tex19 paralogs are new members of the pirna pathway...
TRANSCRIPT

© 2017. Published by The Company of Biologists Ltd.
Tex19 paralogs are new members of the piRNA pathway controlling retrotransposon
suppression
Yara Tarabay1,5,6, Mayada Achour1,6, Marius Teletin1,2, Tao Ye1, Aurélie Teissandier3,
Manuel Mark1,2, Déborah Bourc’his3, Stéphane Viville1,4,*°
1Institut de Génétique et de Biologie Moléculaire et Cellulaire (IGBMC), Institut National de
Santé et de Recherche Médicale (INSERM) U964/ Centre National de Recherche Scientifique
(CNRS) UMR 1704/Université de Strasbourg, 67404 Illkirch, France 2Service de Biologie de la Reproduction, Centre Hospitalier Universitaire, 67000 Strasbourg,
France 3Institut Curie, department of Genetics and Developmental Biology, CNRS UMR3215, INSERM
U934, Paris, France 4Centre Hospitalier Universitaire, 67000 Strasbourg, France 5present address: Institut de génétique humaine (IGH), 141 rue de la Cardonille, 34396
Montpellier, France 6Co-first authors *To whom correspondence should be addressed
° present addresses: Laboratoire de diagnostic génetique, UF3472 – Infertilité, Nouvel Hôpital
Civil, 1 place de l’Hôpital, 67091 Strasbourg cedex and IPPTS, 3, rue Koeberlé, 67000
Strasbourg
Contact information: [email protected]
Tel : +33 3 68 85 37 00
Fax : + 33 3 68 85 38 09
Jo
urna
l of C
ell S
cien
ce •
Adv
ance
art
icle
JCS Advance Online Article. Posted on 2 March 2017

Summary statement
Through in vitro and in vivo experiments, the present study demonstrates that the two TEX19
paralogs belong to the family of piRNA-interacting proteins.
Abstract
Tex19 genes are mammalian specific and duplicated in Tex19.1 and Tex19.2 in some species,
such as the mouse and rat. It has been demonstrated that mutant Tex19.1 males display a
variable degree of infertility whereas they all upregulate MMERVK10C transposons in their
germ line. In order to study the function of both paralogs in the mouse, we generated and
studied double knockout (Tex19DKO) mutant mice. Adult Tex19DKO males exhibited a fully
penetrant phenotype, similar to the most severe phenotype observed in single Tex19.1KO
mice, with small testes and impaired spermatogenesis, defects in meiotic chromosome
synapsis, persistence of DNA double-strand breaks during meiosis, lack of post-meiotic germ
cells and upregulation of MMERVK10C expression. The phenotypic similarities with Piwi
KO mice prompted us to check and then demonstrate, by immunoprecipitation and GST
pulldown followed by mass spectrometry analyses, that TEX19 paralogs interact with PIWI
proteins and their VPTEL domain directly binds piRNAs in adult testes. We therefore
identified two new members of the postnatal piRNA pathway.
Key words:
Meiosis, piRNA pathway, Infertility, Transposable elements
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

Introduction
In all sexually reproducing species, germ cells are the cell lineage that is specialized in
transferring genetic information from one generation to the next (Hajkova, 2011). The
genome of germ cells needs to be maintained stably and protected in all individuals
throughout development. In particular, the genome must be protected against mobile
transposable elements (TEs). These TEs can move, amplify themselves and transpose into
new genomic loci prompting a serious threat to the genome. Although most TEs are inert,
some elements have retained activity and become typically expressed in the germ line where
they can transpose into new copies and pass onto the next generation. TEs account for almost
half of the human and mouse genomes. They are classified into two categories, class I, which
represents the majority of the TE families and class II, which accounts for approximately 3%
of the mammalian genome (Zamudio and Bourc'his, 2010, Goodier and Kazazian, 2008).
Class I TEs are believed to be ancient traces of viral infections; they transpose themselves
using a “copy and paste” mode involving an RNA intermediate, which is converted into
cDNA before its integration. Class II TEs are DNA transposons, which transpose using a “cut
and paste” mode without any transcription needed (Goodier and Kazazian, 2008, Lander et
al., 2001, Zamudio and Bourc'his, 2010).
Despite an undeniable role in evolution (Furano et al., 2004, Ostertag and Kazazian, 2001,
Zamudio and Bourc'his, 2010), TEs can be responsible for drastic mutagenic effects
illustrated by the growing number of human diseases caused by their transposition and
expression (Cowley and Oakey, 2013). Germ cells have developed sophisticated strategies to
control TE activities at the transcriptional level by DNA methylation and the post-
transcriptional level by RNA degradation or by blocking their integration for example (Gasior
et al., 2006, Obbard et al., 2009, Rollins et al., 2006, Yoder et al., 1997, Zamudio and
Bourc'his, 2010). For reasons that remain unclear, the control of TEs in the mammalian germ
line is gender-specific. Indeed, it has been shown that the protection of male germ line
genomes relies on the PIWI family of ARGONAUTE proteins, which produce and bind small
RNAs around 24-30 nucleotides long (named piRNAs) (Ghildiyal and Zamore, 2009). In
mice, the PIWI clade contains three members, namely MIWI (PIWIL1), MILI (PIWIL2) and
MIWI2 (PIWIL4) (Kuramochi-Miyagawa et al., 2004), which interact with germ line-specific
members of the TUDOR family proteins such as TDRD1, TDRD5, TDRD6, TDRD7 and
TDRD9 (Chuma et al., 2003, Hosokawa et al., 2007, Vasileva et al., 2009, Zheng and Wang,
2012). They also interact with other factors such as MOV10L1, MVH (DDX4), MAEL or
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

GASZ (Costa et al., 2006, Ma et al., 2009, Soper et al., 2008, Zheng et al., 2010, Kotaja et al.,
2006, Kuramochi-Miyagawa et al., 2004). Mutation of these piRNA pathway members give
rise to a similar phenotype consisting of male infertility due to meiosis defects that can take
place at different stages of meiosis and lead to an up-regulation of most TEs (Zamudio and
Bourc'his, 2010). Despite a large number of studies, particularly involving loss-of-function
mice, the exact mechanism whereby piRNAs control TE expression and the link between the
loss of TE control and meiosis defects are far from fully understood. Currently, only the male
process is beginning to be deciphered while the female one remains to be discovered.
In 2001, Wang et al described a set of testis expressed (Tex) genes, including Tex19 (Wang et
al., 2001). In a previous study, we showed that Tex19 is a mammal-specific gene, which is
duplicated in the mouse genome into two paralogs, Tex19.1 and Tex19.2. By multiple
sequence alignment of mammalian TEX19 proteins, two highly conserved domains, named
MCP and VPTEL respectively, were identified. However, none of them shares homology
with known proteins, preventing functional prediction (Kuntz et al., 2008). Both genes are co-
expressed in the ectoderm, then in primordial germ cells (PGCs). Later on, they are co-
expressed from embryonic day 13.5 until adulthood in testes. However, only Tex19.1
transcripts are present in developing and adult ovaries as well as in the placenta (Celebi et al.,
2012). Tex19.1 Knockout (KO) mice show a variable degree of male infertility due to
impaired meiosis associated with an up-regulation of MMERVK10C retrotransposon
expression (Ollinger et al., 2008, Tarabay et al., 2013). More recently, it was shown that
Tex19.1 plays an important role in placenta function, as Tex19.1KO mouse embryos exhibit
intra-uterine growth retardation and have small placentas due to reduced number of
spongiotrophoblast, glycogen trophoblast and sinusoidal trophoblast giant cells (Reichmann et
al., 2013, Tarabay et al., 2013). Tex19.2KO mice are fertile presenting only a subtle
phenotype with discrete seminiferous tubule degeneration in adult male testes (unpublished
observations Tarabay et al). The co-expression of Tex19.1 and Tex19.2 in PGCs, gonocytes
and spermatocytes suggests that these two genes could play redundant functions during
spermatogenesis.
By generating a double knockout of both Tex19.1 and Tex19.2 genes (Tex19DKO), we
demonstrate here that TEX19 paralogs exhibit redundant functions, which are essential for
male fertility. We show that the Tex19DKO phenotype is fully penetrant, mimicking the most
severe phenotype observed in single Tex19.1KO mice. All adult DKO males are infertile and
display impaired spermatogenesis : meiosis is blocked at the pachytene stage- during which
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

chromosome synapsis is not correctly formed or incomplete- and this leads to testis
degeneration. Furthermore, MMERVK10C retrotransposon expression is up-regulated during
meiosis in Tex19DKO testes. Through GST pull-down and immunoprecipitation experiments,
we found that TEX19 paralogs interact with PIWI proteins and their accessory partners; they
have moreover the ability to bind RNAs of around 30 nucleotides, a diagnostic size for
piRNAs, but are not required for their biogenesis. Detailed analysis further showed that
TEX19 directly interacts with piRNAs via its VPTEL domain. Altogether, our study provides
novel evidence that TEX19 proteins are new members of the PIWI/piRNA pathway acting
during post-natal spermatogenesis.
Results
Tex19 paralogs play an essential role in male fertility
Tex19.1KO mice show a variable level of male infertility. : one third of the KO male mice are
almost indistinguishable from wild type (WT) mice, one third display a mild phenotype and
the last third a severe phenotype with a complete meiosis arrest at pachytene (Ollinger et al.,
2008, Tarabay et al., 2013). Independently of the severity of the phenotype, a two-fold up-
regulation of MMERVK10C elements was consistently observed among Tex19.1KO animals
(Ollinger et al., 2008, Tarabay et al., 2013). In contrast, Tex19.2KO mice are fertile and their
testes are undistinguishable from controls. However, some seminiferous tubules show
epithelium vacuolization (unpublished observations Tarabay et al). Considering the
overlapping profile of Tex19.1 and Tex19.2 expression (Celebi et al., 2012), we wondered
whether there was any functional redundancy between the two genes in testes. We therefore
generated conditional double KO mice for Tex19.1 and Tex19.2 genes, which exist as tandem
duplication (Fig 1A).
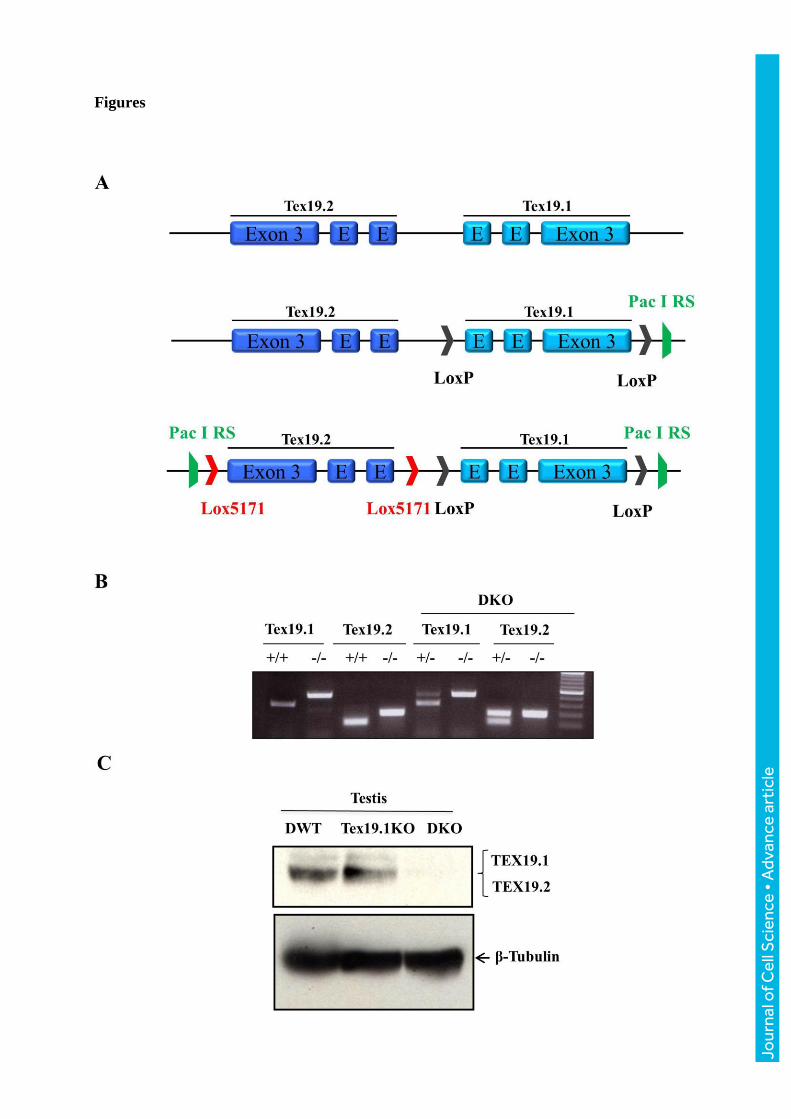
The KO of both alleles (so called Tex19DKO) was confirmed by PCR (Fig 1B) and the
absence of the encoded proteins by western blot using testes extracts from double wild-type
(DWT), Tex19.1KO, Tex19.2KO or Tex19DKO samples (Fig 1C and data not shown).
Interbreeding of Tex19 double heterozygote (DHZ) animals resulted in a statistically
significant deviation of the frequency of double homozygous animals from the expected
Mendelian 1:2:1 ratio (207 (29.3%) Tex19DWT mice, 440 (62.3%) Tex19 double
heterozygote (DHZ) mice and 59 (8.3%) Tex19DKO mice, p<0.05, Student t-test). Yang et
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

al., 2010 previously described a more important female lethality among Tex19.1KO animals,
an observation which was not shared by other independent studies including ours (Ollinger et
al., 2008, Tarabay et al., 2013, Yang et al., 2010). Nonetheless, we found that the number of
Tex19DKO females was significantly reduced when compared to Tex19DKO males (32.2% of
females versus 67.8% males, P<0.05), suggesting a gender-specific lethality. Surviving
Tex19DKO mice did not present gross somatic abnormalities. Females displayed normal
fertility, but males were sterile.
Tex19DKO males displayed a constant and severe reduction in testis size (Fig 2A). The
weight of Tex19DKO adult testes (ranking from 19mg up to 35mg, mean 24,4mg ±10.5mg
per testis) was three-fold less than that of WT testes (ranking from 109mg up to 135mg, mean
113.5mg ±5.52mg per testis) (Fig 2B). In contrast to WT seminiferous tubules (Fig 2C),
Tex19DKO testes histology showed degenerate seminiferous epithelium with tubules
containing only early spermatogenic cells, but a complete lack of post-meiotic germ cells,
suggesting a meiotic arrest (Fig 2D); consequently, the epididymis was free of spermatozoa
(Fig S1A, B). Adult Tex19DKO seminiferous tubules exhibited a drastically increased level of
apoptosis when compared to WT (Fig S1C). This phenotype is comparable to the most severe
phenotype observed in the single Tex19.1KO (Tarabay et al., 2013). Thus, Tex19 paralogs are
required for male meiosis and are essential for male fertility.
To further investigate the meiotic defect of Tex19DKO mutants, we tested the assembly of the
synaptonemal complex by co-immunostaining of the central and lateral elements with
antibodies against SYCP1 and SYCP3, respectively. In contrast to WT spermatocytes where
all autosomes were fully synapsed at pachytene (Fig 2E), synapsis failed to occur properly in
Tex19DKO, as evidenced by the complete or partial absence of the SYCP1 signal (Fig 2F).
Furthermore, as progression of homologous recombination and chromosome synapsis are
interdependent, we checked whether homologous recombination was also affected by
studying DNA double strand break (DSB) distribution, by immunostaining against
phosphorylated histone γH2AX. γH2AX staining is normally present throughout chromatin
during the zygotene stage. As synapsis proceeds, the DSBs are resolved, resulting in γH2AX
disappearance from autosomes, but not from the sex chromosomes. In WT pachytene cells,
synapsis was complete and only the sex chromosomes were stained for γH2AX (Fig 2G).
However, the incompletely synapsed pachytene chromosomes in Tex19DKO nuclear spreads
exhibited strong diffuse γH2AX staining, suggesting that the disruption of Tex19 paralogs
caused meiotic arrest at the pachytene stage because of the persistence of DSBs (Fig 2H).
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

Alternatively, γH2AX retention in spermatocytes from Tex19DKO may be a consequence of
asynapsis.
In order to date the developmental onset of the Tex19DKO phenotype, we analyzed testis
sections at post-natal (p) day 10, 16, 20 and at 16 weeks of age (Fig 3). At p10, Tex19DKO
were histologically indistinguishable from WT and Tex19DHZ testes (Fig 3A-C). Tex19DKO
showed vacuolization and seminiferous epithelium degeneration as soon as p16 (Fig 3D-F).
Thus, the Tex19DKO testes phenotype became histologically visible between p10 and p16,
corresponding to the timing when the first spermatocytes reach the pachytene stage during the
first post-natal wave of spermatogenesis. This confirmed a spermatogenesis arrest occurring
between the zygotene and the early pachytene stages of meiosis prophase I.
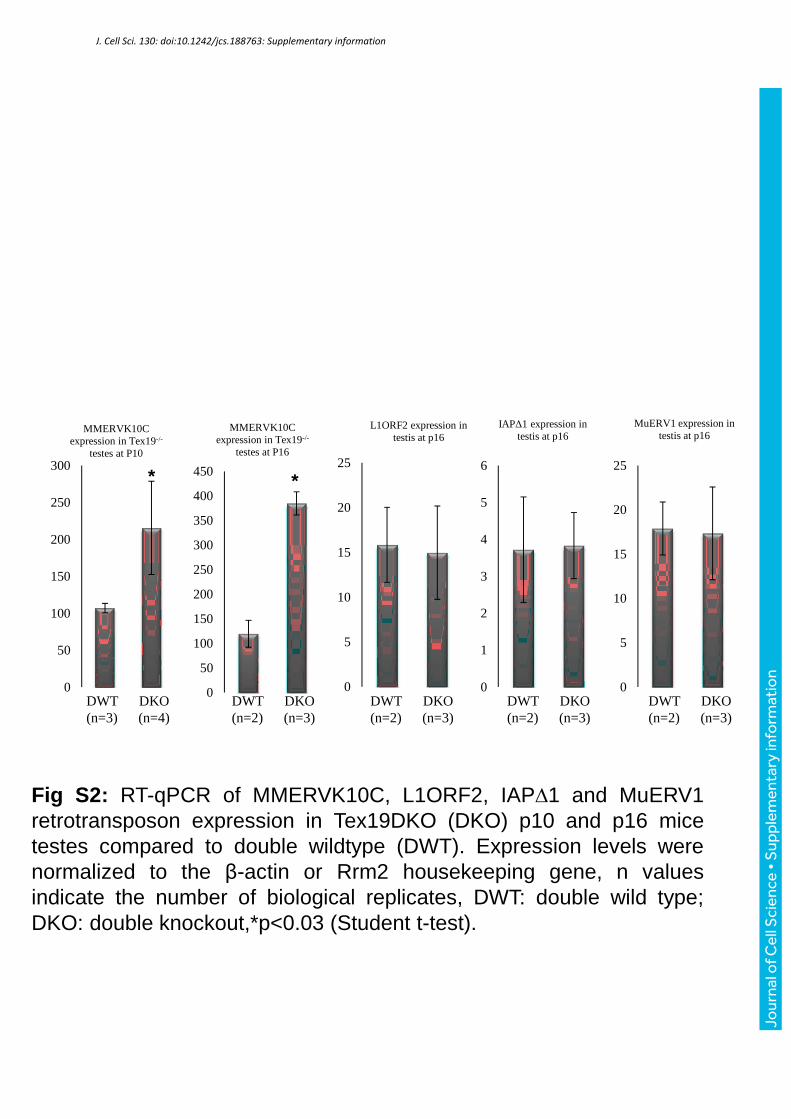
Finally, increased TE expression has been proposed to be responsible for impaired
chromosome synapsis and meiotic defects during spermatogenesis in Dnmt3L, Miwi2 and Mili
mutant mice (Aravin et al., 2007, Bestor and Bourc'his, 2004, Carmell et al., 2007,
Kuramochi-Miyagawa et al., 2004, Zamudio et al., 2015). Overexpression of MMERVK10C
retrotransposons could similarly be responsible for the meiotic defects seen in Tex19.1KO
mice (Ollinger et al., 2008, Tarabay et al., 2013). In Tex19.DKO mice, we observed a two-
fold and four-fold up-regulation of MMERVK10C expression at p10 and p16, respectively;
no upregulation was observed at other TEs (LINEs and IAP) (Fig S2), confirming the specific
impact of Tex19 deficiency in the control of MMERVK10C.
TEX19 associate with PIWI proteins and pachytene piRNAs
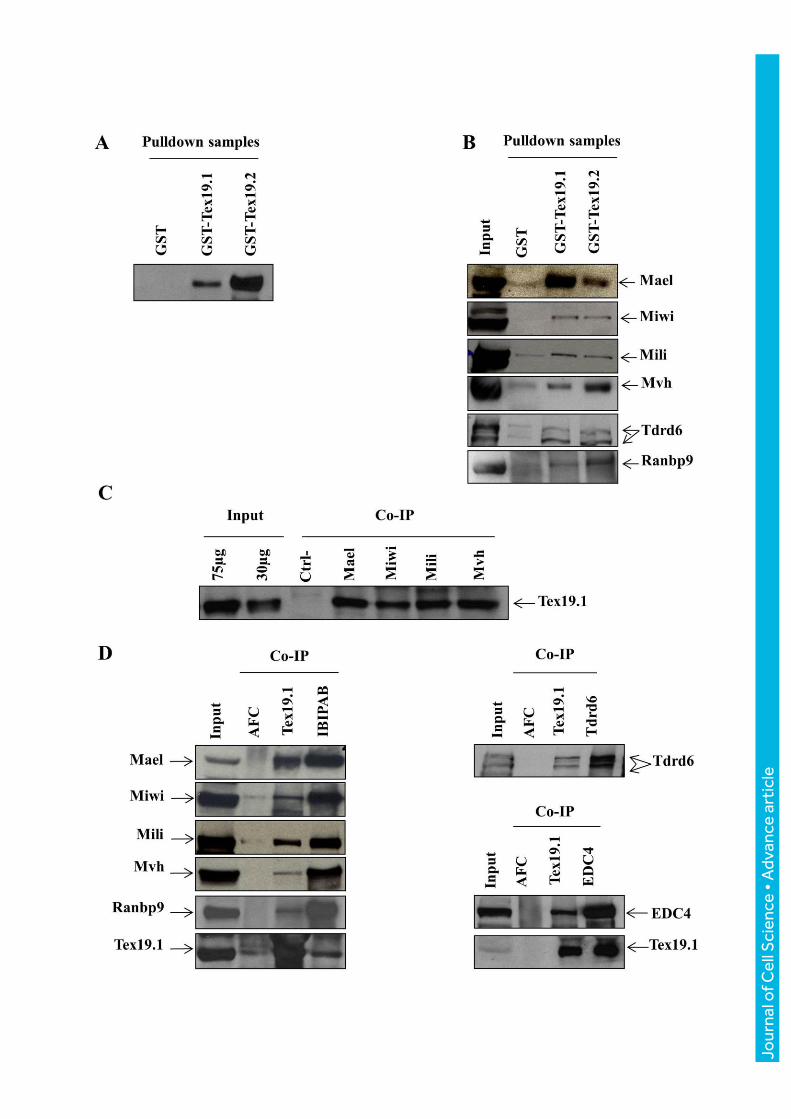
To gain insight into TEX19 function, we went on seeking its protein partners through GST
pull-down and immunoprecipitation (IP) experiments. For the GST-pull-down experiment,
GST alone, GST-TEX19.1 or GST-Tex19.2 was incubated with total testicular protein extract
of adult mice (Fig S3); complexes were then analyzed by mass spectrometry (MS). In GST-
TEX19.1 and GST-TEX19.2 samples, we detected bands corresponding to MAEL, MIWI,
MILI, TDRD6, RANBP9 and MVH while these bands were absent in GST alone (Fig 4B).
These interactions were further confirmed by western blot (WB) in co-immunoprecipitation
experiments. Using a specific anti-TEX19.1 monoclonal antibody, we further immuno-
purified TEX19.1 complexes in adult testes and identified individual polypeptides by MS.
Among the identified proteins, we found MAEL, MIWI, MILI, and MVH which were already
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

identified by GST pull-down, but also TDRD8, TDRD6, EDC4, PABPC2, PSMC3,
RANBP9, ANGEL1, SRPK1, DDX20, DDX46. We confirmed the presence of MAEL,
MIWI, MILI, MVH, EDC4, TDRD6 and RANBP9 in TEX19.1 immuno-purified complexes
isolated from cytoplasmic adult testes extracts by WB (Fig 4C, D). These interactions were
further validated by the reciprocal detection of TEX19.1 in immuno-purified MAEL, MIWI,
MILI, MVH, EDC4 and RANBP9 complexes (Fig 4D). Altogether these results suggest that
TEX19.1 and TEX19.2 interact with proteins of the post-natal PIWI/piRNA pathway, namely
core components such as MIWI, MILI, and accessory proteins such as EDC4, TDRD6,
RANBP9 and MVH.
The identification of TEX19 paralogs as partners of the PIWI proteins along with the
phenotypic similarities observed between Tex19KO mice and piRNA mutant mice prompted
us to check if TEX19s could bind small RNAs, and in particular piRNAs. For this purpose,
we immuno-precipitated TEX19.1, TEX19.2 and MILI (as a positive control) in adult mouse
(aged 12 weeks) testes extracts and assessed for the presence of small RNAs by 32P 5’ end
labeling and gel electrophoresis (Fig 5). As expected, MILI associated with ~26-30 nucleotide
(nt) small RNAs. TEX19.1 purification also revealed an interaction with small RNAs ranging
in size between 26 and 30 nt, migrating similarly to MILI associated piRNAs. Using the
6Tex:4D2 antibody that recognized both TEX19 paralogs on an adult Tex19.1KO testicular
extract from a normal size testis from a 12 weeks old male, we showed that TEX19.2 also
interacted with small RNAs of the same size range (Fig 5). All bands disappeared upon
RNase treatment, confirming the RNA nature of these entities. Furthermore, no such band
was detected in the ascitic fluid control sample or in the sample precipitated with a specific
anti-TEX19.1 monoclonal antibody in Tex19.1KO testicular extract, confirming the
specificity of the interaction between TEX19.1 and small RNAs. Thus both TEX19 paralogs
interact with small RNAs in a size range compatible with piRNAs.
To further identify the small RNA species associated with TEX19 proteins, we generated
small RNA-seq libraries by Illumina sequencing after TEX19 and TEX19.1
immunoprecipitation from adult WT testes. To infer the TEX19.2-bound fraction, we
analyzed TEX19-complexes in Tex19.1KO testes. Size distribution and genomic content were
then compared to available datasets from MILI and MIWI-bound piRNAs from adult testes
(Robine et al., 2009). MIWI was formerly shown to preferentially associate with 29-30nt
piRNA species, while the main piRNA size in MILI complexes is shorter, around 26-27nt.
Interestingly, TEX19-, TEX19.1- and TEX19.2-associated small RNAs were found to
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

segregate in size with MIWI-bound piRNAs, with a diagnostic 30nt size (Fig 6A). TEX19.2
showed a stronger association to 22-23nt long RNAs, which is a diagnostic size for
microRNAs: as small RNA libraries are expressed as relative rather than absolute values, this
may reflect a weaker affinity of TEX19.2 towards piRNAs but a genuine preferential
association with miRNAs cannot be excluded.
Because the three libraries were very similar in size and content, but the TEX19.1 sample had
a better quality, we focused the rest of our analysis on the latter. Out of 1.6 million reads, we
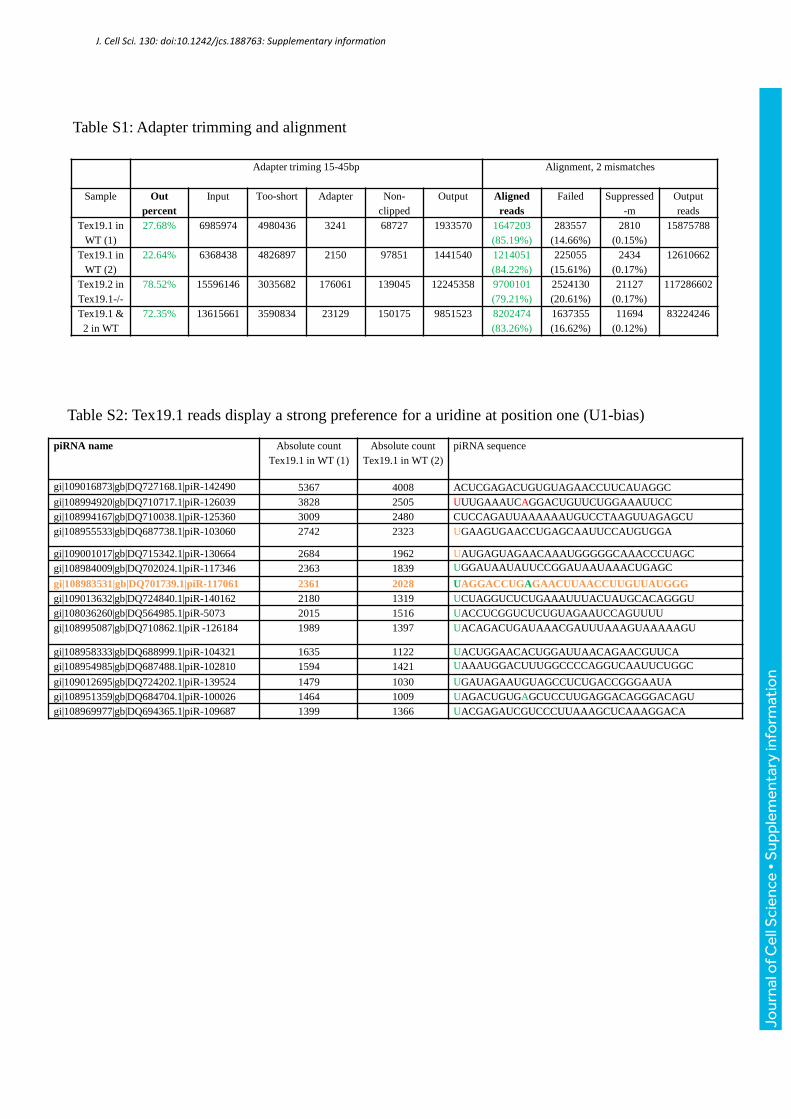
noticed a strong preference (94.42%) for a uridine at position one (U1 bias), in agreement
with the biogenesis mode of piRNAs (Aravin et al., 2006, Frank et al., 2010, Kawaoka et al.,
2011) (Table S2, Fig S4). The genomic distribution appeared identical to both MILI and
MIWI IPs (Fig 6B): the majority (70%) of the TEX19.1-derived reads originated from
intergenic regions, while less than 20% of the reads mapped to repeated elements. This is in
line with the known genomic distribution of pachytene piRNA species, which are produced in
adult testes from a discrete number of unique and intergenic piRNA clusters . Accordingly,
90% TEX19.1-associated reads uniquely mapped onto the genome (Fig 6C), and we moreover
found the large majority of TEX19.1-associated small RNAs to be derived from the same
piRNA loci clusters previously defined as regulated by MILI and MIWI (Fig 6D) (Beyret et
al., 2012). We nonetheless examined the content of the repeated fraction of the TEX19.1
library and found that around 50% were derived from LTR retrotransposons, among which
around 1% belonged to the MMERVK10C class (Fig 6B). This means that along all 25-30nt-
bound RNAs, only 0.1% derived from MMERVK10C elements. Finally, to address the role of
TEX19.1 in the biogenesis of piRNAs, we sequenced whole testes small RNA libraries in
three adult (12 weeks of age) Tex19.1KO animals showing normal size testis: the size
distribution appeared similar to WT testes, and in particular there was no relative
redistribution towards 22-23nt microRNA species, as an indication of impaired piRNA
production (Fig 6E). Enrichment in 30nt-centered small RNAs was even higher in the
Tex19.1KO background compared to WT: this likely reflects different cellular composition,
and in particular an overrepresentation of pachytene cells in Tex19.1KO testes, due to the
meiotic arrest. Altogether, these results suggest that TEX19 proteins sample the same set of
primary, unique piRNA species as MILI and MIWI, at the time of their production at the
pachytene stage in the adult testis. However, they are not required for their synthesis, and
likely act as simple cargo.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

TEX19 paralogs interact directly with piRNAs
In order to assess a direct interaction between TEX19.1, TEX19.2 and piRNAs, we performed
electrophoretic mobility shift assays (EMSA) using GST-TEX19.1 and GST-TEX19.2
proteins as well as different domains of TEX19.1 fused to GST (Fig S5 and S6) against the
piR-117061 piRNA. We choose this piRNA because it has a very significant absolute count
(2361 and 2028) in two biological replicates prepared using anti-TEX19.1 antibody (Table
S3) and it has a U at position one “U1”. We found that the piR-117061 was ‘shifted' up upon
electrophoresis in the presence of GST-TEX19.1, GST-TEX19.2, GST-D2Cter (125-351aa)
or GST-VPTEL of TEX19.1, but not in the presence of GST-alone or GST-MCP (1-124aa) or
GST-Cter (163-351aa) of TEX19.1. This result stresses the specificity of the interaction
between piR-117061 and TEX19.2 and TEX19.1 via the VPTEL domain (Fig 7A). The
specificity of this interaction was further confirmed by showing that a modified form of piR-
117061, in which we replaced U1 and A10 by a C was unable to bind TEX19.1, its VPTEL
domain or TEX19.2 (Fig 7B) and was not able to replace piR-117061 in a competition
experiment (Fig 7C). By adding an increased concentration of the non-radiolabeled piR-
117061 to a fixed concentration of GST-VPTEL (5µM pmole) and radiolabeled piR-117061,
we showed that the non-radiolabeled piR-117061 completely displaced the radiolabeled piR-
117061 at one hundred fold concentration (Fig 7D). Finally, we showed that the quantity of
piR-117061 ‘shift' up was proportional to the quantity of the TEX19.1 VPTEL domain (Fig
7D).
Discussion
Tex19 genes are specific to mammals and can be found as tandem duplication in the mouse
and rat genomes. In the mouse, Tex19.1 and Tex19.2 expression is restricted to the germ line
and the placenta (Celebi et al., 2012). Analyses of Tex19.1KO mice have previously
highlighted a variable phenotype. In the severest form, Tex19.1 deficient males were infertile
and presented an arrest at the pachytene stage of meiosis, in association with a failure to
silence MMERVK10C elements (Ollinger et al., 2008, Tarabay et al., 2013). Tex19.2KO
males are fertile; however a subtle phenotype of testes degeneration could be observed in
adult mice (unpublished observations Tarabay et al, 2013). The present study describes the
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

functional consequences of deleting both TEX19 paralogs and the involvement of these
proteins in the piRNA pathway. Surviving Tex19DKO male and female mice were perfectly
healthy, but males exhibited a fully penetrant phenotype similar to the severest Tex19.1KO
phenotype (Ollinger et al., 2008, Tarabay et al., 2013). Indeed, Tex19DKO males have highly
degenerated testes, with a lack of germ cells beyond the pachytene stage, linked to defects in
chromosome synapsis during meiosis. Compared to Tex19.1KO testes (Ollinger et al., 2008,
Tarabay et al., 2013), we showed here a stronger up-regulation of MMERVK10C
retrotransposon in double Tex19DKO testes, which was two-fold at p10 and four-fold at p16
compared to WT counterparts. This fully penetrant phenotype comforts the hypothesis of a
functional redundancy between Tex19.1 and Tex19.2. Moreover, this phenotype bears striking
resemblance with the Dnmt3L, Miwi2 and Mili mutant phenotypes (Aravin et al., 2007, Bestor
and Bourc'his, 2004, Carmell et al., 2007, Kuramochi-Miyagawa et al., 2004). One
explanation was proposed by Zamudio et al, who suggested that improper TE silencing may
alter the meiotic process through local remodeling of the chromatin landscape and (Zamudio
et al., 2015).
The essential role of TEX19 may extend beyond the context of spermatogenesis as we report
here a severe lethality of Tex19DKO animals, with only 8.3%, instead of the expected 25% of
mutant animals. Moreover, this lethality was sexually dysmorphic as among the survivors,
only one third were females. We did not observe such lethality in our single Tex19.1KO
mouse models (Tarabay et al., 2013), but Yang et al., 2010 also reported such female-specific
lethality in their Tex19.1KO mice and suggested that the variable severity among different
studies could be linked to strain specificities (Yang et al., 2010, Kwon et al., 2003). However,
our studies on Tex19.1KO or Tex19DKO mice were conducted onto the same genetic
background (C57Bl/6). A strong bias in sexual lethality was also observed in C57Bl/6 mice
by Reichmann et al, but only when females were lactating and nursing a pre-existing litter
during pregnancy (Reichmann et al., 2013). Yet, none of our studies were conducted under
such lactating conditions. Consequently, this difference between Tex19.1KO and Tex19DKO
phenotypes is quite surprising since Tex19.2 is not expressed in the placenta, neither in WT
nor in Tex19.1KO animals. This early postnatal lethality could be indicative of defects in
placenta (Reichmann et al., 2013, Tarabay et al., 2013), but also suggests a functional
redundancy between Tex19.1 and Tex19.2 during embryo development. The exact reason for
the preferential female lethality of the Tex19DKO mutation remains unclear, but may be
linked to X chromosome-specific effects.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

The main findings provided by our work relate to the identification of the protein partners of
TEX19 in adult testes. Through GST pulldown and IP experiments, we indeed showed for the
first time that TEX19.1 exists in a complex with members of the PIWI/piRNA pathway:
MAEL, MIWI, MILI and MVH. We also confirmed a significant level of interaction between
TEX19.1 and UBR2 by IP, as it had been previously reported (Yang et al., 2010). Most
importantly, we found that both TEX19 paralogs bind small RNAs that have all the
characteristics of piRNAs: around 30nt in size and with a strong preference for a uridine as
the first nucleotide (Frank et al., 2010, Kawaoka et al., 2011, Brennecke et al., 2007). Finally,
this interaction seems to be direct and to occur through the well-conserved VPTEL domain of
TEX19 proteins. We can therefore conclude that TEX19 paralogs are new members of the
family of proteins that bind piRNAs and that the VPTEL domain could be a new small RNA-
binding motif. While this study may reveal the piRNA-binding function of the VPTEL
domain, we cannot speculate about the function of the MCP domain, which lacks homology
with any known proteins. PIWI proteins have, in addition to their ability to bind piRNAs, an
RNA-guided nuclease function linked to a catalytic triad DDH localized in the PIWI domain
and which is required for piRNA biogenesis (Martinez and Tuschl, 2004). No such triad can
be found in TEX19 proteins. Accordingly, the production of 25-30 nt small RNAs is not
altered in Tex19-deficient testes. This suggests that while TEX19 proteins bind to piRNAs,
they are not involved in the catalysis of piRNA synthesis.
Finally, it is now well established that TEX19 paralogs are required for the control of
MMERVK10C transposons in testes (Ollinger et al., 2008, Tarabay et al., 2013). Despite a
consistent effect of Tex19 deficiency on these elements, we found that MMERVK10C-
derived reads only represent 0.1% of all 25-30nt TEX19-bound small RNAs. In contrast,
TEX19 mostly binds unique, non-transposon derived small RNAs in adult testes, which are
similar to the MILI and MIWI-associated piRNAs. This is not surprising, as the post-natal
PIWI machinery is responsible for producing piRNAs derived from discrete intergenic
regions as male germ cells enter the pachytene stage (Aravin et al., 2006, Girard et al., 2006)
although the function of these mammalian post-natal piRNAs remains enigmatic (Goh et al.,
2015). This poor enrichment of TEX19-bound small RNAs in MMERVK10C fragments
raises the question as to whether the reactivation of MMERVK10C in Tex19.1KO and
Tex19DKO mice reflects a direct role of TEX19 proteins in silencing these elements-likely
via post-transcriptional mechanisms- or an indirect effect of the Tex19 mutation. Further work
should be dedicated to resolve this question.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

In conclusion, our analysis of the double-mutant mice for the two TEX19 paralogs
undoubtedly reveals the crucial role of these proteins for male fertility in mammals, likely via
their association with the PIWI/piRNA pathway.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

Material and methods
Generation of Tex19 double knockout mice
The Tex19 double mutant mouse line was established at the MCI/ICS (Mouse Clinical
Institute- Institut Clinique de la Souris-, Illkirch, France; http://www-mci.u-strasbg.fr). For
Tex19.1, the targeting vector was constructed as follows (Fig 1A): three PCR fragments (from
129S2/SvPas ES cell genomic DNA) one 2.2 Kb fragment corresponding to inter-loxP
fragment encompassing exons 2 and 3, a 4.5 kb and a 3.1kb fragment corresponding to the 5’
and 3’ arms respectively were sequentially subcloned in a MCI proprietary vector. This MCI
vector contains a LoxP site as well as a floxed and flipped Neomycin resistance cassette. A
unique PacI restriction site was inserted downstream to the 3’ loxP site. The linearized
construct was electroporated in 129S2/SvPas mouse embryonic stem (ES) cells. After G418
selection, targeted clones were identified by PCR using external primers and further
confirmed by Southern blot with 5’ and 3’ external probes.
For Tex19.2, the targeting vector was constructed as follows: three PCR fragments (from
129S2/SvPas ES cell genomic DNA) a 2.0 Kb fragment corresponding to inter-lox P fragment
encompassing exons 2 and 3, a 4.4 kb and a 3.5kb fragment corresponding to the 5’ and the 3’
arms respectively were sequentially subcloned in a MCI proprietary vector. This MCI vector
contains a Lox5171 site as well as a F3 surrounded Hygromycin resistance cassette. A unique
PacI restriction site was inserted upstream to the 5’ loxP site. The linearized construct was
electroporated in one fully valid Tex19.1 KO clone. After hygromycin selection, targeted
clones were identified by PCR using external primers and further confirmed by Southern blot
with 5’ and 3’ external probes. Restriction digestion by PacI followed by Southern blot
allowed selection of a clone showing targeting of both constructs on the same allele. This
clone was injected into C57BL/6J blastocysts, and male chimeras tested for germline
transmission. The NeoR and HygroR cassettes were deleted by breeding the F1 mice with Flp
mice (Rodriguez et al., 2000). Excision of both Tex19.1 and Tex19.2 inter-Lox fragments was
done by breeding the mice with a Cre deleter line (Dupe et al., 1997). The absence of Tex19.1
and Tex19.2 mRNA and protein was tested by RT-PCR using oligonucleotides described
previously (Tarabay et al., 2013) and by western blotting respectively.
All animal experimental procedures were performed according to the European authority
guidelines.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

Antibody production, Western blotting, preparation of cytoplasmic and nuclear
fractions
Injecting the entire protein or the peptide DLGPEDAEWTQALPWRC into mice allowed the
generation of anti-TEX19 monoclonal antibodies. The specificity of both antibodies was
tested as described elsewhere (Tarabay et al., 2013). The 7Tex:1F11 antibody is TEX19.1
specific, whereas 6Tex:4D2 antibody detects both TEX19.1 and TEX19.2.
Cytoplasmic and nuclear fraction were prepared as described
http://openwetware.org/wiki/Cytoplasm_and_nuclear_protein_extraction
RT-PCR and quantitative RT-PCR
RNA was prepared using the RNeasy mini or micro kit (Qiagen) following the manufacturer’s
instructions. After DNase I digestion (Roche, Mississauga, ON, Canada), 1µg RNA was
reverse-transcribed by random priming using Superscript II (Invitrogen/Life Technologies,
Burlington, ON, Canada). QPCR reaction was performed using SYBR® green JumpStartTM
Taq ReadyMixTM (Sigma-Aldrich, Oakville, ON, Canada) and LightCycler 480 (Roche,
Mississauga, ON, Canada). The efficiency and specificity of each primer pair was checked
using a cDNA standard curve. All samples were normalized to β-actin and Rrm2 expression
(Tarabay et al., 2013).
Histology
Testes were collected and fixed in Bouin’s fluid for 48 hours and then embedded in paraffin.
Adult male testis were dissected at 16 weeks of age. For histological analyses, 5 µm thick
sections were stained with hematoxylin/eosin. All slides were examined using a DMLA
microscope (Leica) with 10X, 20X, 40X and 100X objectives with apertures of 0.3, 0.5, 0.7
and 1.3, respectively. Images were taken with digital camera (CoolSnap; Photometrix) using
CoolSnap v.1.2 software and then processed with Photoshop CS2 v.9.0.2 (Adobe).
Immunostaining of meiotic chromosome spreads
Meiotic chromosome spreads were prepared as described previously (Mark et al., 2008).
TEX19 protein production and purification
Tex19.1 cDNA or its domains were cloned, by RT- PCR starting from a testis cDNA library,
into pCR®4-TOPO (Invitogen/Life Technologies, Burlington, ON, Canada) then into pENTR
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

1A (Life Technologies) then by LR recombinase into pHGGWA or pGGWA. The
recombinant proteins GST-TEX19, MCP, VPTEL-Cter, VPTEL, Cter, and protein GST-alone
were individually expressed in E. coli BL21 cells. The induction by 0.1mM of IPTG was
performed at a cell density of A600 = 0.8 at 20°C overnight and a portion of the proteins
examined by 10% SDS-PAGE. Then, sonication was performed 3s ON and 3s OFF for 5 min
at amplitude 33%. Cleared lysates were prepared in lysis buffer (Tris HCl 50mM pH9, NaCl
400mM, DTT 2mM, triton 1% and protease inhibiting cocktail) and centrifuged for 1 hour at
20000 rpm. The soluble fusion proteins were purified by batch procedure incubating the
lysate with 500µL of Glutathione Sepharose (Amersham Biosciences) for 1 hour at 4°C with
gentle shaking. Then the Glutathione Sepharose was collected by centrifugation and washed
five times with lysis buffer. After boiling, the bound proteins were analyzed by 10% SDS‐
PAGE followed by Western blotting using anti-TEX19 monoclonal antibodies 7Tex:1F11 or
6Tex:4D2.
For GST pulldown assays, total testicular extracts of adult mice (aged 12 weeks) were added
to recombinant protein fixed to Gluthatione Sepharose beads for 2h00 at 4°C. After fives
washes, proteins were separated on 10% SDS–PAGE and subsequently stained with Bio-Safe
Coomassie (Bio-Rad, Hercules, CA, USA). The gel was then sent to Taplin Biological Mass
Spectrometry Facility to be analyzed by LC/MS/MS (Taplin, Boston, MA, USA).
Purified recombinant proteins were used in EMSA assays as described hereafter.
Co-Immunoprecipitation
Antibodies were chemically cross-linked to protein G–Sepharose beads using dimethyl
pimelimidate (DMP; Sigma-Aldrich D-8388) and used to purify TEX19.1, TEX19.2, MAEL,
MIWI, MILI, MVH complexes from adult mouse testes (aged 12 weeks) cytoplasmic
fractions. 30mg of clarified DWT testes cytoplasmic extracts were incubated with 300µl
protein G-Sepharose beads coupled to antibodies; anti-TEX19.1 (7Tex:1F11,100µl), anti-
TEX19.1/TEX19.2 (6Tex:4D2, 10ml of hybridoma supernatant), anti-MAEL (18µg, Abcam,
Toronto, ON, Canada), anti-MIWI (20µg, Aviva System Biology, San Diego, CA, USA),
anti-MILI (40µg, Abcam), anti-MVH (12µg, Abcam), anti-TDRD66 (50µg,
Millipore/Upstate, Etobicode, ON, Canada), anti-EDC4 (12µg, Abcam), anti-RANBP9 (8µg,
Proteintech, Rosemonte, IL, USA) or an ascitic fluid control (negative control, 100µl) in Co-
IP buffer (50 mM Tris, pH 8, 150 mM NaCl, 1Mm EDTA, complete protease inhibitor)
overnight at 4°C. After five washes with Co-IP buffer, the retained proteins were denatured
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

by Laemmli buffer with 5% -Mercaptoethanol and boiled to 100°C, then separated by SDS-
PAGE as described hereafter. If required, gels were stained with Coomassie blue and sent to
Taplin Biological Mass Spectrometry Facility to be analyzed by LC/MS/MS (Taplin, Boston,
MA, USA).
Small RNA immunoprecipitation and purification
TEX19 libraries were prepared as described elsewhere (Reuter et al., 2009).
Electrophoretic Mobility Shift Assay (EMSA)
For 32P-labeling, piR-117061 (5’UAGGACCUGAGAACUUAACCUUGUUAUGGG 3’)
and piR-nonspecific (5’CAGGACCUGCGAACUUAACCUUGUUAUGGG 3’) were
incubated for 60min at 37°C in 20µl kinase buffer containing 5μl RNA (0.5 nmol), 2μl buffer
B, 4µl PEG, 10U T4 PNK kinase (10U/µl, Fermentas), 40U RNAsine (40U/µl), 2 μl [γ-
32P]ATP (6000 Ci/mmole) (Perkin Elmer), 5μl ddH2O.
EMSA assays were performed with 5 µM GST-alone, GST-TEX19.1, GST-MCP, GST-
VPTEL/Cter, GST-VPTEL, GST-Cter and 125 femtomoles (8000cpm) denatured
radiolabeled piRNA in the binding buffer (250µg Herpain, 10Mm MOPS pH 7, 50mM
MgCl2, 1 mM DTT,10% glycerol, 40U RNAsine, 10ng/µl tRNA of E.coli) as described
(Bendak et al., 2012).
Small RNA sequencing and analysis
Small RNA-seq of whole adult testes and TEX19-IP were done using Illumina HiSeq 2000 on
libraries (18-45nt RNAs size-fractionated on acrylamide gel) prepared with the Illumina
TruSeq small RNA protocol. For the analysis, adapters were removed using Cutadapt (Martin,
2011). The trimmed sequencing reads of each library were then mapped and annotated with
ncPRO-seq (Chen et al., 2012) onto the mouse reference genome (mm9). Briefly, read
alignment was done using Bowtie v0.12.8, allowing a sum of qualities of mismatching bases
lower than 50 (-e 50). A number of 5,000 hits per aligned read were allowed. Aligned reads
were then annotated using RepeatMasker and refGene databases, and the positional read
coverage was weighted by mapping site numbers.
The piRNA cluster analysis was performed as described (Beyret et al., 2012). A cluster was
defined as a group of piRNAs where the reads were less than 1,500bp away from each other.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

Clusters with at least 1,000 normalized read count were reported. The UCSC genome tracks
were generated using the HOMER software (v4.3) using unique mapped reads.
Funding
The study was funded by Agence Nationale de la Recherche (ANR-11-BSV2-002
« TranspoFertil ») and l’Agence de BioMédecine (« AMP, diagnostic prénatal et diagnostic
génétique »).
Acknowledgment
Y.T., M.A., M.T., T.Y., A.T., M.M., D.B., S.V.: Data analysis and interpretation, manuscript
writing, final approval of manuscript. We would also like to thank Robert Drillien for his
critical reading of the manuscript. We are grateful to the Institute of Genetics and Molecular
and Cellular Biology (IGBMC) platforms. The study was funded by Agence Nationale de la
Recherche (ANR-11-BSV2-002 « TranspoFertil ») and l’Agence de BioMédecine (« AMP,
diagnostic prénatal et diagnostic génétique »). This work was supported by the French Centre
National de la Recherche Scientifique (CNRS), Institut National de la Santé et de la
Recherche Médicale (INSERM), the Ministère de l’Education Nationale et de l’Enseignement
Supérieur et de la Recherche, the University of Strasbourg, the University Hospital of
Strasbourg.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

References
ARAVIN, A., GAIDATZIS, D., PFEFFER, S., LAGOS-QUINTANA, M., LANDGRAF, P.,
IOVINO, N., MORRIS, P., BROWNSTEIN, M. J., KURAMOCHI-MIYAGAWA, S.,
NAKANO, T., CHIEN, M., RUSSO, J. J., JU, J., SHERIDAN, R., SANDER, C.,
ZAVOLAN, M. & TUSCHL, T. 2006. A novel class of small RNAs bind to MILI
protein in mouse testes. Nature, 442, 203-7.
ARAVIN, A. A., HANNON, G. J. & BRENNECKE, J. 2007. The Piwi-piRNA pathway
provides an adaptive defense in the transposon arms race. Science, 318, 761-4.
BENDAK, K., LOUGHLIN, F. E., CHEUNG, V., O'CONNELL, M. R., CROSSLEY, M. &
MACKAY, J. P. 2012. A rapid method for assessing the RNA-binding potential of a
protein. Nucleic Acids Res, 40, e105.
BESTOR, T. H. & BOURC'HIS, D. 2004. Transposon silencing and imprint establishment in
mammalian germ cells. Cold Spring Harb Symp Quant Biol, 69, 381-7.
BEYRET, E., LIU, N. & LIN, H. 2012. piRNA biogenesis during adult spermatogenesis in
mice is independent of the ping-pong mechanism. Cell Res, 22, 1429-39.
BRENNECKE, J., ARAVIN, A. A., STARK, A., DUS, M., KELLIS, M.,
SACHIDANANDAM, R. & HANNON, G. J. 2007. Discrete small RNA-generating
loci as master regulators of transposon activity in Drosophila. Cell, 128, 1089-103.
CARMELL, M. A., GIRARD, A., VAN DE KANT, H. J., BOURC'HIS, D., BESTOR, T. H.,
DE ROOIJ, D. G. & HANNON, G. J. 2007. MIWI2 is essential for spermatogenesis
and repression of transposons in the mouse male germline. Dev Cell, 12, 503-14.
CELEBI, C., VAN MONTFOORT, A., SKORY, V., KIEFFER, E., KUNTZ, S., MARK, M.
& VIVILLE, S. 2012. Tex 19 paralogs exhibit a gonad and placenta-specific
expression in the mouse. J Reprod Dev, 58, 360-5.
CHEN, C. J., SERVANT, N., TOEDLING, J., SARAZIN, A., MARCHAIS, A.,
DUVERNOIS-BERTHET, E., COGNAT, V., COLOT, V., VOINNET, O., HEARD,
E., CIAUDO, C. & BARILLOT, E. 2012. ncPRO-seq: a tool for annotation and
profiling of ncRNAs in sRNA-seq data. Bioinformatics, 28, 3147-9.
CHUMA, S., HIYOSHI, M., YAMAMOTO, A., HOSOKAWA, M., TAKAMUNE, K. &
NAKATSUJI, N. 2003. Mouse Tudor Repeat-1 (MTR-1) is a novel component of
chromatoid bodies/nuages in male germ cells and forms a complex with snRNPs.
Mech Dev, 120, 979-90.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

COSTA, Y., SPEED, R. M., GAUTIER, P., SEMPLE, C. A., MARATOU, K., TURNER, J.
M. & COOKE, H. J. 2006. Mouse MAELSTROM: the link between meiotic silencing
of unsynapsed chromatin and microRNA pathway? Hum Mol Genet, 15, 2324-34.
COWLEY, M. & OAKEY, R. J. 2013. Transposable elements re-wire and fine-tune the
transcriptome. PLoS Genet, 9, e1003234.
DUPE, V., DAVENNE, M., BROCARD, J., DOLLE, P., MARK, M., DIERICH, A.,
CHAMBON, P. & RIJLI, F. M. 1997. In vivo functional analysis of the Hoxa-1 3'
retinoic acid response element (3'RARE). Development, 124, 399-410.
FRANK, F., SONENBERG, N. & NAGAR, B. 2010. Structural basis for 5'-nucleotide base-
specific recognition of guide RNA by human AGO2. Nature, 465, 818-22.
FURANO, A. V., DUVERNELL, D. D. & BOISSINOT, S. 2004. L1 (LINE-1)
retrotransposon diversity differs dramatically between mammals and fish. Trends
Genet, 20, 9-14.
GASIOR, S. L., WAKEMAN, T. P., XU, B. & DEININGER, P. L. 2006. The human LINE-1
retrotransposon creates DNA double-strand breaks. J Mol Biol, 357, 1383-93.
GHILDIYAL, M. & ZAMORE, P. D. 2009. Small silencing RNAs: an expanding universe.
Nat Rev Genet, 10, 94-108.
GIRARD, A., SACHIDANANDAM, R., HANNON, G. J. & CARMELL, M. A. 2006. A
germline-specific class of small RNAs binds mammalian Piwi proteins. Nature, 442,
199-202.
GOH, W. S., FALCIATORI, I., TAM, O. H., BURGESS, R., MEIKAR, O., KOTAJA, N.,
HAMMELL, M. & HANNON, G. J. 2015. piRNA-directed cleavage of meiotic
transcripts regulates spermatogenesis. Genes Dev, 29, 1032-44.
GOODIER, J. L. & KAZAZIAN, H. H., JR. 2008. Retrotransposons revisited: the restraint
and rehabilitation of parasites. Cell, 135, 23-35.
HAJKOVA, P. 2011. Epigenetic reprogramming in the germline: towards the ground state of
the epigenome. Philos Trans R Soc Lond B Biol Sci, 366, 2266-73.
HOSOKAWA, M., SHOJI, M., KITAMURA, K., TANAKA, T., NOCE, T., CHUMA, S. &
NAKATSUJI, N. 2007. Tudor-related proteins TDRD1/MTR-1, TDRD6 and
TDRD7/TRAP: domain composition, intracellular localization, and function in male
germ cells in mice. Dev Biol, 301, 38-52.
KAWAOKA, S., IZUMI, N., KATSUMA, S. & TOMARI, Y. 2011. 3' end formation of
PIWI-interacting RNAs in vitro. Mol Cell, 43, 1015-22.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

KOTAJA, N., BHATTACHARYYA, S. N., JASKIEWICZ, L., KIMMINS, S., PARVINEN,
M., FILIPOWICZ, W. & SASSONE-CORSI, P. 2006. The chromatoid body of male
germ cells: similarity with processing bodies and presence of Dicer and microRNA
pathway components. Proc Natl Acad Sci U S A, 103, 2647-52.
KUNTZ, S., KIEFFER, E., BIANCHETTI, L., LAMOUREUX, N., FUHRMANN, G. &
VIVILLE, S. 2008. Tex19, a mammalian-specific protein with a restricted expression
in pluripotent stem cells and germ line. Stem Cells, 26, 734-44.
KURAMOCHI-MIYAGAWA, S., KIMURA, T., IJIRI, T. W., ISOBE, T., ASADA, N.,
FUJITA, Y., IKAWA, M., IWAI, N., OKABE, M., DENG, W., LIN, H., MATSUDA,
Y. & NAKANO, T. 2004. Mili, a mammalian member of piwi family gene, is
essential for spermatogenesis. Development, 131, 839-49.
KWON, Y. T., XIA, Z., AN, J. Y., TASAKI, T., DAVYDOV, I. V., SEO, J. W., SHENG, J.,
XIE, Y. & VARSHAVSKY, A. 2003. Female lethality and apoptosis of spermatocytes
in mice lacking the UBR2 ubiquitin ligase of the N-end rule pathway. Mol Cell Biol,
23, 8255-71.
LANDER, E. S., LINTON, L. M., BIRREN, B., NUSBAUM, C., ZODY, M. C., BALDWIN,
J., DEVON, K., DEWAR, K., DOYLE, M., FITZHUGH, W., FUNKE, R., GAGE, D.,
HARRIS, K., HEAFORD, A., HOWLAND, J., KANN, L., LEHOCZKY, J., LEVINE,
R., MCEWAN, P., MCKERNAN, K., MELDRIM, J., MESIROV, J. P., MIRANDA,
C., MORRIS, W., NAYLOR, J., RAYMOND, C., ROSETTI, M., SANTOS, R.,
SHERIDAN, A., SOUGNEZ, C., STANGE-THOMANN, N., STOJANOVIC, N.,
SUBRAMANIAN, A., WYMAN, D., ROGERS, J., SULSTON, J., AINSCOUGH, R.,
BECK, S., BENTLEY, D., BURTON, J., CLEE, C., CARTER, N., COULSON, A.,
DEADMAN, R., DELOUKAS, P., DUNHAM, A., DUNHAM, I., DURBIN, R.,
FRENCH, L., GRAFHAM, D., GREGORY, S., HUBBARD, T., HUMPHRAY, S.,
HUNT, A., JONES, M., LLOYD, C., MCMURRAY, A., MATTHEWS, L.,
MERCER, S., MILNE, S., MULLIKIN, J. C., MUNGALL, A., PLUMB, R., ROSS,
M., SHOWNKEEN, R., SIMS, S., WATERSTON, R. H., WILSON, R. K., HILLIER,
L. W., MCPHERSON, J. D., MARRA, M. A., MARDIS, E. R., FULTON, L. A.,
CHINWALLA, A. T., PEPIN, K. H., GISH, W. R., CHISSOE, S. L., WENDL, M. C.,
DELEHAUNTY, K. D., MINER, T. L., DELEHAUNTY, A., KRAMER, J. B.,
COOK, L. L., FULTON, R. S., JOHNSON, D. L., MINX, P. J., CLIFTON, S. W.,
HAWKINS, T., BRANSCOMB, E., PREDKI, P., RICHARDSON, P., WENNING, S.,
SLEZAK, T., DOGGETT, N., CHENG, J. F., OLSEN, A., LUCAS, S., ELKIN, C.,
UBERBACHER, E., FRAZIER, M., et al. 2001. Initial sequencing and analysis of the
human genome. Nature, 409, 860-921.
MA, L., BUCHOLD, G. M., GREENBAUM, M. P., ROY, A., BURNS, K. H., ZHU, H.,
HAN, D. Y., HARRIS, R. A., COARFA, C., GUNARATNE, P. H., YAN, W. &
MATZUK, M. M. 2009. GASZ is essential for male meiosis and suppression of
retrotransposon expression in the male germline. PLoS Genet, 5, e1000635.
MARK, M., JACOBS, H., OULAD-ABDELGHANI, M., DENNEFELD, C., FERET, B.,
VERNET, N., CODREANU, C. A., CHAMBON, P. & GHYSELINCK, N. B. 2008.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

STRA8-deficient spermatocytes initiate, but fail to complete, meiosis and undergo
premature chromosome condensation. J Cell Sci, 121, 3233-42.
MARTIN, M. 2011. Cutadapt removes adapter sequences from high-throughput sequencing
reads. EMBnet.journal, 17, 10.
MARTINEZ, J. & TUSCHL, T. 2004. RISC is a 5' phosphomonoester-producing RNA
endonuclease. Genes Dev, 18, 975-80.
OBBARD, D. J., GORDON, K. H., BUCK, A. H. & JIGGINS, F. M. 2009. The evolution of
RNAi as a defence against viruses and transposable elements. Philos Trans R Soc
Lond B Biol Sci, 364, 99-115.
OLLINGER, R., CHILDS, A. J., BURGESS, H. M., SPEED, R. M., LUNDEGAARD, P. R.,
REYNOLDS, N., GRAY, N. K., COOKE, H. J. & ADAMS, I. R. 2008. Deletion of
the pluripotency-associated Tex19.1 gene causes activation of endogenous retroviruses
and defective spermatogenesis in mice. PLoS Genet, 4, e1000199.
OSTERTAG, E. M. & KAZAZIAN, H. H., JR. 2001. Biology of mammalian L1
retrotransposons. Annu Rev Genet, 35, 501-38.
REICHMANN, J., REDDINGTON, J. P., BEST, D., READ, D., OLLINGER, R., MEEHAN,
R. R. & ADAMS, I. R. 2013. The genome-defence gene Tex19.1 suppresses LINE-1
retrotransposons in the placenta and prevents intra-uterine growth retardation in mice.
Hum Mol Genet, 22, 1791-806.
REUTER, M., CHUMA, S., TANAKA, T., FRANZ, T., STARK, A. & PILLAI, R. S. 2009.
Loss of the Mili-interacting Tudor domain-containing protein-1 activates transposons
and alters the Mili-associated small RNA profile. Nat Struct Mol Biol, 16, 639-46.
ROBINE, N., LAU, N. C., BALLA, S., JIN, Z., OKAMURA, K., KURAMOCHI-
MIYAGAWA, S., BLOWER, M. D. & LAI, E. C. 2009. A broadly conserved
pathway generates 3'UTR-directed primary piRNAs. Curr Biol, 19, 2066-76.
RODRIGUEZ, C. I., BUCHHOLZ, F., GALLOWAY, J., SEQUERRA, R., KASPER, J.,
AYALA, R., STEWART, A. F. & DYMECKI, S. M. 2000. High-efficiency deleter
mice show that FLPe is an alternative to Cre-loxP. Nat Genet, 25, 139-40.
ROLLINS, R. A., HAGHIGHI, F., EDWARDS, J. R., DAS, R., ZHANG, M. Q., JU, J. &
BESTOR, T. H. 2006. Large-scale structure of genomic methylation patterns. Genome
Res, 16, 157-63.
SOPER, S. F., VAN DER HEIJDEN, G. W., HARDIMAN, T. C., GOODHEART, M.,
MARTIN, S. L., DE BOER, P. & BORTVIN, A. 2008. Mouse maelstrom, a
component of nuage, is essential for spermatogenesis and transposon repression in
meiosis. Dev Cell, 15, 285-97.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

TARABAY, Y., KIEFFER, E., TELETIN, M., CELEBI, C., VAN MONTFOORT, A.,
ZAMUDIO, N., ACHOUR, M., EL RAMY, R., GAZDAG, E., TROPEL, P., MARK,
M., BOURC'HIS, D. & VIVILLE, S. 2013. The mammalian-specific Tex19.1 gene
plays an essential role in spermatogenesis and placenta-supported development. Hum
Reprod, 28, 2201-14.
VASILEVA, A., TIEDAU, D., FIROOZNIA, A., MULLER-REICHERT, T. &
JESSBERGER, R. 2009. Tdrd6 is required for spermiogenesis, chromatoid body
architecture, and regulation of miRNA expression. Curr Biol, 19, 630-9.
WANG, P. J., MCCARREY, J. R., YANG, F. & PAGE, D. C. 2001. An abundance of X-
linked genes expressed in spermatogonia. Nat Genet, 27, 422-6.
YANG, F., CHENG, Y., AN, J. Y., KWON, Y. T., ECKARDT, S., LEU, N. A.,
MCLAUGHLIN, K. J. & WANG, P. J. 2010. The ubiquitin ligase Ubr2, a recognition
E3 component of the N-end rule pathway, stabilizes Tex19.1 during spermatogenesis.
PLoS One, 5, e14017.
YODER, J. A., WALSH, C. P. & BESTOR, T. H. 1997. Cytosine methylation and the
ecology of intragenomic parasites. Trends Genet, 13, 335-40.
ZAMUDIO, N., BARAU, J., TEISSANDIER, A., WALTER, M., BORSOS, M., SERVANT,
N. & BOURC'HIS, D. 2015. DNA methylation restrains transposons from adopting a
chromatin signature permissive for meiotic recombination. Genes Dev, 29, 1256-70.
ZAMUDIO, N. & BOURC'HIS, D. 2010. Transposable elements in the mammalian germline:
a comfortable niche or a deadly trap? Heredity (Edinb), 105, 92-104.
ZHENG, K. & WANG, P. J. 2012. Blockade of pachytene piRNA biogenesis reveals a novel
requirement for maintaining post-meiotic germline genome integrity. PLoS Genet, 8,
e1003038.
ZHENG, K., XIOL, J., REUTER, M., ECKARDT, S., LEU, N. A., MCLAUGHLIN, K. J.,
STARK, A., SACHIDANANDAM, R., PILLAI, R. S. & WANG, P. J. 2010. Mouse
MOV10L1 associates with Piwi proteins and is an essential component of the Piwi-
interacting RNA (piRNA) pathway. Proc Natl Acad Sci U S A, 107, 11841-6.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le
Figures
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

Fig 1: Genetic targeting of Tex19 paralogs in mice. (A) The loci of the Tex19 paralogs are
shown before and after homologous recombination upon removal of the neomycin (Neo) gene
by crossing the mice with the Flp-recombinase strain. LoxP sites surrounding Tex19.1,
Lox5171 sites surrounding Tex19.2 and PacI restriction sites (PacI RS) are depicted; (B)
Examples of PCR genotyping; one example of each genotype, Tex19.1KO (+/+ and -/-),
Tex19.2KO (+/+ and -/-) and Tex19DKO is shown. Wild type alleles are indicated by +/+
whereas mutant alleles are indicated by -/-. (C) Western blot using the 6Tex:4D2 antibody
recognizing both paralogs on adult testes extracts from WT, Tex19.1KO and Tex19DKO
showing the absence of TEX19 proteins in the Tex19DKO. β-Tubulin is used as a loading
control. M: molecular size marker.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

Fig 2: Tex19DKO testicular defects. (A) Testes from Tex19DKO (DKO) mice are smaller
than testes from double wild type (DWT) littermates (8-week old); (B) Adult testes weight in
mg. Mean testes weight is significantly reduced in Tex19DKO mice (Student t-test *p<0.01)
(C, D) Histological sections stained with hematoxylin and eosin through the testes of adult
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

mice. (E-H) Double immunostaining of spread nuclei using antibodies against SYCP1 and
either SYCP3 or γH2AX, as indicated. In E, F, the synaptonemal complexes (SC) appear
yellow from overlapping of SYCP3 (green) and SYCP1 (red) signals. In both DWT and
Tex19DKO spermatocytes at the zygotene stage, short fragments of SC begin to form. In
DWT spermatocytes at the pachytene stage, all nineteen bivalents are fully synapsed. In about
20% of the Tex19DKO spermatocytes at the pachytene stage, only few chromosomes are fully
(arrowheads) or partially (arrow) synapsed, while the majority is unsynapsed (thin green
strands). (I, K) DWT and Tex19DKO spermatocytes at the zygotene stage express γH2AX
(green signal) throughout the nucleus. (J) In DWT spermatocytes at the pachytene stage,
γH2AX expression becomes restricted to the (almost) unsynapsed XY body. (L) In about 80%
of the Tex19DKO spermatocytes at the pachytene stage, γH2AX remains widely distributed
thought the nucleus, even in regions that are apparently synapsed as assessed from the
presence of thick SYCP3-positive strands (in red). Bar (in D): 100 m (in L): 10 m.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le
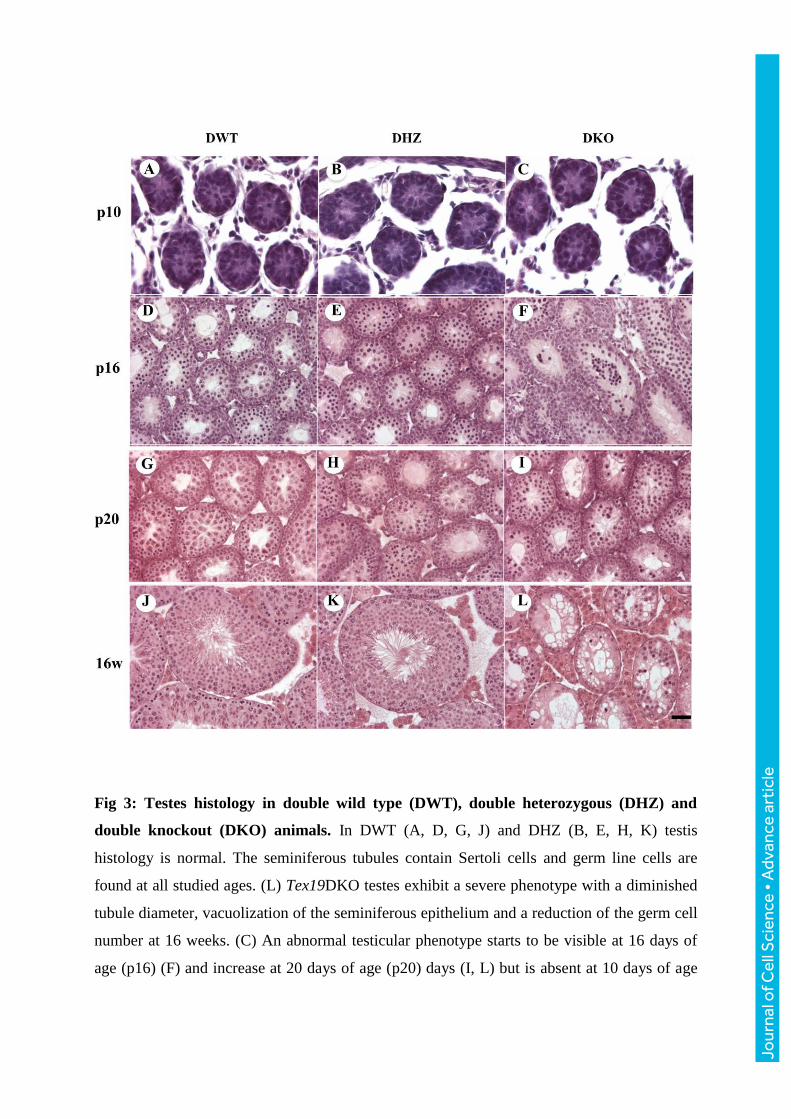
Fig 3: Testes histology in double wild type (DWT), double heterozygous (DHZ) and
double knockout (DKO) animals. In DWT (A, D, G, J) and DHZ (B, E, H, K) testis
histology is normal. The seminiferous tubules contain Sertoli cells and germ line cells are
found at all studied ages. (L) Tex19DKO testes exhibit a severe phenotype with a diminished
tubule diameter, vacuolization of the seminiferous epithelium and a reduction of the germ cell
number at 16 weeks. (C) An abnormal testicular phenotype starts to be visible at 16 days of
age (p16) (F) and increase at 20 days of age (p20) days (I, L) but is absent at 10 days of age
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

(p10) (C). Thus, the histological phenotype starts to be visible between p10 and p16. Bar (in
L): 100 μm.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

Fig 4: GST pull-down and immunoprecipitation of TEX19 paralogs. (A) Western blot
(WB) showing GST-TEX19.1 and GST-TEX19.2. (B) Western blot analysis of the GST
pulldown revealing that MAEL, MIWI, MILI, MVH, TDRD6 and RANBP9 are present in the
analyzed pull down samples. (C) Reciprocal immunoprecipitation experiment confirming the
association of MAEL, MILI, MIWI, MVH, RANBP9, EDC4 with TEX19.1 by WB. (D) Co-
immunoprecipitation of TEX19.1 by anti-TEX19.1 (7Tex: 1F11) and the detection by WB of
either MAEL, MILI, MIWI, MVH, TDRD6, RANBP9 or EDC4. IBIAB (Immuno blot with
IP antibody) is indicated with the corresponding IP antibody. Ascitic fluid from an
unimmunized animal is used as a negative control.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

Fig 5: Interaction of TEX19.1 and TEX19.2 with ~ 30 nt RNAs. MILI, TEX19.1 or
TEX19.2 were immunoprecipited (IP) from two month-old mouse testis protein extract by
specific antibodies and bound RNA were then 32P-end-labeled and separated on a 15%
denaturing urea-polyacrylamide gel. Immunoprecipited RNAs are predominantly ~ 30 nt
long. The size and mobility of the oligoribonucleotide markers is indicated on the right. The
asterisk indicates RNAase treatment. Ctrl- indicates negative control; Ascitic fluid from an
unimmunized animal is used as a negative control.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

Fig 6: PiRNA populations isolated from TEX19 paralogs and MILI complexes were
cloned, sequenced and annotated. A) Size distribution of Small RNA-seq libraries
immunoprecipitated with TEX19 and TEX19.1 from WT adult testes, and with TEX19 from
Tex19.1-ko. These libraries exhibit a peak around 29-30nt, which is reminiscent of MIWI-
associated piRNAs ; in contrast, MILI-associated piRNAs range in size between 26-27nt. (B)
Genomic annotation of TEX19.1-associated reads compared to MILI and MIWI-associated
reads. In agreement with piRNA size, the analysis was restricted to 25-32nt long reads. (C)
The vast majority of TEX19.1 reads have unique hits on the genome, similarly to MILI and
MIWI-associated Small RNAs. (D) Venn diagram distribution of piRNA cluster defined by
the piRNA population associated with TEX19.1 compared to MILI and MIWI. (E) Size
distribution of whole adult testes Small RNA-seq libraries obtained from a WT and three
Tex19.1-KO animals. The 22-23nt peak corresponds to microRNAs, the piRNAs are clustered
in two peaks, at 26-27nt and at 29-30nt.
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

Fig 7: Tex19 paralogs interact directly with ~26-30 nt piRNA: (A) and (B) Electrophoretic
Mobility Shift Assay “EMSA” was performed in the presence of 5µM of GST-alone, GST-
TEX19.1, GST-D1 (1-124aa), GST-D2Cter (125-351aa), GST-D2 (125-163aa “VPTEL”) or
GST-Cter (164-351aa) of TEX19.1 or GST-TEX19.2. All proteins were incubated with 125
femtomoles (8000cpm) of radiolabeled piR-117061 or modified piR-117061. (C) An
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

increased concentration of the modified non-radiolabeled piR-117061(2C) was used with a
fixed concentration (5µM) of TEX19.1 VPTEL domain and fixed quantity of radiolabeled
piR-117061 (8000 cpm). (D) An increased concentration of the non-radiolabeled piR-117061
was used with a fixed concentration of the VPTEL domain of TEX19.1 (5 µM) and a fixed
quantity of radiolabeled piR-117061 (8000 cpm).
Jour
nal o
f Cel
l Sci
ence
• A
dvan
ce a
rtic
le

DWT DKO
Adu
ltp1
2
A B
DWT DKO
C
Fig S1: (A) and (B) Histological sections stained with hematoxylin andeosin through the caudal epididymis of adult mice, note absence ofsperm in the mutant epididymis (C) TUNEL assays: the positive signalwas converted to a red false color and superimposed with the DAPInuclear stain (blue false color); Note a normal, low, proportion ofapoptotic germ cells in DWT testes but an increase in the proportion ofTUNEL- positive germ cells in Tex19DKO testes (black arrows).
J. Cell Sci. 130: doi:10.1242/jcs.188763: Supplementary information
Jour
nal o
f Cel
l Sci
ence
• S
uppl
emen
tary
info
rmat
ion
0
5
10
15
20
25
L1ORF2 expression in testis at p16
0
1
2
3
4
5
6
IAP∆1 expression in testis at p16
0
5
10
15
20
25
MuERV1 expression in testis at p16
DWT DKO(n=3) (n=4)
0
50
100
150
200
250
300
350
400
450
MMERVK10C expression in Tex19-/-
testes at P16
0
50
100
150
200
250
300
MMERVK10C expression in Tex19-/-
testes at P10
* *
DWT DKO(n=2) (n=3)
DWT DKO(n=2) (n=3)
DWT DKO(n=2) (n=3)
DWT DKO(n=2) (n=3)
Fig S2: RT-qPCR of MMERVK10C, L1ORF2, IAP∆1 and MuERV1retrotransposon expression in Tex19DKO (DKO) p10 and p16 micetestes compared to double wildtype (DWT). Expression levels werenormalized to the β-actin or Rrm2 housekeeping gene, n valuesindicate the number of biological replicates, DWT: double wild type;DKO: double knockout,*p<0.03 (Student t-test).
J. Cell Sci. 130: doi:10.1242/jcs.188763: Supplementary information
Jour
nal o
f Cel
l Sci
ence
• S
uppl
emen
tary
info
rmat
ion

GST
-TE
X19
.1
G-s
epha
rose
be
ads
GST
28
36
557295
130150
kDa
GSTTEX19.1 full length
GST-D1 1-124 aa GST-D2 125 -351 aa
BI AI 1 2 3
BI AI 1 2 3 BI AI 1 2 3
BI AI 1 2 3
GST
GSTVPTEL 125-163 aa
28
36
557295
130150
GSTTEX19.2 full lengh
Tex19.2
BI 1 2 3
VPTELD1
D2
GST
Tex19.1
kDa
28
36
557295
130150
kDa
B
A
Fig S3 : (A) Silver staining of GST-Tex19.1 pull down using G-sepharose beads. G-sepharose beads and GST combined to G-sepharose beads are deposited as control. (B) Production andpurification of GST, GST-TEX19.1, GST-TEX19.2, GST-MCP (1-124aa), GST-VPTEL Cter (125-351aa) or GST-VPTEL (125-163aa) ofTEX19. Gel 10% and bleu Coomassie coloration were used.
J. Cell Sci. 130: doi:10.1242/jcs.188763: Supplementary information
Jour
nal o
f Cel
l Sci
ence
• S
uppl
emen
tary
info
rmat
ion

0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
U1 A10
VPTEL (125-163aa)
D2(125-351aa)
MCPP VPTELTEX19.1
MCPPD1(1-124aa)
VPTEL
VPTEL
Cter(164-351aa)
M C P P V S V R H G A R G M S C L Y E A W L Y H L V H G E Q T K I C F A C F K A A F
L L N K L Y L E M G D W Q E E E E E E E E E D A D L L E Y L S E S E S E S E Q E P G P
E Q D A W R G L G S L Y V P Q S V S E G S G V L L P T P V W T Q G I L F S I F V P T E L
F P Q E A V P L D L G P E D A E W T Q A L P W R L D G L F P C S H Q L I P P L T W W D
I F D V M P S P G Q P V L L E L R C H W P L D Q T V A Q S W L Q D Q K F V L L L D S
V Q S R C H L L S M R V R W V V R T Q V Q H W Q V L L D P G E M W V A H F R K E V
G Q H G L Y H Q S L N P W R L S I L T A S E L G M E L L P A T C Y L W N K G F W V G
S F L P W H I N M P E T W S W E P G E R L F I T D A T I C G T D Y H L A Q S F L D S H
P T P H P L L T L T P
TEX19.1
MC
PV
PT
EL
94.42%
30%
TEX19.1 piRNA library
Fig S4: A: Percentage of the distribution of TEX19.1 piRNA. TEX19.1
J. Cell Sci. 130: doi:10.1242/jcs.188763: Supplementary information
Jour
nal o
f Cel
l Sci
ence
• S
uppl
emen
tary
info
rmat
ion
Adapter triming 15-45bp Alignment, 2 mismatches
Sample Out percent
Input Too-short Adapter Non-clipped
Output Aligned reads
Failed Suppressed-m
Output reads
Tex19.1 in WT (1)
27.68% 6985974 4980436 3241 68727 1933570 1647203 (85.19%)
283557 (14.66%)
2810 (0.15%)
15875788
Tex19.1 in WT (2)
22.64% 6368438 4826897 2150 97851 1441540 1214051 (84.22%)
225055 (15.61%)
2434 (0.17%)
12610662
Tex19.2 in Tex19.1-/-
78.52% 15596146 3035682 176061 139045 12245358 9700101 (79.21%)
2524130 (20.61%)
21127 (0.17%)
117286602
Tex19.1 & 2 in WT
72.35% 13615661 3590834 23129 150175 9851523 8202474 (83.26%)
1637355 (16.62%)
11694 (0.12%)
83224246
Table S1: Adapter trimming and alignment
piRNA name Absolute count Tex19.1 in WT (1)
Absolute count Tex19.1 in WT (2)
piRNA sequence
gi|109016873|gb|DQ727168.1|piR-142490 5367 4008 ACUCGAGACUGUGUAGAACCUUCAUAGGCgi|108994920|gb|DQ710717.1|piR-126039 3828 2505 UUUGAAAUCAGGACUGUUCUGGAAAUUCCgi|108994167|gb|DQ710038.1|piR-125360 3009 2480 CUCCAGAUUAAAAAAUGUCCTAAGUUAGAGCUgi|108955533|gb|DQ687738.1|piR-103060 2742 2323 UGAAGUGAACCUGAGCAAUUCCAUGUGGA
gi|109001017|gb|DQ715342.1|piR-130664 2684 1962 UAUGAGUAGAACAAAUGGGGGCAAACCCUAGCgi|108984009|gb|DQ702024.1|piR-117346 2363 1839 UGGAUAAUAUUCCGGAUAAUAAACUGAGC
gi|108983531|gb|DQ701739.1|piR-117061 2361 2028 UAGGACCUGAGAACUUAACCUUGUUAUGGGgi|109013632|gb|DQ724840.1|piR-140162 2180 1319 UCUAGGUCUCUGAAAUUUACUAUGCACAGGGUgi|108036260|gb|DQ564985.1|piR-5073 2015 1516 UACCUCGGUCUCUGUAGAAUCCAGUUUUgi|108995087|gb|DQ710862.1|piR -126184 1989 1397 UACAGACUGAUAAACGAUUUAAAGUAAAAAGU
gi|108958333|gb|DQ688999.1|piR-104321 1635 1122 UACUGGAACACUGGAUUAACAGAACGUUCAgi|108954985|gb|DQ687488.1|piR-102810 1594 1421 UAAAUGGACUUUGGCCCCAGGUCAAUUCUGGC
gi|109012695|gb|DQ724202.1|piR-139524 1479 1030 UGAUAGAAUGUAGCCUCUGACCGGGAAUAgi|108951359|gb|DQ684704.1|piR-100026 1464 1009 UAGACUGUGAGCUCCUUGAGGACAGGGACAGUgi|108969977|gb|DQ694365.1|piR-109687 1399 1366 UACGAGAUCGUCCCUUAAAGCUCAAAGGACA
Table S2: Tex19.1 reads display a strong preference for a uridine at position one (U1-bias)
J. Cell Sci. 130: doi:10.1242/jcs.188763: Supplementary information
Jour
nal o
f Cel
l Sci
ence
• S
uppl
emen
tary
info
rmat
ion