earliest mexican turkeys (meleagris gallopavo) in the … earliest mexican turkeys (meleagris...
TRANSCRIPT

1
Earliest Mexican turkeys (Meleagris gallopavo) in the Maya region: implications for pre-1
Hispanic animal trade and the timing of turkey domestication 2
3
Erin Kennedy Thorntona1, Kitty F. Emeryb, David Steadmanb, Camilla Spellerc,e, 4
Ray Mathenyd, and Dongya Yangc 5
6
a Trent University Archaeological Research Centre, DNA C205, 2140 East Bank Drive, 7
Peterborough, ON K9J 7B8, Canada. 8
9
b Florida Museum of Natural History, University of Florida, Gainesville, FL 32611–7800, USA 10
11
c Ancient DNA Laboratory, Department of Archaeology, Simon Fraser University, Burnaby, BC 12
V5A 1S6, Canada 13
14
d Department of Anthropology, Brigham Young University, Provo, Utah 84042 15
16
e Department of Archaeology, University of Calgary, Calgary, AB T2N 1N4, Canada 17
18
19
1To whom correspondence should be addressed. E-mail: [email protected] 20
21

2
Abstract: 22
Late Preclassic (300 BC–AD 100) turkey remains identified at the archaeological site of El 23
Mirador (Petén, Guatemala) represent the earliest evidence of the Mexican turkey (Meleagris 24
gallopavo) in the ancient Maya world. Archaeological, zooarchaeological, and ancient DNA 25
evidence combine to confirm the identification and context. The natural pre-Hispanic range of 26
the Mexican turkey does not extend south of central Mexico, making the species non-local to the 27
Maya area. Prior to this discovery, the earliest evidence of M. gallopavo in the Maya area dated 28
to approximately one thousand years later. The El Mirador specimens therefore represent 29
previously unrecorded Preclassic exchange of animals from northern Mesoamerica to the Maya 30
cultural region. As the earliest evidence of M. gallopavo found outside its natural geographic 31
range, the El Mirador turkeys also represent the earliest indirect evidence for Mesoamerican 32
turkey rearing or domestication. Although wild and domestic forms of M. gallopavo are 33
morphologically similar, the presence of male, female and sub-adult turkeys, and reduced flight 34
morphologyfurther suggests that the El Mirador turkeys were raised in captivity. This confirms 35
that the origins of turkey husbandry or at least captive rearing date to the Preclassic. 36
37
Keywords: Meleagris gallopavo, turkey, Maya, Mesoamerica, El Mirador, trade 38
39

3
The turkey was a significant animal for the ancient Maya, whose realm stretched from 40
northern Honduras to southern Mexico. Turkeys were not only a source of food, but were also 41
important sacrificial offerings, and their feathers, bones, and other byproducts were used to 42
produce medicines, fans, tools, musical instruments and personal adornments. Until this study, 43
however, the Maya were assumed to have used only the native, wild ocellated turkey (Meleagris 44
ocellata) throughout the Preclassic to Classic period of cultural florescence (ending in AD 1000). 45
The Mexican turkey (Meleagris gallopavo gallopavo), domesticated in central/northern Mexico 46
[1], was presumed not to have been introduced until the Postclassic (AD 1000–1500), the final 47
period of pre-Contact Maya occupation. Our recent identification of M. gallopavo in Late 48
Preclassic (ca. 300 BC–AD 100) deposits from the Maya archaeological site of El Mirador 49
overturns these assumptions. In this collaborative study, we identified the El Mirador turkey 50
specimens through morphology, osteometrics, and ancient DNA (aDNA) analysis. The context 51
and dates were confirmed through archaeology and AMS radiocarbon dating. The results lead us 52
to reconsider the timing of turkey domestication and diffusion throughout Mesoamerica, as well 53
as the nature and extent of Preclassic Mesoamerican trade connections. 54
Today, the domesticated form of M. gallopavo is distributed worldwide, but its wild 55
progenitor was limited to the eastern and southwestern United States and central/northern 56
Mexico north of the Isthmus of Tehuantepec, and thus outside the Maya cultural region [1–3] 57
(Fig.1). The absence of wild populations of M. gallopavo in the Maya area after the end of the 58
Pleistocene is supported by both the paleontological and archaeological records [4–6]. In 59
contrast, the ocellated turkey ranges throughout the northern half of the Maya cultural area 60
including Mexico’s Yucatan Peninsula and northern Belize and Guatemala where it remains 61

4
locally common [3,7]. Although some ocellated turkeys may have been raised in captivity during 62
pre-Hispanic times, there is no evidence that this species was ever domesticated [2,8]. 63
The exact timing and location of New World turkey domestication are still unknown: 64
recent evidence points to at least two separate domestication events in Mexico and the North 65
American Southwest [9]. In central Mexico, archaeological M. gallopavo bones have been 66
identified at sites dating to 800–100 BC [10,11]. It is unclear whether these early specimens 67
represent wild or domestic individuals, but domestic turkeys were likely established in central 68
Mexico by the first half of the Classic Period (ca. AD 200–1000) [12]. Until this study, M. 69
gallopavo had not been identified in any Maya archaeological deposits predating the Postclassic 70
[6,8,13]. The Postclassic Maya specimens are all presumed to represent domesticated individuals 71
[8,13] either imported directly from central/northern Mexico or bred and raised in the Maya 72
world following their initial introduction. 73
The M. gallopavo specimens reported here were recovered from the major archaeological 74
site of El Mirador, located in north-central Petén, Guatemala (SI Text) (Fig. 1). Settlement at the 75
site dates back to at least 600 BC, but population and architectural extent peaked at the site 76
during the Late Preclassic (300 BC–AD 100) when one of the largest assemblages of Maya 77
public architecture was constructed at the site, including but not limited to the Tigre and Danta 78
Pyramids. El Mirador’s Late Preclassic florescence coincided with a time of increasing social, 79
political, and economic complexity in the Maya region when many of the hallmarks of Classic 80
Maya civilization (e.g., institution of kingship, monumental stone architecture, extensive trade 81
networks, and elaborate iconography) were established. At the end of the Late Preclassic, the site 82
was largely abandoned although there was a small presence in the Early Classic and somewhat 83

5
more substantial settlement during the Late Classic. No monumental constructions like those 84
from the Late Preclassic occurred during these later occupations. 85
Zooarchaeological turkey specimens (n=7) from El Mirador were recovered along with 86
other animal remains (n=1116) from the Tigre complex, a large public architectural group on the 87
site’s western edge (Fig. 2). Most of the turkey bones were associated with the Jaguar Paw 88
Temple (Op. 26), a nine meter high platform topped by triadic architecture and decorated with 89
sculptured stucco masks. An additional turkey specimen was recovered from an eight meter high 90
building (Op. 35) located on the east side of the Tigre Plaza. The turkey bones were associated 91
with Late Preclassic ceramics in well-sealed, undisturbed contexts [14, SI Text]. AMS 92
radiocarbon ages from animal bones found in close association with the turkey remains confirm 93
that the deposits are Preclassic (cal 327 BC–AD 54) (Table S2). 94
95
Results 96
The turkey specimens were identified during zooarchaeological analysis of El Mirador 97
animal remains conducted by the Environmental Archaeology Program of the Florida Museum 98
of Natural History (FLMNH-EAP), University of Florida. Comparison with modern FLMNH-99
EAP and FLMNH-Ornithology collections confirmed that six of the seven specimens are M. 100
gallopavo (Table 1). The remaining specimen (a fragmentary femur) could not be identified to 101
the species level because of poor preservation. Morphological characteristics identifying the 102
specimens as M. gallopavo include element size, shape/curvature and robustness as well as 103
spacing of the quill tubercles (also called cubital tubercles or papillae remigiales) on two ulnae 104
(Fig. 3). The quill tubercles, which form where tendons connect the secondary flight feathers to 105
the ulna are also underdeveloped suggesting that the animals were raised in captivity. Age and 106

6
sex characteristics (e.g., skeletal element size, tarsometatarsus spur morphology) further indicate 107
that a minimum of three Mexican turkeys are represented in the assemblage—two males and a 108
female. One of the males is a subadult (<2 years old). 109
The morphological evaluations were supported by osteometric analysis. Five of the seven 110
skeletal elements were complete enough to allow for shaft width and depth measurements. When 111
compared to published M. gallopavo and M. ocellata osteometrics [6], three specimens fall 112
within the range of male domestic Mexican turkeys (Fig. 4). 113
Ancient DNA analysis of four of the turkey bones, conducted in the Simon Fraser 114
Ancient DNA Laboratory, further verified the morphological and osteometric identifications 115
(Table 1). Poor preservation of tropical faunal assemblages is often problematic for aDNA 116
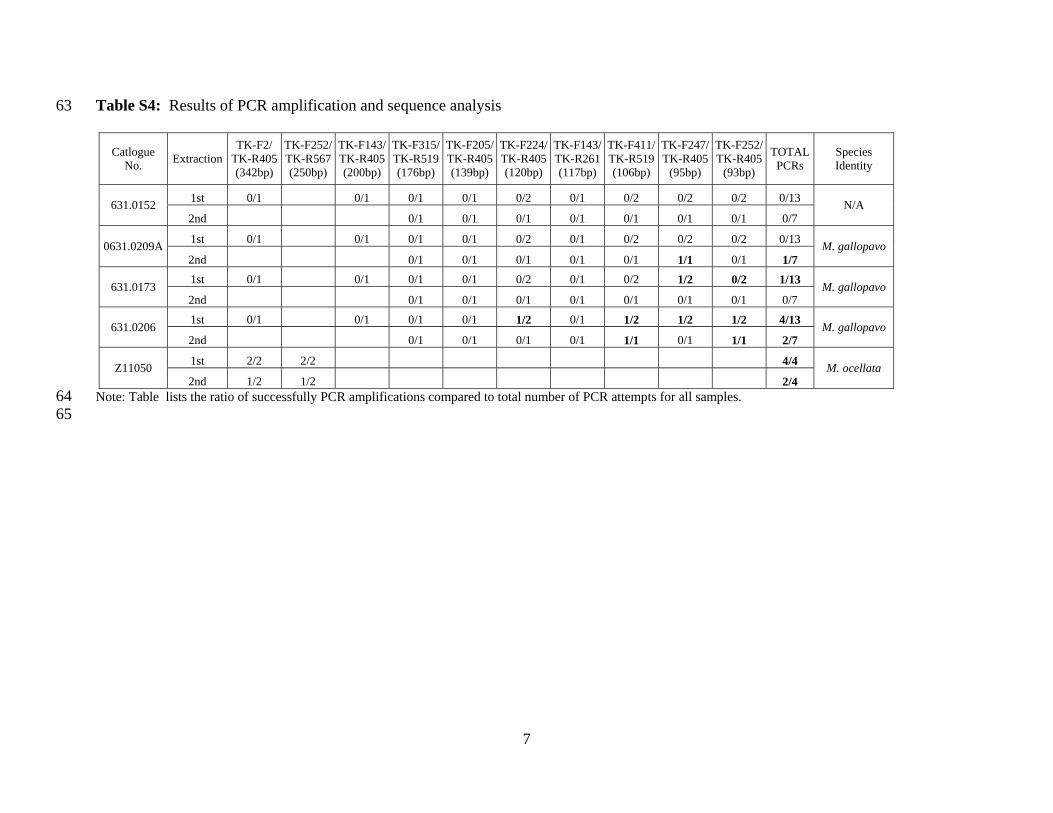
analysis. Nonetheless, numerous PCR amplifications successfully amplified short fragments (93–117
120bp) of Meleagris DNA using a combination of different primer sets (Table S3). A total of 80 118
PCR amplifications were conducted on the ancient bone samples, eight of which yielded PCR 119
amplifications and sequences of expected length (Table S4). While three of the bones produced 120
at least one short Meleagris DNA sequence, only one bone yielded replicable DNA sequences 121
using multiple primer sets. Two different fragments of control region mitochondrial DNA 122
(mtDNA) (121bp and 106bp respectively) were successfully amplified and replicated from both 123
the initial and repeat extraction. All obtained mtDNA sequences matched most closely or 124
identically with M. gallopavo sequences, and were considerably different from those of M. 125
ocellata (Table S5), confirming the species identity as M. gallopavo. 126
127
Discussion 128
Mesoamerican Preclassic Trade Connections 129

7
The combined morphological, osteometric and aDNA evidence confirm the early 130
presence of M. gallopavo at El Mirador in the Maya lowlands. The presence of male, female and 131
subadult turkeys, some with reduced flight capabilities, suggests that the introduced birds were 132
captive reared and/or domesticated. Although we do not yet know the immediate source of the 133
turkey bones, the most reasonable explanation is that a few Mexican turkeys entered the site as 134
exchange goods directly from central/northern Mexico suggesting a close Late Preclassic 135
association between El Mirador and contemporary northern Mesoamerican cultures at sites such 136
as Teotihuacan. Most of the evidence for the exchange of goods and ideas between central 137
Mexico and the Maya region dates to the Classic period several centuries later (ca. AD 250–900) 138
[15]. The El Mirador turkeys therefore add to a relatively sparse record of Preclassic cultural and 139
material exchange between the Maya lowlands and northern Mesoamerica [16–18]. Prior 140
information on Preclassic exchange comes primarily from non-perishable goods such as obsidian 141
and ceramics so the non-local turkeys at El Mirador expand our understanding of the types of 142
goods that were exchanged long distances during this early period of Maya history. Although 143
The El Mirador turkey specimens could represent the transport of dried meat or partial carcasses, 144
the presence of associated upper and lower limb bones suggests that the animals were imported 145
whole and possibly live. The imported turkeys further emphasize that El Mirador’s vast Late 146
Preclassic trade connections extended some 1000 kilometers north into central Mexico, in 147
addition to the site’s better known connections with the Atlantic and Pacific coasts and the Maya 148
highlands [14,19,20]. El Mirador’s participation in interregional trade and cultural interaction 149
was likely pivotal to the site’s accumulation of political and economic power during the Late 150
Preclassic [20,21]. 151
152

8
Timing of Mesoamerican Turkey Domestication and Diffusion 153
The Late Preclassic presence of M. gallopavo in the Maya region has important 154
implications for documenting the timing of Mesoamerican turkey domestication and diffusion. 155
To date, morphological and genetic studies have been unable to distinguish between wild and 156
early domestic forms of M. gallopavo. In the absence of morphological and genetic markers, 157
archaeologists have relied on indirect evidence of domestication such as the presence of pen 158
structures, egg shells and neonates or appearance of the species outside its presumed natural 159
geographic range. Previous to our study, all indirect evidence for Mesoamerican turkey 160
husbandry dated to the Classic period or later [8,22,23]. Since the Preclassic El Mirador turkeys 161
represent movement of M. gallopavo outside its natural geographic range, the specimens 162
represent the earliest indirect evidence of captive turkey rearing or domestication in 163
Mesoamerica. A Preclassic origin for Mexican turkey domestication has been suggested 164
previously [11,24], but archaeological evidence has been lacking. The El Mirador turkey 165
specimens confirm that turkey domestication, or at least captive rearing, dates to the Preclassic. 166
Determining when Mesoamerican cultures started experimenting with turkey rearing and 167
domestication is vital to the larger question of whether the origins of New World turkey 168
husbandry should be attributed to cultures of the American Southwest or Mesoamerica. It was 169
originally believed that the turkey was first domesticated in Mesoamerica and then introduced in 170
domestic form to the American Southwest [1]. More recent archaeological and genetic evidence 171
has overturned this scenario demonstrating that turkeys were independently domesticated in 172
these two regions although the timing of domestication remains unclear [9,25,26]. It is possible 173
that the idea for turkey rearing or husbandry prior to domestication also arose independently in 174
the American Southwest and Mesoamerica. However, the wealth of documented cultural and 175

9
material exchange between the regions supports a model of diffusion for the concept of turkey 176
husbandry as part of early exchange networks. The Late Preclassic El Mirador M. gallopavo 177
specimens provide evidence for the antiquity of Mesoamerican turkey rearing, and support the 178
probable role of interregional turkey exchange in the diffusion of ideas about animal 179
management in the New World. 180
The presence of Late Preclassic Mexican turkeys at El Mirador also confirms that this 181
non-local species was introduced to the Maya region over one thousand years earlier than 182
previously thought. If the El Mirador turkeys are isolated examples of imported captive-183
raised/domestic Mexican turkeys, it raises the question of why the Maya did not broadly adopt 184
the idea of turkey rearing or domestication until the Postclassic. One possibility is that turkey 185
domestication was not widespread or common in any part of Mesoamerica until the later half of 186
the Classic period despite its potentially earlier origins. This suggestion is supported by the 187
relative rarity of M. gallopavo specimens in Preclassic/Formative central Mexican faunal 188
assemblages, and their increasing frequency in Classic and Postclassic deposits [11,22]. An 189
alternative explanation is related to the nature of the Postclassic Maya economy. During the 190
Postclassic, long-distance trade between central Mexico and the Maya area increased with the 191
expansion of maritime trade routes around the Yucatan Peninsula between the Gulf of Mexico 192
and Central America’s Caribbean coast [27,28]. Increased Postclassic exchange throughout 193
Mesoamerica could have facilitated the dispersal of domesticated turkeys to the Maya area 194
through repeated introductions of breeding pairs and transmission of rearing information. In 195
contrast, the rare earlier introduction of the bird might not have been sufficient to fully 196
incorporate the species into the Maya economy. 197

10
Although the El Mirador turkeys may represent isolated imports, it is also possible that 198
M. gallopavo has been under-identified in Preclassic and Classic Maya zooarchaeological 199
assemblages since M. gallopavo and M. ocellata can be difficult to differentiate morphologically 200
when preservation is poor. Researchers also may not have considered the possible presence of M. 201
gallopavo in earlier assemblages due to the longstanding belief that they were not introduced to 202
the Maya region until the Postclassic. It is essential to determine whether the Mexican turkey 203
appeared in the Maya region earlier than previously understood because an earlier introduction 204
would have provided a second domestic vertebrate during the Late Preclassic to Classic period of 205
Maya population expansion and increasing social complexity. During the Preclassic, the Maya 206
relied extensively on the domestic dog (Canis lupus familiaris), which they used for both dietary 207
and ritual purposes [29], although perhaps primarily for ceremonies related to elite display and 208
power negotiations [29,30]. The turkey was another important food and ritual animal among the 209
Maya [31]. Prior models suggest that only local, wild ocellated turkeys were used through the 210
Classic period, but an early demand for domesticated or captive-reared turkey (i.e., M. 211
gallopavo) could have been related to increased elite ceremonial and status-displaying activities 212
as well as the need for meat to feed growing populations during the Late Preclassic to Classic 213
period of population growth and cultural florescence. 214
215
Conclusions 216
Combined zooarchaeological and aDNA analyses identified the earliest non-local 217
Mexican turkey remains in the Maya cultural region at the site of El Mirador. Prior to this 218
discovery, the earliest evidence of M. gallopavo in the Maya area dated to approximately one 219
thousand years later. The El Mirador turkeys may represent rare, or isolated imports from 220

11
central/northern Mexico, but it is also possible that captive/domestic Mexican turkey husbandry 221
was practiced by the ancient Maya much earlier than previously thought. The Maya may 222
therefore have had access to another domestic vertebrate, besides the dog, during the Late 223
Preclassic to Classic period of population expansion and increasing social complexity. 224
Significantly, the El Mirador turkeys also provide the earliest indirect evidence of M. gallopavo 225
captive rearing or domestication in Mesoamerica. Previously, all other indirect evidence of 226
husbandry (e.g., pen structures, egg shells and neonates, or the appearance of the species outside 227
its natural geographic range) dated to the Classic period or later [8,22,23]. 228
The early presence of M. gallopavo at Late Preclassic El Mirador demonstrates a need to 229
reassess the timing of turkey domestication and diffusion in Mesoamerica. Understanding when 230
the Mexican turkey was domesticated and when it was introduced to and fully adopted by the 231
ancient Maya has important consequences for understanding Mesoamerica subsistence systems 232
and long-distance trade connections. The topic also has broader ramifications with respect to the 233
process and timing of New World animal domestication, and the culture-specific motivations for 234
incorporating or not incorporating potential domesticates or managed species into ancient social 235
and economic systems. 236
237
Materials and Methods 238
Zooarchaeology and Osteometrics 239
The archaeological turkey bones were identified within a larger zooarchaeological 240
assemblage from the site (number of identified specimens = 3470). The sample also contained 241
other bird bones that we could only identify to the level of taxonomic subclass (Aves) because 242
they were undiagnostic elements or poorly preserved. Nearly all of the unidentified bird remains 243

12
come from large-bodied species, and some of these may represent additional M. gallopavo 244
elements. 245
Zooarchaeological specimens were identified through comparison with modern skeletons 246
housed in the Florida Museum of Natural History Environmental Archaeology and Ornithology 247
collections (www.flmnh.ufl.edu/museum/collections.htm). Turkey age and sex determinations 248
were based on skeletal element size, osteometrics [6,32], and tarsometatarsus spur morphology. 249
Archaeological bones were measured using standard osteometric measurements and were 250
compared to published metric data available for M. gallopavo and M. ocellata [6: Tables 10, 11, 251
14, 15, 20, 21]. 252
253
Ancient DNA Analysis 254
The four archaeological bird bones were processed in the Ancient DNA Laboratory 255
located in the Department of Archaeology at Simon Fraser University. The ancient DNA 256
laboratory is specifically designed for and dedicated to ancient DNA work - no modern DNA 257
samples have ever been processed in the lab. The lab is equipped with a UV filtered ventilation 258
and positive airflow, with dedicated equipment and bench UV lights. Strict contamination 259
control protocols are followed in the lab, including: 1) the use of protective clothing including 260
Tyvex™ suits, gloves, masks, etc.; 2) the separation of the pre- and post-PCR work (located in 261
two buildings with separate ventilation systems); and 3) the inclusion of multiple blank DNA 262
extractions and negative PCR controls. 263
Two separate DNA extractions were conducted for each bone, with the repeat extractions 264
occurring several months after the initial extractions. For both extractions, the analyzed bone 265
samples weighed approximately 0.5g. Bone samples were subjected to rigorous chemical 266

13
decontamination in order to remove possible surface contamination [33]. The samples were 267
immersed in a 6% sodium hypochlorite solution for 7 minutes, followed by immersion in 1 N 268
HCl solution for 30–60 seconds, then immersion in 1N NaOH for 30–60 seconds, before being 269
rinsed twice in ultra-pure water and UV irradiated in a crosslinker for 30 minutes on two sides. 270
The samples were crushed into powder using a liquid nitrogen grinding mill (6750 SPEX 271
CertiPrep Freezer/Mill). Three additional ancient turkey bones were included in analysis to act as 272
positive controls for both the initial and repeat extractions, as well as the subsequent PCR 273
reaction sets. These three bones were recovered from archaeological sites in Arizona (ca. AD 274
1100–1300) [9] and were processed separately from the El Mirador samples. DNA extraction 275
was performed using a modified silica-spin column technique [9,34], and approximately 100 µl 276
of DNA solution was collected for each sample. 277
PCR amplifications were conducted in a Mastercycler Personal (Eppendorf, Hamburg, 278
Germany) in a 30–50 µL reaction volume containing 50 mM KCl, 10 mM Tris-HCl, 2.5 mM 279
MgCl2, 0.2 mM dNTP, 1.0 mg/ml BSA, 0.3 µM each primer, 3.0–5.0 µl DNA sample and 2.5–280
3.5 U AmpliTaq Gold™ LD (Applied Biosystems). Primers were designed to target fragments of 281
Meleagris mitochondrial DNA of various lengths. Several different primer sets were tested 282
(Table S3). PCR began with an initial 12 minute denaturing period at 95°C, followed by 60 283
cycles at 94°C for 30 seconds (denaturing), 52°C for 30 seconds (annealing), and 72°C extension 284
for 40 seconds. Blank extracts and negative controls were included in each of the PCR reaction 285
sets. Ancient positive controls (Arizona archaeological turkey bone extracts) were also tested to 286
ensure the efficacy of the primer sets and PCR conditions. 287
Five µL of PCR product were visualized via electrophoresis on a 2% agarose gel using 288
SYBR Green™ staining, Successfully amplified PCR products of expected length were purified 289

14
using MinElute™ purification kits (Qiagen, Valencia, CA). Purified products were sequenced 290
using both forward and reverse primers at the Central Facility of the Institute for Molecular 291
Biology and Biotechnology Laboratory at McMaster University (using an ABI 3100) and at 292
Macrogen, Seoul, Korea (ABI 3730XL). The obtained electropherograms were edited, aligned 293
and compiled using ChromasPro software (www.technelysium.com.au). Consensus sequences 294
were developed based on multiple PCR amplifications and sequencing. 295
Once the DNA analysis of the ancient samples was completed, DNA was extracted from 296
a modern M. ocellata phalanx collected from Guatemala (FLMNH catalog number Z11050, 297
Table S4). The M. ocellata samples were processed in the SFU Center for Forensic Research in a 298
lab dedicated to DNA analysis of modern or forensic bone samples. Two 0.5g bone samples 299
were extracted using methods similar to those listed above. The DNA extracts were PCR 300
amplified using primers TK-F2/TK-R405 and TK-F252/TK-R567 (Table S3) and produced 301
amplicons of 254bp and 338bp in length, respectively. The PCR products were sequenced from 302
both directions and consensus sequences matched identically with the GenBank M. ocellata 303
reference sequence AF487120. 304
305
Ancient DNA Extraction Results and Authenticity 306
Sequences were obtained for three (631.0209, 631.0173, 631.0206) of the four bone 307
samples. Only one bone yielded replicable sequences (631.0206) using different primer sets and 308
using both the initial and repeated DNA extracts (Table S4). The obtained ancient DNA 309
sequences were BLAST-compared through GenBank to determine if they would match 310
Meleagris sequences and to ensure that they did not match with any other unexpected species or 311
sequences. Multiple alignments of the sample sequences and published Meleagris mtDNA 312

15
reference sequences were conducted using ClustalW [35] in order to confirm the species 313
identifications. 314
Two replicable DNA fragments were obtained for sample 631.0206 totaling 71bp and 315
55bp respectively once the primer sequences were removed. The two fragments correspond to 316
positions 15731–15800, and 15858–15913 on the M. gallopavo mitochondrial genome (reference 317
NC010195). Despite their short lengths, the sequences clearly match most closely with M. 318
gallopavo rather than M. ocellata, as demonstrated by the multiple alignment (Table S5). 319
Due to the antiquity of the samples and the tropical climate from which they were 320
recovered, a low success rate for DNA extraction and amplification is expected, and provides 321
support for the authenticity of the recovered sequences. The successfully reproduced sample, the 322
carpometacarpus fragment, was the largest of the four bone samples and the best preserved 323
morphologically [36]. PCR targeted fragments of mtDNA varying in length from <100 to 400bp. 324
Only short fragments of DNA could be amplified despite repeat amplifications with longer 325
primer sets designed to detect contamination from modern sources [37]. Multiple blank 326
extractions and negative PCR controls were included in the study, none of which yielded DNA 327
fragments of expected length. Successful amplification of three positive controls (ancient turkey 328
bones from Arizona) demonstrated the efficacy of both the extraction method and PCR primers 329
[9]. 330
The retrieved sequences matched very closely or identically with modern turkey 331
reference sequences in GenBank, and therefore cannot be used to definitively rule out the 332
possibility of contamination from modern sources. However, considering the short length of the 333
retrieved sequences significant differences between the ancient and modern turkeys were not 334
expected. Moreover, the primer sets were designed to target areas maximizing differences 335

16
between M. gallopavo and M. ocellata, rather than polymorphic sites within the M. gallopavo 336
control region. The retrieved sequences themselves, including those unreplicated sequences, 337
demonstrate significant post-mortem damage characterized by C→T transitions (Figure S1). 338
These DNA transitions were likely caused by hydrolytic damage and are anticipated to occur in 339
ancient sequences [37,38]. Finally, the DNA identification of M. gallopavo supports the 340
morphological and osteometric identification of the bones 341
Acknowledgments 342
FLMNH provided laboratory space and access to modern comparative specimens. Ashley Sharpe 343
and Deanne Matheny assisted with manuscript preparation. 344
345
References 346
1. Leopold AS (1959) Wildlife of Mexico: the game birds and mammals. Berkeley: University of 347
California Press. 581 p. 348
349
2. Schorger AW (1966) The wild turkey: its history and domestication. Norman: University of 350
Oklahoma Press. 625 p. 351
352
3. Howell SNG, Webb S (1995) A guide to the birds of Mexico and northern Central America. 353
New York: Oxford University Press. 1010 p. 354
355
4. Camacho-Escobar MA, Jiménez-Hidalgo E, Arroyo-Ledezma J, Sánchez-Bernal EI, Pérez-356
Lara E (2011) Natural history, domestication and distribution of the turkey (Meleagris 357
gallopavo) in Mexico. Revista Universidad y Ciencia, 27: 351–360. 358

17
359
5. Corona-M E (2002) The Pleistocene bird record of México. Acta Zoologica Cracoviensia 360
45(special issue): 293–306. 361
362
6. Steadman DW (1980) A review of the osteology and paleontology of turkeys (Aves: 363
Meleagridae). Contribution of the Science and Natural History Museum of Los Angeles County 364
California 330: 131–207. 365
366
7. Steadman DW, Stull JA, Eaton SW (1979) Natural history of the ocellated turkey. World 367
Pheasant Assoc. J. 4: 15–37. 368
369
8. Hamblin NL (1984) Animal use by the Cozumel Maya. Tuscon: University of Arizona Press. 370
206 p. 371
372
9. Speller CF, Kemp BM, Wyatt SD, Monroe C, Lipe WD, et al. (2010) Ancient mitochondrial 373
DNA analysis reveals complexity of indigenous North American turkey domestication. Proc. 374
Natl. Acad. Sci. U.S.A. 107: 2807–2812. 375
376
10. Álvarez T, Ocaña A (1999) Sinopsis de restos arqueo-zoológicos de vertebrados terrestres - 377
basada en informes del laboratorio de paleozoología del INAH. Mexico City: INAH Colección 378
Científica. 108 p. 379
380

18
11. Flannery KV (1967) Vertebrate fauna and hunting patterns. In Byers DS, editor. The 381
prehistory of the Tehuacan Valley, volume 1. Austin: University of Texas Press. pp. 132–177. 382
383
12. Middleton WD, Feinman GM, Nicholas LM (2002) Domestic faunal assemblages from the 384
Classic period Valley of Oaxaca, Mexico: a perspective on the subsistence and craft economies. 385
J. Archaeol. Sci. 29: 233–249. 386
387
13. Götz CM (2008) Coastal and inland patterns of faunal exploitation in the prehispanic 388
northern Maya lowlands. Quat. Int. 191: 154–169. 389
390
14. Hansen RD (1990) Excavations in the Tigre Complex El Mirador, Peten, Guatemala. Provo, 391
UT: New World Archaeological Foundation. 308 p. 392
393
15. Braswell GE, editor (2003) The Maya and Teotihuacan: reinterpreting Early Classic 394
interaction. Austin: University of Texas Press. 441 p. 395
396
16. Moholy-Nagy H, Nelson FW (1990) New data on sources of obsidian from Tikal, 397
Guatemala. Ancient Mesoam. 1: 71–80. 398
399
17. Pendergast DM (1971) Evidence of early Teotihucan-lowland Maya contact at Altun Ha. 400
Am. Antiq. 36: 455–460. 401
402

19
18. Clayton SC (2005) Interregional relationships in Mesoamerica: interpreting Maya ceramics at 403
Teotihuacan. Lat. Am. Antiq. 16: 427–448. 404
405
19. Neff H, Bishop RL, Arnold DE (1990) A reexamination of the compositional affiliations of 406
Formative period whiteware from highland Guatemala. Ancient Mesoam. 1: 171–180. 407
408
20. Matheny RT, Matheny DG (2011) Introduction to Investigations at El Mirador, Peten, 409
Guatemala. El Mirador Series, Part 1. Provo, UT: New World Archaeological Foundation, 410
Brigham Young University. 240 p. 411
412
21. Reese-Taylor K, Walker DS (2002) The passage of the Late Preclassic into the Early Classic. 413
In Masson M, Freidel DA, editors. Ancient Maya political economies. New York: AltaMira 414
Press. pp. 87-122. 415
416
22. Lapham HA, Feinman GM, Nicholas LM (in press) Animal economies in prehispanic 417
southern Mexico. In Götz C, Emery KF, editors. Archaeology of Ancient Mesoamerican 418
Animals. Oxford: Oxbow Press, David Brown Book Company. 419
23. Storey R (1992) Life and death in the ancient city of Teotihuacan: a modern 420
paleodemographic synthesis. Tuscaloosa: University of Alabama Press. 328 p. 421
422
24. Valadez AR (2003) Domesticación y zootecnia en el México antiguo. Imagen Veterinaria 3: 423
32-45. 424

20
425
25. Breitburg E (1993) The evolution of turkey domestication in the greater Southwest and 426
Mesoamerica. In Woosley AI, Revesloot JC, editors. Culture and contact: Charles C. Di Peso’s 427
gran chichimeca. Albuquerque: University of New Mexico Press. pp. 153-172. 428
429
26. McKusick CR (2001) Southwest birds of sacrifice. Tucson: Arizona Archaeological Society. 430
208 p. 431
432
27. Guderjan TH, Garber JF (1995) Maya maritime trade, settlement, and populations on 433
Ambergris Caye, Belize. San Antonio, TX: Maya Research Program and Labyrinthos. 201 p. 434
435
28. McKillop H, Healy PF, editors (1989) Coastal Maya trade. Peterborough, ON: Trent 436
University Occasional Papers in Anthropology, No. 8. 189 p. 437
438
29. Emery KF (2004) In search of the "Maya" diet: is regional comparison possible in the Maya 439
area? Archaeofauna 13: 37–56. 440
441
30. Teeter WG (2004) Animal utilization in a growing city, vertebrate exploitation at Caracol, 442
Belize. In Emery KF, editor. Maya zooarchaeology: new directions in method and theory. Los 443
Angeles: Institute of Archaeology, UCLA Press. pp 177–191. 444
445

21
31. Pohl MD, Feldman LH (1982) The traditional role of women and animals in lowland Maya 446
economy. In Flannery KV, editor. Maya subsistence: studies in memory of Dennis E. Puleston. 447
New York: Academic Press. pp 295–311. 448
449
32. von den Driesch A (1976) A guide to the measurement of animal bones from archaeological 450
sites. Cambridge, MA: Peabody Museum of Archaeology and Ethnology, Harvard University. 451
137 p. 452
453
33. Yang DY, Cannon A, Saunders SR (2004) DNA species identification of archaeological 454
salmon bone from the Pacific Northwest Coast of North America. J. Archaeol. Sci. 31: 619–631. 455
456
34. Yang DY, Eng B, Waye JS, Dudar JC, Saunders SR (1998) Improved DNA extraction from 457
ancient bones using silica-based spin columns. Am. J. Phys. Anthro. 105: 539–543. 458
459
35. Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W: improving the sensitivity of 460
progressive multiples sequence alignments through sequence weighting, position-specific gap 461
penalties and weight matrix choice. Nucl. Acids Res. 22: 4673–4680. 462
463
36. Haynes S, Searle JB, Bretman A, Dobney KM (2002) Bone preservation and ancient DNA: 464
the application of screening methods for predicting DNA survival. J. Archaeol. Sci. 29: 585–592. 465
466
37. Paabo S (1989) Ancient DNA: extraction, characterization, molecular-cloning, and 467
enzymatic amplification. Proc. Natl. Acad. Sci. U.S.A. 86: 1939–1943. 468

22
469
38. Gilbert MT, Binladen J, Miller W, Wiuf C, Willersley E, et al. (2007) Recharacterization of 470
ancient DNA miscoding lesions: insights in the era of sequencing-by-synthesis. Nucl. Acids Res. 471
35: 1–10. 472
473

23
Figure Legends 474
475
Figure 1: Pre-Hispanic range of M. ocellata and M. gallopavo [2,6] in Mesoamerica, and 476
location of discussed archaeological sites. 477
478
Figure 2: El Tigre Complex showing structures containing turkey bones (circled). Redrawn from 479
original by B. Dahlin. 480
481
Figure 3: Archaeological turkey specimens compared with modern M. gallopavo and M. 482
ocellata: A) right ulnae, and B) left tarsometatarsi. 483
484
Figure 4: Archaeological turkey () osteometrics compared with mean (±2 standard deviations) 485
M. gallopavo (male , female ); and M. ocellata (male , female ) [6]. 486
487

24
Table 1: Provenience dates and zooarchaeological, aDNA, and osteometric identifications of the El Mirador turkey specimens.
Catalog no. Provenience AMS date (calibrated)a
Zooarchaeological identification
aDNA identification
Osteometric identification
Element Sex/Age
631.0209A 26O-25/27 200 BC–AD 3 M. gallopavo M. gallopavo M. gallopavo ulna male
631.0173 26J-4 - M. gallopavo M. gallopavo M. gallopavo tarsometatarsus male (subadult)
631.0152 26J-14 186 BC–AD 54 M. gallopavo no amplification M. gallopavo ulna male
631.0206 35B-5 327–204 BC M. cf. gallopavo M. gallopavob inconclusive carpometacarpus -
631.0210 26O-25/27 200 BC–AD 3 M. cf. gallopavo not tested inconclusive carpometacarpus -
631.0209B 26O-25/27 200 BC–AD 3 M. gallopavo not tested - tarsometatarsus female
631.0341 26K-4 - Meleagris. sp. not tested - femur - a AMS dates from zooarchaeological specimens found in association with the turkey bones (Table S2) b aDNA identification was confirmed through repeat extractions and amplification





1
Supporting Information (SI) 1
El Mirador: excavation history, provenience descriptions, and dating 2
El Mirador, an ancient Maya city located deep in the tropical forest of the northern area 3
of the Department of Peten, Guatemala is famous for its Preclassic cultural remains (Table S1). 4
In 1932 the 16th Carnegie Institution Central American Expedition briefly visited the site. Years 5
later many of the largest architectural features of the site were mapped by Ian Graham and some 6
areas briefly tested by Joyce Marcus; a larger program of excavation and mapping was carried 7
out by Bruce Dahlin and Ray Matheny from 1978–1983, and included the Harvard El Mirador 8
Project in 1982. Excavations were resumed in 2003 by Richard Hansen and are ongoing as part 9
of what is known as the Mirador Basin investigations. The archaeological turkey specimens 10
reported here along with other archaeological animal remains were recovered from excavations 11
between 1980 and 1982 (Fig. 2). These materials were housed by the Brigham Young University 12
Museum of Peoples and Cultures prior to zooarchaeological analysis of the El Mirador animal 13
remains at FLMNH-EAP, which began in 2003. 14
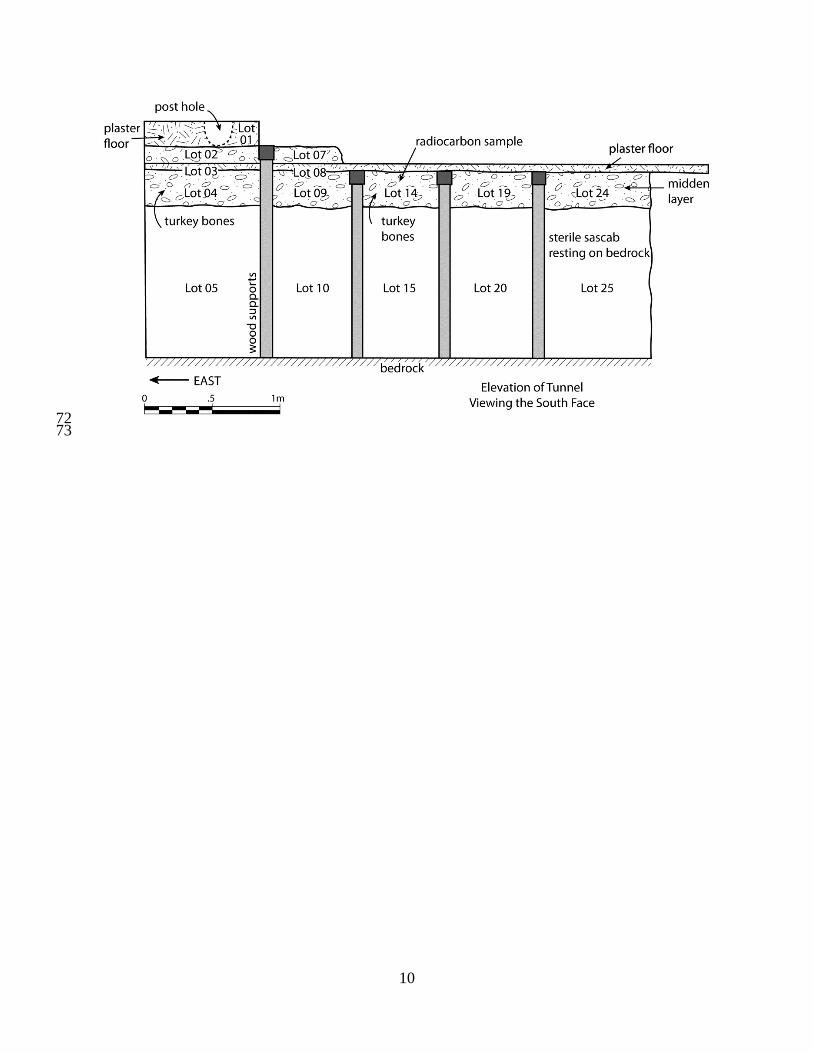
All of the turkey bones recovered in the 1980s came from investigations in the Late 15
Preclassic Tigre Complex, whose major focus is the 56 m tall Tigre Pyramid. Operation 26J, 16
conducted by Hansen [14] under supervision by Matheny, was a tunnel placed under an ancient 17
plaster floor exposed at the base of the east side of the Jaguar Paw Temple. This floor was of 18
interest because it appeared to run under the building platform. Hansen probed under the floor 19
with a 3.7 m tunnel under a second continuous floor (below the first) that sealed a midden layer 20
containing bones, pieces of charcoal, ceramics, and other cultural materials; turkey bones were 21
found in Lots 04 and 14, and radiocarbon sample Beta 241842 was recovered from Lot 14 (Fig. 22
S2). Hansen [14: Table 4] classified all ceramics from the lots below the lower floor as 23

2
Preclassic. The midden layer may have been in place prior to floor construction or brought in 24
from elsewhere as floor preparation. Below the midden layer excavation continued through a 25
substantial layer of sterile sascab (deteriorated limestone) lying on bedrock. 26
Operation 26O was an exploratory 33 m tunnel into the heart of the Jaguar Paw Temple 27
[14: 70–73]. The turkey bones, and the mammal bone used for radiocarbon sample Beta 241844, 28
were found in contiguous Lots 25/27. The specimens were found about 24 meters into the 29
structure, almost directly under the center of the temple above (Fig. S3). The context of the 30
bones was sealed in antiquity by Late Preclassic construction of the temple that also covered 31
Middle Preclassic architectural remains. 32
Operations 35 A and B were in Structure 4D2-1, a prominent 8 m high building located 33
on the east side of the Tigre Plaza, directly east of the main Tigre Pyramid stairway (Fig. 1). This 34
building is the central construction of a triadic group. Operation 35A included the excavation of 35
two large chambers on the summit of the building, including the central room [14, Fig. S4]. 36
Excavations continued below what may have been the latest floor in the room. In Operation 35B, 37
which was a continuation of Operation 35A to a deeper level in the southern portion of the 38
central room, plaster floors were found at -3.18 m and at -4.7 m below datum. The excavations 39
continued beneath the lower plaster floor to -9.05 m in Lot 05 where radiocarbon sample Beta 40
241843 and turkey bones were found. Of an estimated 400 potsherds from Operation 35B, 41
Hansen [14] reported that all were from the early Late Preclassic period in a sealed context. 42
43

3
Figure Legends (Supporting Information) 44
Figure S1: Multiple alignments of obtained sequences demonstrating DNA damage induced 45
transitions. 46
47
Figure S2: Operation 26J baulk profile. Redrawn after [14: Fig. 29]. 48
49
Figure S3: Operation 26O baulk and tunnel profile. Redrawn after [14: Fig. 44]. 50
51
Figure S4: Operations 35A and 35B wall and baulk profile. Redrawn after [14: Fig. 56].52

4
Table S1: Generalized chronology used in the text 53
Mesoamerican cultural period Approximate dates
Early Preclassic/Formative 2000–1000 BC
Middle Preclassic/Formative 1000–300 BC
Late Preclassic/Formative 300 BC–AD 100
Terminal Preclassic/Protoclassic AD 100–250
Early Classic AD 250–600
Late Classic AD 600–800
Terminal Classic AD 800–1000
Postclassic AD 1000–1500
54 55

5
Table S2: AMS Radiocarbon ages from zooarchaeological remains found in association with the 56
archaeological turkey bones. 57
Taxa (scientific name)
Beta Analytic lab no.
Provenience (Op. no.)
Uncalibrated ages (years BP)
2 Sigma calibrated radiocarbon ages (INTCAL09 calendar years)
Odocoileus virginianus Beta-241842 26J-14 2050 +/- 50 186 BC–AD 54 Mammalia Beta-241843 35B-5 2250 +/- 40 330 BC–204 BC Mammalia Beta-241844 26O-25/27 2080 +/- 40 200 BC–AD 3
58

6
Table S3: Meleagris primers for PCR amplification 59
Primer name Direction Coordinates§ Sequence (5' to 3')
TK-F2* Forward 15482–15505 AATTTATTCCCGCTTGGATAAGCC TK-F143* Forward 15624–15650 GCATAATCGTGCATACATTTATATACC TK-F205 Forward 15685-15712 CGTACTAAACCCATTATATGTARACGGA TK-F224* Forward 15704–15729 GTAGACGGACATAACAACCTTTACCCC TK-F247 Forward 15729–15754 CCCATTYCTCCCTAATGACTACTCC TK-F252 Forward 15731-15755 CCATTTCTCCCACAATGACTACTCC TK-F315* Forward 15759–15782 ACATGCCAATGACATTAACTCCTTC TK-F411* Forward 15829–15854 TGGTTACAGGACATACCTCTAAATCT TK-R261* Reverse 15718–15741 AGGGAGRAATGGGGTAAAGGTTGT TK-R405* Reverse 15801–15824 TGTATATGGTCTCTTGRGGGTTGG TK-R519 Reverse 15914–15935 GGGTTGGTGATCTCTCGTGAGG TK-R567* Reverse 15962–15981 GGGAAAGAATGGGCCTGAAG * Previously published results [9] 60 § Coordinates numbered according to GenBank reference sequence accession EF153719. 61 62
7
Table S4: Results of PCR amplification and sequence analysis 63
Catlogue No.
Extraction TK-F2/
TK-R405 (342bp)
TK-F252/ TK-R567 (250bp)
TK-F143/ TK-R405 (200bp)
TK-F315/ TK-R519 (176bp)
TK-F205/ TK-R405 (139bp)
TK-F224/ TK-R405 (120bp)
TK-F143/ TK-R261 (117bp)
TK-F411/ TK-R519 (106bp)
TK-F247/ TK-R405
(95bp)
TK-F252/ TK-R405
(93bp)
TOTAL PCRs
Species Identity
631.0152 1st 0/1 0/1 0/1 0/1 0/2 0/1 0/2 0/2 0/2 0/13
N/A 2nd 0/1 0/1 0/1 0/1 0/1 0/1 0/1 0/7
0631.0209A 1st 0/1 0/1 0/1 0/1 0/2 0/1 0/2 0/2 0/2 0/13
M. gallopavo 2nd 0/1 0/1 0/1 0/1 0/1 1/1 0/1 1/7
631.0173 1st 0/1 0/1 0/1 0/1 0/2 0/1 0/2 1/2 0/2 1/13
M. gallopavo 2nd 0/1 0/1 0/1 0/1 0/1 0/1 0/1 0/7
631.0206 1st 0/1 0/1 0/1 0/1 1/2 0/1 1/2 1/2 1/2 4/13
M. gallopavo 2nd 0/1 0/1 0/1 0/1 1/1 0/1 1/1 2/7
Z11050 1st 2/2 2/2 4/4
M. ocellata 2nd 1/2 1/2 2/4
Note: Table lists the ratio of successfully PCR amplifications compared to total number of PCR attempts for all samples. 64 65

8
Table S5: Multiple alignment of M. gallopavo and M. ocellata control-region reference 66
sequences, with the retrieved ancient sequence 67
1574
0
1574
1
1574
1.1
1576
0
1576
1
1576
2
1576
2.1
1576
3
1577
6
1577
7
1577
8
1578
8
1578
9
1579
0
1579
1
1579
4
1579
5
1579
6
1579
9
1586
0
1586
3
1588
5
1589
6
NC010195 M. gallopavo C T - C A T - C C T C - C C C A T C T T C A G
AF532414 M. gallopavo A - - . . . G . . . . - . . . . . . . . . . .
AJ297180 M. gallopavo . . - . . . - . . . . - . . . . . . . . . . A
AF486875 M.g.silvestris . . - . . . - . . . . - . . . . . . . . . . .
GQ303165 M.g.gallopavo . . - . . . - . . . . - . . . . . . . . . . .
AF487121 M.ocellata . A C T G C - T T C T C A T A C C T G C G - .
AF487120 M.ocellata . A C T G C - T T C T C A T A C C T G C G - .
M. ocellata (cat #Z11050) . A C T G C - T T C T C A T A C C T G C G - .
Specimen 631.0206 . . - . . . - . . . . - . . . . . . . . . . .
68 69

9
70 71
10
72 73

11
74 75

12
76