the role of root anatomy and root architecture in
TRANSCRIPT

Purdue UniversityPurdue e-Pubs
Open Access Theses Theses and Dissertations
January 2016
The Role of Root Anatomy and Root Architecturein Resistance to Ralstonia solanacearumDenise Caldwell CaldwellPurdue University
Follow this and additional works at: https://docs.lib.purdue.edu/open_access_theses
This document has been made available through Purdue e-Pubs, a service of the Purdue University Libraries. Please contact [email protected] foradditional information.
Recommended CitationCaldwell, Denise Caldwell, "The Role of Root Anatomy and Root Architecture in Resistance to Ralstonia solanacearum" (2016). OpenAccess Theses. 1128.https://docs.lib.purdue.edu/open_access_theses/1128

Graduate School Form30 Updated
PURDUE UNIVERSITYGRADUATE SCHOOL
Thesis/Dissertation Acceptance
This is to certify that the thesis/dissertation prepared
By
Entitled
For the degree of
Is approved by the final examining committee:
To the best of my knowledge and as understood by the student in the Thesis/Dissertation Agreement, Publication Delay, and Certification Disclaimer (Graduate School Form 32), this thesis/dissertation adheres to the provisions of Purdue University’s “Policy of Integrity in Research” and the use of copyright material.
Approved by Major Professor(s):
Approved by:Head of the Departmental Graduate Program Date
Denise L. Caldwell
THE ROLE OF TOMATO ROOT ANATOMY AND ROOT ARCHITECTURE IN RESISTANCE TO RALSTONIASOLANACEARUM
Master of Science
Dr. Anjali Iyer-PascuzziChair
Dr. Peter B. Goldsbrough Co-chair
Dr. Mary Alice WebbCo-chair
Dr. Anjali Iyer-Pascuzzi
Dr. Christopher J. Staiger 7/20/2016

i
THE ROLE OF ROOT ANATOMY AND ROOT ARCHITECTURE
IN RESISTANCE TO RALSTONIA SOLANACEARUM
A Thesis
Submitted to the Faculty
of
Purdue University
by
Denise L. Caldwell
In Partial Fulfillment of the
Requirements for the Degree
of
Master of Science
August 2016
Purdue University
West Lafayette, Indiana

ii
I dedicate this thesis to my husband Frank and our children. Frank, you have always
encouraged me to reach for things that I never thought I could achieve, and never once
did you ask me to be false to myself.
~ You are my Soul ~
Parker, I thank you for teaching me about true dedication and leadership. You have
shown me what hard work and pursuit of your dreams can accomplish.
~ You are my Determination ~
Chloe, you have offered me your strength when all of mine had fled. I will never forget
the countless words of encouragement, nor the times that you have told me that you are
proud of me.
~ You are my Strength ~
Gabby, you inspire me every day. You have a natural curiosity about the world around
you and the ability to shrug off anyone who thinks you are different with a smile.
~ You are my Wonder ~
Josslyn, you have the sweetest soul. Many days, I have come home feeling beat down and
exhausted. You cuddle in my lap and restore me.
~ You are my Joy ~
Without each of you, I could not have accomplished this goal. Thank you!

iii
ACKNOWLEDGEMENTS
I would like to thank Dr. Iyer-Pascuzzi for bringing me into her lab and developing my
talents. My wonderful lab members for all of their encouragement and support during my
time in Dr. Anjali Iyer-Pascuzzi’s lab. John Cavaletto for taking me under his supervision
and invoking enthusiasm to teach within me. Deb Lubelski for her countless hours
ensuring that our plants remain alive. And a special thank you to Dr. Christopher Gilpin
and Laurie Muller of the Life Science Microscopy Facility for their countless hours of
instructions.

iv
TABLE OF CONTENTS
Page
LIST OF FIGURES .......................................................................................................... vii
LIST OF ABBREVIATIONS ............................................................................................ ix
ABSTRACT ....................................................................................................................... xi
CHAPTER 1. INTRODUCTION ................................................................................. 1
1.1 Overview and Objectives ..................................................................................... 1
1.2 Introduction to the Pathogen and Bacterial Wilt Disease ..................................... 2
1.3 Tomato Root Anatomy and Root Architecture .................................................... 8
1.4 Lateral Root Development ................................................................................. 11
1.5 Conclusions ........................................................................................................ 12
CHAPTER 2. MATERIALS AND METHODS ........................................................ 14
2.1 Introduction ........................................................................................................ 14
2.2 Root Anatomy Experiment ................................................................................. 14
2.2.1 Seed Preparation ............................................................................................ 14
2.2.2 Stratification & Scarification ......................................................................... 15
2.2.3 Plant growth and Ralstonia solanacearum (K60) culture and infection ....... 15
2.2.4 Sample Collection .......................................................................................... 16
2.2.5 Fixation (Killing & Preserving) ..................................................................... 17
2.2.6 Dehydration ................................................................................................... 18
2.2.7 Infiltration ...................................................................................................... 19
2.2.8 Embedding ..................................................................................................... 20
2.2.9 Mounting Specimen Blocks ........................................................................... 20
2.2.10 Trimming Blocks ........................................................................................... 20

v
Page
2.2.11 Sectioning ...................................................................................................... 21
2.2.12 Deparaffination, Rehydration, Staining and Dehydration ............................. 21
2.2.13 Mounting Slides ............................................................................................. 21
2.2.14 Imaging .......................................................................................................... 22
2.2.15 Measurement .................................................................................................. 22
2.3 Scanning Electron Microscope ........................................................................... 22
2.4 Root Architecture Experiment ............................................................................ 23
CHAPTER 3. RESULTS ............................................................................................ 26
3.1 Introduction ........................................................................................................ 26
3.2 Root Anatomy .................................................................................................... 27
3.2.1 Resistant HA variety of tomato have larger xylem tracheary elements ........ 27
3.2.2 Bacteria are distributed differently within the vascular cylinder in resistant and susceptible plants .................................................................................... 28
3.2.3 Bacteria colonize the root vascular cylinder of resistant plants at 48 hpi ..... 30
3.2.4 Additional resistant and susceptible lines were examined to test whether the pattern of colonization was specific to the resistant variety HA ................... 32
3.2.5 Distribution of R. solanacearum in the stem differs between resistant HA-I and susceptible WV-I .................................................................................... 32
3.3 Root Architecture ............................................................................................... 33
3.3.1 Root architecture of resistant HA-I differs from that of other varieties ........ 33
3.4 Results Summary ................................................................................................ 34
CHAPTER 4. DISCUSSION ...................................................................................... 53
4.1 Introduction ........................................................................................................ 53
4.2 Colonization ....................................................................................................... 54
4.2.1 Delay in bacteria colonization of the HA variety .......................................... 54
4.2.2 Distribution of bacteria within the root ......................................................... 55
4.2.3 Susceptible WV parenchyma cells collapse .................................................. 57
4.3 Architecture ........................................................................................................ 57
4.3.1 Root architecture of resistant HA-I differs from that of other varieties ........ 57
4.4 Area of Vessels ................................................................................................... 59

vi
Page
4.4.1 Resistant HA variety of tomato have larger xylem tracheary elements ........ 59
4.5 Conclusions ........................................................................................................ 60
LITERATURE CITED ..................................................................................................... 61
APPENDICES
Appendix A .................................................................................................................. 68
Appendix B .................................................................................................................. 70
Appendix C .................................................................................................................. 71
Appendix D .................................................................................................................. 72
Appendix E .................................................................................................................. 73
VITA ................................................................................................................................. 74
PUBLICATION ................................................................................................................ 78

vii
LIST OF FIGURES
Figure ............................................................................................................................ Page
Figure 1.1. Cross-section of HA root approximately 24 days after germination showing different root cell-types and tissues .............................................................. 13
Figure 2.1. Example of tomato root tissue used in experiments ..................................... 24
Figure 2.2. Demonstration of how tracheary element size was calculated using imageJ software ........................................................................................................ 25
Figure 3.1. Bacterial wilt disease tomato phenotypes at 144 hours post inoculation (hpi) ...................................................................................................................... 35
Figure 3.2. Comparison of primary root anatomy of HA-MI and WV-MI at 144 hpi .... 36
Figure 3.3. Comparison of primary root T1-MI and T2-MI at 144 hpi........................... 37
Figure 3.4. Comparison of mock-inoculated root tracheary element average area between tomato varieties at 144 hpi ............................................................. 38
Figure 3.5. SEM cross-sections of HA-I at 24hpi ........................................................... 39
Figure 3.6. SEM cross-sections of HA-I at 72 hpi .......................................................... 40
Figure 3.7. SEM cross-sections of HA-I at 144 hpi ........................................................ 41
Figure 3.8. SEM cross-sections of WV-I at 24 hpi ......................................................... 42
Figure 3.9. SEM cross-sections of WV-I at 72 hpi ......................................................... 43
Figure 3.10. SEM cross-sections of WV-I at 144 hpi ..................................................... 44
Figure 3.11. Confirmation of R. solanacearum colonization within the root ................. 45
Figure 3.12. Comparison of bacterial colonization of HA-I and WV-I at 0, 24. 48, 72 and 144 hpi with light microscopy ...................................................................... 46

viii
Figure ............................................................................................................................ Page
Figure 3.13. Comparison of bacterial colonization of T1-I and T2-I at 0, 72 and 144 hpi with light microscopy ................................................................................... 48
Figure 3.14. Magnified SEM cross-section of paraffin embedded hypocotyl comparing bacterial colonization of HA-I and WV-I at 144 hpi ................................... 49
Figure 3.15. Cross-section of paraffin embedded hypocotyl comparing anatomical structure of HA-I and WV-I at 144 hpi with light micrographs ................... 50
Figure 3.16. Comparison of lateral root emergence of mock-inoculated and inoculated resistant and susceptible varieties at 144 hpi ............................................... 51
Figure 3.17. Percent change in lateral root emergence of Inoculated/Mock-Inoculated at 144 hpi by variety ......................................................................................... 52

ix
LIST OF ABBREVIATIONS
ABBREVIATION DESCRIPTION
µg Microgram
BW Bacterial wilt
C Celsius
CFU Colony forming units
cm Centimeter
CPG Casamino acid-Peptone-Glucose
ddH2O Double distilled water
EPS Exopolysaccharides
ETD Everhart Thornley detector
EtOH Ethanol
g Gram
gpt Grams per plant tissue
HA Hawaii 7996 variety
hpi Hours post inoculation
I R. solanacearum inoculated
IAA Auxin
K2O Potassium
kV Kilovolt
M Molar
MI Mock-inoculated
ml Milliliter
mm Millimeter
Mp Megapixel

x
ABBREVIATION DESCRIPTION
N Nitrogen
OD Optical density reading
P2O5 Phosphate
QTL Quantitative trait loci
R Resistant
R. solanacearum Ralstonia solanacearum
RH Relative humidity
ROI Region of Interest
RSSC Ralstonia solanacearum species complex
S Susceptible
SEM Scanning electron microscopy
SiQ2 Silicon Dioxide
T1 CRA-66 variety
T2 Okitsu variety
T3Es Type III effectors
TBA Tert-Butyl Alcohol
TTSS Type III secretion system
WV West Virginia 700 variety
x Magnification

xi
ABSTRACT
Caldwell, Denise L. M.S., Purdue University, August 2016. The Role of Tomato Root Anatomy and Root Architecture in Resistance to Ralstonia solanacearum. Major Professor: Dr. Anjali Iyer-Pascuzzi. Bacterial wilt is a devastating plant disease that can cause upwards of 90% crop loss. The
soil-borne bacterium Ralstonia solanacearum, is the causal agent of bacterial wilt, and
infects over 200 plant species, including tomatoes. Roots are fundamental to resistance in
tomato, as grafting resistant rootstocks to susceptible scions results in resistant plants.
Despite the devastation it causes, there are no known resistance genes in crops to R.
solanacearum, and crop defense mechanisms are unknown. Here, we investigated the
role of root anatomy and root architecture in tomato resistance to R. solanacearum. We
find that bacteria colonize root tissue types of resistant and susceptible plants differently,
and that vascular treachery elements are generally larger in resistant plants. Further, we
find that the production of lateral roots may contribute to mechanisms of resistance. Our
data suggest that differences in root anatomy and architecture are part of plant defense
mechanisms to R. solanacearum.

1
CHAPTER 1. INTRODUCTION
1.1 Overview and Objectives
The soil-borne bacterium Ralstonia solanacearum is the causal agent of bacterial
wilt (BW) and is ranked as the second most destructive bacterial plant pathogen
(Mansfield et al., 2012). Root diseases such as bacterial wilt are among the most
challenging plant diseases to control, as chemical methods are often not effective below
ground where soil-borne pathogens reside (Haas & Defago, 2005). The best means of
disease control of bacterial wilt is through resistant varieties (Yuliar & Toyota, 2015) but
the plant genes underlying resistance in crop varieties are unclear, and no plant resistance
genes to R. solanacearum in crop varieties have been identified. Understanding the
mechanisms through which plants mount an effective defense response is essential to
ensure effective crop protection.
R. solanacearum enters the root through small natural wounds such as those
created during lateral root emergence (Vasse, 1995). The pathogen primarily colonizes
the root xylem of both resistant and susceptible plants (Digonnet et al., 2012a; Digonnet
et al., 2012b; Vasse, 1995). Xylem is a complex water-conducting tissue composed of
both living and dead cells. Defense mechanisms in the root appear to be key to whole
plant resistance, because grafting resistant (R) tomato rootstocks to susceptible (S) scions
significantly reduces bacterial wilt disease (McAvoy et al., 2012; Rivard et al., 2008).

2
Despite the importance of roots in bacterial wilt resistance, and that disease
control is primarily managed through resistant rootstocks and cultivars (Huet, 2014; Scott
et al., 2005) there is little information about root defense mechanisms. Because R.
solanacearum primarily colonizes the root xylem tissue of both resistant and susceptible
tomatoes, this tissue may play an important role in root defense mechanisms. Further,
because R. solanacearum enters the root through wounds created during lateral root
emergence, lateral roots may be an important factor that affects resistance or
susceptibility.
The objectives of this thesis were to test the following hypotheses:
• Bacteria colonize the vascular cylinder of resistant plants more slowly than in susceptible plants
• Bacteria are distributed differently within the vascular cylinder of resistant and susceptible plants
• Resistant plants have larger xylem vessels, tracheary elements and a larger vascular cylinder
• Resistant and susceptible plants undergo different changes to root architecture after infection
1.2 Introduction to the Pathogen and Bacterial Wilt Disease
R. solanacearum was first described by E.F. Smith in 1896 on Solanaceae species
including tomato, potato and eggplant (Álvarez et al., 2010). R. solanacearum infects
over 200 plant species and is particularly devastating to tomatoes and other solanaceous
crops (Hayward, 1991; Scott et al., 2005). The bacteria was named Bacillus
solanacearum, later Pseudomonas solanacearum and currently Ralstonia solanacearum
(Denny, 2006). It is now found on all continents but primarily located in regions with

3
higher temperatures and humidity. It is a severe limiting factor in tomato production in
the Southeastern United States and in Central America (Hayward, 1991; McAvoy et al.,
2012; Rivard & Louws, 2008). This soil-borne pathogen is a class beta-proteobacteria
because it is gram-negative bacteria, rod shaped, and has a polar flagellum.
Due to the diversity within R. solanacearum, the species is known as the
Ralstonia solanacearum species complex (RSSC) (Fegan & Prior, 2005). The RSSC is
divided into four phylotypes (I, IIA/B, III and IV), each corresponding (approximately) to
a different geographic region. The strain used in this research, R. solanacearum K60, is
classified as phylotype IIA, sequevar 7. K60 was isolated in 1953 from tomatoes in North
Carolina, and strains causing current outbreaks in that region are similar to the original
isolate (Remenant et al., 2012). The genome of K60 has been completely sequenced
(Remenant et al., 2012). Recent work has proposed splitting the RSSC into 3 species
(Prior et al., 2016; Remenant et al., 2012) based on genomic and proteomic analysis and
metabolic characterization. The three new species would group phylotype IIA/B as R.
solanacearum, with phylotype I and III as R. pseudosolanacearum and phylotype IV as
R. syzygii.
Disease management of bacterial wilt is very limited as the pathogen is a soil
bacterium. The best means of disease control is through resistant cultivars. Resistance in
tomato is primarily quantitative, resulting from many genes contributing a small amount
to resistance (Danesh, Aarons, McGill, & Young, 1994; Thoquet, Olivier, Sperisen,
Rogowsky, Laterrot, et al., 1996; Young & Danesh, 1994). Although several quantitative
trait loci (QTL) for resistance to R. solanacearum have been identified (Carmeille et al.,
2006; Thoquet et al., 1996; Wang et al., 2013; Wang et al., 2000), none have been fine-

4
mapped or cloned. HA7996 is considered one of the most highly resistant tomato
genotypes to a range of R. solanacearum strains (Scott et al., 2005). This genotype shows
no disease symptoms, yet maintains a high pathogen load.
Many factors contribute to the spread of bacterial wilt disease. For example,
infected machinery can transfer the disease to unaffected soil, as can water runoff.
Control of infected plants in the field is challenging because there are no initial infection
symptoms and once symptoms are visible the disease progresses rapidly. Chemical
treatment with bactericides has not been effective because once symptoms are observed,
the chemicals will not be able to move throughout the plant system to eradicate the
bacteria (Hölttä & Nikinmaa, 2013). Removing infected plants and surrounding soil can
prevent rainwater runoff from carrying the bacteria to nearby plants although in large
field populations, this method is not practical or economical.
Grafting resistant rootstocks to susceptible scions is an effective means of disease
control and is used in the Southern United States as well as in South Korea and Japan
(McAvoy et al., 2012; Rivard et al., 2012). Scions of the susceptible BHN 602 tomato
line were grafted onto rootstocks of various tomato lines with resistance to bacterial wilt
(McAvoy et al., 2012). When disease pressure was low, BW incidence of both grafted
and non- or self-grafted was low. When BW disease pressure was high, both fruit yield
and BW incidence of grafted hybrids was much better than non-grafted and self-grafted
hybrids (McAvoy et al., 2012).
Grafting resistant rootstocks had similar effects on resistance in susceptible scions
in a field study in North Carolina (Rivard et al., 2012). These studies demonstrate the
importance of roots for resistance and tomato yield.

5
R. solanacearum likely gains access to its host through natural wounds caused by
the emergence of lateral roots, and through wounds acquired from growing through the
soil. After entering the root, bacteria infect the root cortex, and then move to the root
vasculature where they multiply in the xylem and subsequently move into the shoot
(Digonnet, et al., 2012; Vasse, 1995). The pathogen secretes exopolysaccharides (EPS),
and increasing amounts of EPS are thought to lead to physical blockage of xylem that
prevents transpiration, resulting in wilting and eventual death (Denny, 2000; Denny &
Baek, 1991; McGarvey et al., 1999; Saile et al., 1997; Schell, 2000). Xylem colonization
appears to be critical to disease progress, because mutants impaired in xylem colonization
cannot cause wilting in plants (Plener et al., 2010; Vasse et al., 2000).
Experiments on Medicago truncatula have illustrated that there are four basic
steps involved in the process of infection by a R. solanacearum (Turner et al., 2009).
Senescence of the root is first observed followed by the bacteria entering the intercellular
spaces of the root cortex and proceeding to the vascular cylinder to allow colonization of
the vascular system to take place. Once the bacteria are able to access the central vascular
system they are able to rapidly reproduce within the plant’s xylem tissue.
Once in this prime location, the bacteria secrete EPS and cause disease symptoms to be
present (Grimault & Prior, 1993). Tissue weakened by secondary root branches allow
more sites for infection with R. solanacearum (Vasse, 1995).
Several studies have shown that R. solanacearum colonizes both resistant and
susceptible plants (Ishihara et al., 2012; McGarvey et al., 1999; Nakaho & Allen, 2009).
For example, R. solanacearum colonized roots, hypocotyls, and the lowest petiole of both
susceptible variety Marion and resistant variety Hawaii 7996 (HA) at 2 dpi (McGarvey et

6
al., 1999). Colonization was 100-fold higher in the petioles and mid-stems of the
susceptible varieties compared to the resistant HA variety, yet the resistant plant
displayed no disease symptoms. Although resistant tomato plants can maintain high
pathogen levels (Ishihara et al., 2012), several reports suggest that bacteria in resistant
plants are limited in their ability to broadly colonize tissues in the stem (Grimault et al.,
1994; Grimault & Prior, 1993; Nakaho et al., 2004). For example, resistant varieties
restricted bacteria to the primary stem xylem, possibly through thickenings of pit
membranes in stem xylem tissue (Nakaho et al., 2004). However, in the susceptible line,
bacteria were also found in secondary xylem tissues of the stem (Nakaho et al., 2004).
Although grafting resistant rootstocks to susceptible scions significantly reduces
bacterial wilt disease (McAvoy et al., 2012; Rivard et al., 2012), bacterial distribution in
roots of resistant plants has not been examined. Further, infection differs in the root of
resistant and susceptible lines, and how bacteria are distributed within the different root
tissues of the root is not clear.

7
Bacteria produce different quantities of EPS in both resistant and susceptible
plants. Susceptible plants had approximately 10 μg EPS per gram of plant tissue (10 μg
/gpt) in the taproot, hypocotyl and mid-stem regions compared to resistant HA plants,
which never exceeded 1 μg/gpt in any of the sampled regions. When the maximum
number of bacterial was reached in resistant HA plants, EPS production did not increase
further. In susceptible plants, however, EPS production continued to increase by as much
as 10-fold. Surprisingly, although EPS production per gram of plant tissue was less in
resistant plants compared to susceptible plants, the amount of EPS per bacterial cell was
by about 15-fold higher in the resistant variety compared to the susceptible plants
(McGarvey et al., 1999).
R. solanacearum produces phytohormones such as auxin (IAA) and ethylene
(Valls et al., 2006). Susceptible and resistant tobacco plants were found to have large
increases of auxin when infected with R. solanacearum (Sequeira & Kelman, 1962).
Phytohormones work to regulate many phases of plant development including growth,
reproduction, environment adaptations and disease resistance (Pieterse, Leon-Reyes, Van
der Ent, & Van Wees, 2009). The role of the hormones produced by R. solanacearum in
defense is not understood.
To maintain virulence, R. solanacearum is equipped with several pathogenicity
factors, including EPS (Denny & Baek, 1991; McGarvey et al., 1999; Milling et al.,
2011; Saile et al., 1997), cell wall degrading enzymes such as cellulase and pectic lyase
(Allen et al., 1997; Huang & Allen, 1997; Liu et al., 2005) and type III effectors (T3Es).
Like many bacterial plant pathogens, R. solanacearum uses the type III secretion system
(TTSS) to inject its T3Es into the host cell, causing disease in susceptible plants and

8
promoting defense responses in resistant plants (Angot et al., 2006; Coll & Valls, 2013;
Jacobs et al., 2013; Remigi et al., 2011; Sole et al., 2012; Turner et al., 2009; Vasse et al.,
2000). Although the function of most T3Es is unknown, (Coll & Valls, 2013), genetic
analyses have demonstrated that several T3Es have roles in R. solanacearum virulence or
avirulence.
The expression of T3Es is dependent on hrp genes. Two hrp genes, hrpG and
hrpB, are necessary for tomato root infection, although at different stages of the process
(Vasse et al., 2000). HrpB is required for xylem colonization (Vasse et al., 2000). The
hrpB regulon is expressed in planta in the initial stages of plant infection, during
colonization of xylem by bacteria, and in later stages of wilting (Jacobs et al., 2013;
Monteiro et al., 2012; Zhang et al., 2013).
1.3 Tomato Root Anatomy and Root Architecture
Given the important role that roots play in resistance to R. solanacearum, root
anatomical and architectural analyses will likely provide insight about tomato
mechanisms of resistance or susceptibility to R. solanacearum. Root anatomy can be
defined as the internal cellular arrangement, morphology, and distribution (Jung &
McCouch, 2013) while root architecture focuses on the pattern, length, orientation and
diameter of different branches on the root system (Fitter & Atkinson, 1991).

9
The three main types of roots formed in tomato cultivars are primary, lateral and
adventitious roots. Primary root development occurs at the time the embryo is
developing, with lateral and adventitious roots forming postembryonically. Each type of
root consists of at multiple tissues and cell types. These include the epidermis, cortex,
endodermis, pericycle, xylem tissue, and phloem tissue. Because the xylem tissue is the
major focus of this work, it will be discussed in more detail.
Primary growth of the root vasculature is less complex as the radial system has
not developed yet. Once secondary growth has initiated, the complexity of cells within
the xylem tissue increases. The primary function of xylem is conductance of water and
there are many different cell types that contribute to the proper function of the xylem
tissue. One statement that is often made about xylem tissue is that it is dead at maturity
but this is misleading, as only certain cells within the tissue are dead. Xylem parenchyma
is comprised of living parenchyma cells that are responsible for translocation and storage
of materials such as carbohydrates, proteins, crystals, fats and oils. The non-vascular
tissues called the pericycle are usually made of parenchyma cells. In tomato, the
cambium is interrupted by two protoxylem poles that are directly opposite from one
another and are the site of lateral root initiation (Esau, 1960).
The vascular cambium develops between the phloem and xylem and the plant
undergoes secondary growth. As the vascular cambium grows, in a diarch root such as
tomato, two bands meet with the pericycle and together girdle the xylem core. The
activation of the vascular cambium is the point of activity for secondary growth. The
vascular cambium acts as a meristematic tissue to increase the circumference of the root.
The water conducting elements or tracheary elements are made of two cell types,

10
tracheids and vessel members and are dead upon maturity. The phloem becomes
dispersed within the cambial tissue. During the activation of secondary growth the xylem
core increases in diameter, pushing the endodermis, pericycle and cortex to the outer
edge where it will eventually be crushed and slough off in the soil (Esau, 1960).
Tracheids are imperforate cells with water flowing between the bordered pit-pairs
that connect them. Vessel members have perforations with apertures at one or both ends
of the cell as well as pits to allow water to flow freely. Vessels are a collection of vessel
members joined end to end. The tracheary cells closest to the pericycle are the first to
mature but remain the narrowest, named protoxylem (primary growth) with cell wall
thickenings that are annular to helical in structure. In the center, the metaxylem elements
begin to form with increasing vessel diameter as the vascular cylinder becomes more
distant from the root apical meristem (Esau, 1960).
A cross section of a root from HA7996 was used to create a model showing
different root cell-types and tissues of a root 24 days after germination (Fig. 1.1). At this
stage, secondary growth of the cambial tissue is apparent. In this section it is unclear if
any epidermal tissue remains attached to the external surface of the root. The xylem is a
complex tissue composed of multiple cell types interspersed with each other: parenchyma
cells (living), tracheids (non-living) and vessels (non-living). Approximate regions were
used because cells become interspersed during secondary growth and identification of a
specific cell type is difficult.

11
1.4 Lateral Root Development
Lateral roots are branches that extend from the primary root, and are a major
component of root architecture. Lateral roots explore the surrounding soil and enhance
the plant’s ability to respond to the envioronment. Lateral root formation can be broadly
generalized into 5 steps: lateral root priming, lateral root initiation, lateral root primordia
formation, lateral root meristem outgrowth/emergence and lateral root elongation
(Malamy & Benfey, 1997). The first three stages impact the number and orientation of
lateral roots. In tomato, lateral root formation begins with specification of specific cells in
the pericycle. These pericycle cells are located at the xylem poles. Once the pericycle is
initiated a lateral root primordia is formed and will push through the surrounding tissue of
the cortex and epidermis. After the lateral root has emerged, it will elongate.
Auxin plays a major role in lateral root development and is the key factor in
specifying the pericycle cells that become lateral roots. R. solanacearum is frequently
found colonizing the lateral root primordia (Vasse, 1995), or lateral root-like structures
that occur on petunia plants inoculated with R. solanacearum (Zolobowska & Van
Gijsegem, 2006).
Like lateral roots, adventitious roots are lateral branches. However, adventitious
roots emerge from the stem and are therefore shoot-borne roots. They can form either as a
result of a normal developmental program or can be induced in propagation by wounding
or applications of hormones (Bellini et al., 2014). For example, adventitious roots can be
stimulated at preformed root initials (Jackson, 1985) during flooding stress (McNamara
& Mitchell, 1990).

12
Because R. solanacearum enters the root in part through wounds created when
lateral roots emerge, and because the pathogen can colonize lateral root primordia, we
hypothesized that inoculated resistant plants had a different root architecture compared to
susceptible plants.
1.5 Conclusions
The root is an important source of resistance to R. solanacearum, but the
mechanisms that mediate this resistance are unknown. Because R. solanacearum
colonizes the vascular cylinder, this region of the root may be key to resistance.
Therefore the work in this thesis examined several different aspects of the root vascular
cylinder after infection with R. solanacearum in resistant and susceptible varieties.

CortexEndodermisPericyclePhloem TissuePhloem ParenchymaCambial TissueXylem Tissue
Figure 1.1. Cross-section of HA root approximately 24 days after germination showing different root cell-types and tissues. Secondary growth from the cambial tissue is apparent. It is unclear if any epidermal tissue is present on this cross-section. The xylem is a complex tissue composed of multiple cell types interspersed with each other: parenchyma cells (living), tracheids (non-living) and vessels (non-living). False coloring was added to image using Photoshop.
13

14
CHAPTER 2. MATERIALS & METHODS
2.1 Introduction
Plant histology is a powerful tool that enables a researcher to observe changes in
plant tissues at specific time points during the developmental or infection process.
Fixation of the sample is the most critical step in the histological process as it determines
the quality of the section used for imaging. There are multiple recipes available for use in
plant histology and understanding and modification of these recipes is essential in the
fixation process. Jensen and the Ruzin provided basic recipes that were modified during
this investigation (Jensen, 1962; Ruzin, 1999).Root Anatomy Experiment
2.1.1 Seed Preparation
Seeds were removed from mature fruit of the parent plants by squeezing fruit into
clear water and sieving away debris. The seeds were surfaced sterilized with 15% bleach
for 5 minutes, dried, packed in non-airtight envelopes and then stored in a small plastic
container containing drierite desiccant. The containers were placed in the dark at 4° C
until use. Hawaii (HA-7996) and T1 (CRA-66) are the two resistant cultivars and West
Virginia (WVa-700) and T2 (Okitsu) are the two susceptible cultivars used in this study.
Five replications of Hawaii (HA) and West Virginia (WV) and three replications of
CRA-66 (T1) and Okitsu (T2) were completed for this study.

15
2.1.2 Stratification & Scarification
Seeds were placed in falcon tube with a 30% bleach solution and oscillated for 5
minutes. The seeds were then rinsed 8 times with distilled water or until the bleach aroma
could no longer be detected. The seeds were then placed in clean ddH2O water and placed
at 4° C overnight.
2.1.3 Plant growth and Ralstonia solanacearum (K60) culture and infection
Seeds were planted at 0.5 to 1 mm depth in rectangular containers (plug flats, 36
cell inserts) each containing 25g – 27g of Propagation Mix by Sun Gro Horticulture. The
propagation mix was comprised of 55% - 56% Canadian Sphagnum peat moss,
vermiculite, dolomite lime, wetting agent and 0.0001% Silicon Dioxide (SiQ2) from
calcium silicate.
Plants were grown in greenhouse from August 2015 to March 2016 with
supplemental lighting from high-pressure sodium lamps for 12 hours during daylight
hours. Trays were put under humidity domes until germination at which time the domes
were removed. The seedlings were bottom watering and fertilized with Peters Excel 15-5-
15 Cal-Meg Special (15% Nitrogen (N), 5% Phosphate (P2O5), 15% Potassium (K2O)) 10
days after germination.

16
The K60 strain of Ralstonia solanacearum was streaked and grown on Casamino
acid-Peptone-Glucose (CPG) medium plates and placed in a 28° C incubator for 48
hours. Bacterial colonies were scraped from plates and resuspended in sterile ddH2O. The
bacterial concentration was determined by an optical density reading, OD600nm = 0.1 as 1
x 108 Colony Forming Units per milliliter (CFU/ml). Performed titer plates were used to
confirm CFU/ml density.
Tomato plants at the three-leaf stage were either inoculated (I) with R.
solanacearum, K60 strain (1.0 x 108 cfu/ml) or soaked in sterile ddH2O (MI - mock-
inoculated). Seedlings were transferred to a 10” x 14” plastic container and inoculum was
then poured into the container so that it came up to root-shoot junction and allowed to
soak for 5 minutes. Seedlings were transferred back to their rectangular containers and
placed into a Conviron growth chamber under light conditions for 16 hours at 28°C,
relative humidity (RH) of 77% and 100 u lumen/second. For the remaining 8 hours plants
were left in the dark at a temperature of 28°C and RH of 70%.
2.1.4 Sample Collection
Mock inoculated tomato plants were collected at the three-leaf stage (approx. 15
days after planting) that represented 0 hours post inoculation (hpi). Mock inoculated (MI)
and R. solanacearum inoculated (I) plants samples were taken at 24, 48, 72 and 144 hpi.

17
The majority of the soil was removed from the root zone via a series of water dips
using an up and down motion. Plants were placed in a shallow container of distilled water
and the remainder of soil was brushed from roots using a 1-inch paintbrush in a parallel
fashion careful not to damage the primary roots. The plants were transferred to a labeled
storage container filled with water covering the root zone to ensure the roots did not dry
out.
Under a dissecting scope the stem was severed from the root at the root-shoot
junction in a perpendicular cut using a fresh double edge razor blade (Personna Stainless
Steel “PTFE coated at .004 thickness). The root-shoot junction was determined by the
color change between the stem and the root, and to the distinct line of trichomes present
at the base of the stem. The lateral roots were removed from the primary root using
MicroPoint Scissors leaving approximately 0.5 to 1 mm of lateral root material attached.
Roots were then perpendicular cut (cut 1) with a fresh double edge razor blade at a
distance of 2.5 cm from the root-shoot junction (where the dense line of trichomes
appear). Measuring 2 mm from cut 1 (cut 2) back towards the root-shoot junction another
perpendicular cut was made (Fig. 2.1).
2.1.5 Fixation (Killing & Preserving)
The 2 mm root samples were transferred to a 20 ml scintillation vial with 5 ml of
fresh 1.5% Glutaraldehyde/2% Formaldehyde in a 0.1 M Cacodylate buffer fixative
solution using forceps. Careful attention was made to ensure no pinching pressure was
applied to the root. Instead the cohesive properties of water helped adhere the root
sections to the forceps and transfer them to the fixative solution. The 2.4 cm sample

18
(between cut 1 and cut 3) was placed into a labeled 50 ml falcon tube containing 70%
ethanol and stored in 4° C refrigerator until processing at a later date for the root
architecture experiment. The scintillation vial with the 2 mm sample was then placed in a
polycarbonate vacuum desiccator on low vacuum pressure for 2 hours. It is important to
note that cuts should be administered immediately prior to submersion into fixative in
order to minimize cellular changes before the killing fixative can take effect. At the end
of two hours the vacuum pressure was slowly released on the scintillation vials. The
fixative was removed from the vials and rapidly replaced with a rinsing solution of 0.1 M
Cacodylate buffer. The samples were not allowed to dry out at any time. Small amounts
of fixative solution remaining around the tissue will be of little consequence once diluted
in new solution. The scintillation vials were placed under low vacuum pressure for 15
minutes. The rinsing procedure was performed a total of three times.
2.1.6 Dehydration
Under low vacuum pressure a gradual dehydration of specimens was performed.
15 minutes was used for each dehydration step. Solutions used in this series were 20%
Ethanol (EtOH), 50% Ethanol (EtOH), TBA I and TBA II. The TBA II solution was then
changed to TBA III and placed under low vacuum pressure for 15 minutes. After the
allotted time the vacuum pressure was shut off but kept attached with pressure inside for
a minimum of four hours (and often overnight). Because the tissue was so small it was
often hard to locate once embedded in paraffin. To assist in locating the tissue TBA IV
was made with 0.1% Safranin O stain. After removal of TBA III, TBA IV with 0.1%
Safranin O stain was added to the sample. Sample was replaced under low vacuum

19
pressure for 15 minutes. Vacuum pressure was shut off but kept attached with low
pressure inside for a minimum of four hours. Samples were dehydrated using the same
method for TBA IV and V under low vacuum pressure for 15 minutes and shutting off
vacuum pressure while keeping it attached with low pressure inside for a minimum of
four hours. After removal of the TBA V, it was replaced with 100% TBA. Scintillation
vials were capped and placed on top of paraffin oven overnight as TBA is solid at 25.5°
C. The entire process was repeated two more times.
2.1.7 Infiltration
The solution in the vial was replaced with enough fresh 100% TBA to cover all
the tissue and liquid paraffin (melted Paraplast X-tra) was added to fill vials. Since the
TBA will evaporate from the vial by pulling the paraffin into the tissue, it is essential to
ensure that you have more paraffin to TBA ratio. The vials were recapped and place into
paraffin oven set to 56°C and let sit overnight or until melted. Once 100% TBA/paraffin
mixture had completely liquefied, the vials were gently swirled to ensure mixing of the
TBA/paraffin uncapped and put back into the paraffin oven for a minimum of 24 hours.
After 24 hours, the paraffin was removed from vials and replaced with fresh molten
paraffin two more times with at least 24 hours between changes.

20
2.1.8 Embedding
A glass petri dish was placed on a standard hot plate at between 80°C - 100°C.
Scintillation vial was removed from paraffin oven and the contents of scintillation vial
were poured into the glass petri dish. Fresh melted paraffin wax was poured into a labeled
Peel Away Disposable Embedding Mold. The tissue was then arranged in the desired
orientation taking care not to apply any pressure to the specimen during this process.
Sample was set aside and left to cool at room temperature for 24 hours.
2.1.9 Mounting Specimen Blocks
A Tissue-Tek® Embedding Ring was labeled with a specimen identification
number. The disposable embedding mold was removed and the specimen was oriented so
that it would be on top of block. Excess paraffin was removed so that the area of the
block would situate itself into the embedding ring. Block was set into a plastic hexagonal
weigh boat and melted paraffin wax was poured into any gaps seen between embedding
ring and specimen block. Sample was cooled at room temperature for 2 hours. Excess
paraffin around block base was removed. Using a standard wood burning tool with a
flathead attachment, paraffin was melted where the specimen block meets the embedding
ring ensuring a solid adhesion. Specimen was cooled at room temperature for 2 hours.
2.1.10 Trimming Blocks
A single edge carbon steel razor blade with a .009” thickness was used to trim
around specimen creating a tapered trapezoid being careful to leave a little paraffin
around specimen to enable a ribbon to form.

21
2.1.11 Sectioning
A Leica/Reichert 2035 Microtome with Tissue-Tek® Accu-Edge® Disposable
High Profile Microtome Blades was used to cut samples at a thickness of 7 μm. After the
sectioning a ribbon of 10 to 15 sections, several drops of ddH2O were added atop a 3 in x
1 in Rite-On™ Microslides and the ribbon was placed atop the water with its shiny side
down. Slides were then allowed to dry (4 hours to overnight) and then placed in a
horizontal glass slide rack that had an open bottom.
2.1.12 Deparaffination, Rehydration, Staining and Dehydration
In a chemical fume hood, a series of solutions was used to deparaffinize,
rehydrate, stain and then dehydrate once again. The slide rack was placed in a coplin
staining jar for 5 minutes. The following solutions were used: 100% xylene, 50%
xylene/50% EtOH, 100% EtOH, 50% EtOH, aqueous 0.05% Toluidine Blue stain (10
minutes), 50% EtOH, 100% EtOH, 50% xylene/50% EtOH and 100% xylene.
2.1.13 Mounting Slides
Forceps were used to remove a slide from xylene, the slide was placed on a flat
surface with two drops of Permount™ Mounting Medium on slide surface. A 22 mm x 50
mm cover glass was placed onto surface and let dry for 10 minutes. Two glass vial shells
(2 dram - 7.4ml) were place on top of glass to weight glass cover flatly onto slide and
allowed to dry for 24 hours in the chemical fume hood.

22
2.1.14 Imaging
Light microscope images were captured on an Olympus BX43 upright microscope
with a SPOT Idea 5.0 Mp Color Digital CMOS camera.
2.1.15 Measurement
Calculation of dimensions from the microscope images was performed using a
micrometer to determine the number of pixels in 10 μm; 4x – 8.6683 pixels, 10x –
21.6897 pixels, 20x – 45.167 pixels, 40x – 91.2276 pixels and 100x – 232.6108 pixels.
ImageJ software was used to measure the area of the vascular cylinder by locating
the endodermis layer and outlining the inner tissue boundary. The color threshold tool
was then used to outline the cell walls. The magic wand tool was used to select the
tracheary element to be measured and the Region of Interest (ROI) tool was used to
determine the area. Our experimental data measured the 5 largest lignified tracheary
elements areas (Fig. 2.2).
2.2 Scanning Electron Microscope
Some paraffin-embedded samples were de-paraffinized and processed further to
allow viewing of samples at higher resolution in the SEM (Webb & Arnott, 1983).
Paraffin samples were cut at 30 μm, placed on a drop ddH2O situated atop a 25 mm
circular coverslip, and allowed to dry for 4 hours. After drying, the samples were then
deparaffinized in 100% fresh xylene for 5 minutes, then repeated 3 additional times.
Samples were allowed to dry (30 seconds) then placed on a 25.4 mm aluminum slotted
specimen mount with double sided tape. A line of silver conductive coating was laid

23
down between the pin and the top of the coverslip. Samples were then sputter coated
using a Cressington turbo-pumped sputter coater. SEM samples were viewed using a FEI
NOVA nanoSEM Field Emission Scanning Electron Microscope under high vacuum 5.00
kV, Spot Size 3.0 and an Everhart Thornley detector (ETD) for imaging.
2.3 Root Architecture Experiment
The 2.4 cm root samples (Fig. 2.1) that had been stored in 50 ml falcon tube
containing 70% ethanol at 4° C were used for this experiment. The 70% EtOH was
discarded and replaced with ddH2O for 30 minutes at room temperature. This removed
the hydrophobic response when the tissues were placed into staining solution. Cleaned
roots were placed into 0.05% Toluidine Blue stain for 30 seconds and washed six times
with ddH2O at 5-minute intervals.
Labeled square petri dishes were place onto the Epson Flatbed Scanner and filled
with ddH2O. The stained samples were then placed into corresponding petri dish and
scanned at 400 dpi. Samples were then returned to the falcon tube containing ddH2O and
examined using the dissecting microscope. Roots were sliced longitudinally from root-
shoot junction to bottom cut with a double edge razor blade. Forceps were used to grasp
the root and an extended needle was used to locate lateral roots and roots were counted.

Figure 2.1. Example of tomato root tissue used in experiments. A. Example of 2.5 cm of root tissue used in our experiments. B. Illustration of regions used in our experiments. A perpendicular cut was made 2.5 cm from the root-shoot junction (cut 1). Another perpendicular cut was made 2 mm above cut 1 (cut 2). The 2 mm tissue (T) was then placed in vial containing fixative for the root anatomy experiment. The (RA) tissue was used for the root architecture experiment where number of emerged lateral roots were counted.
Root-Shoot Junction
Primary Root Tip
2.5 cm
Root-Shoot Junction
Primary Root Tip
0 cm
T
RA
A B
Cut 2
Cut 12 mm
24

Figure 2.2. Demonstration of how tracheary element size was calculated using imageJ software. The total area of the vascular cylinder area was measured along with the larger xylem tracheary elements. The area of the top 5 largest tracheary elements was used to calculate the average tracheary size.
25

26
CHAPTER 3. RESULTS
3.1 Introduction
The goal of this research was to investigate the role of root anatomy and root
architecture in resistance to R. solanacearum. I hypothesized that: 1) the size of
tracheary elements in resistant varieties would differ from that in susceptible
varieties, 2) resistant varieties would have a delay in the onset of bacterial
colonization of the root vascular cylinder, 3) the regions of bacterial distribution
within the root vascular cylinder would differ compared to the susceptible varieties,
and 4) the root architecture of resistant varieties would be altered after inoculation
with R. solanacearum. To address these hypotheses I used a histological approach.
For this work I used four tomato varieties shown in Figure 3.1. Hawaii
HA7996 (HA) and CRA-66 (T1) are the two resistant cultivars and West Virginia
WVa700 (WV) and Okitsu (T2) are the two susceptible cultivars used in this study.
Mock-inoculated (MI) and inoculated (I) plant varieties were sampled at 144 hpi.
Figure 3.1A-D shows a side-by-side comparison depicting tomato phenotypes of R.
solanacearum inoculated varieties and mock-inoculated varieties at 144 hpi. The
resistant variety HA-I and T1-I do not show wilting at 144 hpi while the susceptible
WV-I and T2-I show wilting of the aerial portion of the plant.

27
My results, described in detail below, show that bacteria colonize the root
vascular cylinder of resistant HA variety differently and at a later time point than the
susceptible WV. I also found that the resistant HA variety had significantly larger
tracheary elements than that of the susceptible varieties. Finally, I observed that the
HA variety does alter its root architecture at 144 hours after inoculation with R.
solanacearum.
3.2 Root Anatomy
3.2.1 Resistant HA variety of tomato have larger xylem tracheary elements
I hypothesized that resistant varieties would have larger tracheary elements
than susceptible varieties to aid in resistance to a bacterial invasion. Larger tracheary
elements may allow water transport to the aerial portions of the plant thus reducing
wilting incidence even in the presence of bacteria.
Measurement of the average area of tracheary elements in HA-MI (Fig. 3.2A),
WV-MI (Fig. 3.2B), T1-MI (Fig. 3.3A) and T2-MI (Fig. 3.3B) was completed using
the ImageJ software (see materials and methods). Statistical analysis shows
significant differences in the area of tracheary elements between varieties (Fig. 3.4).
The resistant HA-MI variety had larger xylem tracheary elements than the susceptible
varieties WV-MI and T2-MI at 144 hpi. Tracheary elements of the resistant T1-MI
were not significantly different than that of HA-MI, WV-MI or T2-MI. The variation
in the tracheary elements for T1-MI could be due to fewer replicates in this variety.
Additional analysis with more replicates may reduce the standard error and show a
clearer difference between the other varieties.

28
3.2.2 Bacteria are distributed differently within the vascular cylinder in resistant and susceptible plants
Another hypothesis for my research was that plant resistance could be a result
of differences in the distribution of bacteria within the vascular cylinder. If R.
solanacearum is localized to specific areas within the root tissue this could minimize
the transfer of bacteria to other regions of the plant thus reducing wilt incidence. To
test this, I used scanning electron microscopy to view distribution of bacterial
colonization within the root with paraffin fixed tissue samples at 24, 72 and 144 hpi.
HA-I
At 24 hpi, bacteria are not observed within the tracheary elements of the
vascular cylinder, but are found on the surface of the root (Figure 3.5). Figure 3.5A
shows a root cross-section of HA-I. The red box indicates the magnified view of
tracheary elements within the vascular cylinder shown in Fig. 3.5B. Figure 3.5 C is a
magnified image of the yellow box seen in figure 3.5A and shows the root external
surface with bacteria present (Fig. 3.5C). The red arrow is pointing to an example of
bacteria. These data show that at 24 hpi R. solanacearum does not colonize the
vascular cylinder of the root but is present on the external surface.
At 72 hpi, I observed masses of bacteria present in several tracheary elements
(Fig. 3.6A) of HA-I. Closer magnification (the red box shown in Fig 3.6B) shows
bacteria colonization in detail of several tracheary elements (Fig. 3.6B). A magnified
view of the yellow box seen in 3.6A shows a large xylem vessel with bacteria lining
the cell wall with many not directly attached to it (Fig. 3.6C).

29
At 144 hpi, the tracheary elements of HA are colonized with bacteria.
However, their distribution is restricted to a smaller area of the vascular cylinder (Fig.
3.7A). Figure 3.7B shows a magnified view of red box in 3.7A. This magnified image
reveals colonies of R. solanacearum in the metaxylem located between the two
protoxylem poles. The bacteria are forming a mass within the tracheary elements but
are mostly not adhering to the cell walls (Fig. 3.7B).
These data show that R. solanacearum colonizes the vascular cylinder by 72
hpi and that colonization increases by 144 hpi.
WV-I
As shown in Figure 3.8, R. solanacearum colonization is observed in the
vascular cylinder of WV-I at 24 hpi. The WV-I root cross section in Fig. 3.8A shows
a red box magnified in Fig. 3.8B. The magnified view shows unattached R.
solanacearum colonizing the tracheary elements. Figure 3C is a magnified view of
the highlighted yellow box from Figure 3.8A. This shows R. solanacearum
colonizing the external root surface and cortical cells (Fig. 3.8C).
At 72 hpi, multiple tracheary elements within the vascular cylinder of the
WV-I root are lined with bacteria attached to the cell walls (Fig. 3.9A). The red box
from 3.9A shows bacteria directly attached to the cell wall of a neighboring xylem
parenchyma cell (Fig. 3.9B). Magnified view of yellow box from Figure 3.9A shows
a tracheary element with bacteria lining the cell walls (Fig. 3.9C). Several of the
bacteria appear to have undergone binary fission as indicated by the red arrow.

30
At 144 hpi, large amounts of bacteria are observed in the root cortex cells of
WV-I (Fig. 3.10A). In Figure 3.10B, the red box from 3.10A is magnified and large
masses of bacteria can be observed within the cells of the root cortex (Fig. 3.10B). A
magnified view of the yellow box from 3.10A shows bacteria lining a tracheary
element next to pit membranes (Fig. 3.10C).
Together these data show that R. solanacearum colonizes the vascular
cylinder of the susceptible variety WV-I at 24 hpi, but at this same time point in the
HA resistant variety, bacteria are only found on the external surface of the root. At 72
hpi, R. solanacearum colonizes tracheary elements of both the HA resistant and
susceptible variety. At 144 hpi, restriction of bacterial colonization is observed in the
root vascular cylinder of the HA resistant variety compared to the WV susceptible
variety. These data suggest that resistance is due both to a delay in bacterial
colonization of the root vascular cylinder and a restriction of the vascular cylinder
area colonized by R. solanacearum.
3.2.3 Bacteria colonize the root vascular cylinder of resistant plants at 48 hpi
R. solanacearum was not found in the resistant HA-I vascular cylinder at 24
hpi but was by 72 hpi. To have a better understanding of when the bacteria entered
the root vascular cylinder, I used light microscopy to capture bacterial invasion into
the vascular cylinder at five time-points (0, 24, 48, 72, 144 hpi) after infection in HA-
I and WV-I.
First, confirmation was needed to be certain that the mass of rod shaped
organism observed with the light microscope were in fact R. solanacearum
colonization (Fig. 3.11A-C). I confirmed R. solanacearum colonization within the

31
root by taking R. solanacearum K60 from pure culture and streaking it onto a glass
slide followed by staining with 0.05% toluidine blue. A glass coverslip was applied
and then the edges were sealed with clear nail polish to restrict any spread of the
bacteria. The slide was then viewed in a light microscope at 100x magnification (Fig.
3.11D). Rod shaped bacterial bodies were seen similar to those in figures 3.11A-C.
Paraffin fixed R. solanacearum were viewed in the SEM attached to the walls of
xylem tracheary elements (Fig. 3.11E) or released from paraffin embedded tissue
(Fig. 3.11F). Together, these data confirmed that the masses observed in root sections
using light microscopy were R. solanacearum colonies.
Figure 3.12 shows a time course of 0-144 hpi using light microscopy. These
light microscope data confirm that bacteria colonize vascular cylinder more slowly in
the HA resistant variety. The red star in Figure 3.12 marks locations where bacteria
were found within the root at 0, 24, 48, 72 and 144 hpi. HA-I (Fig. 3.12A) and WV-I
(Fig. 3.12B) at 0 hpi are free of bacteria colonization in the xylem. As seen with
SEM, bacteria are not present in the vascular cylinder of HA-I resistant variety at 24
hpi (Fig. 3.12C) but are present in the larger tracheary elements of the WV-I at 24 hpi
(Fig. 3.12D). The light microscopy shows that bacteria are beginning to line the walls
of the tracheary elements in HA-I at 48 hpi (Fig. 3.12E) whereas in WV-I several
large masses of bacteria are already seen within the tracheary elements at this time
point (Fig. 3.12F). Consistent with the SEM results at 72 hpi, both HA-I and WV-I
show an increase in bacteria numbers within the vascular cylinder (Fig. 3.12G)
compared to 48 hpi. WV-I increase in the number of tracheary elements that are
colonized (Fig. 3.12H) to include large and smaller tracheary elements. At 144 hpi in

32
HA-I, the central portion of the xylem has blocked tracheary elements mainly in the
metaxylem between the protoxylem poles (Fig. 3.12I). WV-I has tracheary elements
containing bacteria as well as the cortical cells (fig. 3.12J).
These data show that R. solanacearum enters the vascular cylinder of the
resistant HA variety between 24 and 48 hpi.
3.2.4 Additional resistant and susceptible lines were examined to test whether the pattern of colonization was specific to the resistant variety HA
To examine whether the pattern of colonization in the resistant variety HA
was similar in other resistant varieties, I used light microscopy to examine bacterial
colonization in the resistant CRA-66 (T1) and susceptible Okitsu (T2) at 0, 72 and
144 hpi (Fig. 3.13). No colonization was seen at 0 hpi for either resistant T1-I (Fig.
3.13A) or susceptible T2-I (Fig. 3.13D). At 72 hpi, the area of the vascular cylinder
colonized by R. solanacearum appeared larger in T1-I roots (Fig. 3.13B) while the
susceptible T2-I had a smaller area colonized (Fig. 3.13E). At 144 hpi, the resistant
T1-I showed colonization of the tracheary elements in the vascular cylinder along
with colonization of the cortical cells (Fig. 3.13C) while colonization in the vascular
cylinder of susceptible T2-I had increased in area (Fig. 3.13F). These data show that
colonization of the resistant variety T1-I appears to be different than HA-I. Neither
T1 nor T2 were investigated with SEM.
3.2.5 Distribution of R. solanacearum in the stem differs between resistant HA-I and susceptible WV-I
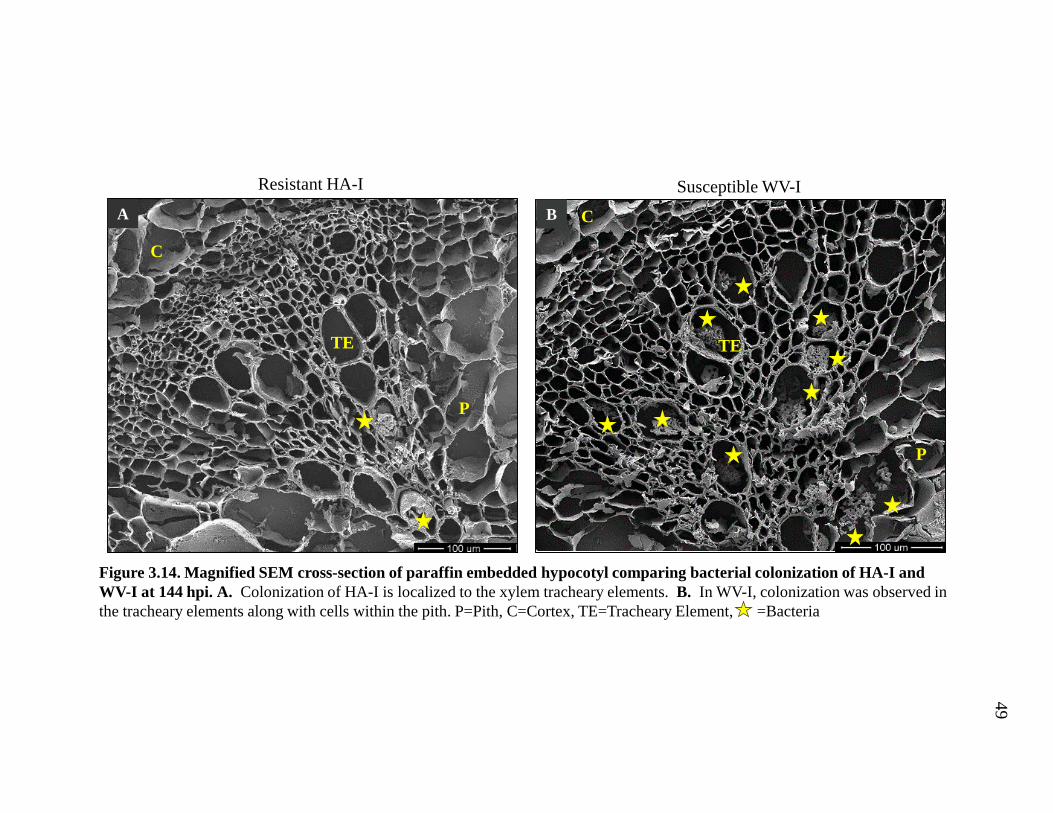
We examined whether the restricted distribution of R. solanacearum observed
in the vascular cylinder of tomato roots was also found in the stem of the resistant

33
variety HA. The pattern of bacterial colonization was observed in the hypocotyl of
HA-I and WV-I at 144 hpi. The distribution was similar in both the SEM and with
light microscopy. In HA-I, R. solanacearum colonization was restricted to the larger
xylem tracheary elements (Fig. 3.14A, 3.15A). In contrast, bacteria colonization in
WV-I was observed in the xylem tracheary elements as well as the parenchyma cells
in the pith (Fig. 3.14B, 3.15B). These data show that, like in the resistant root
vascular cylinder, bacterial colonization is restricted in the stem of the resistant
variety.
3.3 Root Architecture
3.3.1 Root architecture of resistant HA-I differs from that of other varieties
Emerged lateral roots were counted under a dissecting scope from the root-
shoot junction to 2.4 cm below (Fig. 3.16). The percent change of emerged lateral
roots was calculated by comparison of inoculated to mock-inoculated (Fig. 3.17).
Emerged lateral roots increased nearly 29.67% in inoculated resistant HA compared
to mock-inoculated HA roots. The other three varieties, resistant T1, and susceptible
WV and T2 all had a suppression of emerged lateral roots after inoculation. These
data suggest that changes in root architecture may be important for resistance in HA.

34
3.4 Results Summary
In summary, we found that the resistant HA-MI variety of tomato has larger
xylem tracheary elements. Not only did we observe that the distribution of bacteria
differs in resistant HA-I compared to the susceptible WV-I variety but bacterial
colonization is delayed in HA-I varieties. In the resistant HA-I variety, colonization is
restricted to fewer tracheary elements and surrounding parenchyma cells, while the
susceptible WV-I variety has more colonization of cortical tissues. Furthermore we
saw that the distribution of bacteria differs in hypocotyls of resistant HA compared to
that of the susceptible WV hypocotyl. Finally, resistant HA increases its lateral root
growth approximately 30% six days after inoculation with R. solanacearum.
Together, these data suggest that resistance in HA is likely due to a combination of
factors, including delayed and restricted bacterial colonization in the root vascular
cylinder, and an increase in lateral root emergence after inoculation.

Figure 3.1. Bacterial wilt disease tomato phenotypes at 144 hours post inoculation (hpi). Mock-inoculated (A-B) and inoculated (C-D). Wilting was observed in the susceptible varieties (C-D). T1 displays some stunting but there is no wilting is occurring.
A
WV-MI HA-MI
C
WV-I HA-I
Inoc
ulat
edM
ock-
Inoc
ulat
ed
T1-MIT2-MI
B
D
T1-IT2-I
Susceptible Resistant Susceptible Resistant
35

Figure 3.2. Comparison of primary root anatomy of HA-MI and WV-MI at 144 hpi. The scale bar is equivalent to 100 μm. C=Cortex, TE=Tracheary Element
Susc
eptib
le W
V-M
IR
esis
tant
HA
-MI
B
C
TE
A
TE
C
36

Figure 3.3. Comparison of primary root T1-MI and T2-MI at 144 hpi. The scale bar is equivalent to 100 μm. C=Cortex, TE=Tracheary Element
Susc
eptib
le T
2-M
IR
esis
tant
T1-
MI
B C
TE
A
C
TE
37

0
400
800
1200
1600
2000
HA T1 T2 WV
Aver
age
Are
a of
Lar
gest
5 X
ylem
Tr
ache
ary
Elem
ents
(µm
2 ) in
Roo
t
Comparison of Xylem Tracheary ElementsArea Between Varieties at 144 hpi
A
AB
BB
Figure 3.4. Comparison of mock-inoculated root tracheary element average area between tomato varieties at 144 hpi. The area of five largest xylem tracheary elements was measured using imageJ (see materials and methods). One-way ANOVA showed that variety was significant at p < 0.05, and differences between varieties were determined by Tukey’s test at a p<0.05. Varieties with the same letter above them are not significant at p<0.05. Statistical analysis was performed on log transformed vessel size values in order to meet homogeneity of variance assumption for ANOVA. Statistical analysis was done by Elizabeth French in Dr. Iyer-Pascuzzi’s lab.
Level -Level Difference Std Err Dif Lower CL Upper CL p-ValueHA T2 0.838351 0.253417 0.15056 1.526144 0.0121 ++++++++HA WV 0.697258 0.216115 0.11071 1.283809 0.0149 +++++++T1 T2 0.674844 0.253417 -0.01295 1.362636 0.0560 ++++++T1 WV 0.533750 0.216115 -0.05280 1.120301 0.0849 +++++
HA T1 0.163508 0.226663 -0.45167 0.778688 0.8878 ++WV T2 0.141094 0.244028 -0.52122 0.803404 0.9379 +
38

Figure 3.5. SEM cross-sections of HA-I at 24hpi. A. Root cross-section of HA-I at 24 hpi. B. Magnified view of red square from 3.5A showing tracheary elements in the vascular cylinder of HA at 24 hpi. No bacteria were observed in the tracheary elements or xylem parenchyma cells. C. Magnified view of yellow box from 3.5A showing bacteria (red arrow) present on external surface of the root. C=Cortex, TE=Tracheary Element, S=Root External Surface
A B
TE
C
S
C
39

Figure 3.6. SEM cross-sections of HA-I at 72 hpi. A. Root cross-section of HA-I at 72 hpi. B. Magnified view of red box seen in 3.6A showing bacteria masses in the smaller tracheary elements but mostly lacking attachment to the cell wall. C. Magnified view of yellow box from 3.6A shows a large tracheary element with bacteria along the wall but not attached to it. Arrow=Bacteria, TE=Tracheary Element
A B
TE
C
TE
40

Figure 3.7. SEM cross-sections of HA-I at 144 hpi. A. Root cross-section of HA-I at 144 hpi. B. Magnified view of red box seen in 3.7A showing colonies of R. solanacearum in the metaxylem located between the protoxylem poles. The bacteria are forming a mass within the tracheary elements but are not adhering to the cell walls. C=Cortex, TE=Tracheary Element, Arrow=Bacteria
B
TE
A
C
TE
41

Figure 3.8. SEM cross-sections of WV-I at 24 hpi. A. Root cross-section of WV-I at 24 hpi. B. Magnified view of red box seen in 3.8A showing mostly unattached R. solanacearum populating tracheary elements. C. Magnified view of yellow box from 3.8A showing R. solanacearum colonizing the external root surface and cortical cells. C=Cortex, TE=Tracheary Element, S=Root External Surface, Arrow=Bacteria
B C
C
S
A
TE
42

Figure 3.9. SEM cross-sections of WV-I at 72 hpi. A. Root cross-section of WV-I at 72 hpi. B. Magnified view of red box seen in 3.9A showing bacteria mostly attached to the walls of the tracheary elements. C. Magnified view of yellow box from 3.9A showing tracheary element obstructed with bacteria.. C=Cortex, TE=Tracheary Element, Arrow=Bacteria
BTE
A
C
TE
C
43

Figure 3.10. SEM cross-sections of WV-I at 144 hpi. A. Overview of WV-I root with large amounts of bacteria in cortex cells. B. Magnified view of red box seen in 3.10A showing a large mass of bacterial cells within a cell. C. Magnified view of yellow box from 3.10A showing unattached bacteria lining a tracheary element next to pit membranes. C=Cortex, TE=Tracheary Element, Arrow=Bacteria
BA
TE
C
C
44

Figure 3.11. Confirmation of R. solanacearum colonization within the root. A-C. R. solanacearum colonizes a xylem tracheary element of inoculated HA-I at 144 hours post inoculation (hpi), 100x magnification. D. R. solanacearum K60 isolated from a petri dish and streaked onto a glass slide followed by staining with 0.05% toluidine blue. The rod shaped bacterial bodies were seen at 100x magnification. E. Scanning electron microscope (SEM) images of inoculated WV-I at 72 hpi show bacteria colonizing xylem vessels. F. R. solanacearum released from deparaffinizedsample viewed under SEM. Mock-inoculated root sample not shown.
A B
C D
E F
45

Susceptible WV-IResistant HA-I
0 hp
i24
hpi
72 h
pi48
hpi
144
hpi
A B
C D
E F
G H
I J
Figure 3.12. Comparison of bacterial colonization of HA-I and WV-I at 0, 24. 48, 72 and 144 hpi with light microscopy.
46

Figure 3.12. Comparison of bacterial colonization of HA-I and WV-I at 0, 24, 48, 72 and 144 hpi with light microscopy. Light microscopy was used to investigate if bacteria colonized the HA-I root at a slower rate than that of WV-I. This was done by marking locations with red star in locations bacteria were found within the root at 0, 24, 48, 72 and 144 hpi. A-B. HA-I (A) and WV-I (B) at 0 hpi are free of bacteria colonization in the xylem tissue. C. Bacteria are not present in the vascular cylinder of HA-I resistant variety at 24 hpi. D. Bacteria are present in the larger tracheary elements of the WV-I at 24 hpi. E. Bacteria are beginning to line the walls of the tracheary elements in HA-I at 48 hpi. F. In WV-I at 48 hpi, several large masses of bacteria are seen within the tracheary elements. G. At 72 hpi, HA-I shows an increase in bacteria numbers within the vascular cylinder. H. WV-I increase in the number of tracheary elements that are colonized to include large and smaller tracheary elements. I. At 144 hpi in HA-I, the central portion of the xylem has blocked tracheary elements mainly in the metaxylem between the protoxylem poles. J. At 144 hpi, WV-I has tracheary elements containing bacteria as well as the cortical cells. C=Cortex, TE=Tracheary Element,
=Bacteria
47

Res
ista
nt T
1-I
0 hpiA
C
TE
144 hpiC
72 hpiB
Susc
eptib
le T
2-I
D
TE
CFE
Figure 3.13. Comparison of bacterial colonization of T1-I and T2-I at 0, 72 and 144 hpi with light microscopy. A. T1-I at 0 hpi is free of bacteria colonization in the xylem. B. At 72 hpi, a large number of tracheary elements in T1-I roots are colonized with bacteria. C. At 144 hpi, the resistant T1-I had high colonization of the tracheary elements in the vascular cylinder along with colonization of the cortical cells. D. T2-I at 0 hpi is free of bacteria colonization in the xylem. E. The susceptible T2-I at 72 hpi has a relatively small amount of tracheary elements colonized. F. The susceptible T2-I has increased the number of tracheary elements colonized. C=Cortex, TE=Tracheary Element, =Bacteria 48
Resistant HA-I
A
TE
P
C
Susceptible WV-IB C
TE
P
Figure 3.14. Magnified SEM cross-section of paraffin embedded hypocotyl comparing bacterial colonization of HA-I and WV-I at 144 hpi. A. Colonization of HA-I is localized to the xylem tracheary elements. B. In WV-I, colonization was observed in the tracheary elements along with cells within the pith. P=Pith, C=Cortex, TE=Tracheary Element, =Bacteria
49

Figure 3.15. Cross-section of paraffin embedded hypocotyl comparing anatomical structure of HA-I and WV-I at 144 hpi with light micrographs. A. Bacterial colonization of the HA-I hypocotyl is localized to the xylem tracheary elements. B. WV-I bacteria colonization is seen in the tracheary elements as well as the parenchyma cells within the pith. P=Pith, C=Cortex, TE=Tracheary Element, =Bacteria
Resistant HA-IA
C
P
TE
Susceptible WV-I
B
P
C
TE
50

Figure 3.16. Comparison of lateral root emergence of mock-inoculated and inoculated resistant and susceptible varieties at 144 hpi. Lateral roots were counted under a dissecting scope from the root shoot junction to 2.4 cm. A-B. At 144 hpi, HA-I (B) emergence of lateral roots increased in number compared to HA-MI (A). C-H. The remainder of the varieties did not show this increase in lateral root emergence after inoculation with R. solanacearum.
InoculatedMock-InoculatedA B
C D
E F
G H
Res
ista
nt H
ASu
scep
tible
WV
Susc
eptib
le T
2R
esis
tant
T1
51

Figure 3.17. Percent change in lateral root emergence of Inoculated/Mock-Inoculated at 144 hpi by variety. Number of emerged lateral roots on the uppermost 2.4 cm of primary root of resistant or susceptible plants 144 hours after soil inoculation with R. solanacearum or mock-inoculated with water. The percent change of emerged lateral roots was calculated by the ratio of inoculated to non-inoculated plants. Inoculated HA (resistant) plants had an increase of 29.67% in emerged lateral roots compared to mock-inoculated. The other three varieties show a suppression of lateral roots after inoculation.
-50%
-40%
-30%
-20%
-10%
0%
10%
20%
30%
40%
50%
HA T1 T2 WV29.67% -18.00% -33.67% -21.00%
Perc
ent c
hang
e of
late
ral r
oot e
mer
genc
e of
In
ocul
ated
/Moc
k-In
ocul
ated
at 1
44 h
pi
52

53
CHAPTER 4. DISCUSSION
4.1 Introduction
This work investigated the role of the root vascular cylinder in tomato
resistance to R. solanacearum. Specifically, the objectives of this thesis were to test
four hypotheses: first, that there are differences in the timing of colonization of the
vascular cylinder in resistant plants, second that bacteria are distributed differently
within the vascular cylinder of resistant plants, third that resistant plants have larger
xylem vessels, and fourth that resistant and susceptible plants undergo different
changes to root architecture after infection with R. solanacearum.
We addressed the first three hypotheses using scanning electron (SEM) and
light microscopy. We found that bacterial colonization of the vascular cylinder is
delayed in inoculated resistant HA lines. The distribution of bacteria also differs in
resistant HA compared to the susceptible WV variety. Colonization is restricted in the
resistant HA variety to fewer tracheary elements and surrounding parenchyma cells
while the susceptible WV variety have more colonization of cortical tissues. SEM
showed that parenchyma cells in the susceptible WV variety collapse faster than in
the resistant HA variety. The distribution of bacteria also differs in hypocotyls of
resistant HA compared to that of the susceptible WV hypocotyl. Our work also
showed that the resistant variety HA of tomato has larger xylem tracheary elements.

54
To address the last hypothesis, we examined lateral root structure using a
stereomicroscope. We found that the production of lateral roots increases
approximately 30% in resistant HA tomato plants six days after inoculation with R.
solanacearum compared to the mock-inoculated control.
4.2 Colonization
4.2.1 Delay in bacteria colonization of the HA variety
Light and scanning electron microscopy showed the progression of bacterial
colonization of resistant and susceptible plant varieties. At 24 hpi, bacterial
colonization of susceptible WV had increased enough to become visible in light
micrographs. SEM showed a small amount of colonization of the WV root vascular
cylinder and cortex, while heavy accumulation was observed on the external surface.
Here, the external surface is either epidermal cells or cortex that is exposed after
epidermal shedding that occurs as the roots grow through the rhizosphere. In HA at
24 hpi, a small amount of bacteria accumulate on the external surface but areas of
high colonization are not observed within the vascular cylinder or cortex. These
results show that there is a delay in the bacterial colonization of the root xylem tissues
in the resistant plant.
In another study of R. solanacearum colonization of tomato, Vasse et al.
found that at 24 hpi, bacterial colonization mainly occurs on the surface of the
susceptible root at the extremities and axils of secondary roots (Vasse, 1995). After
48-72 hpi plants had colonization in the cortex, and colonization of the vascular
cylinder was observed after 72 hpi. This group did not observe colonization in a

55
resistant root. The differences between their study and the work here in the timing of
colonization of the root vascular cylinder could be due to several factors. These
include using a different susceptible cultivar and bacterial strain, as well as
differences in the growth medium for tomato plants and the method of inoculation.
Additionally, because this group only used light microscopy, they may have been
unable to observe the low level of vascular cylinder colonization that we observed at
24 hpi in susceptible WV.
We examined additional resistant (T1) and susceptible (T2) tomato varieties to
test if the patterns of colonization we observed in resistant HA and susceptible WV
were variety specific. Surprisingly, bacterial colonization of resistant T1 plants as less
restricted than in the resistant variety HA or the susceptible varieties WV or T2. This
suggests resistant tomatoes have different root defense mechanisms that promote
resistance to R. solanacearum.
4.2.2 Distribution of bacteria within the root
Our work showed that the distribution of bacteria in the resistant HA root is
different than that in the susceptible variety WV. In the resistant HA variety
colonization was restricted to a smaller area in the root vascular cylinder while the
susceptible WV variety had more colonization of cortical tissues. Colonization of
tissues in WV increased at 72 hpi to include multiple tracheary elements in vascular
cylinder and cortical cells. At 72 hpi, bacteria in HA were observed colonizing
smaller protoxylem, and xylem parenchyma cells are obstructed.
In WV, the bacteria are attached to the cell walls of larger and small xylem
tissue and obstruction appeared more dense in WV’s tracheary elements than in the

56
HA variety at 72 hpi. In HA, bacteria appeared to line the cell walls of large xylem
vessels without attachment while neighboring cells can either be devoid of or
obscured by bacteria.
Localization of bacteria within the root central vascular cylinder allows for its
vertical transfer but restricts its horizontal transfer. Vertical movement of bacteria
through the axial system is characteristic of primary xylem and the secondary xylem
moves in both axial and ray systems (Esau, 1960). Nakaho (2000) observed that in
susceptible Ponderosa plants, bacteria traveled vertically (axial) and horizontally (ray
system) within the vascular bundle of the hypocotyl, possibly through degenerated pit
openings (Nakaho, Hibino, & Miyagawa, 2000).
Similar to our work, in roots of a susceptible Arabidopsis accession, R.
solanacearum was observed within intercellular spaces of the root cortex at 6 dpi
(Digonnet et al., 2012), and colonization of the vascular cylinder was observed soon
after that. Interestingly, Diggonet et al., used transmission electron microscopy to
show that R. solanacearum often colonized the periplasm in Arabidopsis – the region
between the cell wall and plasma membrane. Whether this occurs in tomato is not
known.

57
4.2.3 Susceptible WV parenchyma cells collapse
Secretions of cell wall degrading enzymes are an important virulence factor
for R. solanacearum (Allen et al., 1997). Consistent with this, we observed that xylem
parenchyma cells collapse in the susceptible WV variety but not in the resistant HA
variety at the same time-point. Other groups have also observed cell wall degradation.
For example, in Arabidopsis, R. solanacearum degrades the cell walls, allowing for
further penetration of R. solanacearum (Digonnet et al., 2012) into the root.
Cell wall degrading enzymes are secreted by the T2SS secretion system in R.
solanacearum (Genin & Denny, 2012). The T2SS, together with the T3SS secretion
system that transfers the bacteria’s proteins or effectors from the bacteria’s cytoplasm
to the host cell cytoplasm, enables browning, necrosis and collapse of the stem.
4.3 Architecture
4.3.1 Root architecture of resistant HA-I differs from that of other varieties
Restriction of R. solanacearum movement in HA reduces injection of cell wall
degrading enzymes into the ray system, leaving the structural integrity of the stem
intact. We observed a 30% increase in lateral root formation near the root-shoot
junction of R. solanacearum inoculated HA plants at 144 hpi compared to mock
inoculated HA plants. In both the susceptible varieties, WVa and T2, as well as a
second resistant variety, T1, lateral root production was suppressed. T2 and HA have
resistance that was introgressed from different wild species (Lebeau et al., 2011).
These data suggest that an increase in lateral root production is a part of the resistance
mechanism used by HA.

58
Currently, it is not clear how the increase in lateral root production may
promote resistance in HA. However, since the primary root becomes necrotic and
eventually dies, we speculate that the rapid onset of lateral roots offers the plant an
architectural mechanism to circumvent diseased primary root tissue, rerouting water
and oxygen to the plant.
Whether the increase in lateral root emergence is a direct or indirect effect of
R. solanacearum is not clear. It is possible that effectors from the T3SS could
manipulate the lateral root machinery to suppress lateral root emergence in the
susceptible line. Alternatively, hormones produced by R. solanacearum could alter
lateral root emergence. Auxin and ethylene both regulated lateral root development,
and both are produced by R. solanacearum. Susceptible and resistant tobacco plants
were found to have large increases of IAA when infected with Pseudomonas
solanacearum (Sequeira & Kelman, 1962), and plants infected with R. solanacearum
show symptoms when disruption of the internal auxin pool cause a system imbalance.
An increase in IAA levels can result in epinasty of the leaf, adventitious root
formation, stunting, internal structural changes such as tyloses, vessel collapse and
parenchyma proliferation (Buddenhagen & Kelman, 1964).
However, the increase in lateral root emergence could also be due to changes
in the primary root and thus indirectly to the pathogen. For example, in Arabidopsis,
damage to the primary root can promote the production of adventitious roots at the
root-shoot junction (Lucas et al., 2011).

59
Another possible mechanism for the increase in lateral root formation is an
internal hypoxic environment. Experiments with flooding of tomato showed that
tomato plants can adapt to this abiotic stress in a similar way (McNamara & Mitchell,
1990). Damaging root conditions can develop in response to hypoxic environments.
At the root-shoot junction of the stem, unrestricted access to oxygen allows rapid
adventitious root formation. Dawood et al. (2014) found that root system Solanum
dulcamara declines rapidly upon flooding and adventitious root primordia are
activated within 24 hours to begin to replace the former roots and were visible by eye
between the second and fourth day after flooding (Dawood et al., 2014). During
infection with R. solanacearum, the oxygen levels inside the root xylem decrease
(Dalsing, Truchon, Gonzalez-Orta, Milling, & Allen, 2015). This hypoxic
environment may lead to stimulation of root branching.
4.4 Area of Vessels
4.4.1 Resistant HA variety of tomato have larger xylem tracheary elements
Our hypothesis for this research was that anatomical differences between
resistant and susceptible lines would differ and thus affect the susceptibility of that
plant to the plant pathogen R. solanacearum. Calculating the average area the largest
5 tracheary elements from we found that the size of the resistant HA and T1 was
statistically different than that of the susceptible WV and T2. Visual data has
confirmed that the root of HA is larger than that of WV. Larger root tracheary
elements may increase the area in which bacteria are able to occupy. Bacteria would
be dispersed in a much larger area and not able to aggregate to one another.

60
Although these data suggest that resistant plants have larger xylem vessels,
additional work is needed with more resistant varieties to test whether there is a clear
relationship between vessel size and resistance.
4.5 Conclusions
Together, our data reveal that root-mediated resistance in the HA variety both
delays R. solanacearum colonization of the tomato vascular cylinder and restricts it to
fewer vessels. Additionally, the HA variety prevents colonization of the cortex.
Intriguingly, HA, which is highly resistant to many different R. solanacearum strains,
alters its root architecture after infection to increase lateral root production. How this
occurs, and what stage of lateral root development is affected will be the subject of
future work.

8
LITERATURE CITED

61
LITERATURE CITED
Allen, C., Gay, J., & Simon-Buela, L. (1997). A regulatory locus, pehSR, controls polygalacturonase production and other virulence functions in Ralstonia solanacearum. Mol Plant Microbe Interact, 10, 1054-1064. doi: 10.1094/MPMI.1997.10.9.1054
Angot, A., Peeters, N., Lechner, E., Vailleau, F., Baud, C., Gentzbittel, L., Genin, S. (2006). Ralstonia solanacearum requires F-box-like domain-containing type III effectors to promote disease on several host plants. Proc Natl Acad Sci U S A, 103, 14620-14625. doi: 0509393103 [pii] 10.1073/pnas.0509393103
Álvarez, B., Biosca, E. G., & López, M. M. (2010). On the life of Ralstonia solanacearum, a destructive bacterial plant pathogen. Current research, technology and education topics in applied microbiology and microbial biotechnology. Volume, 1.
Bellini, C., Pacurar, D. I., & Perrone, I. (2014). Adventitious roots and lateral roots: similarities and differences. Annual Review of Plant Biology, 65, 639-666.
Buddenhagen, I., & Kelman, A. (1964). Biological and physiological aspects of bacterial wilt caused by Pseudomonas solanacearum. Annual Review of phytopathology, 2(1), 203-230.
Carmeille, A., Caranta, C., Dintinger, J., Prior, P., Luisetti, J., & Besse, P. (2006). Identification of QTLs for Ralstonia solanacearum race 3-phylotype II resistance in tomato. Theor Appl Genet, 113, 110-121. doi: 10.1007/s00122-006-0277-3
Coll, N. S., & Valls, M. (2013). Current knowledge on the Ralstonia solanacearum type III secretion system. Microb Biotechnol, 6, 614-620. doi: 10.1111/1751-7915.12056 10.1111/1751-7915.12056. Epub 2013 Apr 26.
Dalsing, B. L., Truchon, A. N., Gonzalez-Orta, E. T., Milling, A. S., & Allen, C. (2015). Ralstonia solanacearum uses inorganic nitrogen metabolism for virulence, ATP production, and detoxification in the oxygen-limited host xylem environment. MBio, 6, e02471. doi: 10.1128/mBio.02471-14 10.1128/mBio.02471-14.

62
Danesh, D., Aarons, S., McGill, G. E., & Young, N. D. (1994). Genetic Dissection of Oligogenic Resistance to Bacterial Wilt in Tomato. Mol Plant Microbe Interact, 7, 464-471.
Dawood, T., Rieu, I., Wolters-Arts, M., Derksen, E. B., Mariani, C., & Visser, E. J. (2014). Rapid flooding-induced adventitious root development from preformed primordia in Solanum dulcamara. AoB plants, 6, plt058.
Denny, T. (2006). Plant pathogenic Ralstonia species Plant-associated bacteria (pp. 573-644): Springer.
Denny, T. P. (2000). Ralstonia solanacearum--a plant pathogen in touch with its host. Trends Microbiol, 8, 486-489.
Denny, T. P., & Baek, S. (1991). Genetic evidence that extracellular polysaccharide is a viurlence factor of Pseudomonas solanacearum. Mol Plant Microbe Interactions, 4, 198-206.
aDigonnet, C., Martinez, Y., Denance, N., Chasseray, M., Dabos, P., Ranocha, P., . . . Goffner, D. (2012). Deciphering the route of Ralstonia solanacearum colonization in Arabidopsis thaliana roots during a compatible interaction: focus at the plant cell wall. Planta, 236, 1419-1431. doi: 10.1007/s00425-012-1694-y 10.1007/s00425-012-1694-y. Epub 2012 Jun 24.
bDigonnet, C., Martinez, Y., Denancé, N., Chasseray, M., Dabos, P., Ranocha, P., . . . Goffner, D. (2012). Deciphering the route of Ralstonia solanacearum colonization in Arabidopsis thaliana roots during a compatible interaction: focus at the plant cell wall. Planta, 236(5), 1419-1431.
Esau, K. (1960). Anatomy of Seed Plants. New York: Wiley.
Fegan, M., & Prior, P. (2005). How complex is the Ralstonia solanacearum species complex? In P. P. a. A. C. H. C. Allen (Ed.), Bacterial Wilt: The Disease and the Ralstonia solanacearum Species Complex (pp. 449-461). St. Paul, MN, USA: American Phytopathological Society Press.
Fitter, A., & Atkinson, D. (1991). The ecological significance of root system architecture: an economic approach. Plant root growth: an ecological perspective., 229-243.
Genin, S., & Denny, T. P. (2012). Pathogenomics of the Ralstonia solanacearum species complex. Annual review of phytopathology, 50, 67-89.
Grimault, V., Anais, G., & Prior, P. (1994). Distribution of Pseudomonas solanacearum in the stem tissues of tomato plants with different levels of resistance to bacterial wilt. Plant Pathology, 43(4), 663-668.

63
Grimault, V., & Prior, P. (1993). Bacterial wilt resistance in tomato associated with tolerance of vascular tissues to Pseudomonas solanacearum. Plant Pathology, 42(4), 589-594.
Haas, D., & Defago, G. (2005). Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat Rev Microbiol, 3, 307-319. doi: 10.1038/nrmicro1129
Hayward, A. (1991). Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum. Annual review of phytopathology, 29(1), 65-87.
Huang, Q., & Allen, C. (1997). An exo-poly-alpha-D-galacturonosidase, PehB, is required for wild-type virulence of Ralstonia solanacearum. J Bacteriol, 179, 7369-7378.
Huet, G. (2014). Breeding for resistances to Ralstonia solanacearum. Front Plant Sci, 5, 715. doi: 10.3389/fpls.2014.00715 10.3389/fpls.2014.00715. eCollection 2014.
Hölttä, T., & Nikinmaa, E. (2013). Modelling the effect of xylem and phloem transport on leaf gas exchange. Paper presented at the IX International Workshop on Sap Flow 991.
Ishihara, T., Mitsuhara, I., Takahashi, H., & Nakaho, K. (2012). Transcriptome analysis of quantitative resistance-specific response upon Ralstonia solanacearum infection in tomato. PLoS One, 7, e46763. doi: 10.1371/journal.pone.0046763 PONE-D-12-04108 [pii] 10.1371/journal.pone.0046763. Epub 2012 Oct 5.
Jackson, M. B. (1985). Ethylene and responses of plants to soil waterlogging and submergence. Annual Review of Plant Physiology, 36(1), 145-174.
Jacobs, J. M., Milling, A., Mitra, R. M., Hogan, C. S., Ailloud, F., Prior, P., & Allen, C. (2013). Ralstonia solanacearum requires PopS, an ancient AvrE-family effector, for virulence and To overcome salicylic acid-mediated defenses during tomato pathogenesis. MBio, 4, e00875-00813. doi: 10.1128/mBio.00875-13 10.1128/mBio.00875-13.
Jensen, W. A. (1962). Botanical histochemistry.
Jung, J. K. H., & McCouch, S. (2013). Getting to the roots of it: genetic and hormonal control of root architecture. Frontiers in Plant Science, 4. doi: 10.3389/fpls.2013.00186
Lebeau, A., Daunay, M. C., Frary, A., Palloix, A., Wang, J. F., Dintinger, J., . . . Prior, P. (2011). Bacterial wilt resistance in tomato, pepper, and eggplant: genetic resources respond to diverse strains in the Ralstonia solanacearum species complex. Phytopathology, 101, 154-165. doi: 10.1094/PHYTO-02-10-0048 10.1094/PHYTO-02-10-0048.
Liu, H., Zhang, S., Schell, M. A., & Denny, T. P. (2005). Pyramiding unmarked deletions in Ralstonia solanacearum shows that secreted proteins in addition to plant cell-wall-degrading enzymes contribute to virulence. Mol Plant Microbe Interact, 18, 1296-1305. doi: 10.1094/MPMI-18-1296

64
Lucas, M., Swarup, R., Paponov, I. A., Swarup, K., Casimiro, I., Lake, D., . . . Whitworth, M. (2011). Short-Root regulates primary, lateral, and adventitious root development in Arabidopsis. Plant Physiology, 155(1), 384-398.
Malamy, J. E., & Benfey, P. N. (1997). Down and out in Arabidopsis: the formation of lateral roots. Trends in plant science, 2(10), 390-396.
Mansfield, J., Genin, S., Magori, S., Citovsky, V., Sriariyanum, M., Ronald, P., . . . Foster, G. D. (2012). Top 10 plant pathogenic bacteria in molecular plant pathology. Mol Plant Pathol, 13, 614-629. doi: 10.1111/j.1364-3703.2012.00804.x 10.1111/j.1364-3703.2012.00804.x. Epub 2012 Jun 5.
McAvoy, T., Freeman, J. H., Rideout, S. L., Olson, S. M., & Paret, P. L. (2012). Evaluation of Grafting Using Hybrid Rootstocks for Managment of Bacterial Wilt in Field Tomato Production. HortScience, 47, 621-625.
McGarvey, J., Denny, T., & Schell, M. (1999). Spatial-temporal and quantitative analysis of growth and EPS I production by Ralstonia solanacearum in resistant and susceptible tomato cultivars. Phytopathology, 89(12), 1233-1239.
McNamara, S. T., & Mitchell, C. A. (1990). Adaptive stem and adventitious root responses of two tomato genotypes to flooding. HortScience, 25(1), 100-103.
Milling, A., Babujee, L., & Allen, C. (2011). Ralstonia solanacearum extracellular polysaccharide is a specific elicitor of defense responses in wilt-resistant tomato plants. PLoS One, 6, e15853. doi: 10.1371/journal.pone.0015853 10.1371/journal.pone.0015853.
Monteiro, F., Genin, S., van Dijk, I., & Valls, M. (2012). A luminescent reporter evidences active expression of Ralstonia solanacearum type III secretion system genes throughout plant infection. Microbiology, 158, 2107-2116. doi: 10.1099/mic.0.058610-0 10.1099/mic.0.058610-0. Epub 2012 May 18.
Nakaho, K., & Allen, C. (2009). A Pectinase-deficient Ralstonia solanacearum Strain Induces Reduced and Delayed Structural Defences in Tomato Xylem. J. Phytopathol, 157, 228-234.
Nakaho, K., Hibino, H., & Miyagawa, H. (2000). Possible mechanisms limiting movement of Ralstonia solanacearum in resistant tomato tissues. Journal of Phytopathology, 148(3), 181-190.
Nakaho, K., Inoue, H., Takayama, T., & Miyagawa, H. (2004). Distribution and multiplication of Ralstonia solanacearum in tomato plants iwth resistance derived from different origins. Journal of General Plant Pathology, 70, 115-119.
Pieterse, C. M., Leon-Reyes, A., Van der Ent, S., & Van Wees, S. C. (2009). Networking by small-molecule hormones in plant immunity. Nature chemical biology, 5(5), 308-316.

65
Plener, L., Manfredi, P., Valls, M., & Genin, S. (2010). PrhG, a transcriptional regulator responding to growth conditions, is involved in the control of the type III secretion system regulon in Ralstonia solanacearum. J Bacteriol, 192, 1011-1019. doi: 10.1128/JB.01189-09 10.1128/JB.01189-09. Epub 2009 Dec 11.
Prior, P., Ailloud, F., Dalsing, B. L., Remenant, B., Sanchez, B., & Allen, C. (2016). Genomic and proteomic evidence supporting the division of the plant pathogen Ralstonia solanacearum into three species. BMC Genomics, 17, 90. doi: 10.1186/s12864-016-2413-z 10.1186/s12864-016-2413-z.
Remenant, B., Babujee, L., Lajus, A., Medigue, C., Prior, P., & Allen, C. (2012). Sequencing of K60, type strain of the major plant pathogen Ralstonia solanacearum. J Bacteriol, 194, 2742-2743. doi: 194/10/2742 [pii] 10.1128/JB.00249-12 10.1128/JB.00249-12.
Remigi, P., Anisimova, M., Guidot, A., Genin, S., & Peeters, N. (2011). Functional diversification of the GALA type III effector family contributes to Ralstonia solanacearum adaptation on different plant hosts. New Phytol, 192, 976-987. doi: 10.1111/j.1469-8137.2011.03854.x 10.1111/j.1469-8137.2011.03854.x. Epub 2011 Sep8.
Rivard, C. L., & Louws, F. J. (2008). Grafting to manage soilborne diseases in heirloom tomato production. HortScience, 43(7), 2104-2111.
Rivard, C. L., O'Connell, S., Peet, M. M., Welker, R. M., & Louws, F. J. (2012). Grafting Tomato to Manage Bacterial Wilt Caused by Ralstonia solanacearum in the Southeastern United States. Plant Disease, 96, 973-978. doi: Doi 10.1094/Pdis-12-10-0877
Rivard, C. L., O’Connell, S., Peet, M. M., & Louws, F. J. (2008). Grafting as a viable tool to manage major tomato diseases in the southeastern USA. Paper presented at the Proc.
Ruzin, S. E. (1999). Plant microtechnique and microscopy (Vol. 198): Oxford University Press New York.
Saile, E., McGarvey, J. A., Schell, M. A., & Denny, T. P. (1997). Role of extracellular polysaccharide and endoglucanase in root invasion and colonization of tomato plants by Ralstonia solanacearum. Phytopathology, 87(12), 1264-1271.
Schell, M. A. (2000). Control of Virulence and Pathogenicity Genes of Ralstonia Solanacearum by an Elaborate Sensory Network. Annu Rev Phytopathol, 38, 263-292. doi: 10.1146/annurev.phyto.38.1.263
Scott, J. W., Wang, J. F., & Hanson, P. M. (2005). Breeding tomatoes for resistance to bacterial wilt, a global view. Proceedings of the 1st International Symposium on Tomato Diseases, 161-172.

66
Sequeira, L., & Kelman, A. (1962). Acccumulation of Growth Substacnces in Plants Infected by Pseudomonas solanacearum. Phytopathology, 52(5), 439-&.
Sole, M., Popa, C., Mith, O., Sohn, K. H., Jones, J. D., Deslandes, L., & Valls, M. (2012). The awr gene family encodes a novel class of Ralstonia solanacearum type III effectors displaying virulence and avirulence activities. Mol Plant Microbe Interact, 25, 941-953. doi: 10.1094/MPMI-12-11-0321 10.1094/MPMI-12-11-0321.
Thoquet, P., Olivier, J., Sperisen, C., Rogowsky, P., Laterrot, H., & Grimsley, N. (1996). Quantitative trait loci determining resistance to bacterial wilt in tomato cultivar Hawaii7996. Molecular Plant-Microbe Interactions, 9, 826-836. doi: Doi 10.1094/Mpmi-9-0826
Thoquet, P., Olivier, J., Sperisen, C., Rogowsky, P., Prior, P., Anais, G., . . . Grimsley, N. (1996). Polygenic resistance of tomato plants to bacterial wilt in the French West Indies. Molecular Plant-Microbe Interactions, 9, 837-842. doi: Doi 10.1094/Mpmi-9-0837
Turner, M., Jauneau, A., Genin, S., Tavella, M.-J., Vailleau, F., Gentzbittel, L., & Jardinaud, M.-F. (2009). Dissection of bacterial wilt on Medicago truncatula revealed two type III secretion system effectors acting on root infection process and disease development. Plant physiology, 150(4), 1713-1722.
Valls, M., Genin, S., & Boucher, C. (2006). Integrated regulation of the type III secretion system and other virulence determinants in Ralstonia solanacearum. PLoS Pathog, 2(8), e82.
Vasse, J. (1995). Microscopic studies of intercellular infection and protoxylem invasion of tomato roots by Pseudomonas solanacearum. Mol Plant-Microbe Interact, 8, 241-251.
Vasse, J., Genin, S., Frey, P., Boucher, C., & Brito, B. (2000). The hrpB and hrpG regulatory genes of Ralstonia solanacearum are required for different stages of the tomato root infection process. Mol Plant Microbe Interact, 13, 259-267. doi: 10.1094/MPMI.2000.13.3.259
Wang, J.-F., Ho, F.-I., Truong, H. T. H., Hugan, S.-M., Balatero, C. H., Dittapongpitch, V., & Hidayati, N. (2013). Identification of major QTLs associated with stable resitance of tomato cultivar 'Hawaii 7996' to Ralstonia solanacearum. Euphytica, 190, 241-252.
Wang, J. F., Olivier, J., Thoquet, P., Mangin, B., Sauviac, L., & Grimsley, N. H. (2000). Resistance of tomato line Hawaii7996 to Ralstonia solanacearum Pss4 in Taiwan is controlled mainly by a major strain-specific locus. Molecular Plant-Microbe Interactions, 13, 6-13. doi: Doi 10.1094/Mpmi.2000.13.1.6
Webb, M. A., & Arnott, H. J. (1983). Inside plant crystals: a study of the noncrystalline core in druses of Vitis vinifera endosperm. Scanning Electron Microscopy, 1759-1770.

67
Young, N. D., & Danesh, D. (1994). Understanding Bacterial Wilt Resistance in Tomato through the Use of DNA Genetic-Markers. Bacterial Wilt, 145-155.
Yuliar, N. Y. A., & Toyota, K. (2015). Recent trends in control methods for bacterial wilt diseases caused by Ralstonia solanacearum. Microbes Environ, 30, 1-11. doi: 10.1264/jsme2.ME14144 10.1264/jsme2.ME14144. Epub 2015 Mar 26.
Zhang, Y., Chen, L., Yoshimochi, T., Kiba, A., Hikichi, Y., & Ohnishi, K. (2013). Functional analysis of Ralstonia solanacearum PrhG regulating the hrp regulon in host plants. Microbiology, 159, 1695-1704. doi: 10.1099/mic.0.067819-0 10.1099/mic.0.067819-0. Epub 2013 May 23.
Zolobowska, L., & Van Gijsegem, F. (2006). Induction of lateral root structure formation on petunia roots: a novel effect of GMI1000 Ralstonia solanacearum infection impaired in Hrp mutants. Molecular plant-microbe interactions, 19(6), 597-606.

APPENDICES

TE
C
TE
CE F
C D
24 h
pi72
hpi
144
hpi
Resistant HA-I Susceptible WV-I
Appendix A. Delay in colonization seen between HA-I and WV-I in SEM at 24, 72 and 144 hpi.
A B
TE
C
TE
TE
TE
CC
C
68

69
Appendix A. Delay in colonization seen between HA-I and WV-I in SEM at 24, 72 and 144 hpi. A. At 24 hpi, bacterial colonization is not seen in the vascular cylinder of HA-I. B. Bacterial colonization of 24 hpi WV-I is observed in the vascular cylinder, cortex and on the external surface of the root. C. At 72 hpi in HA-I, bacterial colonization can be observed in the xylem tissue within the vascular cylinder. The largest tracheary elements are free of large obstructions caused by R. solanacearum while smaller tracheary elements now are obstructed. D. WV-I has bacterial colonization seen in the vascular cylinder and cortical tissues at 72 hpi. Large obstructions can be seen in larger tracheary elements. E. At 144 hpi, HA-I’s larger tracheary elements are still unobstructed but the smaller tracheary elements are clogged with R. solanacearum. F. Large pockets of bacteria occupy the parenchyma cells in the cortex of WV-I at 144 hpi. C=Cortex, TE=Tracheary Elements, =Bacteria

Appendix B. Comparison of primary root anatomical changes during development of HA-MI and WV-MI at 0, 72 and 144 hpi. Due to size differences in the vascular cylinder area of resistant and susceptible, different magnifications were needed to allow for visual clarity in measurement of tracheary elements. The scale bar is equivalent to 100 μm. C=Cortex, TE=Tracheary Element
Susceptible WV-MIResistant HA-MI
0 hp
i72
hpi
144
hpi
A B
C D
E FC
TE
TE
C
70

Appendix C. Comparison of primary root anatomical changes during development of T1-MI and T2-MI at 0, 72 and 144 hpi. Due to size differences in the vascular cylinder area of resistant and susceptible, different magnifications were needed to allow for visual clarity in measurement of tracheary elements. The scale bar is equivalent to 100 μm. C=Cortex, TE=Tracheary Element
Susceptible T2-MIResistant T1-MI
0 hp
i72
hpi
144
hpi
A B
C D
E F
C
C
TETE
71

Appendix D. SEM cross-section of paraffin embedded hypocotyl comparing anatomical structure of HA-MI and WV-MI at 72 hpi. A. Cross-section of hypocotyl of HA-MI at 72 hpi. B. Hypocotyl of WV-MI at 72 hpi. Differences in vasculature structure of the hypocotyl were not seen. P=Pith, C=Cortex, TE=Tracheary Element
Resistant HA-MI
A
TEP
Susceptible WV-MI
B
TE
P
C
72

Appendix E. Cross-section of paraffin embedded hypocotyl comparing anatomical structure of HA-MI and WV-MI at 144 hpi with light micrographs. A. Hypocotyl of HA-MI at 144 hpi. B. Hypocotyl of WV-MI at 144 hpi. Differences in vasculature structure of the hypocotyl were not seen. P=Pith, C=Cortex, TE=Tracheary Element
Resistant HA-MI
P
TE
C
ASusceptible WV-MI
P TE
C
B
73

13
VITA

74
VITA
Denise Lynn Caldwell
EDUCATION
2013–Present MS., Botany & Plant Pathology, Purdue University
• Master’s Degree expected in August 2016• Advisor: Dr. Anjali Iyer-Pascuzzi, Purdue University, Botany & Plant Pathology• GPA: 3.20
2012 B.A., Horticulture Science, Purdue University
RESEARCH EXPERIENCE
2015-Present Research Assistant, Department of Botany & Plant Pathology, Purdue University. Mentor: Dr. Anjali Iyer-Pascuzzi: Investigating the role of root anatomy and root architecture in resistance to Ralstonia solanacearum using paraffin processing and histological techniques.
2013-2015 Research Assistant, Department of Botany & Plant Pathology, Purdue University. Mentor: Dr. Mary Alice Webb: Investigated calcium oxalate crystals in Aptenia cordifolia.
2013 Research Assistant, Department of Botany & Plant Pathology, Purdue University. Mentor: Dr. Nicolas Carpita: Investigated Flax (Linum usitatissimum) seed coat development in regards to mucilage development and expression.
2013 Lab Technician, Department of Botany & Plant Pathology, Purdue University. Mentor: Dr. Catherine Aime. Mycology media preparation and bacterial and fungal culture transfer.

75
2012 Research & Discovery Intern, Dow AgroSciences LLC. Indianapolis, IN. Mentor: Dr. Yang: Evaluated performance of light emitting diode (LED) as sole/primary light source I growing soybean seedlings and seedlings of other crops such as wheat and weed species in a conviron growth chamber.
2010-2012 Research Lab Assistant, Department of Horticulture, Purdue University. Mentor: Dr. Steve Weller: Managed heirloom tomato competition trial comparing the growth and disease resistance of 13 popular heirloom tomato varieties. Researched breaking seed dormancy in Ambrosia trifida via stratification, scarification, hormones, light and temperature. Assisted with studies on Ambrosia trifida including experiments on competition, propagation, herbicide resistance, and DNS comparisons of resistant and susceptible varieties.
AWARDS & HONORS
2016 Nominated for Excellence in Teaching Award 2015 Teaching Academy Graduate Teaching Award 2014 Botany & Plant Pathology Travel Award
LEADERSHIP, TEACHING & MENTORING
Teaching Assistantships 2016 Spring BTNY 110, Introduction to Plant Science, Lab Leader 2015 Fall BTNY 110, Introduction to Plant Science, Lab Leader 2015 Summer BTNY 110, Introduction to Plant Science, Lab Leader 2015 Spring BTNY 110, Introduction to Plant Science, Lab Leader 2014 Fall BTNY 316, Plant Anatomy 2014 Spring BTNY 110, Introduction to Plant Science, Lab Leader 2013 Fall BTNY 316, Plant Anatomy
Guest Lecture 2016 Spring BTNY 420, Plant Cellular and Developmental Biology
Outreach 2016 High School Purdue Agribusiness & Science Academy (PASA) sponsored by
the Office of Multicultural Programs within the College of Agriculture 2016 Agriculture workshop student leader to rural elementary schools 2015 Agriculture workshop student leader to rural elementary schools
USA Biology Olympiad 2014 Plant Anatomy, Teaching Assistant 2013 Plant Anatomy, Teaching Assistant

76
Mentorship Mina Rostamza Post-Doctoral Student Histological Techniques Seval Taskin Graduate Student Histological Techniques Kristina Gans Graduate Student Histological Techniques Mateo Serna International Student Histological Techniques Alyssa Delong Lab Technician Plant Anatomy Techniques Andrew Katz Undergraduate Student Plant Anatomy Techniques Jorge Diaz Graduate Student Plant Anatomy Techniques Kim Bong-Suk Lab Technician Histological Techniques Frank Surez Graduate Student Microscopy Techniques Khalil Jahed Graduate Student Microscopy Techniques Dr. Jeff Stuart Professor Microscopy Imaging
MICROSCOPY, COMPUTER & LABORATORY EXPERTISE
Microscopes • Transmission Electron Microscopy (TEM), Scanning Electron Microscopes
(SEM), CryoSEM, Electron Dispersive X-ray analysis (EDX), Light, Stereo, Polarized, Dark-field and Fluorescent
Microscopy Support • Cressington turbo-pumped sputter coater, Pelco Oven for microwave fixation,
Critical point drying, resin and paraffin fixation and embedding, plant tissue clearing, free-hand sectioning, microtomy, ultramicrotomy
Computer • Mac OS, Windows and MS Office suite, Statistical packages: JMP, Imaging
software: Spot, imageJ, Adobe Photoshop, Adobe Illustrator, Adobe InDesign
Laboratory • RNA extraction, tissue culture, sterile technique, northern blot, plant nucleic acid
purification, protoplast isolation, electrophoresis, osmolality, media preparation, bacterial and fungal culture, spectroradiometery, microscopy
Field • Weed and plant identification, herbicide application

77
PUBLICATIONS • Kim, B. S., French, E., Caldwell, D., Harrington, E. J., & Iyer-Pascuzzi, A. S.
(2016). Bacterial wilt disease: Host resistance and pathogen virulencemechanisms. Physiological and Molecular Plant Pathology, 95, 37-43.
MANUSCRIPTS IN PREPARATION
• Caldwell D., Iyer-Pascuzzi A (2016). The role of root anatomy and rootarchitecture in resistance to Ralstonia solanacearum.
• Kim, B.S., French, E., Caldwell, D., Iyer-Pascuzzi A. (2016). Transcriptionalresponse of tomato roots to R. solanacearum infection.
PROFFESSIONAL MEETINGS AND CONGERENCES ATTENDED
2014 Gordon Research Conference on Biomineralization (GRS) 2013 M&M Microscopy and Microanalysis
REFERENCES
Dr. Anjali Iyer-Pascuzzi Department of Botany and Plant Pathology 915 West State St., West Lafayette, IN 47907 Email: [email protected]
Dr. Peter Goldsbrough Department of Botany and Plant Pathology 915 West State St., West Lafayette, IN 47907 Email: [email protected]
Dr. Jeffrey Stuart Department of Entomology 901 West State St., West Lafayette, IN 47907 Email: [email protected]

PUBLICATION

Bacterial wilt disease: Host resistance and pathogen virulencemechanisms*
Bong-Suk Kim, Elizabeth French, Denise Caldwell, Emily J. Harrington,Anjali S. Iyer-Pascuzzi*
Department of Botany and Plant Pathology, Purdue University, West Lafayette, IN 47906, USA
a r t i c l e i n f o
Article history:Received 15 December 2015Received in revised form23 February 2016Accepted 29 February 2016Available online 2 March 2016
Keywords:Plant disease resistanceTomatoQuantitative disease resistanceBacterial pathogen
a b s t r a c t
Bacterial wilt (BW), caused by Ralstonia solanacearum, is one of the most destructive bacterial diseases ofSolanaceous species worldwide. The species infects plants in more than 200 species and 50 families andwas ranked second in a list of the top 10 most scientifically and economically important bacterial plantpathogens [1]. The molecular mechanisms underlying resistance and the functions of R. solanacearumeffectors are beginning to be uncovered, and much remains to be discovered. In this mini-review, weprovide a summary of host resistance and R. solanacearum virulence mechanisms, with a focus ontomato.
© 2016 Elsevier Ltd. All rights reserved.
1. Introduction
Ralstonia solanacearum infects over 200 plant speciesthroughout the world, but is particularly devastating in the Sol-anaceae [1,2,3]. The pathogen is soil-borne and thrives in hot, hu-mid environments. In Central America and the Southeastern UnitedStates, R. solanacearum is a severe limiting factor to tomato pro-duction, where it regularly reduces crop yields. A newly emergedstrain, race 3, biovar 2 is also a severe threat in more temperateregions [4,5].
Bacteria enter the tomato root either in the elongation zone orthrough wounds created at the site of lateral root emergence [6].Once in the root, bacteria enter the vasculature, which allows themtomove from the root to the shoot. R. solanacearum produce severalpathogenicity factors, including type III effectors, exopolysaccharide(EPS) [7e11] and cell wall degrading enzymes such as cellulase andpectic lyase [9,12e15]. As bacteriamultiply in thexylem, the amountof EPS increases, ultimately preventing water flow to the shoot, andresulting in shoot wilting and eventual plant death.
Bacterial wilt disease management is difficult, and the bestmeans of control has historically been through resistant cultivars.Resistance in tomato is primarily quantitative, the result of manygenes contributing a small amount to resistance [16e23]. Althoughseveral QTL for resistance to R. solanacearum have been coarse-mapped [16e23], none have been fine-mapped or cloned.
R. solanacearum is an extremely diverse species. Older classifi-cation schemes divided the pathogen into races and biovars [2]. Therace designation was based on the host range of the pathogen.Within races, R. solanacearum was divided into biovars, based inpart on the ability of the pathogen to degrade specific carbohy-drates [24]. However, as the diversity of the species became clearthrough phylogenetic analyses, the race-biovar designations werechanged [25,26]. Currently, R. solanacearum is classified accordingto phylotype and sequevar and is known as the R. solanacearumspecies complex (RSSC) [25]. The RSSC is divided into four phylo-types, which each correspond (approximately) to a differentgeographic region. Phylotype I is found in Asia, Phylotype II (A andB) in the Americas, Phylotype III in Africa, and Phylotype IV in theAustraliaeIndonesia region. Phylotypes subsequently are dividedinto sequevars. Strains in phylotype IIA are found in the south-eastern United States and the Caribbean, and typically infect plantsin the Solanaceae. Strains in Phylotype IIB include the select agentstrain of R. solanacearum (old designation race 3 biovar 2) that caninfect plants at lower temperatures than other strains [4,5].
* This article is part of a special issue entitled “The U.S.-Japan Scientific Seminar:Molecular Contact Points in Host-Pathogen Co-evolution”.* Corresponding author.
E-mail addresses: [email protected] (B.-S. Kim), [email protected](E. French), [email protected] (D. Caldwell), [email protected](E.J. Harrington), [email protected] (A.S. Iyer-Pascuzzi).
Contents lists available at ScienceDirect
Physiological and Molecular Plant Pathology
journal homepage: www.elsevier .com/locate/pmpp
http://dx.doi.org/10.1016/j.pmpp.2016.02.0070885-5765/© 2016 Elsevier Ltd. All rights reserved.
Physiological and Molecular Plant Pathology 95 (2016) 37e43

Phylogenetic analyses have shown little correlation between hostrange and phylotypes [10,26].
The genomes of at least 14 different RSSC strains have beencompletely sequenced [27]. These 14 include representatives fromall four RSSC phylotypes, including the phylotype II type strain K60[28], and the phylotype I strain GMI1000 frequently used in labo-ratory studies [29]. Sequencing has been instrumental not only inclassification but also in the identification of virulencedeterminants.
In this mini-review, we provide an overview of host geneticresistance and bacterial virulence mechanisms. For more detailedreviews see Refs. [10,26,30,31].
2. Host resistance mechanisms
2.1. Genetic mechanisms of resistance
BW resistance in tomato is largely quantitative (polygenic).Although in Arabidopsis, resistance to R. solanacearum is governedby at least one major gene, RRS1-R [32], no major genes for BWresistance have been identified in tomato. Several QTL for BWresistance have been mapped to regions of the tomato genome[16e23]. These are both strain-specific and broad-spectrum QTLand include a large QTL on chromosome 12 to a Taiwanese phylo-type I strain [21,23], weaker QTL on chromosome 6 to both phylo-type I and II strains [17e23], and additional small possibly strain-specific QTLs on chromosomes 3, 4 and 8 [22]. Most of these QTLwere identified using the same recombinant inbred line (RIL)population, Hawaii7996 (HA) x WestVirginia700 (WVa700). HA isunusual in that it has strong resistance in a wide variety ofgeographic locations throughout the world [3]. The published QTLdata suggest that HA contains multiple loci contributing to resis-tance to several R. solanacearum strains.
Thus far, the strongest QTL for BW resistance to any R. sol-anacearum strain maps to a 2.8 cM region on chromosome 12. Bwr-12 is effective against phylotype I (Asian) strains, accounting forapproximately 18e56% of the resistance [21,23]. This QTL does notappear to be active against phylotype II strains [22].
The other major QTL for BW resistance, Bwr-6, localizes to a15.5 cM region on the short arm of tomato chromosome 6 and iseffective against both phylotype I and II strains [17e23]. QTL in thisregion have mapped to slightly different areas of this region in fourseparate experiments (all in the HA � Wva700 RILs). It is unclearexactly how many QTL are in this region. Bwr-6 explains approxi-mately 11.5e22% of the variation in resistance to R. solanacearum[23]. This region appears to be a ‘hot-spot’ for both qualitative andquantitative disease resistance genes. Resistance genes that map tothis region include Mi for resistance to root-knot nematodes andpotato aphids [33e35], Cf2 and Cf5 for resistance to Cladosporiumfulvum [36], and the QTL Ol-1 for resistance to Oidium lycopersicon[37], Ty-1 for resistance to Tomato yellow leaf curl virus [38], and Am,for resistance to Alfalfa mosaic virus [39].
In Arabidopsis, resistance to R. solanacearum is conferred by thecombined action of two NB-LRR genes, RRS1-R (resistance to Ral-stonia solanacearum 1) and RPS4 (resistance to Pseudomonassyringae 4) [40e46]. Together these act to confer recognition of thetype III effectors (T3Es) PopP2 from R. solanacearum and AvrRps4from Pseudomonas syringae. RRS1-R is unusual in that it not onlyencodes the only known recessive NB-LRR R gene but the proteinalso contains a WRKY DNA binding domain [32]. The T3E PopP2, anacetyltransferase, directly acetylates a specific lysine within the C-terminal WRKY transcription factor domain of RRS1-R [47]. Thisdisrupts the connection between RRS1-R and DNA, leading to theactivation of RPS4-induced defenses [45,46]. Thus, RRS1-R acts asan ‘integrated decoy’: it uses theWRKY domain as an effector target
but can activate defense signaling through a different NBS-LRRprotein. Thus, it is able to prevent effector virulence and activatedefense genes [45,46,48].
2.2. Host whole genome transcription profiling
Given the importance of R. solanacearum, surprisingly little in-formation is known regarding the transcriptional responses ofplants to R. solanacearum infection. A time-course of transcriptionalchanges viewed bymicroarray analysis of R. solanacearum infectionin Arabidopsis found that many hormone-related genes wereaffected [49]. At early wilting stages (5 days after inoculation), 471genes were differentially expressed in the compatible interactioncompared to the incompatible. Most (353) were upregulated, and40% of these genes were involved in the biosynthesis or signaling ofthe plant hormone abscisic acid (ABA). This is consistent with theidea that during early wilting stages bacteria-produced EPS re-stricts water flow throughout the plant, and ABA plays a major rolein water stress. Many down-regulated genes at the early wiltingstage were related to auxin and cytokinin signaling [49].
Because tomato has a different type of resistance to R. sol-anacearum compared to Arabidopsis (poly-vs. monogenic) andpathogenesis of the bacteria within each host appears to bedifferent [49], work directly in tomato is crucial for understandingresistance and disease mechanisms. Examination of tomato tran-scriptional responses in a resistant and susceptible line withmicroarrays 24 h after inoculation showed no change in geneexpression in the inoculated susceptible plant relative to the mock-inoculated susceptible plant, but identified 146 up-regulated and10 down-regulated genes in the resistant plant [50]. Many differenthormone-related genes were differentially expressed in the resis-tant cultivar, including those with roles in the jasmonic acid, auxin,and ethylene signaling pathways. PR (pathogenesis-related) genesas well as lignin and hydroxyl cinnamic acid biosynthesis geneswere also upregulated, suggesting that gene expression changesleading to alterations in cell wall structure within xylem vesselsmay be important for resistance [50]. In both the transcriptionalprofiling reports, shoots were sampled for gene expression, butsince roots are part of the natural infection process, and are animportant part of resistance [51,52], it is likely that knowledge ofroot transcriptional reprogramming will provide important infor-mation for deciphering resistance mechanisms.
2.3. Roots are an important part of resistance
Studies with grafting have demonstrated that roots are key toresistance. Grafting resistant rootstocks to susceptible scions is aneffective means of disease control in the Southern United States aswell as South Korea and Japan [51,52]. Scions of the susceptible BHN602 tomato line were grafted onto rootstocks of various tomatolines with resistance to bacterial wilt [51]. When disease pressurewas low, BW incidence of both grafted and non- or self-grafted waslow. When BW disease pressure was high, both fruit yield and BWincidence from grafted hybrids performed much better than non-grafted and self-grafted hybrids [51]. Grafting resistant rootstockshad similar effects on resistance in susceptible scions in a fieldstudy in North Carolina [52]. These studies demonstrate theimportance of roots for resistance and tomato yield in the field.
2.4. Insights to resistance from histological studies
Several groups have used histological studies to provide insightinto how plants are affected by R. solanacearum. R. solanacearum isthought to enter the root system through secondary roots, root capor through wounds acquired in the soil [2]. In susceptible tomatoes,
B.-S. Kim et al. / Physiological and Molecular Plant Pathology 95 (2016) 37e43

R. solanacearum entry into roots occurs in four stages [6]: Bacteriafirst colonize the root surface either in the elongation zone or at thepoint of emerging lateral roots. During the second stage of coloni-zation, 2e3 days after inoculation, significant numbers of bacteriainfect the cortex, an inner layer of the root, with smaller numbersfound in the epidermis, an outer root layer. Bacteria enter thevasculature, including the xylem, and begin to degrade cell wallsduring the third stage of infection, about 3 days after inoculation. Inthe fourth stage, bacteria multiply and begin to move into theshoot. The increase in EPS in the root and shoot plugs the xylem,preventing water movement, and leading to wilt, the most recog-nizable disease symptom.
The infection process is less well understood in resistant plants.Multiple reports have shown that resistant cultivars have signifi-cant numbers of bacteriawithin both roots and shoots, although themultiplication of bacteria in resistant plants may be slower [9], andin general, susceptible plants have higher levels of bacteria in thestem. Several reports suggest that bacteria in resistant plants aredistributed differently than in susceptible plants, and are limited intheir ability to broadly colonize tissues [53]. Grimault et al. [53,54]showed that resistance was significantly correlated with bacterialcolonization in the mid-stem. Consistent with this, QTL controllingR. solanacearum race 3 colonization in tomato stems were recentlyidentified [22]. There appear to be two types of resistance mecha-nisms in the stem that both block bacterial movement, but indifferent ways. Examination of bacteria distribution in stems ofresistant and susceptible lines found that resistant lines confinedbacteria to the primary xylem, possibly through thickenings of pitmembranes in stem xylem tissue [55]. In the susceptible line bac-teria were found in secondary xylem tissues of the stem or hadmoved into the pith [55]. A second means of preventing bacterialmovement may be through the accumulation of tyloses in stemxylem vessels [53]. Tyloses are outgrowths of xylem parenchymacells that balloon into the lumen of adjacent xylemvessels, creatinga physical barrier that may prevent bacterial movement.
Experiments inMedicago truncatula showed that as in tomato, R.solanacearum enters the intercellular spaces of the root cortex andproceeds to the vascular cylinder to allow colonization of thevascular system [56]. Bacteria were identified in both resistant andsusceptible cultivars but with higher numbers in susceptible plants.As in tomato, once bacteria accessed the central vascular systemthey were able to rapidly reproduce within the xylem vessel.
The infection process appears to be somewhat similar in Ara-bidopsis thaliana roots grown on agar media. Digonnet et al. [57]found that six days post inoculation bacteria are found withinintercellular spaces of the root cortex. Cell wall degradationallowed the bacteria to enter into the vascular system andmultiply,and bacterial pockets to form within xylem vessels [57] of sus-ceptible plants.
3. Environmental mechanisms of resistance
3.1. Biocontrol
In recent years, many studies have investigated the use ofbeneficial microorganisms as biocontrol agents (BCAs) for plantdiseases. The most commonly studied microorganisms for thepurpose of controlling BW are bacteria [58]. Most researchers havefocused on avirulent strains of R. solanacearum, Pseudomonas spp,and Bacillus spp., although other species have been used as well.BCAs can suppress disease through multiple mechanisms includingcompetition, production of cell-wall degrading enzymes, antibi-otics, or siderophores, and induction of systemic resistance [58]. Acommon strategy for identifying potential BCAs is to isolate themfrom the rhizospheres of healthy plants [59]. However, isolating
microorganisms from the rhizospheres of R. solanacearum-infectedplants to enrich for bacteria with biocontrol capabilities against BWmay be a more effective strategy [60].
Hyakumachi et al. [61] showed that soil pretreatment witheither a cell culture or cell-free filtrate of Bacillus thuringiensissuppresses tomato bacterial wilt and induces expression ofdefense-related genes. In a follow-up study, they examined tran-scription patterns in tomato roots pretreated with B. thuringiensiscell-free filtrate and subsequently challenged with R. solanacearum[62]. Salicylic acid (SA)-related defense genes were induced only inmain roots, but not in lateral roots, while jasmonic acid (JA)-dependent defense genes were either suppressed or not induced.Ethylene (ET)-responsive genes were induced in both main andlateral roots. Taken together, these results suggest that B. thur-ingiensismediated disease suppression involves co-activation of SAand ET defense hormone signaling pathways and simultaneoussuppression of the JA pathway, which is different than the classicmodel of induced systemic resistance mediated by activation of JAand ET [63]. In an early study on biocontrol of BW, pretreatmentwith the oomycete Pythium oligandrum transiently induced ETproduction and expression of ET-related genes and subsequentlyreduced wilt symptoms [64].
Various rhizosphere-inhabiting fungi have also been found toinduce resistance against tomato bacterial wilt. Many studies havefocused on mycorrhizal fungi, which develop a symbiotic rela-tionship with plant roots and have also been shown in some casesto induce systemic resistance against plant disease. An early studyfound that colonization by Glomus versiformae induced phenolproduction in tomato roots and protected them against R. sol-anacearum infection [65]. A more recent investigation character-ized three different endomycorrhizal fungi for their biocontrolcapabilities and found that bacterial wilt disease suppression variedbetween fungal species [66].
3.2. Soil amendments
Several different types of soil amendments have also beenshown to provide efficient control for tomato bacterial wilt. To-matoes grown in soil amended with 20% (v/v) municipal wastebiochar, a charcoal-like substance for soil application in agriculture,had a reduced disease incidence compared to tomatoes grown inun-amended soil or soil amended with a wood biochar [67].Addition of bioorganic fertilizer decreased soil R. solanacearumpopulations and reduced disease incidence in tomato field plotscompared to plots treated with either an inorganic or organic NPKfertilizer [68]. The bioorganic fertilizer, which contains its ownpopulations of bacteria, increased the diversity and richness of thebacterial populations in the amended soil, which was correlatedwith increased disease resistance. Soil application of systemicresistance elicitors - silicon and chitosan - also reduced bacterialwilt disease incidence and severity and increased expression ofdefense-related genes, such as chitinases and peroxidases [69].Another study examining silicon-induced resistance to tomatobacterial wilt showed that silicon amendment primed tomatoplants for JA-/ET- and reactive oxygen species-dependent defensegene expression, increasing their resistance to disease [70]. For amore comprehensive review of biocontrol and soil amendmentdisease control for bacterial wilt, see Yuliar et al., [58].
4. Pathogen effectors and other virulence factors
4.1. Non-type III effector virulence factors
R. solanacearum uses numerous virulence factors during thedisease process. The pathogen secretes exopolysaccharide (EPS1), a
B.-S. Kim et al. / Physiological and Molecular Plant Pathology 95 (2016) 37e43

polymer of N-acetylated sugars and a major virulence factor forinfection in tomato [10]. EPS1 deficient strains cause significantlyless disease in tomato regardless of whether they are inoculatedonto unwounded root systems or directly into the xylem [9,11]. Inresistant tomato plants, wild type R. solanacearum induced greaterdefense responses than the eps- mutant, suggesting that resistantplants can recognize this pathogen trait [11]. Coincident with this,R. solanacearum growing in roots of infected resistant tomato plantsproduce less EPS per gram of tissue than those found in susceptibleplants [8]. The production of EPS1 is regulated by cell density, and ishigher when pathogen density is greater than 107 cells/ml [71]. Inaddition to high cell density, EPS1 production requires nitrogen inthe form of nitrate. Mutants defective in nasA, which encodes anitrate reductase, do not produce normal amounts of EPS1 [72].
Other virulence factors in both tomato and Arabidopsis includecell wall degrading enzymes [13,73e75], which are secretedthrough R. solanacearum's type II protein secretion system (T2SS)[26]. Both pectolyctic and cellulolytic wall degrading enzymes in R.solanacearum have been described. A R. solanacearum mutantdefective in the T2SS has only weak virulence [76], and is lessvirulent than mutants defective only in CWDE, suggesting that inaddition to CWDE, other proteins secreted through the T2SS areimportant for virulence [14]. Surprisingly, a triple mutant defectivein three pectolytic enzymeswasmore virulent than either a mutantdefective in six CWDE (both pectolytic and cellulolytic) [75] or twocellulolytic enzymes. The authors attribute this to a lack of plantdefense stimulation in the triple mutant [75]. Similar to infection intomato, CWDE are important for R. solanacearum infection in Ara-bidopsis. Inoculation with R. solanacearum GMI1000 resulted inpectin degradation in the root of wild-type Col-0 Arabidopsis [57].
Another significant aspect of R. solanacearum virulence ismotility. Nonmotile mutants that either lacked flagella or haddefective flagella had reduced virulence when soil-soak inoculatedon tomato [77]. However, when the nonmotile fliC mutant wasinoculated directly into the xylem via petiole inoculation, it wasable to cause disease [77]. These data suggest that flagella areimportant for locating roots but are dispensable once bacteria havereached the xylem. Twitching motility, which provides a flagella-independent form of surface movement for Ralstonia sol-ancearum, also plays a role in virulence [78], as mutants defective inthis ability show reduced virulence in tomato [78].
How R. solanacearum thrives in the oxygen- and nutrient-limited environment of the xylem has been puzzling. However,recent work has demonstrated that the ability to use nitrate is animportant part of R. solanacearum virulence in tomato [72,79,80]. R.solanacearum infection reduces already low levels of oxygen in thexylem [80]. Tomato xylem sap contains significantly high levels ofnitrate. Work by Dalsing et al., 2015 [80] showed that duringinfection, R. solanacearum consumed the available oxygen, and wasable to use inorganic N as an energy source with nitrate as a ter-minal electron acceptor. Consistent with this, the genome ofGMI1000, a phylotype I strain of R. solanacearum, contains a set ofnitrate respiration and denitrification genes (narG, aniA, norB, nosZand hmpX) [29,80]. Further, transcriptomic analysis of bacteriagrowing in planta showed that all of these genes were significantlyexpressed during R. solanacearum pathogenesis in tomato [79].Single mutant analysis showed that two mutants with defects inthe nitrate detoxification pathway, aniA and hmpX, caused less wiltdisease thanwild type R. solanacearum [80]. Thus, the ability to useinorganic N is an important part of the pathogen's ability to colo-nize the tomato host and cause disease.
4.2. Pathogen effectors and the T3SS
Like most plant pathogens, R. solanacearum uses the type III
secretion system (T3SS) to inject type III effectors (T3Es) into thehost cell, causing disease in susceptible plants and defense re-sponses in resistant plants [30]. hrp genes are induced in planta, andR. solanacearum is thus far the only species in which induction ofhrp genes has been shown to require physical contact between thebacterium and the plant host cell wall [81e84]. In R. solanacearum,an outer membrane receptor known as PrhA recognizes a non-diffusible plant cell wall molecule and initiates a transcriptionalcascade that results in the activation of the master regulator hrpG,followed by hrpB. HrpB then directly controls the expression of theT3SS machinery and T3E expression [26]. In addition to its role inactivating the T3SS through hrpB, HrpG also directs the expressionof hundreds of hrpB-independent genes that are important forpathogen virulence [15,85]. The hrpB regulon is expressed in plantaboth at early stages of plant infection and during later wiltingstages [79,86]. Both hrpB and hrpG genes are necessary for tomatoroot infection, although at different stages of the process [87].
Genomic sequencing has revealed that R. solanacearum strainshave, on average, between 60 and 75 different T3Es [27,31], severalof which are part of multigene families. About 50% of the predictedT3Es in the RSSC are thought to be specific to this species complex[26], and many are highly conserved within phylotypes. Forexample, within phylotype II, 54 effectors are conserved amongfour sequenced strains [26]. Thirty-two effectors are conservedacross at least 10 of the fully sequenced RSSC genomes [31],although a minimal set of “core” effectors has not yet been iden-tified. Despite the effector conservation within phylotype, thereappears to be little correlation between host range and phylotype[26].
Of the predicted 110 T3E gene families in the RSSC [26], onlyabout two dozen have a known function [30]. Genetic analyses havedemonstrated that several T3Es have roles in R. solanacearumvirulence. For example, PopS and GALA4 suppress plant defenses intomato [79,88], while GALA7 is required for virulence in M. trun-catula [56]. In tobacco cells, PopA localizes to the membrane andforms an ion-conducting pore, which may promote translocation ofbacterial proteins from the apoplast into the host cell cytoplasm[89]. Effectors in the RSSC also have roles in avirulence. AvrA is amajor avirulence factor in tobacco [90,91], while PopP2 interactswith the Arabidopsis resistance protein RRS1-R [32]. However,effector function is dependent on host species, as both AvrA andPopP2 contribute to pathogen growth in tomato (AvrA and PopP2)[92] or M. truncatula (AvrA) [56]. Depending on the strain, the R.solanacearum genome also contains effectors that are homologs ofXanthomonas TAL effectors, known as RipTALs in R. solanacearum[27,93,94].
In part due to the prevalence of gene families among RSSC ef-fectors, multiplemutations are often necessary to see a reduction invirulence. For example, knocking out all five members of the AWRgene family causes reduced virulence in tomato [95]. A triplemutant in members of the HLK family also shows a reduction invirulence [96]. The GALA T3E family contains eight genes thatencode proteins with F-box motifs [97]. Four members of thisfamily interact with Arabidopsis Skp-1 like proteins, suggestingtheymay be involved in proteasome-mediated degradation of planthost targets [97]. Mutants lacking all seven GALA genes in GMI1000have reduced virulence in Arabidopsis, tomato, and eggplant [88].For a more detailed description of T3Es in R. solanacearum, thereader is referred to [10,26,30,31].
Understanding the R. solanacearum genes used during patho-genesis is helping to shed light on virulence mechanisms in planta.For example, using a strain of R. solanacearum expressing a lumi-nescent reporter Montiero et al. [86] showed that the gene con-trolling EPS biosynthesis is activated in bacteria that colonize thexylem. In a different study, examination of gene expression from
B.-S. Kim et al. / Physiological and Molecular Plant Pathology 95 (2016) 37e43

two strains of R. solanacearum in planta identified genes that weredifferentially expressed in tomato during early bacterial wilt dis-ease [79]. Surprisingly, 45% of HrpB- regulated genes were upre-gulated in planta, suggesting that T3SS has an important rolethroughout the disease cycle. Virulence factors such as EPS1biosynthesis genes and mobility genes (pilA and fliC) wereexpressed highly during pathogenesis, as were pathways for su-crose uptake and metabolism. Sucrose regulon genes were upre-gulated significantly in planta. A mutant defective in the scrA genewas less virulent than the wild type pathogen when inoculated tounwounded tomato roots, but not when introduced to the xylemthrough cut petioles. However, calculation of the population size ofthe wild type and scrA mutant in the xylem during wilting showeda reduction in scrA growth, suggesting that the scrA mutant had areduction in fitness during pathogenesis [79]. Knowledge of the R.solanacearum genes active during infection will lead to greaterunderstanding of host resistance and susceptibility.
5. Conclusions
In conclusion, R. solanacearum is a diverse pathogen that uses abattery of virulence mechanisms to cause disease. While there isstill much to be learned about host plant resistance, mechanisms ofpathogen virulence are becoming clearer. Future studies willcontinue to investigate pathogen virulence mechanisms and willgenerate insight into host plant resistance through QTL cloning,examination of the molecular mechanisms of resistance in roots,and through the identification of additional plant proteins targetedby R. solanacearum effectors.
Acknowledgments
We thank members of the Iyer-Pascuzzi lab for comments andPurdue University for funding. EF is supported by a NSF (grant no:DGE-1333468) graduate research fellowship.
References
[1] J. Mansfield, S. Genin, S. Magori, V. Citovsky, M. Sriariyanum, P. Ronald,M. Dow, V. Verdier, S.V. Beer, M.A. Machado, I. Toth, G. Salmond, G.D. Foster,Top 10 plant pathogenic bacteria in molecular plant pathology, Mol. PlantPathol. 13 (6) (2012) 614e629.
[2] A.C. Hayward, Biology and epidemiology of bacterial wilt caused by pseudo-monas-solanacearum, Annu. Rev. Phytopathol. 29 (1991) 65e87.
[3] J.W. Scott, J.F. Wang, P.M. Hanson, Breeding tomatoes for resistance to bac-terial wilt, a global view, Proc. 1st Int. Symposium Tomato Dis. 695 (2005)161e172.
[4] D.J. Norman, M. Zapata, D.W. Gabriel, Y.P. Duan, J.M. Yuen, A. Mangravita-Novo, R.S. Donahoo, Genetic diversity and host range variation of Ralstoniasolanacearum strains entering North America, Phytopathology 99 (9) (2009)1070e1077.
[5] C.R. Clarke, D.J. Studholme, B. Hayes, B. Runde, A. Weisberg, R. Cai,T. Wroblewski, M.C. Daunay, E. Wicker, J.A. Castillo, B.A. Vinatzer, Genome-enabled phylogeographic investigation of the quarantine pathogen ralstoniasolanacearum race 3 biovar 2 and screening for sources of resistance againstits core effectors, Phytopathology 105 (5) (2015) 597e607.
[6] J. Vasse, P. Frey, A. Trigalet, Microscopic studies of intercellular infection andprotoxylem invasion of tomato roots by pseudomonas-solanacearum, Mol.Plant-Microbe Interact. 8 (2) (1995) 241e251.
[7] T. Denny, S. Baek, Genetic evidence that extracellular polysaccharide is aviurlence factor of Pseudomonas solanacearum, Mol. Plant Microbe Interact. 4(1991) 198e206.
[8] J.A. McGarvey, T.P. Denny, M.A. Schell, Spatial-temporal and quantitativeanalysis of growth and EPSI production by Ralstonia solanacearum in resistantand susceptible tomato cultivars, Phytopathology 89 (12) (1999) 1233e1239.
[9] E. Saile, J.A. McGarvey, M.A. Schell, T.P. Denny, Role of Extracellular Poly-saccharide and Endoglucanase in Root Invasion and Colonization of TomatoPlants by Ralstonia solanacearum, Phytopathology 87 (12) (1997) 1264e1271.
[10] S. Genin, Molecular traits controlling host range and adaptation to plants inRalstonia solanacearum, New Phytol. 187 (4) (2010) 920e928.
[11] A. Milling, L. Babujee, C. Allen, Ralstonia solanacearum extracellular poly-saccharide is a specific elicitor of defense responses in wilt-resistant tomatoplants, PLoS One 6 (1) (2011) e15853.
[12] C. Allen, J. Gay, L. Simon-Buela, A regulatory locus, pehSR, controls poly-galacturonase production and other virulence functions in Ralstonia sol-anacearum, Mol. Plant Microbe Interact. 10 (9) (1997) 1054e1064.
[13] Q. Huang, C. Allen, An exo-poly-alpha-D-galacturonosidase, PehB, is requiredfor wild-type virulence of Ralstonia solanacearum, J. Bacteriol. 179 (23) (1997)7369e7378.
[14] H. Liu, S. Zhang, M.A. Schell, T.P. Denny, Pyramiding unmarked deletions inRalstonia solanacearum shows that secreted proteins in addition to plant cell-wall-degrading enzymes contribute to virulence, Mol. Plant Microbe Interact.18 (12) (2005) 1296e1305.
[15] M. Valls, S. Genin, C. Boucher, Integrated regulation of the type III secretionsystem and other virulence determinants in Ralstonia solanacearum, PLoSPathog. 2 (8) (2006) e82.
[16] N.D. Young, D. Danesh, Understanding bacterial wilt resistance in tomatothrough the use of DNA genetic-markers, Bact. Wilt (1994) 145e155.
[17] D. Danesh, S. Aarons, G.E. McGill, N.D. Young, Genetic dissection of oligogenicresistance to bacterial wilt in tomato, Mol. Plant Microbe Interact. 7 (4) (1994)464e471.
[18] P. Thoquet, J. Olivier, C. Sperisen, P. Rogowsky, H. Laterrot, N. Grimsley,Quantitative trait loci determining resistance to bacterial wilt in tomatocultivar Hawaii7996, Mol. Plant-Microbe Interact. 9 (9) (1996) 826e836.
[19] P. Thoquet, J. Olivier, C. Sperisen, P. Rogowsky, P. Prior, G. Anais, B. Mangin,B. Bazin, R. Nazer, N. Grimsley, Polygenic resistance of tomato plants to bac-terial wilt in the French West Indies, Mol. Plant-Microbe Interact. 9 (9) (1996)837e842.
[20] B. Mangin, P. Thoquet, J. Olivier, N.H. Grimsley, Temporal and multiplequantitative trait loci analyses of resistance to bacterial wilt in tomato permitthe resolution of linked loci, Genetics 151 (3) (1999) 1165e1172.
[21] J.F. Wang, J. Olivier, P. Thoquet, B. Mangin, L. Sauviac, N.H. Grimsley, Resis-tance of tomato line Hawaii7996 to Ralstonia solanacearum Pss4 in Taiwan iscontrolled mainly by a major strain-specific locus, Mol. Plant-MicrobeInteract. 13 (1) (2000) 6e13.
[22] A. Carmeille, C. Caranta, J. Dintinger, P. Prior, J. Luisetti, P. Besse, Identificationof QTLs for Ralstonia solanacearum race 3-phylotype II resistance in tomato,Theor. Appl. Genet. 113 (1) (2006) 110e121.
[23] J.-F. Wang, F.-I. Ho, H.T.H. Truong, S.-M. Hugan, C.H. Balatero,V. Dittapongpitch, N. Hidayati, Identification of major QTLs associated withstable resistance of tomato cultivar ‘Hawaii 7996’ to Ralstonia solanacearum,Euphytica 190 (2013) 241e252.
[24] T.P. Denny, A.C. Hayward, Ralstonia, third ed., APS Press, 2001.[25] M. Fegan, P. Prior, How complex is the Ralstonia solanacearum species com-
plex?, in: C. Allen, A.C. Hayward (Eds.), Bacterial Wilt: the Disease and theRalstonia Solanacearum Species Complex, American Phytopathological Soci-ety Press, St. Paul, MN, USA, 2005, pp. 449e461.
[26] S. Genin, T.P. Denny, Pathogenomics of the Ralstonia solanacearum speciescomplex, Annu. Rev. Phytopathol. 50 (2012) 67e89.
[27] N. Peeters, S. Carrere, M. Anisimova, L. Plener, A.C. Cazale, S. Genin, Repertoire,unified nomenclature and evolution of the Type III effector gene set in theRalstonia solanacearum species complex, BMC genomics 14 (2013) 859.
[28] B. Remenant, L. Babujee, A. Lajus, C. Medigue, P. Prior, C. Allen, Sequencing ofK60, type strain of the major plant pathogen Ralstonia solanacearum,J. Bacteriol. 194 (10) (2012) 2742e2743.
[29] M. Salanoubat, S. Genin, F. Artiguenave, J. Gouzy, S. Mangenot, M. Arlat,A. Billault, P. Brottier, J.C. Camus, L. Cattolico, M. Chandler, N. Choisne,C. Claudel-Renard, S. Cunnac, N. Demange, C. Gaspin, M. Lavie, A. Moisan,C. Robert, W. Saurin, T. Schiex, P. Siguier, P. Thebault, M. Whalen, P. Wincker,M. Levy, J. Weissenbach, C.A. Boucher, Genome sequence of the plant path-ogen Ralstonia solanacearum, Nature 415 (6871) (2002) 497e502.
[30] N.S. Coll, M. Valls, Current knowledge on the Ralstonia solanacearum type IIIsecretion system, Microb. Biotechnol. 6 (6) (2013) 614e620.
[31] L. Deslandes, S. Genin, Opening the Ralstonia solanacearum type III effectortool box: insights into host cell subversion mechanisms, Curr. Opin. plant Biol.20 (2014) 110e117.
[32] L. Deslandes, J. Olivier, N. Peeters, D.X. Feng, M. Khounlotham, C. Boucher,I. Somssich, S. Genin, Y. Marco, Physical interaction between RRS1-R, a proteinconferring resistance to bacterial wilt, and PopP2, a type III effector targetedto the plant nucleus, Proc. Natl. Acad. Sci. U. S. A. 100 (13) (2003) 8024e8029.
[33] M. Rossi, F.L. Goggin, S.B. Milligan, I. Kaloshian, D.E. Ullman, V.M. Williamson,The nematode resistance gene Mi of tomato confers resistance against thepotato aphid, Proc. Natl. Acad. Sci. U. S. A. 95 (17) (1998) 9750e9754.
[34] S.B. Milligan, J. Bodeau, J. Yaghoobi, I. Kaloshian, P. Zabel, V.M. Williamson, Theroot knot nematode resistance gene Mi from tomato is a member of theleucine zipper, nucleotide binding, leucine-rich repeat family of plant genes,Plant Cell. 10 (8) (1998) 1307e1319.
[35] P. Vos, G. Simons, T. Jesse, J. Wijbrandi, L. Heinen, R. Hogers, A. Frijters,J. Groenendijk, P. Diergaarde, M. Reijans, J. Fierens-Onstenk, M. de Both,J. Peleman, T. Liharska, J. Hontelez, M. Zabeau, The tomato Mi-1 gene confersresistance to both root-knot nematodes and potato aphids, Nat. Biotechnol. 16(13) (1998) 1365e1369.
[36] M.J. Dickinson, D.A. Jones, J.D. Jones, Close linkage between the Cf-2/Cf-5 andMi resistance loci in tomato, Mol. Plant Microbe Interact. 6 (3) (1993)341e347.
[37] J.G. van der Beek, G. Pet, P. Lindhout, Resistance to powdery mildew (Oidiumlycopersicum) in Lycopersicon hirsutum is controlled by an incompletely-dominant gene Ol-1 on chromosome 6, Theor. Appl. Genet. 89 (4) (1994)
B.-S. Kim et al. / Physiological and Molecular Plant Pathology 95 (2016) 37e43

467e473.[38] D. Zamir, I. Ekstein-Michelson, Y. Zakay, N. Navot, M. Zeidan, M. Sarfatti,
Y. Eshed, E. Harel, T. Pleban, H. van-Oss, N. Kedar, H.D. Rabinowitch,H. Czosnek, Mapping and introgression of a tomato yellow leaf curl virustolerance gene, TY-1, Theor. Appl. Genet. 88 (2) (1994) 141e146.
[39] G. Parrella, A. Moretti, P. Gognalons, M.L. Lesage, G. Marchoux, K. Gebre-Selassie, C. Caranta, The Am gene controlling resistance to alfalfa mosaic virusin Tomato is located in the cluster of dominant resistance genes on chro-mosome 6, Phytopathology 94 (4) (2004) 345e350.
[40] M. Narusaka, K. Shirasu, Y. Noutoshi, Y. Kubo, T. Shiraishi, M. Iwabuchi,Y. Narusaka, RRS1 and RPS4 provide a dual Resistance-gene system againstfungal and bacterial pathogens, Plant J. Cell Mol. Biol. 60 (2) (2009) 218e226.
[41] M. Narusaka, Y. Kubo, T. Shiraishi, M. Iwabuchi, Y. Narusaka, A dual resistancegene system prevents infection by three distinct pathogens, Plant Signal.Behav. 4 (10) (2009) 954e955.
[42] M. Narusaka, Y. Kubo, K. Hatakeyama, J. Imamura, H. Ezura, Y. Nanasato,Y. Tabei, Y. Takano, K. Shirasu, Y. Narusaka, Breaking restricted taxonomicfunctionality by dual resistance genes, Plant Signal. Behav. 8 (6) (2013)e24244.
[43] M. Narusaka, K. Hatakeyama, K. Shirasu, Y. Narusaka, Arabidopsis dual resis-tance proteins, both RPS4 and RRS1, are required for resistance to bacterialwilt in transgenic Brassica crops, Plant Signal. Behav. 9 (7) (2014) e29130.
[44] K.H. Sohn, C. Segonzac, G. Rallapalli, P.F. Sarris, J.Y. Woo, S.J. Williams,T.E. Newman, K.H. Paek, B. Kobe, J.D. Jones, The nuclear immune receptorRPS4 is required for RRS1SLH1-dependent constitutive defense activation inArabidopsis thaliana, PLoS Genet. 10 (10) (2014) e1004655.
[45] C. Le Roux, G. Huet, A. Jauneau, L. Camborde, D. Tremousaygue, A. Kraut,B. Zhou, M. Levaillant, H. Adachi, H. Yoshioka, S. Raffaele, R. Berthome,Y. Coute, J.E. Parker, L. Deslandes, A receptor pair with an integrated decoyconverts pathogen disabling of transcription factors to immunity, Cell 161 (5)(2015) 1074e1088.
[46] P.F. Sarris, Z. Duxbury, S.U. Huh, Y. Ma, C. Segonzac, J. Sklenar, P. Derbyshire,V. Cevik, G. Rallapalli, S.B. Saucet, L. Wirthmueller, F.L. Menke, K.H. Sohn,J.D. Jones, A plant immune receptor detects pathogen effectors that targetWRKY transcription factors, Cell 161 (5) (2015) 1089e1100.
[47] C. Tasset, M. Bernoux, A. Jauneau, C. Pouzet, C. Briere, S. Kieffer-Jacquinod,S. Rivas, Y. Marco, L. Deslandes, Autoacetylation of the Ralstonia solanacearumeffector PopP2 targets a lysine residue essential for RRS1-R-mediated im-munity in Arabidopsis, PLoS Pathog. 6 (11) (2010) e1001202.
[48] S. Cesari, M. Bernoux, P. Moncuquet, T. Kroj, P.N. Dodds, A novel conservedmechanism for plant NLR protein pairs: the “integrated decoy” hypothesis,Front. plant Sci. 5 (2014) 606.
[49] J. Hu, X. Barlet, L. Deslandes, J. Hirsch, D.X. Feng, I. Somssich, Y. Marco,Transcriptional responses of Arabidopsis thaliana during wilt disease causedby the soil-borne phytopathogenic bacterium, Ralstonia solanacearum, PLoSOne 3 (7) (2008) e2589.
[50] T. Ishihara, I. Mitsuhara, H. Takahashi, K. Nakaho, Transcriptome analysis ofquantitative resistance-specific response upon Ralstonia solanacearuminfection in tomato, PLoS One 7 (10) (2012) e46763.
[51] T. McAvoy, J. Freeman, S. Rideout, S. Olson, P. Paret, Evaluation of graftingusing hybrid rootstocks for managment of bacterial wilt in field tomatoproduction, HortScience 47 (5) (2012) 621e625.
[52] C.L. Rivard, S. O'Connell, M.M. Peet, R.M. Welker, F.J. Louws, Grafting tomato tomanage bacterial wilt caused by Ralstonia solanacearum in the SoutheasternUnited States, Plant Dis. 96 (7) (2012) 973e978.
[53] V. Grimault, B. Gelie, M. Lemattre, P. Prior, J. Schmit, Comparative histology ofresistant and susceptible tomato cultivars infected by Pseudomonas-Sol-anacearum, Physiol. Mol. Plant Pathol. 44 (2) (1994) 105e123.
[54] V. Grimault, P. Prior, Bacterial wilt resistance in tomato associated withtolerance of vascular tissues to Pseudomonas-Solanacearum, Plant Pathol. 42(4) (1993) 589e594.
[55] K. Nakaho, H. Inoue, T. Takayama, H. Miyagawa, Distribution and multiplica-tion of Ralstonia solanacearum in tomato plants iwth resistance derived fromdifferent origins, J. General Plant Pathol. 70 (2) (2004) 115e119.
[56] M. Turner, A. Jauneau, S. Genin, M.J. Tavella, F. Vailleau, L. Gentzbittel,M.F. Jardinaud, Dissection of bacterial Wilt on Medicago truncatula revealedtwo type III secretion system effectors acting on root infection process anddisease development, Plant Physiol. 150 (4) (2009) 1713e1722.
[57] C. Digonnet, Y. Martinez, N. Denance, M. Chasseray, P. Dabos, P. Ranocha,Y. Marco, A. Jauneau, D. Goffner, Deciphering the route of Ralstonia sol-anacearum colonization in Arabidopsis thaliana roots during a compatibleinteraction: focus at the plant cell wall, Planta 236 (5) (2012) 1419e1431.
[58] Nion YA. Yuliar, K. Toyota, Recent trends in control methods for bacterial wiltdiseases caused by Ralstonia solanacearum, Microbes environ./JSME 30 (1)(2015) 1e11.
[59] H. Kurabachew, K. Wydra, Characterization of plant growth promoting rhi-zobacteria and their potential as bioprotectant against tomato bacterial wiltcaused by, Ralst. Solanacearum. Biol. Control 67 (2013) 75e83.
[60] J. Huang, Z. Wei, S. Tan, X. Mei, S. Yin, Q. Shen, Y. Xu, The rhizosphere soil ofdiseased tomato plants as a source for novel microorganisms to control bac-terial wilt, Appl. Soil Ecol. 72 (2013) 79e84.
[61] M. Hyakumachi, M. Nishimura, T. Arakawa, S. Asano, S. Yoshida, S. Tsushima,H. Takahashi, Bacillus thuringiensis supresses bacterial wilt disease caused byRalstonia solanacearum with systemic induction of defense-related geneexpression in tomato, Microbes environ./JSME 28 (2013) 128e134.
[62] H. Takahashi, K. Nakaho, T. Ishihara, S. Ando, T. Wada, Y. Kanayama, S. Asano,S. Yoshida, S. Tsushima, M. Hyakumachi, Transcriptional profile of tomatoroots exhibiting Bacillus thuringiensis-induced resistance to Ralstonia sol-anacearum, Plant Cell. Rep. 33 (1) (2014) 99e110.
[63] C.M. Pieterse, C. Zamioudis, R.L. Berendsen, D.M. Weller, S.C. Van Wees,P.A. Bakker, Induced systemic resistance by beneficial microbes, Annu. Rev.Phytopathol. 52 (2014) 347e375.
[64] S. Hase, A. Shimizu, K. Nakaho, S. Takenaka, H. Takahashi, Induction of tran-sient ethylene and reduction in severity of tomato bacterial wilt by Pythiumoligandrum, Plant Pathol. 55 (4) (2006) 537e543.
[65] H.H. Zhu, Q. Yao, Localized and systemic increase of phenols in tomato rootinduced by Glomus versiformae inhibits Ralstonia solanacearum,J. Phytopathol. 152 (2004) 537e542.
[66] M.M. Tahat, K. Siiam, R. Othman, The potential of endomycorrhizal fungi incontrollng tomato bacterial wilt Ralsotnia solanacearum under glasshousecoditions, Afr. J. Biotechnol. 11 (2012) 13085e13094.
[67] M. Nerome, K. Toyota, T.M. Islam, T. Nishimima, T. Matsuoka, K. Sato,Y. Yamaguchi, Suppression of bacterial wilt of tomato by incorporation ofmunicipal biowaste charcoal into soil, Soil Microorg. 59 (1) (2005) 9e14.
[68] L. Liu, C. Sun, S. Liu, R. Chai, W. Huang, X. Liu, C. Tang, Y. Zhang, Bioorganicfertilizer enhances soil suppressive capacity against bacterial wilt of tomato,PLoS One 10 (4) (2015) e0121304.
[69] L.M. Kiirika, F. Stahl, K. Wydra, Phenotypic and molecular characterization ofresistance induction by single and combined application of chitosan and sil-icon in tomato against, Ralst. solanacearum. Physiol. Mol. Plant Pathol. 81(2013) 1e12.
[70] H. Ghareeb, Z. Bozso, P.G. Ott, C. Repenning, F. Stahl, K. Wydra, Transcriptomeof silicon-induced resistance against Ralstonia solanacearum in the siliconnon-accumulator tomato implicates priming effect, Physiol. Mol. Plant Pathol.75 (2011) 83e89.
[71] Y. Kang, E. Saile, M.A. Schell, T.P. Denny, Quantitative immunofluorescence ofregulated eps gene expression in single cells of Ralstonia solanacearum, Appl.Environ. Microbiol. 65 (6) (1999) 2356e2362.
[72] B.L. Dalsing, C. Allen, Nitrate assimilation contributes to Ralstonia sol-anacearum root attachment, stem colonization, and virulence, J. Bacteriol. 196(5) (2014) 949e960.
[73] M.A. Schell, D.P. Roberts, T.P. Denny, Analysis of the Pseudomonas sol-anacearum polygalacturonase encoded by pglA and its involvement in phy-topathogenicity, J. Bacteriol. 170 (10) (1988) 4501e4508.
[74] J. Tans-Kersten, Y. Guan, C. Allen, Ralstonia solanacearum pectin methyl-esterase is required for growth on methylated pectin but not for bacterial wiltvirulence, Appl. Environ. Microbiol. 64 (12) (1998) 4918e4923.
[75] E.T. Gonzalez, C. Allen, Characterization of a Ralstonia solanacearum operonrequired for polygalacturonate degradation and uptake of galacturonic acid,Mol. Plant Microbe Interact. 16 (6) (2003) 536e544.
[76] Y. Kang, J. Huang, G. Mao, L. He, M.A. Schell, Dramatically reduced virulence ofmutants of Pseudomonas solanacearum defective in export of extracellularproteins across the outer membrane, Mol. Plant Microbe Interact. 7 (1994)370e377.
[77] J. Tans-Kersten, H. Huang, C. Allen, Ralstonia solanacearum needs motility forinvasive virulence on tomato, J. Bacteriol. 183 (12) (2001) 3597e3605.
[78] H. Liu, Y. Kang, S. Genin, M.A. Schell, T.P. Denny, Twitching motility of Ral-stonia solanacearum requires a type IV pilus system, Microbiology 147 (Pt 12)(2001) 3215e3229.
[79] J.M. Jacobs, A. Milling, R.M. Mitra, C.S. Hogan, F. Ailloud, P. Prior, C. Allen,Ralstonia solanacearum requires PopS, an ancient AvrE-family effector, forvirulence and To overcome salicylic acid-mediated defenses during tomatopathogenesis, MBio 4 (6) (2013) e00875e00813.
[80] B.L. Dalsing, A.N. Truchon, E.T. Gonzalez-Orta, A.S. Milling, C. Allen, Ralstoniasolanacearum uses inorganic nitrogen metabolism for virulence, ATP pro-duction, and detoxification in the oxygen-limited host xylem environment,MBio 6 (2) (2015) e02471.
[81] M. Marenda, B. Brito, D. Callard, S. Genin, P. Barberis, C. Boucher, M. Arlat,PrhA controls a novel regulatory pathway required for the specific inductionof Ralstonia solanacearum hrp genes in the presence of plant cells, Mol.Microbiol. 27 (2) (1998) 437e453.
[82] B. Brito, M. Marenda, P. Barberis, C. Boucher, S. Genin, prhJ and hrpG, two newcomponents of the plant signal-dependent regulatory cascade controlled byPrhA in Ralstonia solanacearum, Mol. Microbiol. 31 (1) (1999) 237e251.
[83] D. Aldon, B. Brito, C. Boucher, S. Genin, A bacterial sensor of plant cell contactcontrols the transcriptional induction of Ralstonia solanacearum pathoge-nicity genes, EMBO J. 19 (10) (2000) 2304e2314.
[84] B. Brito, D. Aldon, P. Barberis, C. Boucher, S. Genin, A signal transfer systemthrough three compartments transduces the plant cell contact-dependentsignal controlling Ralstonia solanacearum hrp genes, Mol. Plant MicrobeInteract. 15 (2) (2002) 109e119.
[85] A. Occhialini, S. Cunnac, N. Reymond, S. Genin, C. Boucher, Genome-wideanalysis of gene expression in Ralstonia solanacearum reveals that the hrpBgene acts as a regulatory switch controlling multiple virulence pathways, Mol.Plant Microbe Interact. 18 (9) (2005) 938e949.
[86] F. Monteiro, S. Genin, I. van Dijk, M. Valls, A luminescent reporter evidencesactive expression of Ralstonia solanacearum type III secretion system genesthroughout plant infection, Microbiology 158 (Pt 8) (2012) 2107e2116.
[87] J. Vasse, S. Genin, P. Frey, C. Boucher, B. Brito, The hrpB and hrpG regulatorygenes of Ralstonia solanacearum are required for different stages of the
B.-S. Kim et al. / Physiological and Molecular Plant Pathology 95 (2016) 37e43

tomato root infection process, Mol. Plant Microbe Interact. 13 (3) (2000)259e267.
[88] P. Remigi, M. Anisimova, A. Guidot, S. Genin, N. Peeters, Functional diversifi-cation of the GALA type III effector family contributes to Ralstonia sol-anacearum adaptation on different plant hosts, New Phytol. 192 (4) (2011)976e987.
[89] J. Racape, L. Belbahri, S. Engelhardt, B. Lacombe, J. Lee, J. Lochman, A. Marais,M. Nicole, T. Nurnberger, F. Parlange, S. Puverel, H. Keller, Ca2þ-dependentlipid binding and membrane integration of PopA, a harpin-like elicitor of thehypersensitive response in tobacco, Mol. Microbiol. 58 (5) (2005) 1406e1420.
[90] B.F. Carney, T.P. Denny, A cloned avirulence gene from Pseudomonas sol-anacearum determines incompatibility on Nicotiana tabacum at the hostspecies level, J. Bacteriol. 172 (9) (1990) 4836e4843.
[91] A.E. Robertson, W.P. Wechter, T.P. Denny, B.A. Fortnum, D.A. Kluepfel, Rela-tionship between avirulence gene (avrA) diversity in Ralstonia solanacearumand bacterial wilt incidence, Mol. Plant Microbe Interact. 17 (12) (2004)1376e1384.
[92] A.P. Macho, A. Guidot, P. Barberis, C.R. Beuzon, S. Genin, A competitive indexassay identifies several Ralstonia solanacearum type III effector mutant strainswith reduced fitness in host plants, Mol. Plant Microbe Interact. 23 (9) (2010)1197e1205.
[93] O. de Lange, T. Schreiber, N. Schandry, J. Radeck, K.H. Braun, J. Koszinowski,H. Heuer, A. Strauss, T. Lahaye, Breaking the DNA-binding code of Ralstoniasolanacearum TAL effectors provides new possibilities to generate plantresistance genes against bacterial wilt disease, New Phytol. 199 (3) (2013)773e786.
[94] L. Li, A. Atef, A. Piatek, Z. Ali, M. Piatek, M. Aouida, A. Sharakuu, A. Mahjoub,G. Wang, S. Khan, N.V. Fedoroff, J.K. Zhu, M.M. Mahfouz, Characterization andDNA-binding specificities of Ralstonia TAL-like effectors, Mol. plant 6 (4)(2013) 1318e1330.
[95] M. Sole, C. Popa, O. Mith, K.H. Sohn, J.D. Jones, L. Deslandes, M. Valls, The awrgene family encodes a novel class of Ralstonia solanacearum type III effectorsdisplaying virulence and avirulence activities, Mol. Plant Microbe Interact. 25(7) (2012) 941e953.
[96] L. Chen, M. Shirota, Y. Zhang, A. Kiba, Y. Hikichi, K. Ohnishi, Involvement ofHLK effectors in Ralstonia solancearum disease development in tomato, J. Gen.Plant Pathol. 80 (2014) 79e84.
[97] A. Angot, N. Peeters, E. Lechner, F. Vailleau, C. Baud, L. Gentzbittel, E. Sartorel,P. Genschik, C. Boucher, S. Genin, Ralstonia solanacearum requires F-box-likedomain-containing type III effectors to promote disease on several hostplants, Proc. Natl. Acad. Sci. U. S. A. 103 (39) (2006) 14620e14625.
B.-S. Kim et al. / Physiological and Molecular Plant Pathology 95 (2016) 37e43