the nature of trophallaxis in honeybees
TRANSCRIPT

Insectes Sociaux, Paris 1982, V o l u m e 29, n ~ 2, ZPP. 209-221
�9 Mas s o n , Par is , 1982
TILE NATURE OF TROPHALLAXIS IN HONEYBEES
PJ.A.M. KORST and H.H.W. VELTHUIS
Laboratory o~ Comparative Physiology, University o~ Utrecht, The Netherlands ReCu le 4 f6vrier 1981. Accept6 le 5 juin 1981.
SUMMARY
TrophaUactic in terac t ions bet~veen individual 'worker bees of Apis melli[era mellilera were studied in groups consisting of 2-5 bees. Less than 5 '% of these interact ions result in food transfer . I t is supposed that the major i ty of trophallactic contacts serve commu- nicational purposes ra ther than being idle food t ransfer a t tempts .
Over a for tnight period interindividual differences ,with respect to the nature of trophallactic activity become p rominen t : some workers turn out to be " asking " indivi- duals 'while others tend to specialize in offering. The nature of a ~vorl~ers' trophallactic activity is correlated ~ i t h the trophallactic t r ea tment she receives f rom her associates. These behavioural differentiations concur ~vith physiological dissimilarit ies between wor- kers, viz. ovarian development and rectum content.
The existence of a causal relation bevween the behavioural and physiological differen- t iation is assumed, the intermediate factor being the degree of t rophic advantage. In a normal queenright colony these behavioural differentiat ion processes could cause the age-dependent polyethism.
R E S U M E
Nature de la trophallaxie chez I'Abeille domestique
Les interact ions trophallactiques entre ouvri6res d'abeilles (Apis melli[era mellifera) d'individu ~t individu ont 6t6 6tudi6es darts des groupes de 2 h 5 individus. Moins de 5 % de ces interact ions aboutissent ~t des t ransfer ts de nourri ture. Nous supposons que la majori t6 des contacts trophallactiques servent davantage ~t la communicat ion qu'~t des tentatives de t ransfer t s de nourri ture.
Sur une p6riode de deux semaines, des differences inter-individuelles se d6gagent darts les activit6s t rophaUactiques: quelques ouvri6res se caractdrisent par des sollicita- tions nombreuses tandis que d 'autres se sp6cialisent dans l 'offrande. Les caract6ristiques de l 'activitd trophallact ique d 'une ouvri6re est li4e ~t I' << accueil ~ trophallact ique qu'elle re~oit des autres. Ces diff6rences 6thologiques coincident avec des disparit6s physiologiques dans le d6veloppement des ovaires et d a n s l e contenu de l 'ampoule rectale.

210 P.J.A.M. KORST, H.H.W. VELTHUIS
Nous supposons qu'il existe une relation causale entre les diff6renciations 6thologi- cues et physiologiques, le facteur interm6diaire 6tant le degr6 d'avantage trophique. Darts une ruche norrnale, ces diff6renciations 6thologiques pourrait 6tre h l'origine du poly- 6thisme, qui d6pend de l'fige.
I N T R O D U C T I O N
In a strict sense the exchange of food betxveen members of a colony means that food is s'wopped. However, in t rophallact ic interact ions between workers of an insect society food material is t ransferred f rom one individual to another by oral contact, there is a donor and a receiving individual. Tro- phallaxis is then a mat te r of transition, not of exchange. Giving food to an adult nestmate is giying away the oppor tun i ty to use those materials for other purposes. Therefore, a causal unders tanding of trophallaxis seems to be of interest.
In more primitive societies like Polistes but also in Vespula t rophallact ic interactions usually occur in an agonistic context in which the donor is regarded as the submissive, the receiving animal as the dominant individual (PARDI, 1947; MONTAGNER, 1967). This implies no random give and take between adults in such colonies. On the basis of these interactions a division of labour, including reproductivity, seems probable. In the Halictid bee Lasioglossum zephyrum a similar division of labour is regulated by means of agonistic interactions 'without food exchange (BROTHERS and MICHENER, 1974; MICHENER, 1977; BREED, 1977). One 'way or another, there has to be a causal relation between the availability of energy to the individual and its capacity to realize egg production.
In honeybee societies ~r are often involved in trophallact ic inter- actions. These contacts usually occur 'without overt aggression or submission. Comparing food transmissions between xvorkers of Vespula and Apis, MONTA- GNER and PAIN (1971) note the absence of overt aggressiveness in Apis. They suppose that the contents of the honey s tomachs flow freely between the individual bees. The concept " social s t o m a c h " has been extensively elabo- ra ted upon (see WILSON, 1971). In connect ion 'with such a f requent food exchange between all members a communica t ion system can be imagined, either through the food itself or through the admixture of pheromones (see FREE, 1965). In contras t to Vespula and Polistes trophallaxis in Apis there- fore seems to be a levelling mechanism, characterized by its non-agonistic character.
However, worke r bees that court the queen are only donors or candidate donors in food transfer. The cuticular contact ~vith the queen causes them to become loaded 'with queen pheromones and therefore, after disrupt ion of their part icipat ion in the court, they make a food gain in their contacts -~vith o ther bees (VELTHUIS, 1972 ; SEELEY, 1979). In both cases there is a relation between pheromonal identity and t rea tment by the nestmates.

T R O P H A L L A X I S IN H O N E Y B E E S 211
S o m e o t h e r o b s e r v a t i o n s i n d i c a t e the ex i s t ence of n o n - r a n d o m exchanges .
LINDAUER (1954) r e p o r t s th is fo r w a t e r h o a r d i n g s ; FREE (1957) d e m o n s t r a t e s t h a t the food f l a w is d i r e c t e d to c e r t a i n age ca tegor ies . NIxon a n d RIBBANDS
(1952) and FREE (1957) p r e s e n t d a t a i n d i c a t i n g t h a t exchange w i t h i n an age
c a t e g o r y is m o r e i m p o r t a n t t h a n b e t w e e n age groups .
In th is p a p e r ,we s tudy the t r o p h a l l a c t i c i n t e r a c t i o n s in g r o u p s of 'worke r
bees (Apis mellifera mellifera) of t he s a m e age. Espec ia l ly in s m a l l g roups
'we 'were able to qua l i fy and to q u a n t i f y the t r o p h a l l a c t i c r e l a t i o n s b e t w e e n
the i nd iv idua l 'worke r s and to r e l a t e th is to phys io log i ca l d i f f e r e n t i a t i o n s
d u r i n g the o b s e r v a t i o n pe r iod . We h o p e to u n c o v e r s o m e of t he m e c h a n i s m s tha t g o v e r n these i n t e r i n d i v i d u a l r e l a t ions , .which m i g h t en l a rge t h e ins igh t
in the p r o x i m a t e causes u n d e r l y i n g the o r g a n i s a t i o n of a h o n e y b e e co lony as
a 'whole.
M E T H O D S
T.wo kinds of observations -,vere made: 1. By means of a video set registrations were made of food transmissions between ~vo~ker bees in a queenright observation hive. Aspects of the motor pattern and time characte- ristics were studied. The registrations were made under far-red illumination. Individual bees were followed -,vith the camera through the display of the signal on a small monitor. 2. Worker bees, upon emergence, 'were marked individually and placed ,with 2-5 bees toge- ther in Liebefeld cages (see VELTZ-IUIS, 1972, for description) provided with a single-sided comb on the back 'wall. These cages ,were placed in a box kept at 32~ Interactions between bees could be observed through the frontal glass wall of the box. Observations were made almost daily for a period of 2-3 weeks using the dimmed light of a fluorescent tube. All the observed interactions 'were recorded orally on tape. Three times a .week the bees were allowed to fly out and to defalcate. Some did return voluntarily, others 'were placed back by hand.
At the end of the observation period the bees 'were examined for the activation of their ovaries. Also the appearance of their rectum was recorded.
The first observational method proved to be very suitable for detailed observations on especially the longer food transfers. Since observations in a hive make it difficult to understand ,why a particular bee sometimes offers and at other times asks for food, the second method ,was used for groups in 'which .all the individuals "~vere to be studied simultaneously. The present description of the nature of trophallaxis is based mainly on quantitative data obtained ,with this second method. In analysing these data 'we restrict ourselves to the summed totals over the whole period of observation.
R E S U L T S
1. The elements of trophallaetic interactions
W h e n t w o bees m e e t f r o n t a l l y they may , o r m a y not , e n t e r in an inter-
ac t ion. I f an i n t e r a c t i o n is o b s e r v e d this m a y s t a r t ,with the o f f e r i n g of food
o r 'with the a sk ing fo r it. The p a r t n e r in th is i n t e r a c t i o n m a y i g n o r e th is in i t i a t ive and m a y then, fo r the h u m a n obse rve r , e i t he r no t r e a c t a t all o r

212 P.J.A.M. KORST, H.H.W. VELTHUIS
may show the same behaviour, or that par tner may respond 'with the comple- mentary behaviour, viz. taking the offered food or offering after having been asked. Fur thermore, the intensity of the display varies with the condit ions within the group while the form of the display changes 'with age. Altogether the course of an encounter between two bees is subject to much variation.
In our s tudy the folio'wing behavioural elements were dist inguished on the basis of the motor p a t t e r n :
a) Asking. The asking bee opens the mandibles and prot rudes the tongue. At the same time the vertical plane through the head, t ransverse to the body axis, is tilted. These movements are accompanied by and generally preceded by more or less intense movements of the antennae that are b rought forward in the direction of the bee that is being asked. This pat tern can become more intense, in which ca~e the bee may per fo rm 'with one or bo th f ront legs grab- bing movements .
b) Offering. The offering bee opens the mandibles and lifts the p remen tum but the tongue remains hidden under the head. The vertical plane through the head, transverse to the body axis, is a lmost at right angle to that axis. The antennae are kept more or less downward and generally close to the head. In young bees a droplet of considerable size is kept between the mandibles and the prementum, but 'with increasing age the droplet soon becomes smal- ler and may generally even be invisible. The durat ion of the display shortens with increasing age as 'well. Food offering does not occur exclusively at a frontal meeting ; also 'when the side of a body is met during locomotion offe- ring may occur. Likewise, in a group at rest, bees may be observed to offer.
c) Trophallactic interaction. This is the result of complementary behaviour of the par tner after having been asked or offered. In such an interaction the focus of interest for both animals is the basis of the tongue of the pros- pective d o n o r : the outs t re tched antennae of the prospective receiver and the retracted ones of the prospective donor touch each other frequently. Those of the receiver may touch the contour of the head of the donor as 'well. The outs t re tched tongue of the prospective receiver may come into contact with the p remen tum of the donor. In these interactions the pros- pective donor holds its body closer to the comb surface .while the prospective receiver generally stands a little higher on its legs. Food t ransfer is possible only 'when the tongue of the prospective receiver touches the p r emen tum of the donor. This seems to be achieved through tactile and probably chemical information at tained during antennal and leg contacts. Stable food trans- fers may last up till 100-150 sec. with a mode between 20-30 sec.
However, not necessarily does this contact lead to food transfer. Many interactions of the above nature are of a ra ther short duration, but also in cases where the interactions may last for tens of seconds food t ransfer is impossible because the prospective receiver does not succeed in making a

TROPHALLAXIS IN HONEYBEES 213
stable contact bet.,veen its tongue and the place 'where food may become available. Apparently, in these instances there is an inability of the donor to produce food or, more likely, its un'willingness to regurgitate food and its restraint prevents the making of the proper contact. Especially under these conditions do 'we see the grabbing movements of antennae and legs of the prospective receiver, 'which may eventually lead to food rewards, and the retract ion of the head of the prospective donor (" chin on chest "). I t might be clear that such conflicts arise mainly as a result of the initiative "asking ".
d) Conflicts in trophallactic interactions. As a n extension of the above some types of interactions are even more suggestive of conflict. Observed are :
Tongue play : two bees are asking each other simultaneously. They extend their tongues witl~ the result that one places its tongue superior to the other. The other bee responds by quickly moving its tongue away and by a t tempt ing to attain the superior position. This shifting of tongues may take place a few times in succession after 'which one of the bees starts to offer. Fierce antennal movements and front leg grabbing accompany the tongue movements.
- - Tongue pulling : This can also succeed the tongue play or may occur inde- pendently. After having been asked the bee makes offering movements but upon the introduct ion of the tongue by the asking bee in bet'~veen the man- dibles of the offering bee the latter closes its mandibles and pulls or bites the tongue of its partner. This interaction may lead to tongue play or may lead to food transfer f rom the pulling bee towards the one that s tar ted .with asking. The interaction is fur ther characterized by intense antennal and leg movements of both animals.
In addition to these motor pat terns chemical informat ion is used as well. This can already be inferred from the fact that -~vithin the hive visual information can be excluded, but the FR-video recordings revealed the exis- tence of directed locomotory activity of a bee upon food offering at a distance of 1-2 body lengths. Sometimes the tongue of the approaching bee is already extended before any bodily contact has been made. Whether the odour o f the food droplet or pheromonal information is used remains hidden.
2. Quantitative aspects
In a first series of observations 16 groups of 5 bees each were used. In some of these groups mortal i ty disturbed the pat tern of interactions between the members , in others it xvas not possible to detect the clear-cut differentia- tions among the bees that 'will be described. Therefore, we will concentrate on five of these groups. Hce~vever, the conclusions to be arrived at are, to a lesser extent, also valid for most of the other cages in this series, as 'well as to similar but smaller or larger groups started later on.

214 P.J.A.M. KORST, H.H.W. VEL T H U IS
A l t o g e t h e r in a b o u t 50 % of m e e t i n g s b e t w e e n t w o bees d i d 'we o b s e r v e one of t he b e h a v i o u r a l e l e m e n t s m e n t i o n e d . The f r e q u e n c i e s of a s k i n g a n d o f f e r i n g d i f f e r e d in a c h a r a c t e r i s t i c ' w a y : in s o m e g r o u p s a s k i n g fo r food p } e d o m i n a t e d , in o t h e r s o f f e r i n g was q u i t e c o m m o n as wel l . L i k e w i s e , the o c c u r r e n c e of t o n g u e p l a y a n d t o n g u e p u l l i n g d i f f e r e d r e m a r k a b l y b e t w e e n cages . F u r t h e r m o r e , we n o t e d t ha t c h a n g e s in t he a v a i l a b i l i t y o r in the t ype of f ood g iven in the f e e d e r c o u l d i m m e d i a t e l y a n d d r a s t i c a l l y c h a n g e the b e h a v i o u r w i t h i n t he g roup . T h e r e f o r e , i t a p p e a r s t h a t in t he d y n a m i c s of the i n t e r a c t i o n s the e n v i r o n m e n t a l c i r c u m s t a n c e s a n d i n d i v i d u a l c h a r a c t e - r i s t i c s of the m e m b e r s of the g r o u p p l a y a m a j o r role .
W h a t is the d e g r e e of succes s fo r a c e r t a i n a c t i o n ? This s e e m s to be r e m a r k a b l y low. F o r the f ive cages a n a l y s e d the a s k i n g fo r f o o d l e a d s in 37.6 % of the ca ses to food o f f e r i n g b y the p a r t n e r ; u p o n o f f e r i n g in on ly 18.4 % of t he o b s e r v a t i o n s does the p a r t n e r e x t e n d i t s tongue . S i n c e f o o d t r a n s f e r s e e m s p o s s i b l e on ly w h e n t h e s e o r a l c o n t a c t s l a s t fo r a c e r t a i n min i - m u m of t i m e we d i s t i n g u i s h e d b e t w e e n s h o r t ( less t h a n one s e c ond ) a n d l o n g e r c o n t a c t s (table I). Only in 1-3 % of the O b s e r v e d i n t e r a c t i o n s a food
Table I. - - The probability of food transfers as a result of the initiatives " asking '~ and " offering ".
Tableau I . - Probabilit4 des transferts de nourriture en fonction du r6sultat des initia- tives pour solliciter et offrir.
Type Percentage Number of oral contacts of the of observations
initiative in 'which the initiative leads Total < 1 sec. > 1 sec.
to oral contacts (mean duration)
"offering " 18.4 229 212 17 (2l seconds)
" asking " 32.6 285 262 23 (16 seconds)
t r a n s f e r w o u l d be pos s ib l e , b u t t hen the a v e r a g e d u r a t i o n of the t r o p h a l l a c t i c c o n t a c t a g r e e s w i t h the d u r a t i o n s o b s e r v e d in the q u e e n r i g h t co lony b y m e a n s of t he v ideo r e c o r d i n g .
3. Trophallaxis and behavioural differentiation
I n a g r o u p c o n s i s t i n g of on ly two w o r k e r s i n d i v i d u a l d i f f e r e n c e s in t r o p h a l l a c t i c b e h a v i o u r occur . An e x a m p l e is g iven in table H. I n th i s e x a m p l e d u r i n g the w h o l e o b s e r v a t i o n p e r i o d the g r e e n - m a r k e d b e e a s k e d the r e d - m a r k e d bee m u c h m o r e f r e q u e n t l y t h a n she o f f e r e d her . On the o t h e r h a n d , t he r e d - m a r k e d ~vorker p r e d o m i n a n t l y o f f e r e d to the g r e e n w o r k e r . The g r e e n ,worker g a i n e d in t h e f o o d t r a n s f e r c o n t a c t s : she t o o k 38 t i m e s of the o f f e r i n g s b y red , go t 58 r e w a r d s of h e r o w n a s k i n g s a n d
T R O P H A L L A X I S I N H O N E Y B E E S 215
Table II. - - The trophaltactic activities of individual 'worker bees in a group consisting of two members .
Tableau II . - - Activit6s trophallactiques d'ouvri~res d'abeille dans un groupe de 2 individus.
Worker's Asking Offering Food Food Trophal- identity t ransfer t ransfer lactic
after after advantage asking offering
" green" 252* 26* + 58 - - I + 89 " red " 29* 109" + 6 - - 38 - - 89
*X 2 - test, p ~ 0.005. The X2-value is calculated on the basis of the relative frequency of " asking " and " offe- ring ", respectively ",being asked" and "be ing o f fe red" of an individual ,worker.
a c t e d o n l y 7 t i m e s as a f o o d d o n o r . T h e n e t r e s u l t o f t h i s p o l a r i t y 'was a
d i f f e r e n c e o f 89 f o o d r e c e i p t s a n d d o n a t i o n s r e s p . M o s t o f t h e s e c o n t a c t s
w e r e n o t l o n g l a s t i n g b u t s o m e 'were . T h e r e f o r e i t is n o t p o s s i b l e t o d r a w
c o n c l u s i o n s a b o u t t h e d e g r e e o f t r o p h i c a d v a n t a g e g r e e n e x p e r i e n c e s o v e r
r e d . N e v e r t h e l e s s , a t r o p h i c a d v a n t a g e s e e m s to b e c o n n e c t e d ,wi th t h e t ro -
ph~/ l lac t ic a d v a n t a g e in t h i s c a s e .
T h e s a m e d i f f e r e n c e s i n t r o p h a l l a c t i c a c t i v i t y o f ' w o r k e r s a r o s e in g r o u p s
o f 4-5 i n d i v i d u a l s . I n table I I I t h e d a t a o n o n e s u c h a g r o u p a r e p r e s e n t e d .
I n t h i s c a s e t h e i n t e r i n d i v i d u a l r e l a t i o n s b e t w e e n t h e ' w o r k e r s a r e d i s r e g a r d e d .
T h e t a b l e s h o w s t h a t t h e m e m b e r s o f t h e g r o u p d i f f e r i n t h e n u m b e r o f
Table III. - - The trophaUactic activities of individual 'worker bees in a group consisting of five members .
Tableau I I I . - - Activit6s trophallactiques d'ouvri~res d'abeille dans , u n groupe de 5 individus.
Worker's Total Asking* Offering* Being* Being* Rewarded* Troph, identity number asked offered asking advant.
of t imes involved
in a trophallactic interaction
" green " 155 26 81 30 18 3 - - 31 " r e d " 144 38 33 21 49 11 + 8 "ye l low" 221 52 18 31 120 40 + 68 " blue " 99 3 61 ~7 27 2 - - 17 "ye l low53" 140 24 61 34 21 5 - - 28
XZ-test, * p <( 0.005 (see table H).

216 P.J.A.M. KORST, H.H.W. V E L T H U I S
contacts they are engaged in , in the rat io of initiating the contact and being a " t a r g e t " resp., in the ratio asking versus offering and in the ratio being asked versus being offered. These differences lead to prominent disparities iri the trophallactic advantage or disadvantage. In relation to this it might be of interest that animals that were rarely seen in interaction wi th nest- mates generally died before the terminat ion of the experiment.
In this table I I I is also shown that the degree of success of asking is different for the members of the group : yellow is rewarded 40 times out of 52 times having asked, while green, of which animal 26 times asking has been recorded, is rewarded only 3 times. Apparent ly the bees differ in their compe- tence of eliciting offering behaviour in a par tner after their initiative asking.
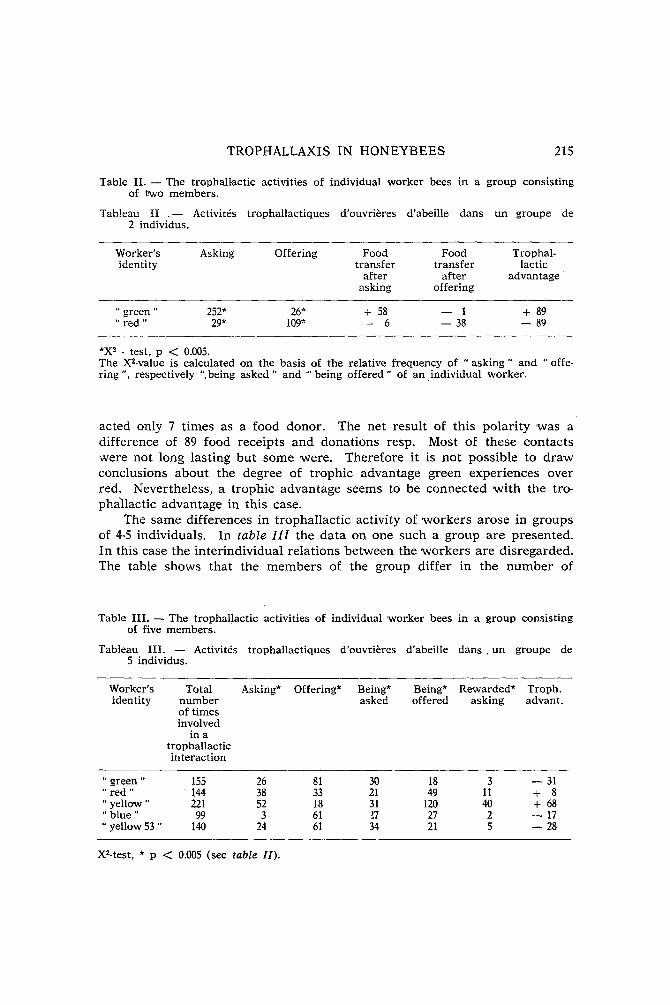
Similar differences but with sometimes quite different numerical values have been found in the other cages. In order to generalize the s t ruc ture in these cages the trolbhallactic activity of the individuals was indexed: the difference between the scores for asking and offering 'was divided by the sum of these activities. Also, the way in 'which the individuals 'were treated by their nestmates was expressed in ratio, the difference between being offered and being asked 'was divided by the sum of the scores for these treat- ments. Both indexes demonstrate a distinct individuality for each bee. Fur thermore , there is a relation between the identity as an actor and the identity as a target (fig. 1). "Asking" bees get offered, are recognizable as such. The reverse holds in a less outspoken way. Obviously, the active and the passive components of trophallactic contacts of individual workers are well-integrated.
The non-randomness of these meetings allows for a description of the hierarchical social s t ructure of such a group in terms of trophallaxis. In this view relatively high scores for offering and being asked and relatively low scores for asking, rewards, being offered and trophallactic advantage are considered as subordinate traits. An argument for such a qualification is that in aggressive encounters between honeybee workers the aggressive indi- vidual (dominant in this respect) asks for f o o d and often gets it, .while the submissive par tner shows those traits that often lead to a food gift to the dominant partner. Much support for this view can be derived from similar phenomena in wasp societies like Vespula (MoNTAaNER, 1967).
For fur ther analysis of differentiation processes between the workers we discerned three classes of individuals within the g r o u p ; for each parti- cular trait a worker was classified as a dominant individual (class 1) when she deviated in a dominant way, as ment ioned above, f rom her group mem- bers. The workers that appeared to be subordinated in an outspoken way were classifield in class 3, the others in class 2. This classification was done either by means of Chi ~ (p ~< 0.05) ( taking the animals close to the average of a certain numerical value of the group as class 2, and, depending on the direction of a significant deviation f rom the mean, divergent animals either as class 1 or 3) ; or in those cases where Chi 2 was not appropriate , by a ranking
T R O P H A L L A X I S I N H O N E Y I ~ E E S 217
+1-
-i- 8
O - �9 �9
o* g�9
O �9
O
IA
Fig. 1 . - The corre la t ion between the na ture of a worker ' s t rophal lact ic ini t iat ives and the na tu re of the init iat ives of the nestmates , directed upon this 'worker. Summat ion of the da ta of five exper imental groups, n = 23. Each dot represents one worker
asking - - offering being offered - - being asked bee. IA = IB =
asking + offering being offered + being asked
Correlat ion coefficient r = 0.4, p < 0.05.
Fig. 1 . - Corr61ation entre le type d ' ini t iat ive t rophal lac t ique d 'une ouvri~re et le type d ' ini t iat ive des autres ouvri~res du m6me nid pa r r appor t h eette ouvri~re. Nombre de donn6es obtenues h pa r t i r de 5 groupes exp6rimentaux, N = 23. Chaque point repr6sente une ouvri~re.
Soll ici tat ions - - off randes Offrandes propos6es - - sol l ici tat ions reques IA = IB =
Soll ici tat ions + offrandes Offrandes propos6es + soll ici tat ions revues
Coefficient de corr61ation r = 0.4 p < 0.05.
m e t h o d . I n a d d i t i o n , t w o o t h e r i m p o r t a n t d i f f e r e n t i a t i o n s a m o n g t h e w o r -
k e r s w e r e t a k e n i n t o a c c o u n t . S o m e ' w o r k e r s i n t h e e x p e r i m e n t a l g r o u p s
d e v e l o p e d a c t i v e o v a r i e s ' w i t h i n t h e o b s e r v a t i o n p e r i o d , u s u a l l y o n e m e m b e r
i n e a c h g r o u p , s o m e t i m e s t w o . I n o n e c a g e egg l a y i n g w a s o b s e r v e d . U p o n
d i s s e c t i o n a t t h e e n d o f t h e o b s e r v a t i o n p e r i o d a l s o d a t a w e r e c o l l e c t e d o n
t h e a s p e c t s o f t h e r e c t u m . W o r k e r s ' w i t h a c t i v e o v a r i e s u s u a l l y h a d a n a l m o s t
e m p t y r e c t u m , s o m e t i m e s c o n t a i n i n g a g r e e n i s h h y a l i n f l u i d i n s t e a d of a
t u r b i d y e l l o w p o l l e n m a s s . O t h e r ' w o r k e r s h a d a r a t h e r s w o l l e n r e c t u m 'wh i l e
i n s o m e b e e s t h e w h o l e a b d o m i n a l c a v i t y w a s f i l l e d u p b y a h e a v i l y l o a d e d
r e c t u m . O v a r i a n a c t i v i t y w a s c l a s s i f i e d as f o l l o w s : c l a s s 1 c o n t a i n s t h e
a n i m a l s h a v i n g e g g s i n t h e s a u s a g e f o r m i n a t l e a s t o n e o v a r y ; c l a s s 2

218 P.J.A.M. KORST, H.H.W. VELTHUIS
a n i m a l s ~vi th l ess a c t i v e o v a r i e s a n d c l a s s 3 t h o s e a n i m a l s 'w i th i n a c t i v e o v a -
r i e s . C o n c e r n i n g t h e r e c t u m c l a s s 1 r e f e r s to i n d i v i d u a l s 'w i th a r e l a t i v e l y
e m p t y r e c t u m , c l a s s 2 to a n i m a l s .with a n o n l y m o d e r a t e l y s w o l l e n r e c t u m a n d c l a s s 3 t h e i n d i v i d u a l s ' w i t h a h e a v i l y l o a d e d r e c t u m .
I n table IV t h e b e h a v i o u r a l a n d p h y s i o l o g i c a l c h a r a c t e r i s t i c s o f t h e ind i -
Table IV. - - The classification of the individual ,workers of a group consisting of five bees, in regard of their behavioural and physiological characteristics.
Tableau IV. - - Classification des ouvri~res d'abeille dans un groupe de 5 individus par rapport ~t leurs caract6ristiques comportementales et physiologiques.
Worker's Class 1 Class 2 Class 3 Parameters identity
" yellow " 0 �9 ['l
" red " A � 9 �9 o D O
" yellc~w 53 " 0["1 / ~ � 9 O 0
" blue " 0 o D
= * Asking/Offering
�9 = * Being asked/Being offered
O = * Rewarded as~king
�9 = ** Troph. advantage
O = *** Ovarian development
U] = *** Rectum
" green " A A 0 o o l 3
* Classification on the basis of contribution to X2-value, table III. ** Classification on the basis of the difference bel:ween the highest profit and the biggest
loss; each class covers one third of this scale. * ~ See text, page 217.
v i d u a l w o r k e r s f r o m table I I I a r e c o m p a r e d b y m e a n s o f t h e s e c l a s s i f i c a t i o n s . T h e b e h a v i o u r a l a n d t h e p h y s i o l o g i c a l t r a i t s a r e c o r r e l a t e d . T h e s a m e p h e n o -
m e n a a r e f o u n d in t h e o t h e r g r o u p s as we l l . T h e i n d i v i d u a l s w i t h t h e
h i g h e s t o v a r i a l d e v e l o p m e n t d i s t i n g u i s h t h e m s e l v e s b y t h e i r t r o p h a l l a c t i c
c h a r a c t e r i s t i c s (table V), b e i n g i n d i v i d u a l s t h a t g e t o f f e r e d a n d w h o s e a s k i n g s a r e r e l a t i v e l y w e l l - r e w a r d e d . B o t h f a c t o r s c o n t r i b u t e t o t r o p h a l l a c t i c a d v a n t a g e .
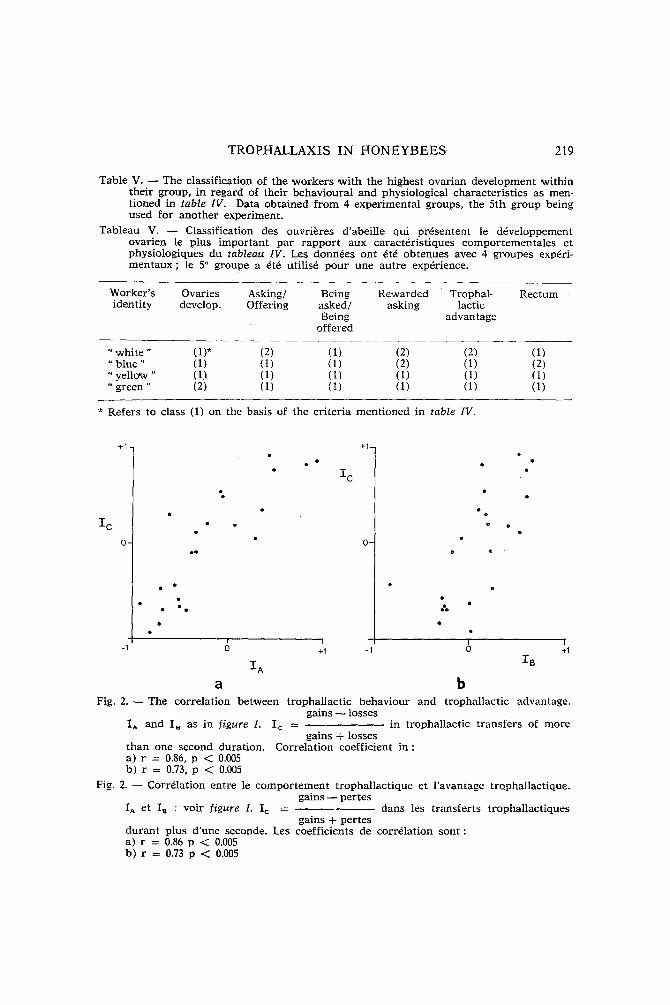
T a k i n g t h e f i v e c a g e s t o g e t h e r i t a p p e a r s t h a t d i f f e r e n c e s in t r o p h a l -
l a c t i c a c t i v i t y a n d t r e a t m e n t r e s u l t i n d i s s i m i l a r i t i e s in t r o p h a l l a c t i c a d v a n -
t a g e b e t w e e n ' w o r k e r s (fig. 2). P r o b a b l y t h i s r e f l e c t s d i s s i m i l a r i t i e s in t r o p h i c a d v a n t a g e , w h i c h c o u l d b e t h e i n t e r m e d i a t e f a c t o r in b e t n v e e n b e h a v i o u r a l d i f f e r e n t i a t i o n a n d o v a r i a l a c t i v i t y .
T R O P H A L L A X I S I N H O N E Y B E E S 219
Table V. -- The classification of the xvorkers ~vith the highest ovarian development xvithin their group, in regard of their behavioural and physiological characterist ics as men- tioned in table IV. Data obtained f rom 4 experimental groups, the 5th group being used for another experiment.
Tableau V. - - Classification des ouvri~res d'abeille qui prdsentent le d~veloppement ovarien le plus important par rappor t aux caract6ristiques comportementales et physiologiques du tableau IV. Les donn6es ont 6t6 obtenues avec 4 groupes exp6ri- men taux ; le 5" groupe a 6t6 utilis6 pour une autre exp6rience.
Worker 's Ovaries Asking/ Being Re'warded Trophal- Rectum identity develop. Offering asked/ asking lactic
Being advantage offered
"wh i t e " (1)* (2) (1) (2) (2) (1) " b l u e " (1) (1) (1) (2) (1) (2) "yelloxv" (1) (1) (1) (I) (l) (1) " green " (2) (1) (1) (1) (1) (1)
* Refers to class (1) on the basis of the criteria ment ioned in table IV.
O-
+I - +I-
T c �9 �9
e
e
s
O-
e
e
e
�9 �9
o
e �9
.%
O
O
J
-1 {3 +1 -1 (] +1
IA IB a b
Fig. 2 . - The correlation between trophallactic behaviour and trophallactic advantage. gains - - losses
IA and I8 as in f igure 1. I c = in trophallactic t ransfers of more gains + losses
than one second duration. Correlation coefficient i n : a) r = 0.86, p < 0.005 b) r = 0.73, p < 0.005
Fig. 2. - - Corr61ation entre le compor tement trophallactique et l 'avantage trophallactique. gains - - pertes
IA et I~ : voir f igure 1. Ir = darts les t ransfer ts trophallactiques gains + pertes
durant plus d'une seconde. Les coefficients de corr61ation son t : a) r = 0.86 p < 0.005 b) r = 0.73 p < 0.005

220 P.J.A.M. KORST, H.H.W. VELTHUIS
D I S C U S S I O N
As described by several authors the trophallactic ne twork in honeybee s6cieties is well-elaborated. NIxoN and RIBBANDS (1952), using a t racer techni- que, demonst ra ted a high rate of food t ransfer between worker bees. Within a day the major i ty of the workers had some of the radio-active mater ia l that was brought into the colony by only six bees.
Other students measured the frequency of oral contacts between workers . ISTOMINA-TZVETKOVA (see WILSON, 1971), following individual workers of five days old in an observation hive, repor ted that such workers engage in about 32 oral contacts per hour, " but many of these contacts were brief and did not necessarily involve transfer of fluid ". ALLEN (1960) and SEELEY (1979) report that the queen is fed by the court ing .workers not more than about six times per hour. In bur experiments we found a relatively small par t of the trophallactic encounters to lead to actual food transport .
These data indicate that food t ransfer is only a part of the trophallact ic activity of 'worker bees. However, such a low rate of real food t ransfers does not contradict the findings of NIXON and RIBBANDS (1952). Supposing that a 'worker gives 10 % of her crop content to a fellow nestmate during a real food transfer (those of at least 10 sec. durat ion) and that each worke r operates as a donor 6 times per hour a proliferation rate as described by NIXON and RIBBANDS is achieved.
Apparently, there is more in trophallaxis than solely food transfer. I t seems unlikely that the 97-99 % of the interactions in which food t ransfer could not take place are to be unders tood as idle t ransfer a t t empts ' the unsuccessfulness being due to an inappropriate st imulation or to lack of motivation of the par tners involved.
WILSON (1971, p. 290), summariz ing the studies on trophallaxis in ants and bees, states that " the role a 'worker assumes in trophallactic contacts largely depends on the state of her crop content on that m o m e n t " . MON- TA6NER and PAIN (1971) conclude the same.
However, in our experiments some workers are asking th roughout the entire observation period 'while others offer and hardly ask for food. Asking individuals even do so to nestmates 'while sitting together on the feeder. We come to the conclusion that whether a 'worker asks or offers to nes tmates depends on the identity of herself and on the identity of the par tner ra ther than merely on her feeding state.
MONTA6NER and PAIN (1971) suggest that in t ransfer contacts next to tactile stimuli chemical ones (pheromones ?) might play a role in the regula- tion of the t ranspor t direction. This intriguing suggestion is suppor ted by our findings that an asking worker is recognized as such and often gets offered from some distance, like court ing bees offer to the queen. CREWE and VELTHUIS (1980) recently repor ted on differences that arise in mandibu la r gland secretions of workers that differentiate -~vith respect to their ovaries.

T R O P H A L L A X I S IN H O N E Y B E E S 221
We a re f aced 'with the q u e s t i o n w h e t h e r these m e c h a n i s m s w o u l d ex is t
on ly in such smal l , queen le s s g r o u p s o r w h e t h e r t hey r e f l ec t p r o c e s s e s t h a t
a re i n v o l v e d in the r e g u l a t i o n of l i fe in a co lony as a 'whole. The t a sks a
�9 w o r k e r ca r r i e s ou t d e p e n d on h e r phys io log i ca l b a c k g r o u n d as ~vell as on the
socia l c o n t e x t she is c o n f r o n t e d wi th . The d i f f e r e n t ac t iv i t i es a r e p e r f o r m e d in d i f f e r e n t a r ea s of the colony. Th is i m p l i e s t h a t the socia l e n v i r o n m e n t
d i f f e r s f r o m p lace to p lace . A r i t u a l i z a t i o n o f the t r o p h a l l a c t i c s y s t e m
p r o v i d e s a c o m m u n i c a t i o n m e c h a n i s m t h a t i n f o r m s an a n i m a l a b o u t the
socia l e n v i r o n m e n t it is i n . A h o n e y b e e co lony t h e n has m u c h in c o m m o n
w i t h a H a l i c t i d l ike Las iog lossum z e p h y r u m , .where b r i e f e n c o u n t e r s betxveen
n e s t m a t e s s t rong ly i n f luence the s t a tu s of the i nd iv idua l s and as s u c h a re r e l a t e d to the t a sk d i f f e r e n t i a t i o n a m o n g t h e m . B e c a u s e of the t h o u s a n d s of
w o r k e r s w i t h i n the h o n e y b e e c o l o n y m a n y of t h e m are in a b o u t t he s a m e
e t h o l o g i c a l p h a s e ; i t is t h r o u g h th is n u m e r i c a l facto/- tha t an a s p e c t of ran-
d o m n e s s c h a r a c t e r i z e s the co lony as a who le .
References
ALLEN M.D., 1960. - - The honeybee queen and her attendants. Anim. Behav., 8, 201-208. BREED M.D., 1977. - Interactions among individuals and queen replacement in a eusocial
Halictine bee. Proc. 8th Int. Congr. IUSSI, ~ageningen, p. 228-231. BROTHERS D.J., M~CHEnER C.D., 1974.- Interactions in colonies of primitively eusocial
bees III. Ethometry of division of labour in Lasioglossum zephyrum. ]. Comp. Physiol., 90, 129-168.
CREWE R.M., VELTHUIS H.H.'W., 1980. - - False queens : a consequence of mandibular gland signals in ~vorker honeybees. Naturwissenscha#en, 67, 467-469.
FREE 3.B., 1957. - - The transmission of food bet',veen ~vor~ker honeybees. Br. 1. Anita. Behav., 5, 41-47.
FREE J.B., 1965. - - The allocation of duties among ,worker honeybees. Syrup. Zool. Soc. Lond., 14, 39-59.
LINDAUER M., 1954. - - Temperaturregulierung und Wasserhaushalt im Bienenstaat. Z. vergl. Physiol., 36, 391-432.
MICHENER C.D., 1977.- Aspects of the evolution of castes in primitively social insects. Proc. 8th Int. Congr. IUSSI, Wageningeri, p, 26.
MONTAGNER H., 1967. - - Le m6canisme et les consequences des comportements trophaltac- tiques chez les Gu~pes du genre Vespa. Bull. Biol. Fr. Belg., I00, 189-323.
MONTAGNER H., PAIN J., 1971.- Etude pr61iminaire des communications" entre ouvri~res d'abeilles au cours de la trophallaxie. Ins. Soc., 18, 177-192.
NIXON H.L., RmBANDS C.R., 1952.- Food transmission within the honeybee community. Proc. Soc., 140, 43-50.
PARDI L., 1948. - - Dominance order in Polistes wasps. Physiol. Zool., 21, 1-13. SEELEY T.D., 1979. - - Queen substance dispersal by messenger ~vor~kers in honeybee colonies.
Behav. Ecol. Sociobiol., 5, 391-416. VELTHUIS H.H.'W., 1972.- Observations on the transmission of queen substances in the
honeybee colony by the attendants of the queen. Behaviour, 41, 105-129. WILSON E.O., 1971. - - The Insect Societies. Belknap Harvard Univ. Press.