the role of prey abundance and flow regulation in the marking behaviour of eurasian otters in a...
TRANSCRIPT

at SciVerse ScienceDirect
Animal Behaviour 84 (2012) 1475e1482
Contents lists available
Animal Behaviour
journal homepage: www.elsevier .com/locate/anbehav
The role of prey abundance and flow regulation in the marking behaviourof Eurasian otters in a Mediterranean catchment
David Almeida a,*, Rafael Barrientos a, Raquel Merino-Aguirre b, David G. Angeler c
aDepartamento de Ciencias Ambientales, Facultad de Medio Ambiente, Universidad de Castilla-La Mancha, Toledo, SpainbDepartment of Ecology, Complutense University of Madrid, Madrid, SpaincDepartment of Aquatic Sciences and Assessment, Swedish University of Agricultural Sciences, Uppsala, Sweden
a r t i c l e i n f o
Article history:Received 31 May 2012Initial acceptance 27 June 2012Final acceptance 7 September 2012Available online 22 October 2012MS. number: 12-00416
Keywords:carnivoreEurasian otterfaecal-marking behaviourfeeding habitflow regulationlatrine featureLutra lutraMediterranean catchmentterritoriality
* Correspondence and present address: D. AlmeiEcology and Environmental Change, Bournemouth U5BB, U.K.
E-mail address: [email protected] (D.
0003-3472/$38.00 � 2012 The Association for the Stuhttp://dx.doi.org/10.1016/j.anbehav.2012.09.020
Scent marking is a common behaviour used to delineate territories by carnivores, yet it remains insuf-ficiently understood as there are several alternative hypotheses, one being that sites are marked whereresources are abundant, thus guaranteeing their exclusive use. Small Mediterranean rivers are highlyvariable environments throughout the year, representing an ideal scenario to test this theory, as preyavailability changes as a function of the spatial and temporal variability of the flow regime. We studiedseasonal variation in the habitat selection of latrine (i.e. scent-marked) sites, prey availability and preyselection of the Eurasian otter, Lutra lutra, between two contrasting fluvial systems, the regulatedmainstem of River Bullaque and four unregulated tributaries (River Guadiana basin, central Spain). In2010, we measured habitat variables, collected otter spraints and used electrofishing to compare the dietwith prey abundance. Latrines were more frequent at shallow sites, and otters did not discriminatebetween fluvial systems across seasons. Also, latrines were associated with sites where the main prey,the calandino, Squalius alburnoides, was more abundant in the tributaries during spring and summer.However, this abundance was not translated into prey selection, with calandino being preyed below itsavailability in the mainstem. This observation suggests that otters mark places where fishing is especiallyeasy (i.e. shallow sites with proportionally more fish). Calandino was substituted by red swamp crayfish,Procambarus clarkii, which was easier to catch but of lower energetic value, where higher water levelsmade fishing more difficult (i.e. the regulated mainstem).� 2012 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.
A major aspect of animal behaviour is intraspecific communi-cation by means of olfactory signals (Gorman 1990). Typically,crucial information potentially transmitted among conspecificswould be the use of resources, which is also a reflection of thehabitat quality and suitability (Ben-David et al. 2005). Thus, thestudy of environmental factors (both biotic and abiotic) affectingthe marking strategy of key species is highly relevant for unveilingother behavioural traits, such as the social organization (Macdonald1980, 1983), especially in habitats in which resources are highlyvariable and more restricted (e.g. Mediterranean fresh waters). Inrelation to this, territoriality is a behaviour consisting of the occu-pation and defence of a territory, within a home range, whichcontains valuable resources, including food, refuge or mates(Gittleman & Harvey 1982). Territories are commonly delimited bymeans of scent marks (Gorman 1990), consisting of faeces, urine
da, Centre for Conservationniversity, Poole, Dorset BH12
Almeida).
dy of Animal Behaviour. Published
and/or secretions of scent glands (Macdonald 1980; Gorman &Trowbridge 1989), which are typically deposited at specific loca-tions known as latrines (reviewed in Brown & Macdonald 1985;Gorman & Trowbridge 1989). Theory posits that carnivores maymark places with higher prey availability to guarantee theirexclusive use (Hutchings & White 2000; Lewis & Moorcroft 2001).Paradoxically, very few studies have tested the latrineeprey avail-ability relationship with empirical data, possibly because suchmeasurements are labour intensive (but see Prenda & Granado-Lorencio 1996; Ben-David et al. 2005).
The Eurasian otter, Lutra lutra, hereafter simply ‘otter’, is mainlysolitary in Mediterranean areas (Ruiz-Olmo et al. 2005a) and tendsto delimit its territory by marking with faeces (spraints), smears orjellies in conspicuous sites (i.e. latrines), which are usually constantthrough time (Macdonald & Mason 1987). In these nonsocial otters,three main hypotheses have been suggested for scent marking(Ralls 1971; Johnson 1973; Kruuk 1992; Ben-David et al. 2005): (1)territory signalling to reduce aggressive encounters with otherindividuals of the same sex; (2) attraction of potential mates, asthey are a good indicator of territory occupation; and (3) signalling
by Elsevier Ltd. All rights reserved.

D. Almeida et al. / Animal Behaviour 84 (2012) 1475e14821476
of resources to optimize their use by holders and exclude strangers.These hypotheses may be difficult to test individually as they arenot mutually exclusive.
Despite some studies having been carried out on otter latrinecharacteristics (e.g. Dubuc et al. 1990; Swimley et al. 1998; DePue &Ben-David 2010), to our knowledge, studies analysing a combina-tion of temporal and spatial variation in latrine features (i.e. thesprainting strategy) are lacking in wild otters (but see Ruiz-Olmoet al. 2005b for female otters in captivity). In particular, seasonalecosystems, like those located in Mediterranean regions, representan ideal study system, as marking should vary in parallel withresource availability throughout the year (Gasith & Resh 1999).Furthermore, hydraulic works (e.g. dikes, weirs and dams), whichare very abundant in Mediterranean areas, alter natural dischargeregimes and modify the ecological interactions of riverine species(Nilsson et al. 2005; Perez-Quintero 2007; Wang et al. 2011). Thissuggests that the influence of human-mediated habitat manage-ment (e.g. flow regulation) on the scent-marking behaviour ofmustelid species can be important, but little is known about it(Hutchings & White 2000).
Some studies have shown that marking behaviour peaks duringwinter and early spring in temperate habitats (Erlinge 1968;Macdonald & Mason 1987; Kruuk 1992), but this is not sopronounced in Mediterranean areas (Delibes et al. 1991; Ruiz-Olmo& Gosálbez 1997; Prigioni et al. 2005). Thus, while otters usuallymark the same sites (i.e. similar structural features), nontraditionalplaces are more commonly marked during seasonal peaks(Macdonald & Mason 1987); hence, we studied whether seasonalchanges in habitat characteristics of latrine sites exist year-round.We also tested the hypothesis about the use of marking to signalpriority of use of resources (Kruuk 1992), since otters feed prefer-entially in certain feeding patches (Kruuk et al. 1990), whichmay beconsidered as key resources from where competitors are repelled(Kruuk 1995). In particular, pools containing water are scarceduring summer drought in nonregulated Mediterranean rivers, andthese points may be valuable patches to be defended. In addition,we aimed to evaluate whether hypothetical latrine selection isrelated to prey availability and also prey selection. In light of thisdearth of information, we studied otter scent marking in a smallMediterranean catchment where two contrasting fluvial systems(regulated mainstem versus nonregulated tributaries) were moni-tored during a 1-year period. More specifically, we hypothesizedthat: (1) otters vary the structural features of latrine sites (e.g. tomake sites more conspicuous) along the seasonal cycle in parallelwith their changing requirements (Ruiz-Olmo et al. 2005a;Barrientos 2006); (2) otters place latrines in pools that retain waterduring summer in tributaries, but not necessarily in the regulatedmainstem because it remains wet throughout the year; and (3)marking is related to both otter diet and prey abundance; hence,latrines should be located at sites where preferred prey are abun-dant (Kruuk 1992).
METHODS
Study Area
The present study was carried out in the 1019 km2 River Bulla-que catchment (altitude: 550e620 m above sea level), which islocated in the River Guadiana basin, central Spain (Fig. 1). The RiverBullaque (94 km length) has a regulated flow regime owing toa dam, which forms the Torre Abraham Reservoir (Fig. 1). The riverhas a constant flow throughout the year with weak seasonal fluc-tuations because of the operation of the dam (Almeida 2008;Almeida et al. 2012a). Four streams are the main drainingtributaries to the River Bullaque (Fig. 1) and share similar
geomorphological characteristics with the mainstem. However,tributaries otherwise have a relatively pristine hydrologicaldisturbance regime and therefore show high flow fluctuations (seeAlmeida et al. 2012a for discharge profiles) associated with theMediterranean climate (Gasith & Resh 1999).
The fish assemblage of the River Bullaque catchment consists of13 species, seven of which are endemic to the Iberian Peninsula:Iberian long-snout barbel, Luciobarbus comizo, Iberian small-headbarbel, Luciobarbus microcephalus, Iberian arched-mouth nase,Iberochondrostoma lemmingii, southern straight-mouth nase, Pseu-dochondrostoma willkommii, calandino, Squalius alburnoides,southern Iberian chub, Squalius pyrenaicus and southern Iberianspined-loach, Cobitis paludica. The other six species, goldfish, Car-assius auratus, common carp, Cyprinus carpio, northern pike, Esoxlucius, eastern mosquitofish, Gambusia holbrooki, pumpkinseed,Lepomis gibbosus, and largemouth bass,Micropterus salmoides, wereinitially introduced in Torre Abraham Reservoir (built in 1997), buthave successfully spread to the downstream area of the River Bul-laque. Red swamp crayfish, Procambarus clarkii, is also an intro-duced species that is very abundant in the whole study area.
General Field Procedure
Like DePue & Ben-David (2010), we defined ‘latrine’ as an ottersprainting site that has three or more faeces (i.e. ensuring thatparticular location was indeed used for marking) and that is iso-lated from other sites by at least 5 m (i.e. to detect subtle differ-ences in the surrounding microhabitat characteristics). To avoidpseudoreplication of information, daily sampling involvedrecording only the variables surrounding latrines on which therewere fresh faeces (i.e. current use), as well as their position usinga hand-held GPS (GARMIN model 72, U.S.A.) with an accuracy of upto 3 m (WAAS system). Surveys were carried out for 2e3 weeks perseason throughout 2010 in River Bullaque, downstream from TorreAbraham dam, and the four tributaries (Fig. 1). For this purpose, wedivided the year into winter, spring, summer and autumn to obtainthe variable called ‘season’. We divided the water courses intomainstem or tributaries, obtaining the variable ‘fluvial system’. Thefive water courses were carefully sampled for otter latrines by ‘zig-zagging’ along the banks and channels (following Lenton et al.1980). Previous studies have measured up to 30 m of river bankfrom the water’s edge (e.g. DePue & Ben-David 2010), since mostotter latrines occur within this distance. However, previous spraintsurveys were carried out seasonally in the study area (Almeida2008) to check the maximum distance from the bank for latrinepresence, which was 8.2 m in spring. Therefore, we restricted thisdistance to 10 m in the current study, as latrines were not foundbeyond this limit. Thus, this longitudinal area (i.e.10 m at each bankalong the river) was considered as the most likely home range, andtherefore territory, for otters. Sampling locations (600e800 mstretches) were separated by 3 km in each water course (i.e. N ¼ 15locations in the River Bullaque and N ¼ 6 locations for every trib-utary). Surveys were located ca. 5 km away from the confluences tominimize influences between regulated and nonregulated systems.We sampled 180 (i.e. 34 in winter, 48 in spring, 59 in summer and39 in autumn) and 172 (i.e. 44, 72, 21 and 35, respectively) latrinesin the River Bullaque and the four tributaries, respectively (meandistance between adjacent latrines at one particular samplinglocation/stretch was about 70 m). At each latrine, all remains werecollected. Spraint removal and subsequent rinsing of the site withwater allowed otters to select the latrine site according to itsenvironmental features, since this procedure partially avoided theeffect of attraction, both visual and odour, by previous spraints onthe otter’s decision to defecate on that site (e.g. number of daysbetween two consecutive defecations at latrine sites without

Bullaque River
N Torre Abraham Reservoir
Guadiana River
Bullaque River
Tamujar Stream
Alcobilla Stream
Piedrala Stream
Bullaquejo Stream
10 km
Figure 1. Map of the study area. The mainstem (River Bullaque) after Torre Abraham Reservoir and the four tributaries were sampled (see details in Methods).
D. Almeida et al. / Animal Behaviour 84 (2012) 1475e1482 1477
spraint removal ¼ 6.50 � 0.51, with spraint removal ¼ 8.27 � 0.45;TukeyeKramer HSD test: P < 0.01). Moreover, 200 and 192 unusedsites in the River Bullaque and the four tributaries, respectively,were similarly sampled for habitat structure and prey availabilityon the same sampling dates (i.e. 39 in winter, 53 in spring, 64 insummer and 44 in autumn in the River Bullaque and 49, 77, 26 and40, respectively in the tributaries). These sites were randomlyselected by generating coordinates with ArcView 3.3 (Esri, Red-lands, CA, U.S.A.) and balanced as much as possible, taking intoaccount the number of latrine points per season and fluvial cate-gory. When a random site landed onwater or>10 m away from thebank, we moved the plot in a straight line to the nearest accessiblesite (e.g. bank or a stone in the water). Otter spraints were used toanalyse the diet in the laboratory. As the ingested biomass wasaveraged in every latrine, we obtained a single value per latrine.According to the preliminary results after otter spraint genotyping(D. Almeida, unpublished data), 22 different individuals (i.e. 12males and 10 females) would have been identified in the study area.The latrine overlap (i.e. one particular latrine site being used bydifferent individuals) was relatively low (i.e. <12% of latrine sites).
Habitat Structure
A total of nine limnological/structural characteristics wereconsidered as key factors for otters to select particular latrine sites.We measured habitat characteristics in a similar way to those re-ported in previous otter latrine studies (Dubuc et al. 1990; Swimleyet al. 1998; DePue & Ben-David 2010). Specifically, five structural
variables were determinedwithin a 2 m radius, pivoting around thepoint with the highest concentration of otter spraints at eachlatrine (Crait & Ben-David 2007): (1) ‘latrine height’ above theground or current water level (cm); (2) ‘water depth’ (cm); (3)‘water velocity’ (cm/s); (4) ‘coarseness’ (i.e. substratum composi-tion); and (5) ‘shade’ (% visually assessed at 1200 hours solar time).The sampling for these five variables was adapted from DePue &Ben-David (2010) to characterize the structure of latrine siteparticularly at a microhabitat scale. Water depth and velocity wererandomly measured at 10 points within a 2 m radius and averaged.Coarseness was calculated following Statzner et al. (1988) fromvisual assessments, according to the particle size classification byPlatts et al. (1983). The remaining structural variables weremeasured at a larger (i.e. mesohabitat) scale in every latrine sitefollowing Prenda & Granado-Lorencio (1996): (6) ‘bankside vege-tation cover’ (%); (7) ‘bankside vegetation width’ (m); (8) ‘wettedwidth’ (m); and (9) ‘latrine distance to the nearest bank’ (% from thebank to the middle channel, with 0% in the bank). Vegetation coverwas assessed by the percentage cover (2 m bankside) of helophytes,shrubs and trees. Short grass and herbaceous vegetation wereexcluded because they offer little cover for otters (Mason &Macdonald 1986; Delibes et al. 1991). Vegetation width wasmeasured to a distance of 10 m from the bank (2 m bank-side � 10 m width). Prenda & Granado-Lorencio (1996) showedthat vegetation assemblage is an important factor determiningotter sprainting activity. However, riparian vegetation was rela-tively homogeneous along the water courses under study andprevious observations did not reveal any influence of plant

Table 1Significant differences between use and availability of latrine sites at 95% afterbinomial regression for habitat characteristics and prey abundances
Habitat Season Fluvial system Latrine Unused
Depth (cm) e e 19.59�0.53 25.95�1.06Distance (%) e e 14.06�1.46 5.86�1.71Prey abundanceCalandino (kg/ha) Spring Tributaries 7.72�0.82 5.51�0.56
Summer Tributaries 6.43�1.04 2.66�0.42Southern Iberian
chub (kg/ha)e Mainstem 1.46�0.35 0.60�0.15e Tributaries 0.83�0.25 1.48�0.28
Results are means � SE.
D. Almeida et al. / Animal Behaviour 84 (2012) 1475e14821478
composition on otter marking behaviour (i.e. nonsignificantdifferences between latrine and unused sites for the mean scores ofthe first two dimensions, axes 1 and 2, derived from a principalcomponents analysis on the vegetation data; TukeyeKramer HSDtests: overall P > 0.05). Therefore, physical development features(i.e. shade, cover and width) were the only variables used forestimating the influence of riparian vegetation. Latrine distancewas calculated as a percentage to control the effect of wettedwidth.This percentage was either positive or negative depending on thelocation of the latrine site in the channel or from the bank to theland, respectively. All the measurements were made by D.A.
Prey Availability
On the same sampling dates, fish and crayfish biomass(measured as kg/ha) was estimated 100 m around every latrine andevery unused site (i.e. 50 m upstream and downstream from thesite), by isolating the stretch with block nets, and electrofishing(2000 W DC generator at 200e250 V, 2e3 A) in an upstreamdirection, following the removal sampling without replacement orZippin method (1956), with three passes made (sampling time foreach pass 20e30 min). Fish and crayfish were immediatelyimmersed in an innocuous solution of anaesthetic (MS-222 at 0.1 g/litre), identified to the species level, counted and weighed (�0.1 g)to calculate their biomass. Fish were kept in a tank and suppliedwith oxygen (two ‘Aera’ aerators, portable battery pump) until fullrecovery before releasing them. All field procedures complied withanimal use and care regulations of Europe and Spain. Specifically,electrofishing was performed by a trained licence holder (D.Almeida), who had already carried out electrofishing in the samestudy area in previous studies (e.g. Almeida et al. 2009). Thus, noadverse effects were caused on the wildlife in the river and allanimals recovered fully from the anaesthetic. The species includedin our analyses were calandino, southern Iberian chub, southernIberian spined-loach, pumpkinseed, largemouth bass and redswamp crayfish. We also detected the other species mentionedabove (see Study Area) either alive or in otter spraints. However, weexcluded these latter species from the analysis as none of themreached the 10% of ingested biomass at any fluvial system or season,and therefore could not be considered as key resources (Barrientos& Virgós 2006).
Dietary Analysis
In the laboratory, otter spraints were soaked in soapy water for24 h and dried in the oven at 60 �C for 1 h. Food items were iden-tified to the lowest possible taxonomic level using a dedicatedreference collection of fish and crayfish remains, as well as thepublished literature (Conroy et al. 1993; Miranda & Escala 2002,2007; Arce 2011). Food itemswere also counted (minimumnumberfor each prey category per spraint). The diagnostic structures of fish(e.g. vertebrae, pharyngeal arches) and crayfish (e.g. exopodites,telson) were measured with a digital calliper to the nearest0.01 mm to extrapolate them to the ingested biomass of everyspecimen by means of regression equations (Conroy et al. 1993;Miranda & Escala 2002, 2007).
Statistical Analyses
Data were transformed by using ln (x þ 1). Particularly forpercentage data, logit transformation was used (Warton & Hui2011). Assumptions of normality of distributions and homoge-neity of variance were verified using ShapiroeWilk and Levenetests, respectively. All statistical analyses were performed with the
statistical software R version 2.10.1 (R Development Core Team2009). The significance level was set at a ¼ 0.05.
We used generalized linear models (GLZ), using the log-likelihood ratio test with a type III hypothesis (ANOVA Type IIIfunction in the ‘car’ package), with binomial distribution (latrinepresence versus absence) and logit link function. We evaluated thepure effects of the continuous variables as well as their interactionswith the two categorical ones (i.e. season and fluvial system), bothone-by-one and both categorical variables at the same time. We didnot explore quadratic or cubic relationships because these have notbeen described for the studied variables in previous work on otters.In ANOVA Type III, a model including the variable of interest iscompared to a model that lacks it. The log-likelihood ratio test usesa chi-square test to test whether these two models are significantlydifferent (Moya-Laraño & Wise 2007). The Bonferroni step-downcorrection (Holm 1979) was applied to the tests generated withthis procedure. The library ‘effects’ was used for calculating partialeffects. In a previous study (Almeida et al. 2012a), no differenceswere found between the four studied tributaries in limnologicalbiotic and abiotic features. Thus, in the present work, we consid-ered all the tributaries as a single unit. However, we formerly testedfor random effects (i.e. river as a random factor), fitting the generallinear mixed models with lmer in the library lme4 from the Rstatistical software. However, as our results remain essentiallyidentical, random effects are not shown in this paper.
As we estimated prey availabilities by using abundance datafrom electrofishing at the unused sites, we could evaluate the preyselection by comparing these data with ingested biomasses fromspraints. Prey selectionwas analysed using a similar GLZ procedure,with binomial distribution (ingested biomass versus availability).This analytical approach minimizes the influence of otter ambula-tion (i.e. defecation in a site different from that where the otter wasforaging; e.g. Ruiz-Olmo et al. 2001; Almeida et al. 2012b). Tostandardize the two biomass measures used for our explanatoryvariables (i.e. kg/latrine for ingested biomass and kg/ha for preyavailability), we adjusted them to represent the percentage of eachprey per latrine or per ha, respectively.
RESULTS
Selection of Latrine Features
The probability of finding a latrine decreased with increasingwater depth (Table 1). This pattern was consistent across seasonsand both fluvial systems, since the interaction between these twofactors was not significant (Table 2). Moreover, the probability offinding a latrine increased with the distance to the bank (Table 1),again in all the seasons and fluvial systems (Table 2).
The relationship between calandino abundance and latrinepresence varied both with season and fluvial system (Table 2). Siteswith more biomass of this species were positively selected forlatrine placement during spring and summer in tributaries

Table 2Results from ANOVA Type III tests separating unused sites from those where latrineswere found
Variable Log likelihood c2 df P
Water depth 28.28 1 <0.001Distance to the nearest bank 13.84 1 0.008Season*fluvial system*calandino 17.30 3 0.024Fluvial system*southern Iberian chub 11.71 1 0.024
Only significant tests after Bonferroni step-down correction are shown.
Table 4Significant differences between use and availability at 95% after binomial regressionfor the main prey of otter
Prey Season Fluvialsystem
Ingestedbiomass
Availability
Calandino (%) e Mainstem 26.07�2.22 41.11�2.40Pumpkinseed (%) e Tributaries 1.21�0.42 <0.01�<0.001Largemouth bass (%) e Tributaries 0.58�0.28 <0.01�<0.001Red swamp crayfish (%) Winter e 17.94�3.62 34.43�4.14
Spring e 38.53�3.30 28.70�3.39Autumn e 43.63�4.42 24.11�3.54
Results are means � SE.
D. Almeida et al. / Animal Behaviour 84 (2012) 1475e1482 1479
(Table 1). However, the abundance of its sister species, the southernIberian chub, showed a different pattern in every fluvial systemwhen compared with the probability of latrine presence (Table 2).Thus, whereas otters placed latrines in those sites with morebiomass of southern Iberian chub in the mainstem (Table 1), sitesselected in tributaries contained a lower abundance of this fishspecies (Table 1).
Prey Selection
Calandino was the only prey that appeared to influence latrineselection (see above) and prey selection (negatively in this case;Tables 3, 4). This fish species was consumed below its availability inthe mainstem (Table 4). The ingested biomass of introduced cen-trarchids, pumpkinseed and largemouth bass also interacted withthe factor ‘fluvial system’ (Table 3). In particular, otters capturedthese two species above their availability in tributaries (Table 4).Finally, the pattern observed for red swamp crayfish was related toseasonal variation (Table 3), with otters consuming this prey belowits availability during winter (Table 4), whereas in spring andautumn otters tended to consume this prey above its availability(Table 4). However, none of these selections were mirrored inlatrine site selection (see above).
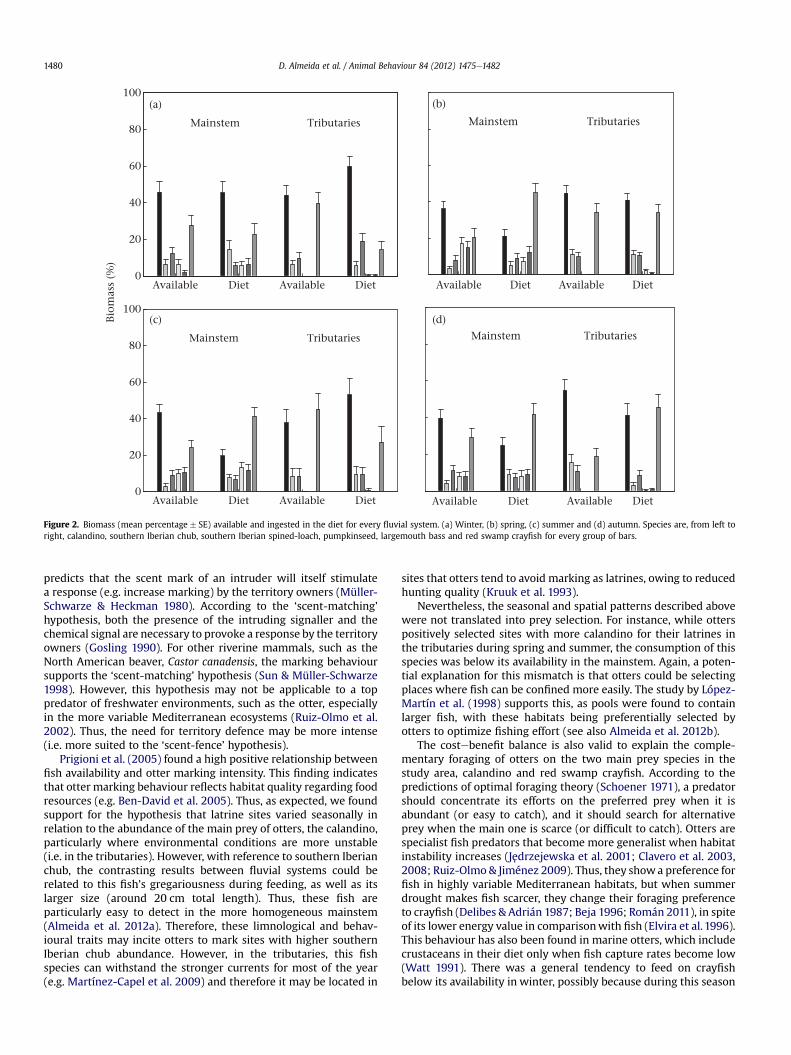
Prey Selection and Diet Output
Despite calandino being the main prey in the study area, witha year-round mean of 38% of ingested biomass (Fig. 2), there wasa general trend to consume it below its availability in the mainstem(Fig. 2, see also above). Indeed, whereas calandino decreased from49% year-round in tributaries to 28% in the mainstem (Fig. 2), redswamp crayfish, the second most important prey, showed theopposite pattern, increasing from 30% to 38%, respectively (Fig. 2).The negative selection of this crustacean during winter was re-flected in the otter’s diet, as the lowest percentages of ingestedbiomass were estimated during winter (Fig. 2). Both introducedpumpkinseed and largemouth bass occurred infrequently in theotter’s diet (Fig. 2), despite otters showing a preference to consumethem more often than expected in tributaries. Pumpkinseed wasa negligible prey in tributaries (mean 1% year-round) and of littleimportance in the mainstem (9%, Fig. 2). Similarly, consumption oflargemouth bass only reached the 1% year-round in tributaries and10% in the mainstem (Fig. 2).
Table 3Results from ANOVA Type III tests differentiating the biomass ingested by ottersfrom the available biomass in the environment
Variable Log likelihood c2 df P
Fluvial system*calandino 19.16 1 <0.001Fluvial system*pumpkinseed 28.74 1 <0.001Fluvial system*largemouth bass 11.20 1 <0.01Season*red swamp crayfish 21.89 3 <0.001
Only significant tests after Bonferroni step-down correction are shown.
DISCUSSION
Regarding habitat characteristics, pools (i.e. deeper locations)had a similar importance at latrine sites in the mainstem andtributaries throughout the year. Therefore, we did not find supportfor our expectation that pools are more important in tributaries (i.e.dry and highly fragmented during the summer drought) than in themainstem. A potential explanation for this mismatch is that otterscould be marking places where fishing is especially easy, with thisfeature being selected elsewhere (e.g. Almeida et al. 2012b). In fact,the ease of prey capture is a key factor in maximizing the energeticbalance (i.e. costebenefit) of foraging (Schoener 1971), and thusotters prefer to forage in shallow locations, either in tributaries orin the mainstem, because prey usually aggregate in small watervolumes, which better facilitates fishing than open water (López-Martín et al. 1998; Ruiz-Olmo et al. 2001, 2007; Barrientos et al.2003). Otters also showed a tendency to mark away from theriver’s edge, often on rocks above the water level, and these areprobably selected to ensure the maximum detection probability offaecal marks and to facilitate the dispersal of odour by wind(Macdonald & Mason 1987; Swimley et al. 1998; DePue & Ben-David 2010). Similarly, latrines in common genets, Genetta gen-etta, are also assumed to have a territorial function as they areplaced in conspicuous places, such as old trees or rocky outcrops(Barrientos 2006; Espírito-Santo et al. 2007), which, in turn,represent important sources of refuge (Espírito-Santo et al. 2007). Arelative of the genet, the meerkat, Suricata suricatta, also uselatrines to defend a territory from neighbouring groups and pro-specting individuals, but latrines are also positioned close to refugesites, that is, they signal a resource (Jordan et al. 2007). Anotherexample of signalling resources by means of scent marks is theEuropean badger, Meles meles, with males visiting boundarylatrines more often than females, particularly during the matingseason to prevent other males from mating with females living intheir territories, the females therefore in this case being a resourceto be defended (Roper et al. 1993). Marking with faeces onconspicuous substrata is also displayed by larger carnivores, such aswolves, Canis lupus, which are able to discriminate plants accordingto their physical properties (Barja 2009). In addition, the selectionof conspicuous substrata is particularly important in otters, as theeffectiveness of marking seems to be short term when it is fooddependent and prey could become scarce relatively soon (Kruuk1992).
Regarding the effectiveness and function of marking, spraint canprovide information on the social status of a territory holder(Rostain et al. 2004), but we could not link scent marks to indi-vidual otters because we did not mark the otters (because of theirlegal status in a nearby Spanish National Park, ‘Cabañeros’) andspraints were not genotyped (as we lacked the resources for pro-cessing the large number of samples, >500 spraints). Thus, wecould not test the so-called ‘scent-fence’ and ‘scent-matching’hypotheses (e.g. Gosling 1982). The ‘scent-fence’ hypothesis
0
20
40
60
80
100
Mainstem
(a)
Tributaries Mainstem
(b)
TributariesB
iom
ass
(%)
0
20
40
60
80
100
Mainstem
(c)
Tributaries
Available Diet Available Diet Available Diet Available Diet
Available Diet Available Diet Available Diet Available Diet
Mainstem
(d)
Tributaries
Figure 2. Biomass (mean percentage � SE) available and ingested in the diet for every fluvial system. (a) Winter, (b) spring, (c) summer and (d) autumn. Species are, from left toright, calandino, southern Iberian chub, southern Iberian spined-loach, pumpkinseed, largemouth bass and red swamp crayfish for every group of bars.
D. Almeida et al. / Animal Behaviour 84 (2012) 1475e14821480
predicts that the scent mark of an intruder will itself stimulatea response (e.g. increase marking) by the territory owners (Müller-Schwarze & Heckman 1980). According to the ‘scent-matching’hypothesis, both the presence of the intruding signaller and thechemical signal are necessary to provoke a response by the territoryowners (Gosling 1990). For other riverine mammals, such as theNorth American beaver, Castor canadensis, the marking behavioursupports the ‘scent-matching’ hypothesis (Sun & Müller-Schwarze1998). However, this hypothesis may not be applicable to a toppredator of freshwater environments, such as the otter, especiallyin the more variable Mediterranean ecosystems (Ruiz-Olmo et al.2002). Thus, the need for territory defence may be more intense(i.e. more suited to the ‘scent-fence’ hypothesis).
Prigioni et al. (2005) found a high positive relationship betweenfish availability and otter marking intensity. This finding indicatesthat otter marking behaviour reflects habitat quality regarding foodresources (e.g. Ben-David et al. 2005). Thus, as expected, we foundsupport for the hypothesis that latrine sites varied seasonally inrelation to the abundance of the main prey of otters, the calandino,particularly where environmental conditions are more unstable(i.e. in the tributaries). However, with reference to southern Iberianchub, the contrasting results between fluvial systems could berelated to this fish’s gregariousness during feeding, as well as itslarger size (around 20 cm total length). Thus, these fish areparticularly easy to detect in the more homogeneous mainstem(Almeida et al. 2012a). Therefore, these limnological and behav-ioural traits may incite otters to mark sites with higher southernIberian chub abundance. However, in the tributaries, this fishspecies can withstand the stronger currents for most of the year(e.g. Martínez-Capel et al. 2009) and therefore it may be located in
sites that otters tend to avoid marking as latrines, owing to reducedhunting quality (Kruuk et al. 1993).
Nevertheless, the seasonal and spatial patterns described abovewere not translated into prey selection. For instance, while otterspositively selected sites with more calandino for their latrines inthe tributaries during spring and summer, the consumption of thisspecies was below its availability in the mainstem. Again, a poten-tial explanation for this mismatch is that otters could be selectingplaces where fish can be confined more easily. The study by López-Martín et al. (1998) supports this, as pools were found to containlarger fish, with these habitats being preferentially selected byotters to optimize fishing effort (see also Almeida et al. 2012b).
The costebenefit balance is also valid to explain the comple-mentary foraging of otters on the two main prey species in thestudy area, calandino and red swamp crayfish. According to thepredictions of optimal foraging theory (Schoener 1971), a predatorshould concentrate its efforts on the preferred prey when it isabundant (or easy to catch), and it should search for alternativeprey when the main one is scarce (or difficult to catch). Otters arespecialist fish predators that become more generalist when habitatinstability increases (Jedrzejewska et al. 2001; Clavero et al. 2003,2008; Ruiz-Olmo & Jiménez 2009). Thus, they showa preference forfish in highly variable Mediterranean habitats, but when summerdrought makes fish scarcer, they change their foraging preferenceto crayfish (Delibes & Adrián 1987; Beja 1996; Román 2011), in spiteof its lower energy value in comparisonwith fish (Elvira et al. 1996).This behaviour has also been found in marine otters, which includecrustaceans in their diet only when fish capture rates become low(Watt 1991). There was a general tendency to feed on crayfishbelow its availability in winter, possibly because during this season

D. Almeida et al. / Animal Behaviour 84 (2012) 1475e1482 1481
red swamp crayfish tend to be inactive because of low tempera-tures (Beja 1996; Gherardi et al. 2000). However, we captured redswamp crayfish by electrofishing, a method found to have catchrates of >60% (Alonso 2001). It is therefore possible that our resultsare likely to be biased in winter.
A final remarkable finding from the dietary results relates to thepreference of otter for centrarchids (i.e. pumpkinseed and large-mouth bass) in the tributaries. These results contrast with thosedescribed by Blanco-Garrido et al. (2008) in the lower RiverGuadiana basin, since both species were rejected by otters probablybecause of the presence of antipredator body structures. However,the year-round ingested biomass in tributaries was negligible forboth species (ca. 1%). Consequently, the statistical significancefound is probably due to these species being almost absent intributaries. It is therefore possible that the prey found in spraintswere caught in other places (Almeida et al. 2012b).
In conclusion, the overall results highlight the importance ofmonitoring the otter’s marking strategy for the entire year to revealaccurate patterns of seasonal variation, with particular emphasis inhighly variable Mediterranean ecosystems. Moreover, we analysedotter marking behaviour at a catchment scale, which allowed thecomparison of the mainstem with near-pristine tributaries andshowed how flow regulation modulated the sprainting strategy.Also, prey selection was influenced by both endemic and invasivespecies. Finally, the study of latrine site selection and prey selectionprovided insights into the behavioural ecology of otters in relationto the suitability of habitat and food availability in strongly fluc-tuating (Mediterranean) environments.
Acknowledgments
D.A. held a postdoctoral fellowship from JCCM-FSE 2009/2011(PO 2007-13) through the University of Castilla-LaMancha. R.B. wassupported by a postdoctoral grant from the JCCM-FSE 2007/2013programme and FEDER funds. We thank J. Moya-Laraño for his helpwith the statistical analyses, I. Barja for improving a first version ofthe manuscript with her comments and G.H. Copp for editing thetext.
References
Almeida, D. 2008. Ecología y conservación de la fauna fluvial en el Parque Nacionalde Cabañeros. Efectos de la degradación del hábitat y de la introducción deespecies exóticas. Ph.D. thesis, Complutense University of Madrid.
Almeida, D., Almodóvar, A., Nicola, G. G. & Elvira, B. 2009. Feeding tactics and bodycondition of two introduced populations of pumpkinseed Lepomis gibbosus:taking advantages of human disturbances? Ecology of Freshwater Fish,18, 15e23.
Almeida, D., Merino-Aguirre, R. & Angeler, D. G. 2012a. Benthic invertebratecommunities in regulated Mediterranean streams and least-impactedtributaries. Limnologica, http://dx.doi.org/10.1016/j.limno.2012.02.003, pub-lished online 27 March 2012.
Almeida, D., Copp, G. H., Masson, L., Miranda, R., Murai, M. & Sayer, C. D. 2012b.Changes in the diet of a recovering Eurasian otter population between the 1970sand 2010. Aquatic Conservation: Marine and Freshwater Ecosystems, 22, 26e35.
Alonso, F. 2001. Efficiency of electrofishing as a sampling method for freshwatercrayfish populations in small creeks. Limnetica, 20, 59e72.
Arce, J. A. 2011. Empleo de excrementos de nutria Lutra lutra (Linnaeus, 1758) parala diagnosis específica de los cangrejos de río en la Península Ibérica. Galemys,23, 3e8.
Barja, I. 2009. Decision making in plant selection during the faecal-markingbehaviour of wild wolves. Animal Behaviour, 77, 489e493.
Barrientos,R.2006.Year-rounddefecationpattern inwildgenets (GenettagenettaL.) ina mountain forest (Toledo, Central Spain). Polish Journal of Ecology, 54, 325e328.
Barrientos, R. & Virgós, E. 2006. Reduction of potential food interference in twosympatric carnivores by sequential use of shared resources. Acta Oecologica, 30,107e116.
Barrientos, R., Gil, T., Hernando, J., Iglesis, A., Jiménez, M. D. & Navarro, E. 2003.Influencia de un embalse sobre la distribución y alimentación de la nutria (Lutralutra L.) en el río Riaza (Segovia). Galemys, 15, 81e90.
Beja, P. R. 1996. An analysis of otter Lutra lutra predation on introduced Americancrayfish Procambarus clarkii in Iberian streams. Journal of Applied Ecology, 33,1156e1170.
Ben-David, M., Blundell, G., Kern, J., Maier, J., Brown, E. & Jewett, S. 2005.Communications in river otters: creation of variable resource sheds forterrestrial communities. Ecology, 86, 1331e1345.
Blanco-Garrido, F., Prenda, J. & Narváez, M. 2008. Eurasian otter (Lutra lutra) dietand prey selection in Mediterranean streams invaded by centrarchid fishes.Biological Invasions, 10, 641e648.
Brown, R. E. & Macdonald, D. W. 1985. Social Odours in Mammals. Oxford: OxfordUniversity Press.
Clavero, M., Prenda, J. & Delibes, M. 2003. Trophic diversity of the otter (Lutra lutraL.) in temperate and Mediterranean freshwater habitats. Journal of Biogeog-raphy, 30, 761e769.
Clavero, M., Prenda, J., Blanco-Garrido, F. & Delibes, M. 2008. Hydrologicalstability and otter trophic diversity: a scale-insensitive pattern? CanadianJournal of Zoology, 86, 1152e1158.
Conroy, J. W. H., Watt, J., Webb, J. B. & Jones, A.1993. A Guide to the Identification ofPrey Remains in Otter Spraint. London: The Mammal Society.
Crait, J. R. & Ben-David, M. 2007. Effects of river otter activity on terrestrial plantsin trophically altered Yellowstone Lake. Ecology, 88, 1040e1052.
Delibes, M. & Adrián, I.1987. Effects of crayfish introduction on otter Lutra lutra foodin the Doñana National Park, SW Spain. Biological Conservation, 42, 153e159.
Delibes, M., Macdonald, S. M. & Mason, C. F. 1991. Seasonal marking, habitat andorganochlorine contamination in otters (Lutra lutra): a comparison betweencatchments in Andalucia and Wales. Mammalia, 55, 567e578.
DePue, J. E. & Ben-David, M. 2010. River otter latrine site selection in arid habitatsof Western Colorado, USA. Journal of Wildlife Management, 74, 1763e1767.
Dubuc, L. J., Krohn, W. B., Owen, J. & Ray, B. 1990. Predicting occurrence of riverotters by habitat on Mount Desert Island, Maine. Journal of Wildlife Manage-ment, 54, 594e599.
Elvira, B., Nicola, G. G. & Almodóvar, A. 1996. Pike and red swamp crayfish: a newcase on predatoreprey relationship between aliens in central Spain. Journal ofFish Biology, 48, 437e446.
Erlinge, S. 1968. Territoriality of the otter Lutra lutra L. Oikos, 19, 81e98.Espírito-Santo, C., Rosalino, L. M. & Santos-Reis, M. 2007. Factors affecting the
placement of common genet latrine sites in a Mediterranean landscape inPortugal. Journal of Mammalogy, 88, 201e207.
Gasith, A. & Resh, V. H. 1999. Streams in Mediterranean Climate Regions: abioticinfluences and biotic responses to predictable seasonal events. Annual Review ofEcology and Systematics, 30, 51e81.
Gherardi, F., Barbaresi, S. & Salvi, G. 2000. Spatial and temporal patterns in themovement of Procambarus clarkii, an invasive crayfish. Aquatic Sciences, 62,179e193.
Gittleman, J. L. & Harvey, P. H. 1982. Carnivore home-range size, metabolic needsand ecology. Behavioral Ecology and Sociobiology, 10, 57e63.
Gorman, M. L. 1990. Scent marking strategies in mammals. Revue Suisse de Zoologie,97, 3e29.
Gorman, M. L. & Trowbridge, B. J. 1989. The role of odor in the social lives ofcarnivores. In: Carnivore Behavior, Ecology and Evolution (Ed. by J. L. Gittleman),pp. 57e139. Ithaca: Cornell University Press.
Gosling, L. M. 1982. A reassessment of the function of scent marking in territories.Zeitschrift für Tierpsychologie, 60, 89e118.
Gosling, L. M. 1990. Scent marking by resource holders: alternative mechanisms foradvertising the costs of competition. In: Chemical Signals in Vertebrates V (Ed. byD. W. Macdonald, D. Müller-Schwarze & S. E. Natynczuk), pp. 315e328. Oxford:Oxford University Press.
Holm, S. 1979. A simple sequentially rejective Bonferroni test procedure. Scandi-navian Journal of Statistics, 6, 65e70.
Hutchings, M. R. & White, P. C. L. 2000. Mustelid scent-marking in managedecosystems: implications for population management. Mammal Review, 30,157e169.
Jedrzejewska, B., Sidorovich, V. E., Pikulik, M. M. & Jedrzejewski, W. 2001. Feedinghabits of the otter and the Americanmink in Bia1owie _za Primeval Forest (Poland)compared to other Eurasian populations. Ecography, 24, 165e180.
Johnson, R. P. 1973. Scent marking in mammals. Animal Behaviour, 21, 521e535.Jordan, N. R., Cherry, M. I. & Manser, M. B. 2007. Latrine distribution and patterns
of use by wild meerkats: implications for territory and mate defence. AnimalBehaviour, 73, 613e622.
Kruuk, H. 1992. Scent marking by otters (Lutra lutra): signalling the use ofresources. Behavioral Ecology, 3, 133e140.
Kruuk, H. 1995. Wild Otters: Predation and Populations. London: Academic Press.Kruuk, H., Wansink, D. & Moorhouse, A. 1990. Feeding patches and diving success
of otters (Lutra lutra L.) in Shetland. Oikos, 57, 68e72.Kruuk, H., Carss, D. N. & Durbin, L. 1993. Otter (Lutra lutra L.) numbers and fish
productivity in rivers in N.E. Scotland. Symposia of the Zoological Society ofLondon, 65, 171e191.
Lenton, E. J., Chanin, P. R. F. & Jefferies, D. J.1980. Otter Survey of England 1977e79.London: Nature Conservancy Council.
Lewis, M. A. & Moorcroft, P. 2001. ESS analysis of mechanistic models for terri-toriality: the value of scent marks in spatial resource partitioning. Journal ofTheoretical Biology, 210, 449e461.
López-Martín, J. M., Jiménez, J. & Ruíz-Olmo, J. 1998. Caracterización y uso delhábitat de la nutria Lutra lutra (Linné, 1758) en un río de carácter mediterráneo.Galemys, 10, 175e190.
Macdonald, D. W. 1980. Patterns of scent marking with urine and faecesamongst carnivore communities. Symposia of the Zoological Society of London,45, 107e139.

D. Almeida et al. / Animal Behaviour 84 (2012) 1475e14821482
Macdonald, D. W. 1983. The ecology of carnivore social behaviour. Nature, 301,379e384.
Macdonald, S. M. & Mason, C. F. 1987. Seasonal marking in an otter population.Acta Theriologica, 32, 449e462.
Martínez-Capel, F., García de Jalón, D., Werenitzky, D., Baeza, D. & Rodilla-Alamá, M. 2009. Microhabitat use by three endemic Iberian cyprinids inMediterranean rivers (Tagus River Basin, Spain). Fisheries Management andEcology, 16, 52e60.
Mason, C. F. & Macdonald, S. M. 1986. Otters, Conservation and Ecology. Cambridge:Cambridge University Press.
Miranda, R. & Escala, M. C. 2002. Guía de Identificación de Restos Óseos de losCiprínidos Presentes en España. Pamplona: Publicaciones de Biología de la Uni-versidad de Navarra.
Miranda, R. & Escala, M. C. 2007. On the use of cyprinid scales in the diet analysisof piscivorous species: how much information is hidden in a fish scale?Transactions of the American Fisheries Society, 136, 1008e1017.
Moya-Laraño, J. & Wise, D. W. 2007. Two simple strategies of analysis to increasethe power of experiments with multiple response variables. Basic and AppliedEcology, 8, 398e410.
Müller-Schwarze, D. & Heckman, S. 1980. The social role of scent marking inbeaver (Castor canadensis). Journal of Chemical Ecology, 6, 81e95.
Nilsson, C., Reidy, C. A., Dynesius, M. & Revenga, C. 2005. Fragmentation and flowregulation of the world’s large river systems. Science, 308, 405e408.
Perez-Quintero, J. C. 2007. Diversity, habitat use and conservation of freshwatermolluscs in the lower Guadiana River basin (SW Iberian Peninsula). AquaticConservation, 17, 485e501.
Platts, W. S., Megahan, W. F. & Minshall, W. G. 1983. Methods for evaluatingstream, riparian, and biotic conditions. United States Department of Agriculture,Forest Service, Intermountain Forest and Range Experiment Station Ogden, GeneralTechnical Report INT, 138, 1e70.
Prenda, J. & Granado-Lorencio, C. 1996. The relative influence of riparian habitatstructure and fish availability on otter Lutra lutra L. sprainting activity in a smallMediterranean catchment. Biological Conservation, 76, 9e15.
Prigioni, C., Remonti, L., Balestrieri, A., Sgrosso, S., Priore, G., Misin, C.,Viapiana, M., Spada, S. & Anania, R. 2005. Distribution and sprainting activityof the otter (Lutra lutra) in the Pollino National Park (southern Italy). EthologyEcology & Evolution, 17, 171e180.
R Development Core Team. 2009. R: A Language and Environment for StatisticalComputing. Vienna: R Foundation for Statistical Computing. http://www.R-project.org.
Ralls, K. 1971. Mammalian scent marking. Science, 171, 443e449.Román, J. 2011. What do otters eat where there is no fish? Mammalian Biology, 76,
237e239.
Roper, T. J., Conradt, L., Butler, J., Christian, S. E., Ostler, J. & Schmid, T. K. 1993.Territorial marking with faeces in badgers (Meles meles): a comparison ofboundary and hinterland latrine use. Behaviour, 127, 289e307.
Rostain,R.R., Ben-David,M.,Groves,P.&Randall, J.A.2004.Whydoriverotters scent-mark? An experimental test of several hypotheses. Animal Behaviour, 68, 703e711.
Ruiz-Olmo, J. & Gosálbez, J. 1997. Observations on the sprainting behaviour of theotter Lutra lutra in the NE Spain. Acta Theriologica, 42, 259e270.
Ruiz-Olmo, J. & Jiménez, J. 2009. Diet diversity and breeding of top predators aredetermined by habitat stability and structure: a case study with the Eurasianotter (Lutra lutra L.). European Journal of Wildlife Research, 55, 133e144.
Ruiz-Olmo, J., López-Martín, J. M. & Palazón, S. 2001. The influence of fishabundance on the otter (Lutra lutra) populations in Iberian Mediterraneanhabitats. Journal of Zoology, 254, 325e336.
Ruiz-Olmo, J., Olmo-Vidal, J. M., Mañas, F. & Batet, A. 2002. Influence of resourceseasonality on the breeding patterns of the Eurasian otter (Lutra lutra) inMediterranean habitats. Canadian Journal of Zoology, 80, 2178e2189.
Ruiz-Olmo, J., Margalida, A. & Batet, A. 2005a. Use of small rich patches byEurasian otter (Lutra lutra L.) females and cubs during the pre-dispersal period.Journal of Zoology, 265, 339e346.
Ruiz-Olmo, J., Batet, A. & Pérez, M. S. 2005b. Comparación de la conducta demarcaje de la nutria (Lutra lutra L.) en cautividad durante el periodo de ges-tación y cuidado de las crías y fuera de éste. Galemys, 17, 53e60.
Ruiz-Olmo, J., Jiménez, J. & Chacon, W. 2007. The importance of ponds for theotter (Lutra lutra) during drought periods in Mediterranean ecosystems: a casestudy in Bergantes River. Mammalia, 71, 16e24.
Schoener, T. W. 1971. On the theory of feeding strategies. Annual Review of Ecologyand Systematics, 2, 369e404.
Statzner, B., Gore, J. A. & Resh, V. 1988. Hydraulic stream ecology: observedpatterns and potential applications. Journal of the North American BenthologicalSociety, 7, 307e360.
Sun, L. & Müller-Schwarze, D. 1998. Beaver response to recurrent alien scents:scent fence or scent match? Animal Behaviour, 55, 1529e1536.
Swimley, T. J., Serfass, T. L., Brooks, R. P. & Tzilkowski, W. M.1998. Predicting riverotter latrine sites in Pennsylvania. Wildlife Society Bulletin, 26, 836e845.
Wang, L., Infante, D., Lyons, J., Stewart, J. & Cooper, A. 2011. Effects of dams inriver networks on fish assemblages in non-impoundment sections of rivers inMichigan and Wisconsin, USA. River Research and Applications, 27, 473e487.
Warton, D. I. & Hui, F. K. C. 2011. The arcsine is asinine: the analysis of proportionsin ecology. Ecology, 92, 3e10.
Watt, J. 1991. Prey selection by coastal otters (Lutra lutra L.). Ph.D. thesis, Universityof Aberdeen.
Zippin, C. 1956. An evaluation of the removal method of estimating animal pop-ulation. Biometrics, 12, 163e189.