transport and metabolism of folates by bacteria* · transport and metabolism of folates by...
TRANSCRIPT

THE Jourmar, OF BIOLOGICAL CHEMISTRY Vol. 250, No. 6, Issue of March 25, pp. 2243-2253, 1975
Printed in U.S.A.
Transport and Metabolism of Folates by Bacteria*
(Received for publication, July 26, 1974)
BARRY SHANE AND E. L. R. STOKSTAD
From the Department of Nutritional Sciences, University of California, Berkeley, California 94720
SUMMARY
Transport of labeled folic acid (PteGlu), pteroylpolygluta- mates (PteGlu,.,), 5-methyl-tetrahydrofolate (5.methyl- HPteGlu), and methotrexate in late-log phase cells of Lacfobacillus casei was active, and subject to inhibition by unlabeled pteroyhnonoglutamates, pteroylpolyglutamates, and iodoacetate, but not glutamate or glutamate dipeptides. Pteroylpolyglutamates were transported without prior hy- drolysis and shared a common uptake system with pteroyl- monoglutamates. The afIinity and maximum velocity of PteGlu, uptake decreased with increasing glutamate chain length (K,: PteGlul, 0.03 PM; PteGlus, 0.32 PM; PteGlur, 1.9 pM; PteGlus, 3.7 PM) and comparisons with growth response curves suggested that polyglutamates were more effectively utilized by Z. casei, once transported, than monoglutamate. No concentration of 5-methyl-HPteGlus.s inside the cells was observed.
The major folate metabolites found in L. casei preloaded with high levels of [3H]PteGlu (0.5 pM) were IO-formyl- HsteGluz and IO-formyl-PteGlu. Both compounds were released, the monoglutamate more rapidly. Pteroyltrigluta- mate formation appeared to be a rate-limiting step in intra- cellular metabolism. No IO-formyl-PteGlu was found in iodoacetate-treated cells and efflux was inhibited. Cells pre- loaded with low levels of [3H]PteGlu (7 m) metabolized the vitamin to polyglutamate forms, the major derivatives being HPteGlu,.
First order exit rates of labeled folate from preloaded L. cosei indicated an inhibition of PteGlu uptake with time. Exit rates dropped from 0.05 mm1 to <0.002 min+ as intra- cellular folate was metabolized from monoglutamate to poly- glutamate derivatives (n 2 3). In the latter case, materials lost by efflux were breakdown products and no folate of glutamate chain length greater than two was released.
Pediococcus cerevisiae actively transported 5.methyl- HPteGlu but did not take up 5-methyl-H4PteGlu3.8. No active accumulation of 5-methyl-HcPteGlu was observed in Streptococcus fuecalis.
Since the discovery of pteroylheptaglutamates in yeast (2), many authors have reported the existence of pteroylpolygluta-
* This research was supported by United States Public Health Service Grant AM-08171 from the National Institutes of Health. A preliminary report of this study has been published (1).
mates in plants (3), algae (4), bacteria (5, 6), and animal tissues (7-9). The recent introduction of specific chromatographic methods for determining polyglutamate chain length (10, 11) coupled with methods for the preparation of synthetic polyglu- tamates for use as reference standards (12, 13) has made possible the exact determination of polyglutamate forms in bacteria and animal tissues (11, 13, 14). For instance, the main folate forms in rat tissues, with the exception of plasma, have been shown to be pentaglutamates (11, 14), while those in Lactobaci/lus casei and Streptococcus juecalis were the octa- and tetraglutamates, respec- tively (13).
With the exception of L. casei, which is able to grow on oxidized pteroylpolyglutamates of chain length at least up to seven (15), all folate-requiring bacteria so far investigated require mono- or diglutamates for growth (16). Intestinal absorption of folate is also limited to mono- and diglutamates (17) and human marrow cells will not transport triglutamates (18). From the limited studies carried out, the reason for intracellular metabolism of pteroyl compounds to polyglutamate forms would seem to be that these compounds have higher affinities for folate-dependent enzymes than the corresponding monoglutamates (19-25). An- other possibilit,y, suggested by lsuehring et al. (13), is that poly- glutamates are better rctaincd by cells than nrc monoglutamate derivatives. They showed that folate in the growth medium of bacteria was apparently of shorter length than that found intra- cellularly.
To investigate some of these problems, uptake, efflux, and metabolism of various mow and polyglutamyl folate derivatives were studied in nonproliferating bacteria. Initially, uptake of 5-methyl-II~I’teGlu,i was studied in S. jueculis and Pediococcus ceretisiae as these organisms have been reported to actively trans- port 5.methyl-H&eGlu (26, 27) although they cannot metab-
olize or utilize this compound. As both organisms proved to be unsuitable for the study of nonmetabolizable polyglutamate up- take, further studies were carried out with L. cusei, which necessi- tated a study of the metabolism of transported vitamin.
EXPERIMENTAL PROCEDURE
MateriuZs-[[3H]PteGlu, labeled in the 9, 3’, and 5’ positions (specific activity, 0.5 or 37 Ci per mmol), (f )-5-[Wjmethyl-HaPte- Glu (specific activity, 54 mCi per mmol), and [3H]methotrexate, labeled in the 9, 3’, and 5’ positions (specific activity, 15 Ci per mmol), were obtained from Amersham-Searle. Labeled folates were purified by chromatography on QAE (quarternary amino- ethyl)-Sephadex A-25 prior to use (13).
1 The abbreviations used are: PteGlu, pteroylglutamic acid, folic acid; PteGlu,, pteroylmono- to octaglutamic acid; n indi- cates the number of glutamic acid residues; HrPteGlu,, 5,6-7,8- tetrahydropteroylmono- to polyglutamic acid.
2243
by guest on April 8, 2019
http://ww
w.jbc.org/
Dow
nloaded from

2244
5-Methyl-HaPteGlu was prepared by reduction of 5, IO-methe- nyl-HaPteGlu with sodium borohydride (28), HaPteGlu was pre- oared bv reduction of PteGlu in glacial acetic acid with sodium hithion;e (29), and 10.formyl-PteGlu was prepared by formyla- tion of PteGlu (30).
Folic acid polyglutamates PteGluz to PteGlu?, PteGlu-[U-14C]- Glu-Glu (specific activity, 310 &i per mmol), Pt,eGlu*-[UJ4C]- Glu-Glu (specific activity, 100&i per mmol), and PteGlus-[U-i4C]- Glu-Glu (specific activity, 410 &i per mmol) were synthesized by the method of Baugh et al. (12). (+)-5-Methyl-[3H]H4PteGlu-8 (specific activity, 37 Ci per mmol) were prepared from [“H]Pte- Glu by the biosynthetic method of Buehring et al. (13). All poly- glutamates were purified by chromatography on QAE-Sephadex and DEAE-cellulose. The identity of each‘compound was unam- biguously confirmed by its chromatographic behavior on Sephadex G-25 and by differential microbiologic response after hog kidney conjugase treatment (13, 15, 31).
r-L-Glutamyl-L-glutamic acid was obtained from Sigma Chemi- cal Co. and or-n-glutamyl-L-glutamic acid was obtained from Schwartz-Mann.
Organisms and Growth Conditions-Lactobacillus casei (ATCC 7469), Streptococcus jaecalis (ATCC 8043), and Pediococcus cere- visiae (ATCC 8081) were cultured bv the arocedure of Bird et al. (31) with slight modifications employed in this laboratory (15). Folates added to the media were PteGlu (1 ng per ml for L. casei, 2 ng per ml for S. fuecalis), or 5-formyl-H4PteGlu (1 ng per ml for P. cerevisiae). In some experiments, S. juecalis medium contained thymidine (2 pgm per ml) instead of PteGlu.
Measurement of Folate Uptake-Bacteria were harvested by centrifugation from growth media in late log phase (20 to 24 hours at 37”), washed twice with double-strength buffer, resuspended in buffer, and stored at O-4” during the course of an experiment until used. Unless indicated otherwise, the buffer used (single strength) was 50 mM KtIIPOd-100 mM sodium acetate-HaPOd, pH 6, contain- ing glucose (lyO)). Cell concentration (dry weight) was estimated by absorbance at 640 nm.
Cells (0.02 to 1.0 mg per ml) were preincubated in a shaking water bath at 37“ for 5 min before addition of labeled vitamin. Aliquots (0.5 or 1.0 ml) were removed at various intervals and filtered on HA filters (Millipore Corp., 25.mm diameter, 0.45~pm pore size). After washing with cold buffer (twice with 1 ml), cells plus filter were added directly to counting vials. Aquasol (New England Nuclear) or Triton X-lOO-toluene (1:2) scintillation mix- ture containing 2,5-diphenyloxazole (PPO) and 1,4-bis[2-(5- phenyloxazolyl)]benzene (POPOP) (10 ml) was added to each vial. Counting efficiencies were determined by external standardization or by a channels ratio method and were about 80% for r4C and 25% for tritium.
Eflux and Metabolism of Intracellular Folate-L. casei (0.2 mg per ml) was preloaded with [H]PteGlu for various times, filtered, washed with buffer, resuspended in fresh buffer (2 volumes for exit rate studies; 0.5 volume for metabolic studies), and incubated with shaking at 37”. In some experiments, resuspension buffer contained iodoacetate (20 mM) plus azide (50 mM) or unlabeled PteGlu (50 PM).
For exit rate studies, aliquots were removed at 2,4,6, and 8 min and filtered, and cell-free filt,rate (1 ml) was added to counting vials to determine released vitamin.
For metabolic studies, aliquots (5 ml) were removed at 5,20, and 60 min and filtered, and cells plus filter were resuspended in buffer (5ml) containingmercaptoethanol (0.2 M). Intracellular folate was extracted by boiling for 5 min and cell debris and filter were re- moved by centrifugation. Mercaptoethanol (0.2 M) was also added to the cell-free medium.
Identification of Labeled Folates-Folates were identified bv their ch”romatographic behavior on Sephadex G-25 and DEAE- cellulose before and after hog kidney conjugase treatment (13).
The chromatographic behavior of folates on Sephadex G-25 has been described in detail bv Shin et al. (10. 11). Columns (200 X 0.75 cm) were eluted with 6.1 in potassium phosphate buffer; pH 7, containing 0.2 M mercaptoethanol. In general, polyglutamates could be separated according to size. The aromatic and heterocy- clic moieties of the folate compounds caused a slower passage through the gel than would be expected by their molecular sizes (for instance, PteGlua was eluted later than glutamic acid). Ilow- ever, formyl derivatives were retarded less than PteGlu, HaPte- Glu, and 5-methyl-HaPteGlu. This was also true for polygluta-
mates of folic acid and for lo-formyl-HdPteGlu, which eluted ap- proximately at the position of I’teGlu,+r.
[3H]Folate samples were chromatographed on Sephadex G-25 columns that. had been previously standardized with authentic folate standards and individual peaks were rerun on Sephadex together with appropriate r4C-labeled or unlabeled folate stand- ards.
Further identification of the various folate derivatives was achieved by chromatography of [3H]folate samples, before and after hog kidney conjugase treatment, on DEAE-cellulose as described by Buehring et al. (13). Columns (25 X 0.9 cm) were eluted by an exponential phosphate gradient formed with 0.01 M
potassium phosphate, pH 6.0 (100 ml), in a closed mixing chamber attached to a reservoir containing 0.6 h* potassium phosphate, pH 6.0. All buffers contained 0.2 M mercaptoethanol. A standard solution containing unlabeled PteGlu and IO-formyl-PteGlu and 5-[14C]methyl-H,PteGlu was added to each [3H]folate sample before application to the column. The 5-[14C]methyl-H4PteGlu standard had been stored for several months at 4’ and, under these conditions, about 50% had been converted to a degradative prod- uct of unknown composition, indicated by X in Figs. 6, 7, and 9. The elution position of this compound, which had no growth-pro- moting activity for L. casei (13),was useful in the interpretation of data from DEAE-cellulose chromatoaraahv. as IO-formvl-HdPte- Glu was elutcd midway between X and iO~formyl-PteGl”u. -
Treatment of Kinetic Data-K, and V,,, values for folate trans- port were calculated by the statistical method of Wilkinson (32). Estimates of K,, and V,,, were obtained by a least squares weighted treatment of the Lineweaver-Burk double reciprocal plot (33) and “best-fitting” the provisional values to a hyperbola. A similar treatment was applied to determine KI values for folate analogs. All data were inspected visually to ensure that they fol- lowed Michaelis-Menten-type kinetics. V,,, values were checked in inhibition studies to ensure that they were unaffected by the presence of inhibitors and that competition was competitive. V,,, values are expressed as micromolar per min increase in intra- cellular vitamin concentration and assume an intracellular water volume of 4 ml per g (dry weight) of cells (34).
As loss of intracellular vitamin followed first order kinetics, rate constants were determined by a least squares log-linear regression method.
RESULTS
5-dlethyl-H4PteGlu Uptake by Streptococcus faecalis-Although intracellular levels of 5-[r4C]methyl-H4PteGlu slowly equilibrated with extracellular vitamin at 37”, the only concentration effects noted could bc explained by intracellular breakdown of labeled vitamin to the degradative product X. These effects were noted in a variety of glucose-containing buffers (20 mM potassium phosphate, pH 6; acetate-phosphate buffer, pH 6), and prcincu- bation of cells in buffer, addition of excess unlabeled PteGlu (50 PM) or azide (50 mM) plus iodoacetate (10 mM), and culturing cells in thymidine instead of PteGlu did not significantly change this uptake pattern. Uptake rates were approximately propor- tional to substrate concentration over the range tested (0.01 to 5.0 PM) and were reduced slightly at 0”. A slight pH effect on uptake with a maximum at pH 6.0 was noted, but as low levels of relatively low specific activity vitamin was being measured, it was difficult to distinguish whether this was a transport phe- nomenon or a nonspecific binding effect.
McElwee and Scott (26) suggested that S. juecalis concen- trated 5-methyl-H~l’teGlu as Lactobacillus casei was unable to grow on used medium containing low levels of PteGlu and 5- methyl-H4PteGlu in which S. fuecalis had previously been grown. As 5-methyl-H&eGlu will support growth of L. casei but not S. jaecalis, it appeared that S. juecalis had removed all of the 5-methyl-H41’teGlu from the medium. We were able to confirm these growth observations but, using 5-[14C]methyl-H41’teGlu, found practically all the radioactive label in the used medium. It appeared that the lack of growth by L. casei was due to the
by guest on April 8, 2019
http://ww
w.jbc.org/
Dow
nloaded from

removal of a non-folate growth factor by S. juecalis or excretion of a growth inhibitor, as has been suggested for Pediococcus ceretisiae (35), rather than lack of folate.
Uptake of Folates by P. cerewisiae-As S. jaecalis did not appear to actively transport 5-methyl-H&eGlu, uptake studies were carried out with P. ceretisiae, an organism that requires reduced folates, with the exception of 5-methyl-HJ’teGlu, for growth.
Table I shows typical effects of time on the uptake of various mono- and polyglutamyl folates by P. cerevisiae. As shown by Mandelbaum-Shavit and Grossowicz (27), the organism will ac- tively transport 5-methyl-H4PteGlu but will not concentrate PteGlu.
It was hoped that this organism could be used for transport studies on 5-methyl-H$teGlu polyglutamates, but only low levels of the Glus to Glus forms were taken up (Table I) and it is possible that these results reflect binding to the cells and filter rather than transport as such.
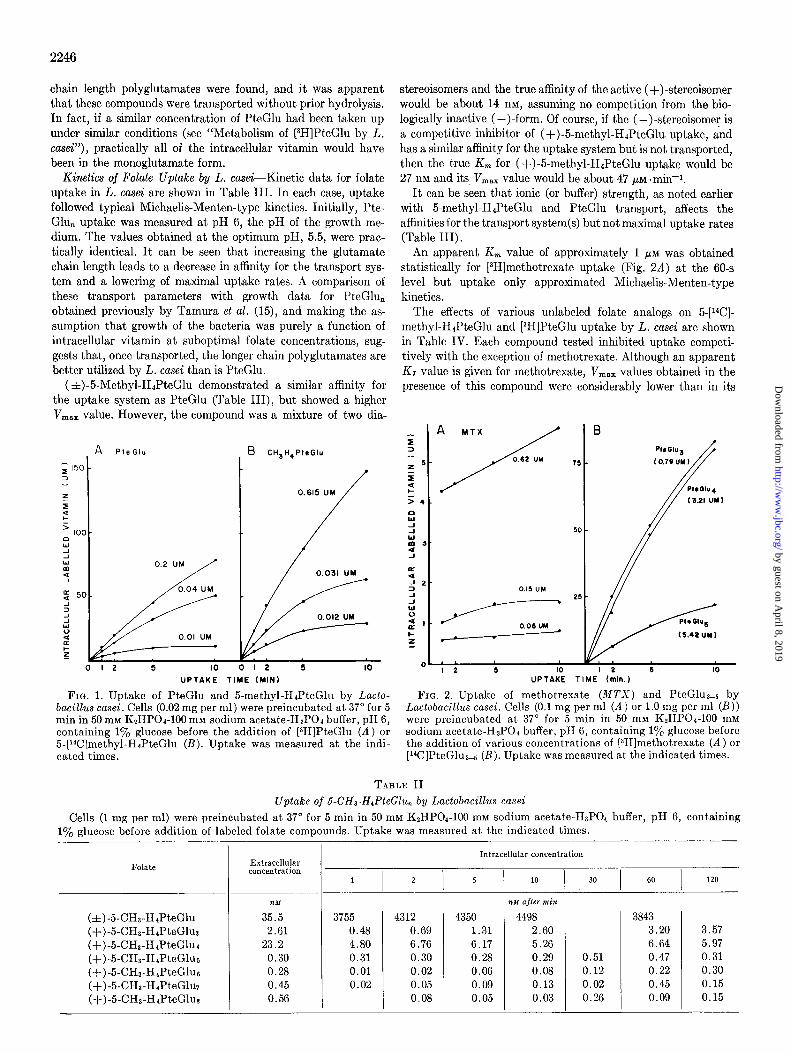
Uptake of Folate ~1Ionoglutamates by L. case%-Fig. 1 shows typical effects of time on the uptake of [3H]I’teGlu and 5-[i4C]- methyl-H&eGlu by L. casei. Intracellular concentrations of greater than 2000.fold over that in the medium were found when low folate concentrations were used, and in the first 5 min at least, most of this transported vitamin had not been metabolized to different forms. The 5.[i”C]met.hyl-HJ’teGlu was a mixture of two diastereoisomers, of which only the (+)-form is bio- logically active. When heavier cell suspensions (0.5 to 1.0 mg per ml) were used, practically half of the radioactivity in the medium was taken up (Table II). Under comparable conditions, L. casei was unable to deplete the medium of [3H]I’teGlu, and an over- shoot effect was often noted in intracellular tritium levels.
Uptake of 5-methyl-HJ’teGlu and PteGlu was proportional to cell concentration up to 0.05 mg per ml (dry weight). Above this concentration, a falling off from linearity was observed.
[3H]Methotrexate was also concentrated by L. casei (Fig. 2A). A rapid initial uptake or binding occurred within 60 s and was followed by a gradual accumulation of tritium label.
Iodoacetate (10 rn$ inhibited initial uptake rates by 40 to 80%. At higher levels (100 mM), intracellular levels of 5.[%I- methyl-H&eGlu equilibrated with extracellular levels and only a slight concentration of [3H]PteGlu was observed. Azide (50 mM), arsenite (20 mM), and arsenate (20 mM) had no apparent effect on monoglutamate uptake in L. casei. Upt.ake at 0” was about 10% of the 37” rates.
2245
5-[i4C]Methyl-H4PteGlu or [3H]PteGlu uptake by L. casei was similar in buffers containing sodium or potassium, phosphate, or acetate, and was reduced in citrate buffer. Uptake was stimu- lated by increased buffer concentrations, maximal rates were found with dipotassium phosphate ( 2 100 mM) or sodium acetate ( 2200 mM). This stimulation was most apparent when low sub- strate concentrations were used.
A broad pH optimum between pH 5 and 6.5 with a peak at pH 5.5 was observed when uptake was measured in the presence of excess 5-[14C]methyl-H41’teGlu or [3H]PteGlu (0.5 PM). Uptake dropped off more rapidly outside this pH range. With lower folate levels (0.03 PM), similar effects were noted, with a more pro- nounced drop below pH 5 and above pH 6.5, suggesting a reduced affinity of the uptake system(s) for folates in these pH ranges. During growth of L. casei, the medium pH drops from 6 to about 5.2. It appears that this change in pH has little effect on the ability of the organism to transport folates.
Uptake of 5-dlethyl-[3H]H.,PteGlu Polyglutamates by L. casei- The uptake of 5-methyl-[3H]H&eG1u3-a by L. casei is shown in Table II. Although a slow accumulation of tritium label was observed, intracellular levels never reached greater than 50% more than extracellular levels. At these low levels of uptake, metabolism or binding could explain this small concentration effect. Iodoacetate (20 mM) had no apparent effect on uptake rates so there was no evidence for an active accumulation of these compounds. However, unlike the results obtained with P. cere- visiae (Table I), it was apparent that 5-methyl-H&eGlu poly- glutamates were being taken up by the bacteria. Essentially identical results were obtained when these uptake experiments were repeated utilizing 5-methyl-H&eGlu polyglutamates syn- thesized on different occasions.
Uptake of [14C]PteGlu Polyglutamates by L. case&The uptake of [%]PteGlu3-5 by L. casei is shown in Fig. 2B. Relatively large amounts of the polyglutamates had to be used as well as high cell concentrations (1 mg per ml) in order to accurately measure the uptake of the extremely low specific activity material. Even un- der these conditions, significant accumulation and intracellular concentration of vitamin occurred. In contrast to PteGlu upt,ake, within 60 min most of the PteGlus-5 had been taken up by the cells. Uptake of these compounds was also subject to inhibition by iodoacetate and was reduced at 0”.
Extracts of cells loaded with [%]PteGlu3 or [14C]PteGlus were chromatographed on Sephadex G-25 columns (Fig. 3). No lower
TABLE I
Uptake of 5-CHs-H4PteGlu,, and PteGlu by Pediococcus cerevisiae Cells (1 mg per ml) were preincubated at 37” for 5 min in 50 mM K,HPOb-100 mM sodium acetate-H8P01 buffer, pH 6, containing
1% glucose before addition of labeled folate compounds. Uptake was measured at the indicated times.
Folate
(f)-5-CHZ-H4PteGlu (+)-5-CH3-H4PteGlu~ (+)-5-CHa-H,PteGlur (+)-5-CX-HIPteGlus (+)-5-CX-HIPteGlus (+)-5-CHS-H4PteGlu7 (+)-5-CH3-H4PteGlus PteGlu
Extracellular concentration
mu
36 2.81
23.0 0.31 0.30 0.49 0.61
14.0 1340
1 I 2
72 0.02 2.78 -cl - -
155 337 0.43 0.2 6.19 3.6, -0 -a
2 2 14 111 32 53
Intracellular concentration
5 I
10 30 60 120
1 1
635 0.71 3.12 -a
- 5
39
-a
-
-
-
2007 0.10 3.51
-a
-
15 186
0.24 1.28
14 254
a Dashes indicate no uptake detected.
by guest on April 8, 2019
http://ww
w.jbc.org/
Dow
nloaded from
2246
chain length polyglutamates were found, and it was apparent that these compounds were transported without prior hydrolysis. In fact, if a similar concentration of PteGlu had been taken up under similar conditions (see “Metabolism of 13H]PteGlu by L.
co.&“), practically all of the intracellular vitamin would have been in the monoglutamate form.
Kinetics of Folate Uptake by L. casei-Kinetic data for folate uptake in L. casei are shown in Table III. In each case, uptake followed typical Michaelis-Menten-type kinetics. Initially, Pte- Glu, uptake was measured at pH 6, the pH of the growth me- dium. The values obtained at the optimum pH, 5.5, were prac- tically identical. It can be seen that increasing the glutamate chain length leads to a decrease in aflinity for the transport sys- tem and a lowering of maximal uptake rates. A comparison of these transport parameters with growth data for PteGlu, obtained previously by Tamura et al. (15), and making the as- sumption that growth of the bacteria was purely a function of intracellular vitamin at suboptimal folate concentrations, sug- gests that, once transported, the longer chain polyglutamates are better utilized by L. casei than is PteGlu.
(zt)-5-Methyl-HJPteGlu demonstrated a similar affinity for the uptake system as PteGlu (Table III), but showed a higher V,,, value. However, the compound was a mixture of two dia-
I
I-
O-
0.
d PieGlu B CH3H,PleGIu
0.2 UM
0.04 UM
j_/ ,..----:nl:
UPTAKE TIME (MINI 5 IO I 2 6 IO
UPTAKE TIME (min.)
FIG. 1. Uptake of PteGlu and 5-methyl-HdPteGlu by Lacto- FIG. 2. Uptake of methotrexate (MTX) and PteGluae5 by bacillus casei. Cells (0.02 mg per ml) were preincubated at 37” for 5 Lactobacihs cusei. Cells (0.1 mg per ml (A) or 1.0 mg per ml (B)) min in 50 rnM KzHPOd-100 mM sodium acetate-HaPOh buffer, pH 6, were preincubated at 37” for 5 min in 50 mM K~HPOI-100 mM containing 1% glucose before the addition of [$H]PteGlu (A) or sodium acetate-HsPOa buffer, pH 6, containing 1% glucose before 5-[r4C]methyl-HaPteGlu (B). Uptake was measured at the indi- the addition of various concentrations of (3H]methotrexate (A) or cated times. [14C]PteGlu3-5 (B). Uptake was measured at the indicated times.
stereoisomers and the true affinity of the active (+)-stereoisomer would be about 14 nM, assuming no competition from the bio- logically inactive (-)-form. Of course, if the (-)-stereoisomer is a competitive inhibitor of ( +)-5-methyl-H4PteGlu. uptake, and has a similar affinity for the uptake system but is not transported, then the true K, for (+)-5-methyl-H4PteGlu uptake would be 27 nM and its V,,,,, value would be about 47 PM +min-1.
It can be seen that ionic (or buffer) strength, as noted earlier with 5-methyl-HJ’teGlu and PteGlu transport, affects the affinities for the transport system(s) but not maximal uptake rates (Table III).
An apparent K, value of approximately 1 PM was obtained statistically for [3H]methotrexate uptake (Fig. 2A) at the 60-s level but uptake only approximated Michaelis-Menten-type kinetics.
The effects of various unlabeled folate analogs on 5-[WI- methyl-HePteGlu and [3H]PteGlu uptake by L. case-i are shown in Table IV. Each compound tested inhibited uptake competi- tively with the exception of methotrexate. Although an apparent KI value is given for methotrexate, V,,, values obtained in the presence of this compound were considerably lower than in its
7!
TABLE II
Uptake of 6-CH3-H,PteGlu, by Luctobucillus cusei
Cells (1 mg per ml) were preincubated at 37” for 5 min in 50 mM KsHPOI-100 mM sodium acetate-HaPO1 buffer, pH 6, containing 1% glucose before addition of labeled folate compounds. Uptake was measured at the indicated times.
Intracellular concentration Folate Extracellular
concentration 1 I 2 I 5 I 10 30 60 I 120
t&K
(f)-5-CHs-HdPteGlu 35.5 3755 4312 (+)-5-CH3-H,PteGlus 2.61 0.48 0.69 (+)-5-CH3-HIPteGlur 23.2 4.80 6.76 (+)-5-CHt-HbPteGlua 0.30 0.31 0.30 (+)-5-CHa-H,PteGlus 0.28 0.01 0.02 (+)-5-CHS-H,PteGluT 0.45 0.02 0.05 (+)-5-CHa-HbPteGlus 0.56 0.08
4350 4498 1.31 2.60 6.17 5.26 0.28 0.29 0.06 0.08 0.09 0.13 0.05 0.03
%+I afler min
0.51 0.12 0.02 0.26
3843 3.20 6.64 0.47 0.22 0.45 0.09
3.57 5.97 0.31 0.30 0.15 0.15
by guest on April 8, 2019
http://ww
w.jbc.org/
Dow
nloaded from

absence. It appeared to be a mixed inhibitor of monoglutamate pounds with the consequent removal of much of the substrate uptake. from the medium.
As all compounds tested had similar affinities for both the 5- [14C]methyl-HJ%eGlu and [3H]PteGlu uptake systems, it seems unlikely that these compounds are transported by separate sys- tems. Also, KI values for PteGlu, are similar to their uptake K, values (Table III), which suggests that all the compounds tested are transported by the same system. The slight difference be- tween K, and Kr values for PteGlu, are easily explained by the high cell concentrations needed to measure uptake of these com-
L-Glutamic acid, cr-r-glutamyl-L-glutamic acid, and y-n-glu- tamyl-L-glutamic acid (100 pM) had no effect on 5-[i4C]methyl- H41’teGlu or [3H]1%eGlu transport.
E&z of Intracellular Vitamin-The efflux of labeled vitamin was studied with L. casei that had been preloaded for various times wi .h [3H]l%eG1u. Exit followed first order kinetics, and rate constants are shown in Table V. With cells that had been pre- loaded for 5 min with relatively large amounts of [3H]I%eGlu (0.5 PM), considerable reuptake of vitamin lost by efflux took place, as evidenced by the increased exit rate when uptake was
FIG. 3. Sephadex G-25 column chromatography of intracellular and extracellular labeled folates after incubating Lactobacillus casei with [i4C]PteGlua (A) or [i4C]PteGlus (B). Cells (2 mg per ml) were preincubated at 37” for 5 min in 50 mM KzHP04-100 mM so- dium acetate-HaPOd buffer, pH 6, containing 1% glucose before the addition of [“C]PteGlua (7 PM, A) or [i4C]PteGlus (5 PM; B). After 60 min, cells were filtered, and intracellular vitamin was extracted with boiling buffer. The elution positions of reference compounds under comparable conditions are indicated beneath the abscissae.
2247
TABLE IV
Inhibition of PteGZu(A) and (A)-~-CH~-H&~GZU(B) transport in Lactobucillus casei at pH 6.5 by
jolute analogs
Cells (0.05 mg per ml) were preincubated at 37” for 5 min in 50 mM K*HPO,-100 mM sodium acetate-HsPOd buffer, pH 5.5, containing 1% glucose before addition of various concentrations of either [aH]PteGlu (A) (0.5 Ci per mmol) or 5-[14C]methyl- HdPteGlu (B) (54 mCi per mmol) together with fixed concentra- tions of the indicated unlabeled folate analogs. Uptake was meas- ured at 1, 2, 5, and 10 min.
Analog KI
A I B
Methotrexate* 0.031 0.033 lo-CHO-PteGlu 0.035 0.040 (&)-HAPteGlu 0.038 0.031 (f)-5-CHa-HdPteGlu 0.033 0.037 PteGlu 0.016 0.020 PteGlut 0.054 0.063 PteGlur 0.16 0.14 PteGlur 1.2 1.4 PteGlus 1.2 1.8 PteGluc 7.0 7.2 PteGluT 10.2 9.7 Glu >500 >500 Glu-m-Glu >500 >500 Gl~-~-Glu >500 >500
a Mixed inhibitor of folate uptake.
TABLE III
Uptake parameters for jolate transport in Luctobacillus casei
Cells (0.02 mg per ml for monoglutamates, 1.0 mg per ml for polyglutamates) were preincubated at 37” for 5 min in 50 mM K~HPOI- 100 mM sodium acetate-HaPOd buffer containing 1% glucose at the indicated pH before addition of labeled folate. Uptake was measured at 1, 2, 5, and 10 min. V,,, was calculated from 1 min uptake data.
Folate PH K, LIZ S.E.
PteGlu 6.0 PteGlua 6.0 PteGlu, 6.0 PteGlus 6.0 PteGlua 5.5 PteGlu 5.5
(+)-5-CHa-H,PteGlu” 5.5 (&)-5-CHa-HtPteGlu 5.5
w
0.028 f 0.005 (7) 0.32 f 0.13 (5) 1.9 f 0.3 (5) 3.7 f 1.2 (5)
0.110 f 0.010 (7) 0.030 * 0.001 (7) 0.071 f 0.004 (7) 0.027 f 0.003 (7)
l’mx zt SE.
/dM~ miK
14.2 rt 1.1 (7) 15.6 f 2.0 (5) 13.5 r!c 2.4 (5)
6.0 f 0.7 (5) 15.1 f 0.6 (7) 14.0 f 0.9 (7) 23.1 f 0.9 (7) 23.7 zt 0.7 (7)
Concentration at 503 maximum growth*
Velocity at 50% maximum growth
nld nu.rnin-
0.93 456 0.93 45.2 1.52 10.8 4.49 7.3
a Uptake in 10 mM K~HPOI-HZPOI buffer containing 1% glucose. b From data of Tamura et al. (15).
by guest on April 8, 2019
http://ww
w.jbc.org/
Dow
nloaded from

2248
TABLE V
First order exit rates (k) of labeled jolate from Lactobacillus casei Cells (0.2 mg per ml) in 50 mM KzHPOd-100 mM sodium ace-
tate-HtPO( buffer, pH 6.0, containing 1% glucose were preloaded at 37” for the indicated times with 0.5 PM 13H]PteGlu (500 mCi per mmol) or 7 nM 13H]PteGlu (37 Ci per mmol) before resuspen- sion in buffer-glucose plus indicated additions at a concentration of 0.1 mg per ml. Efflux was measured at 2, 4, 6, and 8 min. So was the intracellular labeled folate concentration after preload- ing.
PW-
‘%2
min
5
60
120
240
Efflux media
Buffer + azide (50mM) + iodo-
acetate (20 mM) + PteGlu (50 PM)
Buffer + azide (50 mM) + iodo-
acetate (20 mM) + PteGlu
Buffer + azide (50 mM) + iodo-
acetate (20 mM) -l- PteGlu (50 PM)
Buffer + azide (50 mM) + iodo-
acetate (20 mM)
Preloaded with 7 no PteGlu
k
min-1
0.020
6.05 0.0017
7.00 0.0016 6.85 0.008
7.12 0.0024 6.01 0.0015 6.24 0.006
Preloaded with 0.5 pld PkGlU
PM
64.3 59.2
65.4 90.3 92.6
94.6
k
min-’
0.015 0.0043
0.048 0.035 0.0079
0.054
inhibited by excess unlabeled PteGlu. Exit rates were very low in the presence of metabolic inhibitors. Reloading cells for 60 min with [3H]PteGlu (0.5 C(M) did not significantly affect the exit rate of labeled vitamin when reuptake was inhibited by excess unlabeled PteGlu, but the net exit rate in buffer alone was in- creased. It appeared that influx of vitamin was decreased com- pared to cells that had been preloaded for 5 min and suggests some metabolic control over the transport of PteGlu. This would also explain the overshoot effect noted earlier with PteGlu up- take.
Cells preloaded with low levels of [3H]PteGlu (7 nM) for 5 min exhibited a similar exit rate to that found with 0.5 PM PteGlu- loaded cells; so it was apparent that the exit system was not saturated under the conditions of these experiments. With longer preloading times with 7 pM [3H]1%eGlu, and consequent intra- cellular metabolism to polyglutamate forms, the exit rate dropped to very low levels. This was not a result of increased reuptake, as excess unlabeled PteGlu had little effect on these rates. Metabolic inhibitors increased the exit rates under these conditions but the rates were still very low. It should be noted that the intracellular vitamin concentration was similar irrespective of preloading time, so the change in exit rate must have been solely a function of folate form.
ilfetabolism of [3H]PteG/u by L. casei-Buehring et al. (13) have shown that L. casei grown in the presence of [3H]PteGlu (25 nM) for 24 hours contain mainly pteroyloctaglutamates while growth medium apparently contained shorter length polyglutamates. In our studies, preloading times and [3H]PteGlu concentrations were varied to control the degree of metabolism of I’teGlu. Addition of glutamic acid (1 mM) to the preloading and efflux media did not discernibly affect metabolism. Metabolism of intracellular vitamin was investigated in preloaded cells at 0, 5, 20, and 60
FIG. 4. Metabolism of [3H]PteGlu by Lactobacillus casei. Cells (0.2 mg per ml) suspended in 50 rn~ K2HP04-100 rnM sodium ace- tate-HaP04 buffer, pH 6, containing 1% glucose were preloaded at 37” with [3H]PteGlu (0.5 FM; specific activity: 0.5 Ci per mmol). After 5 min, the cells were filtered, a portion extracted with boiling buffer, and intracellular extract (equivalent to 0.4 mg cells) and cell-free medium (1 ml) was applied to Sephadex G-25 columns (200 X 0.7 cm). The elution patterns of intracellular (--) and extracellular (- - -) labeled vitamin are shown in Fig. 4A. The remaining cells were resuspended in buffer (0.4 mg per ml) contain- ing various additions and reincubated at 37” for 60 min. Sephadex G-25 elution patterns of extracted intracellular (0.4 mg cells) and extracellular (1 ml) vitamin are shown in Figs. 4B and 4C, respec- tively. Additions to the buffer were: --, none; .... , unlabeled PteGlu (50 PM); -- - -, iodoacetic acid (20 mM) plus sodium azide (50 mM). Column fractions were collected after the void volume (V,) as indicated by the elution position of blue dextran 2000 and the elution positions of reference compounds under comparable conditions are indicated beneath the abscissae.
min after resuspension in fresh media. Exit rates were checked to ensure that they corresponded to the rates shown in Table V.
The Sephadex G-25 elution patterns of cell extract and medium after a 5-min preloading with [3H]PteGlu (0.5 pM) are shown in Fig. 4A. Practically all of the tritium label was associated with the monoglutamate fraction. The Sephadex G-25 elution pat- terns of intracellular labeled vitamin after resuspension of these cells in fresh buffer for 60 min are shown in Fig. 4B. By this time,
by guest on April 8, 2019
http://ww
w.jbc.org/
Dow
nloaded from

2249
FIG. 5. Metabolism of [sH]PteGlu by Lactobacillus casei. Ex- perimental conditions are as described in the legend to Fig. 4, except cells were preloaded for 60 min with [3H]PteGlu (0.5 PM; specific activity: 0.5 Ci per mmol).
cells incubated in buffer alone had metabolized I’teGlu to several compounds, including some polyglutamates eluting before stan- dard PteGluo. One mctabolite (Peak A in Fig. 4B) eluted just after standard PteGlua at the approximate position of a lo- formyl-PteGlu2 derivative, while another (Peak B in Fig. 4B) eluted at the approximate position of a IO-formyl-PteGlu de- rivative. A similar elution pattern was seen in cells suspended in buffer containing excess unlabeled PteGlu although less labeled vitamin was retained by these cells as reuptake was inhibited (Fig. 4B). Conversely, cells resuspended in buffer plus iodoace- tate contained higher levels of labeled vitamin as efflux was inhibited (Fig. 4B). In this case, a large part of the intracellular vitamin was metabolized to the compound resembling a lo- formyl-PteGluz derivative (Peak A in Fig. 4B) and a smaller polyglutamate fraction was observed. The Sephadex G-25 elution patterns of tritium in the media after resuspending the cells for 60 min are shown in Fig. 4C. Extracellular vitamin released from cells incubated in buffer alone consisted mainly of the IO-formyl- PteGluz derivative (Peak A) plus some monoglutamate (Peak D) while vitamin from cells incubated in buffer containing excess
B
FRACTION NUMBER (3.0ML)
FIG. 6. DEAF-cellulose chromatography of 10.formyl-PteGluz derivatives. [3H]folate peaks eluted from Sephadex G-25 columns at the approximate position of a IO-formyl-PteGlut derivative (Peak A, Figs. 4 and 5) were treated with hog kidney conjugase and chromatographed on DEAE-cellulose as described in the legend to Fig. 7. A, the elution pattern obtained with folate derivative from the medium of cells suspended in buffer alone (Peak A, Fig. 5C); B, the pattern with intracellular derivative from iodoacetate- treated cells (Peak A, Fig. 5B).
unlabeled PteGlu was, as expected, mainly in the monoglutamate form (Peak D) plus a compound with the chromatographic prop- erties of a 10.formyl-PteGlu derivative (Peak B in Fig. 4C). Comparison of this elution pattern with the pattern obtained after a 5-min resuspension of cells in buffer containing excess un- labeled PteGlu (not shown) indicated that practically all of Peak B was released in the first 5 min. i\Iost of the intracellular folate released from the cells after a 60.min resuspension in buffer containing metabolic inhibitors eluted at the position of the lo- formyl-PteGluz derivative (Peak A in Fig. 4C).
Fig. 5, A to C, shows comparable results for cells that had been preloaded for 60 min with [W]l’teGlu (0.5 PM) before resuspen- sion in fresh buffer for 60 min. After preloading, the intracellular vitamin was mainly in the monoglutamate form (Fig. 5A) as was found after a 5-min preloading (Fig. 4A). However, resuspension of the cells led to further metabolism of intracellular vitamin (Fig. 5B). The lo-formyl-PteGlut and lo-formyl-1’teGlu deriva- tives built up (Peaks A and B, Fig. 5B) as well as a compound with the elution position of a PteGluz derivative (Peak C). A similar pattern was found with cells resuspended in buffer con- taining excess unlabeled PteGlu while most of the intracellular vitamin in iodoacetate-treated cells eluted at the position of a IO-formyl-PteGluz derivative (Peak A, Fig. 5B). The Sephadex G-25 elution patterns of label in the media of cells incubated in buffer alone or buffer plus excess unlabeled 1’teGlu (Fig. 5C) showed that the main released forms after 60 min were lo-formyl- PteGlu and PteGluz derivatives (Peaks B and C) with some lo- formyl-PteGlu2 derivative (Peak A) and monoglutamate (Peak D). Again, the net release of the IO-formyl-PteGlu derivative
by guest on April 8, 2019
http://ww
w.jbc.org/
Dow
nloaded from

2250
0 )(A20 I 40 60 ;,;z; CH3t14PlrGIu PtaGlu
FRACTION NUMBER (3.OML 1
FIG. 7. DEAE-cellulose chromatography of the lo-formyl- PteGlu derivative. [$H]folate eluted from Sephadex G-25 columns at the approximate position of a lo-formyl-PteGlu derivative (Peak B, Fig. 5C) was applied to a DEAE-cellulose column (25 X 0.9 cm). The column was eluted by an exponential phosphate gradient formed with 0.01 M potassium phosphate buffer, pH 6, (100 ml) in a closed mixing chamber attached to a reservoir of 0.6 M potassium phosphate buffer, pH 6. The elution positions of refer- ence compounds, applied with the [tH]folate sample, are indicated beneath the abscissae.
(Peak B) appeared to be complete by 5 min. No lo-formyl- PteGlu derivative was found in the medium of iodoacetate- treated cells (Fig. 5C), only a buildup of the IO-formyl-PteGluz derivative (Peak A) plus some monoglutamate.
The identity of the 10.formyl-PteGluz derivative (Peak A, Fig. 4, B and C; Fig. 5, B and C) was confirmed by DEAE-cellu- lose chromatography. The folate eluted as a single peak after conjugase treatment at the position of lo-formyl-HdPteGlu (Fig. 6) indicating it to be 10.formyl-HIPteGlut. Similarly, the lo- formyl-PteGlu derivative (Peak B, Figs. 4C and 5, B and C) was identified to be 10.formyl-PteGlu (Fig. 7).
It was apparent that only a small amount of polyglutamate of chain length greater than two was formed under the described conditions and it appeared that triglutamate synthesis was a rate-limiting step in metabolism of PteGlu. It also appeared that iodoacetate inhibited the synthesis of polyglutamates. In order to study the further metabolism of PteGlu and to study efflux of higher polyglutamates, cells were preloaded for 2 or 4 hours with low levels of [3H]PteGlu (7 nM). Although some variation in polyglutamate composition was found under these conditions, the results shown in Fig. 8, A to C, are typical of the effects noted.
The Sephadex G-25 elution pattern of intracellular labeled vitamin after preloading with [3H]PteGlu (7 nM) for 120 min (Fig. 8A) showed that most of the vitamin consisted of polyglu- tamates with chain length 23. Folate in the medium was of shorter glutamate chain length (Fig. 8A) with considerable amounts of mono- and diglutamate forms. Resuspension of these cells for 60 min in buffer alone led to further chain elongation of intracellular folate (Fig. 8B) and practically no net release of labeled vitamin (Fig. 8C). With cells incubated in buffer con- taining iodoacetate, a small release of mono- and diglutamates was observed as well as a folate which eluted just before standard PteGlua from Sephadex G-25 (Fig. 8C). There was less apparent intracellular chain elongation with iodoacetate-treated cells (Fig. 8B) and in some experiments no chain elongation was observed. Intracellular labeled folate from cells suspended in buffer alone for 60 min was treated with conjugase and chromatographed on DEAE-cellulose (Fig. 9A). Most of the tritium label eluted at
I(
d
A 120min
n NO ADDITION
,o c MEDIUM
0 b - I
L I I :
I I 5 I I , NO ADDITION
FIG. 8. Metabolism of [3H]PteGlu by Lactobacillus casei. Ex- perimental conditions are as described in the legend to Fig. 4, except cells were preloaded for 120 min with PH]PteGlu (7 nM; specific activity, 37 Ci per mmol).
the position of HJ’teGlu (Peak D, Fig. 9A, 72%) with some lo- formyl-HdPteGlu (Peak B, 13%), 5-methyl-HdPteGlu (Peak C, lo%), and a trace of PteGlu (5%). Material eluted straight through the column (Peak A, Fig. 9) represented breakdown products of unknown composition. These breakdown products were always observed when high specific activity (37 Ci per mmol) PteGlu was used (13). Iodoacetate-treated cells, after conjugase treatment, also contained 10.formyl-HIPteGlu (Peak B, Fig. 9B, 19%), 5-methyl-HJ’teGlu (Peak C, Fig. 9B, 6%), HJ’teGlu (Peak D, Fig. 9B, 19%), as well as breakdown products (Peak A, Fig. 9B). However, most of the radioactive label eluted from DEAE-cellulose at the position of X (54oj,), the 5-methyl- H&eGlu breakdown product (Fig. 9B). The material eluted at Peak X was rechromatographed on Sephadex G-25 (Fig. 10) and, although it had been treated with conjugase, still eluted just prior to standard PteGlus. It was apparent from this that the chain lengths of polyglutamates found in iodoacetate-treated cells (Fig. 8B) were lower than their elution positions from Sepha- dex G-25 suggested.
The small amounts of labeled vitamin released from cells SUS-
by guest on April 8, 2019
http://ww
w.jbc.org/
Dow
nloaded from

A
: . known folate forms, and appeared to be breakdown products.
E DISCUSSION
t I .
2 5 c D 5-Methyl-HaPteGlu and PteGlu uptake by nonproliferating
A cells of Lactobacillus casei possessed characteristics of a carrier- mediated active process (34, 36, 37). Vitamin was taken up and retained against a concentration gradient and the transport process was dependent on energy, pH, temperature, ionic effects,
6
t I
and displayed structural specificity and saturation kinetics.
t B ‘2
PteGlus.5 uptake also demonstrated many of the properties of
X6 an active process. As suggested by Buehring et al. (13), PteGlu
2251
3%
? Q ’
i2.
peaks were found, equivalent to the two peaks of Fraction A in Fig. 9. The reason for this conjugase effect was unclear. How- ever, none of the radioactive label released from cells containing polyglutamates of chain length 2 3 corresponded to any of the
FRACTION (3.0 ML)
FIG. 9. DEAE-cellulose chromatography of labeled folates in Lactobacillus casei. Cells (0.2 mg per ml) suspended in 50 mM K2HPOa-100 mM sodium acetate-HsPOd buffer, pH 6, containing 1% glucose were preloaded at 37” for 120 min with [3H]PteGlu (7 nM; specific activity: 37 Ci per mmol), filtered, and reincubated in A. fresh buffer or B. buffer containine: azide (50 mM) and iodoace- take (20 mM) (0.5 vol) for 60 min. Intracellular vitamin was ex- tracted with boiling buffer, treated with hog kidney conjugase, and aliquots (equivalent to 0.4-mg cells) were chromatographed on DEAE-cellulose as described in the legend to Fig. 7.
polyglutamates were transported without prior hydrolysis. Although Cooper (38) suggested that PteGlu and 5-methyl-
HdPteGlu were transported by separate uptake systems in L. casei, the similarities we found between K, values for uptake and K, values measured against both [3H]l’teGlu and 5-[i4C]- methyl-HJ’teGlu uptake suggests that all folates tested were transported by the same system. The K, value of 0.03 PM for PteGlu uptake was similar to that reported by Cooper (38) (0.045 PM). The only qualitative difference between uptake of PteGlu compared to other folates was an inhibition of uptake with time. This apparent metabolic control was probably a con- sequence of culturing the cells with PteGlu as folate source.
A decrease in transport system affinities and maximal trans- port rates was observed with increasing pteroylpolyglutamate chain length and glutamic acid dipeptides did not affect uptake. It appears that the specificity of the uptake system was for the pteroyl moiety of folate compounds.
L I- x 5 E 2
f IO T 20 f 30 40 50? 60 VO PIeGlug PteGlu3
FRACTION NUMBER (1.7 ML)
PteGlu,
FIG. 10. Sephadex G-25 chromatography of Peak X. The conju- Ease-treated [3H]folate derivative from iodoacetate-treated &zctobacillus casei, which eluted at the position of X, the 5CHa- HaPteGlu breakdown nroduct (Fig. 9B), was chromatogranhed on Sephadex G-25. The elution posit&s of reference compounds are indicated beneath the abscissa.
pended in buffer alone and buffer containing iodoacetate (Fig. 8C) and the material eluting just before standard PteGlur (Fig. 8C) were chromatographed on DEAE-cellulose before and after conjugase treatment. Each sample, before conjugase treat- ment, eluted straight through the column at the position of the
The study of the exit mechanism was complicated by metab- olism and reuptake of vitamin. Exit was inhibited by iodoace- tate, suggesting a carrier-mediated system. When cells were loaded with relatively large amounts of [3H]PteGlu, very little folate containing more than 2 glutamate residues was formed and it appeared that triglutamate formation was rate-limiting. Sakami et al. (40), in studies with Neurospora crassa, reported the presence of two polyglutamate synthetases, one specific for
the formation of pteroyldiglutamate and the other for synthesis of longer chain polyglutamates. Our results could also be ex- plained by two synthetases with similar properties. Under con-
ditions of limited triglutamate synthesis, lo-formyl-H&eGluz, 10.formyl-PteGlu, and a pteroyldiglutamate built up. IO-Formyl- PteGlu was rapidly released by L. casei and was not detected in
iodoacetate-poisoned cells, which metabolized PteGlu mainly to 10.formyl-H&eGluz. This latter compound was probably the “early eluting folate” described by Cooper (38) which had simi-
lar properties to, but was distinct from, 10.formyl-PteGlu, and whose formation was inhibited by methotrexate. These results are also in agreement with the observation of Ohara and Silber (41) that the specific activity of L. casei 10.formyl-H4PteGlu synthetase was highest in late-log phase cells. Although some
first part of Peak A, Fig. 9. After conjugase treatment, two differences in exit rates with iodoacetate-treated cells might be
Comparisons of transport parameters with growth data ob- tained previously by Tamura et al. (15) suggests that, although pteroylpolyglutamates are transported at a slower rate, once transported they are more effectively utilized by L. casei for growth than is PteGlu.
[3H]Methotrexate uptake did not follow Michaelis-Menten- type kinetics, probably because the compound is rapidly broken down and released by L. casei (39).
by guest on April 8, 2019
http://ww
w.jbc.org/
Dow
nloaded from

2252
expected due to differences in intracellular metabolism, it was by higher concentrations, although we consider this to be un- apparent that exit per se was inhibited by this compound. Iodo- likely. It may well be that the same carrier is used by L. casei acetate also appeared to inhibit polyglutamate formation in for folate uptake and exit and that this carrier will not transport these cells, which again suggests two polyglutamate synthetases reduced pteroylpolyglutamate compounds, thus .explaining in L. casei. their retention by the cells.
Buehring et al. (13) were unable to detect conjugase activity in L. casei. Our results confirm this observation as the metab- olism of [3H]PteGlu in cells resuspended in buffer plus excess unlabeled PteGlu was similar to that found in cells resuspended in buffer alone. If conjugase had been present, an equilibration of tritium between mono- and polyglutamyl folates would have occurred with a much larger proportion of the label in the mono- glutamate fraction in cells resuspended with excess PteGlu.
Acknowledgments-We would like to thank Drs. Y. S. Shin and K. U. Buehring for providing some of the pteroylpolygluta- mates used in this study.
REFERENCES
’
Sakami et al. (40) reported that only HdPteGlu, compounds were substrates for the polyglutamate synthetases of N. cMssa, while Gawthorne and Smith (42) have reported that 5-formyl- H&eGlu and 5-methyl-H$teGlu would also serve as substrates for the sheep liver enzyme. In our studies, with cells metabolizing low levels of [3H]PteGlu, the major polyglutamates present were H4PteGlu, forms. Although this would suggest a specificity of the polyglutamate synthetase for HdPteGlu, compounds, recent studies by Shin el al. (39) have demonstrated the presence of [3H]PteG1u4 in methotrexate-treated L. casei after incubation with [3H]PteGlu and the formation of methotrexate polygluta- mates by rat liver has been well documented (39, 43). Efflux of labeled vitamin from cells containing pteroylpolyglutamates was very low and the released forms appeared to be breakdown products. Less chain elongation took place in iodoacetate-treated cells containing pteroylpolyglutamates and, although exit rates were increased slightly, only breakdown products were released. The major folate in these poisoned cells was of unknown struc- ture and, after conjugase treatment, exhibited similar chromato- graphic properties on DEAE-cellulose and Sephadex G-25 to the 5-methyl-H4PteGlu breakdown product (X). It is difficult to predict folate form from chromatographic properties (for in- stance, IO-formyl-PteGlu elutes much earlier from Sephadex G-25 and DEAE-cellulose than PteGlu or IO-methyl-PteGlu (10, 39)), but as 5-[14C]methyl-H4PteGlu is labeled in the 5- methyl position, the 5’.carbon must be retained in compound X, suggesting it to be a~5- or lo-substituted folate derivative. The unidentified folate found in iodoacetate-treated cells is probably of similar structure and, although we were unable to carboxy- methylate [3H]PteGlu under the conditions of the experiment, it is quite conceivable that carboxymethylation of one of the naturally occurring reduced folates had taken place. The greater efflux of breakdown products from poisoned cells suggests that this compound was more labile than H4PteGlu polyglutamates.
SHANE, B., AND STOKSTAD, E. L. R. (1974) Fed. Proc. 33,1359 PFIFFNER, J. J., CALI~INS, D. G.. BLOO~I. E. S.. AND O’DELL.
B. L. (1946) J: Am. Chek. Sot. is,1392 ’ ’ ROHRINGER, It., KIM, W. K., AND S~MBORSKI, D. J. (1969) Can.
J. Biochem. 47, 1161-1169 EIUCSON, L. E., WIDOFF, E., AND BANHIDI, Z. G. (1953) Acta
Chem. &and. 7,974-979 HUTCHINS, B. L., STOKSTAD, E. L. R., BOHONOS, N., SLOANE,
N. H., >\ND SU~BARO~, Y. (1948) J. Am. Chem. Sot. 70, l-3
6.
7.
8.
9. 10.
11.
12.
13.
14.
15.
16.
SIROTNAK, F. M., DONATI, G. J., AND HUTCHINSON, D. J. (1963) J. Bacterial. 86,658-665
NORONHA, J. M., AND SILVERMAN, M. (1962)J. Biol. Chem. 237, 3299-3302
NORONH~, J. M., AND ABOO~AKER, V. S. (1963) Arch. Biochem. Biophys. 101, 445-447
WHITE&AD, V. M. (1971) Blood 38,809 SHIN, Y. S., BUEHRING, K. U., AND STOKSTAD, E. L. R. (1972)
J. Biol. Chem. 247, 7266-7269 SHIN, Y. S., WILLIAMS, M. A., AND STOKSTAD, E. L. R. (1972)
Biochem. Biophys. Res. Commun. 47.35-43 BAUGH, C. M.,.&EVENS, J. C., AND KRUMDIECK, C. L. (1970)
Biochim. Biowhus. Acta 212. 116-125 BUEHRING, K. ~.,“TAMuR,~, T:, AND STOKSTAD, E. L. R. (1974)
J. Biol. Chem. 249, 1081-1089 SHIN, Y. S., BUE~RING, K. U., AND STOKSTAD, E. L. R. (1974)
Arch. Biochem. Biophys. 163, 211-224 TAMURA, T., SHIN, Y. S., WILLIAMS, M. A., AND STOKSTAD,
E. L. R. (19721 Anal. Biochem. 49.517-521 BLAKLEY, k. L. ‘(1968) in Frontiers if Biology (NEUBERGER, A.,
AND TATUM, E. L., eds) Vol. 13, p. 29, North-Holland, Amsterdam
17.
18.
BAUGH, C. M., KRUMDIECK, C. L., BAKER, H. J., AND BUTTER- WORTH, C. E., JR. (1971) J. Chin. Invest. 60,2009-2021
HOFFBRAND, A. V., TRIPP, E., HOULIHAN, C. M., AND SCOTT, J. M. (1973) Blood 42,141-146
19. WRIGHT; B. E. (1955) J. Am. Chem. Sot. 77,3930-3932 20. HIM~X. R. H.. AND RABINOWITZ. J. C. (1962) J. Biol. Chem.
21. 237, i903-29i4
\ I
LAI~GE, P. J., AND QUAYLE, J. R. (1963) Biochem. J. 87, 386- 396
22. GUEST. J. It., FRIEDMAN, S.. FOSTER, M. A., TEJERINA, G.,
It appears, therefore, that intracellular metabolism of folates to polyglutamates, in L. casei at least, serves two main purposes. One is the formation of a more effective cofactor for folate- dependent enzymes and the other, the formation of folates which are retained by the cells. No folates of glutamate chain length greater than two were released by L. casei.
23. AND iyoo~s; D. D. (1964) Biochem. J.‘92,4971504
WHITFIELD. C. D.. STEERS. E. J.. JR.. AND WEISSBACH. H.
24.
25. 26.
(1970) J. biol. kern. 246, i90-4Oi ’ CURTHOYS, N. P., STRXJS, L. D., AND RABINOWITZ, J. C.
(1972) Biochemistry 11,345-349 POWERS, S. G., AND SNELL, E. E. (1974) Fed. Proc. 33,137l MCELWEE, P. G., AND SCOTT, J. M. (1972) Biochem. J. 127,
901-905 27.
Although McElwee and Scott (26) reported that S. faecalis actively transported 5-methyl-HdPteGlu, we were unable to confirm their results. Pediococcus cerewisiae, as reported by Mandelbaum-Shavit and Grossowicz (27) did actively transport 5-methyl-H4PteGlu, although it could not metabolize this com- pound, but was unable to transport 5-methyl-HhPteGlus-s. L. casei did take up some 5-methyl-H4PteGlua-8 but there was no evidence supporting an active accumulation. Low concentrations of these compounds were used in uptake studies and the possi- bility remains that an uptake system could have been induced
MANDELBAUM-SHAVIT, F., AND GROSSOWICZ, N. (1970) J. Bac- teriol. 104, l-7
28. 29. 30. 31.
CHANARIN, I., AND PERRY, J. (1967) Biochem. J. 106,633-634 DAVIS, L. (1968) Anal. Biochem. 26,459-460 BLAKLEY, R. L. (1959) Biochem. J. 72, 707-715 BII~D, 0. D., MCGLOHON, M. V., AND VAITKUS, J. W. (1965)
Anal. Biochem. 12, 18-36 32. 33.
WILKINSON, G. N. (1961) Biochem. J. 80, 324-332 LINEWEAVER, H., AND BURK, D. (1934) J. Am. Chem. Sot. 66,
658-666 34. KEPES, A., AND COHEN, G. N. (1962) in The Bacteria
(GUNSALUS, I. C., AND STANIER, R. Y., eds) Vol. 4, pp. 179- 222, Academic Press, New York
by guest on April 8, 2019
http://ww
w.jbc.org/
Dow
nloaded from

2253
35. ZAKRZEWSKI, S. F., AND GRZELAKOWSKA-SZTABERT, B. (1973) 40. SAKAMI, W., RITARI, S. J., BLACK, C. W., AND RZEPKA, J. J. Biol. Chem. 243,2684-2690 (1973) Fed. Proc. 32, 471
36. CIRRILO, V. P. (1961) Ann. Rev. Microbial. 16.197-218 41. OHARA, O., AND SILBER, R. (1969) J. Biol. Chem. 244, 193% 37. HOLDEN, J. T. (1962) in Amino Acid Pools (HOLDEN, J. T., ed) 1993
pp. 566-594, Elsevier Publishing Company, Amsterdam 42. GAWTHORNE, J. M., AND SMITH, R. M. (1973) Biochem. J. 136, 38. COOPER, B. A. (1970) Biochim. Biophys. Acta 208,99-109 295-301 39. SHIN, Y. S., BUEHRING, K. U., AND STOKSTAD, E. L. R. (1974) 43. BAUGH, C. M., KRUMDIECK, C. L., AND NAIR, M. G. (1973)
J. Biol. Chem. 249, 5772-5777 Biochem. Biophys. Res. Commun, 62, 27-34
by guest on April 8, 2019
http://ww
w.jbc.org/
Dow
nloaded from

B Shane and E L StokstadTransport and metabolism of folates by bacteria.
1975, 250:2243-2253.J. Biol. Chem.
http://www.jbc.org/content/250/6/2243Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/250/6/2243.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 8, 2019
http://ww
w.jbc.org/
Dow
nloaded from