synergistic effects of brain-derived neurotrophic factor and ciliary neurotrophic factor on cultured...
TRANSCRIPT

Ž .Developmental Brain Research 115 1999 25–32
Research report
Synergistic effects of brain-derived neurotrophic factor and ciliaryneurotrophic factor on cultured basal forebrain cholinergic neurons from
postnatal 2-week-old rats
Yuka Hashimoto, Yasuhiro Abiru 1, Chika Nishio, Hiroshi Hatanaka )
DiÕision of Protein Biosynthesis, Institute for Protein Research, Osaka UniÕersity, 3-2 Yamadaoka, Suita, Osaka 565-0871, Japan
Accepted 16 March 1999
Abstract
Ž . Ž .Brain-derived neurotrophic factor BDNF , a member of the neurotrophin family, and ciliary neurotrophic factor CNTF , a member ofthe neurocytokine family, are known to have synergistic effects on motoneurons, but such synergistic effect has not been studied in detailespecially in the brain. In the present study, we examined the synergistic effects of BDNF and CNTF on the survival of basal forebrain
Ž .cholinergic neurons cultured from postnatal 2-week-old P2w rats. Although BDNF is well-known to promote the survival of basalŽ .forebrain cholinergic neurons in P2w culture, CNTF had little effect on the survival of choline acetyltransferase ChAT -positive neurons
and did not increase ChAT activity in the culture. However, CNTF enhanced BDNF-mediated promotion of cell survival of cholinergicneurons when added concomitantly. BDNF alone induced only a three-fold increase in ChAT activity in control cultures, but theconcomitant addition of CNTF resulted in an eight-fold increase. CNTF did not enhance BDNF-mediated cell survival of total neuronsfrom the basal forebrain, hippocampus or cerebellum, suggesting that the synergistic effects of CNTF on the BDNF-mediated increase ofviability might be strong in basal forebrain cholinergic neurons. CNTF also enhanced the neurotrophin-4r5-mediated increase of ChAT
Ž .activity, but not the nerve growth factor NGF -mediated one. Furthermore, the BDNF-mediated increase was also enhanced by leukemiainhibitory factor but not by interleukin-6. Similar synergistic pattern between neurotrophins and cytokines were also observed in theinduction of ChAT activity in embryonic basal forebrain culture. These results suggest that TrkB, a functional high-affinity receptor ofBDNF and NT-4r5, and LIFR b, a receptor component contained in CNTF and LIF receptor complex, might be involved in the observedsynergistic effects. q 1999 Elsevier Science B.V. All rights reserved.
Keywords: Neuronal cell death; Neurotrophic factor; Neurotrophin; trk; Cytokine
1. Introduction
There are several families of neurotrophic factors whichare involved in neuronal differentiation, survival and re-
w xgeneration in the nervous system 26 . The neurotrophinŽ .family consists of nerve growth factor NGF , brain-de-
Ž . Ž .rived neurotrophic factor BDNF , neurotrophin-3 NT-3and NT-4r5. The members of the neurotrophin family areall small basic proteins comprised of about 120 aminoacids that have about 50% amino acid sequence identity.Neurotrophins act on distinct neuronal populations both in
) Corresponding author. Fax: q81-6-6879-8626; E-mail:[email protected]
1 Present address: Tokushima New Drug Research Institute, OtsukaPharmaceutical, 463-10 Kagasuno, Kawauchi-cho, Tokushima 771-0192,Japan.
w xthe PNS and CNS 26 . For example in the CNS, NGF andBDNF promote the survival and differentiation of basal
w xforebrain cholinergic neurons 14–16,30 . On the otherhand, BDNF and NT-3, but not NGF, support the survivalof dopaminergic neurons of the developing substantia ni-
w xgra 18 .Another group of neurotrophic factors is the cytokine
Ž .family including ciliary neurotrophic factor CNTF ,Ž . Ž .leukemia inhibitory factor LIF and interleukin-6 IL-6
w x32 . These cytokines are all composed of about 200 aminoacids and have about 24–30% amino acid sequence iden-
w xtity with each other 4 . CNTF originally identified as asurvival factor for chicken ciliary ganglion neurons invitro, has been reported to promote survival of embryonic
w xmotor, sensory and hippocampal neurons in vitro 3,20 ,and to prevent axotomy-induced neuronal death in the
w xseptum, substantia nigra and thalamus in vivo 6,11,12,31 .
0165-3806r99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved.Ž .PII: S0165-3806 99 00047-4

( )Y. Hashimoto et al.rDeÕelopmental Brain Research 115 1999 25–3226
It has also been reported that CNTF augments theeffects of BDNF on motoneurons. The BDNF-inducedincrease in ChAT activity in cultures of E14 rat spinalmotoneurons was shown to be enhanced synergistically by
w xCNTF 35 . Furthermore, degeneration of motor neurons invivo was markedly prevented by the combined infusion ofBDNF and CNTF in comparison with infusion of either
w xmolecule alone 28 . However, there have been few studiesof the synergistic effects of BDNF and CNTF in the brain.In this study, we investigated the effects of concomitantaddition of BDNF and CNTF, as well as other neu-rotrophins and cytokines, on the survival of basal forebraincholinergic neurons cultured from postnatal 2-week-oldŽ .P2w rats.
2. Materials and methods
2.1. Neurotrophins, cytokines and the other chemicals
Neurotrophic factors were obtained from the followingsources. NGF was purified from adult male mouse sub-
w xmandibular glands as described 5,34 . BDNF and NT-4r5,produced in Escherichia coli and purified as describedw x8,9 , were provided by Amgen–Regeneron Partners andRegeneron, respectively. NT-3 and CNTF were also re-combinant human proteins and were provided by Regen-eron. The neurotrophins were dissolved at 50 or 25 mgrmlin PBS containing 1 mgrml BSA as stock solutions.Human recombinant LIF and IL-6 were purchased fromPharma Biotechnologie Hannover. LIF was dissolved at 50mgrml in PBS containing 1 mgrnl BSA as a stocksolutions. IL-6 was dissolved at 5 mgrml in DF mediumŽa 1:1 mixture of Dulbecco’s modified Eagle’s mediumŽ . .DMEM and Ham’s F12 medium; both from Gibco as astock solution.
Anti-MAP2 antibody was a gift from Dr. H. MurofushiŽ .The University of Tokyo . An anti-choline acetyltrans-
Ž .ferase ChAT antibody was purchased from Chemicon.Media for cell culture were obtained from Gibco. Otherchemicals of analytical grade were purchased from Sigma,unless otherwise stated.
2.2. Cell culture
Dissociated primary culture of postnatal rat basal fore-brain, striatal, hippocampal and cerebellar neurons was
w xcarried out by the method described previously 16,23 .Here, we used postnatal 10- to 15-day-old rats as P2w rats.Briefly, tissue fragments of the basal forebrain area con-taining the septum and the vertical limb of the diagonalband, the striatum, the hippocampus and the cerebellum
Žwere dissected out from P2w rats Wistar ST, both sexes,.Shizuoka , and quickly transferred into ice-cold L-15Ž .medium Gibco bubbled with oxygen. The tissue frag-
Ž .ments were digested by papain Worthington, 90 units
twice for 15 min at 378C, then resuspended in 5r5r1rDFmedium containing 5% precolostrum newborn calf serumŽ . Ž .PNCS, Mitsubishi Kasei , 5% horse serum Gibco , 1%
Ž .rat serum prepared in our laboratory , and 89% DFmedium. Cells were then dissociated by gentle drawing
Žthrough plastic tips of two sizes 1.2 mm followed by 0.8.mm in diameter . To remove contaminating fragments of
nerve fibers, cells were layered onto 10 ml of PNCS, andcentrifuged at 900 rpm for 5 min. These cells were resus-pended in the above medium and seeded at a density of
5 2 Ž 23–5=10 cellsrcm onto 48-well plates 0.65 cm rwell;.Sumitomo Bakelite coated as follows. For immunocyto-
chemistry, culture wells were covered with an astrocytemonolayer prepared from 20-day fetal rat cortex and hip-
w xpocampus as described 15 . For measurement of ChATactivity, culture wells were coated with polyethyleneimine.Seeded cells were cultured in a humidified 5% CO incu-2
bator at 378C.After 1 day of culture, the medium was changed to the
following medium and the indicated neurotrophic factorswere added to the wells. For the basal forebrain and striatalcultures, medium was changed to serum-containing
Ž5r5r1rDF medium astrocyte-covered or polyeth-.yleneimine-coated wells or to serum-free TIPrDF medium
consisting of DF medium containing 5 mgrml humanŽ . Ž .transferrin Sigma , 5 mgrml bovine insulin Colab Res. ,
Ž .and 20 nM progesterone Sigma . For hippocampal cul-ture, medium was changed to Minimum Essential MediumŽ .MEM, Gibco containing 30 mM Na SeO , 0.5 mgrml2 4
NaHCO , 2.2 mgrml glucose and 5% horse serum. For3
cerebellar culture, medium was also changed to MEM, andthe cells were cultured in a humidified 10% CO incubator2
at 378C. Cells were cultured further for the indicatedperiods and the medium was changed every 3 days.
2.3. Immunocytochemistry, MTT assay and determinationof ChAT actiÕity
Cholinergic neurons were stained with an antibodyw xagainst ChAT as described 29 . Briefly, cells were first
fixed with 4% paraformaldehyde for 20 min at roomtemperature. The cells were then blocked with phosphate-
Ž .buffered saline PBS , containing 5% normal goat serum,0.3% Triton X-100 and 0.05% sodium azide, followed byovernight incubation at 48C with a rabbit anti-ChAT anti-
Ž .body 1:500; Chemicon . Cells were then incubated withbiotinylated secondary antibody and immunoreactive cells
Ž .were visualized with a Vectastain ABC kit Vector LabsŽ .with nickel II ammonium sulfate intensification. The
whole culture well was scanned when counting ChAT-positive neurons.
Ž .Acetylcholinesterase AChE staining was performed asw xdescribed 29 . Briefly, cells were fixed with 4% para-
Ž .formaldehyde in sodium phosphate buffer pH 7.4 for 20min. Then, cells were incubated for 2 h at room tempera-ture in a reaction mixture containing acetylcholine iodate
( )Y. Hashimoto et al.rDeÕelopmental Brain Research 115 1999 25–32 27
as the substrate and tetraisopropyl pyrophosphoramide as apseudocholinesterase inhibitor. AChE-positive neurons
Ž . Ž .were visualized with DAB Dojindo with nickel Laammonium sulfate intensification.
Total neurons in culture were identified by staining withantibodies against microtubule-associated protein 2Ž . w xMAP2 or by MTT assay as described 22,23 . The im-munocytochemical method for MAP2 staining was carriedout in the same manner as that of ChAT staining exceptthat the concentration of Triton X-100 was 0.1% and the
Ž .antibody used was rabbit anti-MAP2 1:5000 . The numberof MAP2-stained neurons in seven microscopic fields perwell was counted and averaged, then the number per wellwas calculated.
ChAT activity was assayed according to the procedurew x w xof Fonnum 10 with some modifications 13 .
Statistical significance was estimated by Student’s t-test.
3. Results
3.1. CNTF enhanced BDNF-mediated surÕiÕal effect onthe P2w basal forebrain cholinergic neurons
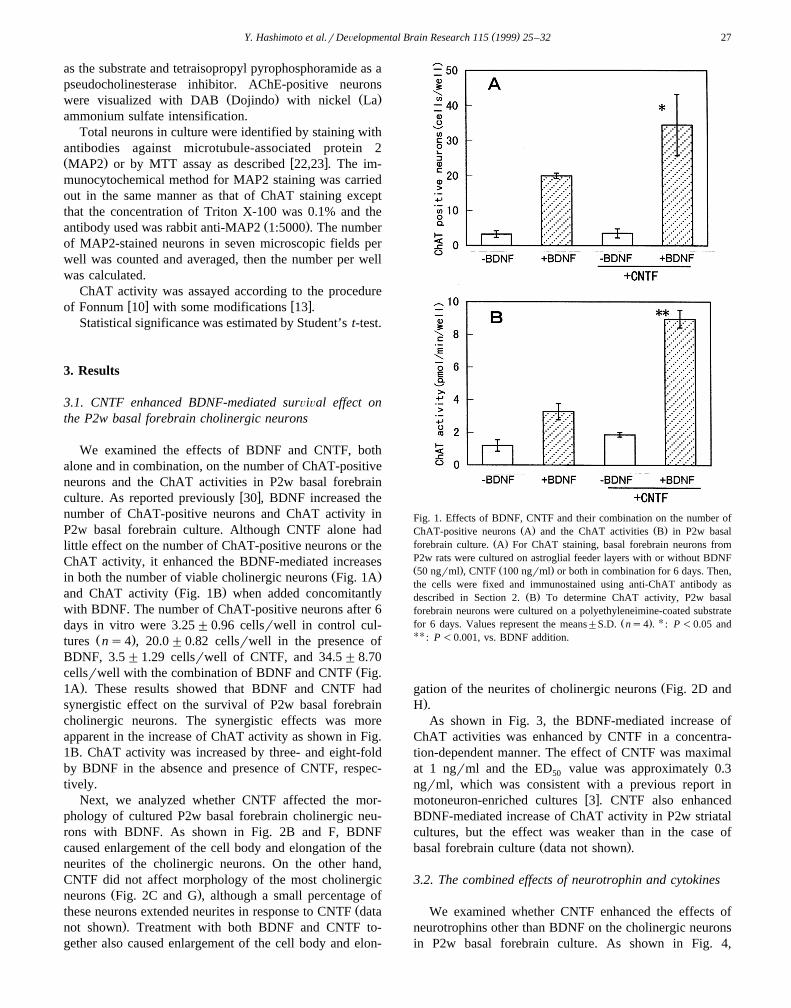
We examined the effects of BDNF and CNTF, bothalone and in combination, on the number of ChAT-positiveneurons and the ChAT activities in P2w basal forebrain
w xculture. As reported previously 30 , BDNF increased thenumber of ChAT-positive neurons and ChAT activity inP2w basal forebrain culture. Although CNTF alone hadlittle effect on the number of ChAT-positive neurons or theChAT activity, it enhanced the BDNF-mediated increases
Ž .in both the number of viable cholinergic neurons Fig. 1AŽ .and ChAT activity Fig. 1B when added concomitantly
with BDNF. The number of ChAT-positive neurons after 6days in vitro were 3.25"0.96 cellsrwell in control cul-
Ž .tures ns4 , 20.0"0.82 cellsrwell in the presence ofBDNF, 3.5"1.29 cellsrwell of CNTF, and 34.5"8.70
Žcellsrwell with the combination of BDNF and CNTF Fig..1A . These results showed that BDNF and CNTF had
synergistic effect on the survival of P2w basal forebraincholinergic neurons. The synergistic effects was moreapparent in the increase of ChAT activity as shown in Fig.1B. ChAT activity was increased by three- and eight-foldby BDNF in the absence and presence of CNTF, respec-tively.
Next, we analyzed whether CNTF affected the mor-phology of cultured P2w basal forebrain cholinergic neu-rons with BDNF. As shown in Fig. 2B and F, BDNFcaused enlargement of the cell body and elongation of theneurites of the cholinergic neurons. On the other hand,CNTF did not affect morphology of the most cholinergic
Ž .neurons Fig. 2C and G , although a small percentage ofŽthese neurons extended neurites in response to CNTF data
.not shown . Treatment with both BDNF and CNTF to-gether also caused enlargement of the cell body and elon-
Fig. 1. Effects of BDNF, CNTF and their combination on the number ofŽ . Ž .ChAT-positive neurons A and the ChAT activities B in P2w basal
Ž .forebrain culture. A For ChAT staining, basal forebrain neurons fromP2w rats were cultured on astroglial feeder layers with or without BDNFŽ . Ž .50 ngrml , CNTF 100 ngrml or both in combination for 6 days. Then,the cells were fixed and immunostained using anti-ChAT antibody as
Ž .described in Section 2. B To determine ChAT activity, P2w basalforebrain neurons were cultured on a polyethyleneimine-coated substrate
Ž . Ufor 6 days. Values represent the means"S.D. ns4 . : P -0.05 andUU : P -0.001, vs. BDNF addition.
Žgation of the neurites of cholinergic neurons Fig. 2D and.H .As shown in Fig. 3, the BDNF-mediated increase of
ChAT activities was enhanced by CNTF in a concentra-tion-dependent manner. The effect of CNTF was maximalat 1 ngrml and the ED value was approximately 0.350
ngrml, which was consistent with a previous report inw xmotoneuron-enriched cultures 3 . CNTF also enhanced
BDNF-mediated increase of ChAT activity in P2w striatalcultures, but the effect was weaker than in the case of
Ž .basal forebrain culture data not shown .
3.2. The combined effects of neurotrophin and cytokines
We examined whether CNTF enhanced the effects ofneurotrophins other than BDNF on the cholinergic neuronsin P2w basal forebrain culture. As shown in Fig. 4,

( )Y. Hashimoto et al.rDeÕelopmental Brain Research 115 1999 25–3228
Ž . Ž .Fig. 2. Effects of BDNF, CNTF and both in combination on the morphology of ChAT A–D and AChE E–H -positive neurons in P2w basal forebrainŽ . Ž .culture. P2w basal forebrain cells were cultured on the astroglial feeder layers without A and E or with 50 ngrml BDNF B and F , 100 ngrml CNTF
. Ž . Ž . Ž .and G or both in combination D and H for 6 days. Then, the cells were stained by ChAT A–D and AChE E–H staining as described in Section 2.Bar, 50 mm.
treatment with NGF and CNTF together resulted in anadditive, increase in ChAT activity. In contrast, treatmentwith CNTF and either BDNF or NT-4r5 showed synergis-tic effects. NT-3 had no apparent effect both in the absenceand presence of CNTF. These results suggested that CNTFcan enhance effects mediated by TrkB, but not by TrkA orTrkC.
We also examined whether LIF or IL-6, a member ofneuropoietic cytokine family, can augment the BDNF-in-duced increase of ChAT activity in P2w basal forebraincultures. As shown in Fig. 5, LIF, similarly to CNTF, didnot increase the ChAT activity by itself, but enhanced the
BDNF-mediated increase in ChAT activity synergistically.On the other hand, the effect of IL-6 was weak both in the
Ž .absence and presence of BDNF Fig. 5 . It has beenreported that the receptor complexes for CNTF or LIFcontain a signal transducer, LIF b, while that for IL-6 doesnot. Thus, these results suggested that LIF b might beinvolved in enhancement of the action of BDNF.
3.3. Combined effects of BDNF and CNTF on CNS neu-rons other than cholinergic neurons
To assess the effects of BDNF and CNTF on CNSneuronal populations other than P2w basal forebrain

( )Y. Hashimoto et al.rDeÕelopmental Brain Research 115 1999 25–32 29
Fig. 3. Concentration-dependent effects of CNTF enhancement onBDNF-mediated increase of ChAT activity in P2w basal forebrain cul-ture. P2w basal forebrain neurons were cultured on a polyethyleneimine-coated substrate with 50 ngrml BDNF in the absence and presence ofvarious concentrations of CNTF for 6 days, then ChAT activities were
Ž . Udetermined. Values represent the means"S.D. ns4 . : P -0.05 andUU : P -0.001, vs. BDNF addition. U : P -0.05 and UU : P -0.001, vs.BDNF addition.
cholinergic neurons, we examined the viability of totalneurons in cultures from the P2w basal forebrain, postnatal
Ž .day 15 P15 hippocampus and P9 cerebellum. The con-comitant addition of BDNF and CNTF did not synergisti-cally promote the survival of total neurons in any cultureexamined. Total neurons were recognized as MAP2-posi-
Ž .tive cells, in cultures from the basal forebrain Fig. 6AŽ . Ž .and hippocampus Fig. 6B . In cerebellar culture Fig. 6C ,
no synergistic effects of BDNF and CNTF was observed
Fig. 4. Effects of CNTF on the ChAT activities induced by neurotrophinfamily in P2w basal forebrain culture. P2w basal forebrain neurons were
Ž .cultured on a polyethyleneimine-coated substrate with NGF 100 ngrml ,Ž .BDNF, NT-3, NT-4r5 all at 50 ngrml in the absence and presence of
Ž .CNTF 100 ngrml for 6 days, then ChAT activities were determined.Ž . UValues represent the means"S.D. ns4 . : P -0.005, vs. BDNF or
NT-4r5 addition.
Fig. 5. Effects of cytokines on BDNF-induced increase of ChAT activi-ties in P2w basal forebrain culture. P2w basal forebrain neurons were
Ž .cultured on a polyethyleneimine-coated substrate with BDNF 50 ngrml ,Ž .in the presence or absence of CNTF, LIF or IL-6 all at 100 ngrml for 6
days, then ChAT activities were determined. Values represent the meansŽ . U"S.D. ns4 . : P -0.001 vs. BDNF addition.
on total neuronal viability evaluated by MTT assay. Theseresults showed that synergistic effects of BDNF and CNTFmight be strong in cholinergic neurons of the basal fore-brain.
In addition, we examined the effects of BDNF andCNTF in combination on embryonic E20 basal forebrain
Ž .cholinergic neurons Fig. 7 . A synergistic increase ofChAT activity was induced by the concomitant addition ofBDNF and CNTF, but not by that of NGF and CNTF, asin the case in P2w cultures.
3.4. The possible role of glial cells in the synergistic actionof BDNF and CNTF on P2w basal forebrain cholinergicneurons
As considerable numbers of glial cells are present inP2w cultures, it is conceivable that the synergistic effectsof BDNF and CNTF observed in the present study mighthave been mediated through modulation or release oftrophic activity in glial cells. To address this issue, weexamined the combined effects of BDNF and CNTF undertwo different culture conditions; one containing large num-bers of glial cells and the other containing only a few suchcells. As the proliferation of non-neuronal glial cells isenhanced by addition of serum and suppressed by removalof serum and addition of AraC, we cultivated P2w basalforebrain cells as sister cultures with serum-containing5r5r1rDF medium and with serum-free TIPrDF mediumcontaining 1 mM AraC. We could construct much differentculture conditions, that is, the former cultures contained alarge number of glial cells, while the latter contained onlysmall number of glial cells. If the enhanced effects ofCNTF were mediated by glial cells, stronger synergistic

( )Y. Hashimoto et al.rDeÕelopmental Brain Research 115 1999 25–3230
effects would be expected in cultures using serum-contain-ing than serum-free medium. Treatment with BDNF andCNTF together showed the same synergistic increase ofChAT activity in both serum-containing and serum-free
Fig. 6. Effects of BDNF, CNTF and both in combination on the survivalŽ . Ž .of total neurons from P2w basal forebrain A , P15 hippocampus B and
Ž . Ž .P9 cerebellum C in culture. A and B Neurons from P2w basalforebrain and P15 hippocampus were cultured on astroglial feeder layerswith BDNF, CNTF or both in combination for 6 days, then total neurons
Ž .were immunostained using anti-MAP2 antibody. C Neurons from P9cerebellum were cultured on the polyethyleneimine-coated substrate withthe above neurotrophic factors for 3 days, then MTT assay was per-
Ž .formed. Values represent the means"S.D. ns4 .
Fig. 7. Effects of CNTF on NGF- and BDNF-induced increase of ChATactivity in E20 basal forebrain cultures. The neurons from E20 basalforebrain were cultured on the polyethyleneimine-coated substrate withNGF and BDNF in the absence or presence of CNTF for 6 days, thenChAT activities were determined. Values represent the means"S.D.Ž . Uns4 . : P -0.05, vs. BDNF addition.
Ž .medium data not shown , suggesting that BDNF andCNTF act directly on neurons.
4. Discussion
Various neurotrophic factors are expressed in the CNS,and they are supplied to neurons not only by retrogradetransport, but also by anterograde transport and autocrine
w xloop 1,2,7,21 . Receptors for neurotrophic factors displaydiscrete distribution patterns, some of which apparently
w xoverlap 26 . Thus, it seems likely that a single neuronmight be affected by several neurotrophic factors under invivo circumstance. Among the various neurotrophic fac-tors, BDNF and CNTF have been reported to have syner-
w xgistic effects on motoneurons 28,35 . However, such co-operative effects of BDNF and CNTF have not yet beenstudied in detail especially in the brain.
It has been suggested that CNTF might have a physio-logical role in neuronal injury. CNTF lacks a classicalsignal sequence for secretion and is located within the
w xcytoplasm 33 . CNTF mRNA is present at very low levelsin the normal adult rat CNS, but increases markedly afterlesioning of the dorsal hippocampus and overlying cortexin the area coincident with reactive astrocytes. The in-creased level of CNTF mRNA in the lesioned hippocam-pus becomes maximal 3 days after lesioning and is sus-
w xtained for up to 20 days 27,31 . In contrast, levels ofBDNF and NT-3 mRNA slightly are decreased during the
w xsame period 19 . In addition, after fimbria–fornix transec-tion the expression of CNTFR a mRNA was shown toincrease in basal forebrain neurons, and CNTF preventsthe degeneration of basal forebrain cholinergic neuronsw x24 . In contrast, BDNF was suggested to play many

( )Y. Hashimoto et al.rDeÕelopmental Brain Research 115 1999 25–32 31
physiological roles during and after the development of theCNS, and also BDNF expression is observed during andafter development with high levels of expression in the
w xhippocampus and cortex 26 . Thus, considering the ex-pression patterns of CNTF and BDNF, it seems likely thatCNTF collaborates with BDNF, especially in case of neu-ronal injury in the hippocampus and the cortex, both ofwhich are target areas of basal forebrain cholinergic neu-rons, especially in the case of neuronal injury.
In this study, we investigated the cooperative effects ofBDNF and CNTF on the survival of basal forebrain cholin-ergic neurons in P2w rat cultures. As shown in Fig. 1A,CNTF enhanced the survival-promoting effect of BDNFon P2w basal forebrain cholinergic neurons, althoughCNTF alone had little effect on survival. Accordingly, itseems likely that the augmentative effect of CNTF on the
ŽBDNF-mediated increase of ChAT activity per well Fig..1B reflects the viable number of ChAT-positive neurons
Ž .per well Fig. 1A at least partially. However, CNTF mayalso enhance neuronal differentiation in addition to neu-ronal survival, because the increase in ChAT activityinduced by CNTF alone was weak but significant in somecases, and because small numbers of cholinergic neurons
Žextended their neurites in response to CNTF data not.shown .
In embryonic basal forebrain cultures, CNTF enhancedthe BDNF-induced increase of ChAT activity as shown inFig. 7. As the increase in ChAT activity in embryonic
w xculture represents cholinergic differentiation 13 , Fig. 7implies that CNTF enhances BDNF-mediated cholinergicdifferentiation in embryonic E20 basal forebrain culture.CNTF is unlikely to be activated during the embryonicstage, although CNTFR a has already been expressedw x33 . Presumably, endogenous ligands other than CNTFcould act on the neurons which contain CNTFR a . Alikely candidate for such a molecule is a chick growth-pro-moting protein, which is structurally similar to CNTF andexpressed during the embryonic period. This ligand or aspecies homologue has been reported to display similar,but not identical, receptor-binding characteristics to CNTFw x7,17,25 .
On the other hand, CNTF did not enhance the BDNF-mediated survival of total neurons in culture from the P2w
Žbasal forebrain, P15 hippocampus or P9 cerebellum Fig..6 . In contrast, CNTF also enhanced the BDNF-mediated
Žincrease of ChAT activity in P2w striatal culture data not.shown . These observations suggested that BDNF and
CNTF collaborate synergistically at least in the case ofcholinergic neurons in the CNS.
Fig. 4 shows that CNTF synergistically enhanced theeffect of NT-4r5 as well as BDNF, but not those of NGFor NT-3. Considering that TrkA, TrkB and TrkC arefunctional high-affinity receptors for NGF, BDNFrNT-
w x4r5 and NT-3, respectively 26 , CNTF may enhance theTrkB-mediated signaling pathway, but not those mediatedby TrkA or TrkC. Although the differences in the signaling
pathways of Trks have not yet been elucidated well, suchdifferences might explain the TrkB-specific augmentativeeffect of CNTF on P2w basal forebrain cholinergic neu-rons.
Fig. 5 shows that LIF as well as CNTF enhanced theeffect of BDNF, while IL-6 did not. The CNTF receptor isa ternary complex of one a component named CNTFR a
w xand two b components named LIFR b and gp130 20 .The IL-6 receptor consists of one a component namedIL-6R a and two gp130s. Considering the compositions ofthese receptor components, LIFR b might be involved inthe enhancement of the effect of BDNF on P2w basalforebrain cholinergic neurons. Alternatively, the heteromerof LIFR b and gp130 might transduce a unique signalwhich is responsible for the synergistic effect.
One plausible explanation for the augmentative effect ofCNTF on the action of BDNF would be the up-regulationof TrkB by CNTF. However, CNTF did not increase thenumber of TrkB-positive cells in P2w basal forebrain
Ž .culture data not shown , which suggests the other mecha-nisms than the above one may be involved. Alternatively,some intracellular molecule might be synergistically aug-mented by co-stimulation with BDNF and CNTF. How-ever, we could not elucidate such possibilities furtherbecause the percentage of cholinergic neurons in basalforebrain or striatal cultures was so small that biochemicalanalysis was not possible. Further analysis using a systemsuited for biochemical analysis will be necessary to under-stand the molecular basis of the cooperative effect ofBDNF and CNTF.
Acknowledgements
This work was supported in part by a Research Grantfor Neurons and Mental Disorders from the Ministry ofHealth and Welfare. We thank Regeneron Pharmaceuticalfor their kind gifts of human recombinant BDNF, NT-3,NT-4r5 and CNTF. We also thank Dr. H. Murofushi forhis kind gift of anti-MAP2 antibody.
References
w x1 A. Acheson, J.C. Conover, J.P. Fandl, T.M. DeChiara, M. Russell,A. Thadani, S.P. Squinto, G.D. Yancopoulos, R. M Lindsay, ABDNF autocrine loop in adult sensory neurons prevents cell death,
Ž .Nature 374 1995 450–453.w x2 C.A. Altar, N. Cai, T. Bliven, M. Juhasz, J.M. Conner, A. Acheson,
R.M. Lindsay, S.J. Wiegand, Anterograde transport of brain-derivedŽ .neurotrophic factor and its role in the brain, Nature 389 1997
856–860.w x3 Y. Arakawa, M. Sendtner, H. Thoenen, Survival effect of ciliary
Ž .neurotrophic factor CNTF on chick embryonic motoneurons inculture: comparison with other neurotrophic factors and cytokines, J.
Ž .Neurosci. 10 1990 3507–3515.w x4 J.F. Bazan, Neuropoietic cytokines in the hematopoietic fold, Neu-
Ž .ron 7 1991 197–208.

( )Y. Hashimoto et al.rDeÕelopmental Brain Research 115 1999 25–3232
w x5 V. Bocchini, P.U. Angeletti, The nerve growth factor: purification asa 30,000-molecular weight protein, Proc. Natl. Acad. Sci. USA 64Ž .1969 787–794.
w x6 R.E. Clatterbuck, D.L. Price, V.E. Koliatsos, Ciliary neurotrophicfactor prevents retrograde neuronal death in the adult central nervous
Ž .system, Proc. Natl. Acad. Sci. USA 90 1993 2222–2226.w x7 F.P. Eckenstein, F. Esch, T. Holbert, R.W. Blacher, R. Nishi,
Purification and characterization of a trophic factor for embryonicperipheral neurons: comparison with fibroblast growth factors, Neu-
Ž .ron 4 1990 623–631.w x8 P.S. DiStefano, B. Friedman, C. Radziejewski, C. Alexander, P.
Boland, C.M. Schick, R.M. Lindsay, S.J. Wiegand, The neu-rotrophins BDNF, NT-3, and NGF display distinct patterns ofretrograde axonal transport in peripheral and central neurons, Neu-
Ž .ron 8 1992 983–993.w x9 J.P. Fandl, N.J. Tobkes, N.Q. McDonald, W.A. Hendrickson, T.E.
Ryan, S. Nigam, A. Acheson, H. Cudny, N. Panayotatos, Characteri-zation and crystallization of recombinant human neurotrophin-4, J.
Ž .Biol. Chem. 269 1994 755–759.w x10 F. Fonnum, A rapid radiochemical method for the determination of
Ž .choline acetyltransferase, J. Neurochem. 24 1975 407–409.w x11 T. Hagg, D. Quon, J. Higaki, S. Varon, Ciliary neurotrophic factor
prevents neuronal degeneration and promotes low affinity NGFŽ .receptor expression in the adult rat CNS, Neuron 8 1992 145–158.
w x12 T. Hagg, S. Varon, Ciliary neurotrophic factor prevents degenerationof adult rat substantia nigra dopaminergic neurons in vivo, Proc.
Ž .Natl. Acad. Sci. USA 90 1993 6315–6319.w x13 H. Hatanaka, H. Tsukui, Differential effects of nerve-growth factor
and glioma-conditioned medium on neurons cultured from variousregions of fetal rat central nervous system, Dev. Brain Res. 30Ž .1986 47–56.
w x14 H. Hatanaka, H. Tsukui, I. Nihonmatsu, Septal cholinergic neuronsfrom postnatal rat can survive in the dissociate culture conditions in
Ž .the presence of nerve growth factor, Neurosci. Lett. 79 198785–90.
w x15 H. Hatanaka, H. Tsukui, I. Nihonmatsu, Developmental change inthe nerve growth factor action from induction of choline acetyltrans-ferase to promotion of cell survival in cultured basal forebrain
Ž .cholinergic neurons from postnatal rats, Dev. Brain Res. 39 198885–95.
w x16 H. Hatanaka, C. Nishio, Y. Kushima, H. Tsukui, Nerve-growth-fac-tor-dependent and cell-density-independent survival of septal cholin-
Ž .ergic neurons in culture from postnatal rats, Neurosci. Res. 8 199069–82.
w x17 S. Heller, J. Huber, T.P. Finn, R. Nishi, H. Rohrer, GPA and CNTFproduce similar effects in sympathetic neurons but differ in receptor
Ž .binding, NeuroReport 5 1993 357–360.w x18 C. Hyman, M. Hofer, Y.-A. Barde, M. Juhasz, G.D. Yancopoulos,
S.P. Squinto, R.M. Lindsay, BDNF is a neurotrophic factor forŽ .dopaminergic neurons of the substantia nigra, Nature 6315 1991
230–232.w x19 N.Y. Ip, S.J. Wiegand, J. Morse, J.S. Rudge, Injury-induced regula-
tion of ciliary neurotrophic factor mRNA in the adult rat brain, Eur.Ž .J. Neurosci. 5 1993 25–33.
w x20 N.Y. Ip, G.D. Yancopoulos, The neurotrophins and CNTF: twofamilies of collaborative neurotrophic factors, Annu. Rev. Neurosci.
Ž .19 1996 491–515.w x21 S. Korsching, The neurotrophic factor concept; a reexamination, J.
Ž .Neurosci. 13 1993 2739–2748.w x22 T. Kubo, T. Nonomura, Y. Enokido, H. Hatanaka, Brain-derived
Ž .neurotrophic factor BDNF can prevent apoptosis of rat cerebellarŽ .granule neurons in culture, Dev. Brain Res. 85 1995 249–258.
w x23 Y. Kushima, C. Nishio, T. Nonomura, H. Hatanaka, Effects of nervegrowth factor and basic fibroblast growth factor on survival ofcultured septal cholinergic neurons from adult rats, Brain Res. 598Ž .1992 264–270.
w x24 M.Y. Lee, T. Naumann, M. Kirsch, M. Frotscher, H.D. Hofmann,Transient up-regulation of ciliary neurotrophic factor receptor-alpha
Ž .mRNA in axotomized rat septal neurons, Eur. J. Neurosci. 9 1997622–626.
w x25 W.D. Leung, A.S. Parent, G. Cachianes, F. Esch, J.N. Coulombe, K.Nikolics, F.P. Eckenstein, R. Nishi, Cloning, expression duringdevelopment, and evidence for release of a trophic factor for ciliary
Ž .ganglion neurons, Neuron 8 1992 1045–1053.w x26 R.M. Lindsay, S.J. Wiegand, C.A. Altar, P.S. DiStefano, Neu-
rotrophic factors: from molecule to man, Trends. Neurosci. 17Ž .1994 182–190.
w x27 M. Mata, C.F. Jin, D.J. Fink, Axotomy increases CNTF receptorŽ .mRNA in rat spinal cord, Brain Res. 610 1993 162–165.
w x28 H. Mitsumoto, K. Ikeda, B. Klinkosz, J.M. Cedarbaum, V. Wong,R.M. Lindsay, Arrest of motor neuron disease in wobbler mice
Ž .cotreated with CNTF and BDNF, Science 265 1994 1107–1110.w x29 M. Nakamura, C. Nishio, T. Nonomura, H. Hatanaka, High potas-
sium and cyclic AMP analog promote neuronal survival of basalforebrain cholinergic neurons in culture from postnatal 2-week-old
Ž .rats, Dev. Brain Res. 81 1994 218–229.w x30 T. Nonomura, C. Nishio, R.M. Lindsay, H. Hatanaka, Cultured basal
forebrain cholinergic neurons from postnatal rats show both overlap-ping and non-overlapping responses to the neurotrophins, Brain Res.
Ž .683 1995 129–139.w x31 M. Sendtner, K.A. Stockli, H. Thoenen, Synthesis and localization
of ciliary neurotrophic factor in the sciatic nerve of the adult rat afterŽ .lesion and during regeneration, J. Cell Biol. 118 1992 139–148.
w x32 K.A. Stockli, F. Lottspeich, M. Sendtner, P. Masiakowski, P. Car-roll, R. Gotz, D. Lindholm, H. Thoenen, Molecular cloning, expres-sion and regional distribution of rat ciliary neurotrophic factor,
Ž .Nature 342 1989 920–923.w x33 K.A. Stockli, L.E. Lillien, N.M. Naher, G. Breitfeld, R.A. Hughes,
M.C. Raff, H. Thoenen, M. Sendtner, Regional distribution, devel-opmental changes, and cellular localization of CNTF-mRNA and
Ž .protein in the rat brain, J. Cell Biol. 115 1991 447–459.w x34 K. Suda, Y.-A. Barde, H. Thoenen, Nerve growth factor in mouse
and rat serum: correlation between bioassay and radioimmunoassayŽ .determinations, Proc. Natl. Acad. Sci. USA 75 1978 4042–4046.
w x35 V. Wong, R. Arriaga, N.Y. Ip, R.M. Lindsay, The neurotrophinsBDNF, NT-3 and NT-4r5, but not NGF, up-regulate the cholinergic
Ž .phenotype of developing motor neurons, Eur. J. Neurosci. 5 1993466–474.