titina y evidencia de un sistema tenso activo
TRANSCRIPT

Tejido Conectivo: Un subsistema pasivo¿Mito o Realidad?
David López PT DC

La estructura neuromusculoesquelética se organiza funcionalmente como una pieza prismatica tridimensional deformable, que se expresa en la variabilidad de su eje de gravedad o linea baricentrica, la cual es una K dependiente de la suma de los cambios espaciales que ocurren a nivel de sus componentes, determinando su postura y equilibrio (tensión, desplazamiento, esfuerzo, deformación).
Deformación
Tensión
Desplazamiento de componentes
Equilibrio Y
Postura
¿Tejido conectivo: Subsistema pasivo?

Relación Tensión – Longitud de los tejidos
carg
as o
Str
ess
Deformación de tensión
NZ EZ PZ
Rango Fisiológico
RangoTraumático
NZ = Neutral ZoneEX = Elastic ZonePZ = Plastic Zone
Rotura
“Injury to ligamentous tissue is injury to mechanoreceptors”
Yamashita et al., Spine, 1993

Cadenas de movimiento
• Representan circuitos en continuidad de dirección y de planos a través de los cuales se propagan las fuerzas del cuerpo se realiza una acción en las tres dimensiones
• Su organización esta dada en torno a la función corporal en relación a la intencionalidad de movimiento.
• Permiten constantes adaptaciones corporales en términos de equilibrio y estabilización requeridas para la realización de una acción minimizando gasto energético
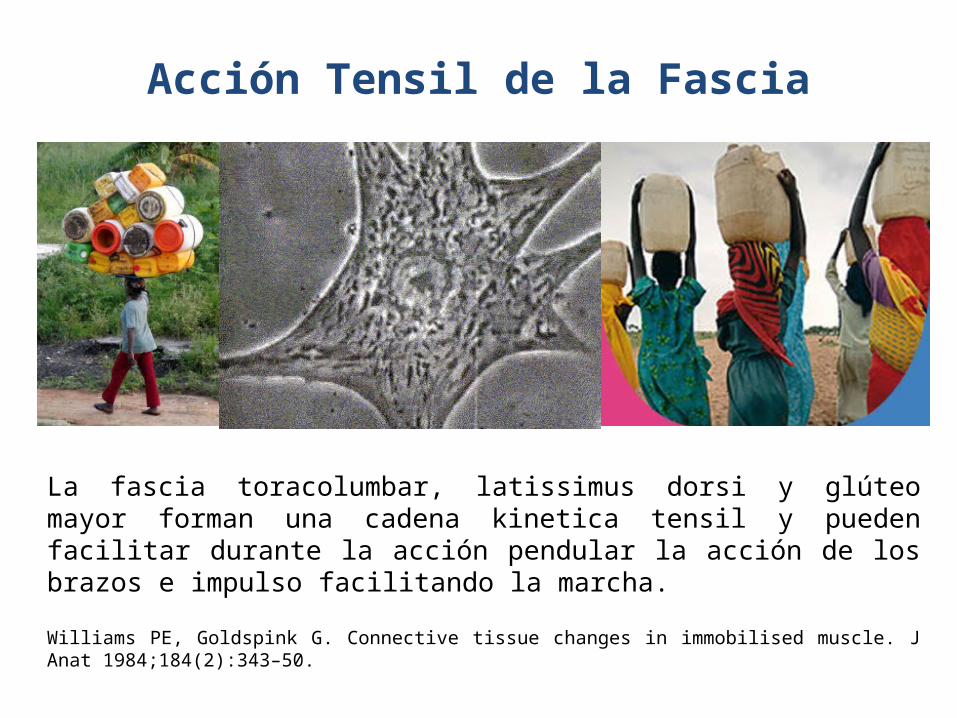
Acción Tensil de la Fascia
La fascia toracolumbar, latissimus dorsi y glúteo mayor forman una cadena kinetica tensil y pueden facilitar durante la acción pendular la acción de los brazos e impulso facilitando la marcha.
Williams PE, Goldspink G. Connective tissue changes in immobilised muscle. J Anat 1984;184(2):343–50.

Tanto la tensión como la compresión fascial pueden variar en el músculo su estado de tensión basal, asimismo el cambio de la tensión de la fascia durante la contracción muscular, será determinante del stiffness muscular al inicio y durante un ejercicio. Role of fascia in maintenance of muscle tension and pressure Garfin et al. J Appl Physiol.1981; 51: 317-320
La fascia puede contraerse como un músculo liso y alterar su comportamiento mecánico
R. Schleip1, W. Klingler1,2 and F. Lehmann-Horn1 Active fascial contractility: fascia maybe able to contract in a smooth muscle-like manner and thereby influence musculoskeletal dynamics. Med Hypotheses 65: 273-277 2005

Acción Muscular
Tensión Fascial
Cambio en la función Fascial tenso activa
Cambio en la función muscular motoactiva
Passive muscle stiffness may be influenced by active contractility of intramuscular connective tissue. Robert Schleip *, Ian L. Naylor, Daniel Ursu, Werner Melzer, Adjo Zorn, Hans-Joachim Wilke, Frank Lehmann-Horn, Werner Klingler

Panjabi 2005. La falla tensil del tejido conectivo y la consecuente corrupción de la señal
propioceptiva mecanorreceptora
¿Rol del Subsitema pasivo?

Desarrollo de la explicación de la contracción muscular
• La contracción muscular se produce exclusivamente por los filamentos contráctiles de actina y miosina durante la acción muscular concéntrica e isométrica (Huxley & Hanson; Huxley & Niedergerke,1954).
• Explicaciones de hace poco más de una década, descubren que las contracciones excéntricas se asocian con una fuerza atribuible al menos en parte a otras causas.
Huxley, H. E., and Hanson, J. (1954). Changes in cross-striations of muscle during contraction and stretch and their structural implications. Nature 173, 973–976. Structural changes in muscle during contraction. Interference microscopy of living muscle fibers. Nature 173, 971–973

Andrew Huxley en 1957 modeló la teoría de los puentes cruzados de la contracción muscular.
Constitucion en doble espiral Unida al disco Z Se fijan a ella dos proteinas:
Tropomiosina Troponina
Huxley, A. F. (1957). Muscle structure and theories of contraction. Prog. Biophys. Biophys. Chem. 7, 255–318.
La teoría de los puentes cruzados

(A) original modelo de 2 estados propuesto por Huxley (1957). (B) Multi-modelo de estado de los puentes cruzados de la contracción muscular con cabezal giratorio según lo propuesto inicialmente por Huxley (1969). (C) Multi-estado basado en la estructura atómica de puentes cruzados y sitios de unión actina como propone Rayment et al. (1993).
Evolución del modelo de los puentes cruzados de la contracción muscular.

En su descripción original de Huxley (1957) fue capaz de predecir con precisión las fuerzas de las contracciones concéntricas mediante un conjunto de constantes de relación fuerza-velocidad concéntrica de los músculos estriados (Hill, 1938).
También la teoría de los puentes cruzados explicó muy bien la fuerza isométrica como una función de longitudes de fibra y del sarcómero (Gordon et al., 1.966).
La teoría de los puentes cruzados
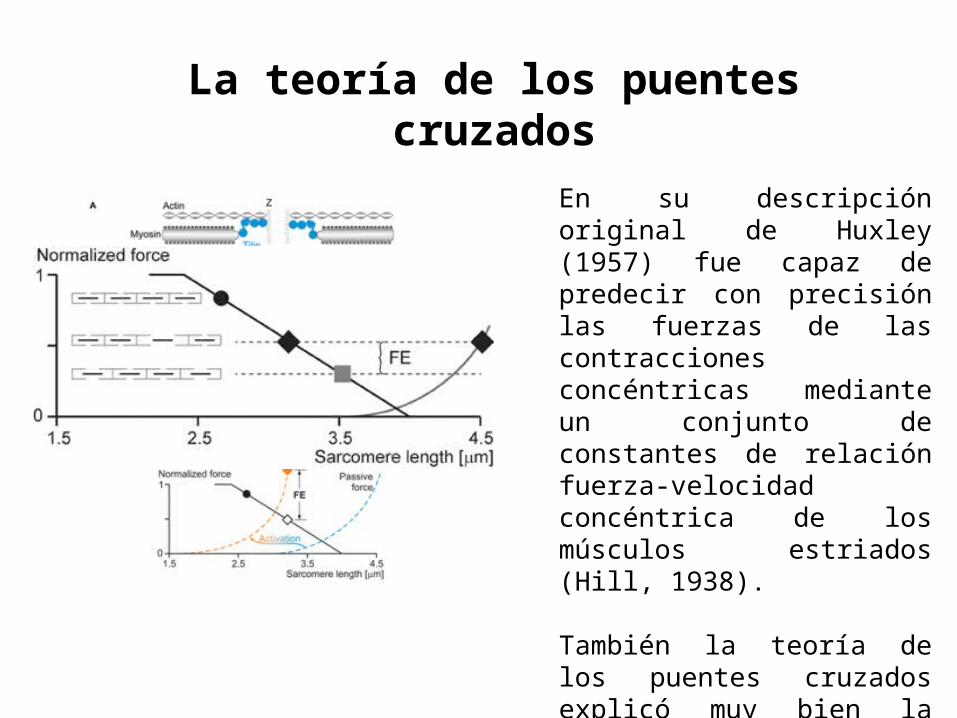
En su descripción original de Huxley (1957) fue capaz de predecir con precisión las fuerzas de las contracciones concéntricas mediante un conjunto de constantes de relación fuerza-velocidad concéntrica de los músculos estriados (Hill, 1938).
También la teoría de los puentes cruzados explicó muy bien la fuerza isométrica como una función de longitudes de fibra y del sarcómero (Gordon et al., 1966).
La teoría de los puentes cruzados

El incremento de fuerza residual después del estiramiento activo de los músculos (Abbott y Aubert, 1952;. Edman et al, 1982) no podía predecirse conceptualmente (Walcott y Herzog, 2008) o explicarse numéricamente por la teoría de los puentes cruzados (Herzog et al, 2012a, b; Herzog, 2014).
La teoría de los puentes cruzados y contracción excéntrica
Walcott, S., and Herzog, W. (2008). Modeling residual force enhancement with generic cross-bridge models. Math. Biosci. 216, 172–186. Herzog, W. (2014). Mechanisms of enhanced force production in lengthening (eccentric) muscle contractions. J. Appl. Physiol. (1985) 116, 1407–1417.

La no uniformidad de longitud del sarcómero aumentan cuando un músculo se estira de forma excéntricamente activa en comparación con las contracciones isométricas (Morgan, 1990, 1994; Allinger et al, 1996). Rassier et Al, 2003b, demostraron que la relación fuerza-longitud durante el estiramiento de sarcómeros dispuestos en serie de no produjo inestabilidades.Los sarcómeros eran perfectamente estables en longitudes muy diferentes en la rama descendente de la relación fuerza-longitud (Rassier et al, 2003b).
Allinger, T. L., Epstein, M., and Herzog, W. (1996). Stability of muscle fibers on the descending limb of the force- length relation. A theoretical consideration. J. Biomech. 29, 627–633Rassier, D. E. (2012). Residual force enhancement in skeletal muscles: one sarcomere after the other. J. Muscle Res. Cell. Motil. 33, 155–165
Teoría de la no uniformidad de longitud del sarcómero (Morgan, 1990)

Si asumimos la que mejora la fuerza en un solo sarcómero, como se observa en la literatura (Leonard et al., 2010), es causada por medio las no uniformidades de longitud del sarcómero y que la fuerza activa es proporcional a la cantidad de filamentos actina-miosina en solapamiento acoplado (contraídos).
Entonces la fuerza activa tendría que comenzar a descender como se indica en la figura (línea gruesa continua negra). La fuerza Pasiva tendría que ser constante.
Controversia Planteada por los hallazgos de Herzog

¿Cómo podemos explicar la mejora fuerza residual post elongación?
• Si no es la tensión residual inducida por la mayor longitud del sarcómero y sus no uniformidades ¿qué la produce?
• Hace una década atrás, descubrimos la existencia de un componente pasivo de la mejora de la fuerza residual que sugiere fuertemente que la titina, una proteína estructural provoca esta directamente relacionada con la mejora de la fuerza pasiva (Herzog et al., 2006)
Herzog, W., Lee, E. J., and Rassier, D. E. (2006). Residual force enhancement in skeletal muscle. J. Physiol. 574, 635–642.

La Titina es una proteína conectiva cuyo rol es activo durante la contracción y elongación
muscular• La Titina aumenta su rigidez con la
activación muscular mediante la unión del calcio sus sitios especializados (Labeit et al., 2003).
• Labeit et al. (2003) identificaron la región llamada PEVK de la titina como un sitio de unión para el calcio rigidizándose.
• La Titina también puede cambiar su longitud mediante la unión de su región proximal a la actina tras la activación y aumentar la fuerza muscular. Leonard y Herzog (2010).

Figure 1. Experimental strategies for measuring interaction forces between PEVK and F-actin. (a) PEVK fragments were attached to the surface of streptavidin-coated latex beads via their C-terminally located cysteine residue using maleimide-biotin cross-linking...
Pasquale Bianco, Attila Nagy, András Kengyel, Dávid Szatmári, Zsolt Mártonfalvi, Tamás Huber, Miklós S.Z. Kellermayer
Interaction Forces between F-Actin and Titin PEVK Domain Measured with Optical Tweezers ☆
null, Volume 93, Issue 6, 2007, 2102–2109
http://dx.doi.org/10.1529/biophysj.107.106153

La Titina aumenta su tensión durante la contracción muscular
Herzog, W., and Leonard, T. R. (2013). Residual force enhancement: the neglected property of striated muscle contraction. J. Physiol. 591(Pt 8), 2221

“El subsistema pasivo no existe; el tejido conectivo es totalmente activo durante la tensión muscular y el movimiento”. David López DC