tumour immunology - oncologypro.esmo.org · curotto de lafaille ma, lafaille jj. immunity. 2009...
TRANSCRIPT

Tumour Immunology
ESMO Preceptorship, Lund, December 2015
Nature reviews cancer April 2012
David GilhamInsttute of Cancer Sciences
The University of Manchester

Schreiber, Lloyd, Smyth (2011). Science 331(6024): 1565
Proposed theory of Cancer immunoeditng
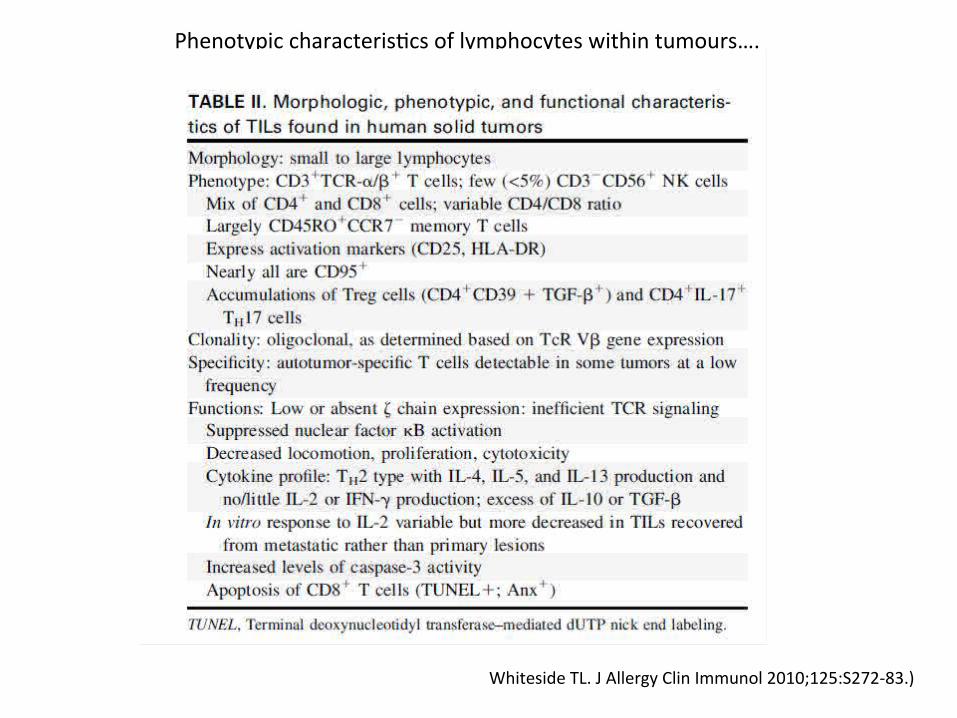
Whiteside TL. J Allergy Clin Immunol 2010;125:S272-83.)
Phenotypic characteristcs of lymphocytes within tumours….

Whiteside TL. J Allergy Clin Immunol 2010;125:S272-83.)
…. And in the periphery

Fridman et al. Nature Reviews Cancer April 2012

Fridman et al. Nature Reviews Cancer April 2012

Immune suppression
T cell recogniton of tumourCell suppression mechanismsProtein / other suppressive factors

Whiteside TL. J Allergy Clin Immunol 2010;125:S272-83.)
Human Tumour Asociated Antgens
Majority are ‘SELF’ antgens
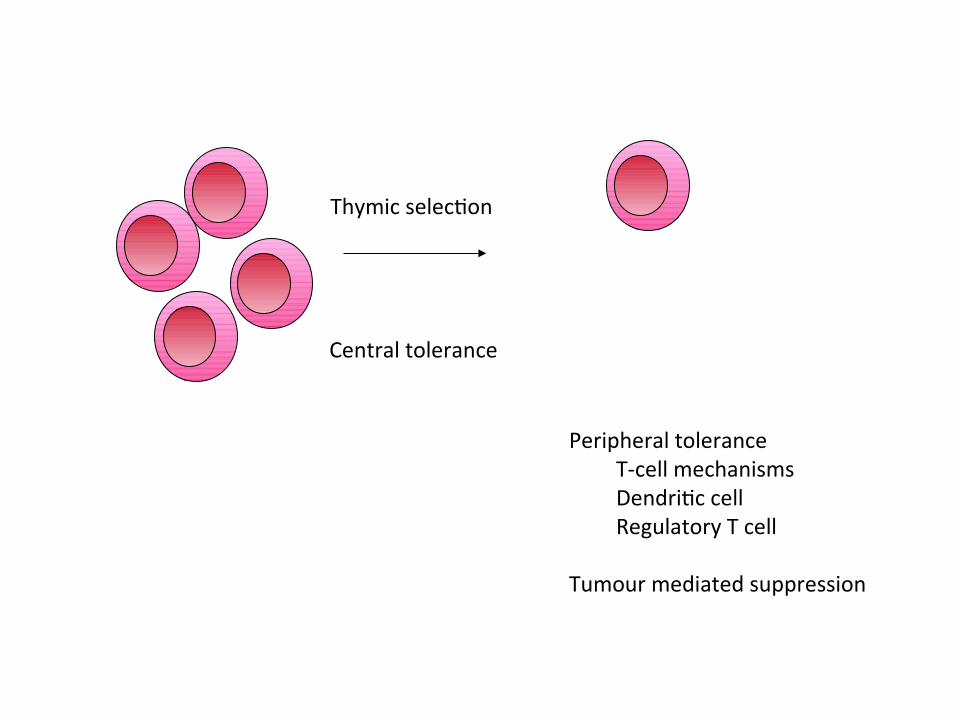
Thymic selecton
Central tolerance
Peripheral toleranceT-cell mechanismsDendritc cellRegulatory T cell
Tumour mediated suppression

htp://info.cancerresearchuk.org/cancerstats/mortality/age/

Immune senescence / skewing of the immune response due to chronic viral infecton

Unfortunately, we all get older……
Gravekamp C.Curr Opin Immunol. 2011 Aug;23(4):555-60.Epub 2011 Jul 18.

T-cell fate under different conditions of TCRengagement.
Alegre et al. Nat Rev Immunol. 2001;1(3):220-8.
Signal 1
Signal 2

Keratin
Bw6
A-locus
A2
Keratin
Bw6
A-locus
A2
Kindly provided by Professor Peter Stern
Tumour evasion on T cell actvity: down-regulaton of HLA

The three categories of costimulatory and coinhibitorysignals at the APC-T cell interface
Ligand-receptor pairs for the B7 family, TNF family, and cytokine family of signals between the APCand T cell. Not all receptors for some B7 family members have been identfed. Some B7 familymembers interact with both costmulatory and coinhibitory receptors
Pardoll. Prendergast & Jafee (Ed):Cancer Immunotherapy, Immune Suppression and Tumour Growth. 2007.

Dong H, Chen L. B7-H1 pathway and its role in the evasion of tumor immunity. J Mol Med (Berl). 2003May;81(5):281-7. Epub 2003 Apr 30.
B7-H1 / PD-L1 expression on tumours

Surface expression of CD80 on transplantable tumor cell lines.
Tirapu I et al. Cancer Res 2006;66:2442-2450
©2006 by American Association for Cancer Research

Increased immunogenicity of cells expressing high levels of CD80.
Tirapu I et al. Cancer Res 2006;66:2442-2450
©2006 by American Association for Cancer Research

T-cell-intrinsic mechanisms of peripheral tolerance.
Walker & Abbas. Nat Rev Immunol. 2002 Jan;2(1):11-9.

Role of DCs in the choice between immunity and tolerance.
Walker & Abbas. Nat Rev Immunol. 2002 Jan;2(1):11-9.

Differentiation of T helper cell subsets.
Zou and Restfo. Nat Rev Immunol. 2010 Apr;10(4):248-56.

Curotto de Lafaille MA, Lafaille JJ. Immunity. 2009 May;30(5):626-35.
Thymic and Peripheral Generaton of Foxp3+ Tregs

Zou. Nature Reviews Immunology 6, 295–307 (April 2006) |
Sources for Treg in the tumour??

Vignali et al. Nat Rev Immunol. 2008 Jul;8(7):523-32.
Immunosuppressive mechanisms used by Tregs.
Treg-cell mechanisms centred around four basic modes of acton. a) Inhibitory cytokines include IL-10, IL-35 and TGF-b. b) Cytolysis includes granzyme-A- and B-dependent and perforin-dependent killing mechanisms. c) Metabolicdisrupton includes high-afnity CD25 (also known as IL-2 receptor)-dependent cytokine-deprivaton-mediatedapoptosis, cyclic AMP (cAMP)-mediated inhibiton, and CD39- and/or CD73-generated, adenosine receptor 2A(A2AR)-mediated immunosuppression. d) Targetng DCs includes mechanisms that modulate DC maturaton and/orfuncton such as lymphocyte-actvaton gene 3 (LAG3; also known as CD223)–MHC-class-II-mediated suppression ofDC maturaton, and CTLA4–CD80/CD86-mediated inducton of indoleamine 2,3-dioxygenase (IDO), which is animmunosuppressive molecule made by DCs.

Curiel et al. Nat Med. 2004 Sep;10(9):942-9.
Tumour Tregs suppress T cell actvaton in vitro.
• CD4+CD25+ Tregs fromtumour ascites, the tumourmass or blood were added tothe culture of T respondercells.
• Tumour ascites and tumourmass Tregs are as efcient asblood Tregs at inhibitng Tcell proliferaton (a) .
• Tumour ascites Tregs inhibitT cell producton of IFN-g (b)and IL-2 (c), but have noefect on IL-4 (d) or IL-10 (e)producton.

p=0.0059*
PBMC TIL
PBMC
TIL
CD4+FoxP3+ Tregs are higher in TILs thanPB of RCC
Grifths et al. Cancer Immunol Immunother. 2007;56(11):1743-53.
FOXP3
0
5
10
15
20
25
30
35
40F
OX
P3+
/CD
4+
%

Prognostc signifcance of Tregs in cancer.
Elkord et al. Expert Opin Biol Ther. 2010 Nov;10(11):1573-86.

• Tumour immunoeditng and progression include three phases, defned as: eliminaton,equilibrium and escape.
• Early in carcinogenesis T cell driven M1 actvated macrophages may contribute toeliminaton.
• During tumour progression a gradual switching of polarizaton, M1 versus M2, is paralleledby the gradual inhibiton of NF-kB actvity.
• These events concur to establish conditons for tumour growth and spread (escape phase).
Macrophage polarizaton (M1 to M2).
Mantovani &Sica. Curr Opin Immunol. 2010 Apr;22(2):231-7.

• Recent studies have identfed myeloid-origin cells that are potent
suppressors of tumour immunity.
• Together with Tregs, MDSCs promote an immunosuppressive environment
in tumour-bearing hosts.
• MDSCs accumulate in the blood, lymph nodes, and bone marrow and at
tumour sites in most patents and experimental animals with cancer
• MDSCs are induced by tumour-secreted and host-secreted factors, many
of which are proinfammatory molecules.
• The inducton of MDSC by proinfammatory mediators led to the
hypothesis that infammaton promotes the accumulaton of MDSCs that
down-regulate immune surveillance and anttumor immunity, thereby
facilitatng tumour growth.
Myeloid-derived suppressor cells (MDSCs)
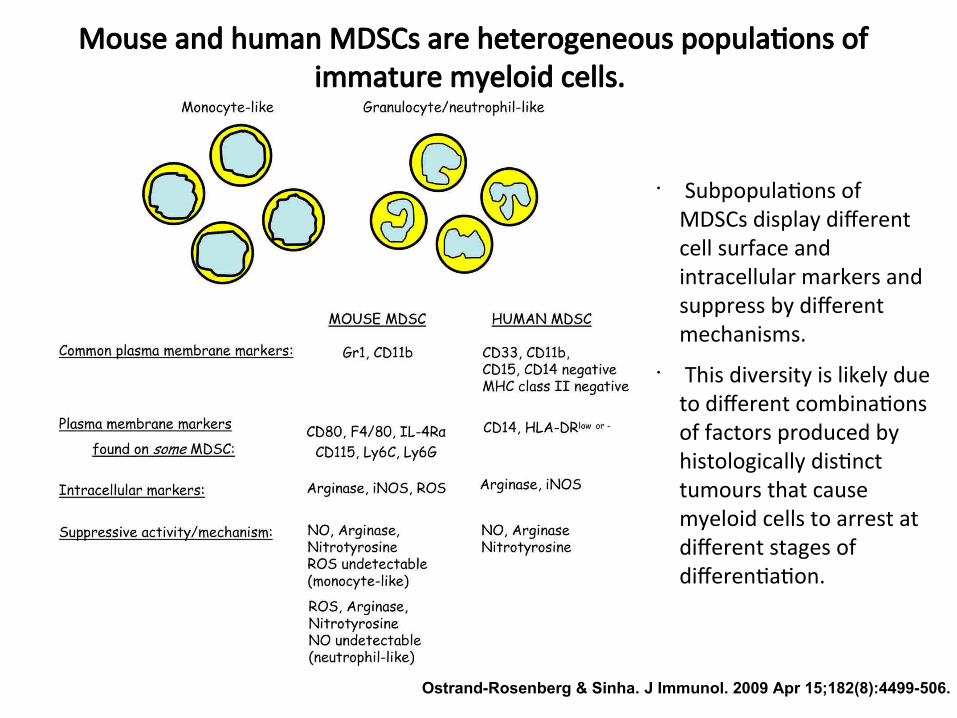
Mouse and human MDSCs are heterogeneous populatons ofimmature myeloid cells.
• Subpopulatons ofMDSCs display diferentcell surface andintracellular markers andsuppress by diferentmechanisms.
• This diversity is likely dueto diferent combinatonsof factors produced byhistologically distncttumours that causemyeloid cells to arrest atdiferent stages ofdiferentaton.
Ostrand-Rosenberg & Sinha. J Immunol. 2009 Apr 15;182(8):4499-506.

PrecursorPartally actvated / stalled / regulatory
Fully actvated / diferentated efector
Block?

MDSCs suppress anttumor immunity through a variety of diversemechanisms.
• T cell actvaton is suppressedby producton of arginase andROS, the nitraton of the TCR,cysteine deprivaton, and theinducton of Tregs.
• Innate immunity is impaired bythe down-regulaton ofmacrophage-produced IL-12,the increase in MDSCproducton of IL-10, and thesuppression of NK cellcytotoxicity.
• Ag presentaton is limited bythe expansion of MDSC at theexpense of DC.
MDSCs suppress adaptve and
innate ant-tumour immunity
Ostrand-Rosenberg & Sinha. J Immunol. 2009 Apr 15;182(8):4499-506.

Whiteside TL. J Allergy Clin Immunol 2010;125:S272-83.)
‘Tumour’ derived factors infuencing the immune system


Whiteside TL (2008) Oncogene 27: 5904-5912

Parmianiz et al. (2007). Journal of Immunology 178:1975-1979
SELF mutated antgens

Wooldridge L et al. J. Biol. Chem. 2012;287:1168-1177

Recognition of 30 peptides drawn from a CPL-based importance sampling set with effectivesize = 1. 66 × 108 (calculated from the sampling entropy) (second set).
Wooldridge L et al. J. Biol. Chem. 2012;287:1168-1177
©2012 by American Society for Biochemistry and Molecular Biology

Whiteside TL (2008) Oncogene 27: 5904-5912

Disis ML. J Clin Oncol. 2010 Oct 10;28(29):4531-8.
T-helper (Th)2 and Th1 tumour-specifc T cell immunity.