ultrastructural study of the blood cells of the beluga whale, delphinapterus leucas
TRANSCRIPT

JOURNAL OF MORPHOLOGY 209:97-110 (1991)
Ultrastructural Study of the Blood Cells of the Beluga Whale, Delphinapterus leucas
CHARLES ROGERS WILLIAMS 111, GEORGE B. CHAPMAN, AND ANDREA S. BLAKE Department of Biology, Georgetown University, Washington, DC 20057
ABSTRACT General cytological and ultrastructural features of cells found in the peripheral blood of three captive beluga whales (Delphinapterus leucas) are described. We noted all major peripheral cell types common to the circulation of mammalian species, including polymorphonuclear neutrophils, eosinophils, basophils, lymphocytes, and monocytes, as well as myelocytes, erythroblasts, and plasma cells. Platelets and two populations (normal and hypochromic) of red blood corpuscles were observed.
The first morphological observations of be- luga whale (Delphinapterus leucas) blood were carried out by Quay (’54) on three whales killed by Inuit (Eskimos) at Churchill, Manitoba. Quay was unable to receive his specimens before the whales died. They had been harpooned (marked) and shot. The amount of time that elapsed before sampling is uncertain and recorded as “less than 12 hours” for two of the whales, and as much as 11 days for the other. Since that time, beluga wiale blood has received more standardized scrutiny by Dhindsa et al. (’741, and En- gelhardt (’79). Now, blood is routinely sam- pled from healthy captive individuals and subjected to extensive biochemical analysis. The object of this paper is not to add to the increasing data on cetacean hematology and blood biochemistry, but to continue the pio- neering morphological work of Quay. We have been unable to find previous ultrastructural studies on whale blood, except for the brief report of Taylor and Farrell (’73) on killer whales (Orcinus orca). Therefore, this report is an update of structural analysis on the blood cells of Cetacea, with particular refer- ence to the beluga.
MATERIALS AND METHODS
The three whales (Delphinapterus leucas, Pallas 1776) whose blood is the subject of this study were captured from the Churchill River in Manitoba in 1985 (Illamar and An- ore) and 1987 (Kia). They were kept at the National Aquarium in Baltimore since cap- ture except for a period when Illamar and Anore were lent to the New York Aquarium. Their diet consisted of herring (Alosa aestiua- lis [Mitchill]), capelin (Mullotus uillosus),
some smelt (Osmerus eperlanus [Linnaeus]) and squid, (Loligo pealei), and was supple- mented with seaTAb brand marine mammal vitamins. (The vitamin content of the diet provided these whales was changed since this study began.) The whales were subject to monthly blood tests. They were conditioned to this procedure, and it appeared to cause no undue stress. All three whales were appar- ently healthy at the time of sampling and were asymptomatic.
Approximately 7 ml of blood (provided through the courtesy of the staff of the Na- tional Aquarium in Baltimore) was drawn, by fluke venipuncture, from whales re- strained in a sling for sampling. Samples were immediately placed in heparinized tubes and stored on ice. Within 1 hr the samples were transferred to a laboratory near the captivity tanks and centrifuged (IEC Clinical centrifuge model CL) for 25 min at 700 rpm. The white blood cells (WBC) accumulated in a buffy coat, 1-2 mm thick. The WBC were removed by carefully pipetting out the buffy coat. The cells were transferred to tubes con- taining4.5 ml of dH,O, 4.5 ml double strength 0.1 M sodium cacodylate buffer, and 1 ml monomeric glutaraldehyde 25% (Karnovsky, ’65) for 1.5-2 hr. The samples were then transferred to new tubes containing 5-6 ml of buffer. To accomplish the transfer, the original tubes were agitated slightly so the WBC would enter suspension while the red blood corpuscles (RBC) were left behind. The cells were centrifuged for another 30 min to pack them down and facilitate further treat- ment. Three additional washes of 15 min each, in buffer, were followed by a secondary
o 1991 WILEY-LISS, INC.

98 C. ROGERS WILLIAMS ET AL
fixation in 1% osmium tetroxide (Palade, '52) in buffer for 1.5-2 hr at 6°C. The samples were washed 4 times for 15 min in buffer, and then dehydrated in a graded ethanol series for 15 min at each concentration (25%, 50%, 70%, 80%, 95%, loo%, 100%). To en- sure complete dehydration, the cells were treated with two 15 min baths of propylene oxide. The cells were infiltrated with an accel- erated Epon LX-112 mixture according to Luft ('611, 1:l (Epon LX-112: propylene ox- ide) solution for 8 hr, 3:l for 8 hr, and finally dispensed into Beem Capsules (that were cen- trifuged for 15 min after encapsulation) or embedding trays in the accelerated mixture. Polymerization was for 48 hr at 60°C. The embedded tissue was sectioned using a Sor- vall Porter-Blum MT2-B Ultramicrotome with a DuPont diamond knife. Thin sections (0.5-1.0 p.m) were collected and stained for light microscopy according to the method of Richardson et al. ('60). Ultrathin sections (pale gold) were collected onto uncoated 200- mesh copper grids. The sections were stained in a 5% uranyl acetate in 50% ethanol solu- tion (Gibbons and Grimstone, '60) for 10 min, followed by 0.25% lead citrate solution (Venable and Coggeshall, '65) in a C0,-free environment, for 5 min. The ultrathin sec- tions were examined with an RCA EMU-2D electron microscope.
RESULTS
We observed all of the formed elements typically found in mammalian peripheral blood: polymorphonuclear neutrophils (polys), eosinophils, basophils, lymphocytes, monocytes, platelets, and erythrocytes. In addition, we found, occasionally, in the circu- lating blood, maturational stages in the for- mation of myeloid (metamyelocytes), lym- phoid (plasmablasts, plasma cells), and erythroid (polychromatophilic erythroblasts,
normoblasts, reticulocytes) lines of blood cells, usually associated with marrow or lymphoid tissue. Table 1 consists of our own cell mea- surements, those of Quay ('54), who mea- sured cell diameters from a smear, and text- book values (Bloom and Fawcett, '86) for human blood in most instances measured both from a smear and in suspension.
Granulocyte precursors Metamyelocytes, not usually present in pe-
ripheral blood, were identified (Fig. l) in one of the samples. The determination was based on nuclear heterochromatin patterns, gran- ule composition, and quantity and distribu- tion of rough surfaced endoplasmic reticu- lum (RER). The cell contains a nucleus, nucleolus, rough surfaced endoplasmic retic- ulum, mitochondria, a Golgi zone, and a cen- triole and is, thus, typical of animal cells in general.
Granulocytes Polymorphonuclear neutrophils (Fig. 2)
compose about 62% of the cells examined and their morphology is typical of mammalian neutrophils in general. Developing neutro- phils show parallel stacks of RER, cut trans- versely (Fig. 3) and obliquely (Fig. 4). Band forms (Fig. 5) of immature neutrophils make up 4% of the WBC in a differential count.
Eosinophils (Fig. 6) compose about 8.3% of the WBCs. They usually present a bilobed nucleus, as they do in other mammals, but seldom display the paracrystalline rods found in the specific granules of some mammalian species. When present (Fig. 7) , the rods occur in fusiform granules. Specific granules with- out crystals are usually spherical. Incomplete and multiple crystallizations are also ob- served. The eosinophilic granules are usually nearly spherical and homogeneous, but sev- eral different kinds of intragranular inclu-
TABLE 1. Cell diameters (pm) in human and Beluga whale blood
Beluga Beluga Human Human
(suspension) (smear) (suspension) (smear) maximum diameter (Quay, 54) (Bloom and Fawcett, '86) (Bloom and Fawcett, '86)
Neutrophils 10 17.0 Eosinophils 9 14.5 Basophils 9 17.0 Lymphocytes 9 8.2 Monocytes 10 13.1 Platelets 3 RBC 8.4' 9.0'
-
7 9 -
9-12
10-12 12 10
7-g3 -
17 2-33 - 7 3 -
'Average for 3 whales (n = 25). 'Average taken from the individual with the smallest standard deviation 'Dimension(s) given without reference to type preparation.

ULTRASTRUCTURE OF BELUGA WHALE BLOOD CELLS 99 -
Fig. 1. Ultrastructure of a beluga whale metamyelo- cyte or lymphoblast. Notice the nucleus (N), nucleolus (Nu), rough surfaced endoplasmic reticulum (RER), and mitochondria (M) scattered throughout the cytoplasm. A
sions are seen. Some are circular and appear to be in contact with the limiting membrane of the specific granule (Fig. 6, arrow). These inclusions are slightly more dense than the cytoplasm. A few stand out by their conspicu- ous similarity in size to mitochondria (Fig. 6, double arrow). Still others (not illustrated) are slightly more dense than the granule internum. A concentric lamellar substruc- ture was observed on one occasion next to a central crystal.
Although no basophils were observed in differential counts, five did occur in our blood samples (Figs. 8-11). They vary from having a simple nucleus to demonstrating as many as three lobes in ultrathin section. The most conspicuous feature of these cells is the heter- ogeneity of the cytoplasmic granules (Figs. 9-11). Granules are homogeneous and elec-
Golgi zone (G) surrounds a centriole ( C ) cut in cross section. This cell lacks some of the characteristic hetero- chromatin clumping of mature cells. Bar: 1 pm. x 10,000.
tron dense (Gl), homogeneous and less dense (G2), electron lucent and contain smaller granules (G3), very small and contain a dense core (G4), and contain several dense masses (G5). Crystal and myeloid figures occur in some granules. The cytoplasm contains exten- sive arrays of loosely organized or associated bundles of microfilaments sectioned ob- liquely, longitudinally, and, occasionally, transversely (Fig. 8, Mi).
Agranulocytes Lymphocytes (Fig. 12) compose about
23.6% of the WBCs. They are similar in ap- pearance to human lymphocytes. Often, their cytoplasmic RER is in close ( < 0.5 pm) asso- ciation with mitochondria. They contain a Golgi body oriented near a pair of centrioles. Morphological distinction between T and B
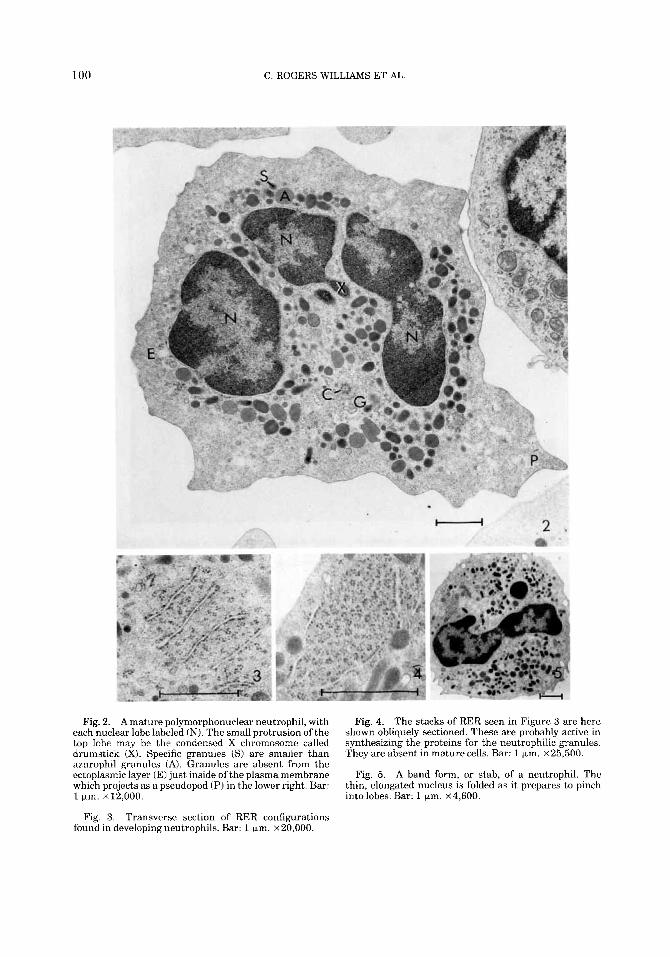
100 C. ROGERS WILLIAMS ET AL.
Fig. 2. A mature polymorphonuclear neutrophil, with each nuclear lobe labeled (N). The small protrusion of the top lobe may be the condensed X chromosome called drumstick (X). Specific granules ( S ) are smaller than azurophil granules (A). Granules are absent from the ectoplasmic layer (E) just inside of the plasma membrane which projects as a pseudopod (P) in the lower right. Bar: 1 pm. ~ 1 2 , 0 0 0 .
Transverse section of RER configurations Fig. 3. found in developingneutrophils. Bar: 1 ym. ~ 2 0 , 0 0 0 .
Fig. 4. The stacks of RER seen in Figure 3 are here shown obliquely sectioned. These are probably active in synthesizing the proteins for the neutrophilic granules. They are absent in mature cells. Bar: 1 pm. ~25 ,500 .
Fig. 5. A band form, or stab, of a neutrophil. The thin, elongated nucleus is folded as it prepares to pinch intolobes. Bar: 1 pm. ~ 4 , 6 0 0 .

ULTRASTRUCTURE OF BELUGA WHALE BLOOD CELLS
.. . .
101
I
-L
Fig. 6. An eosinophilic granulocyte. The nucleus has two lobes (N), and a cytoplasm dominated by eosinophilic specific granules (GI. Also present are small amounts of endoplasmic reticulum and an occasional mitochondrion (MI. Different types of inclusions inside of the specific granules are labeled with arrows. Bar: 1 pm. x 17,500.
Fig. 7. A higher magnification of a different eosino- phi1 to demonstrate the presence of paracrystalline disk- like inclusions or internum (I) which are embedded in the externum (E). Bar: 1 pm. x 20,000.

102 C. ROGERS WILLIAMS ET AL
. , . . .. - '.. .!-
7 . ' ..g
8
Figs. 8-11. Basophils demonstrate a great deal of granule heterogeneity (Gl-G5). Occasionally, a myeloid figure (MD was encountered inside a granule. G4 repre- sents a small granule found in all the basophils. Inclu- sions, such as the crystal (arrow), are common. The
nucleus (N) is either bilobed or bent as it appears here. Notice the microfilaments (Mi) coursing throughout the cytoplasm and the occasional mitochondrion (M). Bar: 1 km. Fig. 8, ~ 1 7 , 0 0 0 ; Fig. 9, ~26 ,000 ; Fig. 10, ~27 ,000 ; Fig. 11, ~ 1 7 , 0 0 0 .

ULTRASTRUCTURE OF BELUGA WHALE BLOOD CELLS 103
2
Fig. 12. The lymphocyte nucleus (N) contains abundant heterochromatin (H), euchromatin (E), and a nucleolus (Nu). Notice how the heterochromatin clumps around the inner surface of the nuclear envelope. The rough surfaced endoplasmic reticulum (RER) is located close to the mitochondria (MI. Bar: 1 Fm. x 18,500.
lymphocytes was not possible. However, plas- mablasts and fully mature plasma cells (Fig. 13) are evident in small numbers, thus con- firming a B cell population. Plasma cells are often in close ( < 1 pm) association with extra- cellular dense spherical granules (Fig. 13G). These granules also occur intracellularly and are membrane limited.
Monocytes (Figs. 14-16) compose only about 2% of the differential count. Occasion- ally, a granule is seen, or a cytoplasmic inclu- sion reminiscent of a nebenkern (Fig. 14, central unlabeled mass), an aggregation of mitochondria found in some invertebrates, especially the male gametes of insects (Wil- son, '24). The monocyte in Figure 15 has phagocytized an entire platelet. Lipid mate-
rial, accumulated by both monocytes and neu- trophils, is also noted (Fig. 16).
Platelets Whale platelets exhibit typical mammalian
ultrastructure. The peripherally located mi- crotubule components of their cytoskeleton are notable (Figs. 17, 18, Mt).
Erythrocytes Normoblasts (Fig. 19) are observed in the
peripheral blood, but an accurate count was not obtained. A polychromatophilic erythro- blast (Fig. 20) reveals several stages (Fig. 20, 11-14) in the presumed pinocytosis of an electron lucent material in coated vesicles. Mitochondria (M) are degenerating and ap-

104 C. ROGERS WILLIAMS ET AL.
Fig. 13. In this fully differentiated B-lymphocyte, called a plasma cell, the distended rough surfaced endoplasmic reticulum (RER) fills the cytoplasm. The associated granules (G) are seen along the plasma membrane at the lower left and right of the figure. M = mitochondrion; N = nucleus. Bar: 1 pm. x 12,500.
pear distended. Beluga whale reticulocytes (Fig. 21) show nothing remarkable to the species.
Two of the whales presented RBCs, the majority of which were typically electron dense (Fig. 23), but a few of which were electron lucent (Figs. 22, 24). The electron lucent corpuscles were located at the inter- face of the packed cells and plasma, upon centrifugation. They were slightly microcytic when compared to normal density cells and had a coarser granularity to their cytoplasm. They often contained very electron dense in- clusions located at the periphery of the cor- puscle and in clumps randomly scattered in the cytoplasm (Fig. 22D). Some of the corpus-
cles in this subpopulation present an irregu- lar shape (Fig. 22); others are typically bicon- cave (Fig. 24).
DISCUSSION
Morphologically, and by differential count, the WBC distribution in the peripheral blood of Delphinapterus leucas is similar to that of terrestrial mammals. This suggests that, while adapting to their aquatic environment, the blood-borne immunological components were qualitatively and quantitatively suffi- cient to meet the different demands.
The differences we found in cell measure- ments from those reported by Quay ('54)

L
ULTRASTRUCTURE OF BELUGA WHALE BLOOD CELLS 105
. ..
. .
1 C H
Fig. 14. A monocyte nucleus (N) is kidney shaped and displaced from the center. There are few mitochondria (MI and small amounts of RER. An aggregation of objects resembling mitochondria occupies the center of the cell and is reminiscent of a nebenkern. Bar: 1 pm. x 13,000.
Fig. 16. This monocyte has formed a large central lipid (L) droplet to the left of its nucleus (N). The cell at the left of the figure appears to be competing with it in the phagocytosis of a pool of extracellular material. Note the pseudopod (P). The cell at the top is a neutrophil. Bar: 1 Fm. ~4,000.
Fig. 15. A monocyte with a phagocytized platelet (Pl). Both the platelet plasma membrane and the phagosome membrane are discernible. Note the monocyte nucleus (N). Bar: 1 pm. ~6,000.

106 C. ROGERS WILLIAMS ET AL
_-I_
'I
.. . . . .
Figures 17-20

ULTRASTRUCTURE OF BELUGA WHALE BLOOD CELLS 107
(Table 1) are believed to reflect methodology. His measurements were carried out on a smear, where the cells have flattened out and attached to the substratum. Our cells were suspended in a liquid fixative and probably more nearly represent their actual in vivo size. As Table 1 shows, similar differences occur between the measurements of human blood cells derived from suspension and smears. The RBC diameters given by Quay and found by us are in close agreement with that of Cornell et al. ('88), i.e., 8.8 ? 0.4 Fm, also for belugas. Our WBC differential per- centages also agree fairly closely with those of Cornell et al. ('88) and are given here for ready comparison (ours first): neutrophils- 62/56; eosinophils-8.318; lymphocytes- 23.6130; monocytes-2/6. Cornell et al. ('88) also present useful data on serum chemistry. To date, no correlation between body size and erythrocyte diameter has been estab- Iished.
Myelocytes occurred in the peripheral cir- culation of Delphinus and Eschrichtius, on at least one occasion (De Monte and Pilleri, '79). Our observations confirm this report. The parallel stacks of RER are characteristic of neutrophilic metamyelocytes (Bloom and Fawcett, '86). Whether the presence of these cells represents a particular pathologic state in these cases is unknown. Their premature release in mammals is, of course, known to be related to the presence of an inflammatory process. Certainly, many early studies used
Fig. 17. A platelet which clearly shows the microtu- bules (Mt), here cut in cross section. Note the biconvex shape. Bar: 1 km. X28,500.
Fig. 18. Another platelet showing the microtubules (Mt) in longitudinal section. They form a circular ring at the periphery of the platelet. Bar: 1 pm. x 16,000.
Fig. 19. This normohlast presumably was about to extrude its nucleus. No cytoplasmic organelles are present and only a few polyribosomes persist. The lumen of the perinuclear cisterna is dilated but nuclear pores remain intact. The plasma membrane has buckled at the left side of the cell. Bar: 1 pni. ~9,000.
Fig. 20. A polychromatophilic erythroblast fixed while performing one of its last synthetic functions, i.e., trans- porting iron across the membrane. The pinocytotic vesi- cles are coated, presumably with transferrin, and can be seen pinching off from the plasma membrane in various stages (11-141. Polyribosomes (Po) are scattered in the cytoplasm and the mitochondria (M) are undergoing deg- radation. The nuclear pores (PI are prominent and the lumen of the perinuclear cisterna is slightly distended. Bar: 1 pm. x 17,000.
the blood of stressed or even dead wild ani- mals, which can account for a variety of changes in the circulating blood. According to Campbell and Murru ('go), some fish have hematopoietic function in their peripheral blood. The extent of myeloid tissue that is present in the circulation of healthy beluga whales as a species is unclear. That two dif- ferent classes of the subphylum vertebrata would show this phenomenon is interesting, but its possible significance is unexplored.
The presence of stabs, the band form of polymorphonuclear neutrophils, is typical of mammals and is probably indicative of nor- mal maturation. An excellent synopsis of clin- ical analysis of cetacean neutrophilic re- sponse was presented by Hyne et al. ('82).
Belugas have a higher percentage of eosino- phils than do humans, but not above the normally accepted range for many domesti- cated mammals (Schalm et al., '75). Eosino- philia is most pronounced among the smaller Cetacea (Medway and Geraci, '64, '65; En- gelhardt, '79; Nielsen and Andersen, '82). The larger captive odontocites, such as the killer whale (Cornell, '83) and beluga, have blood cell counts similar to those of terres- trial mammals. Eosinophilia has been attrib- uted to parasitic infections (Campbell and Murru, '90) known to affect most wild spe- cies, while a decrease in eosinophils can be related to stress (Schalm et al., '75). This latter point is somewhat surprising as one might expect parasitic infection to generate stress. Elevated eosinophils were reported in the lymph nodes of dolphins which had no identified parasites a t the time of death (Britt and Howard, '83). The whales in Baltimore did not carry any known parasitic load. Whether an elevated eosinophil count is a species specific adaptation of certain Cetacea is not know. The eosinophil nuclei reported here did not show increased segmentation as is sometimes the case in humans suffering from eosinophilia (Zucker-Franklin et al., '81). An interesting observation in the eosino- phils is the occurrence of intragranule inclu- sions that closely resemble eosinophil mito- chondria in size and approach them in structure, density and texture. Whether these inclusions represent mitochondria phagocy- tized by the granules, mitochondria about which the specific granule material has accu- mulated, or some other possibility remains to be determined.
Basophils from beluga whales are rare and routinely absent from clinical complete blood

108 C. ROGERS WILLIAMS ET AL
-- Fig. 21. In this reticulocyte, a few organelles are still
present and the cytoplasm has increased in density rela- tive to the cell in Figure 19. Bar: 1 pm. X 6,500.
Fig. 23. A red blood corpuscle illustrating its charac- teristic biconcave shape and electron density. Beluga whale red blood corpuscles are slightly larger than those of humans. Bar: 1 pm. ~9,000.
Fig. 22. An electron lucent red blood corpuscle of a captive beluga whale showing very dense inclusions (D). This one may have just extruded its nucleus. Bar: 1 pm, ~6,300.
Fig. 24. A biconcave electron lucent red blood corpus- cle demonstrating the considerable difference in density between itself and the typical corpuscle in the adjacent figure. Bar: 1 pm. x 13,000.
counts. Their cytoplasmic granules described by Quay ('54) possessed different shades of purple in his smears. Our observations con- firm granule heterogeneity. Until the con- tents of these presumably specific granules are ascertained, and their function deter- mined, it is recommended that clinicians merely try to determine precisely their nor-
mal percentage in cetacean blood, and their percentage during known clinical episodes. Basophils have been identified in only 7 of the 21 species of Cetacea examined (Camp- bell and Murru, '90) in one summary. The small granules with a dense center (Figs. 9, 11, G4) are hitherto unreported, and their significance is unknown.

ULTRASTRUCTURE OF BELUGA WHALE BLOOD CELLS 109
Stimulation of beluga lymphocytes, by pokeweed mitogen (PWM) and phytohemag- glutinin mitogen (PHA), led Mumford et al. ('75) to imply that there may be a greater number of B cells than T cells among ceta- cean lymphocytes. The fully mature plasma cell confirms the presence of B cells, but our study cannot morphologically distinguish be- tween B and T cells and is thus unable to offer further information in this area. The significance of the granules found in associa- tion with the plasma cells is unknown. Ac- cording to De Monte and Pilleri ('79), Knoll identified plasma cells from Znia geoffrensis and Physter catodon, where they are consid- ered atypical in peripheral circulation.
Morphologically, it is sometimes difficult to distinguish large lymphocytes from mono- cytes. The differential values for monocytes (11.4%) and lymphocytes (39.3%), provided by Quay ('541, fall in the accepted range for many domestic mammals (Schalm et al., '75). These data should probably be evaluated in light of the quality of cell preservation and the fact that the cells were taken from dead animals. Mean values of 19-34% for lympho- cytes and 2% for monocytes were recorded for the beluga by Englehardt ('79). The most interesting feature of the monocytes re- ported here was the nebenkern-like inclusion (Fig. 14). The components of the aggregate resemble the adjacent mitochondria in tex- ture and density. Also interesting is the phagocytized platelet (Fig. 15). Platelet phagocytosis by tissue macrophages is not uncommon, especially in the spleen, accord- ing to Sacher (personal communication), who also noted that such phagocytosis may occur in disorders where platelets are coated with immunoglobulins.
According to De Monte and Pilleri ('791, normoblasts are not normally found in the bloodstream of Cetacea. However, 1.6% retic- ulocytes were reported in Tursiops truncatus by Harkness and Grayson ('69). In our belu- gas, both normoblasts and reticulocytes were present in peripheral blood. The presence of polychromatophilic erythroblasts in circula- tion may be indicative of a pathologic condi- tion. However, the mechanics of beluga whale erythropoiesis have yet to be completely worked out. The final stage of erythrocyte maturation has been speculated to occur in the peripheral blood in fish (Campbell and Murru, '90). If the speculation is correct, one
might wonder how fish and aquatic mam- mals would benefit from such a shift.
The stages in presumptive pinocytosis in coated vesicles of electron lucent material by a polychromatophilic erythroblast (Fig. 20) are especially interesting. In the absence of cytochemical data, we can only suggest, fol- lowing the lead of Zucker-Franklin et al. ('81), that the stages represent the absorption of iron, in the form of transferrin, for hemoglo- bin production. If this is correct, the iron is presumably accumulated at the surface of the vesicles for, as noted, their content is electron lucent.
The hypochromic and microcytic RBCs present hematologists interested in cetacean blood with another perplexing condition. The subpopulation of such corpuscles is small and exists only at the interface of the buffy coat and packed cell volume after centrifugation. An accurate count could not be made. They were demonstrated in only two of the whales, but it is conceivable that they were missed in the third whale due to their isolated location and small population size. There is no basis for regarding them artifactually, but the pos- sibility exists. The common intracellular granules are consistent with the description of Howell-Jolly bodies, previously reported in captive Cetaceans (Quay, '54; Harkness and Grayson, '69; Cornell, '83). The inclusions also match the description of siderocytes (Da- cie and Mollin, '66; Deiss et al., '66).
We have found these studies of cetacean blood cell morphology enlightening and poten- tially clinically valuable. As aquariums con- tinue to increase in popularity, we hope that this study will spark further research to ben- efit both the captive and wild Cetaceans.
ACKNOWLEDGMENTS
We are indebted to Dr. Ronald Sacher, Georgetown University Hospital, 3800 Reser- voir Road, N.W., Washington, DC 20007, for his insights into blood physiology, and to the staff of the National Aquarium in Baltimore, Maryland, for their exemplary cooperation in this study.
LITERATURE CITED Bloom, W., and D.W. Fawcett (1986) A Textbook of
Histology, 1 l t h ed. Philadelphia: W.B. Saunders Com- pany.
Britt, J.O., Jr . , and E.B. Howard (1983) The hemopoietic system. In E.B. Howard (ed): Pathobiology of Marine Mammal Diseases, Vol. 11. Boca Raton: CRC Press, pp. 65-74.
Campbell, T., and F. Murru (1990) An introduction to fish hematology. Compend. Contin. Educ. Pract. Vet. 12:525-533.

110 C. ROGERS WILLIAMS ET AL
Cornell, L.H. (1983) Hematology and clinical chemistry values in the killer whale, Orcinus orca. J. Wild. Dis. 19:259-264.
Cornell, L.H., D.S. Duffield, B.E. Joseph, and B. Stark (1988) Hematology and serum chemistry values in the beluga (Delphinapterus leucas). J. Wild. Dis. 24.220- 224.
Dacie, J.V., and D.L. Mollin (1966) Siderocytes, sidero- blasts and sideroblastic anaemia. Acta Med. Scand. ISuppl.l445:237-255.
Deiss, A,, D. Kurth, G.E. Cartwright, and M.M. Wintrobe (1966) Experimental production of siderocytes. J. Clin. Invest. 45353-364.
De Monte, T., and G. Pilleri (1979) Cetacean hematology. 11. The blood cells. Invest. Cetac. 10.289-306.
Dhindsa, D.S., J. Metcalfe, A S . Hoversland, and R.A. Hartman (1974) Comparative studies of the respira- tory functions of mammalian blood. Killer whale (Orci- nus orca Linnaeus) and beluga whale (Delphinapterus leucas). Respir. Physiol. 20t93-103.
Engelhardt, F.R. (1979) Haematology and plasma chem- istry of captive pinnipeds and cetaceans. Aquat. Mamm. 7:11-20.
Gibbons, I.R., and A.V. Grimstone (1960) On flagellar structure in ccrtain flagellates. J. Biophys. Biochem. Cytol. 7:697-719.
Harkncss, D.R., and V. Grayson (1969) Erythrocyte me- tabolism in the bottle-nosed dolphin Tursiops trunca- tus. Camp. Biochem. Physiol. 28:1289-1301.
Hyne, R.H.J., E.E. Lepherd, and J.L. Van Everhrock ( 1982) Clinical significance of hematologic examina- tions of the dolphin (Tursiops sp.) in captivity. J. 2001. Ann. Med. 13:95-100.
Karnovsky, M.J. (1965) A formaldehyde-glutaraldehyde fixative of high osmolality for use in electron micros- copy. J. Cell Biol. 27:137A, 138A.
Luft, J.H. (1961) Improvements in epoxy resin embed- ding methods. J. Biophys. Biochem. Cytol. 9,409-414.
Medway, W., and J.R. Geraci (1964) Hematology of the bottlenose dolphin (Tursiops truncatus). Am. J. Phys- iol. 207:1367-1370.
Medway, W., and J.R. Geraci (1965) Blood chemistry of the bottlenose dolphin (Tursiops truncatus). Am. J. Physiol. 209: 169-172.
Mumford, D.M., G.D. Stockman, P.B. Barsales, T. Whit- man, and J.R. Wilbur (1975) Lymphocyte transforma- tion studies of sea mammal blood. Experientia 31:498- 500.
Nielsen, E., and S.H. Andersen (1982) Clinical chemistry and hematologic findings in the harbour porpoise (Pho- coena phocoena) from Danish waters. Aquat. Mamm. 9: 1-3.
Palade, G. (1952) A study of fixation for electron micros- copy. J. Exp. Med. 95285-298.
Quay, W.B. (1954) The blood cells of Cetacea with partic- ular reference to the beluga (Delphinapterus leucas Pallas, 1776). Saugetierkundliche Mitteil. 2:49-54.
Richardson, K.C., L. Jarett, and E.H. Fink (1960) Embed- ding in epoxy resins for ultrathin sectioning in electron microscopy. Stain Technol. 35313-323.
Schalm, O.W., N.C. Jain, and E.J. Carroll (1975) Veteri- nary Hematology. Philadelphia: Lea & Febiger.
Taylor, R.F., and R.K. Farrell (1973) Light and electron microscopy of peripheral blood neutrophils in a killer whale affected with Chediak-Hieashi svndrome. Fed. " Proc. 32t3043.
Venable, J.M., and R. Coggeshall(1965) A simplified lcad citrate stain for use in &ctron microscopy. J. Cell Biol. 25:407408.
Wilson, J.M. (1924) The Cell in Development and Hered- itv. 3rd ed.. with corrections. 5th minting. 1953. New
, I Y _
Y b r k Macmillan Company. Zucker-Franklin, D., M.F. Greaves, C.E. Grossi, and A.M.
Marmont (1981) Atlas of Blood Cells. Function and Pathology. Milano: E. Ermes.