the calm mouse: an animal model of stress reduction
TRANSCRIPT

6 0 6 | G U R F E I N E T A L . | M O L M E D 1 8 : 6 0 6 - 6 1 7 , 2 0 1 2
INTRODUCTIONEnvironmental stress is a pervasive
dimension of life that results in complexbiological changes. Evidence suggeststhat chronic stress can exert negative effects on general health, disease suscep-tibility, and progression of existing ill-ness (1). Networks that are stimulatedby stress include: the hypothalamic– pituitary–adrenal (HPA) axis, the sympa-
thetic adrenal medullary axis and sym-pathetic and parasympathetic nerve pro-jections that directly innervate secondarylymphoid organs (2–4). Repeated or pro-longed exposure to stress-related neu-roendocrine factors, such as glucocorti-coids and catecholamines, can potentlyinfluence immune function and is be-lieved to underlie the adverse health out-comes associated with chronic stress
(2,3). For example, human studies haveshown that chronically stressed individu-als exhibit poorer influenza vaccine re-sponses (5,6), enhanced susceptibility toexperimental rhinovirus infection (7) andaccelerated human immunodeficiencyvirus disease progression (8). Researchemploying animal models has demon-strated that chronic stress impairs innateand adaptive immunity (9–11) and canyield enhanced tumor growth (12,13) orincreased simian immunodeficiencyvirus–associated mortality (14). Otherwork has also linked stress and glucocor-ticoid production with hippocampal at-rophy and memory dysfunction in bothhumans and animals (15–17).
To mitigate the negative physiologiceffects of stress, stress management ap-proaches are often advocated by healthprofessionals. Stress reduction tech-
The Calm Mouse: An Animal Model of Stress Reduction
Blake T Gurfein,1,2 Andrew W Stamm,2 Peter Bacchetti,3 Mary F Dallman,4 Nachiket A Nadkarni,5,6
Jeffrey M Milush,2 Chadi Touma,7 Rupert Palme,8 Charles Pozzo Di Borgo,6 Gilles Fromentin,6Rachel Lown-Hecht,2 Jan Pieter Konsman,9 Michael Acree,1 Mary Premenko-Lanier,2 Nicolas Darcel,5,6
Frederick M Hecht,1 and Douglas F Nixon2
1Osher Center for Integrative Medicine, 2Division of Experimental Medicine, 3Department of Epidemiology and Biostatistics,4Department of Physiology, University of California, San Francisco, California, United States of America; 5Chaire ANCA, FoodNutrition and Eating Behavior, 6INRA, Unit 914 Nutrition Physiology and Ingestive Behavior, AgroParisTech, Paris, France;7Research Group of Psychoneuroendocrinology, Max Planck Institute of Psychiatry, Munich, Germany; 8Department of BiomedicalSciences/Biochemistry, University of Veterinary Medicine, Vienna, Austria; 9Psychoneuroimmunology, Nutrition, and Genetics,CNRS UMR 5226, Victor Segalen Bordeaux 2 University, Bordeaux, France
Chronic stress is associated with negative health outcomes and is linked with neuroendocrine changes, deleterious effects oninnate and adaptive immunity, and central nervous system neuropathology. Although stress management is commonly advo-cated clinically, there is insufficient mechanistic understanding of how decreasing stress affects disease pathogenesis. Therefore,we have developed a “calm mouse model” with caging enhancements designed to reduce murine stress. Male BALB/c micewere divided into four groups: control (Cntl), standard caging; calm (Calm), large caging to reduce animal density, a cardboardnest box for shelter, paper nesting material to promote innate nesting behavior, and a polycarbonate tube to mimic tunneling;control exercise (Cntl Ex), standard caging with a running wheel, known to reduce stress; and calm exercise (Calm Ex), calmcaging with a running wheel. Calm, Cntl Ex and Calm Ex animals exhibited significantly less corticosterone production than Cntlanimals. We also observed changes in spleen mass, and in vitro splenocyte studies demonstrated that Calm Ex animals had in-nate and adaptive immune responses that were more sensitive to acute handling stress than those in Cntl. Calm animals gainedgreater body mass than Cntl, although they had similar food intake, and we also observed changes in body composition, usingmagnetic resonance imaging. Together, our results suggest that the Calm mouse model represents a promising approach tostudying the biological effects of stress reduction in the context of health and in conjunction with existing disease models.Online address: http://www.molmed.orgdoi: 10.2119/molmed.2012.00053
ND, FH, and DN are co–senior authors.Address correspondence to Blake T Gurfein, 1001 Potrero Avenue, Building 3 Room 609,UCSF Division of Experimental Medicine, San Francisco, CA 94110. Phone: 415-206-4981;Fax: 415-826-8449; E-mail: [email protected] February 10, 2012; Accepted for publication February 28, 2012; Epub(www.molmed.org) ahead of print February 29, 2012.

R E S E A R C H A R T I C L E
M O L M E D 1 8 : 6 0 6 - 6 1 7 , 2 0 1 2 | G U R F E I N E T A L . | 6 0 7
niques, such as mindfulness-based stressreduction, have been shown to reduceanxiety and depression, influence theHPA axis and may affect immunity(18–22). Although clinical studies ofstress management are informative, theydo not identify the mechanistic pathwaysthat are modulated in response to re-duced stress exposure, leaving uncer-tainty about how stress reduction influ-ences biology and which pathways areimportant to influence for specific healthoutcomes. Furthermore, prior animalstudies have focused primarily on in-creasing stress, and few animal modelsof stress reduction have been fully developed.
Here, we describe a “calm mousemodel” of stress reduction in which wehave altered several facets of the mouse-caging environment. Environmental en-richment protocols have been shown toreduce the negative effects of both psy-chogenic and neurogenic stress in rodentmodels (for review see 23). Studies prob-ing the stress-related effects of environ-mental enrichment have demonstratedthat animals housed in enriched cagingexhibit enhanced stress resiliency (24)and that reduced expression of amyg-dalar corticotropin releasing factor recep-tor type 1 is linked to the anxiolytic ef-fects of an enriched environment (25).Interestingly, published studies in whichenvironmental enrichment protocolswere used have revealed both increased(26–28) and decreased (25,29) corticos-terone production, a physiologic indica-tor of stress. We hypothesized that theincreased corticosterone levels found insome of these studies may have been dueto excessive cage complexity or frequentrotation of enrichment items within acage. Thus, we omitted those elementsfrom our design and carefully selectedcaging enhancements with the intentionof minimizing stress levels. For example,increasing the amount of floor space peranimal within a cage is associated withreduced glucocorticoid production andadrenal gland mass (30). Other work hasshown that mice prefer cages that areequipped with enhanced nesting mate-
rial and that shredded paper strips aremost similar to materials that mice use tobuild three-dimensional nests in naturalsettings (31). Flat-roofed cardboard nestboxes and polycarbonate tubes provideauxiliary hiding spaces and increase cagecomplexity, characteristics that mice pre-fer (32). Lastly, in humans, cardiovascu-lar exercise is associated with stress re-duction (33), and animal studies haveshown that mice will carry out tasks, likebar pressing, to engage in voluntarywheel running, which suggests that ac-cess to a running wheel is a rewardingenhancement (34).
In this study, male BALB/c mice wereorganized into four arms: control (Cntl),standard caging; calm (Calm), largecaging, a cardboard nest box, papernesting material, and a polycarbonatetube; control exercise (Cntl Ex), stan-dard caging with a running wheel; andcalm exercise (Calm Ex), Calm cagingwith a running wheel. We examinedphysiological, biological and behavioralparameters in mice housed in each con-dition to investigate whether Calmcaging, access to an exercise wheel, orthe combination of both caging en-hancements could effectively reducestress levels in mice. Findings from thisstudy indicate that the Calm mousemodel achieved reductions in physio-logic stress measures and is a promisingapproach to modeling the biological ef-fects of stress reduction.
MATERIALS AND METHODS
AnimalsForty 8-wk-old male BALB/c mice
were purchased from Charles River (Hol-lister, CA, USA) and housed in the Labo-ratory Animal Resource Center (LARC)at San Francisco General Hospital. Cageswere kept in a temperature-controlledroom (22°C) with a light–dark 12:12 cycle(light on 0600–1800 h). All studies wereapproved by the University of CaliforniaSan Francisco Institutional Animal Careand Use Committee and were conductedin accordance with national guidelines ofhumane laboratory animal care.
DietAll cages were provided with water
and PicoLab Rodent Diet 20 (LabDiet,PMI Nutrition International, St. Louis,MO, USA) ad libitum. Percent caloriesprovided by macronutrients were: pro-tein, 24.651%; fat, 13.205%; carbohy-drates, 62.144%; and crude fibercontents: not more than 6.0%.
TimelineAfter arrival, all mice were group
housed (five animals per cage) in stan-dard control caging for a period of 14 d(d –14 to 0) to allow them to acclimatizeto the LARC husbandry environment.After acclimatization, animals wereplaced in their respective caging condi-tions and remained there for an addi-tional 70 d. The elevated plus maze(EPM) was carried out at d 42. Animalswere weighed during weekly cage clean-ing and measures of dry food consump-tion were recorded by cage. Additionally,fecal samples were taken from each ani-mal at regular intervals and kept at–20°C for batched fecal corticosteroneanalysis (see below).
Caging and EnrichmentMice were randomly assigned to four
groups (n = 10 per group): Cntl, Calm,Cntl Ex or Calm Ex. For all groups, eachcage housed five animals (two cages pergroup). All caging, bedding and enrich-ment items were autoclaved before useor sterilized with Coverage disinfectantspray (Steris, Mentor, OH, USA). Stan-dard wire-bar lids for food and waterand filter-top bonnets were used for all cages. Cages for the Cntl group (189 mm × 297 mm × 128 mm; 484 cm2
surface area; Allentown Inc., Allentown,NJ, USA) contained standard Paperchipbedding (Shepherd Specialty Papers,Milford, NJ, USA) and lacked enhance-ments. Cages for the Calm group werelarger (257 mm × 483 mm × 152 mm;980 cm2 surface area) and containedstandard bedding, 1 paper nest box (Bio-Serv, Frenchtown, NJ, USA), 1 red poly-carbonate mouse tunnel (Bio-Serv) and236 cm3 (1 cup) of compressed Enviro-
T H E C A L M M O U S E M O D E L O F S T R E S S R E D U C T I O N

6 0 8 | G U R F E I N E T A L . | M O L M E D 1 8 : 6 0 6 - 6 1 7 , 2 0 1 2
T H E C A L M M O U S E M O D E L O F S T R E S S R E D U C T I O N
dri Eco-bedding shredded paper strips(FiberCore, LLC, Clevland, OH, USA).Cages for the Cntl Ex group containedthe same amount of surface area as Cntlcages (312 × 235 × 152 mm; 484 cm2 sur-face area), standard bedding and agreen Silent Spinner running wheel(Super Pet, Elk Grove Village, IL, USA).Cages for the Calm Ex group were thesame size (980 cm2 surface area) andcontained the same enhancements asCalm caging with the addition of agreen Silent Spinner running wheel(Super Pet). Once weekly, cages andwashable enrichment items werecleaned, and bedding, Enviro-dri stripsand nest boxes were replaced. Note:Green running wheels were used inthese experiments; however, identicalrunning wheels in red color were usedfor display in Figures 1D, G, H. To fit therunning wheel in Cntl Ex caging, themetal stand was removed and the plasticwheel mount was sawed and sanded flatto facilitate adhering of the wheel mountto the inner cage wall. The wheel mountwas scored with a utility knife andbonded to the cage wall by use of waterresistant epoxy glue and left to dry for10 d before use.
BehaviorAt d 42 (08:00), each animal was
placed on an EPM and behavior was re-corded via digital video camera for 5 min(Stoelting Co., Wood Dale, IL, USA). Themaze was elevated to a height of 40 cmwith 2 open arms (5 × 35 cm) and2 closed arms (5 × 35 × 15 cm). Lightingconditions were kept low, with the cen-ter platform of the maze measuring<200 lux. At the beginning of each exper-iment the mouse was placed on the cen-ter platform facing an open arm andafter each run the platform was thor-oughly cleaned with disinfectant. Behav-ior analysis was carried out by twoblinded observers and the results wereexpressed as: percent time spent in theopen arms (open-arm time/[open-armtime + closed-arm time]), percent timespent in the closed arms. Placement of allfour limbs within a maze arm or the cen-
ter platform constituted an entry. Dis-tance traveled was measured using semi-automated object tracking in ImageJv1.43n software (ImageJ; National Insti-tutes of Health [NIH], Bethesda, MD,USA). Data were expressed in distancetraveled as percent of Cntl.
Body CompositionWe determined body composition
using magnetic resonance imaging(MRI). At the experimental endpoint, an-imals were killed and then frozen at–20°C until imaging. Animals were fullythawed overnight and imaged at 23°C.Imaging was performed with AgilentVnmrJ software v3.1 and a 7 Tesla(310-mm bore size) superconductingmagnet equipped with actively shieldedimaging gradients (400 mT/m maximumgradient strength, 120-mm inner boresize; Agilent Technologies, Palo Alto, CA,USA). A 72-mm inner diameter quadra-ture 1H birdcage resonator (RAPID Bio-medical GmbH, Rimpar, Germany) wasused for radiofrequency pulse transmis-sion and signal reception. Mice were positioned in the middle of the coil toachieve a complete radiofrequency fieldcovering of the animal. The location ofeach animal was checked with a localizersequence (gradient-echo, repetition time[TR] = 10 ms; echo time [TE] = 3.4 ms;field of view [FOV] = 120 × 120 mm2; ac-quisition matrix = 512 × 512 mm2, slicethickness = 2 mm; three orthogonalslices). A global iterative shim procedurewas used to improve the field homo-geneity. Lipid-sensitive imaging was per-formed by using a three-dimensionalfast-spin–echo sequence with the follow-ing parameters: TR = 200 ms; effectiveecho time (TEeff) = 45.1 ms; TE = 5.64 ms;echo train length = 16; FOV = 100 ×37.5 × 37.5 mm3; acquisition matrix =256 × 96 × 96 mm3; resolution = 390 ×390 × 390 µm3; receiver bandwidth =156.2 kHz; number of averages = 4; num-ber of dummy scans = 2; total acquisitiontime = 7 min 42 s. Image analysis wasfully automatic and performed in Med-ical Imaging Processing, Analysis and Vi-sualization (NIH). The fuzzy c-means al-
gorithm was used to hard-segment theimage into five tissue classes, the bright-est of which was a reliable fat mask. Thiswas converted into a volume of interestand volume measured and expressed aslipid volume per unit body mass(mm3/g).
Fecal Corticosterone MetabolitesDuring cage changing, animals were
temporarily placed in individual contain-ers, and fecal pellets produced at thistime were collected and stored at –20°C.Glucocorticoid metabolite quantificationin fecal samples has been established in alarge number of species and has been ex-tensively validated for laboratory mice(35–37). Collected samples were ana-lyzed for immunoreactive corticosteronemetabolites by use of a 5α-pregnane-3β,11β,21-triol-20-one enzyme immuno-assay (EIA) as described previously(35,38). Briefly, fecal samples were ho-mogenized and aliquots of 0.05 g wereextracted with 1 mL of 80% methanol asdescribed (35). The EIA used a double-antibody technique and was performedon anti-rabbit immunoglobin G–coatedmicrotiter plates. After overnight incuba-tion (4°C) of standards (range: 0.8–200 pg/well) and samples with steroid antibodyand biotinylated label, the plates wereemptied, washed and then blotted drybefore a streptavidin horseradish peroxi-dase conjugate was added. After a 45-min incubation, plates were emptied,washed and blotted dry. The substrate(tetramethylbenzidine) was added andincubated for another 45 min at 4°C be-fore the enzymatic reaction was stoppedwith 1 mol/L sulphuric acid. Then, theoptical density (at 450 nm) was recordedwith an automatic plate reader and thehormone concentrations were calculated.Samples were run in one batch to en-hance data measure consistency. Theintra- and interassay coefficients were8.8% and 13.4%, respectively.
In Vitro StudiesAt the experimental endpoint, d 70,
animals were killed one at a time via car-diac puncture while under deep isoflu-

R E S E A R C H A R T I C L E
M O L M E D 1 8 : 6 0 6 - 6 1 7 , 2 0 1 2 | G U R F E I N E T A L . | 6 0 9
rane anesthesia. This process requiredapproximately 10–15 min per cage.Depth and duration of anesthesia wassimilar for all animals. Sample collectionwas done within 2 h of lights on in thehusbandry facility. Spleens were takenand cells were mechanically dissociated.Cell number was counted with standardmethods after red blood cell lysis (eBio-science, San Diego, CA, USA). Purifiedsplenocytes were plated at 2.5 × 105 cellsper well in 200 µL of culture medium ina flat-bottom or round-bottom 96-wellplate. Standard splenocyte culture me-dium was used with the addition of 5%charcoal-stripped fetal bovine serum (FBS),which contains minimal quantities ofhormonal agonists of the glucocorticoidreceptor. Cells were stimulated with LPS(1 µg/mL; Sigma Aldrich, St. Louis, MO,USA), CD3/CD28-coated Dynal beads (1bead per cell; Invitrogen, Carlsbad, CA,USA), or vehicle control and treatedwith several concentrations of corticos-terone (0.005 µmol/L; 0.025 µmol/L;0.05 µmol/L; 0.075 µmol/L; SigmaAldrich). All conditions were run in du-plicate. At 18 h supernatants were col-lected and frozen at –80°C and laterused to measure quantities of interleukin6 (IL-6) and IL-2 via enzyme-linked im-munosorbent assay (ELISA) according tothe manufacturers’ instructions (R&D,Minneapolis, MN, USA).
Serum AssaysAt the experimental endpoint, whole
blood was collected via cardiac puncture(see In Vitro Studies above), within 2 h oflights on, and supernatants were frozenat –20°C. ELISA kits were used as in-structed by the manufacturer to measureserum levels of corticosterone (IBL Amer-ica, Minneapolis, MN, USA), insulin(Millipore, Billerica, MA, USA) and insulin-like growth factor 1 (IGF-1)(R&D).
Statistical AnalysisAnalyses were performed with Prism
software v5.0a (GraphPad Inc., SanDiego, CA, USA). Repeated measuresanalysis of variance (ANOVA) was used
for body mass analysis with SAS PROCGLM v9.2 (Figure 2A). Unpaired t testswere used to compare experimentalgroups with Cntl for: body mass (Fig-ures 2B–D), adiposity (Figure 2G),serum IGF-1 production (Figure 2H),corticosterone metabolite (CM) level(Figure 3), cytokine production (Fig -ures 4B, C) and behavior (Figures 5A, B).Two-way ANOVA was used for mea-suring differences in energy intake at individual time points (statistical signif-icance displayed for main effect of exer-cise, Figure 2E) and spleen mass
(Figure 4A). Linear regression followedby two-tailed statistical comparison ofregression coefficients was used to com-pare the relationship between serum in-sulin level and final body mass betweengroups (Figure 2J). Similarly, regressionlines were used to illustrate the associa-tion between cytokine production andendpoint serum corticosterone level(Figures 4D, E). In all cases, P < 0.05was considered statistically significant.
All supplementary materials are availableonline at www.molmed.org.
Figure 1. Enhanced caging and experimental design. Enrichment items were placed inexperimental cages depending on condition and included a cardboard nest box (A),shredded paper nesting material (B), polycarbonate tube (C) and running wheel (D).Cage contents were arranged as described in Materials and Methods, and as pictured(E–H). (E) Standard caging (484 cm2 surface area) lacking enrichment items was used tohouse Cntl animals. (F) Larger caging (980 cm2) was used for Calm animals and thesecages contained a corner-fitting cardboard nest box, shredded nesting material and apolycarbonate tube. (G) Cntl Ex caging (484 cm2) was similar to Cntl, but included a run-ning wheel. (H) Calm Ex caging was also larger (980 cm2) and contained a nest box,shredded nesting material, a polycarbonate tube and a running wheel. (I) Experimentaltimeline; 8-wk-old male BALB/c mice (n = 10 per group) acclimatized to the husbandry fa-cility in standard caging for 14 d. Animals were randomized and placed in respective ex-perimental cages (five per cage) for 70 d. EPM studies were carried out at d 42.
6 1 0 | G U R F E I N E T A L . | M O L M E D 1 8 : 6 0 6 - 6 1 7 , 2 0 1 2
T H E C A L M M O U S E M O D E L O F S T R E S S R E D U C T I O N
RESULTS
Generating a Stress-ReducingEnvironment
Items that were placed in enhancedcages included a cardboard nest box,shredded paper nesting material, and apolycarbonate mouse tube (Figures 1A–C).Additionally, certain cages contained asilent running wheel (Figure 1D).Eight-week-old BALB/c mice wereused in these studies. Although femalesare known to be less territorial and ag-gressive, we used male mice to circum-vent estrous cycle variability and its ef-fects on hormonal regulation. Micewere randomly assigned to one of fourcaging groups (n = 10 per group): Cntl,Calm, Cntl Ex or Calm Ex. Cntl animalswere group housed (five animals percage) in standard caging without en-hancements (Figure 1E). Calm animalswere housed in larger caging contain-ing a cardboard nest box for shelter,paper nesting material to promote nest-ing behavior, and a polycarbonate tubeto mimic tunneling (Figure 1F). Ani-mals in Cntl Ex were housed in stan-dard caging equipped with a runningwheel (Figure 1G). Lastly, Calm Excaging included the same enrichmentsas Calm with the addition of a runningwheel (Figure 1H). See Materials andMethods for further details.
Before being placed in their respectiveexperimental caging environments, allmice were kept in Cntl caging for a pe-riod of 14 d to allow them to acclimatizeto the animal husbandry facility (Fig -ure 1I). After acclimatization, animalswere housed in experimental caging, according to their randomized group as-signments, until the end of the experi-ment. On d 42 all animals were sub-jected to assessment of anxiety-relatedbehavior on an EPM. At the end of theexperiment, on d 70, animals were killedfor blood collection, in vitro immuno-logic studies, and assessment of bodycomposition. At regular intervals fecalsamples were collected from each ani-mal and analyzed for corticosterone metabolites.
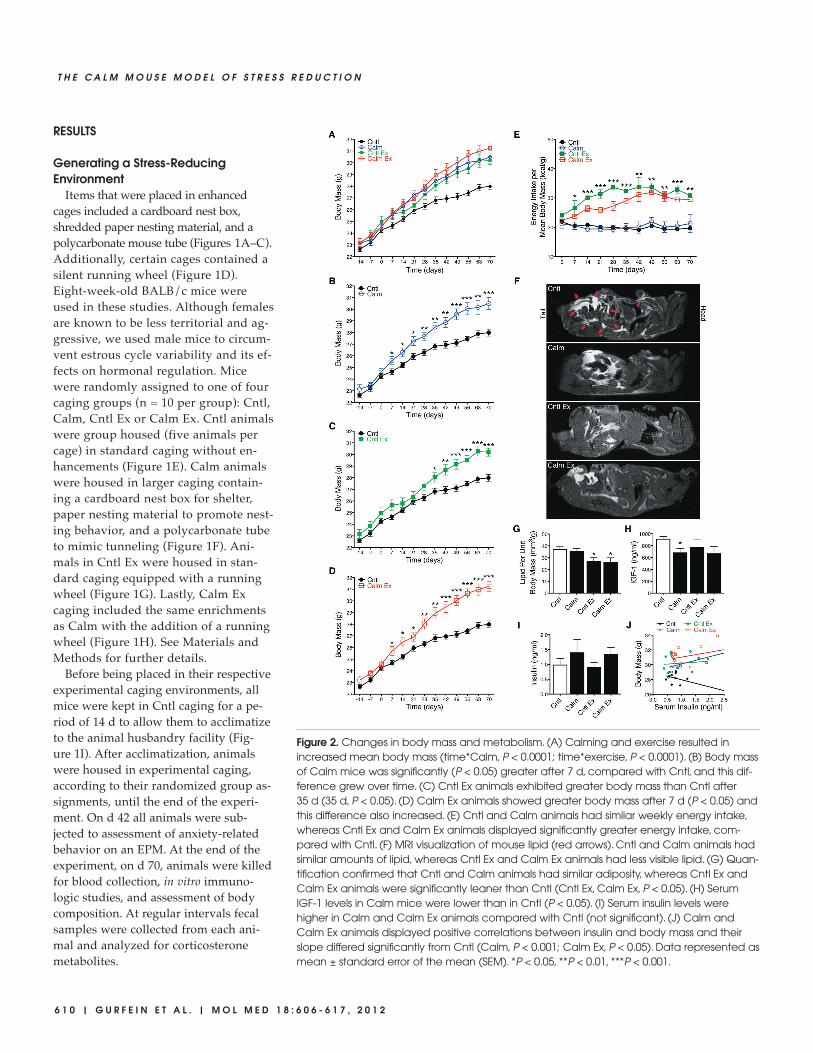
Figure 2. Changes in body mass and metabolism. (A) Calming and exercise resulted in increased mean body mass (time*Calm, P < 0.0001; time*exercise, P < 0.0001). (B) Body massof Calm mice was significantly (P < 0.05) greater after 7 d, compared with Cntl, and this dif-ference grew over time. (C) Cntl Ex animals exhibited greater body mass than Cntl after35 d (35 d, P < 0.05). (D) Calm Ex animals showed greater body mass after 7 d (P < 0.05) andthis difference also increased. (E) Cntl and Calm animals had similar weekly energy intake,whereas Cntl Ex and Calm Ex animals displayed significantly greater energy intake, com-pared with Cntl. (F) MRI visualization of mouse lipid (red arrows). Cntl and Calm animals hadsimilar amounts of lipid, whereas Cntl Ex and Calm Ex animals had less visible lipid. (G) Quan-tification confirmed that Cntl and Calm animals had similar adiposity, whereas Cntl Ex andCalm Ex animals were significantly leaner than Cntl (Cntl Ex, Calm Ex, P < 0.05). (H) SerumIGF-1 levels in Calm mice were lower than in Cntl (P < 0.05). (I) Serum insulin levels werehigher in Calm and Calm Ex animals compared with Cntl (not significant). (J) Calm andCalm Ex animals displayed positive correlations between insulin and body mass and theirslope differed significantly from Cntl (Calm, P < 0.001; Calm Ex, P < 0.05). Data represented asmean ± standard error of the mean (SEM). *P < 0.05, **P < 0.01, ***P < 0.001.
R E S E A R C H A R T I C L E
M O L M E D 1 8 : 6 0 6 - 6 1 7 , 2 0 1 2 | G U R F E I N E T A L . | 6 1 1
Calming and Exercise IncreasedBody Mass
Body mass was monitored weekly inall groups. We found that both calmingand exercise resulted in significantly(time*Calm, P < 0.0001; time*exercise, P <0.0001) increased body mass (Figure 2A).After 7 d of experimental caging, Calmanimals exhibited a significantly greatermean body mass compared with Cntl(7d, difference in means [Md] 0.94 g, 95%confidence interval [CI] 0.04–1.84, P =0.041), and this difference increased (70 d,Md 2.50 g, CI 1.23–3.77, P = 0.0006) overthe course of the experiment (Fig ure 2B).Animals housed in Cntl Ex caging alsohad a greater mean body mass than Cntlanimals (35d, Md 1.23 g, CI 0.16–2.37, P =0.027) (Figure 2C). Similar to Calm, CalmEx animals showed a greater mean bodymass than Cntl animals (7 d, Md 1.05 g, CI 0.10–2.00, P = 0.032), and increased thedifference over the duration of the experi-
ment (70 d, Md 3.26 g, CI 2.17–4.35, P <0.0001). Collectively, these findings sug-gest that both Calm caging and voluntaryexercise can induce rapid gains in bodymass in growing mice.
Energy Intake Was Affected byExercise
To further characterize the differencesin body mass described above, we inves-tigated changes in energy intake, bodycomposition and peptide hormone levelsin experimental and control animals(Figures 2E–J). Dry food consumptionwas monitored and recorded weekly bycage for the length of the experiment.These data are expressed as weeklycage-total energy intake per mean cagebody mass (kcal/g) at the end of eachweek to account for longitudinal in-creases in body mass. Remarkably, al-though we observed similar propor-tional weekly energy intake between the
Calm and Cntl groups (Figure 2E), Calmanimals exhibited significantly greaterbody mass than Cntl animals(Figure 2B). We also observed that CntlEx and Calm Ex animals consumedgreater quantities of food per week thanCntl and Calm animals (Figure 2E). Incomparing the two exercise groups, wenoted that for time points 14, 21, 28 and35 d, Cntl Ex animals displayed greaterenergy intake (Figure 2E) but less bodymass (Figure 2A) than Calm Ex animals,although these differences did not reachstatistical significance.
Calm Caging Had Little Effect on BodyComposition, whereas ExerciseReduced Adiposity
To determine whether the increasedbody mass in experimental groups wasattributable to differences in body com-position, we carried out an MRI protocol(Figures 2F, G). Animals were imaged byusing a three-dimensional fast-spin–echosequence that emphasized the lipid signalcontrast (Figure 2F, red arrows), facilitat-ing subsequent adipose tissue segmenta-tion and quantification (see Materials andMethods).
Cntl and Calm animals had similarquantities of lipid, whereas Cntl Ex andCalm Ex animals had less visible lipidthan Cntl animals (Figure 2F). Quantifi-cation of the proportion of adipose tissuewas expressed as total lipid volume pertotal body mass. These data revealed thatCntl and Calm animals had comparableadiposity (Md 1.56 mm3/g, CI –6.16 to9.29) and similar body composition (Fig-ure 2G). Cntl Ex and Calm Ex animals,however, exhibited a significantlysmaller proportion of adipose tissuecompared with Cntl (Cntl Ex, Md–10.15 mm3/g, CI –18.87 to –1.41, P =0.025; Calm Ex, Md –11.11 mm3/g, CI–21.37 to –0.85, P = 0.035), but were simi-lar to each other (Figure 2G). Together,these findings suggest that increasedbody mass in Cntl Ex and Calm Ex ani-mals may be partially attributable to in-creased lean muscle mass as a result ofvoluntary wheel running. However, Cntland Calm animals appeared to have sim-
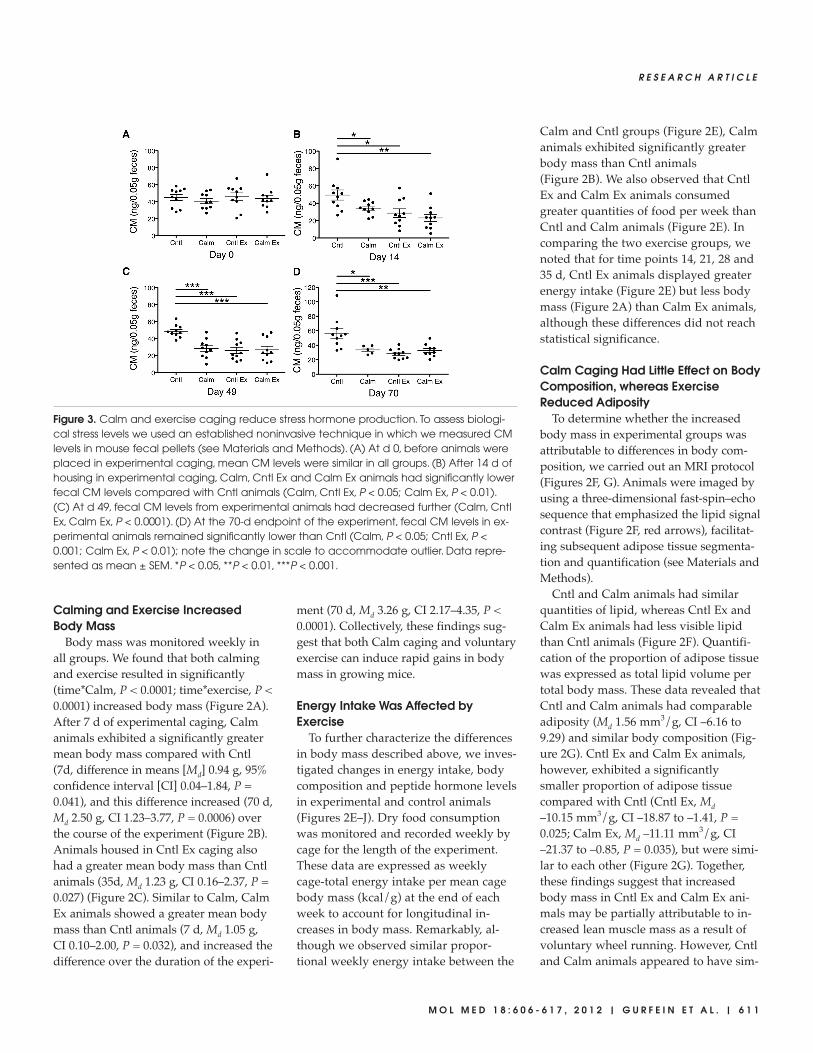
Figure 3. Calm and exercise caging reduce stress hormone production. To assess biologi-cal stress levels we used an established noninvasive technique in which we measured CMlevels in mouse fecal pellets (see Materials and Methods). (A) At d 0, before animals wereplaced in experimental caging, mean CM levels were similar in all groups. (B) After 14 d ofhousing in experimental caging, Calm, Cntl Ex and Calm Ex animals had significantly lowerfecal CM levels compared with Cntl animals (Calm, Cntl Ex, P < 0.05; Calm Ex, P < 0.01). (C) At d 49, fecal CM levels from experimental animals had decreased further (Calm, CntlEx, Calm Ex, P < 0.0001). (D) At the 70-d endpoint of the experiment, fecal CM levels in ex-perimental animals remained significantly lower than Cntl (Calm, P < 0.05; Cntl Ex, P <0.001; Calm Ex, P < 0.01); note the change in scale to accommodate outlier. Data repre-sented as mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001.

6 1 2 | G U R F E I N E T A L . | M O L M E D 1 8 : 6 0 6 - 6 1 7 , 2 0 1 2
T H E C A L M M O U S E M O D E L O F S T R E S S R E D U C T I O N
ilar body composition, a finding thatprovided little explanation of how Calmmice gained significantly more bodymass while maintaining the same pro-portional energy intake as Cntl animals.
Effects of Calming on PeptideHormones
To determine whether Calm-associatedbody mass differences were related tocirculating hormone concentrations, we
collected serum samples at the 70-d end-point and assayed them for IGF-1 and in-sulin (Figures 2H, I). IGF-1, a focal medi-ator of growth hormone, is produced bythe liver and can stimulate growth in
Figure 4. Caging environment affects the murine immune system. At the endpoint of the experiment animals were sacrificed and spleenswere taken for massing (A). Splenocytes were isolated as described in Materials and Methods, and were resuspended in medium con-taining 5% charcoal-stripped FBS, which has minimal hormone content. Cultures were treated with LPS (1 µg/mL), anti-CD3 plus anti-CD28(CD3/CD28) beads (1 bead/cell), or phosphate-buffered saline for 18 h. Some wells contained varying concentrations of corticosteroneto assess glucocorticoid-mediated suppression of cytokine production. Data displayed are from cultures containing 0.005 µmol/L corti-costerone, because this concentration is nearest to physiological levels (B–E). (A) Spleen mass for Calm, Cntl Ex and Calm Ex mice wasgreater than that for Cntl mice (Cntl Ex, P < 0.05; Calm, Calm Ex, P < 0.01). (B) In all groups, LPS-treated splenocytes produced comparablelevels of IL-6. (C) anti-CD3/anti-CD28 stimulation of splenocytes from Calm animals exhibited slightly greater production of IL-2 than Cntl(not significant), whereas Cntl Ex and Calm Ex animals showed little difference from Cntl. (D) LPS-stimulated splenocyte production of IL-6was positively correlated with serum corticosterone level at the time that animals were killed, but this relationship was significant only forCalm Ex animals (r2 = 0.7; P < 0.01). (E) Conversely, CD3/CD28 bead stimulation of IL-2 production was negatively correlated with serumcorticosterone level at the time that animals were killed. This relationship was significant in Cntl and Calm Ex animals (Cntl, r2 = 0.41, P <0.05; Calm Ex, r2 = 0.61, P < 0.01). Data represented as mean ± SEM. *P < 0.05, **P < 0.01.
R E S E A R C H A R T I C L E
M O L M E D 1 8 : 6 0 6 - 6 1 7 , 2 0 1 2 | G U R F E I N E T A L . | 6 1 3
most cell types, including muscle, boneand connective tissue. Previous studiesof enriched environments have reportedupregulation of IGF-1 in the rodent cen-tral nervous system (39), and thus, wehypothesized that serum IGF-1 might beupregulated as a product of Calm cagingor exercise. We found, however, thatcompared with Cntl animals, serum lev-els of IGF-1 were lower in Cntl Ex (Md–278 ng/mL, CI –897 to 342) and CalmEx (Md –489 ng/mL, CI –1032 to 54) andsignificantly lower (Md –459 ng/mL, CI–832 to 86, P = 0.019) in Calm animals(Figure 2H). We also measured serumlevels of insulin, the principal hormoneresponsible for regulating glucose metab-olism. We found that both Calm andCalm Ex animals exhibited trends to-ward higher levels of serum insulin com-pared with Cntl animals (Calm, Md0.42 ng/mL, CI –0.06 to 1.46; Calm Ex,Md 0.28, CI –0.10 to 0.65), although theresults lacked statistical significance (Fig-ure 2I). Given the functional role that in-sulin plays in metabolism and energy ho-meostasis, we reasoned that higher levelsof insulin might be associated with en-hanced ability to derive energy from di-gested dry food. To examine this rela-tionship, we looked at the associationbetween serum insulin level and finalbody mass within each group of ourstudy. We found that a linear regressionfor Cntl animals revealed a negative rela-
tionship between serum insulin level atthe end of the experiment and final bodymass (Cntl insulin effect –0.98 g perng/mL of insulin, CI –0.40 to –1.56, P =0.0018) (Figure 2J). However, Calm, CntlEx and Calm Ex regressions had positiveeffects, indicating that higher insulin lev-els were associated with greater bodymass (Calm insulin effect 0.99 g perng/mL of insulin, CI 0.58 to 1.41, P <0.0001; Cntl Ex insulin effect 0.26 g perng/mL of insulin, CI –1.04 to 1.56, P =0.69; Calm Ex insulin effect 0.67 g perng/mL of insulin, CI –0.43 to 1.78, P =0.22) (Figure 2J). The overall insulin bycaging group interaction was statisticallysignificant (P < 0.0001). These resultssuggest that caging-induced changes inbody mass were related to altered re-sponses to serum insulin level, althoughfurther work is needed to confirm thisassociation.
Both Calming and Exercise ReducedCorticosterone Production
The most widely used biomarker of ro-dent stress is the production of corticos-terone. Collecting blood from an animalto measure circulating corticosterone lev-els, however, may elicit a substantialstress response. For the purposes of as-sessing corticosterone production in thepresent study, we employed a less stress-ful, noninvasive technique in which wemeasured CMs found in fecal pellets
from each animal (Figure 3). CM mea-sures are well correlated with a gradedaverage of corticosterone productionfrom the preceding 10 h, and these re-sults have been extensively validated inlaboratory mice and other species(35–38,40). Fecal pellets were collectedfrom individual mice and were later as-sayed for CM as described in Materialsand Methods. We found that on d 0, be-fore animals were placed in their respec-tive experimental caging environments,the mean levels of CMs were similar inall groups (Figure 3A). After mice werehoused in Calm, Cntl Ex or Calm Excaging for 14 d, they exhibited signifi-cantly lower fecal CM levels (Calm, Md15.1 ng, CI 1.9–28.2, P = 0.027; Cntl Ex,Md 20.8 ng, CI 4.8–36.8, P = 0.014; CalmEx, Md 26.4, CI 11.4–41.4, P = 0.0017)compared with Cntl animals (Figure 3B).On d 49, mean fecal CM levels from ex-perimental animals were even lower(Calm, Md 20.5 ng, CI 11.7–29.4, P <0.0001; Cntl Ex, Md 22.5 ng, CI 13.4–31.5,P < 0.0001; Calm Ex, Md 21.8, CI11.7–32.0, P = 0.0003) and this effect per-sisted through d 70 (Calm, Md 23.1 ng, CI1.3–44.9, P = 0.039; Cntl Ex, Md 28.4 ng,CI 13.2–43.5, P = 0.0010; Calm Ex, Md23.9 ng, CI 7.4–40.4, P = 0.0070) the lastday of the experiment (Figures 3C, D).These findings suggest that both Calmcaging and caging equipped with a run-ning wheel can effectively reduce stresslevels as evidenced by lasting reductionsin CM production.
Immunologic Changes in MiceHoused in Calm and Exercise Caging
To characterize the effects of Calm andexercise caging on murine immune func-tion, we examined spleens and carriedout functional in vitro studies usingsplenocytes (Figure 4). At the experimen-tal endpoint animals were sacrificed andspleens were aseptically removed andmassed. We found that both calming andexercise generated a significant increase(Calm, P = 0.0086; exercise, P = 0.049) inspleen mass (Figure 4A).
Chronic social stress has been shownto affect innate and adaptive immunity
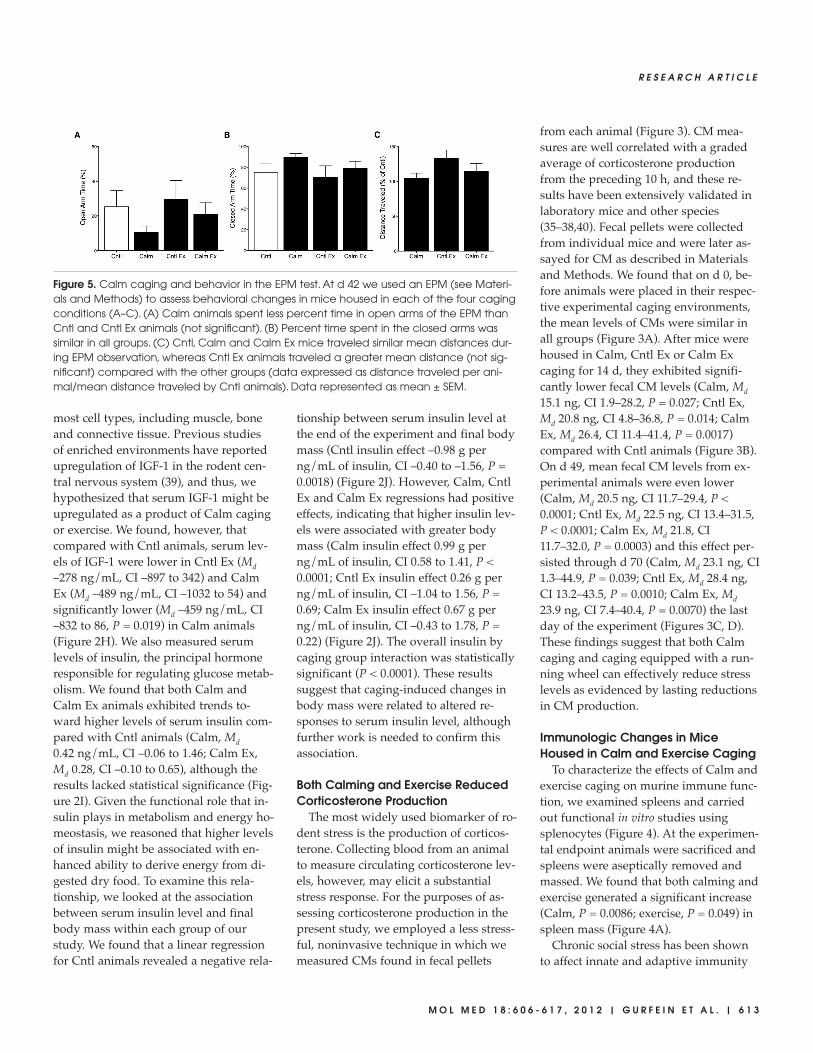
Figure 5. Calm caging and behavior in the EPM test. At d 42 we used an EPM (see Materi-als and Methods) to assess behavioral changes in mice housed in each of the four cagingconditions (A–C). (A) Calm animals spent less percent time in open arms of the EPM thanCntl and Cntl Ex animals (not significant). (B) Percent time spent in the closed arms wassimilar in all groups. (C) Cntl, Calm and Calm Ex mice traveled similar mean distances dur-ing EPM observation, whereas Cntl Ex animals traveled a greater mean distance (not sig-nificant) compared with the other groups (data expressed as distance traveled per ani-mal/mean distance traveled by Cntl animals). Data represented as mean ± SEM.

6 1 4 | G U R F E I N E T A L . | M O L M E D 1 8 : 6 0 6 - 6 1 7 , 2 0 1 2
T H E C A L M M O U S E M O D E L O F S T R E S S R E D U C T I O N
in humans and animals. For example,studies of chronic stress have revealedincreased production of IL-6 afterlipopolysaccharide (LPS) stimulation ofsplenocytes (41), and both reduced lym-phocyte proliferation and suppressedIL-2 production (5,42). In this work, toassess whether there were changes in in-nate or adaptive immune function westimulated splenocyte cultures with LPS(1 µg/mL) or anti-CD3 plus anti-CD28(CD3/CD28) Dynabeads (1 bead/cell),respectively. Some stimulated splenocytecultures were treated with different con-centrations of corticosterone, to assessleukocyte sensitivity to corticosterone-mediated suppression of cytokine pro-duction. All cells were cultured insplenocyte medium containing 5% char-coal-stripped FBS, which minimizesbackground hormone levels commonlyfound in serum and reduces unspecificbinding of the glucocorticoid receptor.Because murine immune cells usuallyfunction in the presence of a small con-centration of corticosterone (43), the re-sults displayed are from cultures con-taining 0.005 µmol/L corticosterone, themost physiologically relevant conditiontested (Figures 4B–E).
LPS-stimulated splenocyte culturesfrom Calm and Cntl Ex animals exhib-ited comparable levels of IL-6 to those ofCntl animals (Figure 4B). Cultures fromCalm Ex animals displayed a slight trendtoward greater production of IL-6 afterstimulation with LPS, but these datawere not statistically significant (Fig-ure 4B). Although splenocyte productionof IL-2 in response to CD3/CD28 beadstimulation was similar among Cntl, CntlEx and Calm Ex groups, cultures fromCalm animals had slightly higher (Md138.0 pg/mL, CI –18.8 to 296.1, P = 0.081)mean levels of IL-2 (Figure 4C).
Acute Stress and ImmunomodulationStress is known to be immunomodula-
tory and may enhance or suppress im-mune responses depending on the typeof stressor, acute or chronic, and the cellpopulations being studied, innate oradaptive (9,11,44,45). For example,
human studies have shown that acutepsychological stress results in increasedproduction of IL-6 in LPS-stimulated pe-ripheral blood mononuclear cells (46). Inthe context of animal studies, the han-dling of mice before sacrifice is a substan-tial acute stressor, resulting in increasedserum levels of stress mediators such ascorticosterone and epinephrine (47). Toassess the level of acute stress at the endof the experiment we measured the quan-tity of corticosterone found in serumsamples taken from mice at the time ofsacrifice. We observed a distinct pattern,in that within individual cages, the ani-mals killed first often had lower levels ofacute stress than subsequent animals, asindicated by serum corticosterone con-centration (Supplementary Figure 1).Thus, in each group there was a broadrange of serum corticosterone levels atthe time of sacrifice (Supplementary Fig-ures 1A–D). The within-group variationin serum corticosterone provided an op-portunity to investigate the effects ofvarying levels of acute stress on func-tional immunity in animals housed ineach of the four caging conditions.
To determine whether the level ofacute stress at the time of sacrifice hadany effect on cytokine production, wecarried out linear regression analyses toexamine the relationship between serumcorticosterone at the time of death and invitro stimulation of splenocyte cytokineproduction (Figures 4D, E). We foundthat all four groups displayed a positiveassociation between serum corticosteroneand LPS-stimulated production ofsplenocyte IL-6, but the correlation was astrong fit and statistically significant (P =0.0029) only for Calm Ex animals (Fig -ure 4D). In contrast, the overall trend forserum corticosterone and IL-2 productionafter CD3/CD28 bead stimulation was anegative correlation (Figure 4E). Similarto the results of IL-6 analysis, we foundthat the strongest and most significant (P = 0.0074) relationship between serumcorticosterone and IL-2 production waswithin the Calm Ex group (Figure 4E).
To investigate whether the above ob-servations were specific to the presence
of 0.005 µmol/L corticosterone in thesplenocyte cultures, we applied similarregression analyses to cultures that con-tained other concentrations of exogenouscorticosterone, including: 0, 0.025, 0.05,and 0.075 µmol/L (SupplementaryTable 1). These analyses showed that thepositive association between serum corti-costerone and splenocyte IL-6 productionremained intact at several other concen-trations of in vitro corticosterone and thatthe strongest relationships were found inCalm Ex animals (SupplementaryTable 1). Similarly, the negative correla-tions observed between serum corticos-terone and IL-2 production were foundat other concentrations of in vitro corti-costerone, and again, the Calm Ex groupdisplayed highly significant correlations(Supplementary Table 1).
In summary, animals housed in Calm,Cntl Ex and Calm Ex caging had greatermean spleen mass than Cntl, and our re-gression analyses suggest that the acutestress of handling had an enhancing ef-fect on innate immune function and asuppressive effect on adaptive immunefunction as determined by changes in cy-tokine production. Finally, Calm Ex ani-mals exhibited the greatest sensitivity toacute stress-mediated alteration of im-mune function.
Caging Environment and EPMBehavior
To assess the behavioral effects of hous-ing animals in enhanced caging, we ap-plied the EPM, a widely used test of ro-dent anxiety (Figure 5). The EPM is araised, plus-shaped apparatus consistingof two open arms and two arms with awalled enclosure, or closed arms. Theprinciple of the assay relies on the innateaversion that rodents have to open ele-vated alleys. Interpretation of EPM behav-ior is derived from original work showingthat rodents given anxiolytic agents spenta greater proportion of time in open armsof the maze (48). In pilot studies of theCalm model, we found that biomarkers ofstress decreased to the lowest levels after6 wks, and we hypothesized that behav-ioral anxiety might be reduced on a simi-

R E S E A R C H A R T I C L E
M O L M E D 1 8 : 6 0 6 - 6 1 7 , 2 0 1 2 | G U R F E I N E T A L . | 6 1 5
lar temporal scale. In the present work, at42 d each animal was placed in the EPMand behavior was digitally recorded for5 min and later analyzed by a blinded ob-server (see Materials and Methods).
We found that Calm animals spent asmaller proportion of time in open armsof the EPM than Cntl animals, althoughnone of the between-groups comparisonsreached statistical significance(Figure 5A). Similarly, the proportion oftime spent in the closed arms of the EPMwas not statistically significantly differ-ent between groups nor were the num-ber of open- or closed-arm entries (Fig-ure 5B, data not shown). Lastly, wemeasured the total distance traveled byeach animal, an indicator of exploration,and found that Cntl Ex animals traveledthe greatest distance during the course ofthe observation period, whereas Cntl,Calm, and Calm Ex mice traversed themaze comparably (Figure 5C).
DISCUSSIONIn this paper, we describe an animal
model that shows promise as a tool formodeling biological effects of stress re-duction. Our data demonstrate that ani-mals housed in stress reduction caging ex-hibited significant and lasting reductionsin fecal CM levels, a measure of physio-logic stress (Figure 3). Also, growing ani-mals housed in Calm, Cntl Ex, and CalmEx caging gained significantly greaterbody mass than Cntl animals (Figure 2).Strikingly, Calm animals gained greaterbody mass than Cntl despite comparableweekly energy intake (Figure 2). Lastly,immunologic studies showed that stress-reduction caging was associated with in-creased spleen mass and that splenocytesfrom Calm Ex animals were more sensi-tive to the immunomodulating effects ofacute stress (Figure 4).
Animal studies of stress commonlycite corticosterone as a key indicator ofstress level and have shown that subject-ing mice to chronic mild stress or chronicstress often results in elevations of corti-costerone level (49,50). In the presentwork, we used an established noninva-sive method in which we measured fecal
corticosterone metabolites from eachmouse. This approach allowed us tomonitor stress hormones longitudinallywithout inducing a stress response andalso provided us with reliable data thatreflect a 10-h graded average of corti-costerone production. Our resultsdemonstrated that Calm, Cntl Ex andCalm Ex animals exhibited significantreductions in fecal CM levels comparedwith Cntl, and that these differenceswere observed throughout the experi-ment. Interestingly, calming and exer-cise were both effective at reducing CMlevels. Toward the beginning of thestudy, at d 14, it appeared that the com-bination of Calm and exercise resultedin more rapid CM reductions than ei-ther approach alone. At subsequenttimes, however, CM seemed similar ineach of the modified caging groups.These findings suggest that a plateau ef-fect on reducing CM may have beenreached over time with each of thesecaging interventions. Collectively, theseresults show that animals housed inCalm and exercise caging exhibited re-duced biomarkers of stress, although,importantly, these results also under-score the relatively higher levels ofHPA-axis activity found in Cntl animals,which were housed in standard caging.
Manipulation of the mouse-caging en-vironment can evoke a variety of othermeasureable physiologic effects. In ro-dent studies, body mass is easily moni-tored and widely interpreted as an indi-cator of health. Chronically stressedanimals exhibit substantial reductions inbody mass, and disease models often usethe extent of body mass loss as a measureof morbidity (51,52). In the present work,we observed greater increases in bodymass in animals housed in Calm, Cntl Ex,and Calm Ex caging. We also measuredweekly energy intake and, as expected,found that Cntl Ex and Calm Ex mice hadgreater intake per week compared withCntl mice. However, after observing thebody mass differences between Cntl andCalm animals, we were surprised to findthat these groups had very similarweekly energy intake per body mass. Our
MRI analyses demonstrated that Cntl andCalm animals had similar proportions oflipid to body mass, indicating similarbody composition. We then hypothesizedthat the body mass changes in Calm micemight be attributable to endocrinechanges. Measurement of serum levels ofinsulin revealed that Calm and Calm Exanimals had higher levels of insulin thanCntl, although the results did not reachstatistical significance. Further analysisshowed that Calm, Cntl Ex and Calm Exanimals had positive correlations be-tween serum insulin level and final bodymass, whereas Cntl animals showed anegative correlation. These results pro-vide evidence for interaction betweencaging condition and the effects of insulinon body mass. This interaction could bedriven by factors such as altered auto-nomic nervous system or HPA activity,and in subsequent studies we aim to fur-ther examine this issue.
Glucocorticoids are known to havecatabolic effects on muscle, skin, adiposeand connective tissues, which results indecreased synthesis and increased degra-dation of protein and fat, and reducedglucose and amino acid uptake (53).Taken together, the reduced levels of glu-cocorticoids combined with increasedlevels of circulating insulin found inCalm animals may explain how theygained significantly more body massthan Cntl animals without increased en-ergy intake per unit body mass. To vali-date this hypothesis, further metabolicand endocrine studies are needed.
Our immunologic studies revealedthat both calming and exercise resultedin greater mean spleen mass. Calm Exanimals had the greatest spleen mass,which suggests there may be an additiveeffect of calming and exercise on im-mune function. Other important workhas shown that chronic restraint stress inmice generated marked reductions inspleen cellularity and that this was dueto CD95-mediated lymphocyte apoptosis(10). Together, these findings may sug-gest that animals in reduced-stresscaging exhibit greater spleen mass as aproduct of reduced leukocyte apoptosis.

6 1 6 | G U R F E I N E T A L . | M O L M E D 1 8 : 6 0 6 - 6 1 7 , 2 0 1 2
T H E C A L M M O U S E M O D E L O F S T R E S S R E D U C T I O N
In our in vitro work, we found that theacute stress of handling before sacrificeincreased the production of splenocyteIL-6 after stimulation with LPS and thedecreased production of IL-2 after CD3/CD28 bead stimulation. These resultsconfirm previous reports in humans thatacute stress can exert enhancing effectson innate immunity (46). Although thesetrends were apparent across all groups,we found that the relationships betweenacute stress and modulation of cytokineproduction were the strongest for CalmEx animals. These data support the con-cept that Calm Ex animals had increasedsensitivity to acute stress-associated im-mune modulation and that this sensitiv-ity may be a result of reduced chronic ex-posure to corticosterone, catecholaminesand other stress-related factors duringthe course of the experiment.
Analysis of the EPM data suggestedlittle difference between groups in anxi-ety-related behavior. Interestingly, otherinvestigators have found that after expo-sures to similar enriched caging environ-ments, BALB/c mice spent less time inthe open arms of the EPM (54), whereasC57BL/6 mice spent greater time in theopen arms (25), suggesting the possibil-ity that mouse strain may interact withthe environment to influence maze be-havior. Additionally, based on the char-acterization of BALB/c as a relativelyanxious mouse strain (55), detection ofchanges in anxious behavior with ade-quate sensitivity may require the appli-cation of alternative tests of rodent anxi-ety. In planned future studies, we willassess anxiety-related behavior inBALB/c mice using a light/dark test andopen-field test and, separately, we planto house other strains of mice, such asC57BL/6, in a Calm environment to fur-ther characterize the importance ofmouse strain on Calm-related reductionof anxious behavior.
The use of murine models for studyingbiological systems and disease is ubiqui-tous in scientific research. Researcherscommonly conduct mouse studies withthe goal of providing insights that can beapplied to improve human health. How-
ever, results in animal models often donot translate successfully to clinical ap-plications. There are a variety of reasonswhy the translation fails, with speciesdifferences being a central explanation.Our results and reports from othergroups (56,57) raise the possibility, how-ever, that standard caging conditionsused in most experimental settings are asource of chronic mild stress that maycontribute to problems in translatingmurine research into human studies.Within this framework, cage density, lackof species-specific behaviors, and contin-uous visibility would be sources of per-sistent stress. We have shown that cagingenvironments that mimic elements of anatural murine habitat and provide ac-cess to an exercise wheel can substan-tially influence various aspects of mousephysiology. The caging conditions wehave studied may represent a “healthier”mouse environment. To enhance the de-gree to which animal studies successfullytranslate to clinical applications, it maybe worth investigating whether standardhousing conditions are influencing theaccuracy of results rooted in mouse re-search, particularly when outcomes thatmay be influenced by stress are involved.
CONCLUSIONIn summary, we describe an animal
model that can be used to study the bio-logical effects of stress reduction. Thistechnique will allow us, and others, tocharacterize the pathways related tostress reduction that are important for influencing health outcomes. This ap-proach can also be applied in conjunctionwith other model systems of disease,such as neurodegeneration or viral infec-tion models, to determine the relevanceof stress reduction on clinical course andpathology. Lastly, we predict that pivotalinsights, concerning stress and humanhealth, will result from comparing Calmmouse findings with results from clinicalstudies of stress reduction.
ACKNOWLEDGMENTSWe thank C Grunfeld, K Laugero
and G John for helpful discussion and
G Melkus and J Hawkins for their assis-tance with MRI acquisition and analysis.This work was supported by the BowesFund for Innovative Research in Integra-tive Medicine; by grants T32 AT003997,K01 AT006153-01A1 and P01 AT005013from the National Center for Comple-mentary and Alternative Medicine; bygrant P30 AR058899 from the NationalInstitute of Arthritis, Musculoskeletal,and Skin Diseases; by Chaire ANCA; andby the Harvey V. Berneking Living Trust.
The content of this article is solely theresponsibility of the authors and doesnot necessarily represent the officialviews of the National Center for Com-plementary and Alternative Medicine orthe National Institutes of Health.
DISCLOSUREThe authors declare that they have no
competing interests as defined by Molec-ular Medicine, or other interests thatmight be perceived to influence the re-sults and discussion reported in thispaper.
REFERENCES1. Cohen S, Janicki-Deverts D, Miller GE. (2007) Psy-
chological stress and disease. JAMA. 298:1685–7.2. Glaser R, Kiecolt-Glaser JK. (2005) Stress-induced
immune dysfunction: implications for health.Nat. Rev. Immunol. 5:243–51.
3. Sternberg EM. (2006) Neural regulation of innateimmunity: a coordinated nonspecific host re-sponse to pathogens. Nat. Rev. Immunol. 6:318–28.
4. Tracey KJ. (2009) Reflex control of immunity. Nat.Rev. Immunol. 9:418–28.
5. Kiecolt-Glaser JK, Glaser R, Gravenstein S,Malarkey WB, Sheridan J. (1996) Chronic stressalters the immune response to influenza virusvaccine in older adults. Proc. Natl. Acad. Sci.U. S. A. 93:3043–7.
6. Vedhara K, et al. (1999) Chronic stress in elderlycarers of dementia patients and antibody responseto influenza vaccination. Lancet. 353:627–31.
7. Cohen S, Tyrrell DA, Smith AP. (1991) Psycholog-ical stress and susceptibility to the common cold.N. Engl. J. Med. 325:606–12.
8. Leserman J. (2000) The effects of depression,stressful life events, social support, and copingon the progression of HIV infection. Curr. Psychi-atry Rep. 2:495–502.
9. Zhang Y, et al. (2008) Toll-like receptor 4 mediateschronic restraint stress-induced immune sup-pression. J. Neuroimmunol. 194:115–22.
10. Yin D, Tuthill D, Mufson RA, Shi Y. (2000)Chronic restraint stress promotes lymphocyte

R E S E A R C H A R T I C L E
M O L M E D 1 8 : 6 0 6 - 6 1 7 , 2 0 1 2 | G U R F E I N E T A L . | 6 1 7
apoptosis by modulating CD95 expression.J. Exp. Med. 191:1423–8.
11. Dhabhar FS, McEwen BS. (1997) Acute stress en-hances while chronic stress suppresses cell- mediated immunity in vivo: a potential role forleukocyte trafficking. Brain Behav. Immun.11:286–306.
12. Sapolsky RM, Donnelly TM. (1985) Vulnerabilityto stress-induced tumor growth increases withage in rats: role of glucocorticoids. Endocrinology.117:662–6.
13. Thaker PH, et al. (2006) Chronic stress promotestumor growth and angiogenesis in a mousemodel of ovarian carcinoma. Nat. Med. 12:939–44.
14. Capitanio JP, Mendoza SP, Lerche NW, MasonWA. (1998) Social stress results in altered gluco-corticoid regulation and shorter survival insimian acquired immune deficiency syndrome.Proc. Natl. Acad. Sci. U. S. A. 95:4714–9.
15. Lupien SJ, et al. (1998) Cortisol levels duringhuman aging predict hippocampal atrophy andmemory deficits. Nat. Neurosci. 1:69–73.
16. Pham K, Nacher J, Hof PR, McEwen BS. (2003)Repeated restraint stress suppresses neurogene-sis and induces biphasic PSA-NCAM expressionin the adult rat dentate gyrus. Eur. J. Neurosci.17:879–86.
17. Luine V, Villegas M, Martinez C, McEwen BS.(1994) Repeated stress causes reversible impair-ments of spatial memory performance. Brain Res.639:167–70.
18. Davidson RJ, et al. (2003) Alterations in brain andimmune function produced by mindfulness med-itation. Psychosom. Med. 65:564–70.
19. Robinson FP, Mathews HL, Witek-Janusek L.(2003) Psycho-endocrine-immune response tomindfulness-based stress reduction in individu-als infected with the human immunodeficiencyvirus: a quasiexperimental study. J. Altern. Com-plement. Med. 9:683–94.
20. Carlson LE, Speca M, Faris P, Patel KD. (2007)One year pre-post intervention follow-up of psy-chological, immune, endocrine and blood pres-sure outcomes of mindfulness-based stress re-duction (MBSR) in breast and prostate canceroutpatients. Brain Behav. Immun. 21:1038–49.
21. Jain S, et al. (2007) A randomized controlled trialof mindfulness meditation versus relaxationtraining: effects on distress, positive states ofmind, rumination, and distraction. Ann. Behav.Med. 33:11–21.
22. Speca M, Carlson LE, Goodey E, Angen M.(2000) A randomized, wait-list controlled clinicaltrial: the effect of a mindfulness meditation-based stress reduction program on mood andsymptoms of stress in cancer outpatients. Psycho-som. Med. 62:613–22.
23. Fox C, Merali Z, Harrison C. (2006) Therapeuticand protective effect of environmental enrich-ment against psychogenic and neurogenic stress.Behav. Brain Res. 175:1–8.
24. Lehmann ML, Herkenham M. (2011) Environ-mental enrichment confers stress resiliency to social defeat through an infralimbic cortex-
dependent neuroanatomical pathway. J. Neurosci.31:6159–73.
25. Sztainberg Y, Kuperman Y, Tsoory M, Lebow M,Chen A. (2010) The anxiolytic effect of environ-mental enrichment is mediated via amygdalarCRF receptor type 1. Mol. Psychiatry. 15:905–17.
26. Moncek F, Duncko R, Johansson BB, Jezova D.(2004) Effect of environmental enrichment onstress related systems in rats. J. Neuroendocrinol.16:423–31.
27. Marashi V, Barnekow A, Ossendorf E, Sachser N.(2003) Effects of different forms of environmentalenrichment on behavioral, endocrinological, andimmunological parameters in male mice. Horm.Behav. 43:281–92.
28. Benaroya-Milshtein N, et al. (2004) Environmen-tal enrichment in mice decreases anxiety, attenu-ates stress responses and enhances natural killercell activity. Eur. J. Neurosci. 20:1341–7.
29. Van Loo PL, et al. (2004) Long-term effects ofhusbandry procedures on stress-related parame-ters in male mice of two strains. Lab Anim.38:169–77.
30. Nicholson A, et al. (2009) The response of C57BL/6J and BALB/cJ mice to increased housing den-sity. J. Am. Assoc. Lab. Anim. Sci. 48:740–53.
31. Hess SE, et al. (2008) Home improvement:C57BL/6J mice given more naturalistic nestingmaterials build better nests. J. Am. Assoc. Lab.Anim. Sci. 47:25–31.
32. Olsson IA, Dahlborn K. (2002) Improving housingconditions for laboratory mice: a review of “envi-ronmental enrichment.” Lab Anim. 36:243–70.
33. Salmon P. (2001) Effects of physical exercise onanxiety, depression, and sensitivity to stress: aunifying theory. Clin. Psychol. Rev. 21:33–61.
34. Banjanin S, Mrosovsky N. (2000) Preferences ofmice, Mus musculus, for different types of run-ning wheel. Lab Anim. 34:313–8.
35. Touma C, Sachser N, Mostl E, Palme R. (2003) Ef-fects of sex and time of day on metabolism andexcretion of corticosterone in urine and feces ofmice. Gen. Comp. Endocrinol. 130:267–78.
36. Touma C, Palme R. (2005) Measuring fecal gluco-corticoid metabolites in mammals and birds: theimportance of validation. Ann. N. Y. Acad. Sci.1046:54–74.
37. Touma C, et al. (2004) Age- and sex-dependentdevelopment of adrenocortical hyperactivity in atransgenic mouse model of Alzheimer’s disease.Neurobiol. Aging. 25:893–904.
38. Touma C, Palme R, Sachser N. (2004) Analyzingcorticosterone metabolites in fecal samples ofmice: a noninvasive technique to monitor stresshormones. Horm. Behav. 45:10–22.
39. Ciucci F, et al. (2007) Insulin-like growth factor 1(IGF-1) mediates the effects of enriched environ-ment (EE) on visual cortical development. PLoSOne. 2: e475.
40. Ibarguen-Vargas Y, Surget A, Touma C, Palme R,Belzung C. (2008) Multifaceted strain-specific ef-fects in a mouse model of depression and of anti-depressant reversal. Psychoneuroendocrinology.33:1357–68.
41. Stark JL, et al. (2001) Social stress induces gluco-corticoid resistance in macrophages. Am. J. Phys-iol. Regul. Integr. Comp. Physiol. 280: R1799–805.
42. Kubera M, et al. (1998) Effect of mild chronicstress, as a model of depression, on the im-munoreactivity of C57BL/6 mice. Int. J. Immuno -pharmacol. 20:781–9.
43. Munck A, Naray-Fejes-Toth A. (1992) The upsand downs of glucocorticoid physiology. Permis-sive and suppressive effects revisited. Mol. Cell.Endocrinol. 90: C1–4.
44. Silberman DM, Wald MR, Genaro AM. (2003)Acute and chronic stress exert opposing effectson antibody responses associated with changesin stress hormone regulation of T-lymphocyte reactivity. J. Neuroimmunol. 144:53–60.
45. Schedlowski M, et al. (1993) Changes of naturalkiller cells during acute psychological stress.J. Clin. Immunol. 13:119–26.
46. Goebel MU, Mills PJ, Irwin MR, Ziegler MG.(2000) Interleukin-6 and tumor necrosis factor-alpha production after acute psychological stress,exercise, and infused isoproterenol: differentialeffects and pathways. Psychosom. Med. 62:591–8.
47. Balcombe JP, Barnard ND, Sandusky C. (2004)Laboratory routines cause animal stress. Con-temp. Top. Lab. Anim. Sci. 43:42–51.
48. Lister RG. (1987) The use of a plus-maze to mea-sure anxiety in the mouse. Psychopharmacology(Berl.). 92:180–5.
49. Ayensu WK, et al. (1995) Effects of chronic mildstress on serum complement activity, saccharinpreference, and corticosterone levels in Flinderslines of rats. Physiol. Behav. 57:165–9.
50. Pitman DL, Ottenweller JE, Natelson BH. (1988)Plasma corticosterone levels during repeatedpresentation of two intensities of restraint stress:chronic stress and habituation. Physiol. Behav.43:47–55.
51. Bilbo SD, et al. (2002) Short day lengths augmentstress-induced leukocyte trafficking and stress-induced enhancement of skin immune function.Proc. Natl. Acad. Sci. U. S. A. 99:4067–72.
52. Harris RB, et al. (2002) Weight loss in rats ex-posed to repeated acute restraint stress is inde-pendent of energy or leptin status. Am. J. Physiol.Regul. Integr. Comp. Physiol. 282: R77–88.
53. Baxter JD, Forsham PH. (1972) Tissue effects ofglucocorticoids. Am. J. Med. 53:573–89.
54. Roy V, Belzung C, Delarue C, Chapillon P. (2001)Environmental enrichment in BALB/c mice: ef-fects in classical tests of anxiety and exposure toa predatory odor. Physiol. Behav. 74:313–20.
55. Belzung C, Griebel G. (2001) Measuring normaland pathological anxiety-like behaviour in mice:a review. Behav. Brain Res. 125:141–9.
56. Wolfer DP, et al. (2004) Laboratory animal wel-fare: cage enrichment and mouse behaviour. Nature. 432:821–2.
57. Wurbel H. (2001) Ideal homes? Housing effectson rodent brain and behaviour. Trends Neurosci.24:207–11.