the role of attention in the preparation of visually guided eye movements in monkey and man
TRANSCRIPT

Psychol Res (1986) 48:251-257 Psychological Research © Springer-Verlag 1986
The role of attention in the preparation of visually guided eye movements in monkey and man
B. Fischer
Department of Clinical Neurology and Neurophysiology, University of Freiburg, Hansastr. 9, D-7800 Freiburg, Federal Republic of Germany
Summary. Neurophysiological data from single cells in the monkey's visual association cortex as well as saccadic reaction time measurements in monkey and man are re- ported. When a monkey directs his attention to a peripher- al light stimulus the visual activation of cortical cells re- sponsive to that stimulus is increased. If visual attention is directed to a particular part of the visual field the saccadic reaction time is long (around 200 ms in monkey and above 200 ms in man). In contrast, if attention is disengaged from any location to which it might have been engaged be- fore, monkeys and men can execute express saccades, that is, saccades after extremely short and stable reaction times (around 75 ms in monkey and 100 ms in man). The results are interpreted in the context of a hypothesis according to which the initiation of voluntary, visually guided saccades includes three premotor mechanisms: the computation of the target location, the decision to make a saccade, and the disengagement of attention.
Introduction
Being interested in the neural operations preceding visual- ly guided eye movement, one constantly comes up against the problem of attention, since gaze is usually directed toward objects of attention or interest. For this reason the direction of gaze and the direction of attention are, most of the time, coincident in space. However, as everybody knows from introspection, those two directions can be dis- sociated by one's own decision or by instruction.
In the research described here we use two approaches. We attempt to demonstrate effects of attention in the dis- charge pattern of single neurons. "Attention" is here taken to be an intrinsic operation of the brain that is not directly related to any change in sensory stimulation, nor to any motor response. We therefore looked for an experimental situation in which an animal has to deal with a (foveal) central stimulus during one period of time and then has to deal with another stimulus continuously visible in the peri- phery in a subsequent period of time, without producing any motor response of the limbs or of the eyes.
The other approach makes use of reaction times. We assume that any neural operation takes a certain amount of time from its initiation to its completion. This time will be included in the motor reaction time following some ex- ternal command signal if the neural operation in question
is initiated by the command to execute the movement. It will not be included in the overall motor response time if the situation does not require this operation or if the situa- tion allows the operation in question to be completed be- fore the final command for the reaction is given.
In using the latter approach we took advantage of the existence of a class of saccades that are characterized by their extremely short and stable reaction times. These "ex- press saccades" have been found in monkey (Fischer & Boch, 1983) as well as in man (Fischer & Ramsperger, 1984). They have reaction times of around 75 ms and 100 ms, respectively.
The results described below will show that the instruc- tion "pay attention" versus "do not pay attention" drasti- cally alters the probability of occurrence of express sac- cades. The details of the conditions under which this change occurs lead us to the conclusion that spatially di- rected visual attention inhibits goal directed saccades. At- tention has to be disengaged from any particular spatial location for the oculomotor system to be released from this inhibition, and thus to enable the next saccade. The time needed for the disengagement of attention either is or is not included in the saccadic reaction time, depending on the attentional instruction as well as on the relative timing of the offset of the attentional stimulus and onset of the target for the next saccade.
General methods
For t~oth monkey and man, visual stimuli were back pro- jected on a large, translucent hemispheric screen 57 cm from the eyes. Two paradigms were used:
(1) Gap: a central fixation point was presented which the subject had to fixate during the first part of each trial. The fixation point was then turned off and only after a temporal gap (usually fixed at 200 ms) a peripheral target was presented (Figure 1). Subjects were instructed (verbal- ly or by use of the dimming, see below) to make saccades to the new target.
(2) Overlap: the paradigm was identical to the gap par- adigm except that the fixation point remained continuous- ly illuminated (Figure 1).
The target location was randomized from trial to trial between 4 degrees to the right and left, or, in another con- dition, it was kept constant. Usually the time from the be- ginning of a trial to the occurrence of the target was con-

252
GAP
I " 'i i
E M ~ 1 1 1
F
T
EM'
OVERLAP
I i I I
i !GQp, SRT, ' / /
/ - -
2 s SRT i
Fig. 1. Time course of visual stimulation in the Gap and in the Overlap paradigm. The upper trace (F) indicates the time course of the fixation point, which was later also used as a target for peri- pheral attention. The middle trace (T) indicates the time course of the saccade target. The lower trace (EM) indicates eye movements. Saccadic reaction times (SRT)are measured from target onset
stant (2 s) but for control this time was also randomized between 0.5 and 3.5 s in the study of human saccades.
The luminance of the fixation point and of the target was about 0.8 log units above threshold in relation to the background luminance of less than 0.5 cd per square me- ter.
In both paradigms (gap and overlap) saccadic reaction times were determined by an automatic movement detec- tor which stopped a millisecond counter triggered by the onset of the target. Further experimental details are de- scribed in the appropriate sections of the results.
R e s u l t s
Neurophysiological recording: reactivation of prelunate cortical cells
This section deals with activation of cells in the visual as- sociation cortex located on the prelunate gyrus well above the tip of the inferior occipital sulcus and posterior to the superior temporal sulcus. Cells in this cortical area repre- sent parafoveal vision in the lower quadrant of the contra- lateral visual field (Fischer & Boch, 1981 a, b). They re- spond to the onset of a small light stimulus in their recep- tive field while the animal is fixating another spot of light. The on-response has a latency of about 80 ms from stimu- lus onset (Boch & Fischer, 1983).
Monkeys (macaca mulatta) were placed in a primate chair with their heads fixed by a permanently implanted metal bar. Eye movements were continuously monitored by an infrared light method (Bach, Bouis, & Fischer, 1983). Single unit activity was recorded by a glass insulat- ed Pt-Ir microelectrode inserted through the intact dura. There were two tasks which were trained, and run, sepa- rately in blocks of trials.
(1) In the fixation task (Figure 2A) the monkey had to press a key with his hand in response to the illumination of a small spot. He had to hold the key down for a randomly varying period of time, after which the luminance of the spot was slightly dimmed. In response to the dimming the
monkey had to release the key within a minimum reaction time of 700 ms to receive a reward. Since the dimming was made difficult to detect, the monkey had to use his foveal vision. Therefore, the small spot will be called "fixation point". During the fixation task other stimuli could be pre- sented in the periphery of the monkey's visual field as in- dicated by the trace labeled S in Figure 2A. Up to 20 identical trials were collected for averaging purposes.
(2) The attention task (Fig. 2 B) started in the same way as the fixation task, but a second stimulus appeared in the periphery at a fixed time (1 or 2 s) after the beginning of each trial and the animal was required to still fixate the central fixation point. After another second the fixation point disappeared. The animal was then free to foveate the new stimulus or to suppress the eye movement. On those trials on which the fixation point had already gone off, it was the dimming of the peripheral stimulus that the animal had to detect. Thus, on those trials on which the monkey did not make a saccade he had to detect the dimming of the stimulus using peripheral vision. In the attention task therefore two types of trials could occur. One in which the monkey made no saccade and the other in which the ani- mal foveated the parafoveal stimulus.
The fixation task was used to test the cells' visual re- sponse properties and to map their receptive fields. The at- tention task was used to find extra-retinal modulations of the cells' impulse activity.
I f the fixation point disappears in the middle of a trial (after termination of the on-response to the peripheral sti- mulus) these cells are activated again even though the monkey makes no eye movement to the stimulus. This reactivation is schematically illustrated in Figure3. The activity occurs about 180 to 200 ms after fixation point off- set and lasts as long as the fixation point remains invisible, unless the monkey makes an eye movement. The eye movement of course terminates the cell's activity because its receptive field no longer coincides with the stimulus. It should be pointed out that the receptive fields of these cells do not include the fovea. They respond to the onset of light, but not the offset. The offset of the fixation point does not change the cell's spontaneous impulse activity if the peripheral stimulus is placed outside its receptive field, for example in the opposite quadrant of the visual field. More details and further aspects of the properties of cells
A FIXATION
F I , dim 1 _ _
s I ]
E M
Key I
B F
S
EM
ATTENTION I I s ',
L I I t
I _ _ Reward
dim i
Key I I _ _ Reward
Fig. 2. Behavioural paradigm used to "train monkeys for central fi- xation (panel A) and peripheral attention (panel B). F Fixation- point; S Stimulus; EM Eye movement

Modulation of the visual response by attention
~ O N
Fixp OFFI
Eye movement
I IIIIIII I i . i . i i i
neural discharges
receptive fietd
S t i m u t u s
IIIIIIIII IIIIIIIIIIIIII [ I I I I I I I I I I I I I I I I I I I I I I I
Attention !
visual _ / ~ response
Association Cortex
Fig. 3. Schematic illustration of the activation of a single neuron in the monkey's visual association cortex (V4, prelunate gyrus). In response to the onset of a stimulus in the cell's receptive field (lower left inset and upper trace) the cell is activated. After offset of the fixation point (second trace) the cell is activated again, even though the monkey does not initiate a saccade to the stimulus but rather keeps the direction of gaze in the center (third trace). The lower right diagram indicates that these cells have access to visual information which can be modulated by attention
253
n I
A
B
express~ GAP m fast
100 2()0 300ms
OVERLAP
s• stow
expre regular
160 200 300ms- Saccadic Reaction Time
Fig. 4. Schematic summary of the distribution of the monkey's saccadic reaction times. A In the gap paradigm the distribution is clearly bimodal with the first peak around 75 ms (labeled express) and the second around 140 ms (labeled fast regular). B In the overlap paradigm well trained monkeys also produce a bimodal distribution. The first peak represents the express saccades and the second occurs at above 200 ms (labeled slow regular)
in the prelunate cortex are publ ished by Fischer & Bach (1985).
These observat ions strongly suggest that input from the ret ina to the visual associat ion cortex can be modu la t ed by an extra-ret inal mechanism that becomes effective when the animal has no f ixat ion point. Since the react ivat ion is related neither to any physical change of light in the cells ' receptive field nor to a motor response, and since the ani- mal correctly detects the d imming of the parafovea l stimu- lus at the end of the trial, it seems reasonable to assume that the state of excitat ion o f the visual associat ion cortex is modula ted by directed visual attention.
Express saccades in monkey and man
In an a t tempt to demonst ra te behavioura l correlates of the two different states of cort ical activity - inactive vs reacti- vated visual associat ion cortex - , we used a task in which the monkey was required to make a goal -di rec ted saccade 200 ms after f ixation poin t offset (gap pa rad igm as de- scr ibed above). The cor responding saccadic react ion t ime (SRT) was then compared to those of saccades which the monkey made to the same target when the f ixat ion poin t remained visible (overlap paradigm).
The results are summar ized in Figure. 4. Using the gap parad igm, the dis t r ibut ion of the SRTs is clearly b imodal . The first peak is rather nar row and has its max imum at about 75 ms under these condit ions. The size of the peak and its exact locat ion depends on the amount of previous t ra ining (Fischer, Bach, & Ramsperger , 1984) as well as on the size, luminance, and eccentrici ty of the saccade target (Bach, Fischer, & Ramsperger , 1984).
The second peak, labeled 'fast regular ' in Figure 4A, occurs at about 140 ms. These are also rather short reac- t ion times as compared to those ob ta ined in the over lap paradigm.
In the over lap pa rad igm (F igu re4B) monkeys also p roduce a b imoda l dis t r ibut ion of SRTs. The first peak re- sembles the express saccades (with a mode a round 90 ms) and the second occurs at about 220 ms labeled slow regu-
lar. Compar i son of Figure 4 A with 4 B shows that the sac- cadic react ion times can have at least three clearly differ- ent moda l values, each, of course, with a certain amount of scatter.
These results have a number of impl icat ions for an un- ders tanding of the processes preceding a visually guided saccade that will be discussed below. An impor tan t f inding is that express saccades also occur in the over lap para- digm, where the f ixat ion poin t remains visible and where - unl ike the gap parad igm - there is no sensory event that could indicate to the animal that the target is about to ap- pear.
The phenomenon of express saccades was first report- ed in monkey by Fischer and Bach (1983). At that t ime there was no indica t ion that a s imilar phe nome non existed in man. Saslow (1967) used the gap pa rad igm but he d id not repor t react ion times be low 130 ms nor did he notice a bi- moda l i ty in the dis t r ibut ion of the SRTs. We have repeated Saslow's exper iment and found that in man there exists a first group of saccades at about 100 ms and a second group with react ion t imes of about 150 ms (Fischer & Ramsperg- er, 1984). These two groups occur in the gap si tuation. Us- ing the over lap parad igm, saccadic react ion t imes are in the order of 220 ms or even more. In conclus ion: monkeys as well as human subjects p roduce at least three different values of their saccadic react ion times.
However , we did not observe in man express saccades in the over lap pa rad igm as long as the subject 's task was to fixate the f ixat ion poin t at tentively until the saccade target appeared . In an a t tempt to obta in express saccades in the over lap pa rad igm we told the subjects not to pay at tent ion to the f ixation poin t but rather, to keep their direct ion of gaze on it. This change in the instruction immedia te ly changed the dis t r ibut ion of the react ion times, which now showed a clear peak at about 100 ms in addi t ion to the sec- ond peak at about 220 ms. The size of the express peak de- p e n d e d on the amount of pract ice the subject previously had.
This observat ion has led to the question, what are the necessary a n d / o r sufficient condi t ions under which ex-
254
N I00'
90
80
70'
6O
50
4Cr
3O
20
10
N 100-
90-
80-
70-
60"
50-
40-
30-
20-
10-
N 100-
90"
80"
70"
60"
50"
40'
30"
20"
10" 5
A
B
Overlap peripherat attention
100 200
Gap
300 400 ms
N 40
36 A 32 28
2~
20
16
12
N40-
36" 32" E~
28-
2/...
20-
16-
12.
8-
4-
N40"
36"
32"
28"
24"
20"
16"
12"
8"
C
P ripheral attention
100 200 300 400 ms'
Overlap peripheral attention
G+ ~ @ T A
100 200 300 40(3 ms
Overtap central fixation
100 2013 300 4~ ms 100 20(] 313(} 400 ms
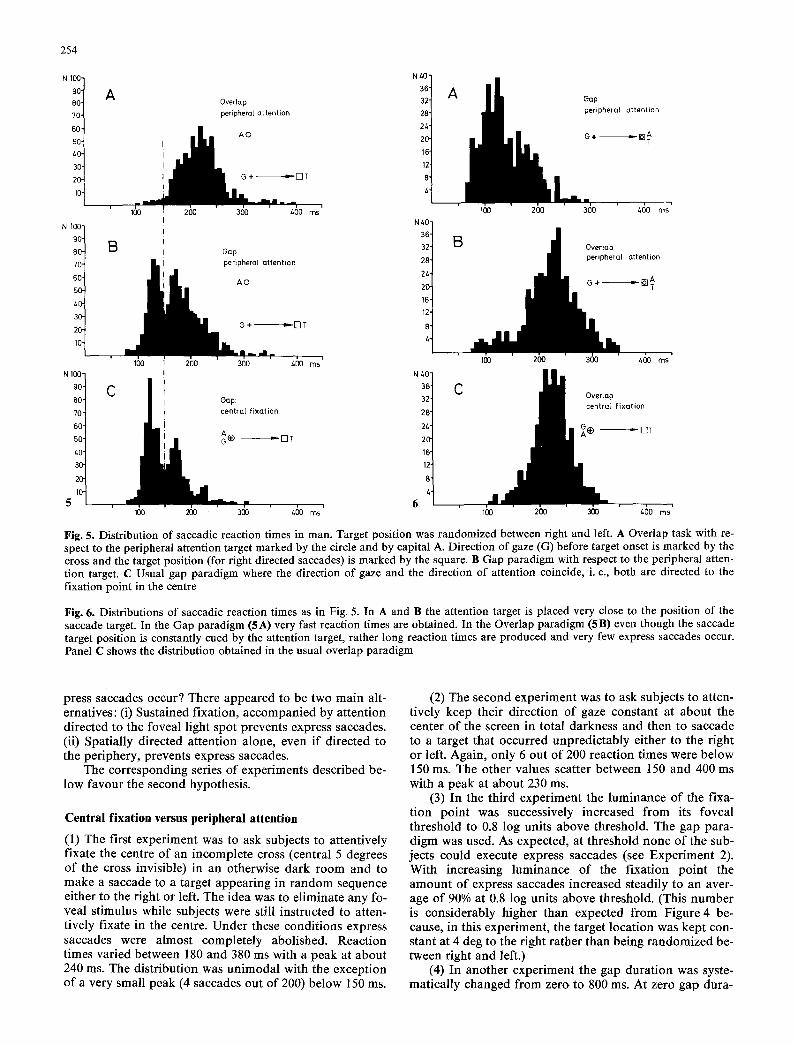
Fig. 5. Distribution of saccadic reaction times in man. Target position was randomized between right and left. A Overlap task with re- spect to the peripheral attention target marked by the circle and by capital A. Direction of gaze (G) before target onset is marked by the cross and the target position (for right directed saccades) is marked by the square. B Gap paradigm with respect to the peripheral atten- tion target. C Usual gap paradigm where the direction of gaze and the direction of attention coincide, i. e., both are directed to the fixation point in the centre
Fig. 6. Distributions of saccadic reaction times as in Fig. 5. In A and B the attention target is placed very close to the position of the saccade target. In the Gap paradigm (5A) very fast reaction times are obtained. In the Overlap paradigm (5B) even though the saccade target position is constantly cued by the attention target, rather long reaction times are produced and very few express saccades occur. Panel C shows the distribution obtained in the usual overlap paradigm
press saccades occur? There appeared to be two main alt- ernatives: (i) Sustained fixation, accompan ied by at tent ion directed to the foveal light spot prevents express saccades. (ii) Spat ial ly directed at tention alone, even if directed to the per iphery, prevents express saccades.
The corresponding series of experiments descr ibed be- low favour the second hypothesis.
Central fixation versus peripheral attention
(1) The first exper iment was to ask subjects to at tentively fixate the centre of an incomple te cross (central 5 degrees o f the cross invisible) in an otherwise dark room and to make a saccade to a target appear ing in r andom sequence either to the right or left. The idea was to el iminate any fo- veal stimulus while subjects were still instructed to atten- t ively fixate in the centre. Under these condi t ions express saccades were almost complete ly abolished. React ion times varied between 180 and 380 ms with a peak at about 240 ms. The dis tr ibut ion was un imoda l with the exception o f a very small peak (4 saccades out o f 200) below 150 ms.
(2) The second exper iment was to ask subjects to atten- t ively keep their direct ion of gaze constant at about the center of the screen in total darkness and then to saccade to a target that occurred unpred ic tab ly either to the right or left. Again, only 6 out of 200 react ion t imes were below 150 ms. The other values scatter between 150 and 400 ms with a peak at about 230 ms.
(3) In the third exper iment the luminance of the fixa- t ion poin t was successively increased from its foveal threshold to 0.8 log units above threshold. The gap para- d igm was used. As expected, at threshold none of the sub- jects could execute express saccades (see Exper iment 2). With increasing luminance o f the f ixat ion po in t the amount of express saccades increased steadily to an aver- age o f 90% at 0.8 log units above threshold. (This number is cons iderably higher than expected from Figure 4 be- cause, in this experiment , the target locat ion was kept con- stant at 4 deg to the right ra ther than being randomized be- tween right and left.)
(4) In another exper iment the gap dura t ion was syste- mat ica l ly changed from zero to 800 ms. At zero gap dura-

255
tion no express saccades occurred (as expected). With in- creasing gap durations the average proportion of express saccades increased rapidly to 75% at gap = 200 ms and then slowly decreased to 20% at gap = 800 ms. When gap durations of more than 1000 ms were used none of the sub- jects could make express saccades any more, in agreement with Experiment 2, above.
We concluded from these results that neither foveal sti- mulation nor central fixation could explain the absence of express saccades. To see whether or not spatially directed attention alone (Hypothesis 2 above) could explain the ab- sence of express saccades, two more experiments were conducted:
(5) The spot that served as a fixation point in the earli- er experiments was used as a target for peripheral atten- tion. It was moved 5 degrees up the vertical meridian and subjects were instructed to dissociate their direction of gaze (keep it in the center of the screen) from their direc- tion of attention (pay attention to the spot). Both the gap and overlap paradigm were used again. When the peri- pheral attention target remained visible after the saccade target appeared (overlap), the distribution of saccadic reaction times was unimodal, ranging from about 150 ms to 350ms with only a few saccades between 100 and 150 ms (Figure 5A). If, however, the gap paradigm was used, the distribution turned bimodal with a large peak be- tween 100 and 150ms (Figure 5B). For comparison, the gap paradigm was used again with the fixation point in the center of the screen (Figure 5 C). The dissimilarity of the distributions in Figures 5 A and 5 B and the similarity of 5 B and 5 C clearly indicate that peripheral attention sup- pressed the execution of express saccades unless the atten- tion target was extinguished shortly (200 ms) before target onset.
This still leaves open the question what would happen if attention were already directed to the target position at the time the target appears.
(6) To find out, we placed the attention target just (1/10 deg) above the saccade target, both at 4 degrees to the right of the center of the screen, and used the gap and the overlap paradigm again. The results are shown in Fi- gure 6. The distribution shown in Figure 6 A was obtained with the gap paradigm. A large peak occurs between 100 and 140 ms which seems to be preceded by still another peak. This early peak (at about 80 ms) is due to anticipato- ry saccades, because in this experiment the subject is con- stantly cued to the target position, a fact which cannot be avoided under these circumstances. In Figure 6 B the over- lap paradigm was used. To our surprise, the number of saccades with reaction times below 150 ms was greatly re- duced. Instead, the distribution peaked at about 240 ms. In fact, it was almost identical to the distribution in Fig. 6C, which was obtained in the usual overlap paradigm with the fixation point in the center of the screen.
Conclusion
Attention directed to any part of the visual field, including the fovea and even including the position for the saccade target, suppresses the execution of express saccades. It seems that directed attention inhibits saccades altogether, as long as attention remains continuously directed to any given part of the visual field. In order to initiate a visually guided saccade, attention must be interrupted (disengaged,
released, defocused or whatever term one wants to use). This disengagement of directed attention is a necessary but not a sufficient condition for a saccade. The disengage- ment takes time to be completed. It is this time which in- creases the reaction times obtained in the overlap para- digm, and which is not included in the reaction times ob- tained in the gap paradigm, because this time can elapse within the duration of the gap.
Discussion
Since we have identified three different groups of saccadic reaction times (both in monkey and man), it seems reason- able to assume that the preparation of a goal directed sac- cade consists of (at least) three operations, each one hav- ing a certain time cost. These three operations may be identified by taking into account three facts: (1) Saccades usually hit the target with an accuracy better than 10% of the saccade distance, which implies that targets are usually localized correctly. Therefore, we assume that one of the operations common to all three SRTs has to do with the computation (C) of the localization of the saccade target. (2) Saccades are voluntary movements that need a decision to be initiated. To make the decision (D) may take more time. (3) Saccades are delayed (in comparison to fast regu- lar and express saccades), if attention is engaged at a spec- ified part of the visual field. To disengage attention (A), another time is needed. Figure 7 shows how these opera- tions may be linked to each other, to the visual input (V), and to the oculomotor output (M).
Each circle in Fig. 7 (except the one which symbolizes the eye) represents a time-consuming operation. Each con- tinuous line sends a READY/NOT READY signal in the direction of the arrow. Each operation is initiated only if all incoming lines have sent their READY signals. This implies that each circle represents also a logical AND- gate. The quantitative results of each operation are made available and can be sent to other places along other lines (not shown in the diagram) only after termination of the operation in question. One feature of such an arrangement is that the operations A, D, and C must take place in se- quence. For example, D cannot be initiated before A is completed even if V has sent its READY signal simultane- ously to C, D, and A. Under these conditions the time costs of A, D, and C are additive. In another condition (e.g., gap task) the offset of the fixation point issues a READY signal to D and A but not to C because C re-
eye
Fig. 7. Hypothetical schematic diagram showing the three differ- ent loops and their connections that may play a role in the prepa- ration and initiation of visually guided saccades. V afferent visu- al; C computation of saccade size and direction; M oculomotor; D decision making; A engagement and disengagement of atten- tion

256
quires the information of target location. In this case, A and D can be completed well before C.
The different reaction times are now explained in the following way: When measured from target onset, the reaction time of the slow regular saccades include all pro- cesses A, D, and C in addition to afferent and efferent de- lays. The fast regular saccades are preceded by D and C. The express saccades are preceded by C only. Note: in the case of an express saccade, processes A and D have been completed before the onset of the saccade target. The need for three processes (A, D, and C) follows from the exis- tence of three different groups of saccadic reaction times.
The additional lines from D to M and A to M over-ex- plain the initiation of the saccade from a logical point of view, because the READY signal from C to M alone would be sufficient. However, from an experimental point of view, we know that visually guided saccades can be me- diated by at least two independent pathways, one includ- ing the superior colliculus, the other including the frontal eye fields (Schiller, True, & Conway 1980). Therefore, at least two of the processes must be directly connected to the oculomotor centers in the brain stem. One of them must be C (computation), because it is the last one in the series (Becker & Juergens 1979). The second can hardly be A (at- tention), because the disengagement of attention does not necessarily trigger a saccade. Therefore, one is left with D (decision). (This argument by itself may be weak. It is, however, supported by the observation of some properties of neurons in the frontal eye fields reported by Bruce and Goldberg (1984). They showed that frontal eye field cells can be activated before purposive saccades which the monkey makes at will, whereas the same cells are not ac- tive before spontaneous saccades made in the dark.) Nevertheless, A (attention) may also directly project to the brain stem (M) in such a way that during engaged atten- tion saccades are inhibited, but during disengaged atten- tion they are allowed.
It is unclear at present which neural structure mediates the attentional effects. Most likely the parietal cortex (A7) is involved but it may well be that the visual complex V4 of the prelunate gyrus is part of the attentional system as well.
Figure 8 schematically shows how, under natural view- ing conditions, the transition of engaged to disengaged vis-
I
eye posi t ion
A t t e n t i o n i move
' i I engaged I disengaged .
I IIIL ,
Illl t Loca l i za t i on . . . . . /
Iden t i f i ca t ion . . . . . . . . . . 7 , [
engaged
Ident i f i ca t ion
Fig. 8. Schematic drawing of a sequence of two saccades and the proposed sequence of engagement - disengagement of attention. The continuous heavy line indicates eye position over time. After a saccade, attention is assumed to be engaged at the new fixated location, from which information about stimulus features is pro- vided to an appropriate identification system. Later - in the mid- dle of a fixation pause - attention is disengaged. During this time information about the localization of peripheral stimuli is pro- vided to the corresponding system. Only then can the eyes and/or attention be moved to a new location
ual attention may take place in the middle of a 'fixation pause' between two saccades. After completion of the first saccade, attention is engaged on the foveal target at which the eye has just arrived. During this period visual informa- tion is accessable for a system which then identifies ob- jects. While the identification still goes on, attention may (but need not necessarily) be disengaged. Peripheral visual information is fed into a localization system, which then provides its information to the motor system, including the saccade system. The eye is then able to move or not, de- pending on whether or not a corresponding decision was made. If the eye does not move (dotted line in Figure 8), attention can still be disengaged, moved, and engaged again at a new position, resulting in a situation of peri- pheral attention. One could also express this notion by saying that a fully programmed but unexecuted saccade re- sults in directed visual peripheral attention.
The diagrams of Figures 7 and 8 are based almost ex- clusively on data from saccadic reaction time studies. However, there is plenty of support from anatomical and physiological data as well. The shortest loop from the reti- na to the oculomotor centers in the brain stem includes the superior colliculus, which therefore may be involved in process C. The superior colliculus receives information about target location in space (Sparks & Mays, 1983; Sparks & Porter, 1983) and its visual activity preceding saccades is enhanced if the visual stimulus is the target (Wurtz & Albano 1980). If the superior colliculus is de- stroyed, express saccades are abolished, but express sac- cades can be made without the frontal eye fields (Sandell, Schiller, & Maunsell, 1984).
The second loop from the retina to the oculomotor centers includes the frontal eye fields. They may be in- volved in process D, because the frontal eye fields are acti- vated before purposive saccades even in the absence of a visual target (Bruce & Goldberg, 1984) and they also re- ceive information about target location (Goldberg & Bruce, 1981). For the interaction between the frontal eye fields and the superior colliculus it is of particular interest that patients with frontal lobe lesions have difficulties in suppressing reflexive glances and produce express sac- cades (Guitton, Buchtel, & Douglas, 1985).
The third loop contains the parietal cortex. It is well known that the posterior parietal cortex is involved in vis- ual attention (Lynch, Mountcastle, Talbot & Yin, 1977; Robinson, Goldberg, & Stanton, 1978; Wurtz, Goldberg, & Robinson, 1980; Bushnell, Goldberg, & Robinson, 1981). Its contribution to the initiation of saccades is not direct, because electrical stimulation of posterior parietal cortex leads to eye movements only if the animal is not at- tentively fixating (Shibutani, Sakata, & Hyv/irinen, 1984).
More specifically, the work of Posner in man suggests that parietal cortex provides a mechanism for covert orien- ting of attention (Posner, Walker, Friedrich, & Rafal, 1984). Posner in fact arrived at a similar conclusion with respect to the attention sequence of engage - disengage - move. Posner reported that patients with parietal lesions have difficulties in directing their attention to the side con- tralateral to the lesion not because of a peripheral deficit in the visual field but because of an inability to withdraw (disengage) their attention from the center. Our results support this notion strongly and add that spatially directed visual attention (engaged attention at a peripheral loca- tion) inhibits saccades.

257
Acknowledgements. This work was supported by the Deutsche Forschungsgemeinschaft, Sonderforschungsbereich "Hirnfor- schung und Sinnesphysiologie (SFB 70, Tp B7).
References
Bach, M., Bouis, D., & Fischer, B. (1983). An accurate and linear infrared oculometer. Journal of Neuroscienee Methods, 9, 9-14.
Becker, W., & Juergens, R. (1979). An analysis of the saccadic system by means of double step stimuli. Vision Research, 19, 967-983.
Boch, R., & Fischer, B. (1983). Saccadic reaction times and activa- tion of prelunate cortex: Parallel observations in trained rhe- sus monkeys. Experimental Brain Research, 50, 201-210.
Boch, R., Fischer, B., & Ramsperger, E. (1984). Express-saccades of the monkey: Reaction times versus intensity, size, duration, and eccentricity of their targets. Experimental Brain Research, 55, 223- 231.
Bruce, C. J., & Goldberg, M. E. (1984). Physiology of the frontal eye fields. Trends in Neuroseiences, 7, 436-441.
Bushnell, M. C., Goldberg, M. E., & Robinson, D. L. (1981). Be- havioral enhancement of visual responses in monkey cerebral cortex. I. Modulation in posterior parietal cortex related to se- lective visual attention. Journal of Neurophysiology, 46, 755-772.
Fischer, B., & Boch, R. (1981 a). Enhanced activation of neurons in prelunate cortex before visually guided saccades of trained rhesus monkeys. Experimental Brain Research, 44, 129-137.
Fischer, B., & Boch, R. (1981b). Selection of visual targets acti- vates prelunate cortical cells in trained rhesus monkeys. Experimental Brain Research, 41, 431-433.
Fischer, B., &Boch, R. (1983). Saccadic eye movements after ex- tremely short reaction times in the monkey. Brain Research, 260, 21-26.
Fischer, B., &Boch, R. (1985). Peripheral attention versus central fixation: Modulation of the visual activity of prelunate corti- cal cells of the rhesus monkey. Brain Research, 345, 111-123.
Fischer, B., Boch, R., & Ramsperger, E. (1984). Express-saccades of the monkey: Effects of daily training on probability of oc- currence and reaction time. Experimental Brain Research, 55, 232-242.
Fischer, B., & Ramsperger, E. (1984). Human express-saccades: extremely short reaction times of goal directed eye move- ments. Experimental Brain Research, 57, 191-195.
Goldberg, M. E., & Bruce, C. J. (1981). Frontal eye fields in the monkey: Eye movements remap the effective coordinates of visual stimuli. Neuroscience Abstracts, 44, 4-131.
Guitton, D., Buchtel, H. A., & Douglas, R. M. (1985). Frontal lobe lesions in man cause difficulties in suppressing reflexive glances and in generating goal-directed saccades. Experimen- tal Brain Research, 58, 455-472.
Lynch, J. C., Mountcastle, V. B. , Talbot, W. H., & Yin, T. C. T. (1977). Parietal lobe mechanisms for directed visual attention. Journal of Neurophysiology, 40, 362-389.
Posner, M. I., Walker, J. A., Friedrich, F. J., & Rafal, R. D. (1984). Effects of parietal injury on covert orienting of attention. Journal of Neuroscience, 4, 1863-1874.
Robinson, D. L., Goldberg, M. E., & Stanton, G. B. (1978). Parie- tal association cortex in the primate: Sensory mechanisms and behavioral modulations. Journal of Neurophysiology, 41, 910-932.
Sandell, J. H., Schiller, P. H., & Maunsell, J. H. R. (1984). The ef- fect of superior colliculus and frontal eye field lesions on sac- cadic latency in the monkey. Perception, 13, A6.
Saslow, M. G. (1967). Effects of components of displacement-step stimuli upon latency of saccadic eye movements. Journal of the Optical Society of America, 57, 1024-1029.
Schiller, P. H., True, S. D., & Conway, J. L. (1980). Deficits in eye movements following frontal eye-field and superior colliculus ablations. Journal of Neurophysiology, 44, 1175-1189.
Shibutani, H., Sakata, H., & Hyv~irinen, J. (1984). Saccade and blinking evoked by microstimulation of the posterior parietal association cortex of the monkey. Experimental Brain Re- search, 55, 1-8.
Sparks, D. L., & Mays, L. E. (1983). Spatial localization of sac- cade targets. I. Compensation for stimulation-induced pertur- bations in eye position. Journal of Neurophysiology, 49, 45-63.
Sparks, D. L., & Porter, J. D. (1983). Spatial localization of sac- cade targets. II. Activity of superior colliculus neurons pre- ceding compensatory saccades. Journal of Neurophysiology, 49, 64-74.
Wurtz, R. H., & Albano, J. E. (1980). Visual-motor function of the primate superior colliculus. Annual Review of Neuroscience, 3, 189-226.
Wurtz, R. H., Goldberg, M. E., & Robinson, D. L. (1980). Be- havioral modulation of visual responses in the monkey: stimulus selection for attention and movement. Progress in Psychobiology, Physiology and Psychology, 9, 43-83.