synaptic targets of pyramidal neurons providing intrinsic horizontal connections in monkey...
TRANSCRIPT

Synaptic Targets of Pyramidal NeuronsProviding Intrinsic Horizontal
Connections in MonkeyPrefrontal Cortex
DARLENE S. MELCHITZKY,1 SUSAN R. SESACK,1,2 MICHELE L. PUCAK,1
AND DAVID A. LEWIS1,2*1Department of Psychiatry, University of Pittsburgh, Pittsburgh, Pennsylvania 15213
2Department of Neuroscience, University of Pittsburgh, Pittsburgh, Pennsylvania 15260
ABSTRACTIn monkey prefrontal cortex, the intrinsic axon collaterals of supragranular pyramidal
neurons extend horizontally for considerable distances through the gray matter and give riseto stripe-like clusters of axon terminals (Levitt et al. [1993] J. Comp. Neurol. 338:360–376).Because understanding the functional role of these connections requires knowledge of theirsynaptic targets, we made injections of biotinylated dextran amine (BDA) into layer 3 ofmacaque prefrontal area 9 and examined the labeled intrinsic axon collaterals by electronmicroscopy. Labeled axon terminals formed exclusively asymmetric synapses, and 95.6% ofthe postsynaptic structures were dendritic spines, presumably belonging to other pyramidalneurons. The remaining postsynaptic structures were dendritic shafts, many of which had themorphological characteristics of local circuit neurons. The prefrontal injections also labeledassociational projections that traveled through the white matter to terminate in other areas ofprefrontal cortex. All of the synapses formed by these associational axons were asymmetric,and 91.9% were onto dendritic spines. The similarities in synaptic targets of the prefrontalintrinsic and associational axon terminals suggested that these projections might arise fromthe same neurons, an interpretation confirmed in dual label, retrograde tracing studies.
To determine the specificity of the synaptic targets of these prefrontal connections, twoadditional comparisons were made. In the posterior parietal cortex (area 7a), 94.2% of thesynapses furnished by BDA-labeled intrinsic collaterals of supragranular pyramidal neuronswere also with dendritic spines. In contrast, only 75.6% of unlabeled asymmetric synapses inthe prefrontal cortex were onto dendritic spines. These comparisons suggest that the axons ofsupragranular pyramidal neurons in primate association cortices are preferentially directedto specific targets.
Finally, after injections of BDA, a small number of retrogradely labeled pyramidalneurons were observed within the anterogradely labeled clusters of intrinsic axon terminals.At the ultrastructural level, synapses between anterogradely labeled axon terminals andretrogradely labeled dendritic spines were identified. These findings suggest that reciprocal,monosynaptic connections may exist between pyramidal neurons located in different stripe-like clusters, providing a potential anatomical substrate for reverberating excitatory circuitswithin the primate association cortices. J. Comp. Neurol. 390:211–224, 1998. r 1998 Wiley-Liss, Inc.
Indexing terms: axon collateral; asymmetric synapse; posterior parietal cortex; local circuit neuron
In the primate brain, the prefrontal cortex (PFC) sub-serves a number of higher-order cognitive functions, includ-ing those involving working memory (Goldman-Rakic,1987; Baddeley, 1992) and the temporal integration ofinformation (Fuster, 1985). Disturbances of these abilitiesin disease states, such as schizophrenia and Alzheimer’s
Grant sponsor: USPHS; Grant numbers:AG05133, MH51234, and MH45156;Grant sponsor: NIMH Independent Scientist Award; Grant number: MH00519.
*Correspondence to: David A. Lewis, Western Psychiatric Institute andClinic, University of Pittsburgh, Biomedical Science Tower, W1651, 3811O’Hara Street, Pittsburgh, PA 15213.E-mail: [email protected]
Received 13 May 1997; Revised 28 July 1997; Accepted 8 August 1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 390:211–224 (1998)
r 1998 WILEY-LISS, INC.

disease, have been associated with abnormalities in pyra-midal neurons, especially those located in supragranularlayers 2 and 3 (Hof et al., 1990; Glantz and Lewis, 1995).The involvement of these excitatory neurons in the patho-physiology of cognitive disorders may be related, at least inpart, to the fact that the majority of their extrinsic axonalprojections are directed to other cortical association re-gions (Schwartz and Goldman-Rakic, 1984; Barbas andPandya, 1989). In addition, the intrinsic axon collaterals ofsupragranular pyramidal neurons in monkey PFC spreadhorizontally for considerable distances and give rise todiscrete, stripe-like clusters of axon terminals that spanlayers 1–3 (Levitt et al., 1993). Retrograde labeling studies(Kritzer and Goldman-Rakic, 1995; Pucak et al., 1996)have demonstrated that the neurons that contribute axoncollaterals to this intrinsic circuitry are also arranged in astripe-like fashion. These clusters of retrogradely labeledneurons are coregistered with anterogradely labeled axonterminals arising from the same injection site (Pucak etal., 1996), suggesting that reciprocal intrinsic connectionslink spatially segregated clusters of supragranular pyrami-dal cells into neuronal networks (Goldman-Rakic, 1995;Lewis and Anderson, 1995).
These networks of supragranular pyramidal neurons inmonkey PFC have been suggested to provide a means forcoordinating the activity of neuronal populations thatshare the same response properties (Levitt et al., 1993;Goldman-Rakic, 1995), analogous to the apparent role ofthe intrinsic horizontal connections in sensory cortices(Lund et al., 1993). For example, in the primary visualcortex, horizontally oriented axon collaterals predomi-nantly interconnect orientation domains of similar prefer-ences, as well as zones of similar ocular dominance (T’so etal., 1986; Gilbert and Wiesel, 1989; Malach et al., 1993;Yoshioka et al., 1996). In addition, the sustained activity ofspecific populations of prefrontal pyramidal neurons dur-ing the delay period of delayed response tasks (Fuster etal., 1982; Funahashi et al., 1989) may depend, at least inpart, on reciprocal intrinsic connections (Lewis and Ander-son, 1995; Pucak et al., 1996). That is, these connectionshave been suggested to provide the anatomical substratefor a reverberating excitatory circuit that maintains thefiring of prefrontal neurons in the absence of externalstimulation, a critical feature of working memory (Funaha-shi et al., 1989; Goldman-Rakic, 1995). However, furthertesting of this hypothesis requires knowledge of the synap-tic targets of these intrinsic axon collaterals.
In other species, the intrinsic axon collaterals of differ-ent subpopulations of pyramidal neurons preferentiallyform synaptic contacts with certain types of structures(Czeiger and White, 1993). For example, in mouse somato-sensory cortex, over 80% of the intrinsic axon collateralsarising from callosally projecting neurons target dendriticspines (Elhanany and White, 1990), whereas dendriticshafts are the postsynaptic targets of over 90% of theintrinsic axon collaterals of corticothalamic neurons (Whiteand Keller, 1987). However, the postsynaptic targets of theintrinsic axon collaterals of supragranular pyramidal neu-rons in primate PFC are not known. Consequently, in thisstudy, we placed injections of the tracer biotinylateddextran amine (BDA) into the superficial layers of monkeyPFC to identify the structures that receive synaptic inputfrom the intrinsic axon collaterals of supragranular pyra-midal neurons. In addition, the relative specificity of thesetargets was assessed through comparisons with the neural
elements that receive synaptic input from other types ofcortical excitatory terminals, both within the PFC and inanother higher-order association region, the posterior pari-etal cortex.
MATERIALS AND METHODS
Injections of biotinylated dextran amine
Surgical procedures. Four male, adult cynomolgusmonkeys (Macaca fascicularis) were used in this study. Allanimals were treated according to the guidelines outlinedin the National Institutes of Health Guide for the Care andUse of Laboratory Animals. After injections of ketaminehydrochloride (25 mg/kg), dexamethasone phosphate (0.5mg/kg), and atropine sulfate (0.05 mg/kg), an endotrachealtube was inserted, and the animal was placed in a stereo-taxic apparatus. Anesthesia was maintained with 1%halothane in 28% O2/air. Guided by stereotaxic coordinates(Szabo and Cowan, 1984), a craniectomy was performedover either the dorsal prefrontal or the posterior parietalcortex. By using a 5.0-µl Hamilton syringe (26-gaugeneedle), one injection (0.3 µl) of 10% BDA (10,000 molecu-lar weight; Molecular Probes, Inc., Eugene, OR) in 0.01 Mphosphate buffer, pH 7.3, was placed into either dorsalarea 9 or area 7a (Fig. 1A). For each animal, one injectionwas made at a depth of approximately 1.0-mm below thepial surface. After tracer injections, the scalp was closed,and the animals were treated with an antibiotic (chloram-phenicol, 15 mg/kg) and an analgesic (hydromorphone,0.02 mg/kg).
Tissue preparation. After a survival time of 8 to12days, monkeys were deeply anesthetized with ketaminehydrochloride (25 mg/kg) and pentobarbital sodium (30mg/kg) and then perfused transcardially with cold 4%paraformaldehyde in phosphate buffer (Pucak et al., 1996).The brain was removed, and coronal blocks (4-mm-thick)were immersed in 0.12 M phosphate buffer, pH 7.3,containing 4% paraformaldehyde and 0.2% glutaralde-hyde, for 2 hours. Tissue blocks were then washed in 0.1 Mphosphate buffer, pH 7.3, and sectioned on a Vibratome at50 µm.
Histochemistry. The histochemical procedures usedto visualize BDA were adapted from those previouslydescribed (Pucak et al., 1996; Woo et al., 1997). Briefly,tissue sections were incubated in 0.05 M phosphate-buffered saline (PBS) containing 4.5% normal humanserum (NHuS), 0.04% Triton X-100, and 0.05 mg/ml bovineserum albumin (BSA) and then placed in 0.05 M PBS, pH7.3, containing avidin-biotin complex (ABC) reagents (Vec-tastain Elite, Vector Laboratories, Burlingame, CA) and4.5% NHuS for approximately 16 hours at 4°C. Sectionswere then rinsed and incubated in 0.1 M phosphate buffer,pH 7.3, containing 0.5 mg/ml diaminobenzidine and 0.006%H2O2 for 5 minutes. After a rinse in 0.1 M phosphate buffer,pH 7.3, sections for electron microscopy were post-fixed in2% osmium tetroxide in 0.1 M phosphate buffer pH 7.3 for1 hour, dehydrated, and embedded in Epon 812. Ultrathinsections were collected, counterstained with uranyl ac-etate and lead citrate, and examined on a Zeiss or JEOLelectron microscope. Sections for light microscopy weremounted onto gel-coated slides and intensified with silveras previously described (Pucak et al., 1996).
212 D.S. MELCHITZKY ET AL.
Definition of neuronal and synaptic elements
Neuronal elements of relevance to this study wereidentified by using the criteria of Peters et al. (1991). Axonterminals were generally greater than 0.2 µm in diameterand contained synaptic vesicles and, occasionally, mitochon-dria. Dendritic shafts were characterized by the presenceof mitochondria, numerous microtubules, and neurofila-ments, as well as postsynaptic specializations. Dendriticspines were identified by the absence of mitochondria andmicrotubules and by the presence of a spine apparatus (inoptimal planes of section). In addition, dendritic spinesinvariably received asymmetric synapses, which werecharacterized by widened and parallel spacing of apposedplasmalemmal surfaces, and a thick postsynaptic density.The axon terminals forming these synapses always con-tained round synaptic vesicles.
Sampling regions and procedures
As previously described (Pucak et al., 1996), BDA-labeled axon terminals within the PFC were organizedinto distinct clusters that could be identified as originatingfrom intrinsic or associational axon projections. Intrinsicclusters were composed of arborizing axons and terminalsconfined to layers 1–3, were associated with labeled preter-minal horizontally oriented axons in the adjacent graymatter, and lacked labeled axons in the underlying whitematter (Figs. 1B, 2A). In contrast, associational clustershad arborizing axons and terminals in both the superficialand deep cortical layers (although the density of labeledaxons in different layers varied across regions), lacked
labeled preterminal axons in the gray matter and ap-peared to arise from labeled axons in the underlying whitematter (Figs. 1B, 2B). Finally, over 95% of the axonclusters with the characteristics of intrinsic projectionswere located within 6 mm of the injection site, whereas90% of the associational clusters were located more than 8mm from the injection site (Pucak et al., 1996).
Three types of labeled axon terminals were examined inthis study: intrinsic prefrontal, associational prefrontaland intrinsic parietal. For each type of terminal, blocks forelectron microscopic analysis were taken from two to fivedifferent clusters of labeled axon terminals per animal. Allintrinsic clusters selected for study were located 1 to 5 mmfrom the injection site. Within each cluster, blocks werecentered in layers 2 and 3. For each block, three ultrathinsections, separated by 10 to 20 sections, were examined.The analysis consisted of identifying all profiles of antero-gradely labeled axon terminals within the entire ultrathinsection, and determining which terminals had synapticspecializations. For those terminals forming synaptic spe-cializations, the postsynaptic structures were recorded.
As an additional check on the relative specificity of thesynaptic targets of the prefrontal axons, unlabeled axonterminals from the general prefrontal neuropil were alsoanalyzed. Random fields from the sections used for sam-pling the intrinsic and the associational prefrontal connec-tions were photographed at 319,000. The total number ofunlabeled asymmetric synaptic profiles in each field werecounted, and the structures postsynaptic to them wererecorded.
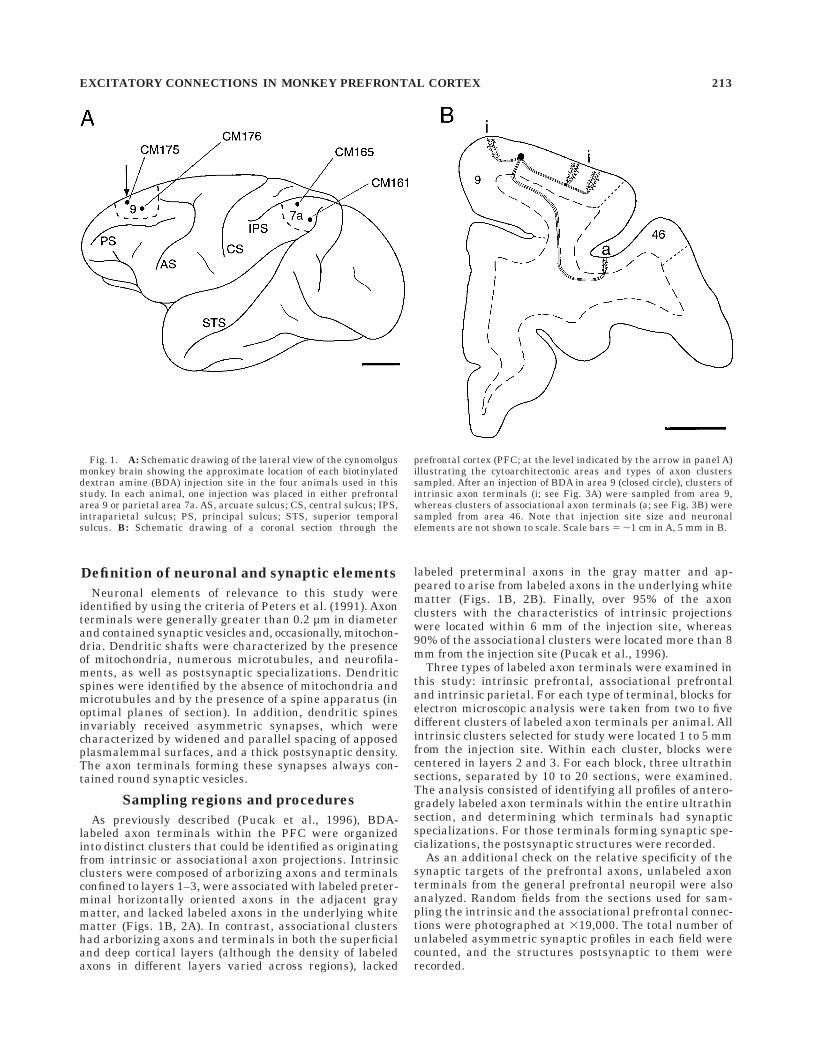
Fig. 1. A: Schematic drawing of the lateral view of the cynomolgusmonkey brain showing the approximate location of each biotinylateddextran amine (BDA) injection site in the four animals used in thisstudy. In each animal, one injection was placed in either prefrontalarea 9 or parietal area 7a. AS, arcuate sulcus; CS, central sulcus; IPS,intraparietal sulcus; PS, principal sulcus; STS, superior temporalsulcus. B: Schematic drawing of a coronal section through the
prefrontal cortex (PFC; at the level indicated by the arrow in panel A)illustrating the cytoarchitectonic areas and types of axon clusterssampled. After an injection of BDA in area 9 (closed circle), clusters ofintrinsic axon terminals (i; see Fig. 3A) were sampled from area 9,whereas clusters of associational axon terminals (a; see Fig. 3B) weresampled from area 46. Note that injection site size and neuronalelements are not shown to scale. Scale bars 5 ,1 cm in A, 5 mm in B.
EXCITATORY CONNECTIONS IN MONKEY PREFRONTAL CORTEX 213
Individual 2 3 2 x2 analyses were performed to comparethe synaptic targets of the intrinsic prefrontal axon termi-nals to those of the associational prefrontal and theintrinsic parietal axon terminals, as well as to comparesynaptic targets of the intrinsic prefrontal and associa-tional prefrontal axon terminals with those of the unla-beled terminals in the prefrontal neuropil that formedasymmetric synapses.
Injections of Fast Blue and cholera toxin B
Surgical procedures. To determine whether the intrin-sic and associational axon terminals in the PFC arise fromthe same population of pyramidal neurons, two additionaladult male cynomolgus monkeys were anesthetized and acraniectomy was performed over the dorsal PFC as de-scribed above. By using a 5.0-µl Hamilton syringe, aninjection (0.3 µl) of 1% cholera toxin (CtB; List Biologicals,West Campbell, CA) in distilled water was placed into theapproximate location of rostral area 9 at a depth of 0.8 mm(Fig. 3A). In addition, as part of other studies (Soloway etal., 1994; Pucak et al., 1996), 9 to 12 injections (0.3 µl) of5% Fast Blue (FB; Sigma Chemical Co., St. Louis, MO) indistilled water were placed caudal to the CtB injection intoipsilateral area 8b (Fig. 3A) as previously described (Oethand Lewis, 1992). After a survival time of 12 to 16 days, theanimals were perfused as described above. The brain wasremoved and coronal blocks containing the CtB or FBinjection sites were immersed in 0.12 M phosphate buffer,
pH 7.3, containing 4% paraformaldehyde for 6 hours.Tissue blocks were washed in a series of cold, gradedsucrose solutions, and coronal cryostat sections (50 µm)were cut and maintained in serial order.
CtB immunocytochemistry. Tissue sections were in-cubated in 0.01 M PBS, pH 7.3, containing 4.5% NHuS,4.5% normal donkey serum (NDS), and 0.3% Triton X-100for 30 minutes. Sections were then incubated in 0.01 MPBS containing 3.0% NHuS, 3.0% NDS, 0.5 mg/ml BSA,0.3% Triton X-100, and a 1:15,000 dilution of a polyclonalgoat anti-CtB antibody (List Biologicals) for 48 hours at4°C. Next, sections were incubated for 1 hour in PBScontaining a biotinylated donkey anti-goat antibody (1:200dilution, Jackson ImmunoResearch, West Grove, PA) with3.0% NHuS, 3.0% NDS, and 0.3% Triton X-100 in PBS.They were then incubated in a solution containing 0.3%Triton X-100 and streptavidin-Texas Red (1:200 dilution,Amersham, Arlington Heights, IL) in PBS for 4 hours.
Sections prepared for CtB immunocytochemistry wereexamined under a Zeiss Axiophot microscope (Zeiss,Oberkochen, Germany) equipped with fluorescent illumi-nation and separate filter sets to visualize Texas Red andFB. As previously described (Pucak et al., 1996), clusters ofCtB-positive neurons were identified as either intrinsic orassociational based on their distance from the CtB injec-tion site (Fig. 3B). For the clusters of CtB-positive neuronsthat also contained FB-positive neurons, the number of
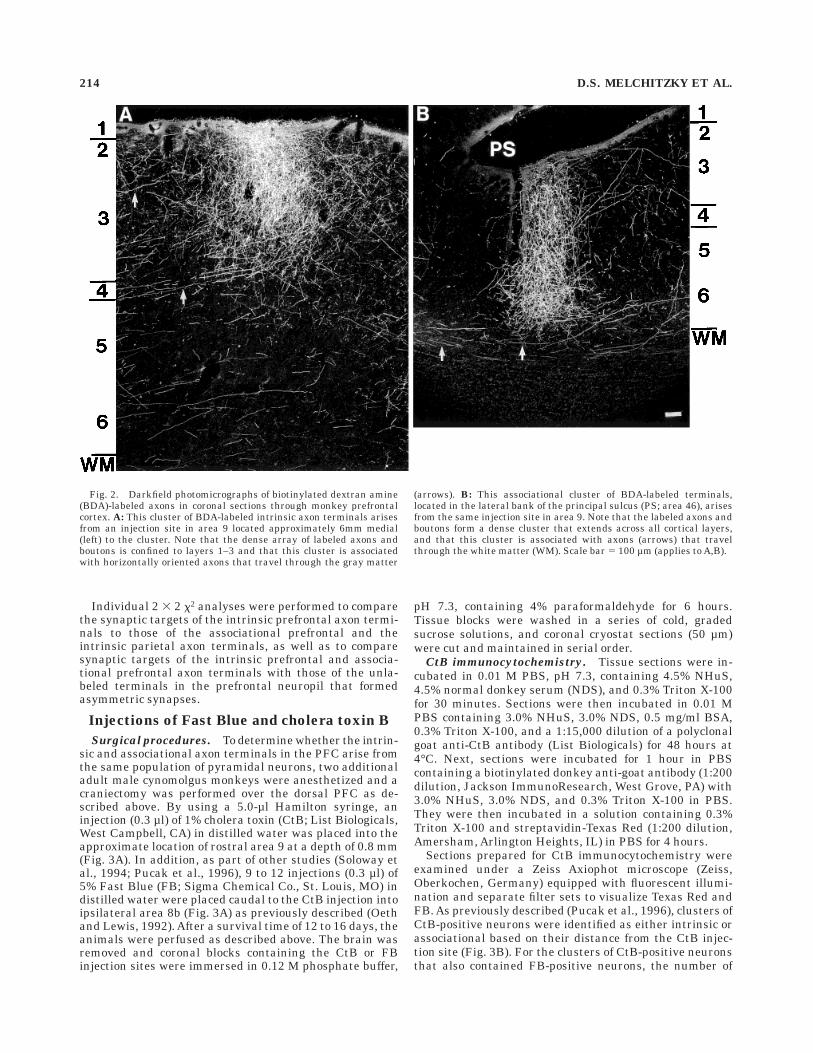
Fig. 2. Darkfield photomicrographs of biotinylated dextran amine(BDA)-labeled axons in coronal sections through monkey prefrontalcortex. A: This cluster of BDA-labeled intrinsic axon terminals arisesfrom an injection site in area 9 located approximately 6mm medial(left) to the cluster. Note that the dense array of labeled axons andboutons is confined to layers 1–3 and that this cluster is associatedwith horizontally oriented axons that travel through the gray matter
(arrows). B: This associational cluster of BDA-labeled terminals,located in the lateral bank of the principal sulcus (PS; area 46), arisesfrom the same injection site in area 9. Note that the labeled axons andboutons form a dense cluster that extends across all cortical layers,and that this cluster is associated with axons (arrows) that travelthrough the white matter (WM). Scale bar 5 100 µm (applies to A,B).
214 D.S. MELCHITZKY ET AL.
FB-positive and double-labeled (FB- and CtB-positive)neurons were counted in layers 2–3.
RESULTS
Distribution of BDA labeling
Comparison of Nissl-stained sections adjacent to thoseprocessed for BDA confirmed that the prefrontal injectionsites were located in dorsal area 9 (Walker, 1940) and thatthe parietal injections were located in area 7a (Von Boninand Bailey, 1947; Neal et al., 1987) (Fig. 1A). In eachanimal, the BDA injection site was marked by a dense coreof intense reaction product (Pucak et al., 1996; Woo et al.,1997) that was centered in layer 3. Surrounding the core ofthe injection was a zone containing labeled axon terminalsand neurons that appeared to have been labeled by theextension of their dendritic tree into the dense core.Horizontally oriented axons spread from this zone andtraversed the supragranular cortical layers, whereas verti-cally oriented axons descended through the infragranularcortical layers and entered the white matter. In both theprefrontal and parietal cortices, BDA-labeled axons gaverise to a number of discrete, stripe-like clusters of arboriz-ing axons and axon terminals (Fig. 2A,B), separated byzones in which labeled axons were much less numerous,exhibited little branching, and had few varicosities. Theaxons within the stripes were highly branched (Fig. 4A)and displayed numerous boutons en passant as well asboutons terminaux (Fig. 4B). As described in the Materialsand Methods section and in previous publications (Pucaket al., 1996; Woo et al., 1997), these clusters of axonterminals could be identified as either intrinsic or associa-
tional (see Figs. 1B, 2A,B). Associational clusters werestudied only in the PFC.
Synaptic targets of BDA-labeled axons
At the ultrastructural level, BDA labeling was seen inaxon terminals and preterminal axons, and occasionally,in myelinated axons in both the prefrontal and parietalcortices. All the BDA-labeled axon terminals located ineither intrinsic or associational clusters contained roundsynaptic vesicles and formed exclusively asymmetric syn-apses (Figs. 5A–D, 6A–C), indicating that they arose frompyramidal cells and not local circuit neurons within theinjection site. As shown in Table 1, approximately one-third of all the labeled terminals examined formed syn-apses in the single plane of section analyzed.
Dendritic spines were the most common postsynaptictargets of the intrinsic axon terminals in the PFC (Fig.5A). Similarly, the labeled terminals that formed prefron-tal associational clusters (Fig. 5B,D), as well as those thatformed intrinsic clusters in the parietal cortex (Fig. 5C),also formed asymmetric synapses predominantly withspines. As shown in Table 1, no differences were present inthe proportion of the intrinsic prefrontal (95.6%), theassociational prefrontal (91.9%), or the intrinsic parietal(94.2%) axon terminals that formed synapses with den-dritic spines. Because most dendritic spines in the neocor-tex are located on pyramidal neurons (Feldman, 1984;DeFelipe and Farinas, 1992), these findings indicate thatother pyramidal neurons are the major recipients of theoutput of supragranular pyramidal neurons in these re-gions.
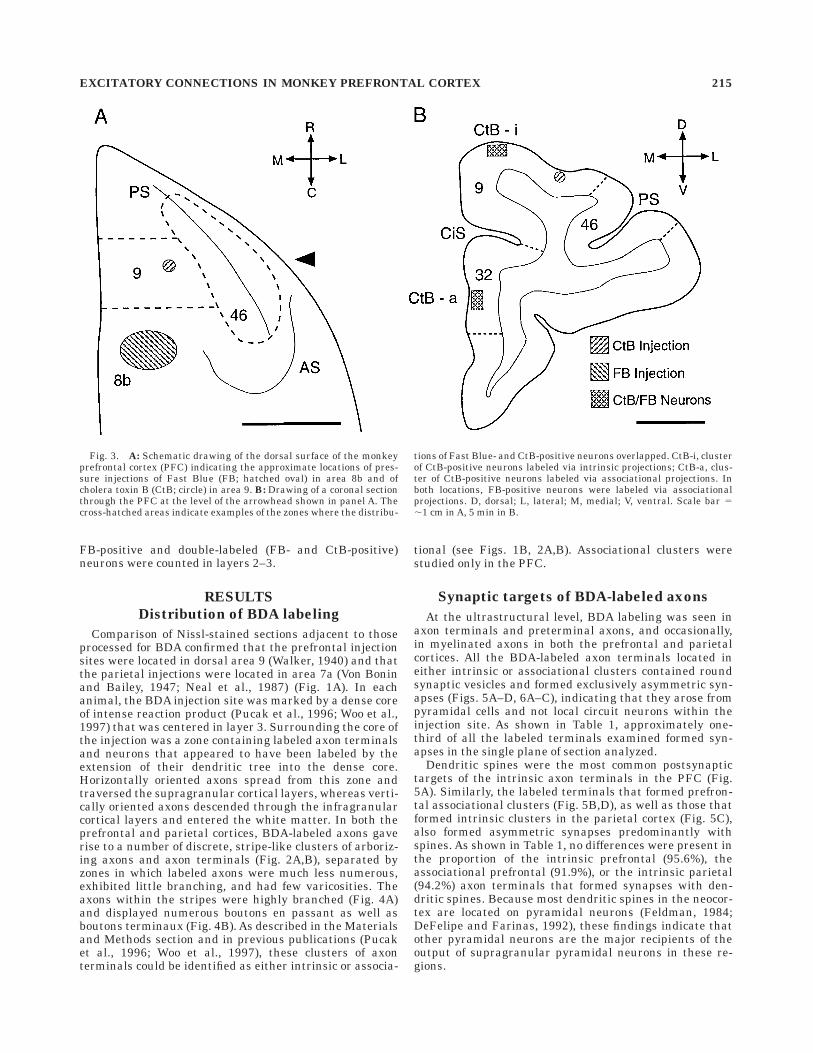
Fig. 3. A: Schematic drawing of the dorsal surface of the monkeyprefrontal cortex (PFC) indicating the approximate locations of pres-sure injections of Fast Blue (FB; hatched oval) in area 8b and ofcholera toxin B (CtB; circle) in area 9. B: Drawing of a coronal sectionthrough the PFC at the level of the arrowhead shown in panel A. Thecross-hatched areas indicate examples of the zones where the distribu-
tions of Fast Blue- and CtB-positive neurons overlapped. CtB-i, clusterof CtB-positive neurons labeled via intrinsic projections; CtB-a, clus-ter of CtB-positive neurons labeled via associational projections. Inboth locations, FB-positive neurons were labeled via associationalprojections. D, dorsal; L, lateral; M, medial; V, ventral. Scale bar 5,1 cm in A, 5 min in B.
EXCITATORY CONNECTIONS IN MONKEY PREFRONTAL CORTEX 215

In the PFC, the remainder of the intrinsic and associa-tional axon terminals formed asymmetric synapses withdendritic shafts (Table 1). Many of these dendritic shaftshad the distinctive morphological features of local circuitneurons (Smiley and Goldman-Rakic, 1993; Sesack et al.,1995). Specifically, these dendritic shafts had a varicosemorphology (Fig. 6A,C) and received a high density ofasymmetric synapses from unlabeled axon terminals(Fig. 6B,C). In the parietal cortex, BDA-labeled axonterminals also formed asymmetric axodendritic as wellas axosomatic synapses. The somata contacted by thelabeled intrinsic axon terminals had the morphologicalcharacteristics of local circuit neurons such as sparsecytoplasm, abundant organelles (especially mitochondria)and numerous asymmetric synaptic contacts (Peters et al.,1991).
The similarities in the targets of the labeled intrinsicprefrontal, intrinsic parietal, and associational prefrontalterminals raised the possibility that the predominance ofaxospinous synapses formed by these terminals reflected ageneral characteristic of the cortical neuropil in primates.To address this question, the types of structures receivingasymmetric synapses from unlabeled terminals in thesupragranular layers of the PFC were counted in the samesections used for analyzing BDA-labeled terminals. Asshown in Table 1, 75.6% of the asymmetric synapses in thegeneral prefrontal neuropil were onto dendritic spines,whereas 24.4% were onto dendritic shafts. Individual x2
analyses revealed that the proportions of intrinsic orassociational prefrontal axon terminals that targeted den-dritic spines differed significantly (x2 5 24.5, P , 0.0001,and x2 5 9.2, P , 0.003, respectively) from unlabeledterminals in the prefrontal neuropil.
Comparison of neurons furnishing intrinsicand associational projections
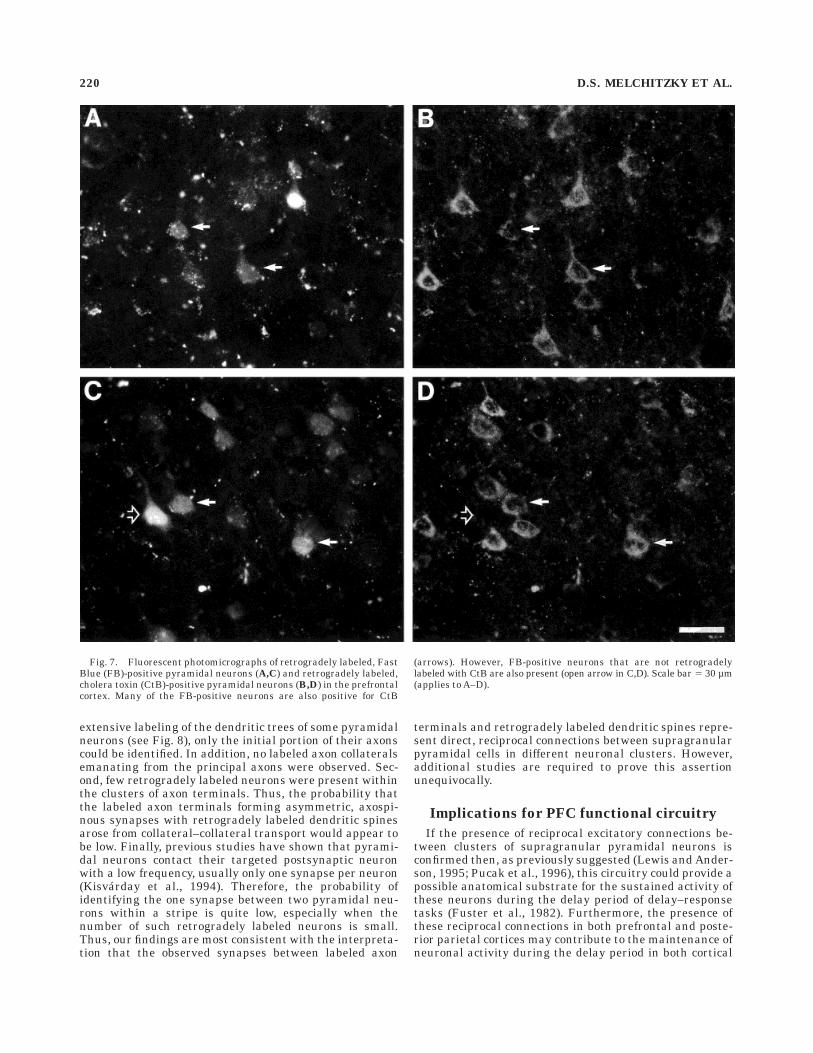
The marked similarities in, and apparent selectivity of,the postsynaptic targets of the prefrontal intrinsic andassociational axons labeled from the same injection sitesuggested that these two projections might arise, at leastin part, from the same subpopulation of prefrontal pyrami-dal neurons. To address this question, injections of CtBand FB were made into different regions of the PFC in twoanimals (Fig. 2A). In both animals, examination of Nissl-stained sections revealed that the CtB injection waslocated in rostral area 9 and that the FB injection waslocated in area 8b. As shown in Figure 2B, the distributionof CtB- and FB-labeled neurons overlapped in some por-tions of the PFC. Based on their cytoarchitectonic locationand distance from the injection sites, two types of clusterscontaining both CtB- and FB-labeled neurons were identi-fied. CtB-positive neurons were identified as retrogradelylabeled via either intrinsic or associational projections aspreviously described (Pucak et al., 1996), whereas for bothof these types of clusters, the FB-positive neurons werelabeled via associational projections. Within the intrinsicclusters of CtB-positive cells, 48% (122/256) and 39%(111/284) of the supragranular FB-labeled neurons in thetwo animals, respectively, also contained CtB (Fig. 7A,B).These findings indicate that many of the pyramidal neu-rons in layers 2 and 3 that send principal axon projectionsthrough the white matter to another prefrontal region alsocontribute axon collaterals to intrinsic stripes. In contrast,less than 5% (7/151) of the FB-positive neurons in theassociational clusters of CtB-labeled neurons were doublelabeled, consistent with previous reports that the principalaxon projections of pyramidal neurons in primate associa-
Fig. 4. Brightfield photomicrographs of biotinylated dextran amine(BDA)-labeled axons and varicosities in monkey prefrontal cortex. A:Arborizing BDA-labeled axon collaterals from the intrinsic stripe
shown in Figure 3A. B: Higher power photomontage of a BDA-labeledaxon. Note the intensely labeled en passant boutons (arrow) andboutons terminaux (arrowhead). Scale bars 5 20 µm in A, 10 µm in B.
216 D.S. MELCHITZKY ET AL.
tion cortices exhibit limited collateralization (Schwartzand Goldman-Rakic, 1984; Andersen et al., 1985; Cipolloniand Pandya, 1989; Morecraft and Van Hoesen, 1993).
Pyramidal cells that receive inputsfrom intrinsic axon collaterals
The presence of reciprocal intrinsic connections amongstripe-like clusters of supragranular prefrontal neurons(Pucak et al., 1996) and the tendency of the intrinsiccollaterals of these neurons to synapse on dendritic spinesraised the question of whether direct synaptic connectionswere present between pyramidal cells located in different,intrinsic stripes. In the present study, injections of BDA inboth the prefrontal and parietal cortices resulted in retro-grade labeling of a small number of pyramidal neurons,although the dendritic labeling of these neurons wastypically quite extensive. Both the apical and basilar
dendritic trees were labeled with BDA (Fig. 8A), anddendritic spines were clearly visible (Fig. 8B). At theultrastructural level, retrogradely labeled dendrites andspines were easily identified by the presence of peroxidasereaction product, although the retrograde labeling wasqualitatively not as dense as the anterograde labeling(compare Figs. 5, 9). Retrogradely labeled spines were alsodistinguished by the presence of a synapse, most often anasymmetric synapse from an unlabeled axon terminal(Fig. 9A,B).
Within the intrinsic clusters of axon terminals in boththe prefrontal and parietal cortices, several examples ofretrogradely labeled dendritic spines that formed asymmet-ric synapses with BDA-labeled axon terminals were found(Fig. 9C,D). These findings suggest that supragranularpyramidal neurons in one intrinsic cluster provide synap-tic input to a pyramidal neuron in another cluster,
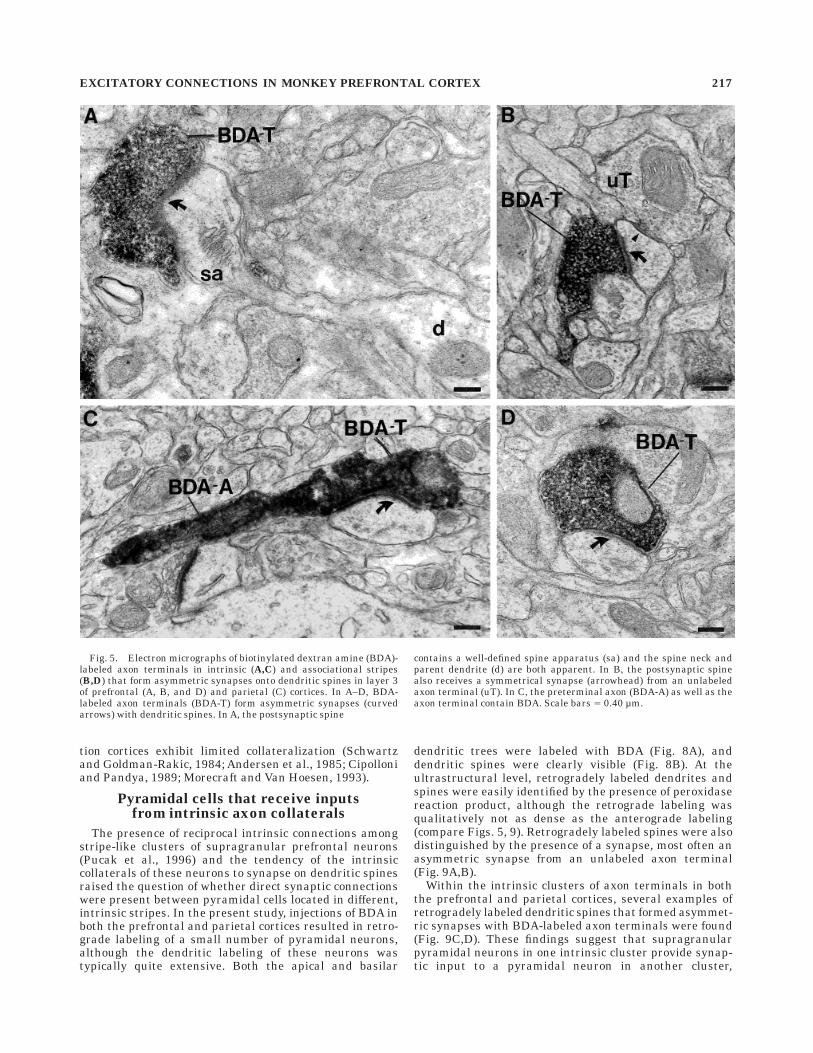
Fig. 5. Electron micrographs of biotinylated dextran amine (BDA)-labeled axon terminals in intrinsic (A,C) and associational stripes(B,D) that form asymmetric synapses onto dendritic spines in layer 3of prefrontal (A, B, and D) and parietal (C) cortices. In A–D, BDA-labeled axon terminals (BDA-T) form asymmetric synapses (curvedarrows) with dendritic spines. In A, the postsynaptic spine
contains a well-defined spine apparatus (sa) and the spine neck andparent dendrite (d) are both apparent. In B, the postsynaptic spinealso receives a symmetrical synapse (arrowhead) from an unlabeledaxon terminal (uT). In C, the preterminal axon (BDA-A) as well as theaxon terminal contain BDA. Scale bars 5 0.40 µm.
EXCITATORY CONNECTIONS IN MONKEY PREFRONTAL CORTEX 217
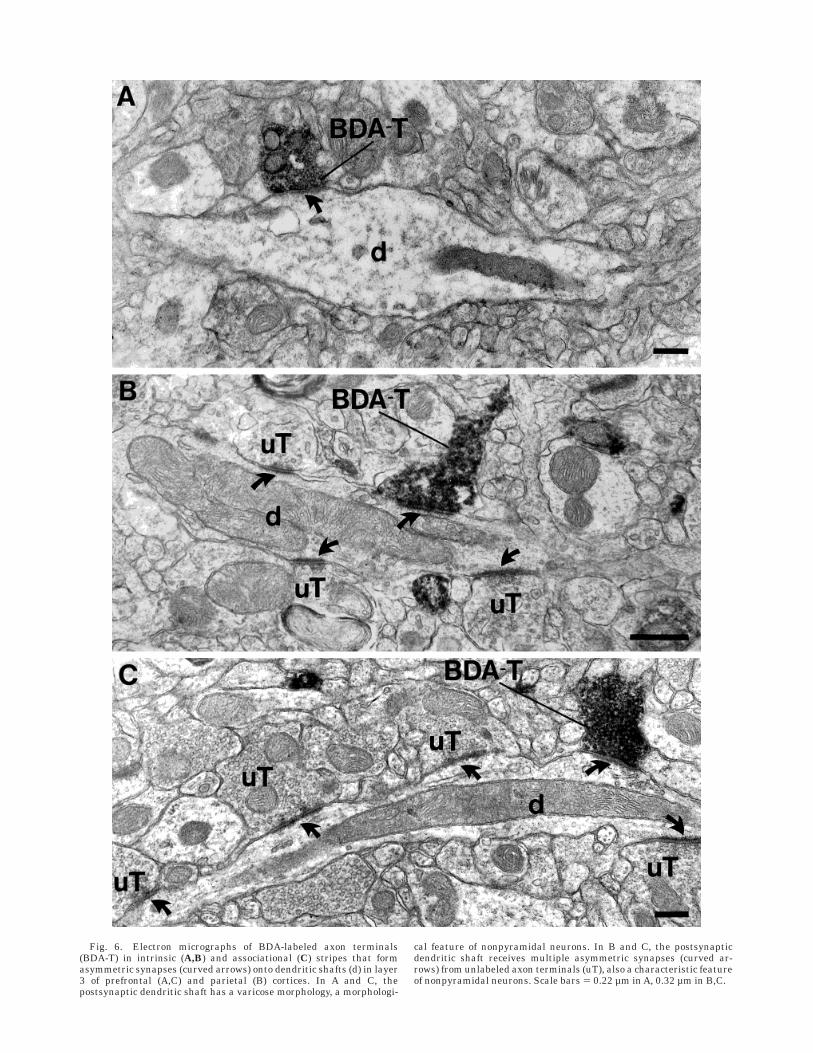
Fig. 6. Electron micrographs of BDA-labeled axon terminals(BDA-T) in intrinsic (A,B) and associational (C) stripes that formasymmetric synapses (curved arrows) onto dendritic shafts (d) in layer3 of prefrontal (A,C) and parietal (B) cortices. In A and C, thepostsynaptic dendritic shaft has a varicose morphology, a morphologi-
cal feature of nonpyramidal neurons. In B and C, the postsynapticdendritic shaft receives multiple asymmetric synapses (curved ar-rows) from unlabeled axon terminals (uT), also a characteristic featureof nonpyramidal neurons. Scale bars 5 0.22 µm in A, 0.32 µm in B,C.

which in turn, sends intrinsic axon projections to the firstcluster (see Fig. 10).
DISCUSSION
Target specificity of supragranularpyramidal neuron projections
The intrinsic axon collaterals of supragranular pyrami-dal neurons in monkey PFC extend horizontally for sub-stantial distances and form stripe-like clusters of branch-ing axons and axon terminals in layers 1–3 (Levitt et al.,1993; Pucak et al., 1996). This study provides the firstultrastructural description of these intrinsic axon termi-nals and their postsynaptic targets, the majority (95%) ofwhich were dendritic spines. These axospinous synapseswere invariably of the asymmetric type (Gray’s Type I) andthus considered to be excitatory (Colonnier, 1968). Thepostsynaptic spines presumably belong to other pyramidalneurons, because they account for most of the spines in thePFC (Feldman, 1984; DeFelipe and Farinas, 1992). Thetargeted neurons likely include pyramidal cells in layers 2and 3, and may include a subpopulation of pyramidal cellsin layers 4–6, which have apical and oblique dendritesextending into the supragranular layers. Thus, the majorrole of the intrinsic axon collaterals of prefrontal supra-granular pyramidal cells appears to be to provide excita-tory drive to other pyramidal neurons.
Understanding the functional significance of this pre-dominant input to other pyramidal neurons depends, inpart, on knowing the relative specificity of this arrange-ment. The similarities in the postsynaptic targets ofprefrontal intrinsic, prefrontal associational, and parietalintrinsic terminals (Table 1) could reflect a general charac-teristic of cortical circuitry. However, because a signifi-cantly smaller proportion of all asymmetric synapses inthe general prefrontal neuropil target dendritic spines, ourfindings suggest that the axon terminals of prefrontalsupragranular pyramidal neurons are preferentially di-rected to the dendritic spines of other pyramidal cells.Given the criteria used to identify neuronal elements, it ispossible that some distal dendritic shafts were misidenti-fied as dendritic spines in the single section analysis usedin this study. However, this number is likely to be verysmall, would not be affected by the type of axon terminalexamined, and thus is not a confound for the data shown inTable 1.
The selective nature of the postsynaptic targets of theaxon terminals labeled in this study is further supportedby comparisons to the literature. For example, 97% of theintrinsic axon collaterals of callosally projecting pyramidal
neurons in mouse visual cortex were reported to contactdendritic spines (Czeiger and White, 1993). Moreover,other investigations (Elhanany and White, 1990; Whiteand Czeiger, 1991) in mouse somatosensory cortex haverevealed that approximately 90% of intrinsic axon collater-als of both associational and callosal pyramidal neuronssynapse onto dendritic spines. In contrast, the percentageof intrinsic axon projections forming excitatory synapseswith dendritic spines appears to be substantially lower inother cortical systems. For example, only 76% of thetargets of the horizontal connections in monkey visualcortex were dendritic spines (McGuire et al., 1991). Inaddition, studies of associational projections have revealeda selectivity of postsynaptic targets. For example, 90% ofthe axons involved in the feed-forward projection from ratprimary visual cortex to extrastriate cortex contact den-dritic spines, whereas 98% of those involved in the respec-tive feedback projection contact dendritic spines (Johnsonand Burkhalter, 1996). Together, these observations areconsistent with White’s proposition ‘‘that different axonalprojections form specific proportions of their synapses withdifferent postsynaptic structures’’ (White, 1989). However,further study is necessary to determine the extent towhich this specificity is a function of species, cortical area,or type of axonal projection.
In the present study, injections of BDA produced retro-grade labeling of a small number of pyramidal neurons inaddition to the anterograde labeling of axons and axonterminals. The presence of retrogradely labeled, BDA-positive neurons in coregistration with the intrinsic clus-ters of anterogradely labeled axons and axon terminals isconsistent with the findings of a previous study (Pucak etal., 1996) in which reciprocal connections, furnished byintrinsic axon collaterals, were present among spatiallysegregated clusters of supragranular pyramidal neurons(Fig. 10). In addition, the demonstration of asymmetricsynapses between BDA-labeled axon terminals and retro-gradely labeled dendritic spines suggests that these connec-tions may occur at the level of individual neurons. That is,the reciprocal connections between neuronal clusters mayinclude monosynaptic excitatory input from a pyramidalcell in one cluster to a pyramidal neuron in a secondcluster that in turn sends intrinsic axon collaterals to thefirst cluster. Such reciprocal excitatory connections be-tween patches of pyramidal neurons have also been demon-strated in cat visual cortex (Kisvarday and Eysel, 1992).However, it is important to note that these observations donot answer the question of whether reciprocity is completeor is affected by the distance between neuronal clusters orother factors.
However, the apparent presence of direct synaptic linksbetween pyramidal neurons located in different clustersmust be interpreted with caution, because it is possiblethat some of the labeled axon terminals observed in thisstudy arose from retrogradely labeled neurons within thestripe. This would require local anterograde transport ofBDA through an axon collateral in neurons that had beenretrogradely labeled through another axon collateral. How-ever, several lines of evidence argue against this interpre-tation. First, although BDA has recently been reported tobe retrogradely transported by a cerebellar granule cellaxon collateral and then anterogradely transported alonganother collateral (Chen and Aston-Jones, 1996), the char-acteristic features of such collateral–collateral transportwere not observed in this study. For example, despite the
TABLE 1. Comparisons of the Synaptic Targets of Prefrontal and ParietalAxon Terminals
Intrinsicprefrontal
cortex1
Associationalprefrontal
cortex1
Intrinsicparietalcortex1
Neuropilprefrontal
cortex2
Total number of termi-nals analyzed 419 243 152 225
Number of terminalsforming synapses 137 (32.7%) 74 (30.4%) 52 (34.2%) 225 (100%)
Synaptic targets-Spines 131 (95.6%) 68 (91.9%) 49 (94.2%) 170 (75.6%)Shafts 6 (4.4%) 6 (8.1%) 1 (1.9%) 55 (24.4%)Somata 0 (0%) 0 (0%) 2 (3.8%) 0 (0%)
1Terminals identified by the presence of biotinylated dextran amine.2Consisted of unlabeled axon terminals that formed asymmetric synapses.
EXCITATORY CONNECTIONS IN MONKEY PREFRONTAL CORTEX 219
extensive labeling of the dendritic trees of some pyramidalneurons (see Fig. 8), only the initial portion of their axonscould be identified. In addition, no labeled axon collateralsemanating from the principal axons were observed. Sec-ond, few retrogradely labeled neurons were present withinthe clusters of axon terminals. Thus, the probability thatthe labeled axon terminals forming asymmetric, axospi-nous synapses with retrogradely labeled dendritic spinesarose from collateral–collateral transport would appear tobe low. Finally, previous studies have shown that pyrami-dal neurons contact their targeted postsynaptic neuronwith a low frequency, usually only one synapse per neuron(Kisvarday et al., 1994). Therefore, the probability ofidentifying the one synapse between two pyramidal neu-rons within a stripe is quite low, especially when thenumber of such retrogradely labeled neurons is small.Thus, our findings are most consistent with the interpreta-tion that the observed synapses between labeled axon
terminals and retrogradely labeled dendritic spines repre-sent direct, reciprocal connections between supragranularpyramidal cells in different neuronal clusters. However,additional studies are required to prove this assertionunequivocally.
Implications for PFC functional circuitry
If the presence of reciprocal excitatory connections be-tween clusters of supragranular pyramidal neurons isconfirmed then, as previously suggested (Lewis and Ander-son, 1995; Pucak et al., 1996), this circuitry could provide apossible anatomical substrate for the sustained activity ofthese neurons during the delay period of delay–responsetasks (Fuster et al., 1982). Furthermore, the presence ofthese reciprocal connections in both prefrontal and poste-rior parietal cortices may contribute to the maintenance ofneuronal activity during the delay period in both cortical
Fig. 7. Fluorescent photomicrographs of retrogradely labeled, FastBlue (FB)-positive pyramidal neurons (A,C) and retrogradely labeled,cholera toxin (CtB)-positive pyramidal neurons (B,D) in the prefrontalcortex. Many of the FB-positive neurons are also positive for CtB
(arrows). However, FB-positive neurons that are not retrogradelylabeled with CtB are also present (open arrow in C,D). Scale bar 5 30 µm(applies to A–D).
220 D.S. MELCHITZKY ET AL.

regions (Fuster, 1989; Funahashi et al., 1989; Koch andFuster, 1989). However, compared with the patch-likearrays of interconnected neurons in the posterior corticalregions, the intrinsic projections in the PFC appear to linkgreater numbers of neurons distributed in larger, stripe-like clusters (Lund et al., 1993). Although this interpreta-tion requires confirmation, these differences suggest thatpyramidal neurons may be connected via intrinsic collater-als to a greater degree in the PFC than in posteriorparietal regions. This more extensive interconnectivitycould provide a possible explanation for the greater resis-tance of prefrontal neurons to the disruption of delay-related activity by intervening stimuli (Di Pellegrino andWise, 1993a, 1993b; Constantinidis and Steinmetz, 1996;Miller et al., 1996).
The intrinsic axon collaterals of supragranular pyrami-dal neurons examined in this study also provided synapticinput to dendritic shafts, many of which exhibited thecharacteristics of local circuit neuron dendrites, such as avaricose morphology and a high degree of synaptic input(Smiley and Goldman-Rakic, 1993; Sesack et al., 1995).Although the percentage of labeled terminals that tar-geted dendritic shafts was relatively low (approximately5%), even a small degree of input to inhibitory neurons canhave a profound influence on the activity of the localcircuitry (Kisvarday et al., 1994). This influence is mostlikely the result of two features of local circuit neurons.First, the typically divergent axons of these neuronsenables them to innervate multiple targets, and second,local circuit neurons often provide multiple synaptic in-puts to each target neuron (Kisvarday et al., 1994).Although it was not possible to determine which subclassof local circuit neurons was targeted by the intrinsic axoncollaterals of supragranular pyramidal neurons, several
characteristics of the wide-arbor class make these neuronsinteresting candidates for this input (Lewis and Anderson,1995). In the PFC, wide-arbor local circuit neurons arepresent in the supragranular layers (Lund and Lewis,1993) and a substantial proportion of their terminals formsymmetric synapses with the cell bodies of pyramidalneurons (Williams et al., 1992). Furthermore, the axonalarbor of these cells may extend for 450 to 500 µm from thecell body (Lund and Lewis, 1993), and based on theirsimilarities to the basket cells of motor cortex, this arbormay extend preferentially in one plane (Marin-Padilla,1974). Thus, as shown in Figure 10, excitatory input towide-arbor neurons located within a given stripe-likecluster of supragranular pyramidal neurons could providea means for inhibiting the activity of pyramidal neuronslocated in the gap or interstripe regions. In this way,activation of pyramidal neurons within a given stripe-likecluster could result in both the sustained firing of pyrami-dal neurons within interconnected stripes, as well as thesuppression of activity in pyramidal neurons located in theinterstripe regions. Such an arrangement might help tomaintain the fidelity of the information encoded by theactivity of the interconnected stripes.
In summary, the intrinsic axon collaterals of supragranu-lar pyramidal neurons in monkey PFC predominantlycontact the dendritic spines of other pyramidal cells lo-cated in spatially segregated clusters. The associationalprojections of these same neurons also preferentially tar-get the dendritic spines of other pyramidal cells. Thesepatterns of connectivity may provide an anatomical sub-strate for the recruitment of distinct networks of neuronswithin a prefrontal region, whose coordinated activitiessubserve the generation of specific patterns of output fromthis region. In addition, the output conveyed via the
Fig. 8. A: Brightfield photomicrograph of a retrogradely labeled,biotinylated dextran amine (BDA)-containing pyramidal neuron lo-cated within a cluster of anterogradely labeled intrinsic axons andterminals in area 9. B: Higher power photomicrograph of the apical
dendrite of the same pyramidal neuron illustrating the extent oflabeling in the dendritic spines (small arrows). Arrowheads point tothe same branch point in both panels. Scale bars 5 50 µm in A, 10 µmin B.
EXCITATORY CONNECTIONS IN MONKEY PREFRONTAL CORTEX 221
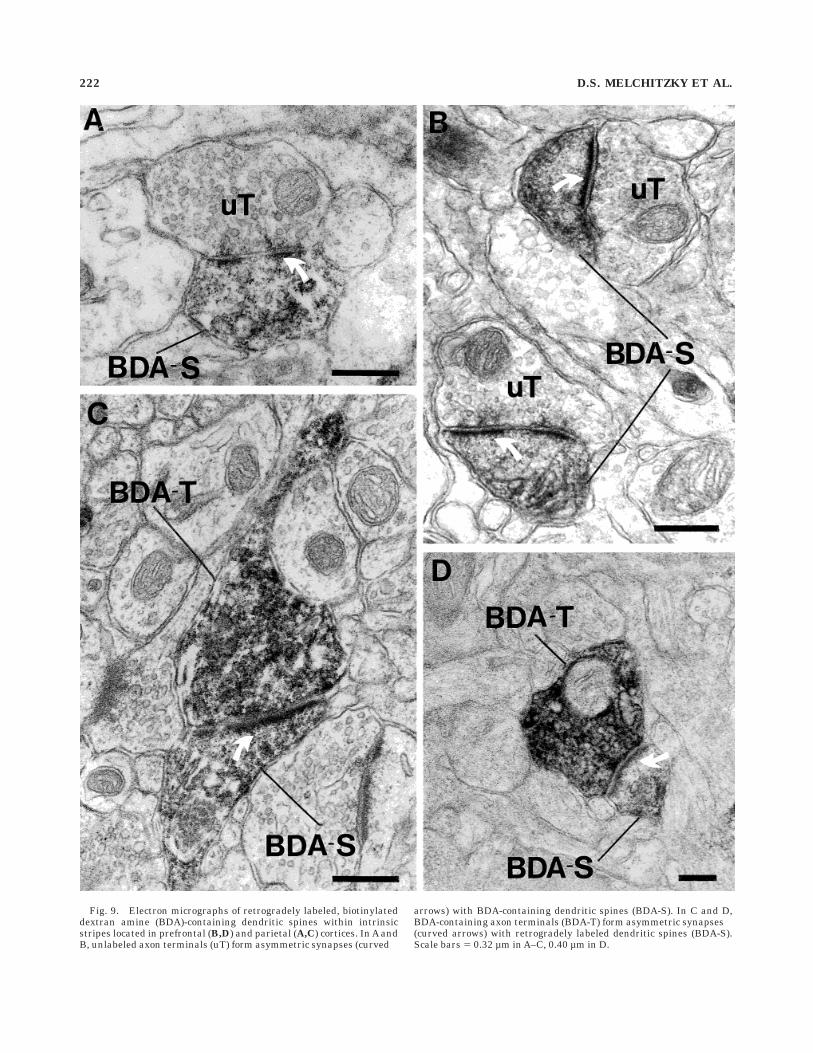
Fig. 9. Electron micrographs of retrogradely labeled, biotinylateddextran amine (BDA)-containing dendritic spines within intrinsicstripes located in prefrontal (B,D) and parietal (A,C) cortices. In A andB, unlabeled axon terminals (uT) form asymmetric synapses (curved
arrows) with BDA-containing dendritic spines (BDA-S). In C and D,BDA-containing axon terminals (BDA-T) form asymmetric synapses(curved arrows) with retrogradely labeled dendritic spines (BDA-S).Scale bars 5 0.32 µm in A–C, 0.40 µm in D.
222 D.S. MELCHITZKY ET AL.
principal axon projections of these supragranular pyrami-dal neurons may serve to activate more broadly distrib-uted neuronal networks in other cortical regions. Suchconnectivity may play a crucial role in working memoryand other cognitive functions mediated by the PFC.
ACKNOWLEDGMENTS
The authors thank Mary Brady for excellent technicalassistance.
LITERATURE CITED
Andersen, R.A., C. Asanuma, and W.M. Cowan (1985) Callosal and prefron-tal associational projecting cell populations in area 7A of the macaquemonkey: A study using retrogradely transported fluorescent dyes. J.Comp. Neurol. 232:443–455.
Baddeley, A. (1992) Working memory. Science 255:556–559.Barbas, H., and D.N. Pandya (1989) Architecture and intrinsic connections of
the prefrontal cortex in the rhesus monkey. J. Comp. Neurol. 286:353–375.Chen, S., and G. Aston-Jones (1996) Biocytin and biotinylated dextran
amine (BDA): False positive anterograde labeling by collateral-collateral transport. Soc. Neurosci. Abstr. 22:1731.
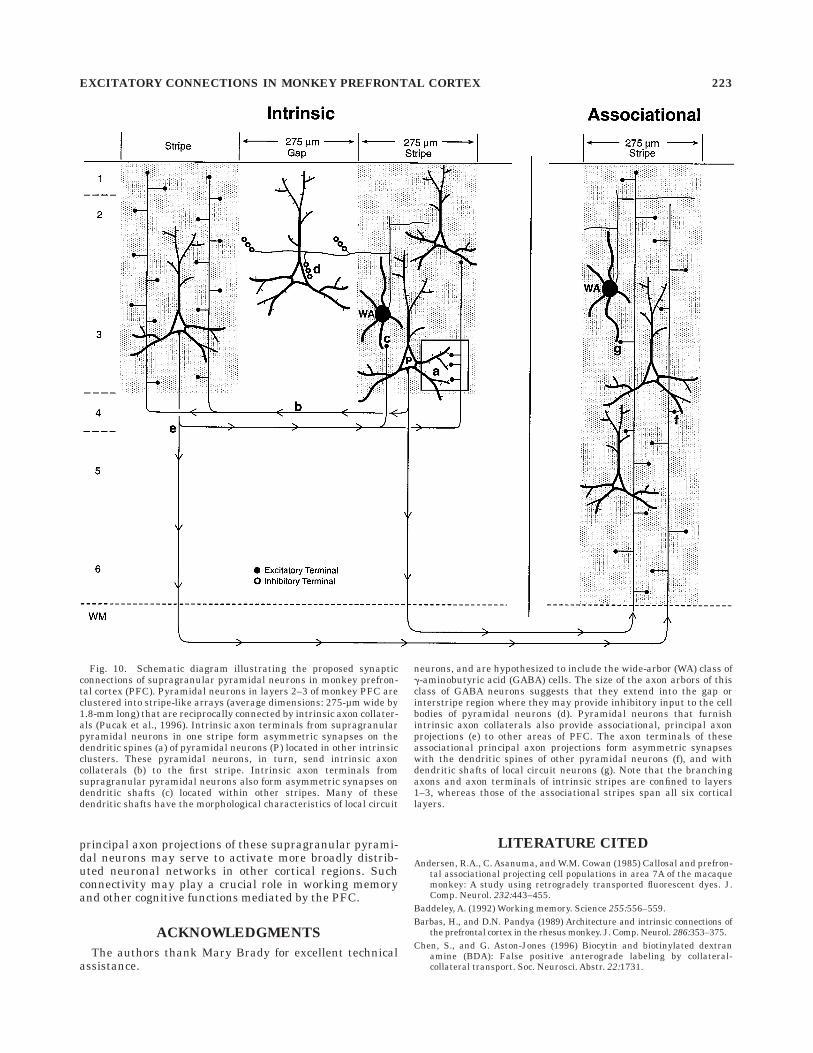
Fig. 10. Schematic diagram illustrating the proposed synapticconnections of supragranular pyramidal neurons in monkey prefron-tal cortex (PFC). Pyramidal neurons in layers 2–3 of monkey PFC areclustered into stripe-like arrays (average dimensions: 275-µm wide by1.8-mm long) that are reciprocally connected by intrinsic axon collater-als (Pucak et al., 1996). Intrinsic axon terminals from supragranularpyramidal neurons in one stripe form asymmetric synapses on thedendritic spines (a) of pyramidal neurons (P) located in other intrinsicclusters. These pyramidal neurons, in turn, send intrinsic axoncollaterals (b) to the first stripe. Intrinsic axon terminals fromsupragranular pyramidal neurons also form asymmetric synapses ondendritic shafts (c) located within other stripes. Many of thesedendritic shafts have the morphological characteristics of local circuit
neurons, and are hypothesized to include the wide-arbor (WA) class ofg-aminobutyric acid (GABA) cells. The size of the axon arbors of thisclass of GABA neurons suggests that they extend into the gap orinterstripe region where they may provide inhibitory input to the cellbodies of pyramidal neurons (d). Pyramidal neurons that furnishintrinsic axon collaterals also provide associational, principal axonprojections (e) to other areas of PFC. The axon terminals of theseassociational principal axon projections form asymmetric synapseswith the dendritic spines of other pyramidal neurons (f), and withdendritic shafts of local circuit neurons (g). Note that the branchingaxons and axon terminals of intrinsic stripes are confined to layers1–3, whereas those of the associational stripes span all six corticallayers.
EXCITATORY CONNECTIONS IN MONKEY PREFRONTAL CORTEX 223

Cipolloni, P.B., and D.N. Pandya (1989) Connectional analysis of theipsilateral and contralateral afferent neurons of the superior temporalregion in the rhesus monkey. J. Comp. Neurol. 281:567–585.
Colonnier, M. (1968) Synaptic patterns on different cell types in thedifferent laminae of the cat visual cortex. An electron microscope study.Brain Res. 9:268–287.
Constantinidis, C., and M.A. Steinmetz (1996) Neuronal activity in poste-rior parietal area 7a during the delay periods of a spatial memory task.J. Neurophysiol. 76:1352–1355.
Czeiger, D., and E.L. White (1993) Synapses of extrinsic and intrinsic originmade by callosal projection neurons in mouse visual cortex. J. Comp.Neurol. 330:502–513.
DeFelipe, J., and I. Farinas (1992) The pyramidal neuron of the cerebralcortex: Morphological and chemical characteristics of the synapticinputs. Prog. Neurobiol. 39:563–607.
Di Pellegrino, G., and S.P. Wise (1993a) Effects of attention on visuomotoractivity in the premotor and prefrontal cortex of a primate. Somatosens.Mot. Res. 10:245–262.
Di Pellegrino, G., and S.P. Wise (1993b) Visuospatial versus visuomotoractivity in the premotor and prefrontal cortex of a primate. J. Neurosci.13:1227–1243.
Elhanany, E., and E.L. White (1990) Intrinsic circuitry: Synapses involvingthe local axon collaterals of corticocortical projection neurons in themouse primary somatosensory cortex. J. Comp. Neurol. 291:43–54.
Feldman, M.L. (1984) Morphology of the neocortical pyramidal neuron. InA. Peters and E.G. Jones (eds): Cerebral Cortex. Cellular Componentsof the Cerebral Cortex. New York: Plenum Press, pp. 123–200.
Funahashi, S., C.J. Bruce, and P.S. Goldman-Rakic (1989) Mnemoniccoding of visual space in the monkey’s dorsolateral prefrontal cortex. J.Neurophysiol. 61:331–349.
Fuster, J.M., R.H. Bauer, and J.P. Jervey (1982) Cellular discharge in thedorsolateral prefrontal cortex of the monkey in cognitive tasks. Exp.Neurol. 77:679–694.
Fuster, J.M. (1985) The prefrontal cortex, mediator of cross-temporalcontingencies. Hum. Neurobiol. 4:169–179.
Fuster, J.M. (1989) The Prefrontal Cortex: Anatomy, Physiology andNeuropsychology of the Frontal Lobe. New York: Raven Press.
Gilbert, C.D., and T.N. Wiesel (1989) Columnar specificity of intrinsichorizontal and corticocortical connections in cat visual cortex. J.Neurosci. 9:2432–2442.
Glantz, L.A., and D.A. Lewis (1995) Assessment of spine density on layer IIIpyramidal cells in the prefrontal cortex of schizophrenic subjects. Soc.Neurosci. Abstr. 21:239
Goldman-Rakic, P.S. (1987) Circuitry of primate prefrontal cortex andregulation of behavior by representational memory. In F. Plum and V.Mountcastle (eds): Handbook of Physiology. Bethesda, MD: AmericanPhysiological Society, pp. 373–417.
Goldman-Rakic, P.S. (1995) Cellular basis of working memory. Neuron14:477–485.
Hof, P.R., K. Cox, and J.H. Morrison (1990) Quantitative analysis of avulnerable subset of pyramidal neurons in Alzheimer’s disease: I. Superiorfrontal and inferior temporal cortex. J. Comp. Neurol. 301:44–54.
Johnson, R.R., and A. Burkhalter (1996) Microcircuitry of forward andfeedback connections within rat visual cortex. J. Comp. Neurol. 368:383–398.
Kisvarday, Z.F., and U.T. Eysel (1992) Cellular organization of reciprocalpatchy networks in layer III of cat visual cortex (area 17). Neuroscience46:275–286.
Kisvarday, Z.F., D.-S. Kim, U.T. Eysel, and T. Bonhoeffer (1994) Relation-ship between lateral inhibitory connections and the topography of theorientation map in cat visual cortex. Eur. J. Neurosci. 6:1619–1632.
Koch, K.W., and J.M. Fuster (1989) Unit activity in monkey parietal cortexrelated to haptic perception and temporary memory. Exp. Brain Res.76:292–306.
Kritzer, M.F., and P.S. Goldman-Rakic (1995) Intrinsic circuit organizationof the major layers and sublayers of the dorsolateral prefrontal cortex inthe rhesus monkey. J. Comp. Neurol. 359:131–143.
Levitt, J.B., D.A. Lewis, T. Yoshioka, and J.S. Lund (1993) Topography ofpyramidal neuron intrinsic connections in macaque monkey prefrontalcortex (areas 9 & 46). J. Comp. Neurol. 338:360–376.
Lewis, D.A., and S.A. Anderson (1995) The functional architecture of theprefrontal cortex and schizophrenia. Psychol. Med. 25:887–894.
Lund, J.S., T. Yoshioka, and J.B. Levitt (1993) Comparison of intrinsicconnectivity in different areas of macaque monkey cerebral cortex.Cereb. Cortex 3:148–162.
Lund, J.S., and D.A. Lewis (1993) Local circuit neurons of developing andmature macaque prefrontal cortex: Golgi and immunocytochemicalcharacteristics. J. Comp. Neurol. 328:282–312.
Malach, R., Y. Amir, M. Harel, and A. Grinvald (1993) Relationship betweenintrinsic connections and functional architecture revealed by opticalimaging and in vivo targeted biocytin injections in primate striatecortex. Proc. Natl. Acad. Sci. USA 22:10469–10473.
Marin-Padilla, M. (1974) Three dimensional reconstruction of the pericellu-lar nests (baskets) of the motor (area 4) and visual (area 17) areas of thehuman cerebral cortex: A Golgi study. Z. Anat. Entwicklungsgesch.144:123–135.
McGuire, B.A., C.D. Gilbert, P.K. Rivlin, and T.N. Wiesel (1991) Targets ofhorizontal connections in macaque primary visual cortex. J. Comp.Neurol. 305:370–392.
Miller, E.K., C.A. Erickson, and R. Desimone (1996) Neural mechanisms ofvisual working memory in prefrontal cortex of the macaque. J. Neuro-sci. 16:5154–5167.
Morecraft, R.J., and G.W. Van Hoesen (1993) Frontal granular cortex inputto the cingulate (M3), supplementary (M2), and primary (M1) motorcortices in the rhesus monkey. J. Comp. Neurol. 337:669–689.
Neal, J.W., R.C.A. Pearson, and T.P.S. Powell (1987) The cortico-corticalconnections of area 7b,PF, in the parietal lobe of the monkey. Brain Res.419:341–346.
Oeth, K.M., and D.A. Lewis (1992) Cholecystokinin- and dopamine- contain-ing mesencephalic neurons provide distinct projections to monkeyprefrontal cortex. Neurosci. Lett. 145:87–92.
Peters, A., S.L. Palay , and D.F. Webster (1991) The Fine Structure of TheNervous System. New York: Oxford University Press.
Pucak, M.L., J.B. Levitt, J.S. Lund, and D.A. Lewis (1996) Patterns ofintrinsic and associational circuitry in monkey prefrontal cortex. J.Comp. Neurol. 376:614–630.
Schwartz, M.L., and P.S. Goldman-Rakic (1984) Callosal and intrahemi-spheric connectivity of the prefrontal association cortex in rhesusmonkey: Relation between intraparietal and principal sulcal cortex. J.Comp. Neurol. 226:403–420.
Sesack, S.R., C.L. Snyder, and D.A. Lewis (1995) Axon terminals immunola-beled for dopamine or tyrosine hydroxylase synapse on GABA-immunoreactive dendrites in rat and monkey cortex. J. Comp. Neurol.363:264–280.
Smiley, J.F., and P.S. Goldman-Rakic (1993) Heterogeneous targets ofdopamine synapses in monkey prefrontal cortex demonstrated by serialsection electron microscopy: A laminar analysis using the silver-enhanced diaminobenzidine sulfide (SEDS) immunolabeling technique.Cereb. Cortex 3:223–238.
Soloway, A.S., M.L. Pucak, and D.A. Lewis (1994) Dendritic morphology ofneurons with different axonal projections in monkey prefrontal cortex.Soc. Neurosci. Abstr. 20:1416.
Szabo, J., and W.M. Cowan (1984) A stereotaxic atlas of the brain of thecynomolgus monkey (Macaca fascicularis). J. Comp. Neurol. 222:265–300.
T’so, D.Y., C.D. Gilbert, and T.N. Wiesel (1986) Relationships betweenhorizontal connections and functional architecture in cat striate cortexas revealed by cross-correlation analysis. J. Neurosci. 6:1160–1170.
Von Bonin, G., and P. Bailey (1947) The Neocortex of Macaca Mulatta.Urbana, IL: The University of Illinois Press.
Walker, A.E. (1940) A cytoarchitectural study of the prefrontal area of themacaque monkey. J. Comp. Neurol. 73:59–86.
White, E.L. (1989) Cortical Circuits. Synaptic Organization of the CerebralCortex. Structure, Function and Theory. Boston, Basel, Berlin:Birkhauser.
White, E.L., and D. Czeiger (1991) Synapses made by axons of callosalprojection neurons in mouse somatosensory cortex: Emphasis on intrin-sic connections. J. Comp. Neurol. 303:233–244.
White, E.L., and A. Keller (1987) Intrinsic circuitry involving the local axoncollaterals of corticothalamic projection cells in mouse SmI cortex. J.Comp. Neurol. 262:13–26.
Williams, S.M., P.S. Goldman-Rakic, and C. Leranth (1992) The synaptol-ogy of parvalbumin-immunoreactive neurons in primate prefrontalcortex. J. Comp. Neurol. 320:353–369.
Woo, T.-U., M.L. Pucak, C.H. Kye, C.V. Matus, and D.A. Lewis (1997)Peripubertal refinement of the intrinsic and associational circuitry inmonkey prefrontal cortex. Neuroscience, 80:1149–1158.
Yoshioka, T., G.G. Blasdel, J.B. Levitt, and J.S. Lund (1996) Relationbetween patterns of intrinsic lateral connectivity, ocular dominance andcytochrome oxidase reactive regions in macaque monkey striate cortex.Cereb. Cortex 6:297–310.
224 D.S. MELCHITZKY ET AL.