
son las mismas, este hecho es una ventaja para procesos de biorremediación dado que en un
mismo medio se produce la máxima cantidad de enzima y fue demostrado que en estas actúan
sinergicamente en procesos de degradación [78].
De las doce cepas que se seleccionaron cualitativamente, solo seis mostraron una buena
actividad enzimática, esto se debe a que muy pocas son las cepas fúngicas con actividad
ligninolítica que están distribuidas ampliamente en el ecosistema.
4.3. OPTIMIZACION DE MEDIOS DE CULTIVO PARA LA PRDUCCION
DE ENZIMAS LIGNINOLITICAS.-
Con el objetivo de llegar a obtener una mayor actividad enzimática fue que se realizo
una segunda modificación de medio de cultivo para producción de enzimas ligninolíticas. Esta
prueba se llevo a cabo con seis cepas seleccionadas: IB-29B (Blasthomyces), IB-18Cb
(Bdellospora), IB-26C (Trinacrium), IB-15B (Stephanoma), IB-15Da (Cephalosporium), IB-6
(Circinutrychum). Esto por los resultados obtenidos en la anterior prueba
4.3.1 Medios de cultivo con variación de concentración de Cobre.-
Se realizo nuevamente un diseño factorial diseño factorial 23, siendo los factores esta
vez: glucosa, cobre, extracto de levadura. Esta prueba se debe a que según bibliografía
revisada, el cobre es un buen inductor de este tipo de enzimas, especialmente lacasas, ya que
estas llevan en su estructura cobre.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
88
a. Manganeso Peroxidasa.-
Se observa que la máxima producción de enzima, se da donde la concentración de
cobre es más baja, Si bien es un hecho común que la MnP se induzca con otro metal que no
sea Mn ya se ha mostrado que en S. hirsutum el Cd induce la síntesis de esta enzima. [77].
También se puede afirmar que la producción de MnP son incrementadas con el agregado de
Cu al medio, tal no es el caso para IB-29B (Blasthomyces) [76].
En la tabla 18 observamos que en las tres cepas con mayor actividad enzimática
(ver anexo C-4), la concentración de cobre es mínima, lo cual confirma, que para la
producción de esta enzima, es limitada la adición de cobre en el medio de cultivo. No
podemos afirmar que esto sea cierto para todas las cepas fúngicas, ya que cada una tiene sus
propios requerimientos.
Tabla 17: Actividad enzimática MnP de diferentes cepas fúngicas con variación Cu. Cepa fúngica Medio Glucosa
(g/L) Cu
(mg/L) Ext. Lev.
(g/L) Tiempo (días)
MnP (UI/L)
IB29B(Blasthomyces) 6 5 40 2 21 245,56
IB15Da(Cephalosporium) 4 5 40 0,5 21 122,84
IB15B (Stephanoma) 1 1 40 0,5 28 121,64
IB18Cb (Bdellospora) 5 1 80 2 28 91,31
IB-26C (Trinacrium) 7 5 80 0,5 28 82,68
IB-6 (Circinutrychum) 5 1 80 2 21 61,03
En IB-29B (Blasthomyces) la fuente de carbono y nitrógeno es muy importante ya que
en el medio el cual se presento la mayor actividad enzimática, se tiene la concentración de
estos alta, de esta forma podemos decir que es muy importante la participación de nitrógeno y
glucosa en el metabolismo de lis hongos para la producción de esta enzima.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
89
IB-29B MnP
0
50
100
150
200
250
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L)
Medio 1Medio 2Medio 3Medio 4Medio 5Medio 6Medio 7Medio 8
Figura 40. Actividad enzimática Manganeso peroxidasa para la cepa fúngica IB-29B (Blasthomyces) de 8
medios evaluados en función de glucosa, extracto de levadura, Cu.
Tabla 18: Análisis de efectos principales ANOVA para la cepa IB-29B (Blasthomyces) para MnP, de 8 medios evaluados en función de glucosa, extracto de levadura, Cu.
Fuente: Elaboración propia con dato
s procesados en Statistica 6.0
Los resultados del análisis de varianza de la cepa IB-26C (Trinacrium) que muestra
(tabla 17) la interacción de los tres componentes como ser: glucosa, extracto de
Factores SS
Suma de cuadrados
DF Grados de
libertad
MS Media de cuadrados
F Valor critico
Prob > F
Interacción 54992,36 1 54992,36 5,832676 0,073150 significativo Glucosa [g/L] 791,59 1 791,59 0,083958 0,786402 significativo
Cu [mg/L] 95,82 1 95,82 0,010163 0,924550 no significativo
Extracto de levadura[g/L] 12859,93 1 12859,93 1,363968 0,307715 significativo
Error 37713,30 4 9428,32
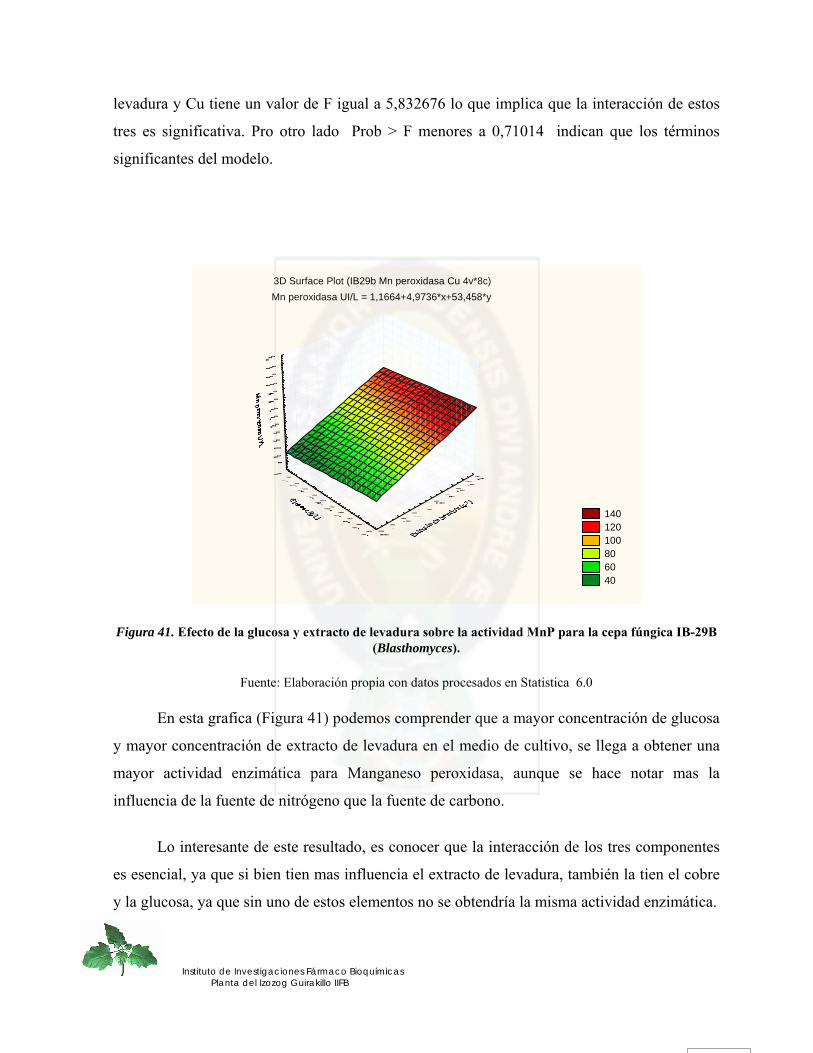
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
90
levadura y Cu tiene un valor de F igual a 5,832676 lo que implica que la interacción de estos
tres es significativa. Pro otro lado Prob > F menores a 0,71014 indican que los términos
significantes del modelo.
3D Surface Plot (IB29b Mn peroxidasa Cu 4v*8c)Mn peroxidasa UI/L = 1,1664+4,9736*x+53,458*y
140 120 100 80 60 40
Figura 41. Efecto de la glucosa y extracto de levadura sobre la actividad MnP para la cepa fúngica IB-29B (Blasthomyces).
Fuente: Elaboración propia con datos procesados en Statistica 6.0
En esta grafica (Figura 41) podemos comprender que a mayor concentración de glucosa
y mayor concentración de extracto de levadura en el medio de cultivo, se llega a obtener una
mayor actividad enzimática para Manganeso peroxidasa, aunque se hace notar mas la
influencia de la fuente de nitrógeno que la fuente de carbono.
Lo interesante de este resultado, es conocer que la interacción de los tres componentes
es esencial, ya que si bien tien mas influencia el extracto de levadura, también la tien el cobre
y la glucosa, ya que sin uno de estos elementos no se obtendría la misma actividad enzimática.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
9
3D Surface Plot (IB29b Mn peroxidasa Cu 4v*8c)Mn peroxidasa UI/L = 78,3716+4,9736*x-0,173*y
80
Figura 42. Efecto de la glucosa y Cu sobre la actividad MnP para la cepa fúngica IB-29B (Blasthomyces).
Fuente: Elaboración propia con datos procesados en Statistica 6.0
En esta grafica (Figura 42) podemos comprender que la concentración de Cu en el
medio de cultivo, no influye en la actividad enzimática para Manganeso peroxidasa. Sin
embargo si comparamos los resultados obtenidos para MnP con la influencia de manganeso,
notaremos claramente que los resultados mejoraron notablemente con la intervención del
cobre, pro lo cual no podemos afirmar que este metal no influya en los resultados obtenidos.
En la cepa fúngica IB-15Da (Cephalosporium) (ver anexo C-4) no influye la
concentración de glucosa, pero en cambio la concentración de manganeso y extracto de
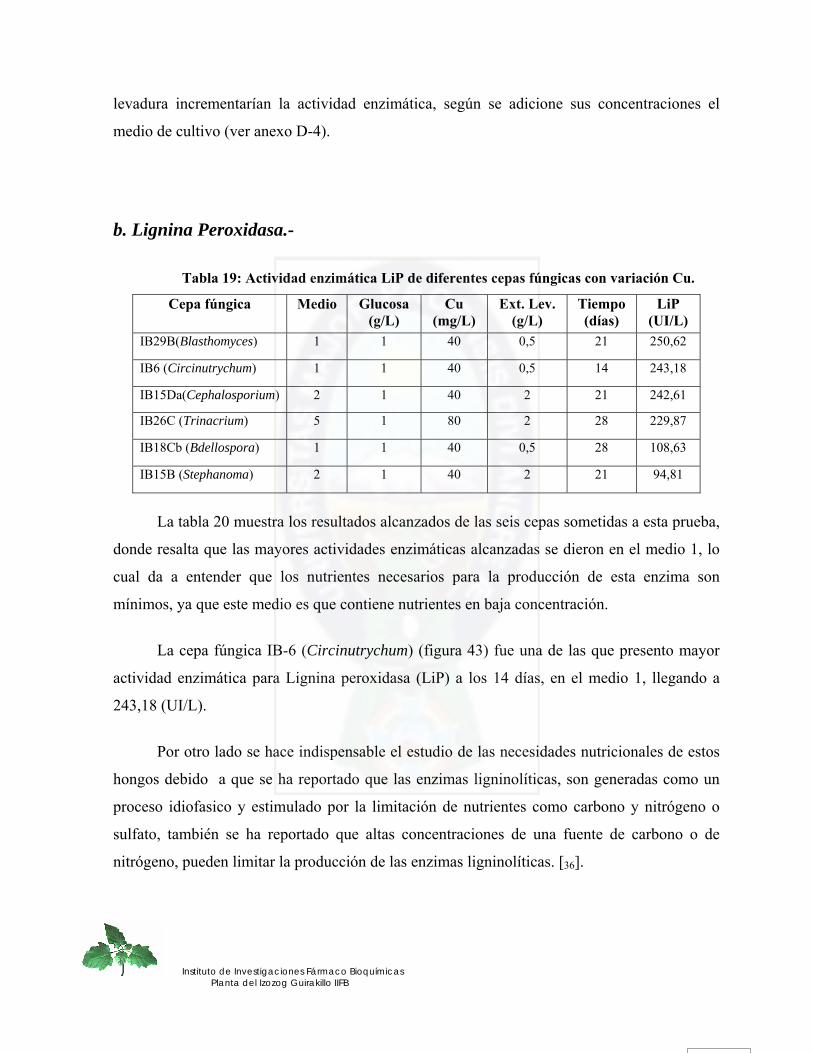
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
92
levadura incrementarían la actividad enzimática, según se adicione sus concentraciones el
medio de cultivo (ver anexo D-4).
b. Lignina Peroxidasa.-
Tabla 19: Actividad enzimática LiP de diferentes cepas fúngicas con variación Cu.
Cepa fúngica Medio Glucosa (g/L)
Cu (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
LiP (UI/L)
IB29B(Blasthomyces) 1 1 40 0,5 21 250,62
IB6 (Circinutrychum) 1 1 40 0,5 14 243,18
IB15Da(Cephalosporium) 2 1 40 2 21 242,61
IB26C (Trinacrium) 5 1 80 2 28 229,87
IB18Cb (Bdellospora) 1 1 40 0,5 28 108,63
IB15B (Stephanoma) 2 1 40 2 21 94,81
La tabla 20 muestra los resultados alcanzados de las seis cepas sometidas a esta prueba,
donde resalta que las mayores actividades enzimáticas alcanzadas se dieron en el medio 1, lo
cual da a entender que los nutrientes necesarios para la producción de esta enzima son
mínimos, ya que este medio es que contiene nutrientes en baja concentración.
La cepa fúngica IB-6 (Circinutrychum) (figura 43) fue una de las que presento mayor
actividad enzimática para Lignina peroxidasa (LiP) a los 14 días, en el medio 1, llegando a
243,18 (UI/L).
Por otro lado se hace indispensable el estudio de las necesidades nutricionales de estos
hongos debido a que se ha reportado que las enzimas ligninolíticas, son generadas como un
proceso idiofasico y estimulado por la limitación de nutrientes como carbono y nitrógeno o
sulfato, también se ha reportado que altas concentraciones de una fuente de carbono o de
nitrógeno, pueden limitar la producción de las enzimas ligninolíticas. [36].
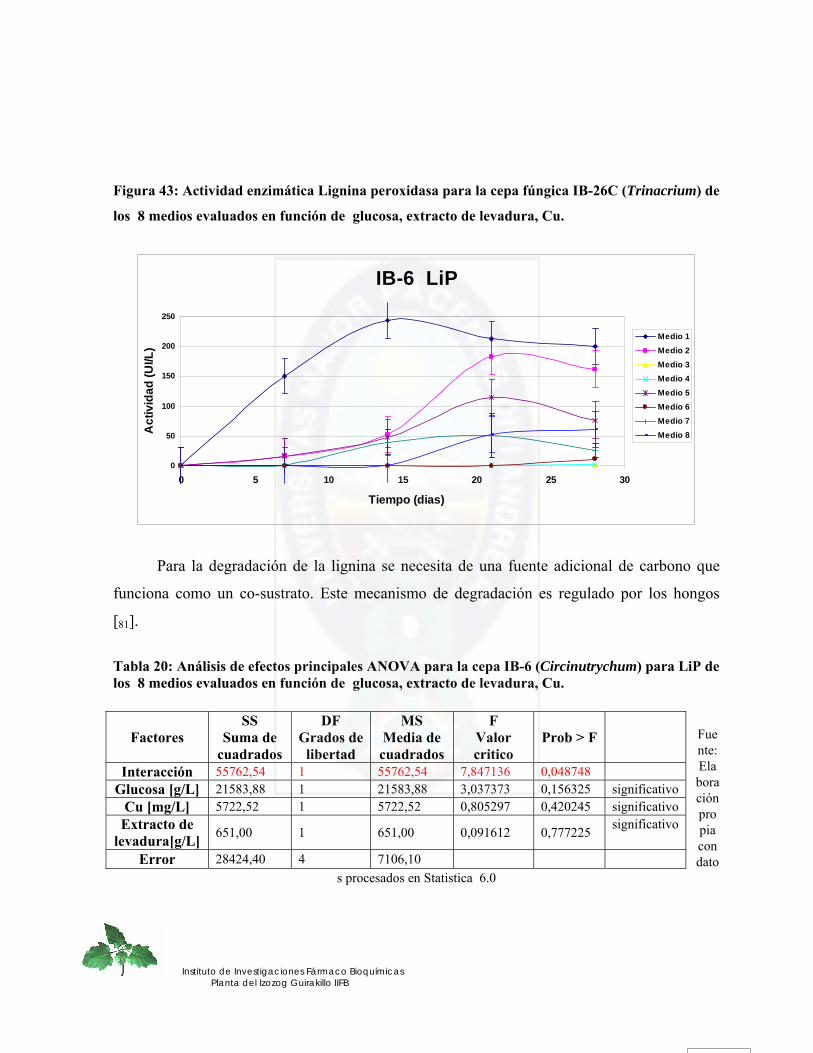
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
93
Figura 43: Actividad enzimática Lignina peroxidasa para la cepa fúngica IB-26C (Trinacrium) de
los 8 medios evaluados en función de glucosa, extracto de levadura, Cu.
IB-6 LiP
0
50
100
150
200
250
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L)
Medio 1Medio 2Medio 3Medio 4Medio 5Medio 6Medio 7Medio 8
Para la degradación de la lignina se necesita de una fuente adicional de carbono que
funciona como un co-sustrato. Este mecanismo de degradación es regulado por los hongos
[81].
Tabla 20: Análisis de efectos principales ANOVA para la cepa IB-6 (Circinutrychum) para LiP de los 8 medios evaluados en función de glucosa, extracto de levadura, Cu.
Fuente: Elaboración propia con dato
s procesados en Statistica 6.0
Factores SS
Suma de cuadrados
DF Grados de
libertad
MS Media de cuadrados
F Valor critico
Prob > F
Interacción 55762,54 1 55762,54 7,847136 0,048748 Glucosa [g/L] 21583,88 1 21583,88 3,037373 0,156325 significativo
Cu [mg/L] 5722,52 1 5722,52 0,805297 0,420245 significativoExtracto de
levadura[g/L] 651,00 1 651,00 0,091612 0,777225 significativo
Error 28424,40 4 7106,10

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
94
Los resultados del análisis de varianza de la IB-6 (Circinutrychum) que muestra (tabla
19) la interacción de los tres componentes como ser: glucosa, extracto de levadura y Cu tiene
un valor de F igual a 7,847136 lo que implica que la interacción de estos tres es significativa.
Pro otro lado Prob > F menores a 0,38678 indican que los términos significantes del modelo.
Figura 44: Efecto de la glucosa y extracto de levadura sobre la actividad LiP para la cepa fúngica IB-6 (Circinutrychum)
3D Surface Plot (IB-6Li peroxidasa Cu 4v*8c)Li peroxidasa UI/L = 146,3669-25,9711*x+12,0278*y
140 120 100 80 60 40 20
Fuente: Elaboración propia con datos procesados en Statistica 6.0
En esta grafica (Figura 44) podemos comprender que a mayor concentración de glucosa
y menor concentración de extracto de levadura en el medio de cultivo, se llega a obtener una
mayor actividad enzimática para Lignina peroxidasa, sin embargo en el medio 5 el cual tien la
misma concentración de extracto de levadura y cobre, y la concentración de glucosa aumenta,
no se obtuvo una actividad enzimática buena, esto puede deberse a que el microorganismo
requiere concentraciones nutricionales mínimas para llegar a su actividad máxima.
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
9
Figura 45: Efecto de la glucosa y Cu sobre la actividad LiP para la cepa fúngica IB-6
(Circinutrychum).
3D Surface Plot (IB-6 Li peroxidasa Cu 4v*8c)Li peroxidasa UI/L = 241,6377-25,9711*x-1,3373*y
180 140 100 60 20
Fuente: Elaboración propia con datos procesados en Statistica 6.0
En esta grafica (Figura 45) podemos comprender que a mayor concentración de
glucosa y menor concentración de Cu en el medio de cultivo, se llega a obtener una mayor
actividad enzimática para Lignina peroxidasa.
IB6 (Circinutrychum) (ver anexo C-5) a mayor concentración de glucosa, y menor
concentración de extracto de levadura obtendría un actividad enzimática máxima, también se
puede denotar que la concentración de cobre no afecta en la producción de enzima (ver anexo
D-5).
c. Lacasa.-
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
96
De acuerdo a los resultados obtenidos en la tabla 22 la producción de esta enzima con
la variabilidad de cobre requiere una concentración mínima de glucosa, y la concentración de
extracto de levadura es muy importante, ya que es muy necesaria la fuente de nitrógeno para
estas enzimas, si comparamos los resultados alcanzados en esta prueba respecto a la
determinación de lacasa con variación de manganeso, notaremos claramente que los resultados
tiene mucha diferencia (ver anexo C-6).
Tabla 21: Actividad enzimática Lac de diferentes cepas fúngicas con variación Cu.
Cepa fúngica Medio Glucosa (g/L)
Cu (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
Lac (UI/L)
IB15Da(Cephalosporium) 1 1 40 0,5 28 287,30
IB29B(Blasthomyces) 1 1 40 0,5 21 265,25
IB26C (Trinacrium) 5 1 80 2 21 220
IB6 (Circinutrychum) 2 1 40 2 28 227.11
IB15B (Stephanoma) 2 1 40 2 21 208,92
IB-18Cb (Bdellospora) 1 1 40 0,5 28 138,49
La cepa fúngica IB-15Da (Cephalosporium) (figura 46) alcanzo la mayor actividad
enzimática para Lacasa (Lac) a los 28 días, en el medio 1, llegando a 287,30 (UI/L).
Figura 46: Actividad enzimática Lacasa para la cepa fúngica IB-15Da (Cephalosporium) de los 8
medios evaluados en función de glucosa, extracto de levadura, Cu.
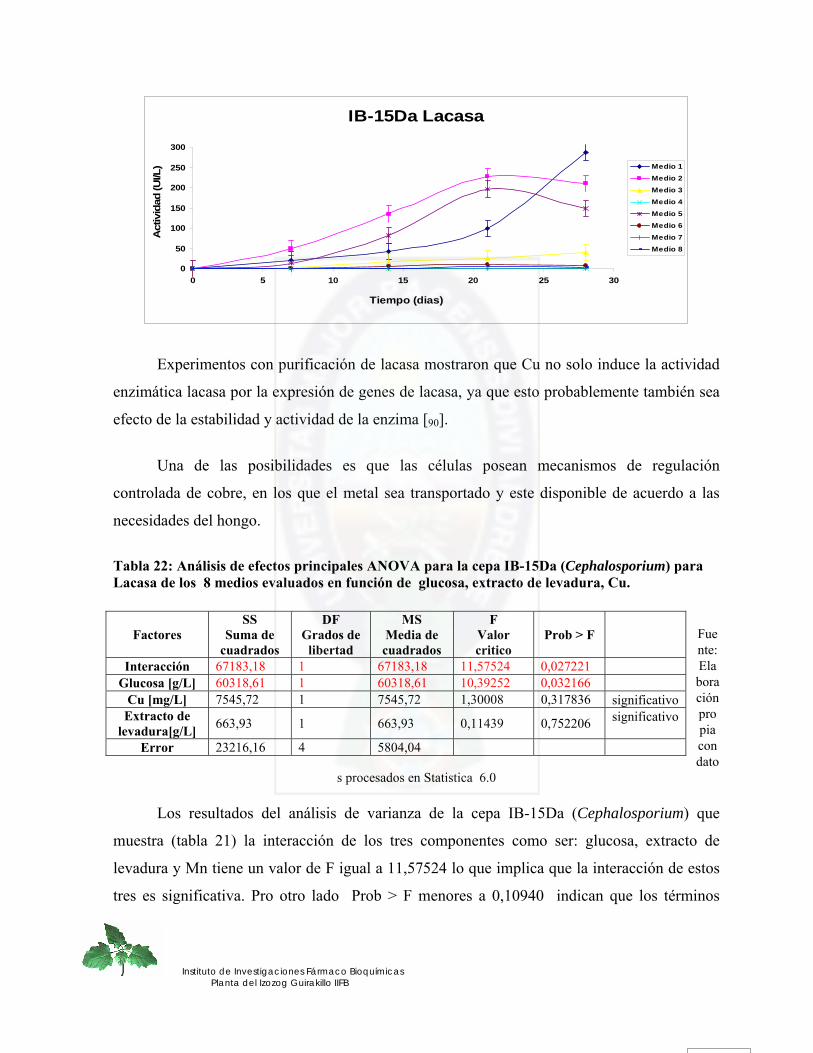
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
9
IB-15Da Lacasa
0
50
100
150
200
250
300
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L) Medio 1
Medio 2Medio 3Medio 4
Medio 5Medio 6Medio 7Medio 8
Experimentos con purificación de lacasa mostraron que Cu no solo induce la actividad
enzimática lacasa por la expresión de genes de lacasa, ya que esto probablemente también sea
efecto de la estabilidad y actividad de la enzima [90].
Una de las posibilidades es que las células posean mecanismos de regulación
controlada de cobre, en los que el metal sea transportado y este disponible de acuerdo a las
necesidades del hongo.
Tabla 22: Análisis de efectos principales ANOVA para la cepa IB-15Da (Cephalosporium) para Lacasa de los 8 medios evaluados en función de glucosa, extracto de levadura, Cu.
Fuente: Elaboración propia con dato
s procesados en Statistica 6.0
Los resultados del análisis de varianza de la cepa IB-15Da (Cephalosporium) que
muestra (tabla 21) la interacción de los tres componentes como ser: glucosa, extracto de
levadura y Mn tiene un valor de F igual a 11,57524 lo que implica que la interacción de estos
tres es significativa. Pro otro lado Prob > F menores a 0,10940 indican que los términos
Factores SS
Suma de cuadrados
DF Grados de
libertad
MS Media de cuadrados
F Valor critico
Prob > F
Interacción 67183,18 1 67183,18 11,57524 0,027221 Glucosa [g/L] 60318,61 1 60318,61 10,39252 0,032166
Cu [mg/L] 7545,72 1 7545,72 1,30008 0,317836 significativo Extracto de
levadura[g/L] 663,93 1 663,93 0,11439 0,752206 significativo
Error 23216,16 4 5804,04

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
98
significantes del modelo.
Figura 47: Efecto de la glucosa y extracto de levadura sobre la actividad Lacasa para la cepa
fúngica IB-15Da (Cephalosporium).
3D Surface Plot (IB-15Da LACASA Cu 4v*8c)Lacasa UI/L = 206,7051-43,4161*x+12,1466*y
200 150 100 50 0
En esta grafica (Figura 47) podemos comprender que a mayor concentración de glucosa
y menor concentración de extracto de levadura en el medio de cultivo, se llega a obtener una
mayor actividad enzimática para Lacasa.
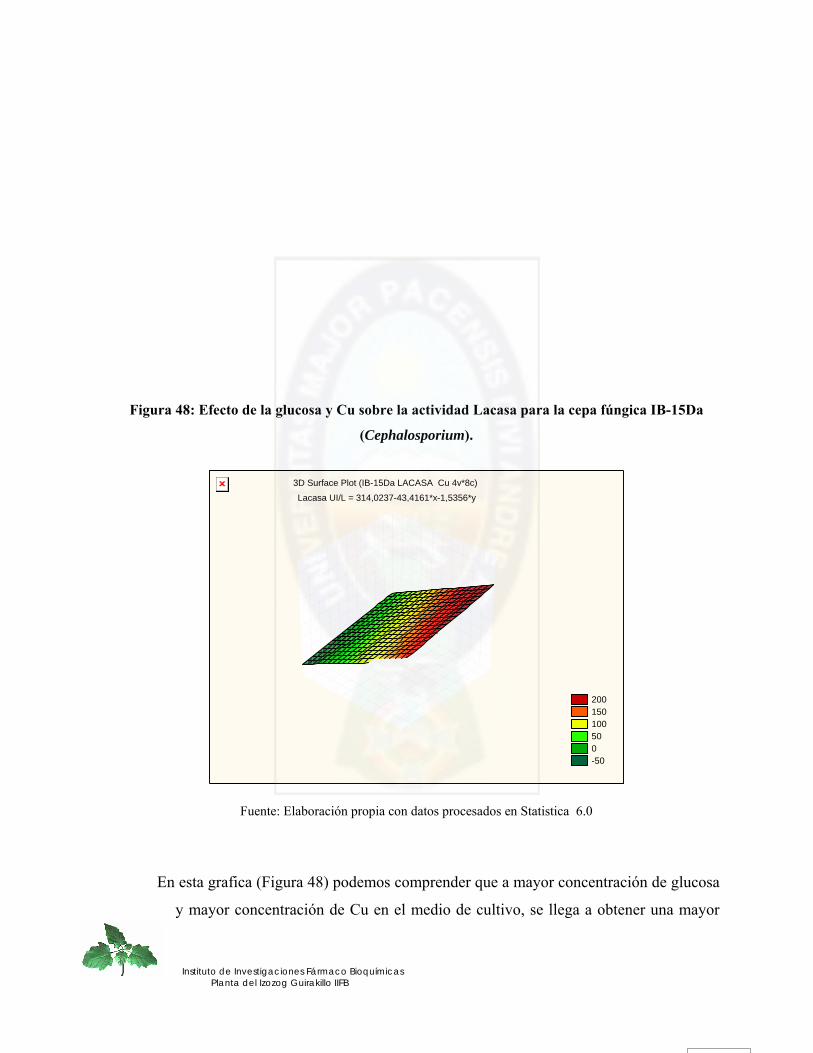
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
99
Figura 48: Efecto de la glucosa y Cu sobre la actividad Lacasa para la cepa fúngica IB-15Da
(Cephalosporium).
3D Surface Plot (IB-15Da LACASA Cu 4v*8c)Lacasa UI/L = 314,0237-43,4161*x-1,5356*y
200 150 100 50 0 -50
Fuente: Elaboración propia con datos procesados en Statistica 6.0
En esta grafica (Figura 48) podemos comprender que a mayor concentración de glucosa
y mayor concentración de Cu en el medio de cultivo, se llega a obtener una mayor

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
00
actividad enzimática para Lacasa. Sin embargo cuando la concentración de glucosa puede
aumentarse esto puede resultar inhibidor de la producción de enzimas que inductor.
Talvez esto pueda deberse a que la lacasa no parece estar transcripcionalmente por el
cobre, puesto que la suplementacion de cobre disminuye la cantidad de transcrito, en
condiciones de exceso de cobre, el aumento observado en cultivos se debe a ala sobre-
expresión de los genes acumulando mRNA [32]. El aumento de la actividad lacasa que se
produce al añadir cobre puede deberse a la incorporación de este a la apoproteína acumulada:
por lo tanto, la actividad lacasa dependería de la disponibilidad de cobre y de la presencia de
apoproteína inactiva.
La IB-29B (Blasthomyces) no tuvo mucha diferencia en su actividad enzimática
respecto a IB-15Da (Cephalosporium) (ver anexo C-6) sin embargo sus requerimientos
nutricionales son muy diferentes ya que esta cepa requiere mayor concentración de fuente de
nitrógeno y vitaminas (ver anexo D-6).
Los resultados obtenidos en la presente prueba son mucho más significativos que los
resultados donde utilizo manganeso, esto nos conduce a afirmar, que el cobre es un mejor
inductor para las cepas fúngicas con las cuales estamos trabajando.
En especial para la actividad enzimática lacasa, se obtuvo resultados óptimos,
probablemente se deba a que esta enzima en su estructura lleve cobre, y este metal sea un
excelente inductor para esta enzima, por esa razón.
Los resultados mas bajos de las tres actividades enzimáticas fueron para manganeso
peroxidas, aun que esto no fue con mucha diferencia significativa, sin embargo con medio de
cultivo Cu, las enzimas llegaron a producir mas de tres veces mas que con el medio de cultivo
Mn.
4.3.2. Medios de cultivo con diferentes sustratos lignocelulosicos.-
Estas enzimas son en términos biotecnológicos, uno de los grupos de enzimas que mas
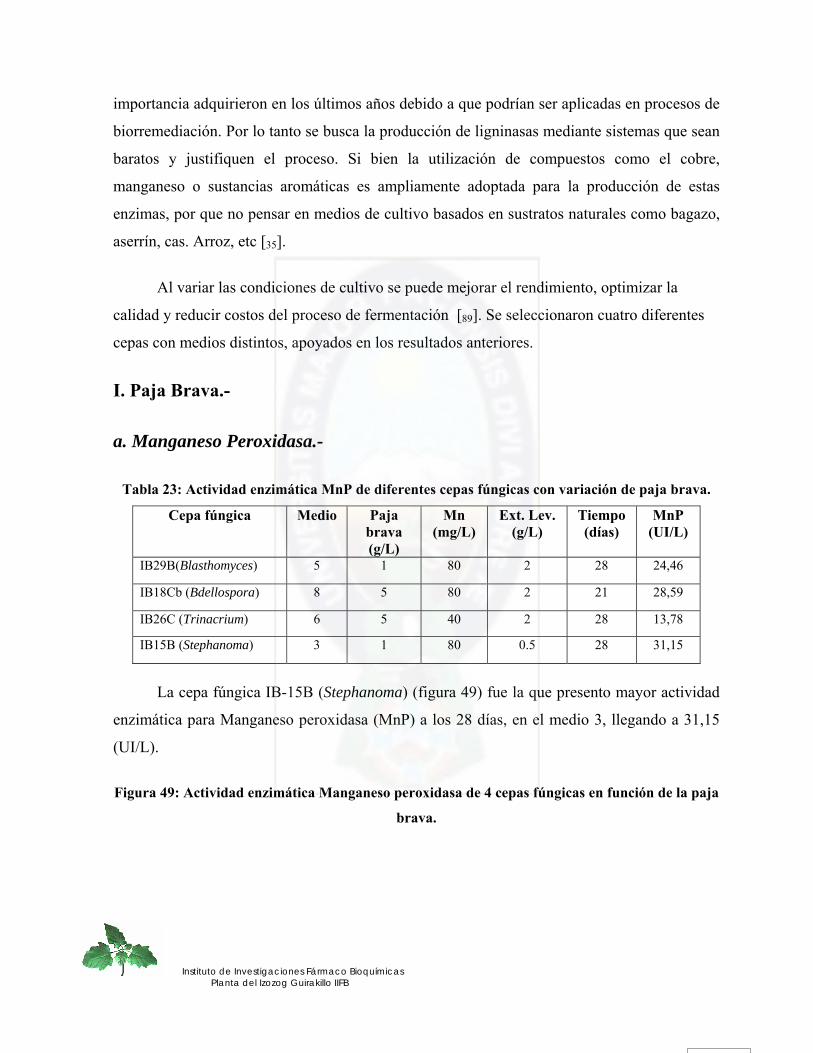
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
0
importancia adquirieron en los últimos años debido a que podrían ser aplicadas en procesos de
biorremediación. Por lo tanto se busca la producción de ligninasas mediante sistemas que sean
baratos y justifiquen el proceso. Si bien la utilización de compuestos como el cobre,
manganeso o sustancias aromáticas es ampliamente adoptada para la producción de estas
enzimas, por que no pensar en medios de cultivo basados en sustratos naturales como bagazo,
aserrín, cas. Arroz, etc [35].
Al variar las condiciones de cultivo se puede mejorar el rendimiento, optimizar la
calidad y reducir costos del proceso de fermentación [89]. Se seleccionaron cuatro diferentes
cepas con medios distintos, apoyados en los resultados anteriores.
I. Paja Brava.-
a. Manganeso Peroxidasa.-
Tabla 23: Actividad enzimática MnP de diferentes cepas fúngicas con variación de paja brava. Cepa fúngica Medio Paja
brava (g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
MnP (UI/L)
IB29B(Blasthomyces) 5 1 80 2 28 24,46
IB18Cb (Bdellospora) 8 5 80 2 21 28,59
IB26C (Trinacrium) 6 5 40 2 28 13,78
IB15B (Stephanoma) 3 1 80 0.5 28 31,15
La cepa fúngica IB-15B (Stephanoma) (figura 49) fue la que presento mayor actividad
enzimática para Manganeso peroxidasa (MnP) a los 28 días, en el medio 3, llegando a 31,15
(UI/L).
Figura 49: Actividad enzimática Manganeso peroxidasa de 4 cepas fúngicas en función de la paja
brava.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
02
MnP (paja brava)
0
10
20
30
40
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad U
I/L
IB-29B IB-18Cb IB-26C IB-15B
Con respecto a los anteriores resultados alcanzados en la pruebas que se adiciono en el
medio glucosa, se alcanzaron valores más altos de actividad enzimática, si bien explicamos
anteriormente que la fuente de carbono es muy importante para la producción de estas
enzimas.
La paja brava en su composición química contiene un 54% de celulosa lo cual es fuente
de carbono, en este caso esa fuente de carbono no esta libre, esta protegida por lignina, lo que
se quizo comprobar es la efectividad de este tipo de enzimas, con este sustrato lignocelulósico.
b. Lignina Peroxidasa.-
Tabla 24: Actividad enzimática LiP de diferentes cepas fúngicas con variación de paja brava.
La
cepa
fúngi
ca
IB-
18Cb (Bdellospora) fue una de las que presento mayor actividad enzimática para Lignina
Cepa fúngica Medio Paja brava (g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
LiP (UI/L)
IB29B(Blasthomyces) 3 1 80 0.5 21 16,19
IB18Cb (Bdellospora) 8 5 80 2 21 133,97
IB-26C (Trinacrium) 8 5 80 2 21 13,84
IB15B (Stephanoma) 8 5 80 2 21 28,54
IB15Da(Cephalosporium) 3 1 80 0.5 21 21,03

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
03
peroxidasa (LiP) a los 21 días, en el medio 8, llegando a 133,97 (UI/L).
Figura 50: Actividad enzimática Lignina peroxidasa de 5 cepas fúngicas en función de la paja
brava.
LiP (paja brava)
0
35
70
105
140
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad U
I/L
IB-29B IB-18Cb IB-26C IB-15B IB-15Da
El resultado alcanzado por la cepa IB-18Cb fue el mas significativo de las 5 cepas,
incluso fue mayor que el alcanzado en la prueba con variación de manganeso. El medio en que
se desarrollo esta actividad enzimática para LiP es aquel en el cual lleva en mayor
concentración sus componentes.
Es posible que esta cepa requiera mayores cantidades de nutrientes para alcanzar una
actividad máxima, también no se deja de lado la hipótesis de que esta actividad se deba a un
producto del metabolismo del microorganismo.
c. Lacasa.-
Tabla 24: Actividad enzimática Lac de diferentes cepas fúngicas con variación de paja
brava.
La
cepa
Cepa fúngica Medio Paja brava (g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
Lac (UI/L)
IB29B(Blasthomyces) 2 1 40 2 21 24,76
IB18Cb (Bdellospora) 3 1 80 0.5 28 37,61
IB-26C (Trinacrium) 8 5 80 2 28 9,52
IB15B (Stephanoma) 5 1 80 2 21 5,20
IB-6 (Circinutrychum) 2 1 40 2 28 9,76
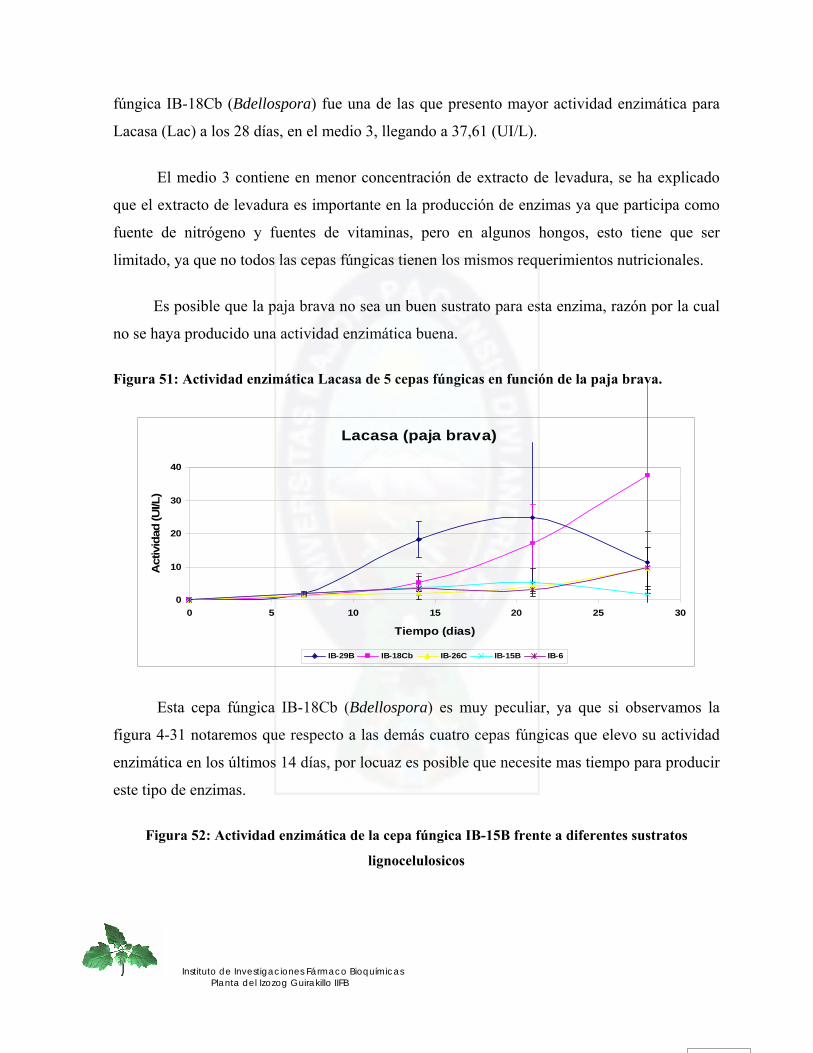
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
04
fúngica IB-18Cb (Bdellospora) fue una de las que presento mayor actividad enzimática para
Lacasa (Lac) a los 28 días, en el medio 3, llegando a 37,61 (UI/L).
El medio 3 contiene en menor concentración de extracto de levadura, se ha explicado
que el extracto de levadura es importante en la producción de enzimas ya que participa como
fuente de nitrógeno y fuentes de vitaminas, pero en algunos hongos, esto tiene que ser
limitado, ya que no todos las cepas fúngicas tienen los mismos requerimientos nutricionales.
Es posible que la paja brava no sea un buen sustrato para esta enzima, razón por la cual
no se haya producido una actividad enzimática buena.
Figura 51: Actividad enzimática Lacasa de 5 cepas fúngicas en función de la paja brava.
Lacasa (paja brava)
0
10
20
30
40
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L)
IB-29B IB-18Cb IB-26C IB-15B IB-6
Esta cepa fúngica IB-18Cb (Bdellospora) es muy peculiar, ya que si observamos la
figura 4-31 notaremos que respecto a las demás cuatro cepas fúngicas que elevo su actividad
enzimática en los últimos 14 días, por locuaz es posible que necesite mas tiempo para producir
este tipo de enzimas.
Figura 52: Actividad enzimática de la cepa fúngica IB-15B frente a diferentes sustratos
lignocelulosicos

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
0
IB-15B
01020304050607080
Paja Brava Bagazo Aserrin ResiduosQuinua
Cascara dearroz
Sustratos
Act
ivid
ad (U
I/L)
MnP UI/L LiP UI/L Lac UI/L
De las seis cepas fúngicas escogidas la que mejor resultado presento para la paja brava
fue la IB-18Cb como se observa en la figura 52, con respecto a otras cepas, fue la cepa que
mejor resultado alcanzo para actividad MnP, sin embargo podemos notar que el bagazo y la
cáscara de arroz presentan una mayor actividad enzimática para LiP, esto depende de los
requerimientos nutricionales de cada hongo , para la expresión de una determinada enzima.
II. Bagazo.-
a. Manganeso Peroxidasa.-
Tabla 25: Actividad enzimática MnP de diferentes cepas fúngicas con variación de bagazo
La
cepa
fúngi
ca
IB-29B (Blasthomyces) fue la que presento mayor actividad enzimática para Manganeso
peroxidasa (MnP) a los 28 días, en el medio 5, llegando a 91,40 (UI/L).
Cepa fúngica Medio Bagazo (g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
MnP (UI/L)
IB29B(Blasthomyces) 5 1 80 2 28 91,40
IB18Cb (Bdellospora) 8 5 80 2 28 11,04
IB26C (Trinacrium) 6 5 40 2 21 16,28
IB15B (Stephanoma) 3 1 80 0.5 21 16,28
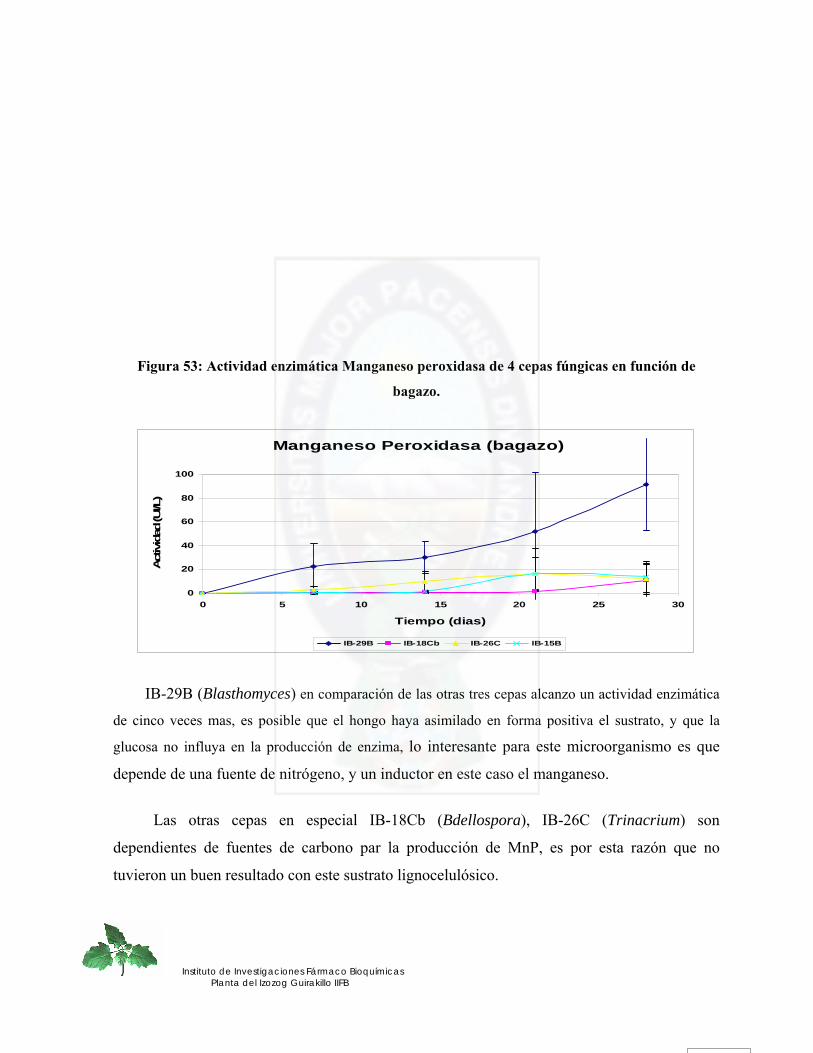
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
06
Figura 53: Actividad enzimática Manganeso peroxidasa de 4 cepas fúngicas en función de
bagazo.
Manganeso Peroxidasa (bagazo)
0
20
40
60
80
100
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L)
IB-29B IB-18Cb IB-26C IB-15B
IB-29B (Blasthomyces) en comparación de las otras tres cepas alcanzo un actividad enzimática
de cinco veces mas, es posible que el hongo haya asimilado en forma positiva el sustrato, y que la
glucosa no influya en la producción de enzima, lo interesante para este microorganismo es que
depende de una fuente de nitrógeno, y un inductor en este caso el manganeso.
Las otras cepas en especial IB-18Cb (Bdellospora), IB-26C (Trinacrium) son
dependientes de fuentes de carbono par la producción de MnP, es por esta razón que no
tuvieron un buen resultado con este sustrato lignocelulósico.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
0
b. Lignina Peroxidasa.-
Tabla 26: Actividad enzimática LiP de diferentes cepas fúngicas con variación de bagazo
La cepa fúngica IB-18Cb (Bdellospora) fue la que presento mayor actividad enzimática
para Lignina peroxidasa (LiP) a los 21 días, en el medio 8, llegando a 487,82 (UI/L).
Según los análisis para esta cepa fúngica, tendría mayor actividad enzimática con la
adición de glucosa, sin embargo el resultado que se obtuvo en esta prueba fue inesperado ya
que alcanzo cinco veces mas actividad enzimática que los resultados obtenidos con glucosa,
esto nos da a entender que la cepa asimilo la fuente de carbono que contiene el bagazo que es
49,58% en celulosa y este estimulo la actividad enzimática de este microorganismo.
Figura 54: Actividad enzimática Lignina peroxidasa de 5 cepas fúngicas en función de la bagazo.
Lignina Peroxidasa (bagazo)
0
100
200
300
400
500
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L)
IB-29B IB-18Cb IB-26C IB-15B IB-15Da
Cepa fúngica Medio Bagazo (g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
LiP (UI/L)
IB29B(Blasthomyces) 3 1 80 0.5 28 26,07
IB18Cb (Bdellospora) 8 5 80 2 21 487,82
IB-26C (Trinacrium) 8 5 80 2 28 44,59
IB15B (Stephanoma) 8 5 80 2 28 69,99
IB15Da(Cephalosporium) 3 1 80 0.5 28 32,10
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
08
De manera peculiar la cepa IB-18Cb (Bdellospora) fue la única de las cinco cepas
testeadas que presento una buena actividad enzimática (figura 54) ya que las demás cuatro no
llegaron ni a la sexta parte de lo que alcanzó esta enzima. Cabe recalcar que la LiP es la más
escasa del sistema ligninolítico.
c. Lacasa.-
Tabla 27: Actividad enzimática Lac de diferentes cepas fúngicas con variación Bagazo
En la tabla 27 veremos el resumen de resultados para el sustito bagazo, comparándolo
con los resultados de variación manganeso, la actividad fue mas baja.
La cepa fúngica IB-18Cb (Bdellospora) fue una de las que presento mayor actividad
enzimática para Lacasa (Lac) a los 21días, en el medio 3, llegando a 56,24 (UI/L).
IB-18Cb (Bdellospora) según análisis realizados para esta cepa fúngica se dice que a
mayor concentración de manganeso, se obtendría mayor actividad enzimática, también es
importante la concentración de extracto de levadura en el medio, pero la concentración de
Cepa fúngica Medio Bagazo (g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
Lac (UI/L)
IB29B(Blasthomyces) 2 1 40 2 21 27,08
IB18Cb (Bdellospora) 3 1 80 0.5 21 56,24
IB-26C (Trinacrium) 8 5 80 2 21 48,56
IB15B (Stephanoma) 5 1 80 2 28 8,18
IB-6 (Circinutrychum) 2 1 40 2 28 11,43

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
09
glucosa no influye en la actividad enzimática.
Figura 55: Actividad enzimática Lacasa de 5 cepas fúngicas en función del bagazo.
Lacasa (bagazo)
0
10
20
30
40
50
60
0 5 10 15 20 25 30
Tiempo (UI/L)
Act
ivid
ad (U
I/L)
IB-29B IB-18Cb IB-26C IB-15B IB-6
Si vemos la figura 4-34 notaremos que la cepa IB-26C (Trinacrium) no tiene mucha
diferencia con IB-18Cb (Bdellospora), pero al contrario esta cepa es dependiente de glucosa, y
por supuesto de manganeso.
La aceptación del sustrato bagazo, por la cepa IB-18Cb, para la producción de LiP
como se observa en la figura 56 fue la que mejor resultado obtuvo, sin embargo el medio que
contiene paja brava resulta ser un medio mucho mas confiable, que el medio del bagazo, esto
por su rango de error que presenta.
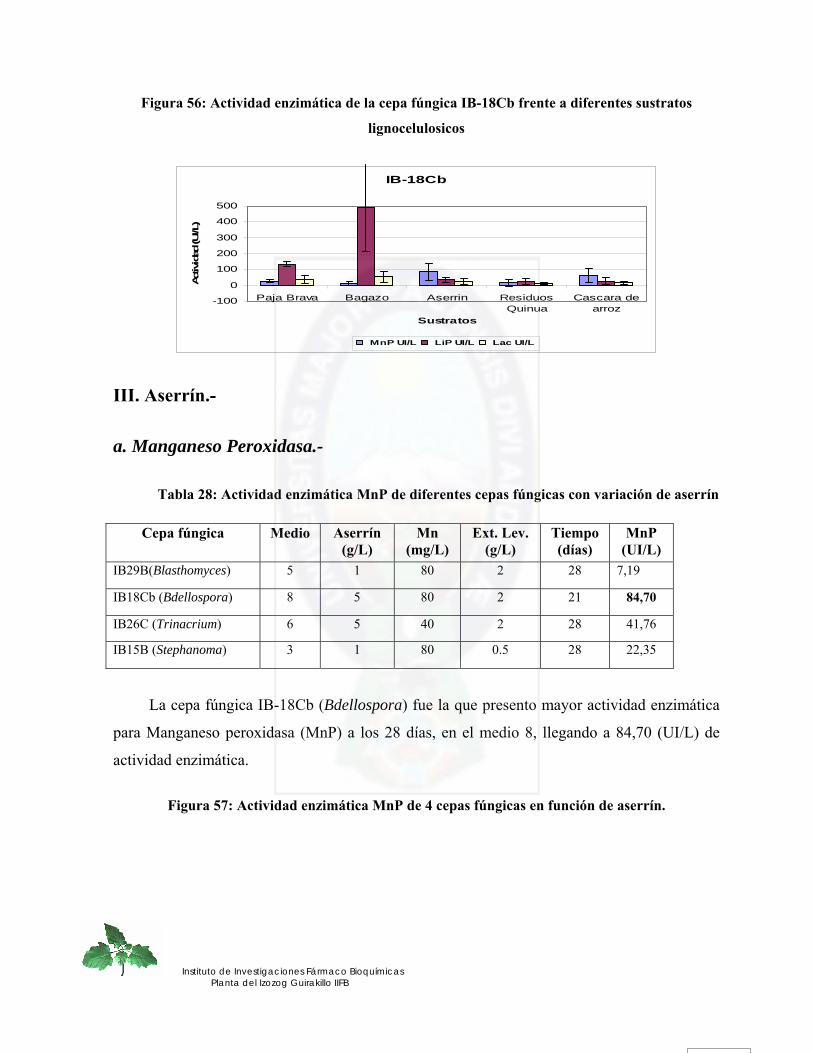
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
0
Figura 56: Actividad enzimática de la cepa fúngica IB-18Cb frente a diferentes sustratos
lignocelulosicos
IB-18Cb
-100
0
100
200
300
400
500
Paja Brava Bagazo Aserrin ResiduosQuinua
Cascara dearroz
Sustratos
Activ
idad
(UI/L
)
MnP UI/L LiP UI/L Lac UI/L
III. Aserrín.-
a. Manganeso Peroxidasa.-
Tabla 28: Actividad enzimática MnP de diferentes cepas fúngicas con variación de aserrín
La cepa fúngica IB-18Cb (Bdellospora) fue la que presento mayor actividad enzimática
para Manganeso peroxidasa (MnP) a los 28 días, en el medio 8, llegando a 84,70 (UI/L) de
actividad enzimática.
Figura 57: Actividad enzimática MnP de 4 cepas fúngicas en función de aserrín.
Cepa fúngica Medio Aserrín (g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
MnP (UI/L)
IB29B(Blasthomyces) 5 1 80 2 28 7,19
IB18Cb (Bdellospora) 8 5 80 2 21 84,70
IB26C (Trinacrium) 6 5 40 2 28 41,76
IB15B (Stephanoma) 3 1 80 0.5 28 22,35

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
Manganeso Peroxidasa (aserrin)
0
15
30
45
60
75
90
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L)
IB-29B IB-18Cb IB-26C IB-15B
En la figura 57 observaremos que hasta los 21 días las cepas IB-18Cb (Bdellospora) y
IB-26C (Trinacrium) se asimilaban en su actividad pero ya a los 28 días IB-26C (Trinacrium)
bajo su actividad enzimática, mientras IB-18Cb (Bdellospora) incrementó notablemente su
actividad enzimática.
No podemos negar que el manganeso juega un papel muy importante en esta actividad
enzimática, si observamos la composición de los medios escogidos, la mayoría tiene
requerimientos nutricionales donde el manganeso tiene una alta concentración.
b. Lignina Peroxidasa.-
Tabla 29: Actividad enzimática LiP de diferentes cepas fúngicas con variación de aserrín
La
cepa
fúngi
ca
IB-15B (Stephanoma) fue una de las que presento mayor actividad enzimática para Lignina
peroxidasa (LiP) a los 28 días, en el medio 8, llegando a 42,25 (UI/L).
Cepa fúngica Medio Aserrín (g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
LiP (UI/L)
IB29B(Blasthomyces) 3 1 80 0.5 21 32,94
IB18Cb (Bdellospora) 8 5 80 2 21 33,91
IB-26C (Trinacrium) 8 5 80 2 21 29,56
IB15B (Stephanoma) 8 5 80 2 28 42,25
IB15Da(Cephalosporium) 3 1 80 0.5 28 29,45

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
2
La cepa que alcanzo la actividad máxima requería una fuente de manganeso alta, pero
esto se presento a los 28 días, ya que de los 15 a 21 días, no tenia buen alcance enzimático
como se puede ver en la figura 4-36.
Las actividades enzimáticas obtenidas en los medios con aserrín, probablemente se
deba a que en las maderas existe una gran variedad de compuestos aromáticos que habrían
pasado a la solución [3]. El manganeso que comúnmente esta presente en las maderas también
pudo haber sido el inductor [88].
Figura 58: Actividad enzimática Lignina peroxidasa de 5 cepas fúngicas en función de la aserrín.
Lignina Peroxidasa (aserrin)
0
10
20
30
40
50
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L)
IB-29B IB-18Cb IB-26C IB-15B IB-15Da
En la figura 58 observaremos que las cinéticas enzimáticas que presentaron las cinco
cepas fúngicas no tuvieron diferencia como en anteriores casos, al parecer este sustrato fue
asimilado por las cinco cepas, aunque esto no estimule una buena actividad enzimática.
c. Lacasa.-

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
3
Tabla 30: Actividad enzimática MnP de diferentes cepas fúngicas con variación de aserrín
La cepa fúngica IB-29B (Blasthomyces) fue la que presento mayor actividad enzimática
para Lacasa (Lac) a los 28 días, en el medio 2, llegando a 30,09 (UI/L).
Las respuestas enzimáticas para cada cepa son diferentes, ya que cuando una variación
se realiza en el medio de cultivo, todas las condiciones cambian, como es le caso de IB-29B
(Blasthomyces) que en su caso, la concentración de manganeso y glucosa no influyen en la
actividad enzimática.
Figura 59: Actividad enzimática Lacasa de 5 cepas fúngicas en función del aserrín.
Lacasa (aserrin)
0
10
20
30
40
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L)
IB-29B IB-18Cb IB-26C IB-15B IB-6
En la tabla 29 se puede notar claramente que los valores de las cinco cepas fúngicas
Cepa fúngica Medio Aserrín (g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
LiP (UI/L)
IB29B(Blasthomyces) 2 1 40 2 28 30,09
IB18Cb (Bdellospora) 3 1 80 0.5 21 25,04
IB-26C (Trinacrium) 8 5 80 2 21 24,25
IB15B (Stephanoma) 5 1 80 2 28 12,72
IB-6 (Circinutrychum) 2 1 40 2 28 26,74

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
4
testeadas no tienen diferencia significativa, con excepción de IB-15B (Stephanoma) que no
alcanzo buena actividad para LiP.
Figura 60: Actividad enzimática de la cepa fúngica IB-26C frente a diferentes sustratos
lignocelulosicos
IB-26C
0
20
40
60
80
100
Paja Brava Bagazo Aserrin ResiduosQuinua
Cascara dearroz
Sustratos
Act
ivid
ad (U
I/L)
MnP UI/L LiP UI/L Lac UI/L
Por la composición química que presenta el aserrín se esperaba que fuera el sustrato
mejor asimilado, en la cepa IB-26C tuvo una mayor actividad enzimática para Mn P , en
cambio para LiP y Lacasa fue superado por el sustrato residuos de quinua.
IV. Residuos de Quinua Real.-
a. Manganeso Peroxidasa.-
Tabla 31: Actividad enzimática MnP de diferentes cepas fúngicas con variación de
residuos de quinua.
Cepa fúngica Medio Residuos quinua (g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
MnP (UI/L)
IB29B(Blasthomyces) 5 1 80 2 21 80,74
IB18Cb (Bdellospora) 8 5 80 2 21 14,55
IB26C (Trinacrium) 6 5 40 2 28 13,07
IB15B (Stephanoma) 3 1 80 0.5 21 26,72
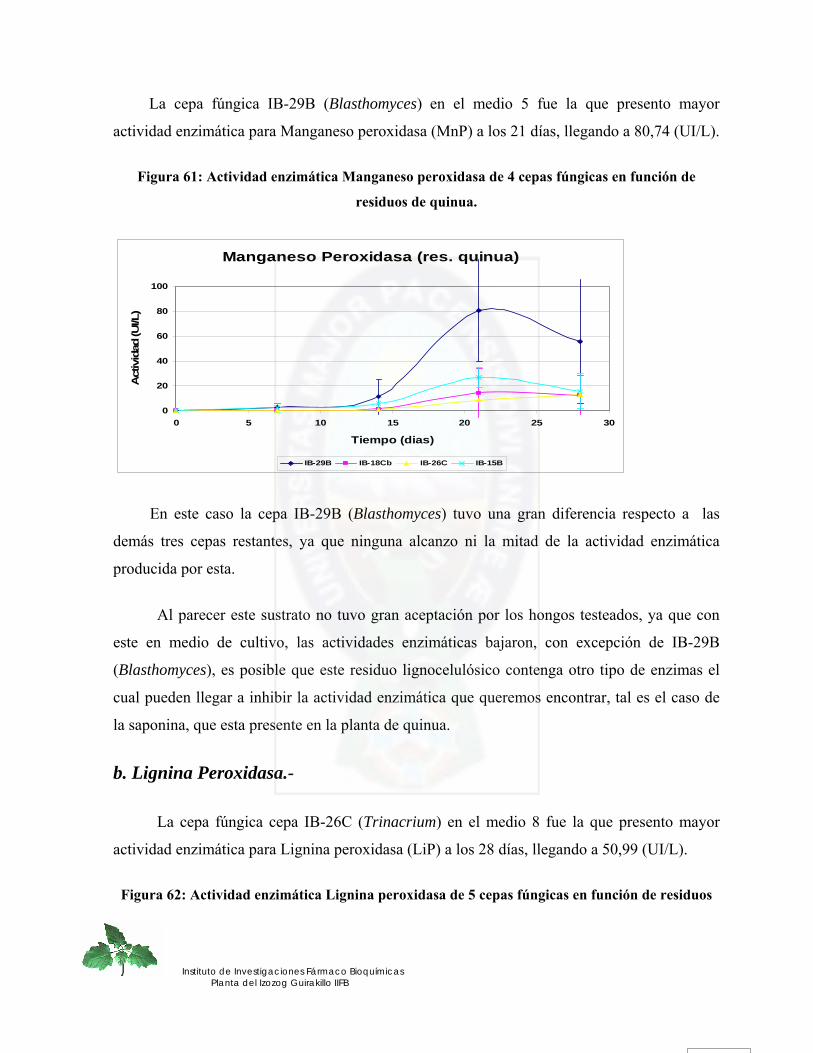
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
La cepa fúngica IB-29B (Blasthomyces) en el medio 5 fue la que presento mayor
actividad enzimática para Manganeso peroxidasa (MnP) a los 21 días, llegando a 80,74 (UI/L).
Figura 61: Actividad enzimática Manganeso peroxidasa de 4 cepas fúngicas en función de
residuos de quinua.
Manganeso Peroxidasa (res. quinua)
0
20
40
60
80
100
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L)
IB-29B IB-18Cb IB-26C IB-15B
En este caso la cepa IB-29B (Blasthomyces) tuvo una gran diferencia respecto a las
demás tres cepas restantes, ya que ninguna alcanzo ni la mitad de la actividad enzimática
producida por esta.
Al parecer este sustrato no tuvo gran aceptación por los hongos testeados, ya que con
este en medio de cultivo, las actividades enzimáticas bajaron, con excepción de IB-29B
(Blasthomyces), es posible que este residuo lignocelulósico contenga otro tipo de enzimas el
cual pueden llegar a inhibir la actividad enzimática que queremos encontrar, tal es el caso de
la saponina, que esta presente en la planta de quinua.
b. Lignina Peroxidasa.-
La cepa fúngica cepa IB-26C (Trinacrium) en el medio 8 fue la que presento mayor
actividad enzimática para Lignina peroxidasa (LiP) a los 28 días, llegando a 50,99 (UI/L).
Figura 62: Actividad enzimática Lignina peroxidasa de 5 cepas fúngicas en función de residuos
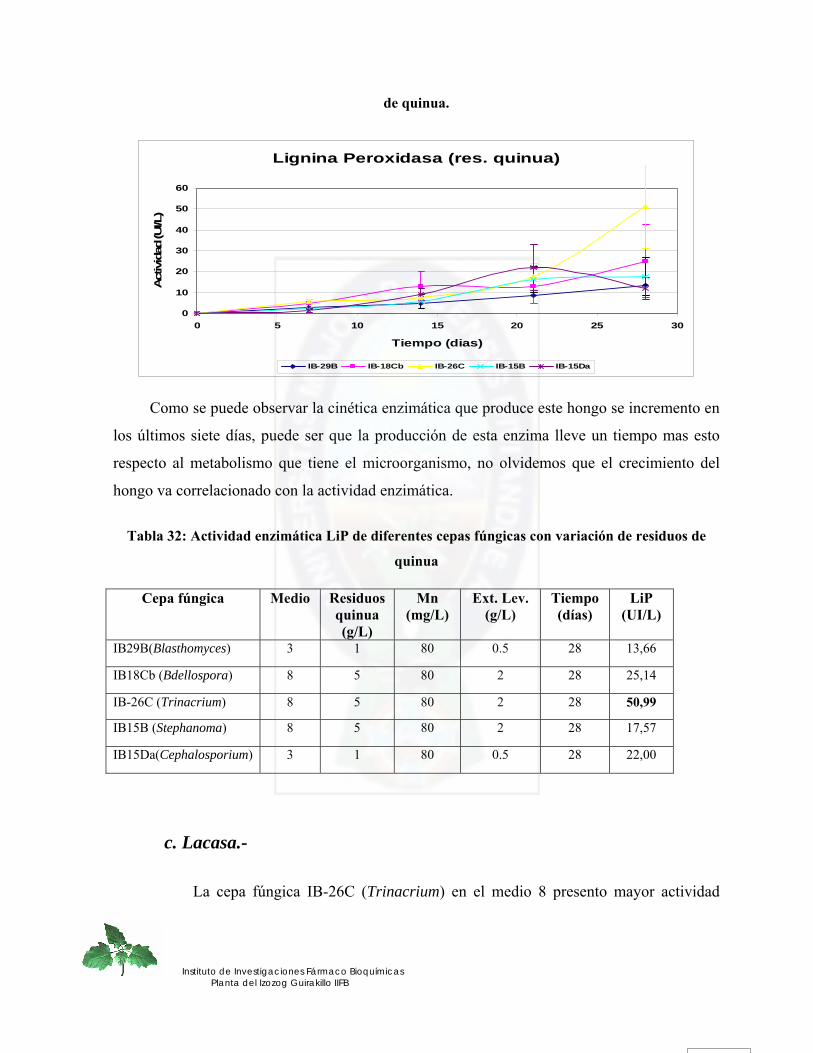
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
6
de quinua.
Lignina Peroxidasa (res. quinua)
0
10
20
30
40
50
60
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L)
IB-29B IB-18Cb IB-26C IB-15B IB-15Da
Como se puede observar la cinética enzimática que produce este hongo se incremento en
los últimos siete días, puede ser que la producción de esta enzima lleve un tiempo mas esto
respecto al metabolismo que tiene el microorganismo, no olvidemos que el crecimiento del
hongo va correlacionado con la actividad enzimática.
Tabla 32: Actividad enzimática LiP de diferentes cepas fúngicas con variación de residuos de
quinua
c. Lacasa.-
La cepa fúngica IB-26C (Trinacrium) en el medio 8 presento mayor actividad
Cepa fúngica Medio Residuos quinua (g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
LiP (UI/L)
IB29B(Blasthomyces) 3 1 80 0.5 28 13,66
IB18Cb (Bdellospora) 8 5 80 2 28 25,14
IB-26C (Trinacrium) 8 5 80 2 28 50,99
IB15B (Stephanoma) 8 5 80 2 28 17,57
IB15Da(Cephalosporium) 3 1 80 0.5 28 22,00

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
enzimática para Lacasa (Lac) a los 28 días llegando a 62,62 (UI/L).
Tabla 33: Actividad enzimática Lac de diferentes cepas fúngicas con variación de residuos de
quinua
IB-26C (Trinacrium) influenciada por la fuente de nitrógeno y vitaminas que contiene
en su medio de cultivo alcanzo una actividad enzimática ala cual supera como cinco veces más
a las de más cuatro cepas fúngicas, no olvidemos que en análisis anteriores de este hongo
estaba inducida por la glucosa, esto puede afecta a que la actividad haya bajado según pruebas
realizadas anteriormente.
Figura 64: Actividad enzimática Lacasa de 5 cepas fúngicas en función de residuos de quinua.
Lacasa (res. quinua)
0
10
20
30
40
50
60
70
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L)
IB-29B IB-18Cb IB-26C IB-15B IB-6
El sustrato bagazo fue el sustrato que asimilaron mejor la mayoría de las cepas fúngicas
Cepa fúngica Medio Residuos quinua (g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
Lac (UI/L)
IB29B(Blasthomyces) 2 1 40 2 28 10,57
IB18Cb (Bdellospora) 3 1 80 0.5 28 12,25
IB-26C (Trinacrium) 8 5 80 2 28 62,62
IB15B (Stephanoma) 5 1 80 2 28 1,42
IB-6 (Circinutrychum) 2 1 40 2 28 9,94
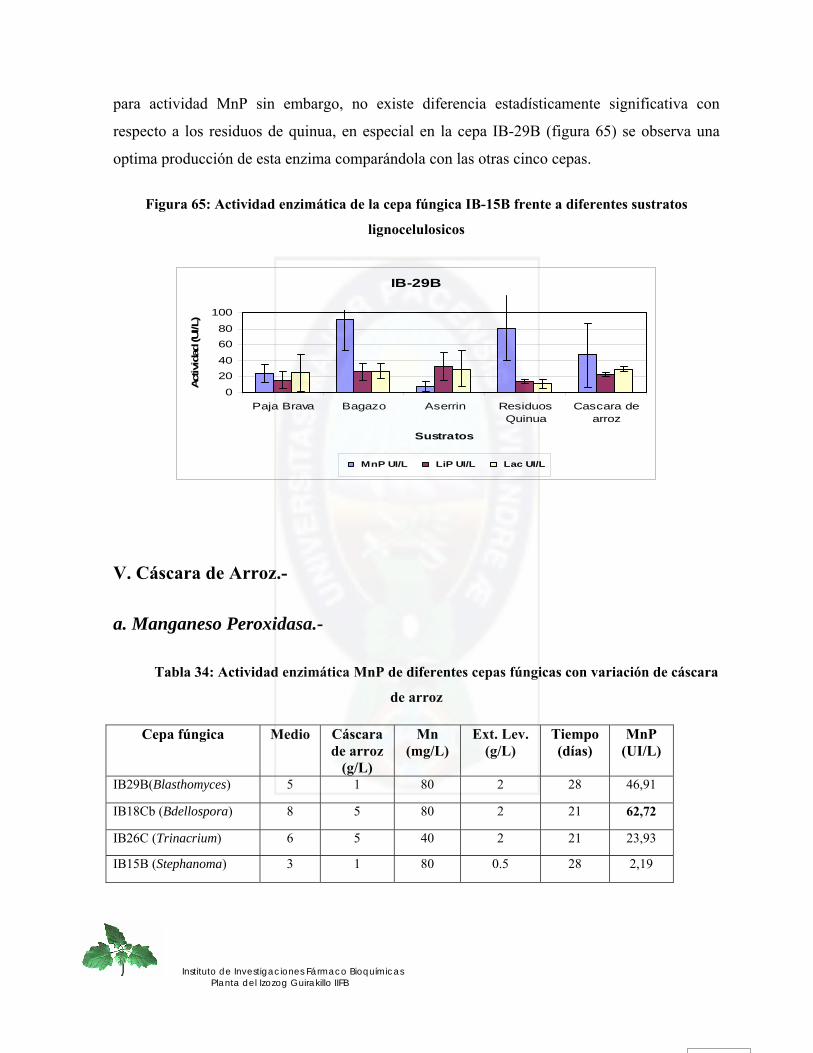
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
8
para actividad MnP sin embargo, no existe diferencia estadísticamente significativa con
respecto a los residuos de quinua, en especial en la cepa IB-29B (figura 65) se observa una
optima producción de esta enzima comparándola con las otras cinco cepas.
Figura 65: Actividad enzimática de la cepa fúngica IB-15B frente a diferentes sustratos
lignocelulosicos
IB-29B
0
2040
6080
100
Paja Brava Bagazo Aserrin ResiduosQuinua
Cascara dearroz
Sustratos
Act
ivid
ad (U
I/L)
MnP UI/L LiP UI/L Lac UI/L
V. Cáscara de Arroz.-
a. Manganeso Peroxidasa.-
Tabla 34: Actividad enzimática MnP de diferentes cepas fúngicas con variación de cáscara
de arroz
Cepa fúngica Medio Cáscara de arroz
(g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
MnP (UI/L)
IB29B(Blasthomyces) 5 1 80 2 28 46,91
IB18Cb (Bdellospora) 8 5 80 2 21 62,72
IB26C (Trinacrium) 6 5 40 2 21 23,93
IB15B (Stephanoma) 3 1 80 0.5 28 2,19

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
9
La cepa fúngica IB-18Cb (Bdellospora) fue la que presento mayor actividad
enzimática para Manganeso peroxidasa (MnP) a los 21 días, en el medio 8, llegando a 62,72
(UI/L).
Figura 66: Actividad enzimática Manganeso peroxidasa de 4 cepas fúngicas en función de
cáscara de arroz.
MnP (cas. arroz)
0
10
20
30
40
50
60
70
80
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L)
IB-29B IB-18Cb IB-26C IB-15B
La cinética enzimática de estas cepas fúngicas se puede observar en la figura 4-41,
podemos notar que entre las cuatro cepas su cinética es muy diferente, para IB-18Cb
(Bdellospora) que alcanzó una actividad tres veces mas que en la prueba donde se utilizo
glucosa, esto quiere decir que este medio es mas optimo para este hongo, se puede decir que la
glucosa, extracto de levadura no influyen en la producción de enzimas.
La cepa IB-29B (Blasthomyces) que depende exclusivamente del extracto de levadura,
y siendo indispensable este en el medio de cultivo para que este microorganismo sea capaz de
producir enzimas.
b. Lignina Peroxidasa.-
Tabla 35: Act enz LiP de diferentes cepas fúngicas con variación de cáscara de arroz
Cepa fúngica Medio Cáscara de arroz
(g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
LiP (UI/L)
IB29B(Blasthomyces) 3 1 80 0.5 28 22,82
IB18Cb (Bdellospora) 8 5 80 2 21 26,74
IB-26C (Trinacrium) 8 5 80 2 21 20,55
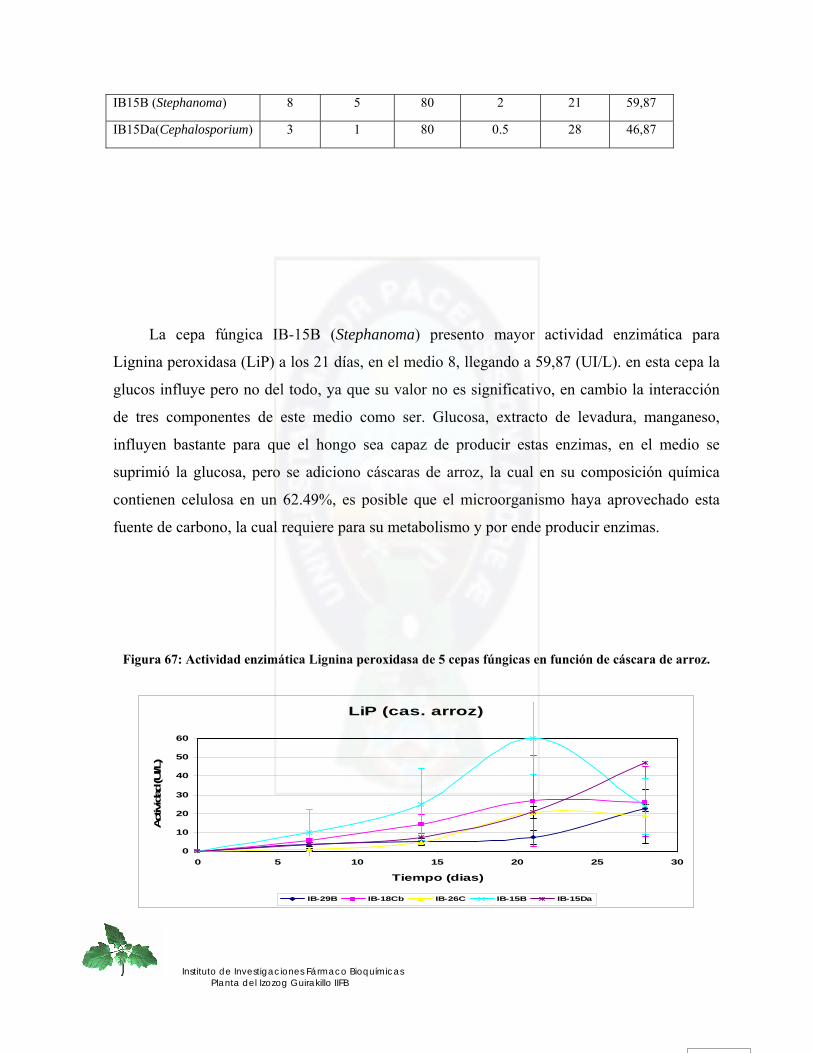
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
20
La cepa fúngica IB-15B (Stephanoma) presento mayor actividad enzimática para
Lignina peroxidasa (LiP) a los 21 días, en el medio 8, llegando a 59,87 (UI/L). en esta cepa la
glucos influye pero no del todo, ya que su valor no es significativo, en cambio la interacción
de tres componentes de este medio como ser. Glucosa, extracto de levadura, manganeso,
influyen bastante para que el hongo sea capaz de producir estas enzimas, en el medio se
suprimió la glucosa, pero se adiciono cáscaras de arroz, la cual en su composición química
contienen celulosa en un 62.49%, es posible que el microorganismo haya aprovechado esta
fuente de carbono, la cual requiere para su metabolismo y por ende producir enzimas.
Figura 67: Actividad enzimática Lignina peroxidasa de 5 cepas fúngicas en función de cáscara de arroz.
LiP (cas. arroz)
0
10
20
30
40
50
60
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L)
IB-29B IB-18Cb IB-26C IB-15B IB-15Da
IB15B (Stephanoma) 8 5 80 2 21 59,87
IB15Da(Cephalosporium) 3 1 80 0.5 28 46,87
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
2
Comparando con el medio que se utilizo glucosa, esta cepa alcanzo un similar
resultado. En las cepas fúngicas restantes como la IB-15Da (Cephalosporium) el componente
que realmente influye en la expresión de enzimas es el manganeso, por lo cual el resultado fue
mas optimo en esta prueba que en la prueba donde se utiliza glucosa.
En cambio en la cepa IB-18Cb (Bdellospora) se obtuvo un resultado mas bajo que el
anterior, esto, por que, para el metabolismo de este hongo, es indispensable una fuente de
carbono, y en concentración alta.
c. Lacasa.-
Tabla 36: Actividad enzimática MnP de diferentes cepas fúngicas con variación de
cáscara de arroz
La cepa fúngica IB-29B (Blasthomyces) en el medio 2 fue una de las que presento
mayor actividad enzimática para Lacasa (Lac) a los 21 días llegando a 28,70 (UI/L). Por lo
visto anteriormente esta cepa no es dependiente de glucosa en concentraciones altas, pero si de
extracto de levadura, el medio en el cual se realizo la prueba contenía a una alta concentración
de fuente de nitrógeno, es por eso que se esperaba un mejor resultado.
Figura 68: Actividad enzimática Lacasa de 5 cepas fúngicas en función de cáscara de arroz.
Cepa fúngica Medio Cáscara de arroz
(g/L)
Mn (mg/L)
Ext. Lev. (g/L)
Tiempo (días)
LiP (UI/L)
IB29B(Blasthomyces) 2 1 40 2 21 28,70
IB18Cb (Bdellospora) 3 1 80 0.5 28 14,51
IB-26C (Trinacrium) 8 5 80 2 28 11,22
IB15B (Stephanoma) 5 1 80 2 21 4,67
IB-6 (Circinutrychum) 2 1 40 2 28 15,99

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
22
Lacasa (cas. arroz)
0
5
10
15
20
25
30
0 5 10 15 20 25 30
Tiempo (dias)
Act
ivid
ad (U
I/L)
IB-29B IB-18Cb IB-26C IB-15B IB-6
Algunos metabolitos que están presentes en el sustrato lignocelulósico pueden
influenciar en la producción de enzimas [81].
Los resultados obtenidos con sustratos lignocelulosicos, reemplazando la glucosa, como
componente del medio de cultivo, en algunos casos como es el de IB-18Cb (Bdellospora) en la
actividad enzimática LiP utilizando como sustrato bagazo, alcanzo una actividad impresiónate,
en otros casos no se obtuvo buenos resultados, pero esto depende de cada cepa fúngicas, ya
que cada una tiene requerimientos nutricionales diferentes, realizando una comparación de la
mejor cepa fúngica para cada sustrato y capacidad de producir enzima podemos ver el anexo
C-7, C-8, C-9, donde se enguanta los hongos que presentaron mejor actividad para cada
enzima.
Podemos resaltar que estos sustratos asimilaron mejor en la actividad lignina
peroxidasa, que las otras dos enzimas
4.4. Determinación de Actividad Celulolítica.-
De los resultados obtenidos para enzimas celulolíticas de actividad combinada, en esta
prueba solo una cepa dio resultados significativos, fue la cepa IB-29B (Blasthomyces) los
cuales se resumen a continuación.
Tabla 37: Actividad celulítica para -29B (Blasthomyces).

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
23
Los resultados obtenidos para las demás cepas fúngicas no fueron significativos, es por
eso que no se los tomo en cuenta.
Para realiza esta prueba se realizo una curva de calibración la cual se puede observar en
anexo E-1.
Esta prueba se realizo queriendo verificar si existen enzimas tipo hidrolasas, ya que esta
enzimas trabajan en conjunto con las enzimas ligninolíticas, teniendo como fin degradar la
celulosa, las primeras, y las otras degradar la lignina.
4.5. Determinación de proteínas totales.-
La absorción a 660 nm se transformo en ug/mL estandarizada con albúmina de suero
bovino (Sigma).
Esta prueba se realizo con el objetivo de evaluar si existía alguna limitación en algunos
nutrientes que estuviera afectando la producción de las actividades obtenidas en ambos medios
[3]. Otro objetivo de era prueba era el de comprobar la existencia de enzimas en el medio , las
cuales son proteínas especializadas.
Los resultados más importantes, fueron siguientes:
Tabla 38: Concentración de proteínas totales para IB-26C (Trinacrium)
IB-26C (Trinacrium) Ci (g/L) Cf (g/L)
Medio 1 0,28 0,4 Medio 2 0,18 0,3 Medio 3 0,19 0,2
IB-29B(Blasthomyces) C (g/L) Medio 6 0,6 Medio 7 0,6
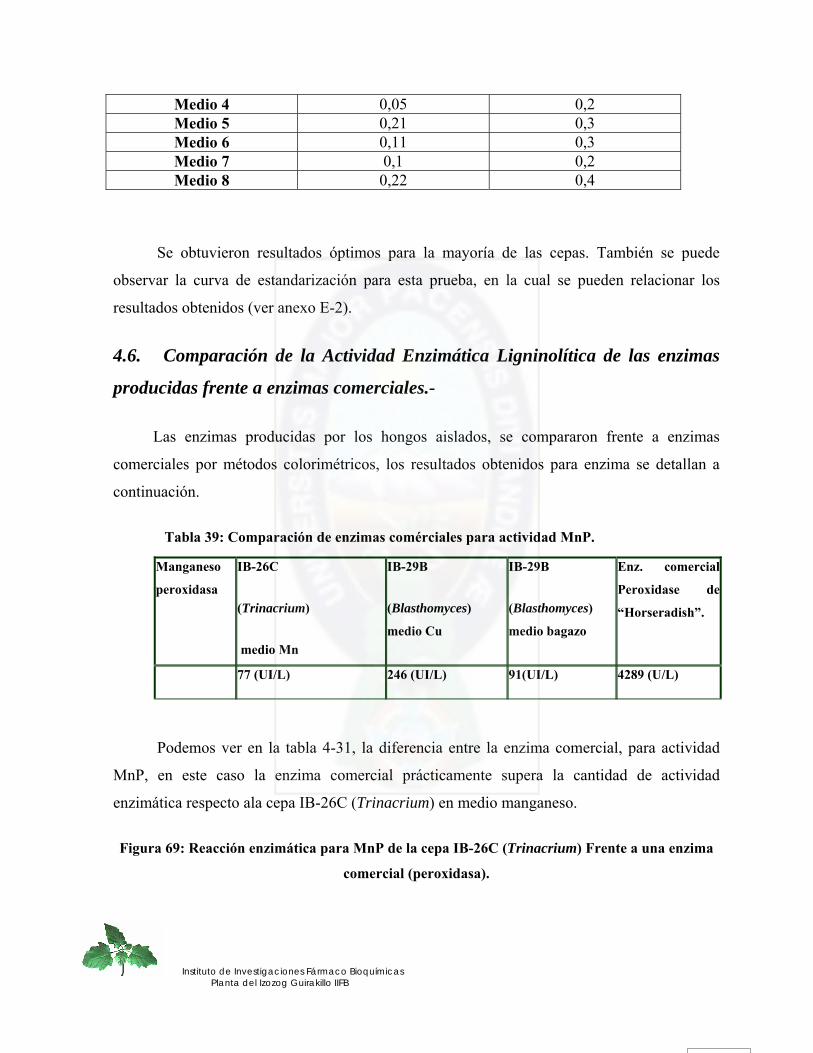
Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
24
Medio 4 0,05 0,2 Medio 5 0,21 0,3 Medio 6 0,11 0,3 Medio 7 0,1 0,2 Medio 8 0,22 0,4
Se obtuvieron resultados óptimos para la mayoría de las cepas. También se puede
observar la curva de estandarización para esta prueba, en la cual se pueden relacionar los
resultados obtenidos (ver anexo E-2).
4.6. Comparación de la Actividad Enzimática Ligninolítica de las enzimas
producidas frente a enzimas comerciales.-
Las enzimas producidas por los hongos aislados, se compararon frente a enzimas
comerciales por métodos colorimétricos, los resultados obtenidos para enzima se detallan a
continuación.
Tabla 39: Comparación de enzimas comérciales para actividad MnP.
Podemos ver en la tabla 4-31, la diferencia entre la enzima comercial, para actividad
MnP, en este caso la enzima comercial prácticamente supera la cantidad de actividad
enzimática respecto ala cepa IB-26C (Trinacrium) en medio manganeso.
Figura 69: Reacción enzimática para MnP de la cepa IB-26C (Trinacrium) Frente a una enzima
comercial (peroxidasa).
Manganeso
peroxidasa
IB-26C
(Trinacrium)
medio Mn
IB-29B
(Blasthomyces)
medio Cu
IB-29B
(Blasthomyces)
medio bagazo
Enz. comercial
Peroxidase de
“Horseradish”.
77 (UI/L) 246 (UI/L) 91(UI/L) 4289 (U/L)

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
2
En la figura 4-44 podemos ver el cambio de coloración que sufrió la reacción gracias a
la enzima MnP, en la cual la que mas oxidación produjo fue la enzima comercial.
Tabla 40: Comparación de enzimas comérciales para actividad LiP.
Para la enzima LiP los resultados de IB-18Cb (Bdellospora) en un medio con variación
de residuos de bagazo alcanza un 10% de la actividad enzimática de la enzima comercial (ver
tabla 4-31).
Figura 70: Reacción enzimática para LIP de la cepa IB-29B(Blasthomyces) frente a una enzima
comercial (peroxidasa).
IB-26C(Trinacrium) medio manganeso
IB-6(Trinacrium) medio cobre
IB18Cb(Bdellospora) medio bagazo
Enzima comercial
76,50926 (UI/L) 251 (UI/L) 488 (UI/L) 4633 (UI/L)

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
26
En la figura 4-45 observamos la oxidación que produce LiP sobre el sustrato ABTS,
como resultado se torna una coloración verde oscura.
Tabla 41: Comparación de enzimas comérciales para actividad Lac.
Para la comparación de actividad enzimática de lacasa, las enzimas comerciales alcanzaron
mayor actividad enzimática que las enzimas producidas.
Para la determinación de la actividad enzimática de las enzimas comerciales, se tuvo
que realizar diluciones 1/10 para que así entren en el rango de linealidad y pueda ser
comparadas con las enzimas producidas en el presente trabajo.
Cabe la pena aclarar que las enzimas producidas, están en bruto, ya que no han sido
purificadas, esto puede influir bastante por que pueden haber elementos que estén inhibiendo
su reacción enzimática, como puede haber elementos que incrementen su actividad
enzimática, son bastantes los factores que pueden estar influyendo.
Figura 71: Reacción enzimática para Lac de la cepa IB-15Da(Cephalosporium) frente a una
IB29B(Blasthomyces) medio manganeso
IB15Da(Cephalosporium) medio cobre
IB26C(Trinacrium) medio res quinua
Enzima comercial
74 (UI/L) 287 (UI/L) 63 (UI/L) 4204 (UI/L)

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
2
enzima comercial (lacasa).
Esta prueba fue realizada, par tener una ida de la actividad enzimática de las enzimas
producidas.
5. CONCLUSIONES
Dentro de todos los objetivos planteados inicialmente se legaron a las siguientes
conclusiones.
Se realizo un viaje de recolección de muestras, en el cual pudimos recolectar 30
muestras diferentes de diferentes fuentes, el viaje se realizo en una parte del Altiplano
boliviano, en los departamentos de La Paz y Oruro, es importante mencionar que un
porcentaje de hongos han sido aislados de diferentes zonas geográficas de Bolivia.
De las muestras recolectadas logramos aislar 31 cepas fúngicas diferentes, las cuales
aun no han sido totalmente estudiadas, de igual forma, no hubiera sido posible realizar el
presente trabajo sin la presencia de hongos. De los dos métodos que se utilizaron, el más
efectivo fue el método indirecto, ya que se logro aislar más variedad de hongos que en el
método indirecto, lo cual nos permite nuevos microorganismos con características
interesantes, que despiertan ideas de investigación.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
28
De las 31 cepas aisladas se identifico y caracterizó a un 80,64%, debido a la diversidad
que presentan los hongos en el ecosistema, la identificación que se realizo solo fue apoyada en
bibliografía, y se estima que sean correctos, por esta razón es necesario confirmarlos por
métodos de biología molecular. Respecto al 19, 35% que no fueron identificadas constituyen
posiblemente a nuevas especies aun no estudiadas.
Debido ala capacidad de producir enzimas con actividad enzimática ligninolítica se
realizo dos selecciones de cepas fúngicas, la primera selección realizada en medios sólidos
específicos, prueba de la cual el 38,7% de los hongos aislados presentaron actividad
ligninolítica, el restos de los hongos 61,3% no presentaron actividad ligninolítica, razón por la
cual no se los tomo en cuenta para otras pruebas en adelante.
Este método cualitativo es muy práctico, par la determinación de actividad enzimática,
ya que permita ahorrar materiales y tiempo en comparación con otras determinaciones
cuantitativas, de todas maneras es indispensable confirmar los resultados con pruebas
espectrofotométricas.
En cuanto a la segunda selección de las doce cepas seleccionadas, solo seis mostraron
una buena actividad enzimática, esto se debe a que muy pocas son las cepas fúngicas con
actividad ligninolítica que están distribuidas ampliamente en el ecosistema. Las seis cepas
seleccionadas fueron: IB-29B (Blasthomyces), IB-18Cb (Bdellospora), IB-26C (Trinacrium),
IB-15B (Stephanoma), IB-15Da (Cephalosporium), IB-6 (Circinutrychum).
Se realizo un diseño factorial 23, siendo los factores: glucosa, manganeso, extracto de
levadura, con esto se obtuvo 8 diferentes medios con diferente composición, lo cual permitió
obtener una mejor actividad enzimática, buscando además el medio especifico para cada cepa
fúngica y para cada enzima.
De acuerdo alas pruebas realizadas por métodos espectrofotométricos logramos
cuantificar la actividad enzimática de las cepas fúngicas seleccionadas. Con estos resultados
obtenidos llegamos a varias conclusiones.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
29
Para actividad MnP y LiP la cepa que presento la mejor actividad enzimática en medio
manganeso fue la IB-26C (Trinacrium) debido ala cinética que presento, podemos afirmar que
esta cepa es capaz de producir enzimas peroxidasas óptimamente.
El hongo capaz de expresar enzimas de actividad lacasa óptimamente es IB-29B
(Blasthomyces).
Los medios de elección en la mayoría de las cepas fúngicas son el 5 y 8, para actividad
MnP, esto se debe a que es importante la fuente de nitrógeno y necesitan un inductor en este
caso es el manganeso para la producción de estas enzimas.
Para una mejor actividad enzimática LiP, los microorganismos requieren una mayor
fuente de nitrógeno, pero la actividad enzimática no se produciría si no existiría la interacción
de los tres compuestos.
La concentración de glucosa no influye en el medio de cultivo para la producción de
enzimas de actividad enzimática lacasa, lo que es indispensable en este medio de cultivo es un
inductor en este caso manganeso.
La mayor actividad enzimática para las enzimas ligninolíticas se da entre los 21 y 28
días, esto para el medio manganeso.
Cada microorganismo tiene diferentes requerimientos nutricionales.
Las tres enzimas ligninolíticas determinas tienen diferentes exigencias para potenciar su
actividad enzimática.
Las cepas que fueron capaces de producir los tres tipos de enzimas fueron: IB-29B
(Blasthomyces), IB-18Cb (Bdellospora), IB-26C (Trinacrium), IB-15B (Stephanoma).
De acuerdo ala segunda selección realizada de los hongos aislados se realizo con un
segundo diseño factorial 23, siendo los factores esta vez: glucosa, cobre, extracto de levadura,
y se observo la actividad enzimática que puedan producir en estos medios.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
30
En la prueba de variación cobre para actividades enzimática de enzimas peroxidasas se
determinó que la concentración de cobre influye en la actividad enzimática, sin embargo no se
necesita altas concentraciones de este metal para optimizar la actividad enzimática, según los
resultados obtenidos, las concentraciones tienen que ser bajas, de lo contrarió puede llegar a
intoxicar al microorganismo e inhibir la síntesis de enzimas.
Con respecto a lacasas se requiere una concentración mínima de glucosa, y la
concentración de extracto de levadura es muy importante, ya que es muy necesaria la fuente de
nitrógeno para estas enzimas
Realizando una comparación de resultados alcanzados con la variación cobre con
respecto a los resultados con variación de manganeso, notamos claramente que los resultados
tiene mucha diferencia, en este sentido podemos afirmar, que el medio de cultivo de cobre es
mas efectivo que el medio de cultivo manganeso.
Se evaluó cinco sustratos lignocelulosicos diferentes para determinar la producción de
enzimas ligninolíticas, en medios donde se suprimió la glucosa por: paja brava, bagazo,
aserrín, residuos de quinua, cáscara de arroz.
Variando las condiciones de cultivo se consiguió mejorar el rendimiento, optimizando
en algunos casos la producción de enzimas, como es el caso de LiP.
Con esta variación de aprovechamiento de sustratos lignocelulosicos se logro reducir
costos del proceso de fermentación.
En los fermentos de cultivo fungicos se encontraron dos tipos de actividda enzimática
celulolítica y redox. Esta observación fue evidente con la cepa fungica Blatomyces.
Se determinó la cuantificación de proteínas, resultados con los cuales afirmamos que
existe una biomasa proteica, equivalente ala actividad enzimática determinada.
Como referencia, se determinó la comparación de la capacidad de la actividad

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
3
enzimática de las enzimas producidas frente a enzimas comerciales.
Las cepas fúngicas aisladas producieron un 52 a 54% de la actividad enzimática
producida por enzimas comerciales.
Las enzimas producidas de actividad enzimática LiP posen mayor actividad enzimática
que las comerciales, sin embargo tomamos en cuanta que estas enzimas producidas en el
presente trabajo no están purificadas.
6. RECOMENDACIONES
- Realizar recolección de muestras explorando más regiones, en especial del altiplano
boliviano, debido ala amplia diversidad del país.
- Hacer pruebas de biología molecular, para la identificación de las cepas fúngicas aisladas,
para verificar, los resultados estimados de esta prueba.
- Realizar estudios profundos sobre las cepas fúngicas aisladas, con respecto a potenciales
biotecnológicos.
- Determinar la actividad enzimática ligninolítica probando un medio de cultivo con otro tipo
de metales que induzcan la actividad enzimática, por ejemplo cadmio.
- Realizar pruebas que confirmen el pH característico para cada enzima, ya que este punto es
muy influyente en la actividad enzimática.
- Buscar la temperatura optima para la fermentación de los medios de cultivo de producción de
enzimas, y para la reacción enzimática.
- Determinar la biomasa de los medios de cultivo de producción de enzimas, por métodos
específicos.
- Profundizar estudios en los medios de cultivo e producción de enzimas, talvez con un diseño

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
32
factorial 33, esto con el fin de mejorar la producción de enzimas.
- Incrementar el tiempo de cultivo para determinar si existen diferencias en la cinética de estas
enzimas, ya que la actividad máxima en algunos casos se dio en los 28 días.
- Probar otro tipo de sustratos sintéticos, para la evaluación de enzimas ligninolíticas, por
ejemplo veratryl alcohol para LiP.
- Se requieren experimentos más precisos para definir la producción de enzimas.
- Por los resultados alcanzados se recomienda realizar la purificaron de dichas enzimas.
- Probar otros sustratos lignocelulosicos, para la producción de estas enzimas, estudiando su
composición química.
- Realizar pruebas de este tipo de enzimas para la producción de alcohol, la aplicación de
industrial como la fabricación de papel, en la industria farmacéutica para la modificación
químico enzimático de fármacos y finalmente para procesos de biorremediación por su
capacidad de oxidantes.
7. BIBLIOGRAFIA

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
33
1. Terrazas-Siles Enrique, Álvarez Teresa, Benoit Guieysse, & Bo. Mattiasson. Isolation and
characterization of a white rot fungus Bjerkandera sp. Strain capable of oxidizing
phenanthrene. Ed 1. Lund Media – Tryck, Lund University 2005; 20: 845-851.
2. Terrazas-Siles Enrique. 2005. Fungal Redox Enzymes Envolved in the oxidation of
organic pollutants. Ed 1. Lund Media – Tryck, Lund University 2005; 20: 10-23.
3. Leandro Papinutti Victor, Luis DIorio, Flavio Forchiassin. Degradación de la madera del
álamo por Fomes sclerodermeus: producción de enzimas ligninolíticas en aserrín de
álamo y cedro. Rev Iberoam Micol 2004; 20: 16-20.
4. Pelaez F, Martinez MJ, Martinez AT. Screening of 68 species of basidiomicetes for
enzymes involved in lignin degradation. Mycol Res 1999; 99: 37-42.
5. Kirt TK, Farrell RL. Enzymatic “combustion”: the microbial degradation of lignin.
Annu Rev Microbiol 1987; 41: 465-505.
6. Kirt TK, Connors WJ, Zeikus JG. Requirement for a growth substrate during lignin
decomposition by two woodroting fungi. Appl Environ Microbiol 1976; 32: 192-194.
7. Perez J, Jeffries TW. Roles of manganese and organic acid chelators in regulating lignin
degradation and biosynthesis of peroxidases by Phanerochaetechrysosporium. Appl
Environ Microbiol 1992; 58: 20402-2409.
8. Solís Sara. Hongos que eliminan contaminantes. Rev Instituto Tecnológico de Mérida
2005; 22: 10-11.
9. Centro de Investigaciones Biológicas de Madrid. El CSIC desarrolla alternativas
ecológicas para eliminar la lignina en la producción de papel. [email protected] 2004; 25:
1-3.
10. Guisado, G., López, M.J., Vargas-García, M.C., Suárez-Estrella, F. y Moreno, J.
Decoloración in vitro de colorantes industriales por ligninasas de

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
34
microorganismos aislados a partir de pilas de compostaje. Departamento de Biología
Aplicada, Universidad de Almería. 2005; 211: 4120-4125.
11. García Torres Angélica María, Rodrigo Gonzalo Torres Sáez. Producción de enzimas
lignolíticas por Basidiomycetes mediante la técnica de fermentación en sustrato sólido.
Rev Universidad Industrial de Santander. 2003; 18: 635-638
12. Dávila Gustavo, Rafael Vázquez-Duhalt. Enzimas Ligninolíticas fúngicas para fines
ambientales. Rev Universidad Nacional Autónoma de México. 2006; 108: 1000-1011.
13. Sánchez Oscar Julián, Carlos Ariel Cardona. Producción biotecnológica de alcohol
carburante I: obtención a partir de diferentes materias primas. Rev Interciencia. 2005;
30: 671-678.
14. Dirección Nacional de Promoción, Subsecretaria de Energía Eléctrica. Energías
renovables a partir de Biomasa. Rev Tecnología de la información Secretaria de energía.
2004; 23: 6-19.
15. Murrieta Dulce Maria, Gerardo Mata, Lourdes Iglesias. Cambios en la producción de
lacasa por el hongo Pleurotas Pulmonarius. Cultivado en pulpa de café en confrontación
con Tricoderma Viride Pers, un moho. Red de Rev Científicas de América Latina y el
Caribe, España y Portugal. 2002; 4: 47-52.
16. Lin SY, Lin IS. Lignin. In: Ullmann’s encyclopaedia of industrial chemistry. 2001; 15:
305-315.
17. Manzano Ana Margarita, Teresa León, Juan Argüelles. Hongos de la podredumbre
blanca con capacidad Ligninolítica y acción decolorante sobre el violeta cristal. Rev
Biología. 2004; 18: 123-127.
18. Campbell CK, Dagmar Schmidt Identification of Pathogenic Fungi
Public Health Laboratory Service. London. 1996; 12: 299-335.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
3
19. Lin, S.Y., Dence, C.W. Methods in lignin chemistry. Springer-Verlag, Berlin. 1999; 41:
465-505.
20. Fuentes Villagran, José Renan. Simulación y modelo matemático de la deslignificacion
selectiva de la madera por hongos blancos en ambiente natural. Rev Temuco Universidad
de la Frontera. 1991; 24: 465-487.
21. Fengel, D., Wengener, G. Chemistry, ultrastructure, reactions. Appl. Environ.
Microbiol. 1999; 23: 3397-3413.
22. Kay RM, Strasberg SM, Petrunka CN. Differential absorption of bile acids by lignins.
In: Inglett GE, Falkenhag I. Dietary fibres: Chemistry and nutrition. London: Academic Press,
1979; 57: 2300-2307.
23. C. Gómez-Dorado, M, Martínez-Salgado, D. Nieto-Mosquera. Estudio del efecto de dos
inductores y un protector enzimático sobre la actividad de las enzimas Mn-peroxidasa y
Lacasa producidas por Trametes versicolor y su efecto en la decoloración de efluentes de
la industria papelera. Rev Facultad de Ciencias. 2005; 10: 37-45.
24. Bial, Aristegui. El reino de los hongos. Rev Iberoam Micol. 2002; 8: 1-4.
25. Lillian Frioni. Procesos microbianos. Ed de la Fundación nacional de Río Cuarto
Argentina. 1999; 11: 723-736.
26. Popoff, Orlando. Reino fungi. Hipertextos del área de la Biología. 2006; 22: 24-29.
27. Enciclopedia Microsoft Encarta Online. Bolivia, consulado en septiembre12, 2006.
28. Poiting, S. B. Feasibility of bioremediation by white-rot fungi. Appl. Microbiol.
Biotechnology. 2001; 57: 20-33.
29. Álvarez Ma. Teresa., Terrazas S. Luis Enrique. Manual para la producción y valoración
de interés biotecnológico. 2002; 1: 1-2.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
36
30. Lehninger A., Nelson D., Principios de Bioquímica. Barcelona ediciones Omega. 2001;
2: 199-210.
31. Vargas Getulio. La magia de las enzimas. Rev énfasis en alimentación. 2006; 2: 75-82.
32. Rodríguez Rincón Francisco. Análisis molecular de la lacasa de Phanerochaete flavido-
alba: Caracterización del gen y regulación por fenoles y metales. Editorial de la
Universidad de Granada. 2006; 84: 338-385.
33. K Griselda., Guillen-Navarro, Facundo J., Márquez-Rocha, José E. Sánchez Vázquez.
Producción de biomasa y enzimas ligninolíticas por Pleurotas ostreatus en cultivo
sumergido. Rev Iberoam Micol.1998; 15: 302-306.
34. M. Martínez-Salgado, A. Pedrosa-Rodríguez, R. Rodríguez-Vázquez. Efecto de la
glucosa y nitratote amonio sobre las enzimas ligninolíticas producidas por Trametes
versicolor inmovilizado en espuma y la decoloración de un efluente papelero en un
biorreactor de lecho fluidazo. Revista de la Facultad de Ciencias. 2005; 10: 2, 27-36.
35. Mouso Nora, Leandro Papinutti, Flavio Forchiassin. Efecto combinado del cobre y pH
inicial del medio de cultivo sobre la producción de lacasa y manganeso peroxidasa por
Stereum hirsutum (Willd) Pers. Rev Iberoam Micol. 2003; 20: 176-178.
36. Alonso-Bozada S., M. Montiel_Gonzales, A. Tomasini, C. Sánchez. Comparación de las
enzimas lacasas de Pleurotas ostreatus producidas por fermentación sólida y sumergida.
Revista de la Universidad Autónoma de Tlaxcala. 2002; 12: 45-49.
37. Arana A., A. Téllez, S. Yague, e. Fermiñan, S. Moreno. Deslignificacion de pasta al
sulfato de Pinus radiata utilizando levaduras recombinantes. Centro de investigación
forestal Madrid. 2004; 18: 34-45.
38. Chávez-López C., J. F. Esparaza-García, Ma. E. Hidalgo-Lara, O: Loera-Corral, R.
Rodríguez-Vázquez. Producción de la enzima lacasa por el hongo Cladosporium

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
3
cladosporioides en presencia de fenantreno. Congreso de la Sociedad Cubana de
Bioingeniería Habana. 2005; 73: 158-163.
39. Andersson, B. E., Lundstedt, S., Tornberg, K., Schunurer, Y., Oberg, L.G., and
Mattiasson, B. Incomplete degradation of polycyclic aromatic hydrocarbons in soil
inoculated with wood-rotting fungi and their effect on the indigenous soil bacteria.
Environ Toxicol Chem. 2003; 22:1238-1243.
40. Maeztu Ramiro de. Biodegradación de Lignina y Compuestos Recalcitrantes. Centro de
Investigaciones Biologicas. 2002; 9: 2840-2845.
41. Barrasa, J. M., Martínez, A. T. Enzimas producidas por nuevos basidiomicetos aislados
en al sierra de Ayllón capaces de degradar colorantes industriales. Departamento de
Biología Vegetal, Universidad de Alcalá. 2003; 17: 372-376.
42. Addleman, K., Archibald, F. Kraft pulp bleaching and delignification by dikaryons
and monokaryons of Trametes versicolor. Appl. Environ. Microbiol. 1993; 59: 266-273.
43. Adler, E. Newer views of lignin formation.Tappi. 1957; 40: 294-301.
44. Aisemberg, G., Grorewold, E., Taccioli, G., Judewiez, N., Major, A. Transcript in the
response of Neurospora crassa to protein synthesis inhibition by cycloheximide. Exp.
Mycol.1989; 13: 121-128.
45. Akileswaran, L., Alic, M., Clark, E., Hornick, J., Gold, M. Isolation and transformation
of uracil autrophs of the lignin degrading bacidiomycete Phanerochaete chrysosporium.
Curr. Genet.1993; 23: 351-356.
46. Higuchi, T. Microbial degradation of lignin: Role of lignin peroxidase, manganese
peroxidase, and laccase. Proceedings of the Japan Academy, Series B: Physical and
Biological Sciences. 2004; 80: 204-214.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
38
47. Glenn, J., Morgan, Mayfield M., Kuwara M., Gold M. An extracellular requiring
enzyme preparation involved in lignin biodegradation by the white-rot bacidiomycete
Phanerochaete chrysosporium. Biochem. Biophys. Res. Común. 2002; 14: 1077-1083.
48. Tien, M., Kirk T. Lignin-degrading enzyme from the hymenomycete Phanerochaete
chrysosporium Burds. Science. 1983; 221: 661-663.
49. Schoemaker, H., Lundell T., Ataca A., Piotek K., The oxidation of veratryl alcohol,
dimeric lignin models and lignin by lignin peroxidase- the redox cycle revisited. FEMS
Microbiol. Rev. 1994; 13: 511-518.
50. Hatakka, A. Lignin-modifying enzymes from selected white-rot fungi: production and
role in lignin degradation. FEMS Microbiol. Rev. 1994; 13: 125-135.
51. Hammel, K., Tardote P., Moen M., Price J. A. Biomimetic oxidation of nonphenolic
lignin models by Mn (II): New observations on the oxidability of guaiacyl and syringyl
substructures. Asch. Biochem. Biophys. 1989; 270: 404-409.
52. Gianfreda, L., Xu, F., Bollag, J-M. Laccases: a useful group of oxidoreductive enzymes.
Bioremed. J. 1999; 3: 1-25.
53. Diamantidis, G., Effosse, A., Potier, P., Bally, R. Purification and characterization of
the first bacterial laccase in the rhizospheric bacterium Azospirillum lipoferum. Soil Biol.
Biochem. 2000; 32: 919-927.
54. Thurston, C. The structure and function of fungal Laccases. Microbiology. 1994; 140:
19-26.
55. Lewis, N., Yamamoto E. Lignin: occurrence, biogenesis and biodegradation. Ann. Rev.
Plant. Physiol. Plant. Mol. Biol. 1990; 92: 455-496.
56. Bollag, J-M., Leonowiez A. Comparative studies of extracellular fungal Laccases.
Appl. Environ. Microbiol. 1984; 48: 849-854.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
39
57. D´Souza, T., Dass, S., Rasooly, A., C. Electrophoretic karyotyping of the lignin-
degrading bacidiomycete Phanerochaete chrysosporium. Mol. Microbiol. 1993; 8: 803-807.
58. Shin, W., Sundaran U., Cole J., Zhang M., Hedman B., Hodgson K., Solomon E.
Chemical and spectroscopic definition of the peroxide level intermediate in the
multicopper oxidases. Relevance to the catalytic mechanism of dioxygen reduction of
water. J. Am. Chem. Society. 1996; 118: 3202-3215.
59. Hess J., Leitner C., Galhaup C. Enhanced formation of extracellular laccase activity by
the white- rot fungus Trametes multicolor. Appl Biochem Biotechnol. 2002; 98-100: 229-
241.
60. Demir G. Degradation of toluene and benzene by Trametes versicolor. J.Environ Biol.
2004; 25:19-25.
61. Clemente A.R., Anazawa, T.A., and Durrant L. R. Biodegradation of polycyclic
aromatic hydrocarbon by soil fungi. Braz J Microbiol. 2001; 32: 255-261.
62. Novtny, C., Erbanova, P., Sasek, V., Kubatova, A., Cajthaml, T., Lang, E., Krahl, J.,
Zadrazil,F. Extracellular oxidative enzyme production and PAH removal in soil by
exploratory mycelium of white rot fungi. Biodegradation. 1999; 10:159-168.
63. Kubatova, A., Erbanova, P., Eichlerova, I., Homolka, L., Nerud, F., and Sasek, V. PCB
congener selective biodegradation by the white-rot fungus Pleurotus ostreatus in
contaminated soil. Chemosphere. 2001; 43: 207-215.
64. Baudette, L. A., Ward, O. P., Pickard, M. A., and Fedorak P.M. Low surfactant
concentration increases fungal mineralization of a polychlorinated biphenyl congener
but has no effect on overall metabolism. Lett Appl Microbiol. 2000; 30:155.
65. Hiratsuka, N., Wariishi, H., and Tanaka, H. Degradation of biphenyl ether herbicides by
the lignin-degrading bacidiomycete Coriolus versicolor. Appl Microbiol Biotechnol. 2001;

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
40
57:563-571.
66. Bending, G. D., Friloux, M., and Walker, A. Degradation of contrasting pesticides by
white rot fungi and its relationship with ligninolytin potential. FEMS Microbiol Lett.
2002; 212: 59-63
67. Kahraman S., Yesilada, O. Decolorization and bioremediation of molasses wastewater
by white-rot fungi in a semi-solid-state condition. Folia Microbiol (Praha). 2003; 48: 525-
528.
68. Wu, J., Xiao, Y. Z., and Yu, H.Q. Degradation of lignin in pulp mill wastewaters by
white-rot fungi on biofilm. Bioresour Technol. 2005; 12:1357-1363.
69. Barnett, H. L., Hunter, B. Illustraded Genera of imperfect fungi. 1972; 3.
70. Rodrigues Rincon Ramiro. Análisis molecular de la lacasa de Phanerochaete flavido-
alba: Caracterización del gen y regulación por fenoles y metales. Ed. De la Universidad
Granada España. 2006; 4: 1026-2006.
71. Álvarez, E. Principales indicadores de la masa sacarificada de aserrín de pino y su
efecto en la alimentación de pollos de ceba, Resúmenes del Primer Taller Internacional
Bioforest. Rev. Pinar del Río.2003
72. Jolkin, Y.I. Tecnología de las producciones hidrolíticas. Editorial Lesnaya Prom,
Moscú. 1989:495 p.
73. Crez, R. Utilización de aserrín como aditivo para mejorar la dinámica de los
elementos nutritivos en el suelo. Rev. 6 Congreso de las ciencias del suelo,
Chile.pp.1996:173-176.
74. Ramírez M. Instructivo técnico funcional de la empresa Agroindustrial ICU S.A.
Centro Nacional de Promoción de la Pequeña Industria y Artesanía. Quito. 1993: 76 p.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
4
75. Prescott, Lasing M. Microbiología. 1999; 4: 539-555.
76. Maganhotto, Celia Maria de Souza Silva. Producao de enzimas Ligninolíticas por
fungos isolados de solos sob cultivo de arroz irrigado.Jaguariuna, SP. 2004; 56:1516-1524.
77. Levin L, Forchiassin F, Ramos AM. Cooper induction of lignin-modifying enzymes in
the white-rot fungus Trametes trogii. Mycologia. 2002; 377-383.
78. Baldrian P, Gabriel J; Nereud F. Effect of cadmium on the ligninolytic activity oh
Stereum hirsutum and Phenerochaete chrysosporium. Folia Microbiol 1996; 41: 363-367.
79. Galliano h, Gas G, Seris JL, Boudet AM. Lignin degradation by Rigidoporus lignosus
involves synergistic action of two oxidizing enzymes: Mn peroxidase and laccase. Enzyme
Microb Technol. 1991; 13: 478-482.
80. Kapich A.N., Prior B.A., Botha A. Effect of lignocellulose-containing substrates on
production of ligninolytic peroxidases in submerged cultures of Phanerochaete
chrysosporium ME-446. Enzyme and microbial Technology. 2004; 34: 187-195.
81. Moreira Neto Sergio Luiz. Enzimas Ligninolíticas produzidas por Psilocybe castanella
CCB444 em solo contaminado com hexaclorobenceno. Instituto de Botanica da Secretaria
de Estado do Meio Ambiente.2006; 110: 376-398.
82. Papinutti, V.L., Diorio, L.A., Forchiassin, F. Production of laccase and manganese
peroxidase by Fomes sclerodermes grow on wheat bran. Journal of Industrial Microbiology
and Biotechnology. 2001; 30: 157-160.
83. Baldrian. P., Valaskova, V., Merhautova, V., Gabriel, P. Degradation of lignocellulose by
Pleurtus ostreatus in the presence of copper, manganeso, lead and zinc. Research in
Microbiology. 2005; 156: 670-676.
84. Harper, H.A., Rodwell, V.W., Mayes, P.A. Manual de Química Fisiológica. 1982; 5:
736.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
42
85. Leatham G.F., Extracellular enzymes produced by the cultivated mushroom Lentinus
edodes during degradation of lignocellulosic medium. Appl Environ Microbiol. 1985; 50:
859-867.
86. Bourbonnais R, Paice MG, Freiermuth B, Bodie E, Bomemen S. Reactivities of various
mediators and Laccases with kraft pulp and lignin model compounds. Appl Environ
Microbiol. 1997; 63: 4627-4632.
87. Soares G.M., de Amorim M.T., Costa Ferreira M. Use of laccase together with redox
mediators to decolourize Remazol Brilliant Blue R. J Biotechnol. 2001; 89: 123-129.
88. Muñoz C., Guillen F., Martinez A.T., Induction and characterization of laccase in the
ligninolytic fungus Pleurotus eryngii. Curr Microbiol. 1997; 34: 1-5.
89. Furushima Y. Okada K., Kawai G. Efficient production of mycelium of Lentinus edodes
by a continuous culture. Murhroom Science. 1991; 13: 721-725.
90 Baldrian, P. Gabriel J. Cooper and cadmium increase laccase activity in Pleurotus
ostreatus. FEMS Microbiol. Lett. 2002; 206: 69-74.
91 Moldes D., Gallego P. Grape seeds: the best lignocellulosic waste to produce laccase by
solid state cultures of Trametes hirsute. Biotechnology Letters. 2003; 25: 491-495.
92. Chávez L., Esparza F., García E. Producción de la enzima lacasa por el hongo
Cladosporium cladosporioides en presencia de fenantreno. Sociedad Cubana de
Bioingeniería. 2005; 73; 76-89.
93. Michniewiez Anna, Ullrich Rene. The white-rot fungus Cerrena unicolor strain 137
produces two laccase isoforms with different physico-chemical and catalytic properties.
Appl Microbiol Biotechnol. 2006; 69: 682-688.
94. Heinfling, A. Transformation of industrial dyes by manganeso peroxidases from
Bjerkandera adusta and Pleurotas eryngii in a manganeso-undependent reaction

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
43
in Applied and Environmental Microbiology .Rev Applied Microbiol 1998; 64: 2788-2793.
95. Koh, S. Plant terpenes and lignin as natural cosubstrates in biodegradation of
polychlorinated bipheyls (PCBs) and polyeyelic aromatic hydrocarbons (HAPs).
Documento de la universidad de la marina de Korea. Korea Pp1-3.
96. Cerniglia C. Fungal metabolism of polyeyelic aromatic hydrocarbons: past, present
and future applications in bioremediation. Journal of Industrial Microbiology &
Biotechnology. 1997; 19: 234-333.
97. Society for Microbiology. Introduction in Journal of Industrial Microbiology &
Biotechnology. 1997; 18:73-74.
98. Ulfig. K. Microbiological aspects of bioremediation. Institute for the ecology of
industrial areas. EE. UU.1995; 76: 765-769.
99. Vidale, M. Bioremediation. An overview in Pure Applied Chemistry. 2001; 7:1163-1172.
100. Viñas, M. Biodegradation of crude oil by three microbial consortia of different
origins and metabolic capabilities. Journal of Industrial Microbiology & Biotechnology.
2002; 28: 252-260.
101. Beaudette, L. Comparison of gas chromatography and mineralization experiments
for measuring loss of selected polychlorinated biphenyl congeners in culture of white rot
fungi. Applied and Environmental Microbiology. 1998; 64:2020-2025.
102. Alvo P, Belkacemi K. Enzymatic saccharification of milled timothy and alfalfa
(Medicago sativa L.). Bio resource Technol. 1997; 61: 185-198
103. Herrera-Mora, J. Rosas Acosta. Estudio preliminar de la producción de enzimas
Ligninolíticas por los hongos Phanerochaete chysosporium, Trametes versicolor y
Pleurotas ostreatus para el tratamiento de efluentes de la industria papelera.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
44
Microbiología Industrial. Bogotá, Colombia. Pontificada Universidad Javeriana, 2003; 151:
342-346.
104. Hawgood N, Evans S. Enhanced ethanol production in multiple batch fermentations
with an auto-flocculating yeast strain. Biomass. 1985; 7: 255-273.
105. Wyman CE, Spindler DD, Grohmann K. Simultaneous saccharification and
fermentation of several lignocellulosic feedstock’s to fuel ethanol. Biomasa Bioenerg.1992;
3: 301-307.
106. Deguchi. T. 1998. Purification and characterization of a Nylon degradation enzyme.
Applied and Environmental Microbiology 64: 1366-1371.
107. Seradati, M. Transformation of high concentrations of chlorosphenols by the white-
rot bacidiomycete Trametes versicolor immobilized on nylon. Electronic Journal of
Biotechnology. 2003; 6: 106-114.
108. Eggert, C. The ligninolytic system of the white rot fungus Pycnoporus cinnabarinus:
Purification and characterization of the laccase. Applied and Environmental Microbiology.
1996; 62:1151-1158.
109. Kumari, M et al. 2000, Secretion of lignin peroxidase by Pencillium citrinum,
fusarium oxysporum and Aspergillus terreus. Indian Journal Experimental Biology. 2000;
40: 802-806.

Instituto de Investigaciones Fármaco Bioquímicas Planta del Izozog Guirakillo IIFB
4