the development of peripheral cold neural circuits based ... · the development of peripheral cold...
TRANSCRIPT

TB
Ya
Lb
Uc
o
AarcdtttcotdyluptlspwBcmcwnsbtfgal
Ko
S(
*v�EArN
Neuroscience 169 (2010) 828–842
0d
HE DEVELOPMENT OF PERIPHERAL COLD NEURAL CIRCUITS
ASED ON TRPM8 EXPRESSIONmepdisbmcwchapanpwTaswn
esp(tnpncdwiHtosrTdia
spm
. TAKASHIMA,a L. MAa,b AND D. D. MCKEMYa,c*
Neuroscience Graduate Program, University of Southern California,os Angeles, CA 90089, USA
Department of Cell and Neurobiology, Zilkha Neurogenetic Institute,niversity of Southern California, Los Angeles, CA 90089, USA
Neurobiology Section, Department of Biological Sciences, Universityf Southern California, Los Angeles, CA 90089, USA
bstract—Afferent nerve fibers of the somatosensory systemre a molecularly diverse cell population that detects a variedange of environmental stimuli, converting these externalues ultimately into a sensory percept. Afferents mediatingetection of thermal stimuli express a repertoire of tempera-ure sensitive ion channels of the TRP family which endowhese nerves with the ability to respond to the breadth ofemperatures in the environment. The cold and menthol re-eptor TRPM8 is responsible for detection of cold and, unlikether thermosensors, detects both innocuous and noxiousemperatures. How this single molecule can perform suchiverse functions is currently unknown, but expression anal-ses in adult tissues shows that TRPM8 neurons are a mo-
ecularly diverse population and it is likely that this diversitynderlies differential functionality. To determine how thishenotype is established, we examined the developmentalime course of TRPM8 expression using a mouse transgenicine in which GFP expression is driven by the TRPM8 tran-criptional promoter (Trpm8GFP). We find that Trpm8GFP ex-ression begins prior to embryonic day 15.5 (E15.5) afterhich expression reaches levels observed in adult neurons.y E18.5, central axons of Trpm8GFP neurons reach the spinalord dorsal horn, but anatomical localization and in vivoeasurements of neural activity suggest that fully functional
old circuits are not established until after the first postnataleek. Additionally, Trpm8GFP neurons undergo a transition ineurochemical phenotype, ultimately reaching adult expres-ion of markers such TRPV1, CGRP, peripherin, and NF200y postnatal day 14. Thus, based on immunochemical, ana-omical and functional criteria, active cold neural circuits areully established by the second week postnatal, thereby sug-esting that important extrinsic or intrinsic mechanisms arective prior to this developmental stage. © 2010 IBRO. Pub-ished by Elsevier Ltd. All rights reserved.
ey words: temperature, TRPM8, cold, menthol, DRG, devel-pment.
ensory neurons of the dorsal root (DRG) and trigeminalTG) ganglia convert environmental stimuli of a chemical,
Correspondence to: D. D. McKemy, 3641 Watt Way, HNB 228, Uni-ersity of Southern California, Los Angeles, CA 90089, USA. Tel:1-213-821-5724.-mail address: [email protected] (D. D. McKemy).bbreviations: CGRP, calcitonin gene-related peptide; DRG, dorsal
boot ganglia; eGFP, green fluorescent protein; IR, immunoreactive;GF, nerve growth factor; Trpm8GFP, trpm8 transcriptional promoter.
306-4522/10 $ - see front matter © 2010 IBRO. Published by Elsevier Ltd. All rightoi:10.1016/j.neuroscience.2010.05.039
828
echanical, or thermal nature into electrical activity, gen-rating distinct percepts including touch, temperature andain (Basbaum et al., 2009). Their remarkable ability toifferentiate between the various modalities, as well as
ntensities, is fundamental for appropriate behavioral re-ponses to environmental changes (McKemy, 2007; Bas-aum et al., 2009). However, the detailed cellular andolecular mechanisms of each of these sensations are not
ompletely understood (Julius and Basbaum, 2001). Aide repertoire of sensory transduction molecules thatonvert external environmental stimuli into neural activityave been recently identified, thereby allowing for the ex-mination of the neurochemical, functional and anatomicalroperties of neurons to which a stimulus modality can bessigned (Basbaum et al., 2009). For example, ion chan-els of transient receptor potential (TRP) family are therimary detectors of thermal stimuli (Jordt et al., 2003),ith the temperatures considered cool to cold mediated byRP melastatin 8 (TRPM8) (McKemy et al., 2002; Peier etl., 2002; Daniels and McKemy, 2007). Thus, an under-tanding of the properties of TRPM8-expressing neuronsill provide insights into the means by which the peripheralervous system detects cold temperatures.
Peripheral sensory neurons in general are specifiedarly in development (Marmigere and Ernfors, 2007), withensory sub-lineages identified even before neural crestrecursors become committed to neuronal or glial fatesZirlinger et al., 2002). However, functionally such distinc-ions require evidence for intact neural circuits in whicheural activity or behaviors to environmental stimuli areroduced. The central projections of DRG neurons termi-ate in a restricted region of the dorsal horn of the spinalord. In growth of putative small-diameter afferents into theeveloping dorsal horn occurs in rodents beginning at E19,ith nociceptive neurons predominantly terminating in lam-
na I and II (Mirnics and Koerber, 1995b; Snider, 1998;unt and Mantyh, 2001; Julius and Basbaum, 2001). While
he establishment of functional spinal neural circuits devel-pmentally is critical for afferent signaling, expression ofensory transduction molecules such as TRPM8 is alsoequired for generation of functional neural responses.herefore, we hypothesized that functional TRPM8-depen-ent cold circuits are established when channel expression
s initiated and when TRPM8-afferent central projectionsre strictly localized at lamina I and IIo.
TRPM8 channels are expressed in functionally distinctubsets of neurons associated with unique neurochemicalrofiles (Takashima et al., 2007; Dhaka et al., 2008). Evenore striking is that many of these cells are only visualized
y TRPM8 expression and thus not easily defined molec-s reserved.

ulmrt1TtgrTnsshtwltermlccttd
I
ApttTbsw2Pprs4Mico
fSwtwlio1eNU
ra
ftSdwalopns
wbLPcttmD
P
TPPmrssnFfaA
Ee
Totpaen2a2mTwreesD6T
Y. Takashima et al. / Neuroscience 169 (2010) 828–842 829
larly (Takashima et al., 2007). In the adult, TRPM8 co-ocalizes with immunoreactivity to the intermediate fila-
ents NF200 and peripherin, markers of A�- and C-fibers,espectively, which provide distinct perceptual cold sensa-ions (Jyvasjarvi and Kniffki, 1987; Simone and Kajander,997; Takashima et al., 2007). Additionally, a cohort ofRPM8 neurons expresses nociceptor markers such as
he painful heat-gated channel TRPV1 as well as calcitoninene-related peptide (CGRP) (McNeill et al., 1988; Cate-ina et al., 1997). However, no co-localization betweenRPM8 and IB4, a neuronal marker for non-peptidergiceurons, is observed which suggests that those pre-umptive non-peptidergic neurons (CGRP-negative) ob-erved are not IB4� neurons. Nevertheless, this strongeterogeneity may account for the broad range of func-ions attributed to TRPM8 (Welberg, 2008). However,hat is not known is when this heterogeneity is estab-
ished developmentally. To begin to answer this ques-ion we used immunochemistry and mouse genetics toxamine the somal expression patterns of TRPM8 neu-ons in the developing mouse embryo, as well as deter-ined when central projections of these afferents are
ikely to establish functional connections in the spinalord dorsal horn. These data suggest that cold neuro-ircuits have the capacity to respond to cold tempera-ures very early in development, but do not form func-ional circuits until late in to the second week of postnatalevelopment.
EXPERIMENTAL PROCEDURES
mmunohistochemistry
ll animal work was performed in accordance with protocols ap-roved by the Institutional Animal Care and Use Committees at
he University of Southern California following the National Insti-utes of Health regulations. Embryos were collected from pregnantrpm8GFP transgenic mice with the plug day designated as em-ryonic day 0.5 (E0.5). Mouse embryos from different embryonictages were cut on a cryostat (Microm HM560). Sections (16 �m)ere processed for immunochemistry as described (Zhao et al.,009). Postnatal animals were transcardially perfused with 0.1 MBS followed with fixative (4% formaldehyde, 0.2% (v/v) saturatedicric acid, 0.1 M PBS, pH 7.3, at 4 °C). DRGs from lumbaregions 4, 5 and 6 were collected and corresponding sections ofpinal cord were carefully dissected and post-fixed overnight at°C in the same fixative solution. Samples were washed with 0.1PBS for 30 min. All tissues were cryo-protected in 30% sucrose
n PBS at 4 °C overnight, quickly frozen in OCT, sectioned with aryostat at 12 �m (DRG) and 20 �m (spinal cord), and mountedn Superfrost Plus slides. Slides were stored at �80 °C.
Frozen slides were dried at 4 °C for 30 min and then at 24 °Cor additional 30 min and washed with deionized water for 30 s.lide-mounted sections were washed three times with PBS, onceith PBS plus 0.3% Triton X-100 (PBS-T) for 45 min, and three
imes with PBS before a blocking step (1 h at room temperatureith 20% normal goat serum in PBS). Primary antibodies were di-
uted in a PBS solution and incubated at 4 °C overnight. Antibod-es and dilution: 1:1000 chicken anti-GFP (GFP-1020; Aves Lab-ratories); 1:500 guinea-pig anti-CGRP (T-5027; Peninsula);:500 rabbit anti-NF200 (N-4142; Sigma); 1:500 rabbit anti-periph-rin (AB-1530; Millipore); 1:500 rabbit anti-TRPV1 (RA-14113;euromics); 1:500 rabbit anti-TrkA (a gift from L. Reichardt,
CSF), 1:500 mouse anti-TrkB (610102; BD Transduction Labo- matories), 1:500 rabbit PV (AF-1404; R&D system), 1:1000 rabbitnti-c-fos (SC-253; Santa Cruz Biotechnology).
Sections were washed three times with PBS and incubatedor 2 h at room temperature with secondary antibodies conjugatedo Alexa-488 or Alexa-568 (Invitrogen) diluted 1:1000 in PBS.lides were then washed three times with PBS, three times witheionized water, and mounted in Vectorshield mounting mediumith DAPI (H-1200; Vector Laboratories, Inc.). Digital images werecquired on an Olympus IX70 microscope. Quantification of over-
ap between green fluorescent protein (eGFP) expression and thatf other neuronal markers was obtained per section and ex-ressed as the percentage of GFP-positive cells that were immu-oreactive for the respective markers with the SEM betweenections obtained from 5 to 12 animals.
For Fos antibody staining, sections were washed three timesith PBS and incubated for 2 h at room temperature with (1:200)iotinylated goat anti-rabbit secondary antibody (PK-6101; Vectoraboratories, Inc.) in PBS-T. Slides were washed three times withBS and incubated for 1 h at room temperature with 1:350 Cy3-onjugated streptavidin (SA1010; Invitrogen) in PBS-T while pro-ected from light. Sections were washed three times with PBS,hree times with deionized water, and mounted in Vectorshieldounting medium with DAPI (H-1200; Vector Laboratories, Inc.).igital images were acquired as described.
eripheral stimulation
rpm8GFP transgenic mice at different ages P3, P10, P14, and35 (P14 and P35 were first anesthetized with isoflurane (Phoenixharmaceutical, Inc.)) were treated with vehicle (70% ethanol) orenthol (1 M; Sigma) applied to the ipsilateral hind paw for 5 s
epeated 10 times with 1 min intervals. After 90 min, mice wereacrificed and spinal cords dissected, fixed, sectioned, andtained with c-fos antibody as described above. Number of c-fosuclei was obtained per field and expressed as the number ofos-positive cells per optical field with the SEM between fields
rom multiple sections obtained from five to eight independentnimals. All datasets were analyzed using two- or one-wayNOVA analysis followed by Tukey’s HSD post hoc analysis.
RESULTS
xpression of TRPM8 in the developing mousembryo
o examine the time course of TRPM8 expression devel-pmentally we used a thoroughly characterized mouseransgenic line in which enhanced GFP (eGFP) is ex-ressed via the Trpm8GFP (Takashima et al., 2007; Carr etl., 2009; Mandadi et al., 2009). Trpm8GFP is robustlyxpressed and recapitulates expression of TRPM8 chan-els in peripheral sensory neurons (Takashima et al.,007), making it a suitable marker for TRPM8 expressionnd analysis of TRPM8-expressing neurons (Welberg,008). We first examined DRG neurons in the developingouse embryo to establish the embryonic stage at whichRPM8 expression begins. At embryonic day 14.5 (E14.5),e observed very sparse eGFP expression in DRG neu-
ons from Trpm8GFP mice, prior to which no transgenexpression was detectable (Fig. 1A). However, by E16.5GFP labeling began to increase followed by more exten-ive labeling by E18.5 (Fig. 1A, B). At E18.5, 8.1�1.0% ofRG neurons (n�3249 cells) were labeled compared to.8�0.6% at P0 (n�2931 cells; Fig. 1B). After birth,rpm8GFP expression was found in a subset of small-to
edium-diameter neurons, similar to that we previously
otseT7Dsdaa
Tt
Webldt
wewtsseto2dpsttfipend
Fem eferenceso
Y. Takashima et al. / Neuroscience 169 (2010) 828–842830
bserved in adult tissues (Takashima et al., 2007). Thisime course is consistent with previous reports of expres-ion of TRPM8 transcripts in mouse DRG (Hjerling-Lefflert al., 2007; Shibasaki et al., 2010). Of note, the extent ofrpm8GFP expression remained consistent postnatal with.7�0.7% (n�2974 cells) and 7.7�0.6% (n�2574 cells) ofRG neurons positive for Trpm8GFP at P3 and P14, re-pectively, even though there was significant neuronaleath during this period (Fitzgerald, 2005). Thus, sensoryfferents are molecularly prepared to detect cold temper-tures at a developmental stage just preceding birth.
RPM8 afferent axons innervate the dorsal horn ofhe spinal cord at approximately E16.5
ith expression beginning on or near E16.5, we nextxamined the central projections of Trpm8GFP afferent fi-ers to determine when these putative cold nerves are
ikely to establish functional connections in the spinal cordorsal horn. In the rat, initial growth of afferent fibers into
ig. 1. Developmental expression of TRPM8. (A) Trpm8GFP fluorescexpression of TRPM8 in small diameter neurons. (B) Quantificationean�SEM (P�0.05) and bars equal 20 �m. For interpretation of the rf this article.
he spinal gray matter occurs at approximately E14–E15, K
ith a second wave of penetration of small diameter affer-nts at E19 (Mirnics and Koerber, 1995b). To determinehen TRPM8 fibers establish their central projections into
he mouse dorsal horn, we examined Trpm8GFP expres-ion in embryonic whole mounts starting at day E15.5, atage of development just prior to that of extensive eGFPxpression (Fig. 1). A few eGFP-positive fibers were ini-ially found in the dorsal roots by E15.5 (Fig. 2A) and thenbserved to reach the medial dorsal horn by E16.5 (Fig.B). By E18.5, Trpm8GFP fibers were still localized pre-ominantly to the medial entry site, but had begun toroject ventrally into the developing dorsal horn by thistage (Fig. 2C). However, the innervation observed at thisime was restricted around the point of entry compared tohat of adults (Figs. 2C, 3A). Our finding that Trpm8GFP
bers enter the dorsal horn by E18.5 is consistent withrevious dye labeling studies in which small-caliber affer-nt fibers, largely considered to be thermosensitive andociceptive, were first observed to enter the developingorsal horn at or near day E18–19 in the rat (Mirnics and
ed thin sections from lumbar DRG taken at the stage indicated showsrcent of neurons within a ganglia that express Trpm8GFP. Data areto color in this figure legend, the reader is referred to the Web version
nce in fixof the pe
oerber, 1995b). By P0, eGFP-labeled fibers were ob-

s(ra3tegv
D
IgEoaan
FddE
Fihr
Y. Takashima et al. / Neuroscience 169 (2010) 828–842 831
erved lateral to the entry site, but still diffusely spreadFig. 3B). However, by P3 eGFP positive fibers began toesolve their projection patterns towards one similar todult tissues (Fig. 3C) and were fully refined by P10 (Fig.D), located in the most superficial laminar structures in
he mouse spinal cord dorsal horn (Fig. 3E, F) (Takashimat al., 2007). Thus, Trpm8GFP fibers reach an adult topo-raphic organization after the first week of postnatal de-elopment.
ig. 2. TRPM8-positive central projections into the spinal dorsal hornorsal roots (arrowheads) as early as E15.5 (A) and increase in densitorsal horn but are restricted to the site of entry (C; arrow). Moreover, t18.5. Bars equal 50 �m.
ig. 3. Distribution of the central projections of TRPM8 fibers in the dnnervating the dorsal horn, but largely restricted to the medial sites (A,
orn (C, D) and by P14 (E) have resolved into an adult-like somatotopic orgeferences to color in this figure legend, the reader is referred to the Web versevelopment of TRPM8 spinal projections
n rodents, the spinal cord dorsal horn formed such that theeneral shape and structure was largely that of the adult by17–E18 (Mirnics and Koerber, 1995b). We have previ-usly reported that Trpm8GFP central afferent projectionsre restricted to lamina I and IIo in the adult (Takashima etl., 2007), consistent with thermosensitive and nociceptiveeurons (Julius and Basbaum, 2001). However, in late
section of Trpm8GFP mice, GFP-positive fibers are observed in the4 h (B). By E18.5, the central projections of Trpm8GFP fibers enter theers of GFP-positive soma are markedly increased between E15.5 and
g spinal dorsal horn. At E18.5 and P0, Trpm8GFP axons are denselyand P10, Trpm8GFP axons have reached lateral regions of the dorsal
. In fixedy within 2he numb
evelopinB). By P3
anizational pattern (F). Bars equal 50 �m. For interpretation of theion of this article.

elteteaiTasd
eeTvvfi4bwHof
FfiP(
Y. Takashima et al. / Neuroscience 169 (2010) 828–842832
mbryonic and postnatal stages, Trpm8GFP fibers wereocated more diffusely and not in a topographic organiza-ion indicative of adult tissues (Fig. 3). To determine thextent of Trpm8GFP innervation in the dorsal horn relevant
o other afferent subtypes, we compared the localization ofGFP-labeled fibers to that of a well characterized temper-ture-sensitive and nociceptive fiber type, those express-
ng the heat-gated channel TRPV1 (Caterina et al., 1997;ominaga et al., 1998). In rats, TRPV1-positive afferentsre known to terminate predominantly in lamina I withparse projections in lamina II between E16.5 to postnatalay 2, but then extend ventrally into lamina II by P10 (Guo
ig. 4. Somatotopic organization of TRPM8 and TRPV1 central projecbers are observed in the dorsal horn, with Trpm8GFP axons projecting
GFP
3 (B), Trpm8 fibers are largely more ventral that TRPV1 axons but beginD) Trpm8GFP and TRPV1 axons are both localized to the same laminar regiont al., 2001). We observed a similar pattern of TRPV1 fiberxpression in Trpm8GFP mice in that the localization ofRPV1-immunoreactive (IR) afferents show a dorsal toentral progression during the first week of postnatal de-elopment (Fig. 4). In contrast, we found eGFP-positivebers extending well past TRPV1 terminals at E18.5 (Fig.A, Ai) and P3 (Fig. 4B). At E18.5, a visible distinctionetween Trpm8GFP and TRPV1-IR fibers was observedith little to no overlap in the projection patterns (Fig. 4Ai).owever, by P10 this extended ventral innervation patternf TRPM8 fibers had retracted dorsally (Fig. 4C) andormed into a pattern indicative of adult innervation and a
e developing dorsal horn. (A) At E18.5, both Trpm8GFP and TRPV1-IRntral than those immunoreactive for TRPV1 (see boxed region Ai). By
tions in thmore ve
to retract and largely overlap with TRPV1 axons by P10 (C). By P14(Di) as is observed in the adult mouse. Bars equal 50 �m.

st
powdaclT1tprPma5stdsmrlfi2aiPswdmda
Esd
TdsihddisPee2otcpm
s((titit(luPrsdcfttfian
Ti
WictTp(Mc(t
cnsmcaaSdn2a9wd2eFef2
Y. Takashima et al. / Neuroscience 169 (2010) 828–842 833
trong overlap in Trpm8GFP and TRPV1-IR central projec-ions (Fig. 4D, Di).
To further define the development of TRPM8 centralrojection we also compared Trpm8GFP labeling to twother lamina specific markers, CGRP and PKC�, each ofhich labels distinct and non-overlapping regions of theorsal horn (McNeill et al., 1988; Polgar et al., 1999; Huntnd Mantyh, 2001; Braz and Basbaum, 2009). CGRPharacteristically labels lamina I and the outer layers ofamina II in the mouse spinal cord and largely overlaps withRPM8 expressing afferents in the adult (McNeill et al.,988; Takashima et al., 2007). We were unable to examinehe locale of CGRP fibers embryonically as peptide ex-ression begins at approximately E18 once afferent fiberseach their peripheral projections (Ai et al., 1998, 1999). By0, CGRP� neuronal projections were localized to theost superficial regions of the dorsal horn and Trpm8GFP
xons largely overlapped with CGRP-positive fibers (Fig.A, Ai). However, by P14, TRPM8 projections were ob-erved to take up a more refined localization suggestinghat they had retracted and moved predominantly moreorsal than CGRP fibers (Fig. 5B, Bi), similar to that ob-erved for TRPV1 afferents (Fig. 4). We next examined aore ventral lamina marker PKC� which labels interneu-
ons located to the inner regions of lamina II as well asamina III and does not overlap with TRPM8 expressingbers in the adult (Zylka et al., 2005; Takashima et al.,007; Seal et al., 2009). At P14, axons labeled for TRPM8re distinctly separated in the dorsal horn from spinal
nterneurons expressing PKC� (Fig. 5D, Di). However, at0, we found eGFP expressing axons to project close topinal neurons expressing PKC� (Fig. 5C, Ci), consistentith a more ventral projection of Trpm8GFP fibers in earlyevelopment. Thus, TRPM8-positive afferents have aore diffuse projection pattern in the mouse spinal cordorsal horn early in development compared to that found indult animals.
stablishment of functional TRPM8-dependentynaptic connections in the spinal cordevelopmentally
he somatotopic organization of Trpm8GFP fibers in theeveloping mouse spinal cord suggests that appropriateynaptic connections between these fibers and spinal res-dent second-order projection neurons and interneuronsave yet to form, or are in the process of being established,uring the first postnatal week. Precedence for temporalevelopment of cold-activated circuits comes from record-
ngs of menthol-evoked glutamatergic synaptic activity inpinal cord slices from postnatal rats (Baccei et al., 2003).rior to P10, menthol does not alter the frequency ofxcitatory postsynaptic currents above baseline, but canvoke robust activity from this stage forward (Baccei et al.,003). As this time course coincides with the progressionf Trpm8GFP spinal projections to a more adult-like pheno-
ype, we asked whether functional menthol-dependent cir-uits were present prior to P10 in vivo using protein ex-ression of the immediate early gene c-fos (Fos) as a
arker of neural activity (Coggeshall, 2005). Thus, we (timulated the hind paw of anesthetized Trpm8GFP miceat days P3, P10, P14 and P35) with menthol unilaterallysee Experimental Procedures) and examined the induc-ion of Fos protein expression in the spinal cord. As shownn Fig. 6, peripheral menthol evokes robust Fos expression inhe dorsal horn at day P14 (30.8�0.7 nuclei per sectionpsilateral and 6.9�0.7 contralateral; P�0.01; n�6) to levelshat are identical (P�0.05) to those observed in adult mice�P35; 32.5�0.8 ipsilateral; n�5). However, at P3 and P10ittle Fos expression was induced by peripheral menthol stim-lation and at levels statistically lower than P14 (8.6�1.6 at3, n�4; 13.6�0.9 at P10, n�4; P�0.01). Thus, as has been
eported for menthol-evoked post-synaptic currents in rodentpinal cords (Baccei et al., 2003), little to no TRPM8 depen-ent activity occurs in the first few postnatal days. Whenonsidered in the context of the developmental delay in theormation of adult-like somatotopic organization of TRPM8erminals in the spinal cord, these data suggest that func-ional cold neural circuits are not established until after therst postnatal week, and that there is likely either trophic orctivity-dependent refinement of these projections to the spi-al cord dorsal horn during this time.
emporal delay in development of themmunochemical phenotype of TRPM8 neurons
e have previously shown that Trpm8GFP neurons aremmunoreactive to many classical somatosensory and no-iceptive markers and exhibit a remarkable diversity inheir neurochemical phenotypes (Takashima et al., 2007).hese results, along with the striking complexity in thehenotype of mice lacking functional TRPM8 channelsBautista et al., 2007; Colburn et al., 2007; Daniels andcKemy, 2007; Dhaka et al., 2007), suggests that the
hannel can serve a multitude of roles in sensory signalingWelberg, 2008). Thus, we examined the developmentalime course that establishes this diversity in functionality.
First, we looked for when eGFP-positive neurons be-ame immunoreactive for TRPV1 and CGRP, markers ofociceptive neurons (Basbaum et al., 2009). Functionaltudies with cultured sensory neurons show that approxi-ately 40–50% of menthol-sensitive neurons are also
apsaicin-sensitive, suggesting overlap between TRPM8nd TRPV1 in adult mice (McKemy et al., 2002; Viana etl., 2002; Xing et al., 2006; Hjerling-Leffler et al., 2007;hibasaki et al., 2010). Consistent with these functionalata, in the adult mouse approximately 40% of Trpm8GFP
eurons are immunoreactive for TRPV1 (Takashima et al.,007). Functional assays (using Ca2� microfluorimetry incutely cultured DRG neurons) showed a greater than0% overlap in the number of menthol-sensitive neuronsith capsaicin-sensitive cells at E18.5, with this numberecreasing to near 50% by P14 (Hjerling-Leffler et al.,007). Thus these data predict a strong overlap in thexpression of TRPM8 and TRPV1 early in development.or TRPV1 we found co-expression with Trpm8GFP asarly as E18.5 and at levels not significantly different thanound in P14 animals and adult mice (Figs. 7A, 8;5.5�1.4% at E18.5, 28.9�3.2% at P14; P�0.05)
Takashima et al., 2007). Surprisingly, at postnatal days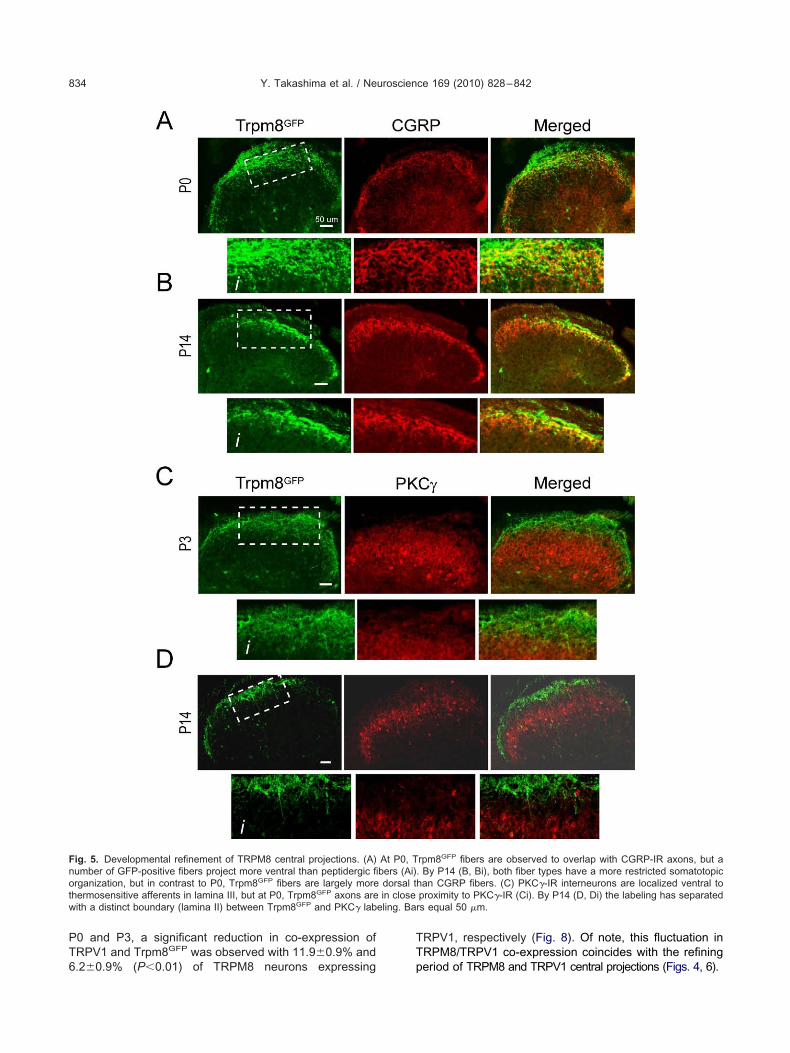
PT6
TT
Fnotw eling. Ba
Y. Takashima et al. / Neuroscience 169 (2010) 828–842834
0 and P3, a significant reduction in co-expression ofRPV1 and Trpm8GFP was observed with 11.9�0.9% and
ig. 5. Developmental refinement of TRPM8 central projections. (A)umber of GFP-positive fibers project more ventral than peptidergic firganization, but in contrast to P0, Trpm8GFP fibers are largely more
hermosensitive afferents in lamina III, but at P0, Trpm8GFP axons areith a distinct boundary (lamina II) between Trpm8GFP and PKC� lab
.2�0.9% (P�0.01) of TRPM8 neurons expressing p
RPV1, respectively (Fig. 8). Of note, this fluctuation inRPM8/TRPV1 co-expression coincides with the refining
rpm8GFP fibers are observed to overlap with CGRP-IR axons, but a. By P14 (B, Bi), both fiber types have a more restricted somatotopichan CGRP fibers. (C) PKC�-IR interneurons are localized ventral toproximity to PKC�-IR (Ci). By P14 (D, Di) the labeling has separatedrs equal 50 �m.
At P0, Tbers (Ai)dorsal tin close
eriod of TRPM8 and TRPV1 central projections (Figs. 4, 6).

nCsncPTma(v(2dentlsafPt1ntm
ttpTmwwca
msrpbtPipt(oCm
Tn
Ninaf12teeplppTesievtrTi1nTdTrpr
gwnbepne
Fdtm(
Y. Takashima et al. / Neuroscience 169 (2010) 828–842 835
For CGRP, as stated previously, expression beginsear E18 in the rodent and thus we saw little overlap inGRP immunoreactivity at E18.5 (not shown) but did ob-erve a graded increase in CGRP expression in Trpm8GFP
eurons at P0 (8.6�1.6%) and P3 (8.0�0.9%) witho-expression reaching levels observed in the adult by14 (20.9�3.5%; Figs. 7B, 8). Since only a fraction ofrpm8GFP neurons labeled with CGRP, it suggests thatany of these cells can be considered non-peptidergicnd should be labeled with the markers IB4 and P2X3Zylka et al., 2005). However, we and others have pre-iously shown that adult TRPM8 neurons do not bind IB4Peier et al., 2002; Takashima et al., 2007; Dhaka et al.,008), an expression phenotype we also observe ineveloping DRG neurons (data not shown). To furtherstablish the neurochemical composition of Trpm8GFP
eurons, we also examined co-expression with the iono-ropic ATP receptor P2X3, which is known to not co-ocalize with CGRP and is almost exclusively localized inmall-diameter non-peptidergic sensory neurons (Vulch-nova et al., 1998). Similar to CGRP labeling, a smallraction of Trpm8GFP neurons were immunoreactive for2X3 at P0 (2.5�0.3%) with expression increasing as
he animal developed (10.8�1.4% at P14; Suppl. Fig.), further demonstrating that TRPM8 afferents expressociceptive markers, but that a significant number ofhese cells cannot be classified by traditional neuronalarkers (Takashima et al., 2007).
A�- and C-fibers are two functionally distinct fiber typeshat can be identified immunochemically by expression ofhe neurofilament markers NF200 and peripherin, res-ectively (Lawson and Waddell, 1991). In adult mice,rpm8GFP-positive neurons were found to express botharkers in adult DRG, albeit many of TRPM8 neuronsere not immunoreactive for either marker and thereforeere unable to be classified as C- or A�-fibers immuno-hemically (Takashima et al., 2007). Nonetheless, we ex-
ig. 6. Menthol-evoked neural activity over the first week of postnatalevelopment. Quantitative data of the number of Fos positive nuclei inhe lumbar spinal cord dorsal horn after hind paw stimulation withenthol at P3 (n�11 sections), P10 (n�32), P14 (n�52), and P35
n�36). ** denote P�0.01.
mined when TRPM8 neurons begin to express these n
arkers during development. As shown in Figs. 9A, 10ubset of Trpm8GFP neurons were immunoreactive for pe-ipherin at E18.5 (5.4�0.8%) and that the percentage oferipherin-positive neurons increased significantly afterirth (13.7�1.9% at P0, P�0.01), reaching levels similar tohat of adult mice by P3 (20.0�1.4% at P3; 18.5�1.3% at14, P�0.05). In contrast, we observed little to no NF200
mmunoreactivity in Trpm8GFP neurons during the firstostnatal week (0.3�0.3% at P0 and 0.2�0.1% at P3), buthen co-expression levels increased to adulthood by P14Figs. 9B, 10; 21.1�0.8%). These data show that a fractionf TRPM8 neurons can be immunochemically classified as-fibers early in development, but fail to express A�-fiberarkers until after the first week of development.
RPM8 neurons exhibit limited expression ofeurotrophic receptors
eurotrophic growth factor receptors and their correspond-ng ligands are the earliest recognized markers of sensoryeuron subtypes and known to control cell developmentnd death, target innervation, and the establishment ofunctional contacts to spinal cord neurons (Smeyne et al.,994; Silos-Santiago et al., 1995; Marmigere and Ernfors,007). In addition, these receptors are critical for the es-ablishment of neuronal phenotypes in that they regulatexpression of both ion channels and peptides. Small diam-ter thermosensory and nociceptive neurons largely ex-ress the nerve growth factor (NGF) receptor TrkA, while
arge diameter low-threshold mechanosensitive and pro-rioceptive neurons express the brain derived neurotro-hic factor (BDNF) and neurotrophin 4 (NT-4) receptorrkB, and the NT-3 receptor TrkC, respectively. Severallegant studies have shown that the majority of DRG sen-ory neurons are dependent on NGF for their survival early
n development (Marmigere and Ernfors, 2007). Thus, wexamined the expression of specific receptor types in de-eloping TRPM8 neurons to determine if the diversity inhe expression pattern of these neurons correlates witheceptor subtype. Surprisingly, only a small percentage ofrpm8GFP neurons are immunoreactive for TrkA embryon-
cally (5.6�2.1% at E16.5 and 9.3�1.0% at E18.5; Fig.1), but the co-expression levels gradually increased post-atal, reaching a peak of 32.8�1.5% at P35 (Fig. 11B).hese data suggest that TRPM8 neurons are likely to beependent on trophic factors other than NGF. However,rpm8GFP neurons are not immunoreactive for the BDNF
eceptor TrkB (data not shown), and only a very smallercentage were immunoreactive for parvalbumin, a sur-ogate marker of TrkC-expressing neurons (Suppl. Fig. 2).
We also examined co-expression of Trpm8GFP with thelial-derived neurotrophic factor (GDNF) receptor Ret, ofhich a subset of labeled neurons are small-diameteron-peptidergic neurons that initially co-express TrkA, butegin to co-express Ret at E16 (Molliver et al., 1997; Chent al., 2006; Luo et al., 2007). These neurons undergo aostnatal switch in trophic factor dependence and elimi-ate TrkA expression, which is complete by P14 (Mollivert al., 1997). Thus, we examined if a subset of TRPM8
eurons go through this differentiation by determining co-
Fo
Y. Takashima et al. / Neuroscience 169 (2010) 828–842836
ig. 7. Immunochemical expression of nociceptive markers in TRPM8 neurons. Fixed sections of DRG from Trpm8GFP mice shows co-labeling of a fraction
f TRPM8 neurons with TRPV1 (A) and CGRP (B). Arrowheads label cells co-expressing Trpm8GFP and the respective marker. Bars equal 50 �M.
lfrnwsf
D
Apioesunt(T2tdtAsaaf
b
oo2reOtib2EsaeTsew2nntsc
Tp
CicecsTcosTstdmmthssautoenthntc
FcCw1a(2*
Y. Takashima et al. / Neuroscience 169 (2010) 828–842 837
ocalization between Trpm8GFP and Ret. However, weound little to no co-localization between Trpm8GFP neu-ons and Ret (Suppl. Fig. 3), suggesting these neurons areot Ret-dependent, non-peptidergic neurons, consistentith observations in mice lacking Ret expression in sen-ory afferents in which TRPM8 expression was not af-ected (Luo et al., 2007).
DISCUSSION
evelopmental profile of TRPM8 expression in DRG
requirement for active sensory transduction is the ex-ression and correct localization of transduction molecules
n the appropriate cell type and cellular domain. In the casef temperature-sensing afferents, select TRP ion channelxpression is fundamental for the establishment of thermalensitivity. In the mouse, neural crest cells coalesce andltimately differentiate into small diameter neurons at orear E11.5 (Lawson and Biscoe, 1979), a subpopulation
hat correlates to nociceptive and thermosensitive neuronsJulius and Basbaum, 2001). Remarkably, 1 day laterRPV1 transcripts can be detected (Hjerling-Leffler et al.,007; Shibasaki et al., 2010), which is then followed shortlyhereafter by protein expression (Funakoshi et al., 2006),emonstrating that a subpopulation of DRG neurons es-ablish their functional identity very early in development.t the cellular level, TRPV1 channels are functional at thistage (Hjerling-Leffler et al., 2007; Shibasaki et al., 2010),nd capsaicin can evoke spinal glutamate release as earlys P0 (Baccei et al., 2003), suggesting the presence ofunctional neural circuits.
In contrast, we find that TRPM8 expression (Trpm8GFP)
ig. 8. Co-expression of TRPM8 with nociceptive markers. Quantifi-ation of the percent of Trpm8GFP neurons that co-label with TRPV1 orGRP-IR. Among Trpm8GFP neurons co-staining for TRPV1 thereere 25.5% at E18.5 (356/1398 in 14 sections), 11.9% at P0 (119/000 in 10 sections), 6.2% at P3 (75/1210 in 12 sections), and 28.9%t P14 (550/1903 in 19 sections). For CGRP there were 8.6% at P052/605 in 6 sections), 8.0% at P3 (168/2100 in 21 sections), and0.9% at P14 (229/1096 in 11 sections). Data are means�SEM and* denotes P�0.01.
egins several days after TRPV1 (E15.5–16.5), a devel- w
pmental time point that coincides with previous analysesf TRPM8 transcript expression (Hjerling-Leffler et al.,007; Shibasaki et al., 2010). Likewise, menthol-evokedesponses are observed at the cellular level near this time,vidence to the functional expression of TRPM8 channels.f note, Hjerling-Leffler and colleagues also report that cold
emperatures (down to 5 °C) evoke activity (measured byncreased intracellular calcium) in a larger percentage of lum-ar DRG neurons (45%) than menthol (Hjerling-Leffler et al.,007). Moreover, cellular cold-sensitivity starts as early as12.5–14.5, a full 2–3 days prior to the onset of menthol-ensitivity and TRPM8 expression. Thus, these expressionnd functional differences suggest that cold-evoked activity inmbryonic neurons may use mechanisms distinct fromRPM8. TRPA1 has also been reported to act as a coldensor in DRG neurons (Story et al., 2003), but transcriptxpression and TRPA1-specific agonist evoked responsesere not observed until after birth (Hjerling-Leffler et al.,007), indicating these early embryonic cold responses wereot TRPA1-mediated. Moreover, the transduction mecha-ism that provides for the increased calcium response to cold
emperatures in this study is not clear, nor is it evident thatuch activity is biologically relevant in the development ofold inputs.
RPM8 dependent cold circuits are establishedostnatal
ellular expression and evidence for channel activationn vitro does not necessarily correlate to functional neuralircuits in the developing nervous system. Therefore, wexamined when TRPM8 afferents likely make synapticonnections with their secondary partner neurons in thepinal cord during development. Our study reveals thatrpm8GFP afferents innervate the dorsal horn of the spinalord starting at approximately E18.5, but likely are not partf a fully functional neural circuit due to an immatureomatotopic organization in the spinal cord dorsal horn.his suggests that fine-tuning of synaptic connections istill possibly occurring at this time. To test this, we inves-igated when the functional synaptic connections are madeuring development. Previous studies show the establish-ent of functional cold circuits in rats by P10 by measuringEPSC frequency in response to menthol by neurons in
he dorsal horn of the spinal cord (Baccei et al., 2003). Weypothesized that TRPM8 afferents establish functionalynaptic connections when the termination pattern is re-tricted within lamina I and IIo. If this is true we should beble to observe markers of synaptic activity, namely Fospon menthol stimulation after the refinement period ratherhan before. Our results show that at all postnatal devel-pmental stages tested in this study except for P3, c-fosxpression in the dorsal horn of the spinal cord was sig-ificantly higher in menthol-treated animals ipsilateral sideo the simulation site than in either the contralateral dorsalorn or in vehicle-treated animals. Moreover, the relativeumber of Fos-positive nuclei observed in P14 was similar to
hose at P35. Therefore, we conclude that functional centralold circuits are established by P14, a stage much later than
hen the peripheral projections of these fibers are likely to
rfiE
Es
Foe
Y. Takashima et al. / Neuroscience 169 (2010) 828–842838
each their peripheral targets. Previous studies show thatbers originating from lumbar DRG reach the hind paw by
ig. 9. Immunochemical expression of A�- and C-fiber markers in TRf a fraction of TRPM8 neurons with peripherin (A) and NF200 (B). Arqual 50 �M.
14.5–E15, and the epidermis of the most distal toes by T
16–E16.5, (Mirnics and Koerber, 1995a) approximately theame time Trpm8GFP expression begins in DRG neurons.
rons. Fixed sections of DRG from Trpm8GFP mice shows co-labelinglabel cells co-expressing Trpm8GFP and the respective marker. Bars
PM8 neurowheads
his indicates that peripheral targeting occurs before central

tm
GT
NismSdeiamsegwewTrdnftwshenvr
crttmttRtTgrgnm
Nd
Wwoesc2adtTgnpCaermtcwiprFfiaerd
tcCoTaso
Ftows(iim
Y. Takashima et al. / Neuroscience 169 (2010) 828–842 839
argeting and suggests that peripheral innervation could beore important for specification of TRPM8 neurons.
rowth factor receptor expression phenotype ofRPM8 neurons during development
eurotrophic growth factor receptors and their correspond-ng soluble ligands are the earliest recognized markers ofensory neuron subtypes and are critical for the establish-ent of neuronal phenotypes (Smeyne et al., 1994; Silos-antiago et al., 1995; Marmigere and Ernfors, 2007). Smalliameter thermosensory and nociceptive neurons largelyxpress the NGF receptor TrkA, and several elegant stud-
es have shown that the majority of DRG sensory neuronsre dependent on NGF for their survival early in develop-ent (Marmigere and Ernfors, 2007). Surprisingly, only a
mall percentage of Trpm8GFP neurons express TrkA inarly development (E16.5 and E18.5) with expressionradually increasing throughout development. These dataere somewhat of surprise for two reasons. First, TrkAxpression is known to decrease during development,hereas here we find that TRPM8 neuronal expression ofrkA increases. Secondly, in adult mice Trpm8GFP neu-ons do not label with IB4, suggesting that these neuronso not fit the classification of “non-peptidergic” by currenteurochemical standards (Takashima et al., 2007). There-ore, we expected a higher percentage of TRPM8 neuronso be TrkA dependent. However, this result is consistentith a previous report where NGF�/�; Bax�/� micehowed reduced expression of TRPM8 in P0 by in situybridization, but TRPM8 expression is not completelyliminated (Luo et al., 2007). Thus, the data suggest thatot all Trpm8GFP neurons are dependent on NGF for sur-ival. Moreover, we did not observe any Trpm8GFP neu-
ig. 10. Co-expression of TRPM8 with A�- and C-fiber markers. Quan-ification of the percent of Trpm8GFP neurons that co-label with peripherinr NF200-IR. Among Trpm8GFP neurons co-staining for peripherin thereere 5.4% at E18.5 (64/1185 in 12 sections), 13.7% at P0 (96/701 in 7ections), 20.0% at P3 (660/3300 in 33 sections), and 18.5% at P14351/1897 in 19 sections). For NF200 there were 0.1% at E18.5 (2/2000n 14 sections), 0.3% at P0 (3/1000 in 10 sections), 0.2% at P3 (10/5000n 57 sections), and 21.1% at P14 (211/1000 in 10 sections). Data are
eans�SEM, * denotes P�0.05 and ** denotes P�0.01.
ons immunoreactive for Ret at P0 and P3 and little to no 2
o-localization at P14. This suggest that Trpm8GFP neu-ons are not part of the TrkA sub-population which switcho Ret dependence after birth, supporting data showinghat TRPM8 expression is unaffected in Retf/f; Wnt1-Cre
ice (Luo et al., 2007). This further supports the hypo-hesis that Ret functions downstream of Runx1, dueo the fact that expression of TRPM8 is eliminated inunx1�/�DRGs (Chen et al., 2006). Additionally, we see little
o no co-localization between Trpm8GFP neurons and eitherrkB or TrkC at any given developmental stage. This sug-ests that TRPM8-expressing neurons are neither mechano-eceptive nor proprioceptive neurons. Moreover, it also sug-ests that TRPM8-expressing neurons do not switch theireurotrophic factors dependency throughout the develop-ent.
eurochemical phenotype of TRPM8 neurons duringevelopment
e have previously shown that TRPM8 neurons express aide cohort of neurochemical markers in adult mice, dem-nstrating that these cells are varied molecularly (Zirlingert al., 2002; Takashima et al., 2007). This phenotype alsouggests that these neurons are also functionally diverseonsistent with TRPM8-null phenotypes (Bautista et al.,007; Colburn et al., 2007; Dhaka et al., 2007; Daniels etl., 2009). We asked if this diversity is present duringevelopment to understand the developmental basis forhe broad range of cold sensations mediated by TRPM8.hermosensitive and nociceptive DRG neurons can berossly categorized into several distinct subsets based oneuronal marker expression. For instance, first and secondain sensations in teeth are attributed to activation of A�- and-fibers, respectively (Jyvasjarvi and Kniffki, 1987; Mengel etl., 1993). These subtypes can be largely identified by thexpression of NF200 and peripherin for A�- and C-fibers,espectively, and we have shown previously that approxi-ately one-quarter of TRPM8 neurons co-express each of
hese markers in the adult (Takashima et al., 2007). Oururrent results show Trpm8GFP neurons do not co-localizeith NF200� until after day P3 (between P3 and P14). This
ncrease in co-localization with NF200 coincides with a re-orted postnatal increase in the number of NF200� rat neu-ons which occurs between days P0 and P3 (Beland anditzgerald, 2001). Conversely, Trpm8GFP expression in C-bers identified by peripherin immunoreactivity is observeds early as E18.5 and increases to adult levels by P3. How-ver, as in the adult, a large percentage of Trpm8GFP neu-ons do not label with either marker and thus cannot beistinguished as either fiber neurochemical phenotype.
In addition tomarkers of fiber-type, we also determinedhe expression patterns of Trpm8GFP DRG neurons inomparison to nociceptor markers, such as TRPV1 andGRP. In adult mice, we have shown that approximatelyne-quarter of Trpm8GFP DRG neurons express eitherRPV1 or CGRP (Takashima et al., 2007). These resultsre in agreement with functional studies in cultured sen-ory neurons from both rat and mouse in which there isverlap in menthol and capsaicin sensitivity (Guo et al.,
001; McKemy et al., 2002; Viana et al., 2002; Xing et al.,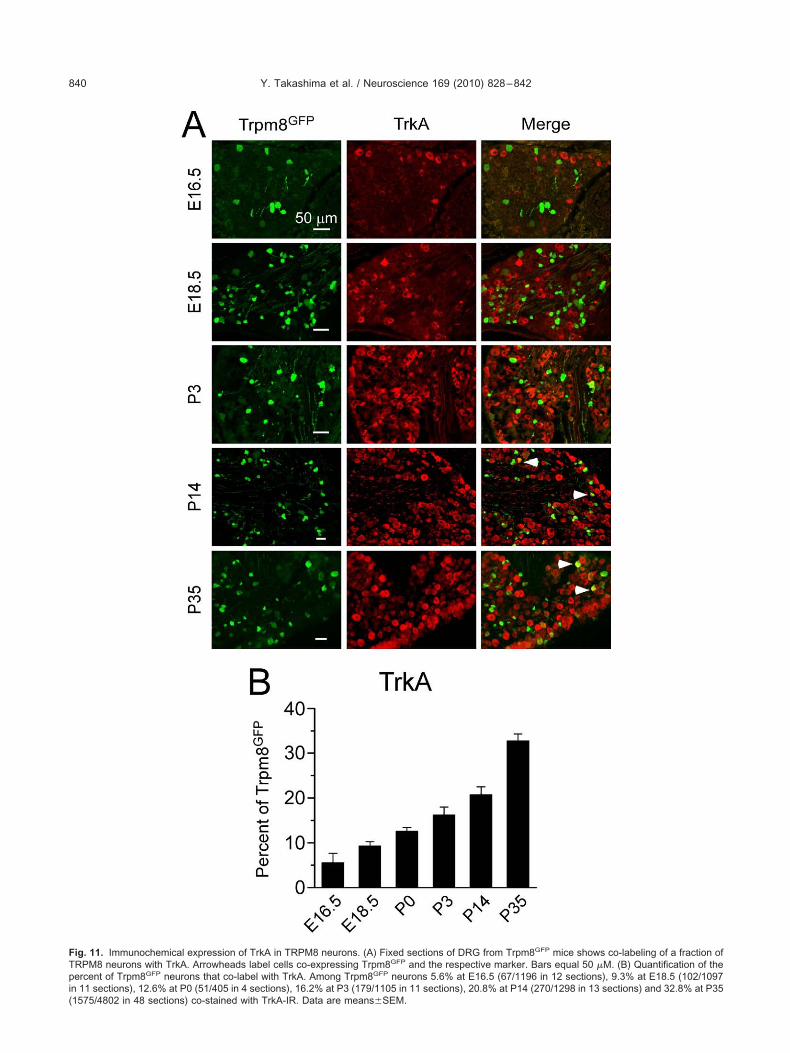
FTpi(
Y. Takashima et al. / Neuroscience 169 (2010) 828–842840
ig. 11. Immunochemical expression of TrkA in TRPM8 neurons. (A) Fixed sections of DRG from Trpm8GFP mice shows co-labeling of a fraction ofRPM8 neurons with TrkA. Arrowheads label cells co-expressing Trpm8GFP and the respective marker. Bars equal 50 �M. (B) Quantification of theercent of Trpm8GFP neurons that co-label with TrkA. Among Trpm8GFP neurons 5.6% at E16.5 (67/1196 in 12 sections), 9.3% at E18.5 (102/1097
n 11 sections), 12.6% at P0 (51/405 in 4 sections), 16.2% at P3 (179/1105 in 11 sections), 20.8% at P14 (270/1298 in 13 sections) and 32.8% at P35
1575/4802 in 48 sections) co-stained with TrkA-IR. Data are means�SEM.
28sldcTesTst
dpaacfiweTotiHTrnncnpeg
ItFcwtetapitatmce1sdr
ftsn
Atttm
A
A
B
B
B
B
B
C
C
C
C
C
D
D
D
D
D
F
Y. Takashima et al. / Neuroscience 169 (2010) 828–842 841
006; Hjerling-Leffler et al., 2007). Indeed, as many as0% of acutely cultured P0 DRG neurons that are menthol-ensitive are reported to also be capsaicin sensitive (Hjer-
ing-Leffler et al., 2007). This overlap in agonist sensitivityrops to nearly 50% by P7 after which these values stayonstant. Thus, these data suggest that TRPM8 andRPV1 neurons arise from an embryonically related lin-age (Hjerling-Leffler et al., 2007). However, our datahow a dramatic decrease in co-localization betweenRPM8 and TRPV1 after birth. Interestingly, co-expres-ion of Trpm8GFP and TRPV1 was biphasic in nature inhat by day P14, co-expression returned to adult levels.
Unlike the other markers examined, CGRP expressionoes not begin until late in embryonic development (ap-roximately at E18) and coincides with peripheral DRGxons reaching their target tissues, such as the skin (Ai etl., 1999). Thus, as predicted we observed little to noo-expression of Trpm8GFP and CGRP at E18.5, but didnd that these two markers are co-expressed postnatally,ith expression reaching adult levels by P14 (Takashimat al., 2007). These data suggest that the phenotype ofRPM8 neurons during the first week of postnatal devel-pment is largely non-nociceptive, but that by P14 a frac-ion of these neurons have developed qualities character-stic of nociceptors, which are maintained into adulthood.owever, although our data suggests that early postnatalRPM8 neurons are presumptive non-nociceptive neu-ons, the IB4, cRet, and P2X3 data suggest that they areot “classical” non-peptidergic neurons. It is interesting toote that a significant proportion of Trpm8GFP neuronsannot be classified into a particular subset in that they doot express markers such as TRPV1, CGRP, NF200, oreripherin, suggesting that this small population of affer-nts (�2% of all afferent neurons within a respective gan-lia) can only be distinguished by TRPM8 expression.
CONCLUSION
n conclusion, our data from TRPM8 axon projection pat-erns and functional synaptic connection determined byos expression show that TRPM8-dependent functionalold circuits are not established until at least the secondeek of postnatal development. Moreover, during postna-
al development the neurochemical phenotype of TRPM8-xpressing neurons changes dramatically until the same
ime period when functional cold circuits are established,pproximately at P14. This suggests that neurochemicalhenotype correlates with the establishment of cold-sens-
ng circuits. Thus, our results from the time course oferminal formation, the functional synaptic connections,nd the developmental neurochemical phenotype suggest
hat maturation of cold circuitry is completed at P14 inice. This process seems to be completed soon after
onnections are formed between A-fiber dorsal root affer-nts, interneurons, and motor neurons (Narayanan et al.,971; Dahlhaus et al., 2005). All of the data combineduggests that maturation of cold circuits could be activity-ependent. Additionally, our data suggest that those neu-
ons which express TRPM8 in DRG are committed to theirate before they form contacts with their future centralargets. Here, we provided an insight that brings us onetep closer to determining how different types of sensoryeurons could be established during development.
cknowledgments—This work was supported by a National Insti-utes of Health Grant NS054069 (D.D.M.). We thank members ofhe McKemy lab for their helpful insights and discussions duringhis study and also thank Q. Wu for help with embryonic wholeounts. We thank L. Reichardt for providing TrkA antibodies.
REFERENCES
i X, Cappuzzello J, Hall AK (1999) Activin and bone morphogeneticproteins induce calcitonin gene-related peptide in embryonic sen-sory neurons in vitro. Mol Cell Neurosci 14:506–518.
i X, MacPhedran SE, Hall AK (1998) Depolarization stimulates initialcalcitonin gene-related peptide expression by embryonic sensoryneurons in vitro. J Neurosci 18:9294–9302.
accei ML, Bardoni R, Fitzgerald M (2003) Development of nocicep-tive synaptic inputs to the neonatal rat dorsal horn: glutamaterelease by capsaicin and menthol. J Physiol 549:231–242.
asbaum AI, Bautista DM, Scherrer G, Julius D (2009) Cellular andmolecular mechanisms of pain. Cell 139:267–284.
autista DM, Siemens J, Glazer JM, Tsuruda PR, Basbaum AI, StuckyCL, Jordt SE, Julius D (2007) The menthol receptor TRPM8 is theprincipal detector of environmental cold. Nature 448:204–208.
eland B, Fitzgerald M (2001) Influence of peripheral inflammation onthe postnatal maturation of primary sensory neuron phenotype inrats. J Pain 2:36–45.
raz JM, Basbaum AI (2009) Triggering genetically-expressed trans-neuronal tracers by peripheral axotomy reveals convergent andsegregated sensory neuron-spinal cord connectivity. Neuro-science 163:1220–1232.
arr RW, Pianova S, McKemy DD, Brock JA (2009) Action potentialinitiation in the peripheral terminals of cold-sensitive neuronesinnervating the guinea-pig cornea. J Physiol 587:1249–1264.
aterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD,Julius D (1997) The capsaicin receptor: a heat-activated ion chan-nel in the pain pathway. Nature 389:816–824.
hen CL, Broom DC, Liu Y, de Nooij JC, Li Z, Cen C, Samad OA,Jessell TM, Woolf CJ, Ma Q (2006) Runx1 determines nociceptivesensory neuron phenotype and is required for thermal and neuro-pathic pain. Neuron 49:365–377.
oggeshall RE (2005) Fos, nociception and the dorsal horn. ProgNeurobiol 77:299–352.
olburn RW, Lubin ML, Stone DJ Jr, Wang Y, Lawrence D, D’AndreaMR, Brandt MR, Liu Y, Flores CM, Qin N (2007) Attenuated coldsensitivity in TRPM8 null mice. Neuron 54:379–386.
ahlhaus A, Ruscheweyh R, Sandkuhler J (2005) Synaptic input of ratspinal lamina I projection and unidentified neurones in vitro.J Physiol 566:355–368.
aniels RL, McKemy DD (2007) Mice left out in the cold: commentaryon the phenotype of TRPM8-nulls. Mol Pain 3:23.
aniels RL, Takashima Y, McKemy DD (2009) Activity of the neuronalcold sensor TRPM8 is regulated by phospholipase C via the phos-pholipid phosphoinositol 4,5-bisphosphate. J Biol Chem 284:1570–1582.
haka A, Earley TJ, Watson J, Patapoutian A (2008) Visualizing coldspots: TRPM8-expressing sensory neurons and their projections.J Neurosci 28:566–575.
haka A, Murray AN, Mathur J, Earley TJ, Petrus MJ, Patapoutian A (2007)TRPM8 is required for cold sensation in mice. Neuron 54:371–378.
itzgerald M (2005) The development of nociceptive circuits. Nat Rev
Neurosci 6:507–520.
F
G
H
H
J
J
J
L
L
L
M
M
M
M
M
M
M
M
M
N
P
P
S
S
S
S
S
S
S
T
T
V
V
W
X
Z
Z
Z
S
S
Y. Takashima et al. / Neuroscience 169 (2010) 828–842842
unakoshi K, Nakano M, Atobe Y, Goris RC, Kadota T, Yazama F(2006) Differential development of TRPV1-expressing sensorynerves in peripheral organs. Cell Tissue Res 323:27–41.
uo A, Simone DA, Stone LS, Fairbanks CA, Wang J, Elde R (2001)Developmental shift of vanilloid receptor 1 (VR1) terminals intodeeper regions of the superficial dorsal horn: correlation with a shiftfrom TrkA to Ret expression by dorsal root ganglion neurons. EurJ Neurosci 14:293–304.
jerling-Leffler J, Alqatari M, Ernfors P, Koltzenburg M (2007) Emer-gence of functional sensory subtypes as defined by transient re-ceptor potential channel expression. J Neurosci 27:2435–2443.
unt SP, Mantyh PW (2001) The molecular dynamics of pain control.Nat Rev Neurosci 2:83–91.
ordt SE, McKemy DD, Julius D (2003) Lessons from peppers andpeppermint: the molecular logic of thermosensation. Curr OpinNeurobiol 13:487–492.
ulius D, Basbaum AI (2001) Molecular mechanisms of nociception.Nature 413:203–210.
yvasjarvi E, Kniffki KD (1987) Cold stimulation of teeth: a comparisonbetween the responses of cat intradental A delta and C fibres andhuman sensation. J Physiol 391:193–207.
awson SN, Biscoe TJ (1979) Development of mouse dorsal rootganglia: an autoradiographic and quantitative study. J Neurocytol8:265–274.
awson SN, Waddell PJ (1991) Soma neurofilament immunoreactivityis related to cell size and fibre conduction velocity in rat primarysensory neurons. J Physiol 435:41–63.
uo W, Wickramasinghe SR, Savitt JM, Griffin JW, Dawson TM, GintyDD (2007) A hierarchical NGF signaling cascade controls Ret-dependent and Ret-independent events during development ofnonpeptidergic DRG neurons. Neuron 54:739–754.
andadi S, Nakanishi ST, Takashima Y, Dhaka A, Patapoutian A,McKemy DD, Whelan PJ (2009) Locomotor networks are targets ofmodulation by sensory transient receptor potential vanilloid 1 andtransient receptor potential melastatin 8 channels. Neuroscience162:1377–1397.
armigere F, Ernfors P (2007) Specification and connectivity of neuronalsubtypes in the sensory lineage. Nat Rev Neurosci 8:114–127.
cKemy DD (2007) Temperature sensing across species. PflugersArch 454:777–791.
cKemy DD, Neuhausser WM, Julius D (2002) Identification of a coldreceptor reveals a general role for TRP channels in thermosensa-tion. Nature 416:52–58.
cNeill DL, Coggeshall RE, Carlton SM (1988) A light and electronmicroscopic study of calcitonin gene-related peptide in the spinalcord of the rat. Exp Neurol 99:699–708.
engel MK, Stiefenhofer AE, Jyvasjarvi E, Kniffki KD (1993) Painsensation during cold stimulation of the teeth: differential reflectionof A delta and C fibre activity? Pain 55:159–169.
irnics K, Koerber HR (1995a) Prenatal development of rat primaryafferent fibers: I. Peripheral projections. J Comp Neurol 355:589–600.
irnics K, Koerber HR (1995b) Prenatal development of rat primaryafferent fibers: II. Central projections. J Comp Neurol 355:601–614.
olliver DC, Wright DE, Leitner ML, Parsadanian AS, Doster K, WenD, Yan Q, Snider WD (1997) IB4-binding DRG neurons switch fromNGF to GDNF dependence in early postnatal life. Neuron 19:849–861.
arayanan CH, Fox MW, Hamburger V (1971) Prenatal developmentof spontaneous and evoked activity in the rat (Rattus norvegicusalbinus). Behaviour 40:100–134.
eier AM, Moqrich A, Hergarden AC, Reeve AJ, Andersson DA, StoryGM, Earley TJ, Dragoni I, McIntyre P, Bevan S, Patapoutian A(2002) A TRP channel that senses cold stimuli and menthol. Cell
108:705–715. tolgar E, Fowler JH, McGill MM, Todd AJ (1999) The types of neuronwhich contain protein kinase C gamma in rat spinal cord. Brain Res833:71–80.
eal RP, Wang X, Guan Y, Raja SN, Woodbury CJ, Basbaum AI,Edwards RH (2009) Injury-induced mechanical hypersensitivityrequires C-low threshold mechanoreceptors. Nature 462:651–655.
hibasaki K, Murayama N, Ono K, Ishizaki Y, Tominaga M (2010)TRPV2 enhances axon outgrowth through its activation by mem-brane stretch in developing sensory and motor neurons. J Neurosci30:4601–4612.
ilos-Santiago I, Molliver DC, Ozaki S, Smeyne RJ, Fagan AM, Bar-bacid M, Snider WD (1995) Non-TrkA-expressing small DRG neu-rons are lost in TrkA deficient mice. J Neurosci 15:5929–5942.
imone DA, Kajander KC (1997) Responses of cutaneous A-fibernociceptors to noxious cold. J Neurophysiol 77:2049–2060.
meyne RJ, Klein R, Schnapp A, Long LK, Bryant S, Lewin A, Lira SA,Barbacid M (1994) Severe sensory and sympathetic neuropathiesin mice carrying a disrupted Trk/NGF receptor gene. Nature368:246–249.
nider WD (1998) How do you feel? Neurotrophins and mechano-transduction. Nat Neurosci 1:5–6.
tory GM, Peier AM, Reeve AJ, Eid SR, Mosbacher J, Hricik TR,Earley TJ, Hergarden AC, Andersson DA, Hwang SW, McIntyre P,Jegla T, Bevan S, Patapoutian A (2003) ANKTM1, a TRP-likechannel expressed in nociceptive neurons, is activated by coldtemperatures. Cell 112:819–829.
akashima Y, Daniels RL, Knowlton W, Teng J, Liman ER, McKemyDD (2007) Diversity in the neural circuitry of cold sensing revealedby genetic axonal labeling of transient receptor potential melastatin8 neurons. J Neurosci 27:14147–14157.
ominaga M, Caterina MJ, Malmberg AB, Rosen TA, Gilbert H, Skin-ner K, Raumann BE, Basbaum AI, Julius D (1998) The clonedcapsaicin receptor integrates multiple pain-producing stimuli. Neu-ron 21:531–543.
iana F, de la Pena E, Belmonte C (2002) Specificity of cold thermo-transduction is determined by differential ionic channel expression.Nat Neurosci 5:254–260.
ulchanova L, Riedl MS, Shuster SJ, Stone LS, Hargreaves KM, BuellG, Surprenant A, North RA, Elde R (1998) P2X3 is expressed byDRG neurons that terminate in inner lamina II. Eur J Neurosci10:3470–3478.
elberg L (2008) Sensory perception: one TRPM8 fits all. Nat RevNeurosci 9:79.
ing H, Ling J, Chen M, Gu JG (2006) Chemical and cold sensitivity oftwo distinct populations of TRPM8-expressing somatosensoryneurons. J Neurophysiol 95:1221–1230.
hao Z, Wang Z, Gu Y, Feil R, Hofmann F, Ma L (2009) Regulate axonbranching by the cyclic GMP pathway via inhibition of glycogensynthase kinase 3 in dorsal root ganglion sensory neurons. J Neu-rosci 29:1350–1360.
irlinger M, Lo L, McMahon J, McMahon AP, Anderson DJ (2002)Transient expression of the bHLH factor neurogenin-2 marks asubpopulation of neural crest cells biased for a sensory but not aneuronal fate. Proc Natl Acad Sci U S A 99:8084–8089.
ylka MJ, Rice FL, Anderson DJ (2005) Topographically distinct epi-dermal nociceptive circuits revealed by axonal tracers targeted tomrgprd. Neuron 45:17–25.
APPENDIX
upplementary data
upplementary data associated with this article can be found, in
he online version, at doi:10.1016/j.neuroscience.2010.05.039.(Accepted 16 May 2010)(Available online 24 May 2010)