summer 1993 microbial diversity - marine biological … · enrichment medium. ... castenholz and h....
TRANSCRIPT

Project Abstract / Adhesion-Mediated Enrichment of Purple Sulfur Bacteria
Carol BarfordMicrobial Diversity
Summer 1993
An independent research project was conducted which investigated the effects of adhesion to
different solid substrata on bacterial enrichments from an environmental inoculum. Stainless
steel, flint glass, polystyrene and tissue-culture polystyrene were chosen for their ready
availability and differing surface wettabilities, a characteristic which is known to influence
biological adhesion. Purple sulfur bacteria were enriched from samples of the pink layer of
Great Sippewisset Marsh microbial mat in anaerobic cultures to which the solid substrata were
added as 0.5 x 8.0 cm strips. The solid substrata were passaged through three stages of
enrichment medium. At each stage the culture supernatants and the attached microbial
community were examined by phase contrast microscopy. At the first and third stages
subsamples of the solid strips were taken for bacteriochiorophyll a (Bchl a) determination and
preparation for scanning electron microscopy. Bchl a content was also determined for the
third stage supernatants (bulk phase). Polystyrene appeared to be a favorable substratum for
the adhesion and growth of a Chromarium strain. By the third stage of enrichment a relatively
dense, homogeneous biofllm of a putative Chromarium had developed on polystyrene, which
was one of the less wettable substrates. This biofilm contained high amounts of Bchl a
(normalized against protein content), perhaps due to light attenuation by the thick biofilm.
Bacterial attachment to other substrata was less dense and resulted in mixed-morphotype
aggregates. In all cases the final bacterial enrichment in the bulk phase differed from the
biofilm.
0

Adhesion-Mediated Enrichment of Purple Sulfur Bacteria
Carol BarfordMicrobial Diversity
Summer 1993
Introduction
Adhesion of bacteria to solid surfaces is important in many natural microenvironments,
including soil crumbs, suspended particulate organic matter, and intra-animal surfaces
(Costerton et al. 1985). Physiology, morphology and community structure can differ between
attached and bulk-phase bacteria (Fletcher 1985). Such differences have practical implications
for clinical medicine, e.g. biofilm formation on implanted materials; water delivery and
wastewater treatment systems, and many other environments (Characklis and Marshall 1990).
Bacterial adhesion is a complex phenomenon which depends on surface characteristics
of the solid substrate and the bacterial cell, as well as environmental parameters such as pH
(Marshall 1976). Because it depends on several physical and chemical factors, adhesion
behavior is difficult to model. However, crude indices of surface characteristics can be used
for preliminary correlation with biological adhesion (Gerhart et a]. 1992). In this project solid
substrata with different wettabilities were incubated with environmental inocula under
conditions known to enrich for purple sulfur bacteria. It was hypothesized that bacterial
adhesion to the substrata would result in enrichments of different purple sulfur bacterial
communities after a few passages through the selective medium. Purple sulfur bacteria were
chosen for enrichment because of their growth habit in Great Sippewisset Marsh, where they
are present as a distinct, cohesive layer of pink sediment. This suggests the ability to adhere
and constitutes a targeted inoculum (Gibson et al, 1984).
0
,a. -a

Materials and MethodsCSolid substrata with a range of surface wettabilities were chosen. Table I lists
published values for wettability as well as measured values obtained by the same procedure
(Gerhart et al. 1992).
Table 1. Surface Wettabilities
Surface Wettability Wettability
(measured) (published)Stainless Steel 20.8
Flint Glass 68.9 95.0* borosilicate glassPolystyrene 20.5 25.6Tissue Culture Polystyrene 30.9 46.3
Solid substrata were cut into 0.5cm x 8cm strips and cleaned by 2 washes with 70% ethanol
and one wash with distilled water before inoculation. The inoculum was prepared from the
pink sediment layer of Sippewisset Marsh mat by combining approximately equal volumes of
sediment and sterile seawater in a screwcap tube with one drop of 1 M Na2S and vortexing.
Two ml of the resulting suspension were added to each of four 50 ml Pfennig bottles
containing 1) Pfennig’s purple sulfur medium (marine) with sodium acetate added to a final
concentration of 5mM (Pfennig and Truper 1992) and 2) each of the four substrate strips (four
bottles, sixteen strips). The enrichments were incubated in bright incandescent light at room
temperature for 5 days, which was sufficient time for the strips to be heavily colonized. After
the first incubation,every strip from three bottles was gently washed and subsampled for 1)
bacteriochlorophyll a (BehI a) determination , 2) preparation for scanning electron microscopy
(SEM), and 3) transfer to sterile purple sulfur medium containing a clean strip of the same

material. Two more iterations of the subsampling and transfer were made at 5 day intervals.
However SUM and Bchl a analyses were not performed at the second enrichment stage. At
the third enrichment stage Bchl a was also determined for the supernatants. The fourth bottle
at each stage of the enrichment was used for phase microscopic examination of the culture
supematants and attached bacteria. Bchl a was determined by A of acetone extracts and
normalized against total cell protein, which was determined by a modified Folin assay.
Results
1. Phase microscopy. In the first enrichment stage, heterogeneous microbial aggregates
formed on each solid substrate. These included purple sulfur bacteria and cyanobacteria,
spirochaetes, ciliates, diatoms and unidentified rods and spirillae. Attached microbial biomass
was visibly higher on the upper side of all strips, perhaps due to greater light availability.
The polystyrene strips supported relatively large, brownish-purple aggregates of Chromatiurnn(possibly C. vinosum). The second enrichment stage was more homogeneous and contained
few eukaryotes.. It was dominated by Chromarium and Thiocapsa strains, which formed very
homogeneous flocs in the bulk phase. At the second stage it became clear that the attached
and bulk phase bacterial communities were qualitatively dissimilar. At the third stage,
colonization of the substrata was slower, and few cells were attached to stainless steel after 5
days. The bulk phase of culture bottles was dominated by Thiocapsa (possibly T.
roseopersicina) with many smaller highly motile rods also present, while the solid substrata
were covered with finer-grained patterns of Chromatium.
2. SUM. In the first enrichment series, microbial colonization and growth on 1-2 cm length
subsamples of solid substrata was heterogeneous enough to preclude the possibility of finding
“representative” fields of view at magnifications higher than about 2,000x. However, SUM
nfl.., -4

enabled visualization of general patterns. Colonization of steel and glass was relatively fine
grained with mixed-morphotype aggregates, while aggregates on polystyrene and tissue-culture
polystyrene (TCPS) were larger and composed of single morphotypes. In the third enrichment
stage glass and polystyrene strips were covered with quite dense, homogeneous films of
Chromarium with easily visible capsular exopolysaccharide. TCPS was covered with a less
continuous biofilm of Chromatium, and steel strips exhibited patchy colonization by several
morphotypes.
3. Bchl a analysis. The results of Bchl a analyses on solid substrata and supematants are
given in Table 2. Normalized Bchl a content of attached bacteria was consistently highest for
polystyrene but no significant differences by substrate were detectable in the third stage
supernatants.
Table 2. Bchl a normalized by protein content (mg Bchl a/g protein).
. Stainless Steel Flint Glass Polystyrene TCPS
1st stage strips 38.1 ± 7.1 38.5 ± 2.5 135 ± 62 49•g*
3rd stage strips protein too low 27.3 ± 2.1 199 ± 43 58.1 ± 28
3rd stage spnt. 18.5 ± 8.9 33.5 ± 18 5.1* 42.3*
* single datum
Conclusions
It appears that polystyrene may be used to enrich for putative Chromatium (vinosum 7)
from a Sippewisset Marsh mat inoculum. Biofllms on polystyrene were homogeneous and
C) relatively dense as visualized with SEM. The enrichments attached to polystyrene differed

from the suspended bacterial community which suggests that aggregates and films found on
polystyrene grow in situ from a few attached cells rather than develop in an attachment-
detachment equilibrium with the bulk phase. This conclusion is also supported by the
homogeneity of bacterial aggregates on polystyrene from the first enrichment stage and the
amount of exopolysaccharide evident in the third stage biofllm SEM’s. Bchl a analysis
suggest that putative Chromatium cells attached to polystyrene may respond to light
attenuation by the thick biofilm by producing more Bchl a. The results suggest that this is a
quantitative response to bioftim thickness rather than a qualitative characteristic of the
attached putative Chromatiwn because the biofilms on glass were also homogeneous but did
not contain as much Bchl a per protein.
References
Characklis, W. and K. Marshall (eds). 1990. Biofilms. John Wiley and Sons, New York.
Costerton, J., T. Marrie and K. Cheng. 1985. Phenomena of bacterial adhesion. pp 3-43. In0. Savage and M. Fletcher (eds), Bacterial Adhesion. John Wiley and Sons, New York.
Gerhart, D., D. Rittschof, I. Hooper, K. Eisenman, A. Meyer, K Baler and C. Young. 1992.Rapid and inexpensive quantification of the combined polar components of surfacewettability: application to biofouling. Biofouling 5: 251-259.
Gibson, 0., E. Leadbetter and H. Jannasch. 1984. Great Sippewisset Marsh: A summary ofprojects carried out by students in the Microbial Ecology Course of the Marine BiologicalLaboratory, Woods Hole, During Summers 1972-1981. pp. 95-100. In Y. Cohen, R.Castenholz and H. Halvorson (eds), Microbial Mats: Stromatolites. Alan It Liss, New York.
Fletcher, M. 1985. Effect of solid surfaces on the activity of attached bacteria. In 0. Savageand M. Fletcher (eds), Bacterial Adhesion. John Wiley and Sons, New York.Marshall, K. 1976. Interfaces in Microbial Ecology. Harvard University Press, Cambridge,MA. 156 pp.
Pfennig, N. and H. Truper. 1992. The Family Chromatiaceae. pp. 3200-3222. In A. Balows,H. Truper, M. Oworkin, W. Harder and K. Schleifer (eds), The Prokaryotes. Vol. 4, SecondEdition. Springer-Verlag, Berlin.
0 Zisman, W. 1964. Relation of the equilibrium contact angle to liquid and solid constitution,
pS-fr 6

pp.1-51. In it Gould (ed),. Contact Angle, Wettability and Cohesion. American ChemicalSociety, Washington, D.C.
0

Project Abstract / Effect of Culture Conditions on Exopolysaccharide Production
Carol BarfordMicrobial Diversity
Summer 1993
Exopolysaccharide production in an enrichment of purple sulfur bacteria was studied.
A pure culture of putative Thiocapsa roseopersicina was isolated from an enrichment from
Great Sippewisset Marsh samples, grown in selective medium, then inoculated into
experimental media in a factorial matrix of three environmental parameters. The parameters
were pH (6.8. 7.2, 7.8), light intensity (200 and 20 uEIm2), and salinity (freshwater and
marine. Following 5 days’ incubation the cultures were subsanipled for total polysaccharide,
protein, and bacteriochiorophyll a content. Due to the short growth interval, protein was too
low to be determined in many of the cultures. Thus, no statistically significant relationships
between environmental parameters and exopolysaccharide (BPS) production were observed.
However, generally higher BPS production occurred at pH 7.8, which may indicate stress on
the bacterial isolate at that pH.
C

flThe Effects of Selected Environmental Parameters on Exopolysaccharide Production by
a Purple Sulfur Bacterium
Carol BarfordMicrobial Diversity
Summer 1993
Introduction
In many environments, bacteria live in aggregates or biofilms which consist largely of
cells in matrices of exopolysaccharide (e.g. Foster 1981, Costerton et al. 1985). The capsular
and peripheral exopolysaccharide (EPS) create protective microenvironments for bacteria.
BPS has been found to protect bacteria from desiccation (Roberson and Firestone 1992), metal
and antibiotic toxicity, shear stress, and starvation conditions (Ralph Mitchell, personal
communication). Conversely, BPS production can increase during physiological stress such as
desiccation (Roberson and Firestone 1992). In this project exopolysaccharide production by a
C) putative Thiocapsa roseopersicina strain was measured following incubation under different
pH, light, and salinity conditions in anaerobic liquid medium. It was hypothesized that
conditions differing from the selective enrichment conditions by which the strain was isolated
would induce BPS production. Salinity, light and pH were chosen because of the ease of
manipulation of these variables over relatively short incubation periods. Purple sulfur bacteria
were chosen as experimental organisms due to apparent BPS production in their growth habits
as aggregates and microbial mats.
Materials and Methods
A pure culture of putative Thiocapsa roseopersicina was isolated from an agar shake
series kindly provided by Minoru Wada, a fellow student. The original enrichment inoculum
was collected from Great Sippewisset Marsh. A cohesive, maroon-colored colony was
0 picked from the agar and tranferred to selective purple sulfur medium (Pfennig and Truper
a _ •.Z.

1992), amended sodium acetate to 5 mM final concentration and marine salts solution. The
culture was grown in bright incandescent light at room temperature for 8 days, then
transferred to flesh medium (backup culture) and inoculated into the experimental media. At
this point BPS could be seen in negative-stained wet mounts by brightfield microscopy. The
experimental culture conditions included duplicates of all factorial combinations of three pH
levels (6.8, 7.2, 7.8), two light levels (200 and 20 uEIm2), and two salinities (freshwater and
marine as defined by Pfennig and Truper, 1992). Experimental cultures were incubated for S
days. At the end of incubation the tubes were vortexed at high speed for 5 minutes to
disrupt the capsular EPS, then centifuged for 8 minutes at 14,000 rpm. The supernatants were
reserved for saccharide determination by the anthrone reaction. This was done by adding 2
ml of anthrone reagent (2 mg/nil in concentrated H2S04)to I ml of supematant, boiling the
mixture for 15 minutes, cooling on ice and measuring A6. The pellet was extracted with
100% acetone for 5 hours in the dark, and the supernatants used for determination of
bacteriochlorophyll a (Bchl a) by measuring A770. The protein content of the pellet was
determined by the Folin assay. BPS production in the cultures was normalized by protein
content and normalized Bchl a content was intended to be an index of light level adaptation.
Results
The results of all determinations are given in Table 1. There were no statistically
different levels of BPS production or Bchl a content between experimental treatments.
Conclusions
Although no statistically significant differences were found, possible trends can be
inferred from the data. In general there was higher normalized BPS production at pH 7.8.
This could indicate low tolerance of high pH in this putative Thiocapsa strain. There was

also a trend of higher protein and normalized Bchl a production, but not EPS production, at
high light level. This could be interpreted as reduced BPS production under favorable growth
conditions. The paucity of data resulted from insufficient incubation time. The Folin protein
assay appeared to be the least sensitive of the three analyses, and because protein content was
used to normalize the other data undetectably low protein values rendered the other values
useless. However, it would be easy to improve upon this project by increasing incubation
time, or possibly by adding solid substrata as incubation variables.
References
Costerton J., T. Marrie and K. Cheng. 1985. Phenomena of bacterial adhesion. pp.3-43. In 0.Savage and M. Fletcher (eds), Bacterial Adhesion. John Wiley and Sons, New York.
Foster, R 1981. Polysaccharides in soil fabrics. Science 214: 665-667.
Pfennig, N, and H. Truper. 1992. The Family Chromatiaceae. pp. 3200-3222. In A. Balows,H. Truper, M. Dworkin, W. Harder and K. Schleifer (eds), The Prokaryotes. Vol. 4, SecondEdition. Springer-Verlag, Berlin.
Roberson, E. arid M. Firestone, 1992. Relationship between desiccation and exopolysaccharideproduction in a soil Pseudomanas sp. Applied and Environmental Microbiology 58: 1284-1291
0
_,
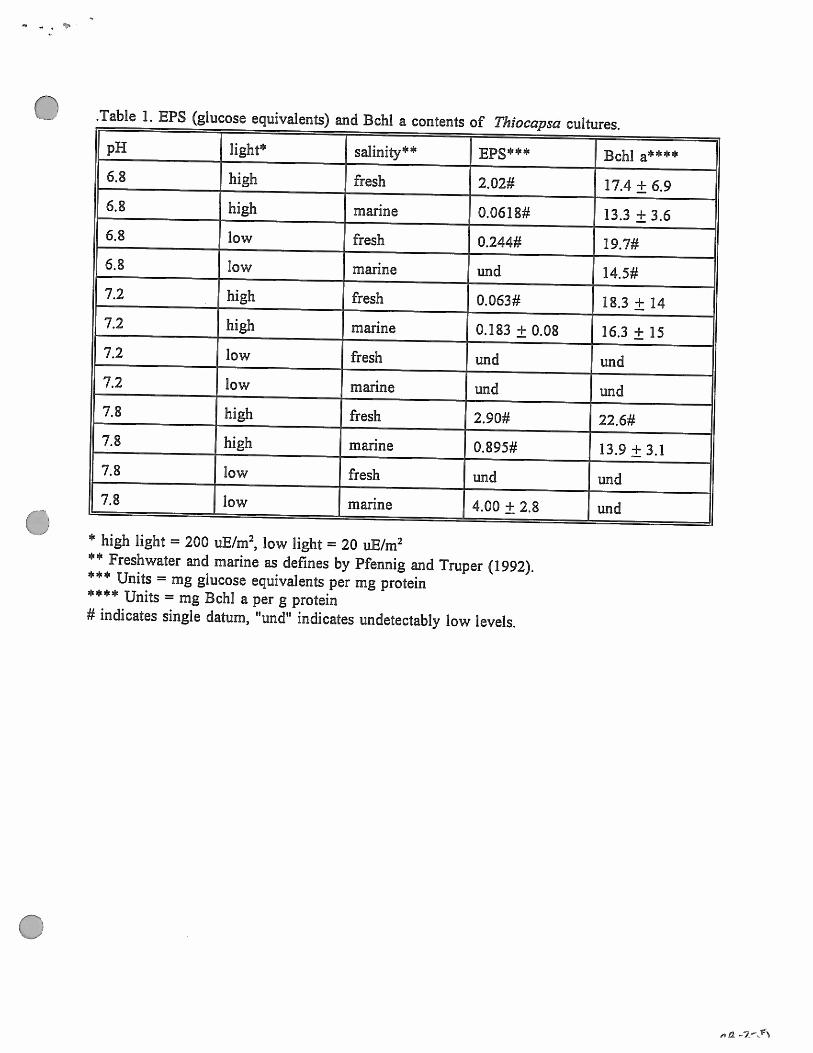
* high light = 200 tiE/rn2, low light = 20 uE/m2** Freshwater and marine as defines by Pfennig and Truper (1992).
Units = mg glucose equivalents per mg protein*44* Units = mg BehI a per g protein# indicates single datum, “und” indicates undetectably low levels.
C) .Table 1. EPS (glucose equivalents) and Echi a contents of Thiocapscz cultures.
pH light4 salinity4 BPS444 Bchl a4446.8 high fresh 2.02* 17.4 ± 6.96.8 high marine 0.0618* 13.3 ± 3.66.8 low fresh O.244# 19.7*6.8 low marine und 14.5*7.2 high fresh 0.063* 18.3 ± 147.2 high marine 0.183 ± 0.08 16.3 ± 157.2 low fresh und und7.2 low marine und und7.8 high fresh 2.90* 22.6*7.8 high marine 0.895* 13.9 ± 3.17.8 low fresh und und7.8 low marine 4.00 ± 2.8 und
C
C