universidad autÓnoma chapingo · rendimiento en canal y propiedades fisico-quimicas de la carne...
TRANSCRIPT

DEPARTAMENTO DE ENSEÑANZA, INVESTIGACIÓN Y
SERVICIO EN ZOOTECNIA
RENDIMIENTO EN CANAL Y PROPIEDADES FISICO-QUIMICAS DE LA CARNE DEL GUAJOLOTE AUTÓCTONO
(Meleagris gallopavo Linn)
QUE COMO REQUISITO PARCIAL PARA OBTENER EL TITULO DE:
INGENIERO AGRÓNOMO ESPECIALISTA EN ZOOTECNIA
PRESENTA:
URIOSTEGUI REYNOSO ERIC
Chapingo, Texcoco, Edo. de México, Septiembre 2009
TESIS PROFESIONAL
UNIVERSIDAD AUTÓNOMA CHAPINGO

AGRADECIMIENTOS
A dios por la oportunidad de llegar a este momento tan importante en mi vida y permitir
compartirlo con mis seres queridos, por estar presente en cada paso que doy, por guiarme y
llevarme de la mano en todas mis decisiones.
A la Universidad Autónoma Chapingo y al departamento de Zootecnia por la oportunidad de
formarme profesionalmente.
A la Ph. D. Elvia López Pérez por la dirección, apoyo, asesoría para la realización de la presente
investigación, además de los consejos tan acertados en esta etapa.
Al Dr. Arturo Pro Martínez por su valiosa colaboración en la dirección, por las observaciones
pertinentes, por su apoyo, sugerencias y disponibilidad para la elaboración de esta tesis.
Al comité revisor: M.C. Ma. Del Rosario Campos Hernández, Ph. D. Marta Elva Ramírez Guzmán,
Dr. José Joel E. Corrales García por su aportación en mi formación profesional y por el tiempo
prestado para la revisión y aprobación de este documento.
A toda mi familia, porque siempre están presentes brindando su apoyo incondicional y a todas
las personas que en algún momento me brindaron su apoyo y comprensión.
A mis papas y hermanos por creer en mí, por que siempre están a mi lado, por que puedo contar
con ustedes, por todos los esfuerzos que han hecho por mí, por su apoyo para hacer mis sueños
realidad.
A los profesores del Departamento de Zootecnia por dejar en mi un poquito de sus conocimientos
que sin duda alguna son una gran herramienta para alcanzar un buen lugar en el mundo
laboral.
A todos mis amigos y compañeros que hicieron de mis estudios una feliz estancia en la
Universidad Autónoma Chapingo.
A todos, Gracias!!! Por hacer de mi una persona diferente, por ayudarme y comprenderme,
corregirme y criticarme, pero sobretodo por no dejarme solo…
i

DEDICATORIA
A mis padres Alfredo y Martha, a quien respeto, admiro y quiero con todo mi ser! Por que siempre
me han dado su amor, porque confían en mí y porque me han enseñado que el trabajo y esfuerzo
son la única manera de salir adelante, por ser el pilar que me motivo para nunca rendirme ante
los obstáculos que hubiesen en el camino, por todos los consejos que me dan en el momento mas
adecuado, por su cariño comprensión y apoyo para lograr lo que me propongo y que no haya
motivo alguno para dejar de luchar, porque gracias a ustedes he llegado a ser quien soy y no hay
palabras suficientes para agradecer todo lo que han hecho por mi, por ser los mejores papás, por
ser mi ejemplo a seguir…
A mis hermanos: Giovas, Negro y Yuli, por que me contagian con sus alegrías y entusiasmo para
hacer las cosas y echarle ganas a pesar de todo, por hacer la vida mas “light” aun en momentos
críticos, por el apoyo que he recibido de ustedes en todos los sentidos y en todo momento, por ser
mis confidentes, por compartir mas de un millón de experiencias buenas y malas, por que son mis
amigos de toda la vida, porque me dan los consejos mas sensatos en los momentos traumantes…
Gracias por su amor y respeto, por ser un punto de apoyo, ternura y comprensión… son lo mejor y
los quiero mucho!!!.
A mis abuelitas, Clementina y Bruna por la confianza depositada en mí, por que han seguido
paso a paso mi formación, porque han dejado en mí muchas enseñanzas que me han ayudado y
me ayudaran a salir adelante, por que han estado conmigo a lo largo de mi vida y siempre será
así…
A mis tíos y primos, porque siempre están presentes en mi vida, porque de ustedes he aprendido
mucho, por los juegos y risas que hemos compartido, por ser una parte fundamental en mi vida…
A mis cuñadas, Fiorela, Karla y mi sobrina Kimberly, que han estado presentes en parte de mi
vida y me han brindado su apoyo.
A mis entrañables amigos del prope 6: Wero, Ana Laura, Malaquías, Beto, Rasta, Chavero, Cuaco,
Jorge, Adair, con ustedes compartí mi primer gran año en esta Universidad, mis primeras
travesías en los viajes de estudio, entre muchas otras aventuras y experiencias que siempre
recordaré. Gracias por seguir en contacto…
A mis amigos: Damaris, Cocho, Caleb, Nengo, Cremas, Parka, teco, Mordelón, Brujo, Tortas. A mis
amigos zootecnistas: Rubén, tun tun, loco, Karim, Chano, Abelardo, Román, Urbano, Zoyejo,
Carlo y a todos los amigos del gremio mejor conocido como “útero”, por brindarme su amistad
desinteresada, en los últimos meses de mi estancia en Chapingo. Gracias amigos, al fin!!!
ii

ÍNDICE GENERAL
Página
AGRADECIMIENTOS……………………………………………………………………...i
DEDICATORIA…………………………………………………………………………….ii
INDICE GENERAL………………………………………………………………………..iii
INDICE DE CUADROS…………………………………………………………………...vi
INDICE DE GRAFICAS……………………………………………………………….... vii
RESUMEN………………………………………………………………………………..viii
SUMMARY….……………………………………………………………………….……..ix
1. INTRODUCCIÓN...……………………………………………………………………...0
2. Revisión de literatura………………………………………………………………….1
2.1. Aspectos generales sobre rendimiento de la canal……………………………..2
2.2. Factores relacionados con el rendimiento en canal………………….……........2
2.2.1. Condiciones ambientales……………………………………………….……2
2.2.2. Conversión alimenticia….……………………………………………………2
2.2.3. Alimentación…..……………..………..………………………………………3
2.2.4. Edad y peso………………………………………………..………………….3
2.2.5. Sexo…………………………………………………………..………………..4
2.2.6. Estirpe……………….……………………………..…………………………..4
2.2.7. Pérdidas ocasionadas por el sacrificio, eviscerado y desplumado…...…5
2.3. Factores que afectan la calidad de la canal……………………………………...5
iii

2.3.1. Rigor mortis…………………………………………..………………………..6
2.3.2. Carne………….…………………………………………………………….….6
2.3.3. Grasa………………..…………………….………………………………..….7
2.3.4. Color…………………...…………………………………………………….…7
2.3.5. Temperatura y pH……………………..………………………….………..…8
2.3.6. Sabor………………...…………………………………………..………….….9
2.3.7. Congelación…………………...……………………………………………....9
3. OBJETIVOS…………...……………………………………………………………….10
3.1. HIPOTESIS……...…………………………………………………………………...10
4. MATERIALES Y METODOS………...……………………………………………….11
4.1. Localización……………...………………………………………………….……..11
4.2. Animales utilizados……………………………………………………………......11
4.3. Pesaje.……………………………………………………………………………...11
4.4. Sacrificio……………………...………………………………………………….…12
4.5. Peso de la canal caliente……………………………………...……………….…13
4.6. Despiezado………………...……………………………………………………....13
4.7. Lavado e identificación……………………………………………………………13
4.8. Inmersión en hielo………………………………………………………………....13
4.9. Registro de pH y temperatura…………...……………………………………….14
4.10. Peso del animal despiezado……………………………………………………14
4.11. Peso de la canal en frio……………………………………………………….…14
iv

4.12. Registro de color…...…………………………………………………………….14
5. VARIABLES DE RESPUESTA………………………………………………………15
5.1. Rendimiento en canal……………………………………………………………..15
5.2. Determinación de pH………...……………………………………………………15
5.3. Determinación de temperatura……...……………………………………….…..15
5.4. Determinación de color……...…………………………………………………....15
6. DISEÑO EXPERIMENTAL……………………………………………………………16
7. RESULTADOS Y DISCUSIÓN……..………………………………..………………19
7.1. Efecto del sexo, edad y peso de guajolotes autóctonos en el rendimiento
de la canal……………………………………………..……………………………19
7.2. Peso de la canal despiezada…………...………………………………………..21
7.3. pH y temperatura en la canal despiezada……...…..………………………..…23
7.4. Efecto del sexo y la edad en el color de la canal del guajolote
autóctono……………………………………………………………….…………...27
7.5. Análisis de varianza univariado………………………………………………….29
7.6. Análisis de varianza multivariado………………………………………………..31
8. CONCLUSIONES……………………………………………………………………...33
9. LITERATURA CITADA……………………………………………………………….34
10. ANEXOS………...………………………………………………………………….…38
v

INDICE DE CUADROS
Cuadro 1. Efecto del sexo, edad y peso de guajolotes autóctonos en el rendimiento de la canal a 7, 12 y 15 meses de edad…………………………………………………………..…...……….. 19
Cuadro 2. Peso de la canal entera y despiezada de guajolote autóctono
hembra y macho a tres diferentes edades (expresada en meses y kg)……………………………………………………………...………….. 22
Cuadro 3. pH promedio de pechuga, muslo, pierna y grasa en guajolotes autóctonos machos.…….…………………………………………….… 23
Cuadro 4. pH de pechuga, muslo, pierna y grasa de hembras a diferente edad
(7, 12 y 15 meses) y horas post mortem...…….………..………….... 24
Cuadro 5. Efecto del sexo de los guajolotes autóctonos en la brillantes (L*),
intensidad (Croma) y ángulo de matiz (Hue °)...……………………. 27
Cuadro 6. Efecto de la edad de los guajolotes autóctonos en la brillantes (L*),
intensidad (Croma) y ángulo de matiz (Hue °)...……………………. 27
vi

INDICE DE GRAFICAS
Gráfica 1. Promedio de pH y temperatura de hembras a diferentes horas post mortem (0h, 5h, 18h, 24h, 48h y 7 días).……………………...……….. 26
Gráfica 2. Promedio de pH y temperatura de machos a diferentes horas post mortem (0, 5, 18, 24, 48 y 7 días)……..……………………..….………. 26
vii

Rendimiento en canal y propiedades físico-químicas de la carne del guajolote autóctono (Meleagris gallopavo Linn)
RESUMEN
El presente estudio se realizó con el objetivo de determinar el rendimiento en
canal y las propiedades físico – químicas de la carne, en particular pH, temperatura
y color, que afectan la calidad de la canal de los guajolotes autóctonos mejor
conocidos como guajolotes criollos. Se utilizaron 18 animales de los cuales 9
fueron hembras y 9 machos a tres diferentes edades (7, 12 y 15 meses), los cuales
se dividieron en tres grupos: animales pequeños (7 meses), animales jóvenes (12
meses) y animales viejos (15 meses). El rendimiento y la calidad de la canal de los
guajolotes se midieron en términos de género, edad, peso, pH, temperatura y color.
Estas mediciones se realizaron tanto en la canal entera como despiezada a
diferentes horas post mortem.
Los resultados indicaron que la edad, el peso y el sexo de los animales
afectó (P<0.05) el rendimiento de la canal caliente y fría. Los pesos más altos en
las tres diferentes edades, así como el rendimiento de la canal al sacrificio se
encontraron en los guajolotes machos a los 15 meses de edad. En el caso de las
hembras, el mayor rendimiento se encontró a los 12 meses de edad. El color de la
canal de hembras y machos a 15 meses de edad presentó diferencias (P<0.05) con
respecto a machos pequeños y jóvenes quienes presentaron una canal más
oscura. Con respecto al pH, se observó que las hembras presentaron mayor pH en
las diferentes piezas de la canal, mostrando diferencias (P<0.05) con respecto a
machos, sin embargo; para temperatura no se encontraron diferencias (P>0.05).
Palabras clave: rendimiento, calidad, canal, pH y color.
viii

ix
Carcass yield and meat´s physical-chemical properties of native mexican turkey (Meleagris gallopavo Linn)
Summary
This study was carried out with the aim to determine carcass yield and
meat´s physical and chemical properties such as pH, temperature and color of
mexican native turkey. We used 18 animals; nine of them were females and 9
males. Animals were arranged in to three groups according to its age: small animals
(7 months), young animals (12 months) and old animals (15 months). Yield carcass
were measured in terms of gender, age and weight. To determine meat quality we
recorded pH, color and temperature.
Results indicated age, weight and animal sex affected (P<0.05) carcass
yield. The highest weight and best carcass yield were found in males at the age of
15 months. Males always weight more than females. In the case of females,
carcass yield was higher at 12 months. Animal sex and age affected (P<0.05) meat
color. Young animals presented darker carcass compared with oldest animals (15
months of age). Respect to pH, females had higher pH compared with males (P
<0.05). There was not differences respect to carcass temperature.
Keywords: Yield, quality, carcass, pH and color.

1 INTRODUCCION
Debido al aumento de la demanda de carne de buena calidad por parte de la
población mexicana y al incremento de los costos de los granos que seguramente
incrementan los costos de producción es necesaria la búsqueda de alternativas que
disminuyan tales costos y suministren a la población, principalmente de origen
rural, proteína de origen animal. Entre estas alternativas esta la cría y explotación
de guajolotes autóctonos mejor conocidos como guajolotes criollos cuya carne
presenta niveles de proteína de hasta 23%, y baja cantidad de grasa. A pesar de
estas cualidades esta especie es poco estudiada y poco ingerida, su consumo es
principalmente en eventos sociales y eventos religiosos, sin embargo, su carne,
puede utilizarse en la elaboración de diversos productos cárnicos (aun de manera
rústica) tales como: jamón, salchicha, mortadela, longaniza, etc. Lo anterior,
representa una alternativa para el consumo humano, principalmente para la
alimentación de poblaciones rurales que crían estos animales a nivel de traspatio.
Una ventaja que tienen los guajolotes autóctonos es que se alimentan de diversos
productos, entre ellos los desperdicios de cocina. Además de que pasan la mayor
parte del día pastoreando, lo que puede disminuir los costos de producción ya que
prácticamente su consumo de granos es muy bajo. Además, son resistentes a
algunas enfermedades exceptuando viruela y newcastle, cuya gravedad puede
causar la muerte total de la parvada, sobre todo cuando los animales son
pequeños.
El propósito de esta investigación fue determinar el rendimiento de la canal y
las propiedades físico - químicas de la carne del guajolote autóctono, a tres
diferentes edades (7, 12 y 15 meses).
0

2. REVISION DE LITERATURA
2.1. Aspectos generales sobre el rendimiento en canal
La canal es la unidad de mayor importancia para determinar el rendimiento
en la producción de carne, ya que establece el valor económico de un animal. En
otras palabras, es el producto final comestible (Berg y Simms, 1960).
El rendimiento se expresa como la relación que existe entre el peso de la canal y el
peso vivo del animal, expresado en porcentaje (Forrest et al., 1975).
Rendimiento de la canal = Peso de canal / peso vivo* 100 (%)
El rendimiento en canal es de gran importancia en estudios sobre producción
de carne y varía dependiendo de la especie, como lo indica Forrest et al. (1975)
quienes encontraron un rendimiento en canal de 58-62% en bovinos, 49-52% en
ovinos, 69-73% en porcinos y 55-60 % en conejos.
En el caso del guajolote criollo el rendimiento en canal no es de uso
cotidiano, esto se debe quizá a que su producción se destina principalmente al
autoconsumo, en eventos sociales principalmente de tipo familiar. La crianza de
esta especie se realiza a nivel de traspatio, poca atención se pone a su
alimentación y mucho menos al rendimiento y calidad de la canal, lo que trae como
consecuencia que trabajos científicos sobre la evaluación y composición de la
canal sean escasos o nulos.
La carne de guajolote, constituye un alimento magro, que contiene un bajo
contenido en grasas saturadas, y un buen contenido en grasas insaturadas
(monoinsaturadas y polinsaturadas). La mayor parte de la grasa del guajolote se
encuentra directamente bajo la piel y se puede retirar fácilmente. El contenido de
1

proteína de esta especie es elevado (20 a 25%). Por otra parte, la carne de
guajolote constituye una excelente fuente de vitaminas, además de que contiene
aminoácidos esenciales que ayudan a la actividad cerebral de quien la consume
(Battula et al., 2008).
2.2. FACTORES RELACIONADOS CON EL RENDIMIENTO DE LA CANAL
2.2.1. Condiciones ambientales
Para alcanzar un buen potencial genético, las aves deben contar con
óptimas condiciones ambientales. Cualquier descuido como una alta densidad de
animales, puede ocasionar disminución en el rendimiento de la canal, debido a que
la temperatura en la caseta incrementa y el flujo de aire disminuye lo cual ocasiona
un aumento en la cantidad de amoníaco y una disminución en el consumo de agua
y alimento, ya que el acceso a estos se ve obstaculizado, lo que trae como
consecuencia una disminución en la tasa de crecimiento, eficiencia alimenticia y
rendimiento de la misma (Puron et al., 1995).
2.2.2. Conversión alimenticia
Según Buddiger y Albers (2000), el principal objetivo de la cría de aves en
los últimos 50 años, ha sido mejorar la tasa de crecimiento, conversión alimenticia y
rendimiento en canal. Estos autores señalan que la conversión alimenticia es muy
importante para determinar el rendimiento en canal y que en el transcurso de
décadas la conversión alimenticia para machos y hembras ha disminuido en 0.49 y
0.51 respectivamente. En general, la industria avícola evalúa el rendimiento de la
canal en pollo de engorda basándose en la conversión alimenticia y ganancia de
peso (Ewart, 1993; Watts y Kennett, 1995).
2

Para muchos productores, el rendimiento de la canal se considera tan
importante como la tasa de crecimiento y la conversión alimenticia, ya que a menor
conversión alimenticia mayor peso de la canal (Leclercq, 1998).
2.2.3. Alimentación
La alimentación constituye uno de los aspectos económicos más importantes
de cualquier explotación avícola, los costos por alimentación representan más del
65% del costo de producción, por lo que es indispensable suministrar a las aves,
alimentos que con un mínimo de gastos, alcancen un máximo rendimiento
(Barbado, 2004). En la producción de carne de guajolote existen diferentes tipos de
mercados que demandan determinadas características que en ocasiones no es
posible satisfacer con una dieta basada exclusivamente en forrajes, por lo que la
calidad de la carne que un mercado exige, obliga al productor a utilizar diferentes
tipos de alimentación (Rubio, 2003). A este respecto, Berg y Simms (1960)
observaron que conforme se incrementaba la cantidad de proteína en la dieta en
pavos de engorda, las canales tenían más carne y menos grasa, por otra parte
Plavnik y Hurwitz (1988) mencionan que la buena presentación del alimento y un
granulo de calidad, mejoran el peso de los pavos al sacrificio de un 5 a un 10%
comparado con el mismo alimento en harina.
En el caso de guajolotes mejorados las líneas actuales están caracterizadas
por su amplia pechuga y un alto porcentaje de músculo, por lo que se considera
que la relación proteína - energía es mayor que en pollos durante las primeras
semanas de vida. Por lo anterior, es necesario un programa de alimentación
adecuado, especialmente durante los primeros días de edad, donde la
presentación del alimento y calidad del granulo repercutirán en gran medida sobre
el rendimiento en canal del guajolote (Nixey, 1988).
2.2.4. Edad y peso
La edad y el peso de las aves son de gran importancia para determinar el
rendimiento en canal de los pavos, en un estudio realizado con pavos de diferentes
edades, Sell (1988) encontró que a mayor edad había un aumento progresivo en el
3

peso vivo de los animales. Por otra parte Scott et al. (1982) observaron que la edad
tiene gran importancia ya que dependiendo de la edad, será la cantidad de
aminoácidos que se adicione, esto principalmente cuando se busca maximizar el
rendimiento de pechuga, pierna y muslo, que son las piezas de interés del
guajolote.
Tulloh (1964), indicó que el peso de la canal repercute en gran medida en el
despiezado del ave. De igual manera, el peso al sacrificio repercutirá en el
rendimiento de la canal, a mayor peso al sacrificio el rendimiento en canal será
mayor.
2.2.5. Sexo
Los machos y hembras difieren en la velocidad del crecimiento y
composición de la canal. El crecimiento de las hembras es lento comparado con el
de machos (Cross et al., 1981). En la cría de aves por sexos, se logra un mayor
crecimiento en los machos, por lo que éstos se pueden vender una semana antes
que las hembras, obteniéndose uniformidad en peso de la parvada hasta del 70%,
a diferencia del 30% obtenido en la cría en conjunto de guajolotes de sexos
diferentes. Algunos criadores desarrollan líneas de hembras, otros producen líneas
de machos para carne, los cuales tienen abundante carne, gran tamaño y rápido
crecimiento (North, 1986).
2.2.6. Estirpe La estirpe del ave ocupa un papel muy importante para determinar el
rendimiento en la canal del guajolote, ya que dependiendo de ésta, el animal
presentará mayor o menor conformación muscular y cantidad de grasa. La elección
de la línea, se basa en el objetivo de producción ya sea para huevo, carne o para
ambos; buscando por lo general en la medida posible, que los animales se adapten
a las condiciones ambientales (Barbado, 2004).
En el transcurso de las décadas se han desarrollado diferentes estirpes
mejoradas de pavos lo que ha dado como resultado trabajar con líneas
especializadas de acuerdo a la finalidad o al objetivo deseado (peso corporal,
4

rendimiento de la carne y eficiencia alimenticia), y esto ha beneficiado al productor
ya que los parámetros productivos han incrementando paulatinamente (Buddiger y
Albers, 2000).
2.2.7. Pérdidas ocasionadas por el sacrificio, eviscerado y desplumado
Las aves que han alcanzado el peso de mercado mediante el consumo de
raciones suficientemente ricas en energía para producir una buena cobertura
adiposa, experimentarán menores pérdidas al ser evisceradas que las aves que no
lo alcanzan. En el caso de pollos de engorda, los rendimientos porcentuales
medios, según el peso congelado de las canales preparadas para cocinar, incluidos
los despojos tales como molleja e hígado, son los siguientes: pechuga, 24%;
piernas, 15 %; muslos, 16%; alas, 13%; dorso, 17%; cuello7.5%, y despojos, 7.5%
(Card y Nesheim 1968).
La sangre y las plumas constituyen las pérdidas ocasionadas por el sacrificio
y el desplumado. La cantidad de sangre perdida se aproxima al 4% del peso vivo.
En el caso de los pollos de engorda, esto representa aproximadamente 48g para
un pollo de engorda que pese 1200 g. La pérdida de peso promedio constituye el
4% del peso vivo y las perdidas por desplume son mayores en hembras que en
machos. En general, las pérdidas producidas por la evisceración dependen de la
gordura y grado de acabado de las canales y, al igual que las pérdidas producidas
por el desplumado, las pérdidas por evisceración tienden a ser mayores en las
aves más pequeñas que en las más pesadas, el rendimiento disminuye en exceso
cuando cada cabeza lleva un trozo excesivo de cuello.
2.3. FACTORES QUE AFECTAN LA CALIDAD DE LA CANAL
La calidad de la canal se refiere a la capacidad para satisfacer las
expectativas de los consumidores. Las propiedades fisicoquímicas tales como
textura, color, sabor de la carne, y propiedades como capacidad de retención de
5

agua, emulsificación y fuerza de cohesión, conjugadas entre sí componen la
calidad sensorial (Ranken, 2003).
2.3.1. Rigor mortis
El rigor mortis es un factor que afecta la calidad de la canal inmediatamente
después de la muerte, se produce en circunstancias ordinarias, un estado de
relajación y flacidez de todos los músculos del cuerpo, que al cabo de cierto
tiempo, aunque en general breve, se inicia un proceso lento de contractura
muscular, ocurrido cuando el ATP (adenosin trifosfato) en el músculo se convierte
en ADP (adenosin difosfato) y AMP (adenosin monofosfato) por la liberación de
energía y es a lo que se le conoce como rigor mortis, el cual finalizará después de
un tiempo, cuando nuevamente se relajen los músculos. El rigor mortis varía de
minutos a horas dependiendo de la especie animal, en el caso del pavo es de 0.5 a
2 horas, terminando de 6 a 24 horas post mortem (Ranken, 2003).
2.3.2. Carne
El músculo o carne magra se compone de la siguiente forma:
• Proteína miofibrilar en forma de múltiples fibrillas, fibras y haces de fibras.
• 10%
• Colágeno y elastina. • 2%
• sarcoplasma compuesto de agua (75%), proteína
sarcosplasmatica (6%) y otras sustancias solubles, tales como mioglobina (color rojo), sales, vitaminas, etc.
• 84.5%
• Grasa, tendones, nervios, vasos sanguíneos, etc • 3.5%
Tomado de Ranken (2003)
6

2.3.3. Grasa
En la tecnología de productos cárnicos “grasa” ordinariamente significa
tejidos grasos. Las grasas también son llamadas lípidos y son el principal
componente de los tejidos grasos (Ranken, 2003).
Los lípidos son productos ternarios, compuestos de carbono, hidrogeno,
oxigeno, que considerándolos desde un enfoque nutricional aportan mayor energía
por unidad de peso debido a que el hidrogeno que contienen en proporción al
oxigeno, es muy superior que el de los glúcidos, se consideran dos grandes
grupos, los glicéridos y los no glicéridos (Pontes y Castelló, 1995).
2.3.4. Color
El color se define como la sensación resultante de una serie de fenómenos
percibidos simultáneamente con longitudes de onda comprendidas entre 380 y 780
nm (Kowaliski, 1978). Para determinar las características de un color se utiliza el
sistema CIE L* a* b*, donde L* determinará la brillantez de un color, de negro a
blanco (0.0) (100.0) y a* b*, determinará las longitudes de onda y la saturación del
color.
El color es uno de los principales indicadores de la calidad, que a menudo es
usado como un indicador de canales PSE Y SFO (Dransfield y Sosnicki 1999). El
color oscuro de la carne de las aves domésticas es más purpura congelada y más
roja cuando están descongeladas. La carne congelada acelerará el cambio de color
por el efecto de la luz, por lo cual la carne deberá estar cubierta durante su
almacenamiento en este estado (Ranken 2003).
Varios investigadores entre ellos Froning et al. (1978); McKee y Sams (1997)
han informado que ocurren cambios en el color, de la carne de pechuga de pavo,
cuando sufre un ritmo acelerado de glucolisis post mortem.
7

2.3.5. Temperatura y pH
El pH y la temperatura están estrechamente relacionados, ya que a un
incremento de temperatura el pH tiende a bajar. Temperaturas post mortem
elevadas (40 oC), aceleran el agotamiento del ATP (Adenosin trifosfato) en el
músculo, así como también un aumento en la tasa de degradación del glucógeno
en el musculo del guajolote (Mckee y Sams, 1997).
El pH ésta relacionado con la calidad de la carne ya que incrementos o
disminuciones en la canal afectan considerablemente sus propiedades
organolépticas tales como (color, sabor, olor y textura). Bendall y Wismer-
Pedersen (1962) mencionan que los rápidos incrementos o rápidas disminuciones
del pH repercuten en gran medida sobre la calidad de la carne de guajolote. Por
otra parte Boulianne y King (1995) encontraron que un pH alto en el músculo está
asociado con la carne oscura, y un pH bajo con un color claro, menor
enrojecimiento y mayor amarillamiento, así como también menor cantidad de
pigmentos, hierro y mioglobina, a lo que se le conoce como carne de tipo pálido,
suave y exudativo (PSE).
La carne tipo pálida, suave, exudativa (PSE) está directamente relacionada
con el pH y la desnaturalización de la proteína del músculo. Owens et al. (2000) en
un estudio realizado con pavos para determinar el por qué de este tipo de carne,
encontró que las proteínas se desnaturalizaban por una rápida disminución del pH,
y que como consecuencia las canales de pavos presentaban un color más pálido.
Cuando el suministro del oxigeno a la corriente sanguínea y el metabolismo
del animal cesa, el glucógeno se convierte en acido láctico, este ocasiona que el
pH del animal baje considerablemente de 7.0 a 5.5, lo cual repercute en la calidad
de la carne y es a lo que se conoce como glucolisis post morten, que da como
resultado carnes pálidas, suave y exudativas (Ranken, 2003).
8

Carne seca, firme y oscura (SFO), ocurre cuando el animal presenta un
incremento en el pH, debido a la poca formación de acido láctico por el poco
glucógeno debido a la inanición, agotamiento o estrés por un periodo prolongado,
lo que ocasiona un color de carne más oscuro de textura cerrada, con mejor
ligazón de agua, pero con calidad microbiológica inferior (Ranken, 2003).
2.3.6 Sabor
El sabor y el olor de la carne pueden verse afectados por cambios químicos,
microbiológicos y oxidativos (Ranken, 2003). Conforme aumenta la edad del animal
al momento del sacrificio el sabor también lo hará, de tal manera que un animal
mayor tendrá mejor sabor que uno joven. En algunos animales como el vacuno,
cordero y pollo con contenido reducido de grasa el sabor será menos palatable que
aquellos que tienen mayor contenido de grasa.
2.3.7 Congelación
La congelación es el enfriamiento de un producto a temperaturas muy bajas,
de tal forma que el agua que constituye parte del producto se transforma en hielo.
La congelación de la carne de manera rápida hace que se formen pequeños
cristales de hielo, lo que ocasiona que haya mucha dispersión de luz, obteniendo
una apariencia opaca y pálida, la carne congelada de manera lenta causa que se
formen cristales de hielo más grandes, dispersara menos luz y tendrá un aspecto
más oscuro y translucido. Al descongelar la carne los cambios de color
desaparecerán (Ranken, 2003).
9

3. OBJETIVOS
Determinar el rendimiento en canal de guajolotes autóctonos hembra y
macho sacrificados a 7, 12 y 15 meses de edad.
Determinar el pH y temperatura de pechuga, muslo, pierna, grasa de
guajolotes autóctonos hembras y machos a diferentes tiempos post mortem a 7, 12
y 15 meses de edad.
Determinar color de la canal de guajolotes autóctonos hembras y machos a
7, 12 y 15 meses de edad.
3.1. HIPOTESIS
El rendimiento en canal, pH y temperatura de la pechuga, muslo, pierna y
grasa de hembras es similar al de machos sacrificados a 7, 12 y 15 meses de
edad.
El rendimiento en canal de guajolotes autóctonos machos y hembras
sacrificados a 7, 12 y 15 meses de edad es similar entre ellos.
El color de la canal de los guajolotes autóctonos machos es similar al de
hembras a 7, 12 y 15 meses de edad.
10

4. MATERIALES Y METODOS
4.1. Localización
El presente trabajo se realizó en las instalaciones del proyecto de Cría de
Guajolotes Criollos de la Granja Experimental del Departamento de Zootecnia, de
la Universidad Autónoma Chapingo, la cual se localiza a 19° 21’ de Latitud Norte y
98°53’ longitud Oeste, a una altura de 2250 msnm. De acuerdo a la clasificación
del clima de Köppen modificada por García (1981), el clima de la región es
C(W0)(w)b(i)g que corresponde a un templado subhúmedo con lluvias en verano, la
precipitación media anual es de 644 mm y la temperatura media anual es de
17.8°C. La temperatura más alta se presenta en mayo, y la más fría en enero.
4.2. Animales utilizados
Se utilizaron 18 animales autóctonos mantenidos en condiciones de
confinamiento en las instalaciones de la granja de Zootecnia de la Universidad
Autónoma Chapingo. Nueve animales fueron machos los cuales se sacrificaron a
tres diferentes edades 7, 12 y 15 meses. Los otros 9 animales fueron hembras las
cuales se sacrificaron a las edades indicadas anteriormente. Los animales fueron
mantenidos con una dieta a base de maíz y pasta de soya. Antes del sacrificio, los
animales fueron separados por sexos y posteriormente se agruparon en grupos de
tres animales cada uno de acuerdo a la edad:
Grupo 1: Animales de 15 meses de edad, grupo 2: animales de 12 meses
de edad y grupo 3: animales de 7 meses de edad.
4.3. Pesaje
Un día antes del sacrificio a los animales se les retiró la comida con la
finalidad de que el buche no estuviera lleno y no se rompiera al momento del
eviscerado y despiezado. Una vez cumplidas las 24 horas de ayuno los animales
fueron pesados en una báscula analítica, y el peso vivo se registró en kilogramos.
11

4.4. Sacrificio
El sacrificio de los animales se realizó en el rastro de conejos ubicado en el
modulo de conejos que se encuentra en la granja experimental de la Universidad
Autónoma Chapingo.
El procedimiento fue el siguiente:
Izamiento
Los animales se sujetaron de las extremidades posteriores para ser
insensibilizados y desangrados.
Insensibilización
Se realizó de forma manual rompiendo la unión de la médula espinal con el
cerebro, el animal se mantuvo con la cabeza hacia abajo, se sujeto de las patas
con la mano izquierda, mientras que con la mano derecha fue sujetado del cuello
inmediatamente por debajo de la cabeza. Una vez en esta posición se efectuó una
tracción hacia abajo hasta que el animal no ofreció resistencia.
Desangrado
Una vez que los animales fueron insensibilizados se desangraron,
permaneciendo colgados por las patas por aproximadamente diez minutos, con el
fin de que se desangraran completamente. La sangre fue recolectada y pesada.
Decapitado y despatado
Consistió en remover la cabeza del animal así como las extremidades
inferiores.
Escaldado
Los guajolotes fueron sumergidos en agua caliente (80oC) por 25 segundos.
12

Desplumado
Se realizó un desplume manual con el fin de dejar la canal sin plumas.
Lavado
Posterior al desplume se realizó el lavado de la canal para eliminar residuos
que pudieran afectar su calidad organoléptica y microbiológica.
Eviscerado
Se practicó una incisión que partió desde la cloaca hacia el esternón, donde
se introdujo la mano en la cavidad abdominal para extraer las vísceras y órganos
internos (Battula et al., 2008).
4.5. Peso de la canal caliente
Después del sacrifico y evisceración, se procedió a pesar la canal. El peso
se expreso en kg.
4.6. Despiezado
Una vez que la canal quedó libre de piel y vísceras se cortó en las
diferentes piezas que la componen.
4.7. Lavado e identificación
Después de que la canal fue despiezada, se procedió a lavar cada una de
las piezas de la canal con agua a presión, así también como a identificarlas.
4.8. Inmersión en hielo
Cuando las diferentes piezas de la canal estuvieron limpias e identificadas,
se pesó cada una en una báscula digital. Después del pesado, las canales se
mantuvieron en hielo donde permanecieron a una temperatura promedio de 13 °C.
13

4.9. Registro de pH y temperatura
Una vez que los animales se sacrificaron, se procedió a desplumarlos para
medir el pH y la temperatura con un potenciómetro portátil marca “Minolta”
previamente calibrado con soluciones amortiguadoras de pH 4.00 y pH 7.00. Las
lecturas de pH y temperatura se tomaron en pechuga, pierna, muslo, grasa, a las
0h, 5h, 12h, 24h, 48h y 7 días postmorten. Durante las primeras 48 horas las
canales permanecieron en hielos a una temperatura promedio de 13oC, pasado
este tiempo, las canales fueron congeladas.
4.10. Peso del animal despiezado
Después de despiezar a cada animal se tomaron mediciones de cada uno de
los órganos tanto en hembras como en machos. Se registró el peso de cabeza,
pechuga, pierna, muslo, alas, patas, piel, grasa y vísceras, así como también se
registraron mediciones de la pechuga (alto y ancho) medición de pH a las 0h, 5h,
12h, 24h, 48h y 7 días después del sacrificio.
4.11. Peso de la canal en frio
El peso de la canal en frio se registro a las 5h, 12h, 24h, 48h y 7 días
después de sacrificados los animales. Los pesos expresados en kg.
4.12. Registro de color
El color de la pechuga, muslo, pierna y grasa fue registrado 48 hrs post
mortem. Para realizar esta actividad se procedió a medir el color con un colorímetro
marca “Hunter lab” las lecturas se efectuaron sobre la parte externa e interna de las
diferentes piezas de la canal, pasando la pieza de la canal por el ojo lector del
colorímetro, para posteriormente registrar la lectura de L*, a* y b*. Obteniéndose 3
repeticiones para posteriormente analizarlas estadísticamente mediante un análisis
de varianza multivariado y univariado.
El cambio de color medido se expresó mediante los índices L∗, ángulo hue y
croma. El índice L∗ se utiliza para evaluar el % de luz que se refleja, hue° el ángulo
14

de matiz y el valor croma representa la intensidad del color puesto que se
correlaciona mejor con el desarrollo visual de la luminosidad en comparación a los
valores CIE (L*, a*, b*) los valores de croma. Luminosidad (L*), rojo / eje verde (a*),
y amarillo / eje azul (b*). Chroma y el ángulo hue0 fueron calculados sobre las
mediciones de a* y b*.
Chroma, C = [ (a*)2 + (b*)2 ]1/2
Angulo hue, h° = arctan−1 (b*/a*).
Un ángulo hue de 0° representa un rojo puro, mientras que un ángulo hue de 90°
representa un amarillo puro.
5. VARIABLES DE RESPUESTA
5.1. Rendimiento en canal
Es la relación entre el peso de la canal caliente y el peso vivo del animal. El
peso vivo representó el 100% de la canal. Esta variable se expresó en porcentaje.
5.2. Determinación de pH
Se midió la evolución del pH después del sacrificio a 0h, 5h, 18h, 24h, 48 h y
7 días.
5.3. Determinación de Temperatura
Se midió la evolución de la temperatura después del sacrificio a 0h, 5h, 12h,
24h, 48 h y 7 días.
5.4. Determinación de color
Se realizaron lecturas de color en pechuga, muslo, pierna y grasa 48 horas
post mortem.
15

6. DISEÑO EXPERIMENTAL
El diseño experimental fue completamente aleatorizado (DCA). El análisis
estadístico fue un factorial 2x3x4, los factores fueron: sexo (Hembra y macho),
Edad (7, 12 y 15 meses), Pieza (muslo, pierna, pechuga y grasa) con tres
repeticiones (3 animales) y tres submuestreos (3 mediciones). El análisis de los
datos se realizó en el programa SAS System, 2000 (Versión 5.1.2600) mediante un
análisis de varianza univariado y multivariado (Anova y Manova).
Univariado. A una sola unidad se le midió una variable.
Multivariado. A una sola unidad se le midieron más de una variable para
posteriormente compararlas entre si.
La prueba de comparación de medias se realizó con la prueba de Duncan y
Medias ajustadas (Steel y Torrie, 1980). Se propone el modelo 1, para estimar si
hay diferencias estadísticas (P<0.05) en las canales analizadas.
Modelo 1, utilizado para analizar sexo, edad, pieza, pH, temperatura y color (L∗,
croma y hue°) :
Yijκ = μ + Si + Ej + Pĸ + (SEP)ij + ξijĸ
Donde:
Yijĸ = Valor de la variable respuesta correspondiente al i-ésimo sexo, j-ésima
edad, l-ésima pH temperatura, m-ésimo color, en la ĸ-ésima observación
μ = Media general
Si = Efecto del i-ésimo sexo (i = 1, 2)
16

Ej = Efecto de la j-ésima edad (j = 1, 2, 3)
Pĸ = Efecto de la ĸ-ésima pieza (j = 1, 2, 3,4)
(SEP)ij = Efecto de la ijĸ-ésima interacción sexo*edad*Pieza (ijĸ = 1, 2, 3)
ξijĸ = Error experimental, ξijĸ ~ NI (0, σ2)
Cuando solo se analizó la edad de los animales, el modelo se redujo a:
Modelo 2
Utilizado para analizar la edad fue el siguiente:
Yijĸ = μ + Si + Ej + ξijĸ
Donde:
Yijĸ = Valor de la variable respuesta correspondiente al i-ésimo tratamiento en la j-
ésima edad en la ĸ-ésima observación
μ = Media general
Si = Efecto del i-ésimo tratamiento (i = 1, 2, 3)
Ej = Efecto de la j-ésima edad (j = 1, 2, 3)
ξijĸ = Error experimental, ξij ĸ ~ NI (0, σ2)
El modelo utilizado para analizar el sexo fue el siguiente:
Modelo 3
Yijĸ = μ + Si + Ej + ξijĸ
17

Donde:
Yijĸ = Valor de la variable respuesta correspondiente al i-ésimo tratamiento en la j-
ésima sexo en la ĸ-ésima observación
μ = Media general
Si = Efecto del i-ésimo tratamiento (i = 1, 2, 3)
Ej = Efecto de la j-ésimo sexo (j = 1, 2, 3)
ξijĸ = Error experimental, ξij ĸ ~ NI (0, σ2)
18

7. RESULTADOS Y DISCUSIÓN
7.1. Efecto del sexo, edad y peso de guajolotes autóctonos en el rendimiento de la canal
El sexo, edad y peso al sacrificio de los animales fue importante para
determinar el rendimiento en canal. Como se observa en el Cuadro 1, los machos
presentaron mayor rendimiento comparado con el de las hembras. Asimismo, los
animales de mayor peso mostraron una tendencia a incrementar el rendimiento de
la canal.
Cuadro 1. Efecto del sexo, edad y peso de guajolotes autóctonos en el rendimiento de la canal a 7, 12 y 15 meses de edad.
NOTA: De los 18 animales que se tenían, 9 hembras y 9 machos a tres diferentes edades, se
obtuvo el promedio general de medias, para obtener un valor para cada edad.
Sexo
Edad (Meses)
Peso vivo (kg)
Peso de la canal (kg)
Rendimiento (%)
Hembra
7
3.087
2.014
65.24
12
3.460
2.333
67.42
15
2.501
1.580
63.17
Macho
7
5.743
4.273
74.40
12
7.710
5.889
76.38
15
7.976
6.250
78.36
19

Resultados similares a las tendencias encontradas fueron reportadas por
Cross et al. (1981) quien señala que el rendimiento de la canal estuvo relacionado
con el sexo del animal y obtuvo mayor rendimiento en los machos. Leclercq (1998)
señala que el peso vivo, el peso de la canal y el rendimiento de canal son
diferentes dependiendo del sexo del animal, estos autores indican que los machos
son más corpulentos y tienen una menor conversión alimenticia, lo cual coincide
con lo señalado por North (1986). Este autor indica que los machos tienen más
carne, debido a su gran tamaño, rápido crecimiento y buena digestión obteniendo
un mayor rendimiento de la canal en comparación con las hembras.
En este estudio, se observó que los machos pesaron casi el doble que las
hembras a las diferentes edades de sacrificio (cuadro 1). En el caso de los machos
se observó que a mayor edad hubo un pequeño aumento en el peso vivo de los
animales, esto mismo es señalado por Sell (1988) quien encontró en un estudio
relacionado con pavos que a mayor edad, los animales tenían un mayor peso vivo.
En el caso de las hembras (cuadro1), se observó un incremento en el peso
conforme avanzó la edad hasta un año, después de este tiempo el peso del animal
comenzó a disminuir. Esta misma tendencia se observó en el peso de la canal y
rendimiento, este comportamiento productivo probablemente sea debido a que los
animales habían pasado por la etapa reproductiva la cual inicia a la edad de 8
meses, para posteriormente a 9 meses la postura de huevos a los 10 meses
empollar y a la edad de 11,12 meses la crianza de polluelos y quizás no habían
recuperado su peso corporal del todo. En el caso de los machos a medida que el
peso del animal mostró incrementos en el peso vivo, el peso de la canal y el
rendimiento también incrementaron, esto quizá es debido a que el animal no sufre
un desgaste como en el caso de las hembras como resultado de actividad
reproductiva y el consumo de nutrientes se acumuló como músculo y grasa. El
comportamiento productivo de estas aves podría indicar que la ganancia de peso
de las hembras es más lento, que la de machos.
20

21
7.2. Peso de la canal despiezada
Los resultados del peso de las diferentes partes de los animales tanto para
hembras como para machos se muestran en el cuadro 2. Las medias mostradas
indican que el peso de los machos es superior al de las hembras (p<0.05) en las
tres diferentes edades y despiezado de los animales. Resultados similares fueron
encontrados por Tulloh (1964) en pavos. Este autor mencionó que el peso de las
aves repercutía en gran medida sobre el rendimiento del despiezado del animal,
concluyendo que a mayor peso, el rendimiento de las aves se incrementaba
considerablemente. Así mismo, Cross et al. (1981) mencionaron que el mayor
crecimiento de los machos favorece el mayor peso y por consiguiente el
rendimiento al sacrificio.
Para las variables de peso vivo (PV), peso del animal recién sacrificado con
vísceras y plumas (PARS CV c/p), peso canal caliente sin vísceras y plumas (PCC
SV s/p), peso canal caliente con cabeza (PCC cc), peso pechuga (PPe), peso
pierna (PPi), peso muslo (PM), peso alas (PA), peso piel (PPl), peso grasa (PG), se
encontraron diferencias (p<0.05) entre machos y hembras. Estos resultados
(Cuadro 2) pueden atribuirse a la mayor proporción de las piezas evaluadas debido
al efecto del crecimiento y desarrollo (Goll et al. 1992), lo que influye en los
rendimientos obtenidos en relación a los animales machos de mayor peso.

Cuadro 2. Peso de la canal entera y despiezada de guajolote autóctono hembra y macho a 3 diferentes edades (expresada en meses y kg.)
Edad1
PV
PARS CV c/p
PCC SV s/p
PCC cc
PPe
PPi
PM
PA
PPl
PG
Hembras 7 3.08b ± 0.03 2.22b ± 0.04 2.01b ± 0.12 2.86b ± 0.06 0.47b ± 0.02 0.31b ± 0.00 0.30b ± 0.03 0.27b ± 0.01 0.16b ± 0.00 0.07b ± 0.02
12 3.46a ± 0.12 2.61a ± 0.12 2.41a ± 0.18 3.23a ± 0.20 0.59a ± 0.03 0.40a ± 0.03 0.43a ± 0.03 0.38a ± 0.02 0.29a ± 0.03 0.19a ± 0.06
15 2.50c ± 0.36 1.67c ± 0.36 1.68c ± 0.32 2.34c ± 0.39 0.39c ± 0.02 0.25c ± 0.02 0.24c ± 0.02 0.23c± 0.02 0.13c ± 0.02 0.04c ± 0.02
Machos 7 5.74c ± 0.21 4.97c ± 0.32 4.27c ± 0.16 5.39c ± 0.09 1.06c ± 0.07 0.53c ± 0.01 0.56c ± 0.00 0.40c ± 0.01 0.33c ± 0.04 0.23c ± 0.06
12 7.62b ± 0.03 6.54b ± 0.12 6.19b ± 0.13 7.38b ± 0.02 1.51b ± 0.01 0.68b ± 0.02 0.76b ± 0.02 0.51b ± 0.06 0.45b ± 0.03 0.31b ± 0.02
15 7.89a ± 0.12 6.78a ± 0.13 6.45a ± 0.12 7.65a ± 0.12 1.69a ± 0.12 0.76a ± 0.01 0.85a ± 0.03 0.59a ± 0.02 0.51a ± 0.09
PV: Peso vivo, PARS CV c/p: Peso animal Recién Sacrificado con vísceras y plumas, PCC SV s/p: Peso canal caliente sin vísceras y plumas, PCC cc: Peso canal caliente con cabeza, PPe: Peso pechuga, PPi: Peso pierna, PM: Peso muslo, PA: Peso alas, PPl: Peso piel, PG: Peso grasa.
0.43a ± 0.03
1Número de animales para cada edad (n=3).
NOTA: Medias en la misma columna con diferente letra muestran diferencias significativas (P < 0.05).
22

Se observa en el cuadro 2, que el sexo y la edad del ave, repercute sobre el
rendimiento de las canales de guajolotes, misma tendencia observada en
animales sacrificados a la misma edad.
7.3. pH y temperatura en la canal despiezada
El pH de la canal despiezada es importante para determinar la calidad de la
carne, incrementos o decrementos de esta propiedad afecta las propiedades
físico-químicas de la canal, como el color (L*, Croma, Hue°) de las distintas piezas
de carne y dá como resultado diferentes niveles de calidad. Diversos autores
hacen referencia a esta propiedad entre ellos Barbut, (1997) y Fletcher (1999)
quienes indicaron que el color pálido de la carne de pavo está asociado con un
menor pH de la carne, similar al de la carne de cerdo cuando se presentan las
canales del tipo pálido, suave y exudativo (PSE).
En este estudio se observó que el pH disminuyó conforme avanzó el tiempo
post sacrificio que fue de 0h a 5h, para posteriormente mantenerse hasta las 48 h,
y después de este lapso tener un ligero incremento a los 7 d en las diferentes
edades (cuadro 3).
Cuadro 3. pH promedio de pechuga, muslo, pierna y grasa de guajolotes
autóctonos machos.
pH pierna pH muslo pH pechuga pH grasa
0 hrs 6.48 6.14 5.99 6.58
5 hrs 5.96 6.03 5.93 6.47
12 hrs 6.0 6.1 5.8 6.4
24 hrs 6.06 6.01 5.88 6.39
48 hrs 6.02 6.08 5.93 6.33
NOTA: Los datos del cuadro muestran el pH promedio para machos. 7 días 6.25 6.28 6.06 6.53
23

Cuadro 4. pH de pechuga, muslo, pierna y grasa de hembras a diferente edad (7, 12 y 15 meses) y horas post mortem. Edad (meses)
Pieza
Hora
pH
Edad (meses)
Pieza
Hora
pH
0 6.01ba±0.19 0 6.53a±0.23 5 5.79d±0.07 5 5.94d±0.11 7 Pechuga 12 5.85dc±0.13 7 Pierna 12 6.09c±0.02 24 5.84cd±0.06 24 6.07c±0.03 48 6.09a±0.08 48 6.23b±0.06 7 días 6.06a±0.32 7 días 6.27b±0.04 0 5.94a±0.06 0 6.49a±0.31 5 5.74c±0.01 5 5.97c±0.04 12 Pechuga 12 5.80b±0.05 12 Pierna 12 5.97c±0.06 24 5.84b±0.04 24 6.03c±0.05 48 5.90a±0.09 48 6.02c±0.09 7 días 5.95a±0.12 7 días 6.17b±0.02 0 5.98b±0.02 0 6.42a±0.12 5 5.78d±0.00 5 5.94d±0.09 15 Pechuga 12 5.83c±0.04 15 Pierna 12 6.02dc±0.08 24 5.97b±0.01 24 6.05dc±0.12 48 5.97b±0.04 48 6.11c±0.13 7 días 6.09a±0.10 7 días 6.24b±0.04 0 6.20a±0.06 0 6.51ba±0.17 5 6.01c±0.05 5 6.38c±0.07 7 Muslo 12 6.06c±0.06 7 Grasa 12 6.35dc±0.06 24 6.00c±0.09 24 6.31dc±0.09 48 6.11b±0.02 48 6.29d±0.12 7 días 6.25a±0.12 7 días 6.64a±0.15 0 6.20ba±0.09 0 6.43a±0.00 5 6.00c±0.12 5 6.31b±0.06 12 Muslo 12 6.06c±0.09 12 Grasa 12 6.30b±0.08 24 6.03c±0.04 24 6.35b±0.06 48 6.26a±0.06 48 6.36b±0.06 7 días 6.25a±0.07 7 días 6.43a±0.01 0 6.22a±0.13 0 6.82a±0.03 5 5.97b±0.09 5 6.56b±0.09 15 Muslo 12 5.99cb±0.12 15 Grasa 12 6.35d±0.04 24 6.07b±0.09 24 6.41c±0.00 48 6.13ba±0.11 48 6.81a±0.02 7 días 6.22a±0.16 7 días 6.84a±0.04
NOTA: Medias en la misma columna con diferente letra muestran diferencias (p<0.05).
24

El pH de las diferentes piezas de la canal disminuyó considerablemente en
las primeras horas después del sacrificio, similar a lo indicado por Obanor et al.,
(2005) en una de sus investigaciones sobre calidad de la carne en pavos post
mortem, donde indicó que después del sacrificio, el pH de los pavos decrecía
rápidamente en las primeras 5 horas para después estabilizarse e incrementar
nuevamente, así mismo Schreuers (2000) menciona que el pH disminuye
gradualmente a partir de 2 a 6 horas después del sacrificio, y se mantiene
constante a partir de las 24 horas post mortem, para posteriormente incrementar.
El pH promedio de la pechuga de hembras y machos a las 0 h fue de 5.99
el cual se encuentran entre los intervalos obtenidos por diversos autores entre
ellos Northcutt et al. (1998) quienes señalan que el pH inicial del músculo de las
aves en un estudio sobre el rendimiento en canal fue de (5.99 ± 0.08) de los
pavos, por otra parte Obanor et al. (2005) encontraron que a 30 min post mortem
el pH de las canales fue de 6.39 ( ± 0.13) y que disminuyó de manera constante a
(6.0 ± 0.17) en las próximas horas post mortem.
La temperatura entre las cuales se mantuvieron las canales despiezadas de
guajolote autóctono fue de 10 a 13 0C (Grafica 1 y 2), un rango apropiado que no
perjudica la calidad de la canal después del sacrificio. Algunos autores entre ellos
Calkins et al. (1982) mencionan que temperaturas elevadas (40 °C) después del
sacrificio producen canales del tipo pálido, suave, exudativo. Los autores
mencionados indicaron que temperaturas mayores a 40 oC post morten presentan
una mayor fatiga en el músculo comparado con temperaturas de 0 y 20 oC. En el
caso de la temperatura, se observó (Grafica 1) una disminución drástica de la
misma a las 5 h post mortem para después mantenerse prácticamente constante
debido probablemente a que la canal fue sumergida en hielo después de tener una
primera medición que fue a las cero horas después del sacrificio, y se mantuvo así
hasta las 48 horas post mortem. Pasadas las 48 hrs las canales fueron
mantenidas en congelación a −5 o C hasta los 7 d, sin embargo no hubo grandes
diferencias en la temperatura y pH de las diferentes piezas de la canal después de
haber sido descongeladas (Grafica 2).
25

Grafica 1. pH y temperatura de hembras a diferentes horas post mortem (0h, 5h, 12h, 24h, 48h y 7 días).
05101520253035
55.25.45.65.86
6.26.46.66.8
0 h 5 h 12h 24 h 48 h 7 d
pH PiernapH MuslopH PechugapH GrasaTemperatura
Temperatura
Tiempo
pH
Grafica 2. pH y temperatura de machos a diferentes horas post mortem (0h, 5h, 12h, 24h, 48h y 7 días).
0
5
10
15
20
25
30
55.25.45.65.86
6.26.46.66.8
0 h 5 h 12 h 24 h 48 h 7 d
pH PiernapH MuslopH PechugapH GrasaTemperatura
Temperatura
Tiempo
pH
NOTA: Cada uno de los resultados que aparecen en la grafica 1 y 2, son el promedio de las medias de los 18 animales hembras y machos.
26

7.4. Efecto del sexo y la edad en el color de la canal del guajolote autóctono
El color de la canal de las hembras presentó significativas (p<0.05), menor
ángulo de matiz de la canal y mas brillantes comparada con la carne de los
machos (Cuadro 5). Lo que nos indica que las hembras tuvieron la carne más
oscura y más roja, comparada con machos.
Cuadro 5. Efecto del sexo de los guajolotes autóctonos en la brillantes (L*), intensidad (Croma) y ángulo de matiz (Hue °).
Sexo
L (%)
Croma (Pureza de color)
Hue (°)
Machos
50.36ª± 0.12
10.35a± 0.36
93.91ª± 0.01
Hembras
46.87b ± 0.36
9.99a± 0.42
89.16b ± 0.12
NOTA: Medias en la misma columna con diferente letra indican diferencias (P<0.05).
Respecto a la edad de los guajolotes autóctonos (cuadro 6), existen diferencias
(p<0.05) en el color de la carne de los animales jóvenes, estos presentan mayor
Cuadro 6. Efecto de la edad de los guajolotes autóctonos en la brillantes (L*), intensidad (Croma) y ángulo de matiz (Hue °).
Edad (Meses)
L* (%)
Croma (pureza de color)
Hue (°)
7
52.66a± 0.58
10.59b± 0.02
91.73b± 0.12
12 53.23a± 0.67 12.09ª± 0.07 100.08ª± 0.06
15 40.25b± 0.32 7.98c± 0.03 82.79 c± 0.03
NOTA: Medias en la misma columna con diferente letra muestran diferencias (P<0.05).
27

porcentaje de luminosidad (L*), pureza de color (Croma), y ángulo de matiz (Hue°).
Las canales tienen una presentación de color mas clara y brillante a 7 y 12 meses
de edad comparado con los animales viejos de 15 meses, esto puede atribuirse a
que los animales viejos tienen mas actividad de las mitocondrias, lo que resulta
en un consumo mayor de oxigeno y por consiguiente una presentación de carne
ligeramente más oscura (Ashmore et al. 1972), lo cual podría demeritar su calidad.
De manera general puede señalarse que el color (L*, Croma y Hue °) de la
canal estuvo relacionado con el sexo y edad del animal.
28

7.5. Análisis de varianza univariado
Ho. Si existen diferencias significativas entre la edad de los animales respecto a:
V. S.
Ha. No existen diferencias significativas entre la edad de los animales respecto a:
V. Resp. Statistic F Value Num DF Den DF Pr > F
Color F- Snedecor 38.61 6 990 <.0001∗∗∗
pH F- Snedecor 2.08 3 1051 0.1794
Temp. F- Snedecor 4.74 6 1327 0.1545
∗∗∗ Altamente significativo (P< 0.001)
∗∗ Significativo (P< 0.05)
Utilizando un nivel de significancia α=0.05, se comprueba que hay
diferencias altamente significativas (P<.0001) entre la edad respecto al color de
los animales, se rechaza la Ha. y se acepta la Ho., para el pH y temperatura se
rechaza la Ho. y se acepta la Ha., por que no presentan diferencias (P>0.05).
Ho. Si existen diferencias significativas entre el sexo de los animales respecto a:
V. S.
Ha. No existen diferencias significativas entre el sexo de los animales respecto a:
V. Resp. Statistic F Value Num DF Den DF Pr > F
Color F- Snedecor 4.30 3 522 <.0001∗∗∗
pH F- Snedecor 4.74 6 1044 0.1545
∗∗∗ Altamente significativo (P< 0.001)
Temp. F- Snedecor 35.29 27 1525 0.1425
∗∗ Significativo (P< 0.05)
29

Se rechaza la Ho. y se acepta la Ha. no existen diferencias significativas
(P> 0.05), según F- Snedecor, entre el sexo respecto al pH y temperatura de los
animales. El color entre el sexo de los animales si presentó diferencias altamente
significativas (P<.0001), se acepta la Ho. y rechazando la Ha.
Ho. Si existen diferencias significativas entre la interacción sexo∗edad de los
animales respecto a:
V. S.
Ha. No existen diferencias significativas entre la interacción sexo∗edad de los
animales respecto a:
V. Resp. Statistic F Value Num DF Den DF Pr > F
Color F- Snedecor 9.86 6 990 <.0001
pH F- Snedecor 7.89 6 1044 0.1362
∗∗∗ Altamente significativo (P< 0.001)
Temp. F- Snedecor 2.24 6 878 0.1652
∗∗ Significativo (P< 0.05)
Se comprueba que no existen diferencias entre la interacción de los
animales respecto al pH y temperatura, se rechaza la Ho. y se acepta la Ha., para
el color respecto a la edad de los animales si presenta diferencias, se acepta la
Ho. y rechazando la Ha.
30

7.6. Análisis de varianza multivariado
Ho. Si existen diferencias significativas entre edad de los animales respecto a pH,
color y temperatura.
V. S.
Ha. No existen diferencias significativas entre edad de los animales respecto a pH,
color y temperatura.
Statistic F Value Num DF Den DF Pr > F
∗∗∗ Altamente significativo (P< 0.001)
Wilks' Lambda 39.07 6 1016 <.0001∗∗∗
∗∗ Significativo (P< 0.05)
Utilizando un nivel de significancia α=0.05, se comprueba que si hay
diferencias altamente significativas en la edad de los animales (P <.0001), según
Wilks' Lambda se acepta la hipótesis nula (Ho.) Y se rechaza la hipótesis alterna
(Ha.)
Ho. Si existen diferencias significativas entre el sexo de los animales respecto a
pH, color y temperatura.
V. S.
Ha. No existen diferencias significativas entre el sexo de los animales respecto a
pH, color y temperatura.
Statistic F Value Num DF Den DF Pr > F
Wilks' Lambda 5.37 3 508 0.0012∗∗
∗∗∗ Altamente significativo (P< 0.001)
∗∗ Significativo (P< 0.05)
31

Se acepta la Ho. y se rechaza la Ha., si hay diferencias significativas
(P<0.05), según Wilks' Lambda respecto al sexo de los animales.
Ho. Si existen diferencias significativas entre la interacción sexo∗edad de los
animales respecto a pH, color y temperatura.
V. S.
Ha. No existen diferencias significativas entre la interacción sexo∗edad de los
animales respecto a pH, color y temperatura.
Statistic F Value Num DF Den DF Pr > F
∗∗∗ Altamente significativo (P< 0.001)
Wilks' Lambda 9.95 6 1016 <.0001∗∗∗
∗∗ Significativo (P< 0.05)
Según Wilks' Lambda se comprueba que si existe una interacción
sexo∗edad de los animales. Se acepta la Ho. Y se rechaza la Ha.
32

8. CONCLUSIONES
De acuerdo a los resultados obtenidos se concluye que el rendimiento en
canal es mayor para los guajolotes autóctonos machos, respecto a hembras. El
rendimiento incremento gradualmente conforme incrementó el peso vivo y la edad
del animal.
De forma cualitativa se observó, que el color de la canal de hembras y
animales machos a 15 meses de edad (animales viejos) es más oscura respecto a
machos pequeños y jóvenes (7 y 12 meses).
El pH de los guajolotes autóctonos machos y hembras, disminuye conforme
avanza el tiempo post sacrificio en las primeras horas (0 a 5h) para mantenerse
hasta las 48 h, después de este lapso tener un ligero incremento a los 7 días en
las diferentes edades.
La temperatura no influyó en el rendimiento de la canal, debido a que fue
similar para hembra y macho en las tres diferentes edades de los animales (7, 12
y 15 meses).
33

9. LITERATURA CITADA
Ashmore, C. R., W. Parker, and L. Doerr. 1972. Respiration of mitochondria
isolated from dark-cutting beef: postmortem changes. J. Anim. Sci. 34:46-
48.
Battula, V, M. W. Schilling, Y. Vizzier-Thaxton, J. M. Behrends, J. B. Willimas and
T. B. Schimidt. 2008. The effects of low atmosphere stunning and deboning
time on broiler breast meat quality. Poultry Sci. 87:1202-1210.
Barbado J. L. 2004. Cría de aves. “Gallinas ponedoras y pollos parrilleros”.
Editorial Albatros. 1era. Edición. Buenos aires, Argentina.
Barbut, S., 1997. Ocurrence of pale soft exudative meat in mature turkey hens. Br.
Poult. Sci 38:355-358.
Bendall, J. R., and J. Wismer-Pedersen, 1962. Some properties of the fibrillar
proteins of normal and watery pork muscle. J. Food Sci. 27:144-159.
Berg, B. N., and H. S. Simms, 1960. Nutrition and longevity in the rat. II. Longevity
and onset of disease with different levels of food intake. J. Nutr. 71:255–
259.
Boulianne, M., and A. J. King. 1995. Biochemical and color characteristics of
skinless boneless pale chicken breast. Poult. Sci. 74:1693–1698.
Buddiger, N. y G. Albers G. 2000. Futuras tendencias en la producción de pavos.
Industria Avícola. 47(5)28-30.
Calkins, C. R.; T. R. Dutson; G. C. Smith and Z. L. Carpenter. 1982. Concentration
of creatine phosphate, adenine nucleotides and their derivatives in
electrically stimulated and non-stimulated beef muscle. J. Food Sci.
47:1350–1353.
Card, E. L., and C. M. Nesheim, 1968. Nutrition of the chicken. Producciòn avìcola.
197-215.
34

Cross, H. R., R. L. West, and T. R. Dutson, 1981. Comparison of methods for
measuring sarcomere length in beef Semitendinosus muscle. Meat Sci.
5:261–266.
Dransfield, E., and A. A. Sosnicki. 1999. Relationship between muscle growth and
poultry meat quality. Poult. Sci. 78:743–746.
Ewart, J. 1993. Evaluation of genetic selection techniques and their application in
the next decade. Br. Poult. Sci. 34:3–10.
Forrest, J. C., E. D. Aberle, H. B. Hedrick, M. D. Judge, and R. A. Merkel, 1975.
Properties of fresh meat. Pages 174–189 in: Principles of Meat Science. W.
H. Freeman and Co., San Francisco, CA.
Fletcher, D. L. 1999. Broiler Breast Meat Color Variation, pH, and Texture. Poultry
Sci.78:1323–1327.
Froning, G. W., A. S. Babji, and F. B. Mather. 1978. The effect of preslaughter
temperature, stress, struggle and anesthetization on color and textural
characteristics of turkey muscle, Poultry Sci. 57:630-633.
García, E. 1981. Modificaciones al sistema de clasificación de Köppen. Impreso
México. México, D. f.
Goll, D. E., V. F. Thompson, R. G. Taylor, and J. A. Christiansen. 1992. Role of the
calpain system in muscle growth. Biochimie 74:225–237.
Kowaliski, P. 1978. Mesura y visión del color. Editorial parís, Masson, New York.
Leclercq, B. 1998. Lysine: Specific effects of lysine on broiler production:
Comparison with threonine and valine. Poult. Sci. 77:118–123.
McKee, S. R., and A. R. Sams. 1997. The effect of seasonal heat stress on rigor
development and the incidence of pale, exudative turkey meat. Poutry. Sci.
76: 1616-1620.
Nixey Y, C. 1988. En: Recent Adv. in Anim. Nutr. W. Haresign y D.W.A. Cole
(Eds.). Butterworths, Reino Unido. pp. 87-98.
Northcutt, J. K., R. J. Buhr, and L. L. Young. 1998. Influence of Preslaughter
Stunning on Turkey Breast Muscle Quality. Poultry Sci. 77:487–492.
35

North O. M. 1986. Manual de producción avícola. Editorial el manual moderno,
Segunda edición. México, Df.
Obanor, F., J. D. Morton, G. H. Geesink, and R. Bickerstaffe. 2005. Effect of
Processing on Turkey Meat Quality and Proteolysis. Poultry Sci. 84:1123–
1128.
Owens, C. M., E. M. Hirschler., R. Martinez-Dawson and A. R. Sams. 2000. The
characterization and incidence of Pale, Soft, Exudative Turkey Meat in a
Commercial Plant. Poultry Sci. 79: 553-558.
Pontes P. M.; J.A. Castelló. 1995. Alimentación de las aves. Ed. Real Escuela de
Avicultura. Barcelona, España. 540 p.
Plavnik, I., and S. Hurwitz. 1988. Early feed restriction in chicks: effect of age,
duration, and sex. Poult. Sci. 67:384–390.
Puron, D., R. Santamaria, J. C. Segaura, and J. L. Alamilla. 1995. Broiler
performance at different stocking densities. J. Appl. Poult. Res. 4:55–60.
Ranken, M. D. 2003. Manual de industrias de la carne. Mundi-Prensa. Madrid,
España.
Rubio, M.A., 2003. Comparación del valor nutricional de los distintos tipos de
carne. II Seminario Internacional Fundisa. Ed. FUNDISA (Fundación Ibérica
para la Seguridad Alimentaria).
Scott, L. M., C. M. Nesheim, and J. R. Young. 1982. Nutrition of the chicken.
Ithaca, New York. 410-427.
Schreuers, F. J. G. 2000. Post-mortem changes in chicken muscle. World’s Poult.
Sci. J. 56:319–346.
SAS Institute. 2000. SAS/STAT User’s Guide. Version 8. SAS Inst. Inc., Cary, NC.
Sell, J. 1988. How age and sex affect carcass yield. Turkey World 64(3):24–25.
Steel, R.G.D., and J. H. Torrie. 1980. Principles and Procedures of Statistics: A
Biometrical Approach (2nd Ed.). McGraw-Hill Book Co., New York.
Tulloh, N. M. 1964. The carcass composition of sheep, cattle and pies as functions
of body weight. Symposium on Carcass Composition and Appraisal of Meat
36

Animals, The Commonwealth Scientific and Industrial Research
Organization, East Melbourne, Victoria, Australia.
Watts, G., and C. Kennett. 1995. The broiler industry. Poult. Tribune (Sep.):6–18.
37

Anexo 1: Análisis de varianza de mínimos cuadrados para rendimiento en canal
en guajolotes autóctonos.
Fuente DF CM F-Valor Pr > F R-cuadrado
Coef. Var
Peso vivo
Modelo 5 17.090 102.33 <.0001 0.977 8.067
Error 12 0.167
Sangre
Modelo 5 0.050 13.39 0.0001 0.848 31.240
Error 12 0.003
Peso sin plumas
Modelo 5 13.810 44.79 <.0001 0.949 11.643
Error 12 0.308
Peso patas
Modelo 5 0.010 57.68 <.0001 0.960 9.756
Error 12 0.000
Peso canal
Modelo 5 12.741 88.25 <.0001 0.973 10.163
Error 12 0.144
38

Anexo 2: Análisis de varianza de mínimos cuadrados para rendimiento en canal
en guajolotes autóctonos.
Fuente DF CM F-Valor Pr > F R-cuadrado
Coef. Var
Peso retazo
Modelo 5 0.613 45.77 <.0001 0.950 11.296
Error 12 0.013
Peso grasa
Modelo 5 0.551 41.49 <.0001 0.945 26.372
Error 12 0.013
Peso piel
Modelo 5 0.1534 20.35 <.0001 0.894 23.446
Error 12 0.007
Vísceras
Modelo 5 0.201 20.09 <.0001 0.893 10.575
Error 12 0.010
Peso cabeza
Modelo 5 0.013 24.43 <.0001 0.910 17.730
Error 12 0.000
39
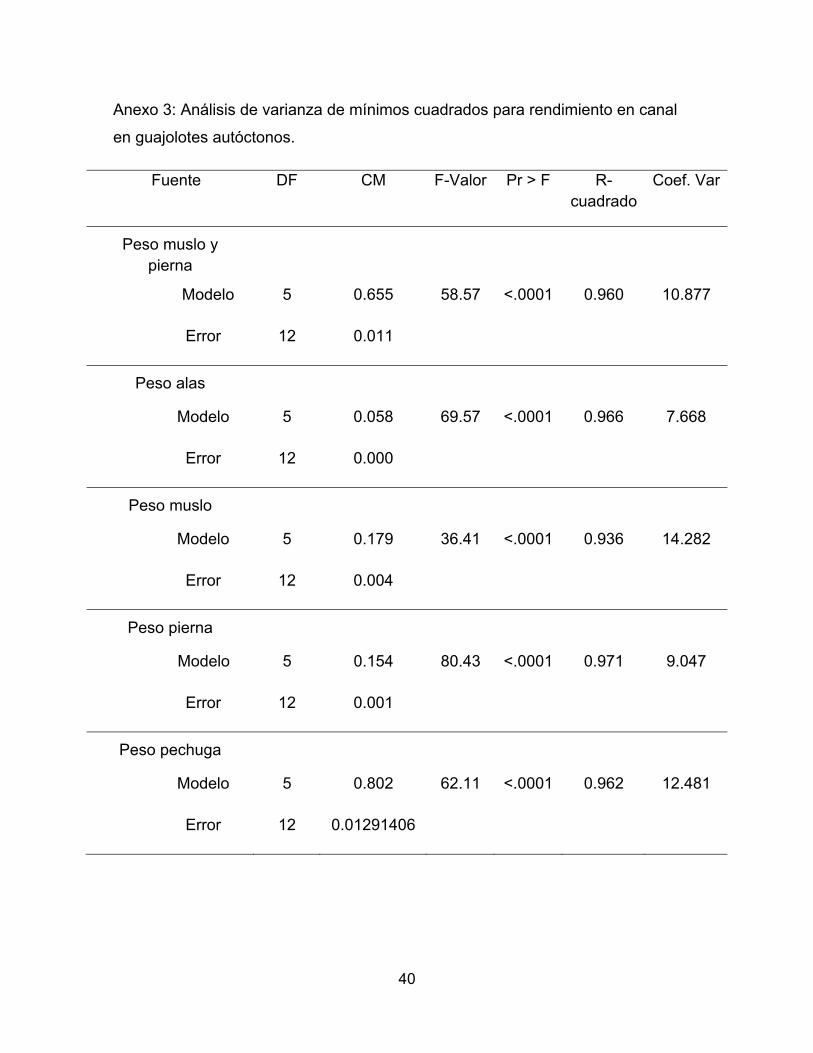
Anexo 3: Análisis de varianza de mínimos cuadrados para rendimiento en canal
en guajolotes autóctonos.
Fuente DF CM F-Valor Pr > F R-cuadrado
Coef. Var
Peso muslo y pierna
Modelo 5 0.655 58.57 <.0001 0.960 10.877
Error 12 0.011
Peso alas
Modelo 5 0.058 69.57 <.0001 0.966 7.668
Error 12 0.000
Peso muslo
Modelo 5 0.179 36.41 <.0001 0.936 14.282
Error 12 0.004
Peso pierna
Modelo 5 0.154 80.43 <.0001 0.971 9.047
Error 12 0.001
Peso pechuga
Modelo 5 0.802 62.11 <.0001 0.962 12.481
Error 12 0.01291406
40

Anexo 4: Análisis de varianza de mínimos cuadrados para rendimiento en canal
en guajolotes autóctonos.
Fuente DF CM F-Valor Pr > F R-cuadrado
Coef. Var
Peso huesos pechuga
Modelo 5 0.035 1.40 <.0001 0.368 99.015
Error 12 0.025
Peso huesos pierna
Modelo 5 0.009 15.50 <.0001 0.865 19.643
Error 12 0.000
Peso huesos muslo
Modelo 5 0.001 7.98 0.001 0.768 19.229
Error 12 0.000
Peso huesos alas
Modelo 5 0.006 21.95 <.0001 0.901 11.672
Error 12 0.000
Peso huesos retazo
Modelo 5 0.040 8.87 <.0001 0.787 20.251
Error 12 0.004
41
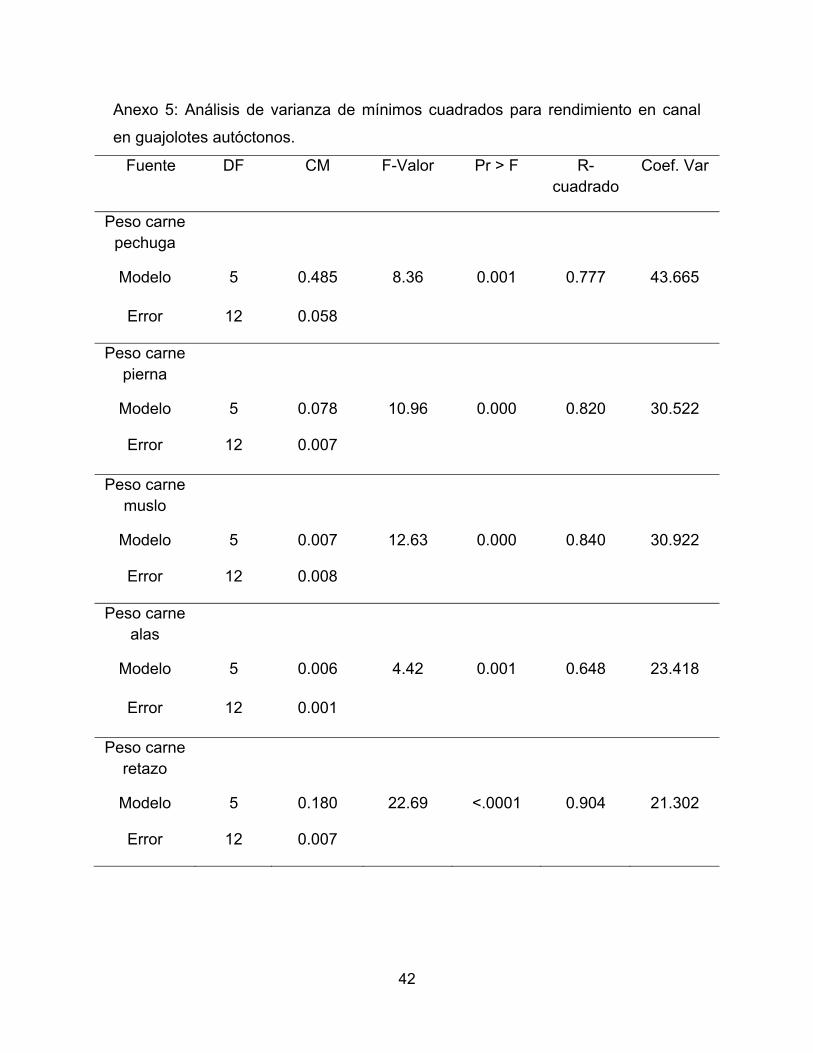
Anexo 5: Análisis de varianza de mínimos cuadrados para rendimiento en canal
en guajolotes autóctonos.
Fuente DF CM F-Valor Pr > F R-cuadrado
Coef. Var
Peso carne pechuga
Modelo 5 0.485 8.36 0.001 0.777 43.665
Error 12 0.058
Peso carne pierna
Modelo 5 0.078 10.96 0.000 0.820 30.522
Error 12 0.007
Peso carne muslo
Modelo 5 0.007 12.63 0.000 0.840 30.922
Error 12 0.008
Peso carne alas
Modelo 5 0.006 4.42 0.001 0.648 23.418
Error 12 0.001
Peso carne retazo
Modelo 5 0.180 22.69 <.0001 0.904 21.302
Error 12 0.007
42

43
Anexo 6: Análisis de varianza de mínimos cuadrados para color en guajolotes
autóctonos.
Fuente DF CM F-Valor Pr > F R-cuadrado Coef. Var
L
Modelo 29 3014.62 27.54 <0.0001 0.610 21.518
Error 510 109.462
CROMA
Modelo 29 278.424 23.98 <.0001 0.576 33.318
Error 510 11.610
HUE
Modelo 29 0.867 16.31 <.0001 0.481 25.194
Error 510 0.053