suhteellisen ilmankosteuden ja kasvualustan...
TRANSCRIPT

SUHTEELLISEN ILMANKOSTEUDEN JA KASVUALUSTAN
TYPPIPITOISUUDEN VAIKUTUKSET RAUDUSKOIVUN
(Betula pendula Roth) LEHTIEN METABOLIITTEIHIN
VIRVE VILKMAN
Pro Gradu -tutkielma
Itä-Suomen yliopisto
Ympäristö- ja biotieteiden laitos
Biologia
2017

ITÄ-SUOMEN YLIOPISTO
Ympäristö- ja biotieteiden laitos, Biologia
VILKMAN, VIRVE: Suhteellisen ilmankosteuden ja kasvualustan typpipitoisuuden
vaikutukset rauduskoivun (Betula pendula Roth) lehtien
metaboliitteihin.
Pro Gradu -tutkielma (40 op), 47 s., 1 liite
Joulukuu 2017
___________________________________________________________________________
TIIVISTELMÄ
Useat ilmastonmuutosmallit ennustavat Pohjois-Euroopan vuotuisen sademäärän kasvavan,
sadepäivien ja pilvisyyden lisääntyvän sekä keskilämpötilan nousevan tämän vuosisadan
aikana, mikä johtaisi suhteellisen ilmankosteuden lisääntymiseen. Korkeassa suhteellisessa
ilmankosteudessa kasvin lehdissä tapahtuva haihdunta sekä veden sisäänotto juuristossa vähe-
nevät, jolloin myös ravinteiden imeytyminen maaperästä ja kuljetus kasvissa heikentyy. Tällöin
kasvin tarvitsee tehdä merkittäviä muutoksia aineenvaihdunnassaan ja kasvustrategioissaan,
jotta se sopeutuisi ja menestyisi muuttuvassa ilmastossa. Primaarimetaboliitit, kuten sokerit,
sokerialkoholit, orgaaniset hapot ja aminohapot, ovat välttämättömiä kasvin kasvulle ja kehi-
tykselle, sillä ne osallistuvat keskeisiin metaboliaprosesseihin, kuten glykolyysiin ja sitruuna-
happokiertoon. Primaarimetaboliitit toimivat myös esiasteina useille sekundaariyhdisteille,
kuten fenoleille ja terpenoideille, joilla on tärkeitä tehtäviä muun muassa kasvin puolustus-
mekanismeissa.
Tämän pro gradu -tutkielman tavoitteena oli selvittää, mitä metaboliitteja kammiokokeessa
kasvatettujen rauduskoivujen (Betula pendula Roth) lehdet ovat tuottaneet, sekä kuinka
suhteellisen ilmankosteuden ja kasvualustan typpipitoisuuden kaksi eri tasoa vaikuttivat näiden
metaboliittien pitoisuuksiin. Lehtinäytteet uutettiin ja derivatisoitiin analysoitaviksi kaasu-
kromatografi-massaspektrometrilla (GC-MS). Analyysituloksia ja kammiokokeen raudus-
koivujen kasvuaineistoa tarkasteltiin tilastollisilla menetelmillä. Metaboliitti- ja kasvuaineiston
välistä lineaarista riippuvuutta tarkasteltiin parametrisella Pearson-korrelaatiolla.
Kohotettu suhteellinen ilmankosteus vaikutti voimakkaasti rauduskoivun lehden metaboli-
aan. Useimpien primaarimetaboliittien tuotanto aleni kohotetussa suhteellisessa ilmankosteu-
dessa kasvatettujen rauduskoivujen lehdissä, ainoastaan raffinoosin pitoisuus nousi. Sekundaa-
rimetaboliiteista kversetiinin ja flavonoliglykosidin pitoisuudet nousivat ja salidrosidin
pitoisuus aleni kohotetussa suhteellisessa ilmankosteudessa kasvatettujen rauduskoivujen
lehdissä. Kohotettu suhteellinen ilmankosteus vaikutti koivuntaimen kasvuun lisäämällä
suhteellista pituus- ja paksuuskasvua sekä alentamalla lehden typpipitoisuutta ja klorofylli-
indeksiä. Kasvatus kasvualustan kohotetussa typpipitoisuudessa nosti malaatin ja sukkinihapon
sekä alensi 3-trans-kumaryyli-kiinihapon pitoisuuksia rauduskoivun lehdissä. Kasvualustan
typpipitoisuudella ei havaittu vaikutusta koivuntaimen kasvuun eikä lehtien ominaisuuksiin.
Kammiokokeen lyhyt kesto sekä istutusaineksen typen lähtötaso ovat voineet vaikuttaa siihen,
ettei kasvualustan typpipitoisuuden vaikutusta lehtien metaboliaan tai koivuntaimen kasvuun
juuri havaittu. Suhteellisen ilmankosteuden ja kasvualustan typpipitoisuuden yhdysvaikutusta
ei havaittu metaboliitti- eikä kasvuaineistossa. Tutkimus osoittaa suhteellisen ilmankosteuden
olevan hyvin tärkeä tekijä, joka vaikuttaa hiilen ja typen sitomiseen sekä niiden ohjaamiseen
eri metaboliareiteille kasveissa.

UNIVERSITY OF EASTERN FINLAND
Department of Environmental and Biological Sciences, Biology
VILKMAN, VIRVE: The effects of relative humidity and soil nitrogen level on
metabolites of silver birch (Betula pendula Roth) leaves.
MSc. Thesis (40 ECTS), 47 pp., 1 appendice
December 2017
___________________________________________________________________________
ABSTRACT
Several climate change scenarios predict significant increases in annual precipitation, number
of rainy days, cloudiness and average temperatures in Northern Europe during this century,
which in total would lead to elevated relative humidity. This will cause transpiration rate of
leaves and uptake of water in the roots to decline, thus decreasing absorption of mineral
nutrients from the soil and transportation within the plant. Significant alterations will be needed
in plant metabolism and growth strategies in order to acclimate successfully to changing
climate. Primary metabolites, such as carbohydrates, sugar alcohols, organic acids and amino
acids, are necessary for plant growth and development due to their involvement in pivotal
metabolic processes, such as glycolysis and TCA cycle. Primary metabolites also serve as pre-
cursors to secondary compounds, such as phenolics and terpenoids, which have important roles
in plant defence.
Aim of this MSc Thesis was to study which metabolites are produced in the leaves of silver
birches (Betula pendula Roth) grown in a chamber experiment under two different levels of
relative humidity and soil nitrogen level, and how these treatments affect the concentrations of
the metabolites. Leaf samples were extracted, derivatized and analyzed with a gas chromato-
graph coupled with mass spectrometer (GC-MS). Statistical analysis of the metabolite data and
growth parameter data was performed. Parametric Pearson-correlation tests were conducted to
explore linear correlation between metabolite and growth parameter data.
Elevated relative humidity had remarkable impacts on the metabolism of silver birch leaves.
Concentrations of most primary metabolites decreased, only raffinose content of leaves
increased. The concentrations of secondary metabolites quercetin and flavonol glycoside
increased, and salidroside decreased. Relative height and diameter growth of silver birches
increased, and nitrogen content and chlorophyll index of leaves decreased under elevated rela-
tive humidity. Elevated soil nitrogen level affected leaf metabolism by increasing the
concentrations of malic acid and succinic acid, and by decreasing the concentration of 3-trans-
coumaroyl-quinic acid. Elevated soil nitrogen level had no significant effect on growth para-
meters. It is possible, that the short duration of the chamber experiment and the nitrogen content
of the planting material have diminished the effects of elevated soil nitrogen level on leaf
metabolism and on growth parameters. There was no interaction found between relative
humidity and soil nitrogen level towards metabolites or growth parameters. This study shows
that relative humidity strongly affects accumulation of carbon and nitrogen and their allocation
towards different metabolic routes in plants.

SISÄLLYSLUETTELO
1 JOHDANTO ....................................................................................................................... 2
2 KASVISOLUN AINEENVAIHDUNTA ........................................................................... 3
2.1 Primaarimetabolia ........................................................................................................ 3
2.2 Typen sidonta orgaanisiin molekyyleihin.................................................................... 4
2.3 Sekundaarimetaboliitit ................................................................................................. 5
3 TUTKIMUKSEN TAVOITTEET JA TUTKIMUSKYSYMYKSET ................................ 5
4 AINEISTO JA MENETELMÄT ........................................................................................ 6
4.1 Näytemateriaalin tuottaminen kammiokokeessa ......................................................... 6
4.2 Metaboliatutkimus ....................................................................................................... 8
4.2.1 Homogenisointi .................................................................................................. 8
4.2.2 Uutto ................................................................................................................... 9
4.2.3 Derivatisointi .................................................................................................... 12
4.2.4 Kaasukromatografis-massaspektrometrinen analysointi .................................. 13
4.2.5 Yhdisteiden identifiointi ja kvantifiointi .......................................................... 14
4.2.6 Tilastollinen analyysi ....................................................................................... 15
5 TULOKSET ...................................................................................................................... 18
5.1 Näytteiden sijoittuminen pääkomponenttianalyysissa............................................... 18
5.2 Kohotetun suhteellisen ilmankosteuden vaikutus metaboliitteihin ........................... 19
5.3 Kasvualustan kohotetun typpipitoisuuden vaikutus metaboliitteihin ........................ 20
5.4 Koivun kasvu kammiokokeessa ................................................................................ 30
5.5 Rauduskoivun taimien kasvu- ja lehtien ominaisuuksien sekä lehtinäytteistä
määritettyjen metaboliittipitoisuuksien välinen Pearson-korrelaatio.…………....…… 32
6 TULOSTEN TARKASTELU ........................................................................................... 34
6.1 Kohotetun suhteellisen ilmankosteuden vaikutus metaboliitteihin ........................... 36
6.2 Kasvualustan kohotetun typpipitoisuuden vaikutus metaboliitteihin ........................ 38
6.3 Rauduskoivun kasvu kammiokokeessa ..................................................................... 38
6.4 Korrelaatio ................................................................................................................. 39
6.5 Kammiokoetulosten vertailu kenttäkokeen tuloksiin ................................................ 40
7 JOHTOPÄÄTÖKSET ....................................................................................................... 41
8 KIITOKSET ...................................................................................................................... 43
KIRJALLISUUSLUETTELO .................................................................................................. 43
LIITTEET ................................................................................................................................. 47

2
Ilmastonmuutoksen vaikutusten arviointiin, määrittelyyn ja ennustamiseen käytetään useita eri
skenaariomalleja, jotka perustuvat mitattuihin päästömääriin, vuosikymmenien sääseurantaan
ja tähän mennessä havaittuihin muutoksiin ilmakehässä (Stocker ym. 2013, Jaagus & Mändla
2014). Esimerkiksi tarkasteltuna 1950-luvulta 2010-luvulle on ilmakehän ja meriveden lämpö-
tila kohonnut, lumen ja jään määrä vähentynyt sekä merenpinta noussut (Stocker ym. 2013).
Hiilidioksidin, metaanin ja typen oksidien pitoisuudet ilmakehässä ovat korkeampia kuin
koskaan ennen. Äärisääilmiöt, kuten äärimmäinen kylmyys ja kuumuus sekä paikalliset rankka-
sateet, ovat yleistyneet. Myös paikalliset tuulituhot ovat lisääntyneet.
Ilmastonmuutoksen ennustetut vaikutukset vaihtelevat käytetystä ilmastonmuutosmallista ja
arvioidusta kasvihuonekaasujen tasosta riippuen (Stocker ym. 2013). Vuotuisen keskisade-
määrän on ennustettu kasvavan Pohjois-Euroopassa 5-30 %, sadepäivien ja pilvisyyden lisään-
tyvän sekä vuotuisen keskilämpötilan nousevan 2-4 °C nykytasosta vuoteen 2100 mennessä
(Kont ym. 2003). Talvisateiden määrän on ennustettu samalla aikavälillä kasvavan 20-50 %.
Kesäsateiden määrän muutoksen on ennustettu vaihtelevan -10 % ja + 20 % välillä (Kont ym.
2003, Jaagus & Mändla 2014). Talvikuukausien keskilämpötilojen on ennustettu nousevan 4-7
°C ja kesäkuukausien keskilämpötilojen nousevan 1-2 °C nykytasosta (Jylhä ym. 2004). Lämpi-
mämpi ilma voi lisätä haihduntaa ja pidättää enemmän vesihöyryä, jolloin suhteellisen ilman-
kosteuden (relative humidity, RH) voidaan odottaa kohoavan edellä mainittujen tekijöiden
yhteisvaikutuksesta. Suhteellinen ilmankosteus tarkoittaa prosenttilukua, joka ilmaisee, kuinka
paljon ilmassa on vesihöyryä suhteessa siihen, mitä se voi kyseisessä lämpötilassa enimmillään
sisältää (Ilmatieteenlaitos 2017). Jos ilmastonmuutokseen liittyy lämpenemisen ja ilman hiili-
dioksidipitoisuuden kohoamisen ohella myös suhteellisen ilmankosteuden nousu, voivat
kasvien reaktiot ilmastonmuutokseen erota siitä, mitä olisi odotettavissa pelkästään ilmaston
lämpenemisen ja hiilidioksidipitoisuuden nousun perusteella. Tätä taustaa vasten on tärkeää
selvittää mahdollisimman monipuolisesti, kuinka kasvien aineenvaihdunta reagoi muuttuvaan
ilmastoon.
Suhteellinen ilmankosteus voi vaikuttaa merkittävästi lehtipuiden aineenvaihduntaan ja
kasvuun, sillä korkeassa suhteellisessa ilmankosteudessa kasvin ylemmissä osissa tapahtuva
haihdunta, nestevirtaus kasvissa ja niiden myötä juuriston veden ja ravinteiden otto vähenevät
(Tullus ym. 2012, Sellin ym. 2013, Rosenvald ym. 2014, Sellin ym. 2015, Lihavainen ym.
2016a,b). Toisaalta korkea suhteellinen ilmankosteus voi lisätä kasvin lehden ilmarakojen auki-
oloa ja sen myötä edistää hiilidioksidin sitomista ilmasta (Gislerød & Nelson 1989, Fordham
1 JOHDANTO

3
ym. 2001, Lihavainen ym. 2016a). Kasvualustan korkea ravinnepitoisuus voi vaikuttaa siihen,
kuinka voimakkaasti korkeammasta suhteellisesta ilmankosteudesta aiheutuva heikentynyt
ravinteiden otto hidastuu (Lihavainen ym. 2016a). Muutokset kasvuolosuhteissa, erityisesti
kasvualustan ravinnepitoisuudessa (Keinänen ym. 1999, Keski-Saari & Julkunen-Tiitto 2003,
Lihavainen ym. 2016a,b), ilmakehän kaasukoostumuksessa ja suhteellisessa ilmankosteudessa
(Lihavainen ym. 2016a,b), vaikuttavat voimakkaasti kasvien primaari- ja sekundaarimetabo-
liaan (Geiger ym. 1999, Scheible ym. 2004, Lihavainen ym. 2016a,b). Primaarimetaboliassa
tuotetaan kasville välttämättömiä hiilihydraatteja muun muassa tukemaan kasvin kasvua ja
kehitystä sekä solurakenteen ylläpitoa. Sekundaarimetaboliassa tuotetut yhdisteet osallistuvat
viestinvälitykseen kasvin ja ympäristön välillä, toimien esimerkiksi kasvin puolustusyhdisteinä
(Kutchan ym. 2015).
Koivut (Betula L.), erityisesti rauduskoivu (Betula pendula Roth), ovat valoisalla paikalla
viihtyviä nopeakasvuisia pioneeripuita (Hynynen ym. 2010), jotka valtaavat avohakkuualueet
ja kesannolle jätetyt pellot ja niityt muutamassa vuodessa (Viherä-Aarnio 2008). Rauduskoivu
ja hieskoivu (Betula pubescens Ehrh.) ovat Pohjois-Euroopan yleisimpiä ja kaupallisesti
merkittävimpiä lehtipuita, kattaen Suomessa noin 15 % metsäalasta (Hynynen ym. 2010).
Nopeakasvuisina puina koivut ovat otollisia tutkimuskohteita ulkoisten stressitekijöiden
muutoksen vaikutusten selvittämiseen, sillä selviytyäkseen mahdollisimman vähin vaurioin
koivujen on sopeuduttava muuttuvaan ympäristöön nopeasti.
Pro gradu -tutkielmani tarkoituksena oli selvittää mitä primaari- ja sekundaarimetaboliitteja
kammiokokeessa kasvatetut rauduskoivun lehdet ovat tuottaneet, sekä kuinka suhteellinen
ilmankosteus (RH) ja kasvualustan typpipitoisuus (N) vaikuttavat näiden metaboliittien
pitoisuuksiin ja rauduskoivujen kasvuun.
2.1 Primaarimetabolia
Kasvien aineenvaihdunnassaan tuottamat yhdisteet eli metaboliitit voidaan jaotella primaari- ja
sekundaarimetaboliitteihin (mm. Taiz & Zeiger 2012, Buchanan ym. 2015). Primaarimetabo-
liitit, kuten hiilipohjaiset sokerit, sokerialkoholit ja orgaaniset hapot sekä typpeä sisältävät
aminohapot, osallistuvat keskeisiin metaboliaprosesseihin ja ovat siten välttämättömiä kasvin
kasvulle ja kehitykselle. Primaarimetaboliittien tuotantoon tarvitaan hapen ja vedyn lisäksi
hiiltä, joka sidotaan ilmakehän hiilidioksidista. Lehden alapinnan ilmarakojen kautta lehden
2 KASVISOLUN AINEENVAIHDUNTA

4
soluvälitilaan otetun ilmakehän hiilidioksidin sitominen hiiliyhdisteisiin käynnistyy viherhiuk-
kasen Calvin-Benson syklissä (Taiz & Zeiger 2012). Fotosynteesin monivaiheisessa
reaktioketjussa tuotetaan hiilirunkoja muun muassa glykolyysiin ja sitruunahappokiertoon
(kuva 1). Näiden reaktioketjujen välituotteet osallistuvat lukuisiin prosesseihin, esimerkiksi
amino- ja rasvahapposynteesiin, pigmentti- ja hormonisynteesiin sekä typen sidontaan. Esimer-
kiksi sikimihaposta syntetisoitavat aromaattiset aminohapot (fenyylialaniini, tryptofaani ja
tyrosiini) (kuva 1, Maeda & Dudareva 2012) toimivat esiasteina lukuisille sekundaarimetabo-
liiteille, kuten flavonoideille.
Kuva 1. Yksinkertaistettu malli primaarimetaboliasta: Glykolyysi, sitruunahappokierto ja aminohappo-
jen tuotanto. (Sweetlove & Fernie 2013, Lihavainen ym. 2016a,b mukaillen).
2.2 Typen sidonta orgaanisiin molekyyleihin
Typpi on kasveille välttämätön alkuaine, jota tarvitaan muiden muassa amino- ja nukleiini-
happojen, proteiinien, klorofyllin, soluseinän komponenttien sekä hormonien ja vitamiinien

5
tuotannossa (Krapp 2015). Kasvit saavat typpeä pääasiallisesti nitraatteina (NO3-) ja ammo-
niumioneina (NH4+) juuriston kautta maaperästä muun muassa orgaanisesta maannoksesta,
symbionttisilta typensitojamikrobeilta ja lannoitteesta, sekä vähäisessä määrin typenoksideina
ilmakehästä lehtien ilmarakojen kautta (Taiz & Zeiger 2012). Maaperän typpi siirretään juuris-
ton kautta nilakuljetuksella lehden fotosynteettisiin mesofyllisoluihin, joissa kasvin tarvitsemat
typpiyhdisteet muodostetaan (Taiz & Zeiger 2012, Krapp 2015).
2.3 Sekundaarimetabolia
Sekundaarimetaboliitit ovat orgaanisia yhdisteitä, jotka osallistuvat kasvin ja sen ympäristön
väliseen kommunikointiin (Kutchan ym. 2015) toimien esimerkiksi kasvin väri-, tuoksu-,
houkutin-, puolustus- ja suoja-aineina. Kasveilla on lajityypillisiä sekundaariyhdisteitä eriä-
vissä määrin usein varsin huomattavinakin pitoisuuksina, ja niiden merkitystä kasveille on
pyritty useissa tutkimuksissa selvittämään (mm. Bi & Felton 1995, Lavola ym. 1998, Keinänen
ym. 1999, Lavola ym. 2000, de la Rosa ym. 2001, Keski-Saari ym. 2005, Akula & Ravishankar
2011, Schreiner ym. 2012). Sekundaariyhdisteitä esiintyy myös stressittömässä tilassa perusva-
rantona (Keski-Saari & Julkunen-Tiitto 2003), mutta erityisesti kasvin puolustukseen liittyvien
sekundaariyhdisteiden tuotanto lisääntyy vasteena ulkoisiin stressitekijöihin, kuten ympäristön
muutokseen tai herbivoriaan (mm. Bi & Felton 1995, Akula & Ravishankar 2011). Sekundaa-
riyhdisteiden on esitetty toimivan kasveissa myös oksidaatio- ja UV-vaurioiden estäjinä (mm.
Bi & Felton 1995, Lavola 1998, Lavola ym. 2000, Mewis ym. 2012, Schreiner ym. 2012,
Kurepa ym. 2016).
Sekundaarimetaboliitit luokitellaan typpipohjaisiin, kuten alkaloidit, ja hiilipohjaisiin, kuten
muun muassa fenoliset yhdisteet ja terpenoidit (Kutchan ym. 2015). Rauduskoivun (Betula
pendula Roth) sekundaariyhdisteet ovat hiilipohjaisia ja niiden pitoisuudet voivat olla merkit-
täviä (Keinänen ym. 1999). Koivunlehtien sekundaariyhdisteiden tiedetään vaikuttavan
esimerkiksi niiden kelpaavuuteen ja soveltuvuuteen herbivorien ravinnoksi (Hartley & Firn
1989, Martemyanov ym. 2015).
Tämän pro gradu -tutkielman tavoitteena on tuottaa uutta tietoa rauduskoivun (Betula pendula
Roth) sopeutumisesta ilmastonmuutokseen liittyvän suhteellisen ilmankosteuden kasvuun. Pro
gradu -tutkimus on osa Kasvien ekofysiologian tutkimusryhmän tutkimusprojektia ”Koivun ja
3 TUTKIELMAN TAVOITTEET JA TUTKIMUSKYSYMYKSET

6
haavan molekulaariset vasteet muuttuvalle ilmankosteudelle”, ja siitä saatuja tuloksia hyödyn-
nettiin osana FT Jenna Lihavaisen väitöstutkimusta (Lihavainen ym. 2016a).
Pro gradu -tutkimuksen tavoitteena oli kaasukromatografis-massaspektrometrisin (GC-MS)
menetelmin selvittää, mitä primaari- ja sekundaarimetaboliitteja kammiokokeen aikana kasva-
neet rauduskoivun lehdet ovat tuottaneet. Lisäksi saatiin tarkasteltavaksi kammiokokeen
rauduskoivuista mitattu kasvuparametriaineisto (Lihavainen ym. 2016a). Pro gradu -tutkiel-
massani tarkastelen vaikuttaako suhteellinen ilmankosteus ja kasvualustan typpipitoisuus
tutkittujen metaboliittien pitoisuuksiin ja kasvuparametreihin. Tilastollisin menetelmin analy-
soidaan käsittelyjen pää- ja yhdysvaikutuksia rauduskoivun lehtien metaboliaan sekä
kasvuparametreihin. Lisäksi tarkastellaan, löytyykö metaboliittien pitoisuuksien ja kasvupara-
metrien välisiä lineaarisia riippuvuuksia.
Tutkimuskysymykset:
Kuinka kohotettu suhteellinen ilmankosteus vaikuttaa rauduskoivun lehtien primaari- ja
sekundaarimetaboliaan?
Kuinka kasvualustan kohotettu typpipitoisuus vaikuttaa rauduskoivun lehtien primaari-
ja sekundaarimetaboliaan?
Onko suhteellisella ilmankosteudella ja kasvualustan typpipitoisuudella yhdysvaiku-
tusta rauduskoivun lehtien primaari- ja sekundaarimetaboliaan?
Kuinka kohotettu suhteellinen ilmankosteus ja kasvualustan kohotettu typpipitoisuus
vaikuttavat koivuntaimien kasvuun kammiokokeen aikana?
Havaitaanko koivuntaimista määritettyjen kasvuparametrien ja koivunlehtien metabo-
liittipitoisuuksien muutosten välillä korrelaatiota?
4.1 Näytemateriaalin tuottaminen kammiokokeessa
Pro gradu -tutkimuksessa käytetty lehtinäytemateriaali oli peräisin heinä-elokuussa 2012 suori-
tetusta kammiokokeesta (Lihavainen ym. 2016a), johon en ole itse osallistunut. Kammiokoetta
varten mikrolisäyksellä tuotettuja Vehmersalmi 14 -genotyyppiä edustavia rauduskoivun
(Betula pendula Roth) taimia oli istutettu lannoitettuun turve-vermikuliitti -seokseen (1:1 v/v)
1,6 litran muoviruukkuihin. Kasvihuoneolosuhteissa kuuden kuukauden ikäisiksi kasvaneet
taimet siirrettiin ruukuissaan kasvihuoneelta kasvukammioon. Kammiokoe kesti 30 päivää,
4 AINEISTO JA MENETELMÄT

7
joista neljä päivää kului akklimaatioon eli kasvien totuttamiseen uusiin olosuhteisiin siirrettä-
essä ne kasvihuoneelta kasvukammioihin, ja 26 päivää varsinaiseen kokeeseen (Lihavainen ym.
2016a). Ennen koekäsittelyjen aloittamista taimista mitattiin rungon alkupituus ja alkupaksuus.
Kammiokokeessa 80 taimea oli jaettu neljään kammioon (20 taimea/kammio), joista kahteen
kammioon asetettiin suhteelliseksi ilmankosteudeksi 60 % ja toiseen kahteen kammioon
suhteellinen ilmankosteus 95 %. Tässä pro gradu -tutkielmassa käytän suhteellisen ilmankos-
teuden prosenttilukuja (lyhenteinä RH60 ja RH95) merkitsemään käsittelyjen eri ilmankos-
teustasoja. Kokeen aikana taimien paikkaa kammion sisällä vaihdettiin viikoittain, sekä yhden
kerran rinnakkaisesta kammiosta toiseen, jotta vältyttäisiin paikka- ja kammiovaikutukselta
(Lihavainen ym. 2016a).
Jokaisessa kammiossa olleista 20 koivuntaimesta kymmenelle taimelle annettiin viikoittain
peruslannoitetta [0,2 % Superex-9 -ravinneliuos (19:5:20 N:P:K sisältäen mikroravinteet,
Kekkilä, Suomi)] (typen saanto 15 mg typpeä/viikko) ja kymmenelle taimelle peruslannoitteen
lisäksi typpilisä (typen saanto 15+15 mg typpeä/viikko) 0,08 % ammoniumnitraattiliuoksena
(NH4NO3, JT Baker) (Lihavainen ym. 2016a). Lannoitustasot vastasivat vuotuista typpisaantoa
35 kg N/ha peruslannoituksella (40 taimea), mikä edustaa typen optimitasoa, ja 70 kg N/ha
typpilisällä (40 taimea), mikä edustaa typen ylimäärää. Tässä pro gradu -tutkielmassa käytän
vuotuisen typpisaannon lukuarvoja (lyhenteinä N35 ja N70) merkitsemään käsittelyjen eri
typpitasoja. Kokeen aikana annettu kokonaistyppimäärä per taimi oli 112 mg peruslannoituk-
sella ja 172 mg kasvualustan kohotetussa typpipitoisuudessa (Lihavainen ym. 2016a).
Kammioiden lämpötila (23 °C valossa/16 °C pimeässä) ja valojakson pituus (22 h valoa/2 h
pimeyttä) asetettiin vastaamaan koivuntaimien esikasvatuksessa käytettyjä kasvihuoneolo-
suhteita sekä vuodenajan mukaisia keskiarvoja (Lihavainen ym. 2016a). Valon maksimi-inten-
siteettinä käytettiin 450 µmol/m2/s, jolloin kaikki lamput olivat päällä. Tunti ennen ja jälkeen
pimeyttä puolet lampuista oli sammutettuina.
Kammiokokeen päätteeksi taimista mitattiin loppupituus ja loppupaksuus (Lihavainen ym.
2016a). Suhteellinen pituus- ja paksuuskasvu saatiin jakamalla loppuarvo ennen koetta mita-
tulla alkuarvolla (suhdeluku = loppuarvo
alkuarvo). Lehden typpipitoisuus (mg/g kuivapainoa kohti)
määritettiin Kjeldahlin menetelmällä (Lihavainen ym. 2016a). Lehden klorofylli-indeksi (CCI)
määritettiin kahden aallonpituuden (653 nm ja 931 nm) funktiona (Richardson ym. 2002): CCI
= transmittanssi 931 nm:ssa
transmittanssi 653 nm:ssa, mitä varten näiden aallonpituuksien optista absorbanssia mitattiin
kammiokokeen aikana kasvaneista lehdistä (jokaisen taimen viides lehti taimen kärjestä lukien)
CCM200 -klorofyllimetrillä (Opti-Sciences, Hudson, Yhdysvallat) (Lihavainen ym. 2016a).

8
Lehden pinta-ala (cm2) määritettiin yhdestä kokeen aikana kasvaneesta lehdestä per taimi
LAMINA -ohjelmistolla (Bylesjö ym. 2008) (Lihavainen ym. 2016a). Lehden vesipitoisuuden
määrittämiseksi taimista kerättiin yksi lehti, lehtien tuorepaino punnittiin ja lehdet kuivattiin 2
vuorokautta 50 °C:ssa, minkä jälkeen punnittiin lehtien kuivapaino (Lihavainen ym. 2016a).
Lehden vesipitoisuus (%-osuus tuorepainosta) laskettiin jakamalla lehtien tuorepainon (FW) ja
kuivapainon (DW) erotus tuorepainolla: FW−DW
FW ja kertomalla saatu lukuarvo 100 %:lla.
Metaboliatutkimusta varten kerättiin latvasta luettuna neljäs, viides ja kuudes lehti, joista
leikattiin halkaisijaltaan yhden senttimetrin (Ø 1 cm) kokoisia kiekkoja á 3 kpl/Eppendorf -
putki (Lihavainen ym. 2016a). Lehtikiekot laitettiin numeroimalla yksilöityihin, tyhjänä punnit-
tuihin putkiin. Kasvimateriaalia sisältävät näyteputket jäädytettiin nopeasti nestetypessä ja
säilytettiin jatkotutkimuksia varten -70 °C pakastimessa.
4.2 Metaboliatutkimus
Pro gradu -tutkimus suoritettiin Itä-Suomen yliopistossa Biologian laitoksella Metabolomiikka-
laboratoriossa kesä-heinäkuussa 2014. Metaboliatutkimuksiin varatut lehtikiekkoja sisältävät
näyteputket punnittiin jäisinä ennen tutkimuksen aloittamista, jolloin saatiin laskettua näyttei-
den tuorepaino. Näytteille tehtyjä työvaiheita olivat homogenisointi, uutto, derivatisointi ja
kaasukromatografis-massaspektrometrinen (GC-MS) analyysi. Uutettuja näytteitä valmistettiin
kaksi rinnakkaista sarjaa sekä erilliset varanäytteet. Näyteputket pidettiin homogenisoinnin ajan
nestetypessä ja uuton ajan jäähauteella kasvimateriaalin sulamisen estämiseksi ja esimerkiksi
entsymaattisten ja muiden ei-toivottujen kemiallisten reaktioiden välttämiseksi.
4.2.1 Homogenisointi
Syväjäädytettynä säilytetty kasvimateriaali homogenisoitiin Qiagen -kuulamyllyssä (Qiagen
TissueLyser, Retsch®) ravistellen (kuva 2a). Jokaiseen näyteputkeen lisättiin Qiagen -kuula-
pyssyllä Ø 5 mm metallikuula helpottamaan kasvimateriaalin murskaantumista (kuva 2b).
Näyteputkia ravisteltiin 3 kertaa 15 s ravistelunopeudella 15 ×/s, jäädyttäen putket ravistelujen
välillä nestetypessä. Viimeisen ravistelukerran jälkeen putkien annettiin olla nestetypessä niin
kauan, kunnes ensimmäinen uuttoliuos lisättiin näyteputkiin, jottei kasvimateriaali pääsisi sula-
maan.

9
Kuva 2. a) Homogenisointi Qiagen -kuulamyllyssä. b) Metallikuula näyteputken kannessa.
4.2.2 Uutto
Homogenisoitu kasvimateriaali uutettiin kahdesti, ensin 100 % metanolissa ja sitten 80 % meta-
nolissa. Uuton tarkoituksena oli saada metanoliin liukenevat sokeriyhdisteet ja veteen liuke-
nevat aminohapot erilleen homogenisoidusta kasvimateriaalista. Uuttovaiheessa näytteisiin
lisättiin myös sisäistä standardia, joka on seos yhdisteitä joita näyte ei luontaisesti sisällä ja
joiden kemialliset ominaisuudet ovat samanlaisia tunnistettavien yhdisteiden kanssa (Hill &
Roessner 2013). Sisäisellä standardilla normalisoidaan uutosta tuleva yhdisteiden pitoisuuksien
lievä vaihtelu (Fiehn ym. 2000), jolloin saadaan keskenään vertailukelpoisia tuloksia eli pysty-
tään havaitsemaan kammiokokeen ilmankosteus- ja typpikäsittelyjen vaikutukset rauduskoivun
lehtien metaboliittipitoisuuksien muutoksiin.
Sisäisenä standardina käytettiin yhdisteseosta, joka sisälsi tilavuussuhteessa 4:2:1:2 reagens-
seja bentsoehappo-d5 (1,2464 mg/ml dimetyylisulfoksidissa) (Benzoic Acid-d5, Campro
Scientific, Saksa ja Hollanti), alaniini-d4 (5,033 mg/ml 50 % metanolissa) (DL-Alanine-2,3,3,3-
d4, Isotec, Miamisburg, Yhdysvallat), glyseroli-d8 (4,900 mg/ml dimetyylisulfoksidissa)
(Glycerol-d8, Campro Scientific, Saksa ja Hollanti) sekä 4-metyyliumbelliferoni (10,32 mg/ml
100 % metanolissa) (4-methylumbelliferone, Sigma-Aldrich, Saksa ja Yhdysvallat). Reagenssit
punnittiin mikroanalyysivaa’alla ja liuotettiin sopivalla liuottimella 10 ml:n mittapulloissa.
Liuottimena käytetty dimetyylisulfoksidi oli hankittu VWR:lta (VWR International, Ranska) ja
metanoli Merck:lta (LiCrhosolv® Methanol for Liquid Chromatography, Merck, Saksa).
Uuttoliuosten valmistamiseen käytettiin suodatettua, ultrapuhdasta milliporevettä (Simplicity®
UV, Millipore, Molsheim, Ranska). Standardiseosta säilytettiin uuttopäivien välillä pakastet-
tuna -20 °C:ssa. Standardiseos sulatettiin vesihauteessa ja vorteksoitiin aina ennen käyttöä.
Homogenisoituihin näytteisiin pipetoitiin ensimmäisessä uutossa 1 ml 100 % metanolia sekä
100 µl sisäistä standardiseosta, minkä jälkeen näyteputkia vorteksoitiin. Lisäksi valmistettiin
jokaisena uuttopäivänä niin kutsuttu näyteblankki (SB), joka sisälsi vain uuttoliuokset sekä
a) b)

10
sisäisen standardin. Näin saataisiin poistettua uuttoliuosten mahdollisesti aiheuttama tausta
massaspektristä. Uuttoliuoksen ja homogenisoidun kasvimateriaalin sekoittumisen varmista-
miseksi ja uuton edistämiseksi suoritettiin Qiagen -kuulamyllyssä ravistelu 1 × 15 s ravistelu-
nopeudella 15 ×/s. Ensimmäinen uutto suoritettiin Eppendorf Thermomixer:lla (Thermomixer
comfort, Eppendorf, Saksa) 15 min 6 °C 1400 rpm (kuva 3a), minkä jälkeen näyteputket sentri-
fugoitiin ultrasentrifugilla (Jouan MR23i, Thermo Electron Corporation, Milford, Yhdysvallat)
3 min 10 °C 13 000 rpm (kuva 3b). Supernatantti (kuva 4a) pipetoitiin sakan päältä ja otettiin
talteen 2 ml:n Eppendorf -putkeen. Supernatanttiputkia säilytettiin uuttovaiheiden välissä jäillä
Eppendorf -telineessä.
Kuva 3. a) Uutto Eppendorf Thermomixer:ssä. b) Uutetut näytteet ultrasentrifugissa.
Toisen uuton tarkoituksena oli maksimoida ja varmistaa haluttavien yhdisteiden, erityisesti
vesiliukoisten aminohappojen uuttuminen homogenaatista. Ensimmäisestä uutosta jääneen
sakan päälle pipetoitiin toista uuttoa varten 1 ml 80 % metanolia. Näytteet vorteksoitiin ja ravis-
teltiin Qiagen -kuulamyllyssä 1 × 15 s ravistelunopeudella 15 ×/s. Toinen uutto suoritettiin
Eppendorf Thermomixer:llä 5 min 6 °C 1400 rpm. Uutokset ultrasentrifugoitiin kuten ensim-
mäisessä uutossa, 3 min 10 °C 13 000 rpm. Erottunut supernatantti (kuva 4b) pipetoitiin talteen
ja yhdistettiin ensimmäisestä uutosta saadun supernatantin kanssa. Yhdistetyt supernatantit
vorteksoitiin tasakoostumuksellisen liuoksen aikaansaamiseksi. Kunkin näytteen yhdistetystä
supernatantista pipetoitiin 2 × 100 µl näytteet vialeissa oleviin insertteihin (kuva 5). Viali on
kaasukromatografiassa käytettävä lasinen 2 ml:n näytepullo. Insertti on vialin sisään laitettava
200 µl:n asetin, jonne tutkittava liuos pipetoidaan. Vialit suljettiin Ø 11 mm kumitulpallisilla
alumiinikorkeilla käyttäen korkinsulkijaa, jolloin vialista tulee ilmatiivis. Varsinainen näyte-
sarja pipetoitiin ruskeisiin vialeihin ja rinnakkainen näytesarja kirkkaisiin vialeihin (kuva 5).
a) b)

11
Kuva 4. Sentrifugoitu näyte a) ensimmäisen uuton jälkeen, b) toisen uuton jälkeen.
Kuva 5. Supernatantin pipetoiminen vialissa olevaan inserttiin.
Ensimmäisenä uuttopäivänä tehtiin myös laadunvarmistusnäyte (QC), mitä varten kunkin
näytteen yhdistetystä supernatantista pipetoitiin 100 µl erilliseen 2 ml Eppendorf -näyteput-
keen. Tällöin saatiin luotua kaikkia käsittelyjä edustava 16 näytteen (4 näytettä/käsittely) super-
natanttien seos, jolla voidaan varmentaa jokaisena ajopäivänä suoritetun derivatisoinnin ja GC-
MS -analyysin laatua sekä GC-MS -analyysistä saatavien tulosten tarkkuutta ja luotettavuutta
sillä oletuksella, että QC-näytteet edustavat likimain koko näyteaineiston keskiarvoja. QC -
vialeja tehtiin yhteensä kahdeksan, jotta jokaiselle ajopäivälle riittäisi varmasti oma laadun-
varmistusnäyte.
Uutetut näytteet haihdutettiin kuiviin vakuumikonsentraattorilla 30 min 35 °C (vakuu-
misentrifugi SPD121P ja vakuumipumppu OFP400, Thermo Electron Corporation, Milford,
Yhdysvallat). Kuivatuksen jälkeen näytteet altistettiin typpikaasuvirtaukselle 1 min ajan,
jolloin typpikaasu syrjäyttää insertin ilmatilassa olevan hapen. Tällä toimenpiteellä pyrittiin
estämään uutetun näytteen reagoiminen hapen kanssa. Näytevialit korkitettiin mahdollisimman
nopeasti typpialtistuksen jälkeen, aseteltiin pakastusrasioihin ja vietiin -70 °C pakastimeen.
Supernatanteista pipetoitiin lisäksi 500 µl varanäytteet 1,5 ml:n Eppendorf -näyteputkiin.
Varanäytteet haihdutettiin kuiviin vakuumikonsentraattorilla 120 min 35 °C, altistettiin typpi-
kaasuvirtaukselle 1 min ajan ja vietiin korkit suljettuina -70 °C pakastimeen. Jäljelle jäänyt
supernatantti pakastettiin sellaisenaan muita julkaisuja varten tehtäviä tutkimuksia ajatellen.
a) b)

12
4.2.3 Derivatisointi
Uutetuille näytteille suoritettiin kaksivaiheinen derivatisointi, minkä avulla kasvimateriaalista
uuttuneet orgaaniset yhdisteet pyrittiin saamaan kaasuuntuvaan muotoon ja muokkaantumaan
metoksi- ja silyylijohdannaisiksi, jotta yhdisteet voitaisiin havaita massaspektrissä. Derivati-
soinnin tarkoituksena on lisäksi vähentää yhdisteissä esiintyvien hydroksyyli-, amino- ja
karboksyyliryhmien polaarisuutta (Hill & Roessner 2013). Derivatisointia edeltävästi huoneen-
lämmössä vialitelineellä yhden tunnin ajan sulatettuihin näytteisiin pipetoitiin 50 µl dikloori-
metaania (DCM for HPLC, Sigma-Aldrich, Steinheim, Saksa) eVol® XR -automaattipipetillä
(SGE Analytical Science, Australia) sulatuksen aikana insertteihin mahdollisesti kondensoitu-
neen veden poistamiseksi. Lisätty dikloorimetaani ja mahdollinen kondensaatiovesi haihdu-
tettiin kuiviin vakuumikonsentraattorissa 5 min 35 °C.
Ensimmäisessä vaiheessa derivatisointireagenssina käytettiin pyridiiniin (Sigma-Aldrich,
Steinheim, Saksa) liuotettua metoksiamiinihydrokloridia (20 mg/ml) (MAHC, Sigma-Aldrich,
Steinheim, Saksa; valmistettu Sveitsissä), jota valmistettiin tuore liuos jokaisena derivatisointi-
päivänä. MAHC:ta punnittiin analyysivaa’alla 4 ml:n ruskeaan lasipulloon 20 mg ja liuotettiin
se 1000 µl pyridiiniin. Jokaiseen näytteeseen pipetoitiin 40 µl MAHC-pyridiini -liuosta ja vialit
korkitettiin välittömästi pipetoinnin jälkeen liuoksen haihtumisen estämiseksi. Ensimmäisen
vaiheen derivatisointi suoritettiin 180 min 30 °C lämpöblokissa 170 rpm tasaisessa ravistelussa.
Toisessa vaiheessa derivatisointireagenssina käytettiin N-metyyli-N-trimetyylisilyyli-
trifluoroasetamidi + 1 % trimetyyliklorosilaania (MSTFA + 1 % TMCS, ThermoScientific,
Bellefonte, Yhdysvallat), jota pipetoitiin näytteisiin 80 µl. Samanaikaisesti derivatisointi-
reagenssin kanssa näytteisiin pipetoitiin 20 µl retentioaikastandardina käytettyä alkaanisarjaa
[saturoituja alkaaneja C7-C40 (Supelco, Bellefonte, Yhdysvallat) heksaanissa (Merck, Saksa)].
Alkaanisarjan avulla lasketaan kaikkien analysoitavien piikkien retentioindeksit, mikä on
välttämätöntä yhdisteiden tunnistamiselle. Näytteiden derivatisoinnin yhteydessä valmistettiin
samanaikaisesti niin kutsuttu reagenssiblankki (RB), joka sisälsi vain derivatisointireagenssit.
Tämän avulla poistetaan derivatisointireagenssien aiheuttama tausta GC-MS -kromatogram-
mista. Toisen vaiheen derivatisointi suoritettiin 120 min 60 °C lämpöblokissa 170 rpm tasai-
sessa ravistelussa. Derivatisoinnin jälkeen näytevialit poimittiin vialitelineelle ja aseteltiin
satunnaisessa järjestyksessä GC-MS -laitteiston (kuva 6) näyteautomaattiin.

13
4.2.4 Kaasukromatografis-massaspektrometrinen analysointi
Kaasukromatografi yhdistettynä massaspektrometriin (GC-MS) sopii laitteistona hyvin sellais-
ten metaboliittien analysoitiin, joiden molekyylipaino, poolisuus ja kiehumispiste ovat alhaiset,
ja jotka ovat herkästi haihtuvia tai derivatisoitavissa herkästi haihtuviksi (Hill & Roessner 2013,
Xue ym. 2015). Tällaisia yhdisteitä ovat erityisesti primaarimetaboliitit, kuten muun muassa
aminohapot, sokerit ja orgaaniset hapot (Hill & Roessner 2013). GC-MS on yksi eniten käyte-
tyistä analyyttisistä menetelmistä kasvimetabolomiikassa sen avulla saatavan korkean
resoluution, korkean herkkyyden (sensibility), hyvän toistettavuuden (reproducibility), standar-
dispektrikirjastojen laajuuden sekä suhteellisen alhaisten kustannusten vuoksi (Xue ym. 2015).
Derivatisoidut näytteet analysoitiin tietokoneohjatulla GC-MS -laitteistolla [(Agilent 6890N
kaasukromatografi, 5973 massaspektrometri, 7683 injektori ja näyteautomaatti (Agilent
Technologies, Palo Alto, Yhdysvallat), kuva 6]. GC-MS -laitteistoa ohjaa ChemStation -ajo-
ohjelmisto (MSD ChemStation E.02.01.1177, Agilent Technologies, Palo Alto, Yhdysvallat),
joka muun muassa säätelee GC-MS -laitteiston eri osien lämpötiloja ja kontrolloi ajoa. Kroma-
togrammeja ja piikkien massaspektrejä voidaan tarkastella ajo-ohjelmistoon liittyvällä
Enhanced Data Analysis -ohjelmalla.
Ajoparametreinä käytettiin räätälöityä metodia VIRVESplit30.M, jossa näytettä injektoitiin
1 µl split-suhteella 30:1. Tällöin injektoidusta näytteestä yksi kolmaskymmenesosa (1/30)
kulkee kantajakaasun mukana kolonnin läpi detektoitavaksi massaspektillä. Kantajakaasuna
käytettiin heliumia (H2), jonka virtaus kolonnin läpi oli 1.0 ml/min. Kolonnina oli Restek, USA
Rxi®-5Sil MS 30 m, 0.25 mmID, 0.25 µm df, jossa on 10 m Integra-esikolonni. Linerina oli
Restek Split Precision® deaktivoitu liner 4 mm x 6.3 x 78.5. Injektointilämpötilana käytettiin
260 ºC:ta. Lämpötilaohjelma oli seuraava: 1 min isoterminen 70 ºC, 6 ºC/min nosto 330 ºC:een,
jossa pito 5 min. Jäähdytys 70 ºC:een, jossa pito 3 min (postrun). 7 min tasapainotus 70 ºC:ssa
ennen seuraavaa ajoa. MSD interface (AUX) 290 ºC, MS lähde 230 ºC, MS quad 150 ºC.
Massa-alue (scanning range) m/z 55 - 550, joka 35 minuutin jälkeen nostettiin m/z 55 - 650.
Massasignaalin kerääminen aloitettiin 5,5 min viiveellä, jotta liuottimet ehtivät kulkea kolon-
nista massadetektorille ja sen ohi. Ajopäivien välissä injektointiruisku (Agilent 10 µl) pestiin
pyridiini - heksaani - metanoli - asetoni -liuotinsarjalla. Injektorilla (kuva 6c) ruiskun neulaosan
pesussa käytettiin pyridiiniä ja heksaania.

14
Kuva 6. GS-MS -analyysilaitteisto, Agilent: a) 6890N kaasukromatografi, b) 5973 massaspektrometri,
c) 7683 injektori ja d) näyteautomaatti; massaspektrometrin ionilähde (ylhäällä vasemmalla) ja kaasu-
kromatografin kolonniuuni (ylhäällä oikealla).
4.2.5 Yhdisteiden identifiointi ja kvantifiointi
GC-MS -analysoitujen näytteiden sisältämät metaboliitit tunnistettiin ja kvantifioitiin kohden-
netusti kvantifiointi-ionin, retentioindeksin (Liite 1: taulukko L1) ja massaspektrin perusteella
sekä vertaamalla yhdistespektrejä ja retentioindeksejä NIST Mass Spectral Database- (version
2.2, Agilent Technologies, National Institute of Standards and Technology, USA) ja Golm
Metabolome Database (Kopka ym. 2004, Schauer ym. 2005, Hummel ym. 2010) -tietokantoi-
hin. Metaboliittien tunnistamiseen ja piikkien pinta-alojen laskemiseen käytettiin
MetaboliteDetector 2.0 -ohjelmistoa (Hiller ym. 2009) erillisellä koivukammiokirjastolla
(Lib_chamber_birch, J. Lihavainen). Piikkejä etsittiin niiden korkeuden ja dekonvoluutiole-
veyden erilaisilla yhdistelmillä. Dekonvoluutio erottelee yksittäiset piikit tilanteessa, jossa
saman retentioajan sisällä tulee monta yhdistettä päällekkäin (Hiller ym. 2009, Chen & Dai
2015). MetaboliteDetector 2.0 -ohjelmistolla suoritetun kvantifioinnin tulokset koottiin Micro-
soft Office Excel 2013 -taulukkopohjaan (Microsoft, Seattle, Yhdysvallat). Yhdisteen
a)
b)
c)
d)

15
pitoisuuden normalisoitu arvo saatiin jakamalla yhdisteen piikin pinta-ala sisäisen standardin
piikin pinta-alalla (bentsoehappo-d5) ja näytteen tuorepainolla. Tällöin saatiin vertailukelpoisia
tuloksia eli pystyttiin havaitsemaan kammiokokeen ilmankosteus- ja typpikäsittelyjen vaiku-
tukset rauduskoivun lehtien metaboliittipitoisuuksiin. Glutamiinilla, fruktoosilla, glukoosilla ja
sedoheptuloosilla havaittiin kaksi lähekkäisillä retentioajoilla eluoituvaa saman yhdisteen
piikkiä, joten näiden metaboliittien kohdalla pitoisuusarvona käytettiin kahden kvantifioidun
piikin pinta-alojen yhteenlaskettua arvoa (Liite 1: taulukko L1). Kolme näytettä jätettiin tilas-
tollisen analyysin ulkopuolelle arvojen normalisoinnin yhteydessä havaitun sisäisen standardin
vähäisen pitoisuuden (kaksi näytettä) tai alhaisen tuorepainon (yksi näyte) vuoksi.
Tilastollisesti analysoitava aineisto koostui lehtinäytteistä (n = 41), jotka oli kerätty
alemmassa suhteellisessa ilmankosteudessa ja kasvualustan optimaalisessa typpipitoisuudessa
kasvatetuista taimista, joita käytettiin vertailukohtana muihin käsittelyihin näiden olosuhteiden
ollessa samankaltaiset kasvihuoneolosuhteiden kanssa (näytteet C = RH60 N35, n = 10), koho-
tetussa suhteellisessa ilmankosteudessa ja kasvualustan optimaalisessa typpipitoisuudessa
kasvatetuista taimista (näytteet H = RH95 N35, n = 10), alemmassa suhteellisessa ilmankosteu-
dessa ja kasvualustan kohotetussa typpipitoisuudessa kasvatetuista koivuista (näytteet N =
RH60 N70, n = 11) sekä kohotetussa suhteellisessa ilmankosteudessa ja kasvualustan kohote-
tussa typpipitoisuudessa kasvatetuista taimista (näytteet HN = RH95 N70, n = 10). Metabo-
liittien havaintoyksiköiden lukumäärä (n) käsittelyittäin on taulukoitu tutkielman liitteeksi
(Liite 1: taulukko L1).
4.2.6 Tilastollinen analyysi
Aineiston tarkkuutta, luotettavuutta ja vertailukelpoisuutta voidaan tarkastella tilastollisin
menetelmin. Pro gradu -tutkimuksessa GC-MS:lla analysoidulle metaboliittiaineistolle suori-
tettiin pääkomponenttianalyysi (PCA) SIMCA-P+ 12.0.1 -ohjelmistolla (Umetrics, Uumaja,
Ruotsi) aineiston sisäisen vaihtelun ja hajonnan tarkastelemiseksi. SPSS 21 -ohjelmistolla (IBM
SPSS Statistics, IBM, Yhdysvallat) laskettiin metaboliitti- ja kasvuparametriaineiston muuttu-
jakohtaiset keskiarvot ja keskivirheet. Kaksisuuntaisella ANOVA -varianssianalyysillä saatiin
selville käsittelyvaikutusten merkitsevyysasteet. Näitä vaikutuksia havainnollistettiin BoxPlot
-kuvaajilla sekä suoritettiin metaboliitti- ja kasvuparametriaineiston välinen parametrinen
Pearson-korrelaatioanalyysi.
PCA on ohjaamaton (unsupervised) monimuuttujamenetelmä, jolla pelkistetään monimut-
kaisen aineiston vaihtelu muutamalle pääkomponentille sekä etsitään suurimman vaihtelun

16
sijoittumista aineistossa (Worley & Powers 2013). PCA:ta varten osalle metaboliiteista tehtiin
SIMCA-P+ -ohjelmistolla pitoisuusarvojen kymmenkantainen logaritmikorjaus (log10) jakau-
mien normalisoimiseksi. SIMCA-P+ -ohjelmisto tekee aineistolle automaattisesti UV -
skaalauksen, jossa kunkin muuttujan arvojen keskiarvo asettuu arvoon 0 (mean centering) ja
keskihajonta asettuu arvoon 1 (unit variance). PCA:n score plot havainnollistaa, kuinka eri
näytteiden välinen kokonaishajonta asettuu PCA-ordinaatioon suhteessa pääkomponentti-
akseleihin (Worley & Powers 2013). Pääkomponentit muodostetaan niin, että aineiston moni-
ulotteisuuden vähentämiseksi ohjelmisto pyrkii sisällyttämään mahdollisimman suuren
osuuden aineiston vaihtelusta ensimmäiseen pääkomponenttiin (Ranta ym. 1994, Zar 1999).
Sitä vastaan kohtisuoraan sijoittuva toinen pääkomponentti sisältää mahdollisimman suuren
osuuden aineiston jäännösvaihtelusta, jota ensimmäinen pääkomponentti ei selitä. Kaukana
origosta sijaitsevat pisteet edustavat aineiston keskiarvosta eniten poikkeavia havaintoja.
Metaboliitti- ja kasvuparametriaineistolle laskettiin SPSS 21 -ohjelmistolla käsittelykoh-
tainen näytelukumäärä (n), tutkittavien metaboliittien pitoisuuksien ja kasvuparametrien
keskiarvo (ka.) kussakin käsittelyryhmässä sekä keskiarvojen keskivirheet (SE). Suhteellisen
ilmankosteuden (RH) ja kasvualustan typpipitoisuuden (N) pää- ja yhdysvaikutuksia (RH×N)
metaboliittipitoisuuksiin ja kasvuparametreihin testattiin kaksisuuntaisella varianssianalyysillä
(ANOVA eli Univariate Analysis of Variance). ANOVA:n kiinteinä tekijöinä (fixed factor)
käytettiin suhteellisen ilmankosteuden ja kasvualustan typpipitoisuuden kahta eri tasoa (RH60
ja RH95 sekä N35 ja N70). ANOVA testaa aineistoa vertaamalla suhteellisen ilmankosteuden
tasoja toisiinsa (näytteet C ja N verrattuna näytteisiin H ja HN) ja etsii näiden kahden tason
väliltä merkitseviä eroavaisuuksia tarkasteltavissa muuttujissa, jolloin havaitaan mahdollinen
suhteellisen ilmankosteuden päävaikutus kussakin muuttujassa. Vastaavasti ANOVA testaa
aineistoa vertaamalla kasvualustan typpitasoja toisiinsa (näytteet C ja H verrattuna näytteisiin
N ja HN) ja etsii näiden kahden tason väliltä merkitseviä eroavaisuuksia tarkasteltavissa
muuttujissa, jolloin havaitaan mahdollinen kasvualustan typpipitoisuuden päävaikutus kussakin
muuttujassa. Lisäksi ANOVA testaa kiinteiden tekijöiden yhdysvaikutusta toisiinsa. Mikäli
kiinteiden tekijöiden yhdysvaikutus on merkitsevä, tällöin kohotetun suhteellisen ilmankosteu-
den vaikutus muuttujiin olisi erisuuntainen eri typpitasoilla sekä kasvualustan kohotetun typpi-
pitoisuuden vaikutus muuttujiin olisi erisuuntainen eri ilmankosteustasoilla. Mikäli kiinteiden
tekijöiden yhdysvaikutus ei ole merkitsevä, tällöin tekijän mahdollinen päävaikutus määrittää
tuloksia eli esimerkiksi kohotettu suhteellinen ilmankosteus vaikuttaa muuttujaan samalla
tavalla kasvualustan molemmilla typpitasoilla.

17
Niille metaboliiteille, joille oli PCA:ta varten tehty kymmenkantainen logaritmimuunnos
(log10) jakauman normalisoimiseksi, käytettiin ANOVA:ssa samoja logaritmimuunnettuja
arvoja. Kasvuparametriaineistosta havaittiin Kolmogorov-Smirnovin sekä Shapiro-Wilkin
testillä, etteivät suhteellisen pituuskasvun arvot täyttäneet normaalijakaumaoletuksia kohotetun
suhteellisen ilmankosteuden ja kasvualustan alhaisemman typpipitoisuuden osalta, joten
suhteellisen pituuskasvun arvoille tehtiin log10 -muunnos ANOVA:a varten. Muunnoksen
jälkeen suhteellisen pituuskasvun arvot poikkesivat edelleen normaalijakaumasta kasvualustan
alemman typpipitoisuuden osalta, mutta enempää muunnoksia ei tehty.
Tarkasteltuihin metaboliitteihin liittyvillä BoxPlot -kuvaajilla (kuvat 10-17) havainnollis-
tettiin käsittelyjen vaikutuksia metaboliittipitoisuuksiin sekä pitoisuusarvojen hajontaa käsitte-
lyryhmien välillä. Vastaavasti kasvuparametriaineistoon liittyvillä BoxPlot -kuvaajilla (kuvat
18-20) havainnollistettiin käsittelyjen vaikutuksia kyseisiin kasvuparametreihin sekä arvojen
hajontaa käsittelyryhmien välillä. Arvoille ei tehty kymmenkantaista logaritmikorjausta (log10)
BoxPlot -kuvaajia varten, joten nämä kuvat edustavat muuttujien luonnollista jakaumaa.
SPSS-ohjelmisto asettaa havaintojoukon arvot suuruusjärjestykseen, minkä jälkeen BoxPlot -
kuvaajat muodostuvat seuraavin periaattein (Disraeli 1996, kuva 7): Pylvään sisällä oleva
vaakaviiva osoittaa havaintojoukon mediaaniarvon (Q2). Pylväs sisältää keskimmäiset 50 %
havaintojoukon arvoista. Pylvään ylä- ja alapuolella olevat pystyviivat osoittavat havainto-
arvoja, jotka ovat ylimmässä tai alimmassa 25 %:ssa tapauksista niissä havaintojoukoissa joissa
ei ole ääriarvoja. Q4 on tällöin havaintojoukon maksimiarvo ja Q0 havaintojoukon minimiarvo.
Kvartiiliväli (IQR) on yläkvartiilin korkeimman arvon (Q3) ja alakvartiilin alimman arvon (Q1)
erotus [IQR = Q3 - Q1]. Ympyrällä (o) merkitty läheinen ääriarvo on arvoltaan suurempi kuin
[Q1 - 1.5 x IQR] tai [Q3 + 1.5 x IQR]. Tähdellä () merkitty kaukainen ääriarvo on arvoltaan
suurempi kuin [Q1 - 3 x IQR] tai [Q3 + 3 x IQR].
Lineaarisia riippuvuuksia kammiokokeessa kasvetuista rauduskoivuista määritettyjen
kasvuominaisuuksien (suhteellinen pituus- ja paksuuskasvu) ja lehtinäytteiden ominaisuuksien
(näytteen tuorepaino, lehden pinta-ala, vesipitoisuus, typpipitoisuus ja klorofylli-indeksi)
(Lihavainen ym. 2016a) sekä tässä pro gradu -tutkimuksessa määritettyjen metaboliittipitoi-
suuksien välillä tarkasteltiin Pearson-korrelaation avulla. Pearson-korrelaatio kuvaa kahden
muuttujan välisen keskinäisen lineaarisen riippuvuuden voimakkuutta (Ranta ym. 1994, Zar
1999). Korrelaatiokerroin voi saada arvoja välillä -1…0...+1. Positiivinen korrelaatio osoittaa
samansuuntaista lineaarista riippuvuutta, negatiivinen korrelaatio puolestaan osoittaa vastak-
kaissuuntaista lineaarista riippuvuutta. Kahden muuttujan välillä voidaan sanoa olevan
voimakas lineaarinen riippuvuus, kun korrelaatiokertoimen arvo on suurempi kuin ± 0,7.

18
Korrelaatiokertoimen arvoilla ± 0,7…0,3 muuttujien välillä on kohtalainen lineaarinen riippu-
vuus. Korrelaatiokertoimen arvon lähestyessä nollaa, muuttujien välillä oleva lineaarinen
riippuvuus on hyvin vähäistä. Havaintoyksiköiden lukumäärä (n) vaikuttaa sekä korrelaatio-
kertoimen arvoon että korrelaation merkitsevyyteen eli p-arvoon.
Kuva 7. BoxPlot-kuvaajan tulkinta. Havaintojoukon minimiarvo (Q0), alakvartiili (Q1), mediaani (Q2),
yläkvartiili (Q3), maksimiarvo (Q4) sekä ääriarvot (o, ).
5 TULOKSET
Tässä pro gradu -tutkielmassa tarkastellaan suhteellisen ilmankosteuden ja kasvualustan typpi-
pitoisuuden kahden eri tason vaikutuksia rauduskoivun (Betula pendula Roth) taimien lehti-
näytteistä (n = 41) GC-MS:lla analysoitujen ja niistä identifioitujen metaboliittien (n = 36)
pitoisuuksiin (Liite 1: taulukko L1). Näihin metaboliitteihin lukeutui muiden muassa sokereita,
sokerialkoholeja, orgaanisia happoja, aminohappoja ja fenolisia yhdisteitä, joita löytyi suurim-
masta osasta näytteitä, ja ne edustavat keskeisiin metaboliareitteihin (kuva 1) liittyviä yhdiste-
ryhmiä. Tutkielmassa tarkastellaan lisäksi koekäsittelyjen vaikutuksia samoista taimista mitat-
tuihin kasvuparametreihin ja lehtien ominaisuuksiin.
5.1 Näytteiden sijoittuminen pääkomponenttianalyysissa
Pääkomponenttianalyysilla pyrittiin selvittämään metaboliittiaineiston sisäistä vaihtelua sekä
löytämään eroja koekäsittelyjen välillä sen suhteen, määrittääkö tuloksia enemmän kohotettu
suhteellinen ilmankosteus vai kasvualustan kohotettu typpipitoisuus. Ensimmäinen pääkompo-
nentti [t(1)] selitti aineiston kokonaisvaihtelusta 26,9 % ja toinen pääkomponentti [t(2)] 19,1 %
(kuva 8). Kolmannen pääkomponentin selitysaste oli 8,6 % ja neljännen pääkomponentin 6,5

19
%, joten melko vähäisinä ja vaikeasti tulkittavina ne jätettiin pois tarkastelusta. Käsittelyryhmät
eivät selvästi erottuneet toisistaan PCA-ordinaatiossa ensimmäisen ja toisen pääkomponentin
suhteen. Kun ensimmäinen ja toinen pääkomponentti yhdistettiin (kuvien 8 ja 9 diagonaalinen
katkoviiva), saatiin uusi tarkastelutaso, jolloin havaittiin näytteiden erottuvan suhteellisen
ilmankosteuden suhteen (kuva 8). Kohotetussa suhteellisessa ilmankosteudessa kasvatetut
näytteet (H ja HN) ryhmittyivät yhteen ja erottuivat suurimmasta osasta alemmassa suhteelli-
sessa ilmankosteudessa kasvatetuista lehtinäytteistä (C ja N) (kuva 8). Kasvualustan
kohotetussa typpipitoisuudessa kasvatetut lehtinäytteet (N ja HN) eivät erottuneet alemmassa
typpipitoisuudessa kasvatetuista näytteistä (C ja H), eli voidaan todeta, ettei kasvualustan
typpipitoisuus määrittänyt tuloksia (kuva 8).
Tulosten ja analyysimenetelmien luotettavuutta osoittaa laadunvarmistusnäytteiden (QC)
toisiinsa nähden läheinen sijoittuminen pääkomponenttianalyysissä sekä keskittyminen PCA-
ordinaatiossa lähelle origoa (kuva 8). Tällöin QC -näytteiden välinen vaihtelu on vähäistä ja
näin ollen voidaan sanoa tulosten olevan tarkkoja ja luotettavia.
5.2 Kohotetun suhteellisen ilmankosteuden vaikutus metaboliitteihin
Metaboliittipitoisuuksien muutosta suhteessa ilmankosteuskäsittelyihin voitiin tarkastella
yhdistämällä analysoidut metaboliitit samaan kuvaan tutkittujen näytteiden kanssa niin kutsu-
tuksi Loadings Bi Plot:iksi (kuva 9). Tästä kuvasta sekä myöhemmistä analyysivaiheista on
poistettu QC -näytteet, joten selitysasteet muuttuivat hieman verrattuna PCA:n tuloskuvaajaan
(kuva 8), ollen ensimmäisellä pääkomponentilla 27,3 % ja toisella pääkomponentilla 19,8 %
(kuva 9). Metaboliitit, joiden pitoisuus aleni kohotetussa suhteellisessa ilmankosteudessa
kasvatetuissa lehdissä, ryhmittyivät alemmassa suhteellisessa ilmankosteudessa kasvatettuja
lehtinäytteitä (C ja N) edustavalle puolelle PCA-ordinaatiota (kuva 9 yläoikea puolisko).
Metaboliitit, joiden pitoisuus nousi kohotetussa suhteellisessa ilmankosteudessa kasvatetuissa
lehdissä, ryhmittyivät kohotetussa suhteellisessa ilmankosteudessa kasvatettuja lehtinäytteitä
(H ja HN) edustavalle puolelle PCA-ordinaatiota (kuva 9 alavasen puolisko). Metaboliitit,
joiden pitoisuus ei juurikaan muuttunut käsittelyjen vaikutuksesta, sijoittuivat PCA-ordinaa-
tiossa lähelle origoa (kuva 9).
SPSS:n kaksisuuntaisella ANOVA:lla tarkasteltuna 36 tutkitusta metaboliitista yhteensä 14
metaboliitin pitoisuus aleni merkitsevästi rauduskoivun lehdissä kohotetussa suhteellisessa
ilmankosteudessa kasvatettuna verrattuna alemmassa suhteellisessa ilmankosteudessa kasva-
tukseen (taulukko 1). Nämä merkitsevyydet on merkitty vastaavasti myös metaboliittiaineis-

20
tosta laadittuihin BoxPlot -kuvaajiin (kuvat 10-17). Tällaisia metaboliitteja olivat glukoosi
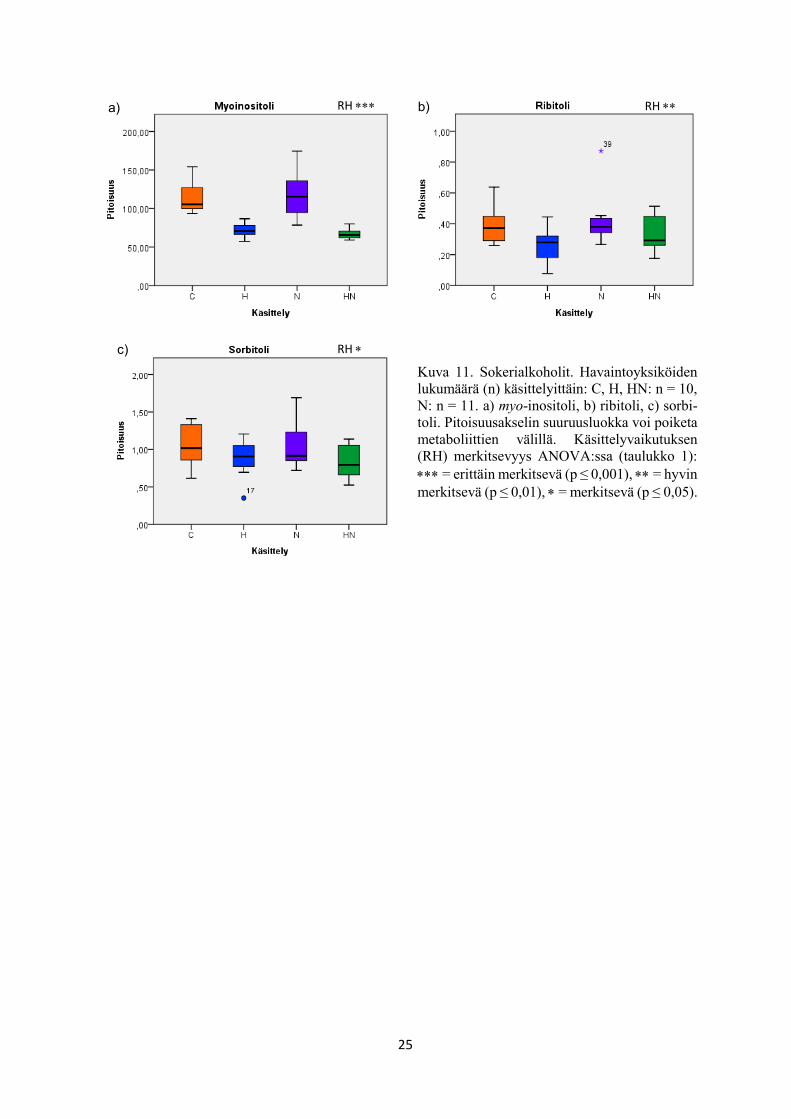
(taulukko 1, kuva 10c), sakkaroosi (taulukko 1, kuva 10f), myo-inositoli (taulukko 1, kuva 11a),
ribitoli (taulukko 1, kuva 11b), sorbitoli (taulukko 1, kuva 11c), malaatti (taulukko 1, kuva 12c),
sikimihappo (taulukko 1, kuva 12d), sitruunahappo (taulukko 1, kuva 12e), sukkinihappo
(taulukko 1, kuva 12f), treonihappo (taulukko 1, kuva 12g), treonihappo-1,4-laktoni (taulukko
1, kuva 12h), treoniini (taulukko 1, kuva 13e), β-sitosteroli (taulukko 1, kuva 14a) ja salidrosidi
(taulukko 1, kuva 16b).
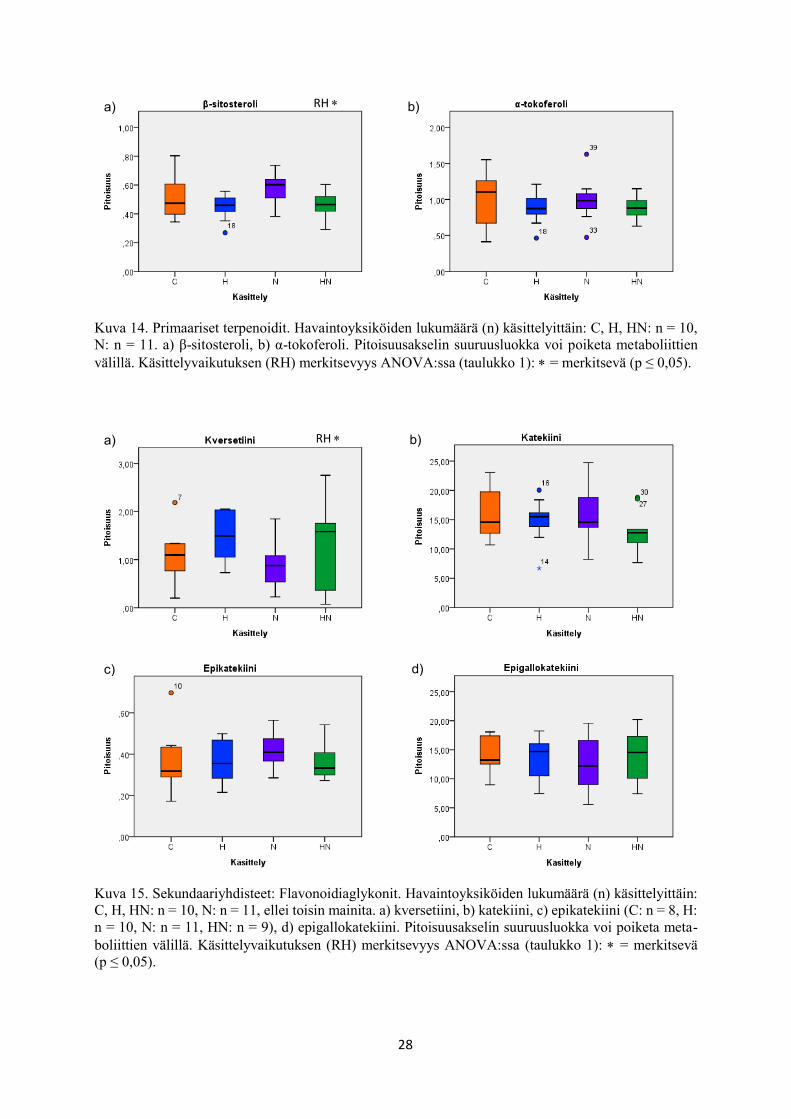
Kolmen metaboliitin pitoisuus nousi merkitsevästi rauduskoivun lehdissä kohotetussa
suhteellisessa ilmankosteudessa kasvatettuna verrattuna alemmassa suhteellisessa ilmankosteu-
dessa kasvatukseen. Tällaisia olivat raffinoosi (taulukko 1, kuva 10e), kversetiini (taulukko 1,
kuva 15a) ja flavonoliglykosidi (taulukko 1, kuva 16a).
5.3 Kasvualustan kohotetun typpipitoisuuden vaikutus metaboliitteihin
SPSS:n kaksisuuntaisella ANOVA:lla (taulukko 1) tarkasteltuna malaatin (taulukko 1, kuva
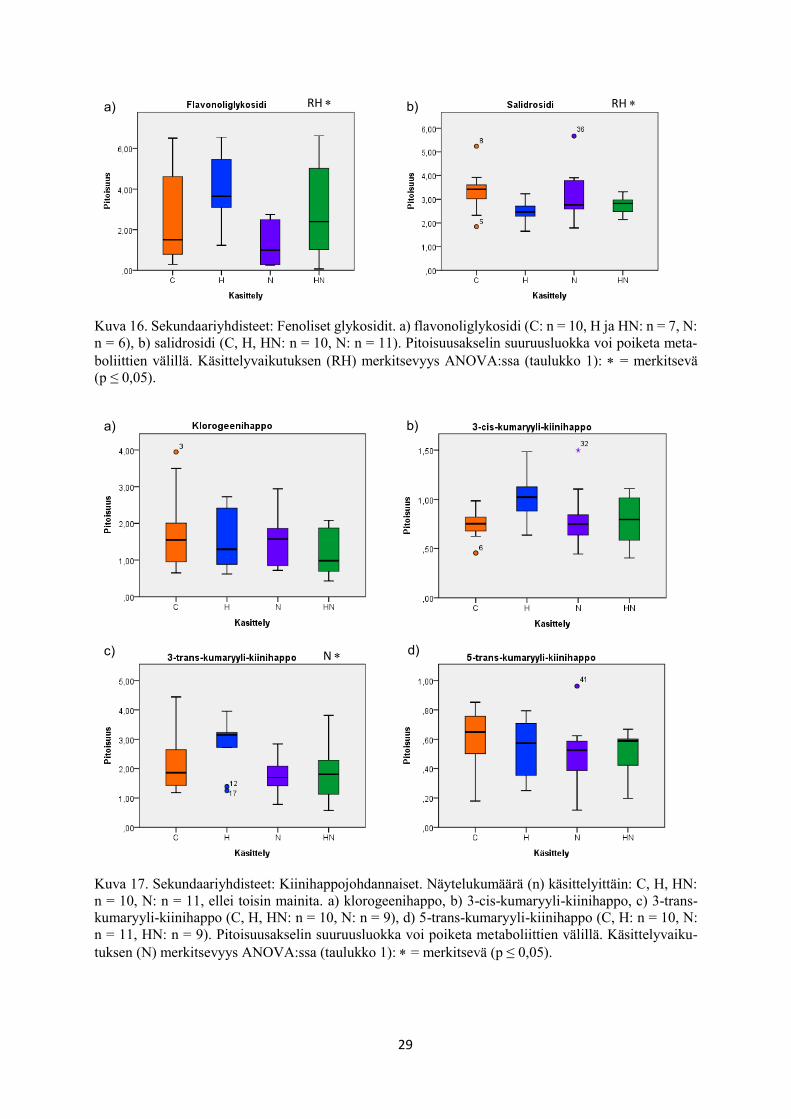
12c) ja sukkinihapon (taulukko 1, kuva 12f) pitoisuudet nousivat ja 3-trans-kumaryyli-kiini-
hapon (taulukko 1, kuva 17c) pitoisuudet alenivat kasvualustan kohotetussa typpipitoisuudessa
kasvatetuissa rauduskoivun lehdissä verrattuna kasvualustan alemmassa typpipitoisuudessa
kasvatukseen.
Suhteellisella ilmankosteudella ja kasvualustan typpipitoisuudella ei havaittu yhdysvaiku-
tusta metaboliittiaineistossa (taulukko 1). Tällöin kohotetun suhteellisen ilmankosteuden vaiku-
tus tarkasteluihin metaboliitteihin on ollut sama riippumatta kasvualustan typpitasosta, ja
vastaavasti kasvualustan kohotetun typpipitoisuuden vaikutus tarkasteltuihin metaboliitteihin
on ollut sama riippumatta suhteellisen ilmankosteuden tasosta.
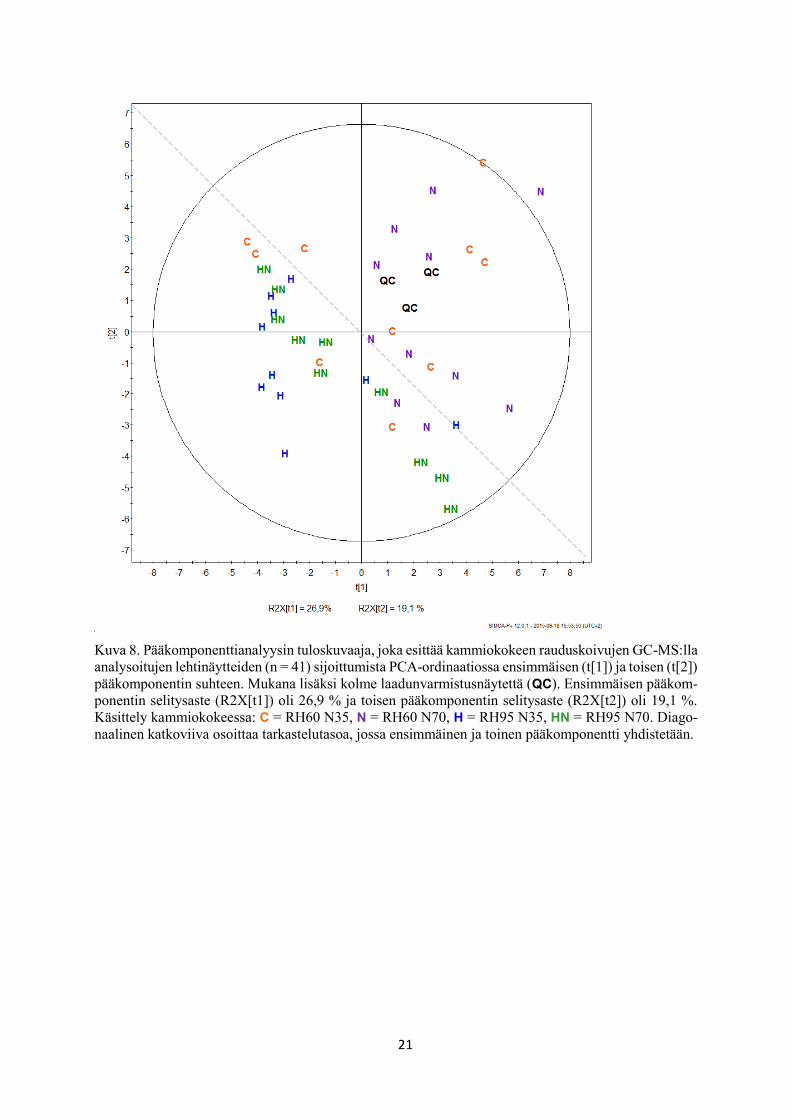
21
Kuva 8. Pääkomponenttianalyysin tuloskuvaaja, joka esittää kammiokokeen rauduskoivujen GC-MS:lla
analysoitujen lehtinäytteiden (n = 41) sijoittumista PCA-ordinaatiossa ensimmäisen (t[1]) ja toisen (t[2])
pääkomponentin suhteen. Mukana lisäksi kolme laadunvarmistusnäytettä (QC). Ensimmäisen pääkom-
ponentin selitysaste (R2X[t1]) oli 26,9 % ja toisen pääkomponentin selitysaste (R2X[t2]) oli 19,1 %.
Käsittely kammiokokeessa: C = RH60 N35, N = RH60 N70, H = RH95 N35, HN = RH95 N70. Diago-
naalinen katkoviiva osoittaa tarkastelutasoa, jossa ensimmäinen ja toinen pääkomponentti yhdistetään.

22
Kuva 9. Pääkomponenttianalyysin tuloskuvaaja yhdistettynä metaboliittiaineistoon (Loadings Bi Plot).
Kammiokokeen rauduskoivujen GC-MS:lla analysoidut lehtinäytteet (n = 41) ja niistä identifioidut
metaboliitit (n = 36) yhdistettynä samaan PCA-ordinaatioon. Ensimmäisen pääkomponentin selitysaste
(R2X[t1]) oli 27,3 % ja toisen pääkomponentin selitysaste (R2X[t2]) oli 19,8 %. Diagonaalinen katko-
viiva osoittaa tarkastelutasoa, jossa ensimmäinen ja toinen pääkomponentti yhdistetään. Käsittely
kammiokokeessa: C = RH60 N35, N = RH60 N70, H = RH95 N35, HN = RH95 N70. Metaboliittien
lyhenteiden selitys taulukossa 1.
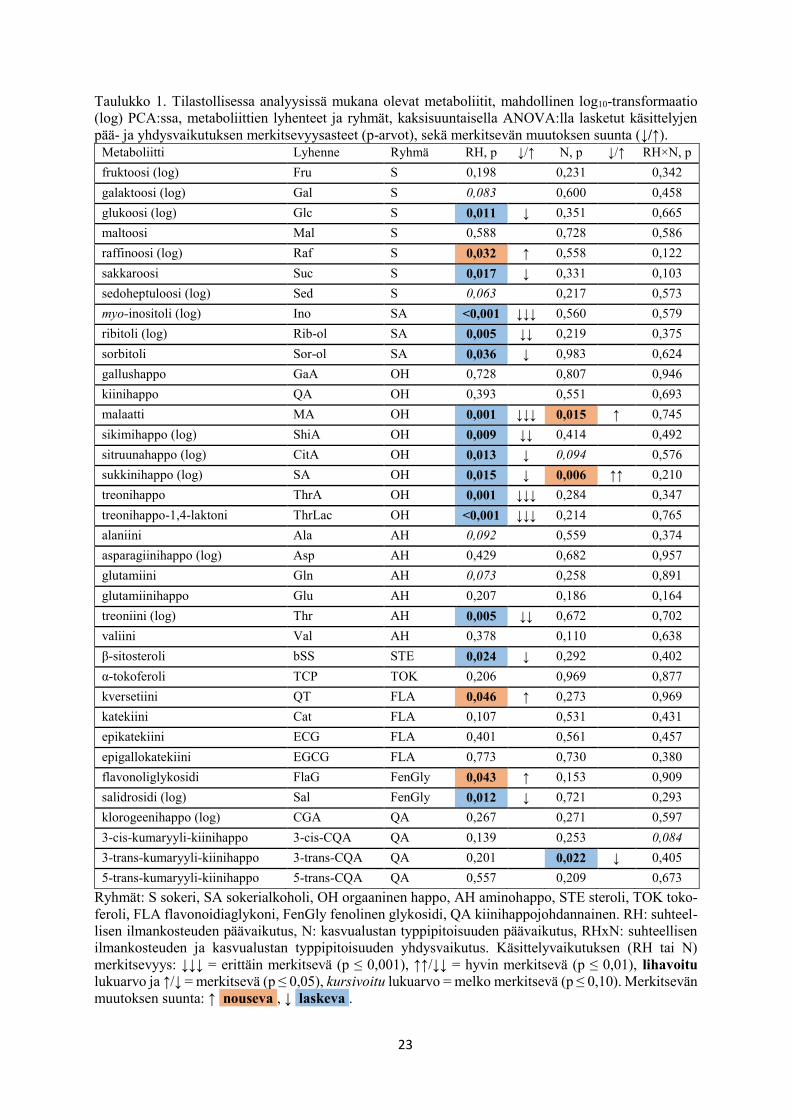
23
Taulukko 1. Tilastollisessa analyysissä mukana olevat metaboliitit, mahdollinen log10-transformaatio
(log) PCA:ssa, metaboliittien lyhenteet ja ryhmät, kaksisuuntaisella ANOVA:lla lasketut käsittelyjen
pää- ja yhdysvaikutuksen merkitsevyysasteet (p-arvot), sekä merkitsevän muutoksen suunta (↓/↑).
Metaboliitti Lyhenne Ryhmä RH, p ↓/↑ N, p ↓/↑ RH×N, p
fruktoosi (log) Fru S 0,198 0,231 0,342
galaktoosi (log) Gal S 0,083 0,600 0,458
glukoosi (log) Glc S 0,011 ↓ 0,351 0,665
maltoosi Mal S 0,588 0,728 0,586
raffinoosi (log) Raf S 0,032 ↑ 0,558 0,122
sakkaroosi Suc S 0,017 ↓ 0,331 0,103
sedoheptuloosi (log) Sed S 0,063 0,217 0,573
myo-inositoli (log) Ino SA <0,001 ↓↓↓ 0,560 0,579
ribitoli (log) Rib-ol SA 0,005 ↓↓ 0,219 0,375
sorbitoli Sor-ol SA 0,036 ↓ 0,983 0,624
gallushappo GaA OH 0,728 0,807 0,946
kiinihappo QA OH 0,393 0,551 0,693
malaatti MA OH 0,001 ↓↓↓ 0,015 ↑ 0,745
sikimihappo (log) ShiA OH 0,009 ↓↓ 0,414 0,492
sitruunahappo (log) CitA OH 0,013 ↓ 0,094 0,576
sukkinihappo (log) SA OH 0,015 ↓ 0,006 ↑↑ 0,210
treonihappo ThrA OH 0,001 ↓↓↓ 0,284 0,347
treonihappo-1,4-laktoni ThrLac OH <0,001 ↓↓↓ 0,214 0,765
alaniini Ala AH 0,092 0,559 0,374
asparagiinihappo (log) Asp AH 0,429 0,682 0,957
glutamiini Gln AH 0,073 0,258 0,891
glutamiinihappo Glu AH 0,207 0,186 0,164
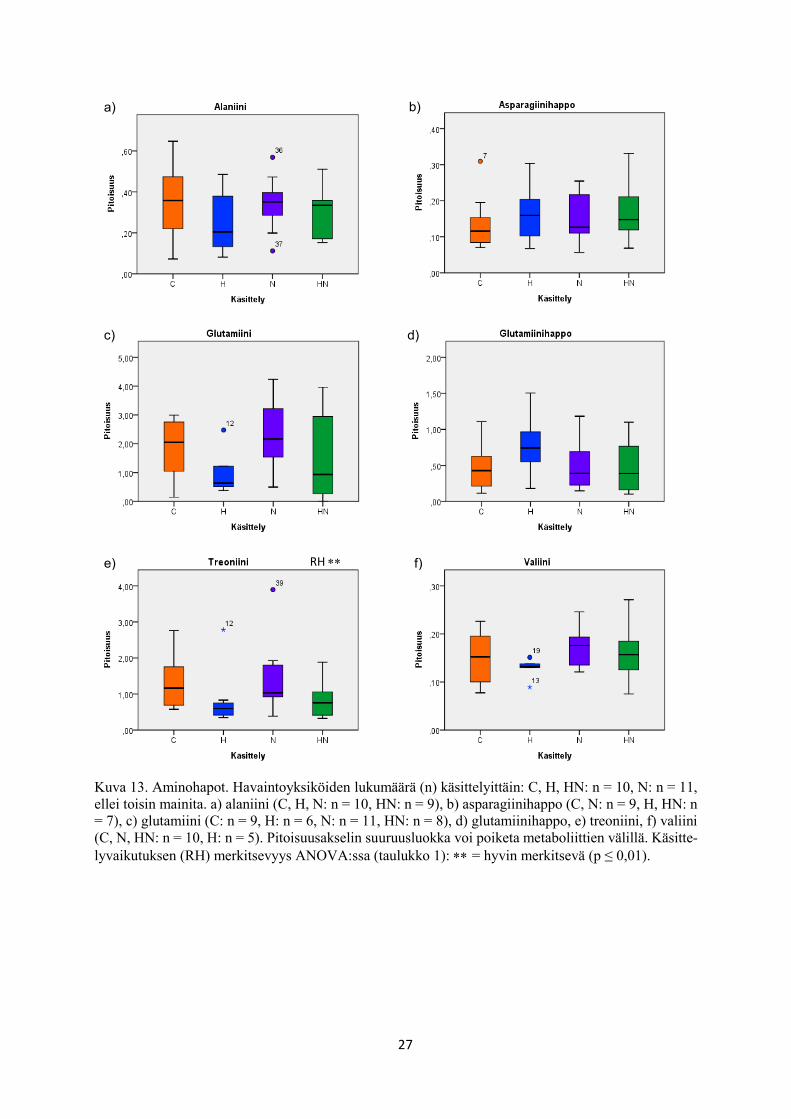
treoniini (log) Thr AH 0,005 ↓↓ 0,672 0,702
valiini Val AH 0,378 0,110 0,638
β-sitosteroli bSS STE 0,024 ↓ 0,292 0,402
α-tokoferoli TCP TOK 0,206 0,969 0,877
kversetiini QT FLA 0,046 ↑ 0,273 0,969
katekiini Cat FLA 0,107 0,531 0,431
epikatekiini ECG FLA 0,401 0,561 0,457
epigallokatekiini EGCG FLA 0,773 0,730 0,380
flavonoliglykosidi FlaG FenGly 0,043 ↑ 0,153 0,909
salidrosidi (log) Sal FenGly 0,012 ↓ 0,721 0,293
klorogeenihappo (log) CGA QA 0,267 0,271 0,597
3-cis-kumaryyli-kiinihappo 3-cis-CQA QA 0,139 0,253 0,084
3-trans-kumaryyli-kiinihappo 3-trans-CQA QA 0,201 0,022 ↓ 0,405
5-trans-kumaryyli-kiinihappo 5-trans-CQA QA 0,557 0,209 0,673
Ryhmät: S sokeri, SA sokerialkoholi, OH orgaaninen happo, AH aminohappo, STE steroli, TOK toko-
feroli, FLA flavonoidiaglykoni, FenGly fenolinen glykosidi, QA kiinihappojohdannainen. RH: suhteel-
lisen ilmankosteuden päävaikutus, N: kasvualustan typpipitoisuuden päävaikutus, RHxN: suhteellisen
ilmankosteuden ja kasvualustan typpipitoisuuden yhdysvaikutus. Käsittelyvaikutuksen (RH tai N)
merkitsevyys: ↓↓↓ = erittäin merkitsevä (p ≤ 0,001), ↑↑/↓↓ = hyvin merkitsevä (p ≤ 0,01), lihavoitu
lukuarvo ja ↑/↓ = merkitsevä (p ≤ 0,05), kursivoitu lukuarvo = melko merkitsevä (p ≤ 0,10). Merkitsevän
muutoksen suunta: ↑ nouseva , ↓ laskeva .
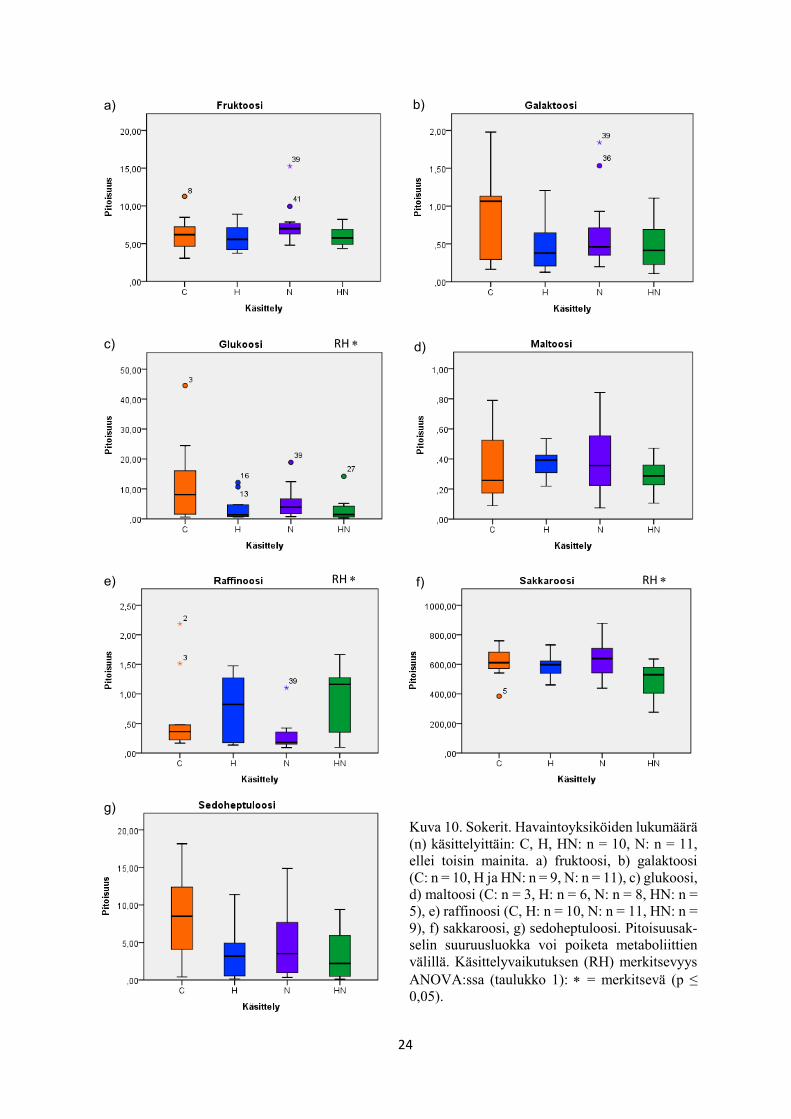
24
Kuva 10. Sokerit. Havaintoyksiköiden lukumäärä
(n) käsittelyittäin: C, H, HN: n = 10, N: n = 11,
ellei toisin mainita. a) fruktoosi, b) galaktoosi
(C: n = 10, H ja HN: n = 9, N: n = 11), c) glukoosi,
d) maltoosi (C: n = 3, H: n = 6, N: n = 8, HN: n =
5), e) raffinoosi (C, H: n = 10, N: n = 11, HN: n =
9), f) sakkaroosi, g) sedoheptuloosi. Pitoisuusak-
selin suuruusluokka voi poiketa metaboliittien
välillä. Käsittelyvaikutuksen (RH) merkitsevyys
ANOVA:ssa (taulukko 1): = merkitsevä (p ≤
0,05).
RH
a) b)
RH
RH
c) d)
e) f)
g)
25
Kuva 11. Sokerialkoholit. Havaintoyksiköiden
lukumäärä (n) käsittelyittäin: C, H, HN: n = 10,
N: n = 11. a) myo-inositoli, b) ribitoli, c) sorbi-
toli. Pitoisuusakselin suuruusluokka voi poiketa
metaboliittien välillä. Käsittelyvaikutuksen
(RH) merkitsevyys ANOVA:ssa (taulukko 1):
= erittäin merkitsevä (p ≤ 0,001), = hyvin
merkitsevä (p ≤ 0,01), = merkitsevä (p ≤ 0,05).
a) b)
c)
RH RH
RH RH
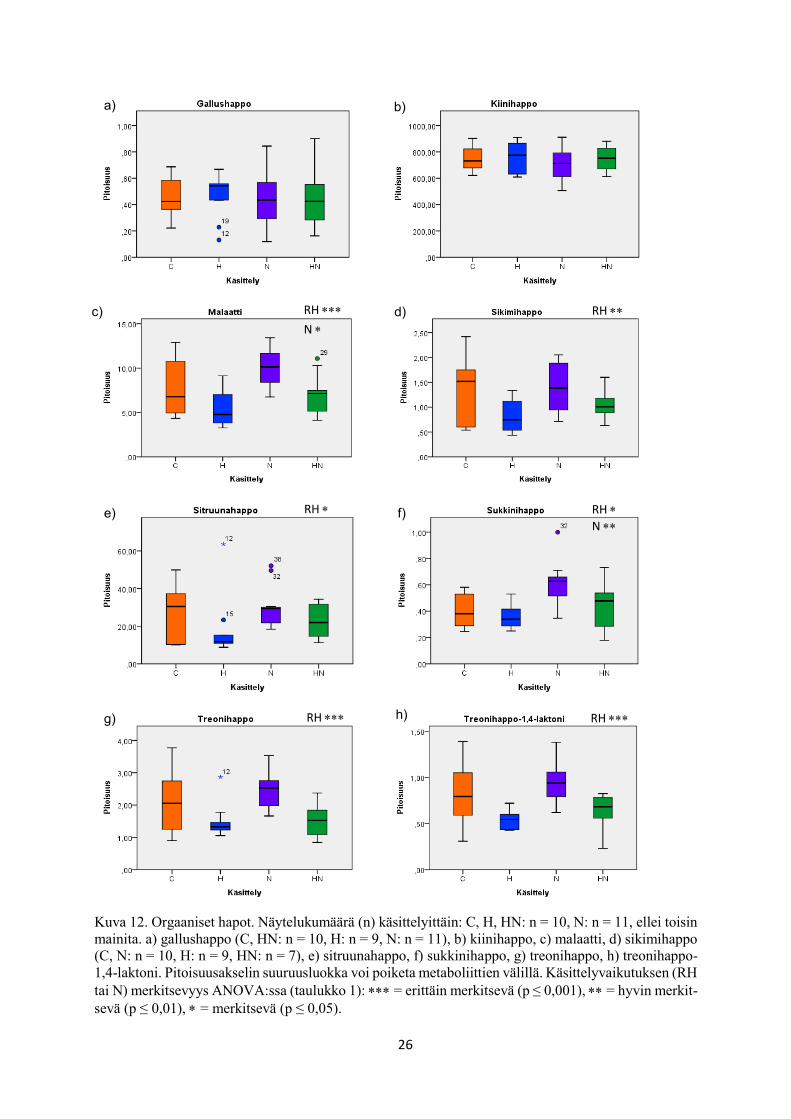
26
Kuva 12. Orgaaniset hapot. Näytelukumäärä (n) käsittelyittäin: C, H, HN: n = 10, N: n = 11, ellei toisin
mainita. a) gallushappo (C, HN: n = 10, H: n = 9, N: n = 11), b) kiinihappo, c) malaatti, d) sikimihappo
(C, N: n = 10, H: n = 9, HN: n = 7), e) sitruunahappo, f) sukkinihappo, g) treonihappo, h) treonihappo-
1,4-laktoni. Pitoisuusakselin suuruusluokka voi poiketa metaboliittien välillä. Käsittelyvaikutuksen (RH
tai N) merkitsevyys ANOVA:ssa (taulukko 1): = erittäin merkitsevä (p ≤ 0,001), = hyvin merkit-
sevä (p ≤ 0,01), = merkitsevä (p ≤ 0,05).
RH
N
RH
RH
RH
RH
N
a)
c) d)
e)
b)
f)
g) RH h)
27
Kuva 13. Aminohapot. Havaintoyksiköiden lukumäärä (n) käsittelyittäin: C, H, HN: n = 10, N: n = 11,
ellei toisin mainita. a) alaniini (C, H, N: n = 10, HN: n = 9), b) asparagiinihappo (C, N: n = 9, H, HN: n
= 7), c) glutamiini (C: n = 9, H: n = 6, N: n = 11, HN: n = 8), d) glutamiinihappo, e) treoniini, f) valiini
(C, N, HN: n = 10, H: n = 5). Pitoisuusakselin suuruusluokka voi poiketa metaboliittien välillä. Käsitte-
lyvaikutuksen (RH) merkitsevyys ANOVA:ssa (taulukko 1): = hyvin merkitsevä (p ≤ 0,01).
a) b)
c) d)
e) f) RH
28
Kuva 14. Primaariset terpenoidit. Havaintoyksiköiden lukumäärä (n) käsittelyittäin: C, H, HN: n = 10,
N: n = 11. a) β-sitosteroli, b) α-tokoferoli. Pitoisuusakselin suuruusluokka voi poiketa metaboliittien
välillä. Käsittelyvaikutuksen (RH) merkitsevyys ANOVA:ssa (taulukko 1): = merkitsevä (p ≤ 0,05).
Kuva 15. Sekundaariyhdisteet: Flavonoidiaglykonit. Havaintoyksiköiden lukumäärä (n) käsittelyittäin:
C, H, HN: n = 10, N: n = 11, ellei toisin mainita. a) kversetiini, b) katekiini, c) epikatekiini (C: n = 8, H:
n = 10, N: n = 11, HN: n = 9), d) epigallokatekiini. Pitoisuusakselin suuruusluokka voi poiketa meta-
boliittien välillä. Käsittelyvaikutuksen (RH) merkitsevyys ANOVA:ssa (taulukko 1): = merkitsevä
(p ≤ 0,05).
a) b)
a) b)
c) d)
RH
RH
29
Kuva 16. Sekundaariyhdisteet: Fenoliset glykosidit. a) flavonoliglykosidi (C: n = 10, H ja HN: n = 7, N:
n = 6), b) salidrosidi (C, H, HN: n = 10, N: n = 11). Pitoisuusakselin suuruusluokka voi poiketa meta-
boliittien välillä. Käsittelyvaikutuksen (RH) merkitsevyys ANOVA:ssa (taulukko 1): = merkitsevä
(p ≤ 0,05).
Kuva 17. Sekundaariyhdisteet: Kiinihappojohdannaiset. Näytelukumäärä (n) käsittelyittäin: C, H, HN:
n = 10, N: n = 11, ellei toisin mainita. a) klorogeenihappo, b) 3-cis-kumaryyli-kiinihappo, c) 3-trans-
kumaryyli-kiinihappo (C, H, HN: n = 10, N: n = 9), d) 5-trans-kumaryyli-kiinihappo (C, H: n = 10, N:
n = 11, HN: n = 9). Pitoisuusakselin suuruusluokka voi poiketa metaboliittien välillä. Käsittelyvaiku-
tuksen (N) merkitsevyys ANOVA:ssa (taulukko 1): = merkitsevä (p ≤ 0,05).
a) b)
c) d)
b) a) RH
N
RH
30
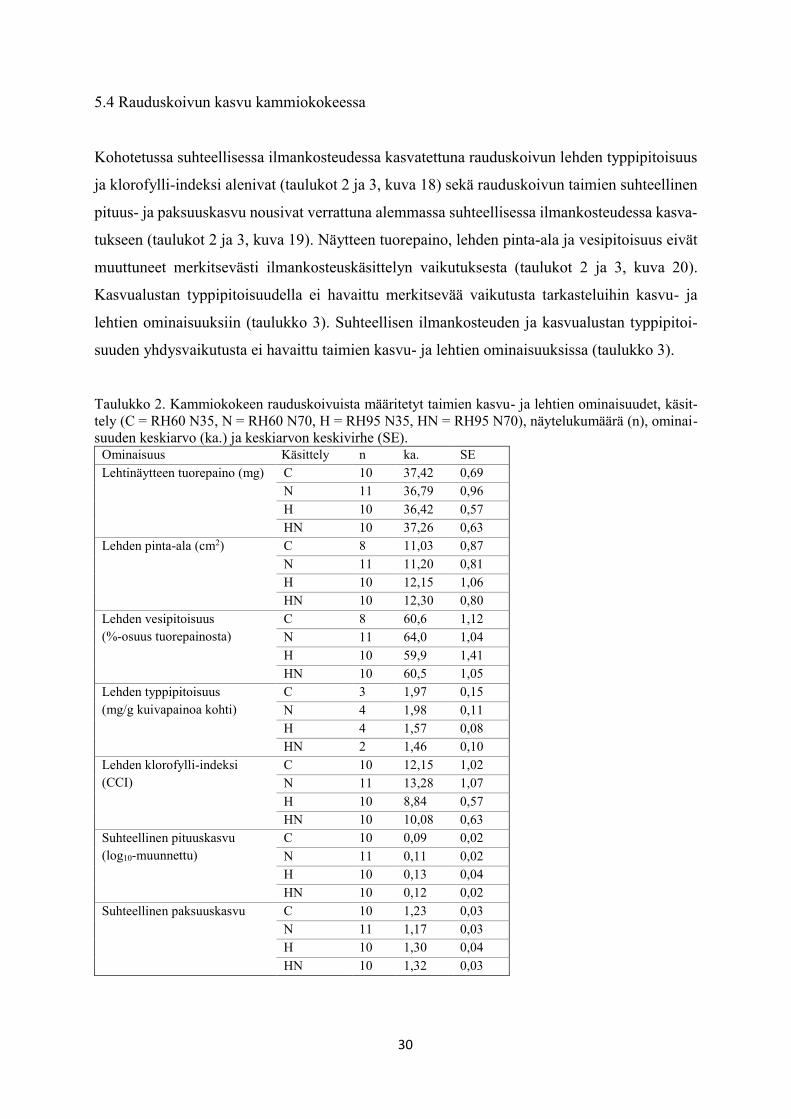
5.4 Rauduskoivun kasvu kammiokokeessa
Kohotetussa suhteellisessa ilmankosteudessa kasvatettuna rauduskoivun lehden typpipitoisuus
ja klorofylli-indeksi alenivat (taulukot 2 ja 3, kuva 18) sekä rauduskoivun taimien suhteellinen
pituus- ja paksuuskasvu nousivat verrattuna alemmassa suhteellisessa ilmankosteudessa kasva-
tukseen (taulukot 2 ja 3, kuva 19). Näytteen tuorepaino, lehden pinta-ala ja vesipitoisuus eivät
muuttuneet merkitsevästi ilmankosteuskäsittelyn vaikutuksesta (taulukot 2 ja 3, kuva 20).
Kasvualustan typpipitoisuudella ei havaittu merkitsevää vaikutusta tarkasteluihin kasvu- ja
lehtien ominaisuuksiin (taulukko 3). Suhteellisen ilmankosteuden ja kasvualustan typpipitoi-
suuden yhdysvaikutusta ei havaittu taimien kasvu- ja lehtien ominaisuuksissa (taulukko 3).
Taulukko 2. Kammiokokeen rauduskoivuista määritetyt taimien kasvu- ja lehtien ominaisuudet, käsit-
tely (C = RH60 N35, N = RH60 N70, H = RH95 N35, HN = RH95 N70), näytelukumäärä (n), ominai-
suuden keskiarvo (ka.) ja keskiarvon keskivirhe (SE). Ominaisuus Käsittely n ka. SE
Lehtinäytteen tuorepaino (mg) C 10 37,42 0,69
N 11 36,79 0,96
H 10 36,42 0,57
HN 10 37,26 0,63
Lehden pinta-ala (cm2) C 8 11,03 0,87
N 11 11,20 0,81
H 10 12,15 1,06
HN 10 12,30 0,80
Lehden vesipitoisuus
(%-osuus tuorepainosta)
C 8 60,6 1,12
N 11 64,0 1,04
H 10 59,9 1,41
HN 10 60,5 1,05
Lehden typpipitoisuus
(mg/g kuivapainoa kohti)
C 3 1,97 0,15
N 4 1,98 0,11
H 4 1,57 0,08
HN 2 1,46 0,10
Lehden klorofylli-indeksi
(CCI)
C 10 12,15 1,02
N 11 13,28 1,07
H 10 8,84 0,57
HN 10 10,08 0,63
Suhteellinen pituuskasvu
(log10-muunnettu)
C 10 0,09 0,02
N 11 0,11 0,02
H 10 0,13 0,04
HN 10 0,12 0,02
Suhteellinen paksuuskasvu C 10 1,23 0,03
N 11 1,17 0,03
H 10 1,30 0,04
HN 10 1,32 0,03
31
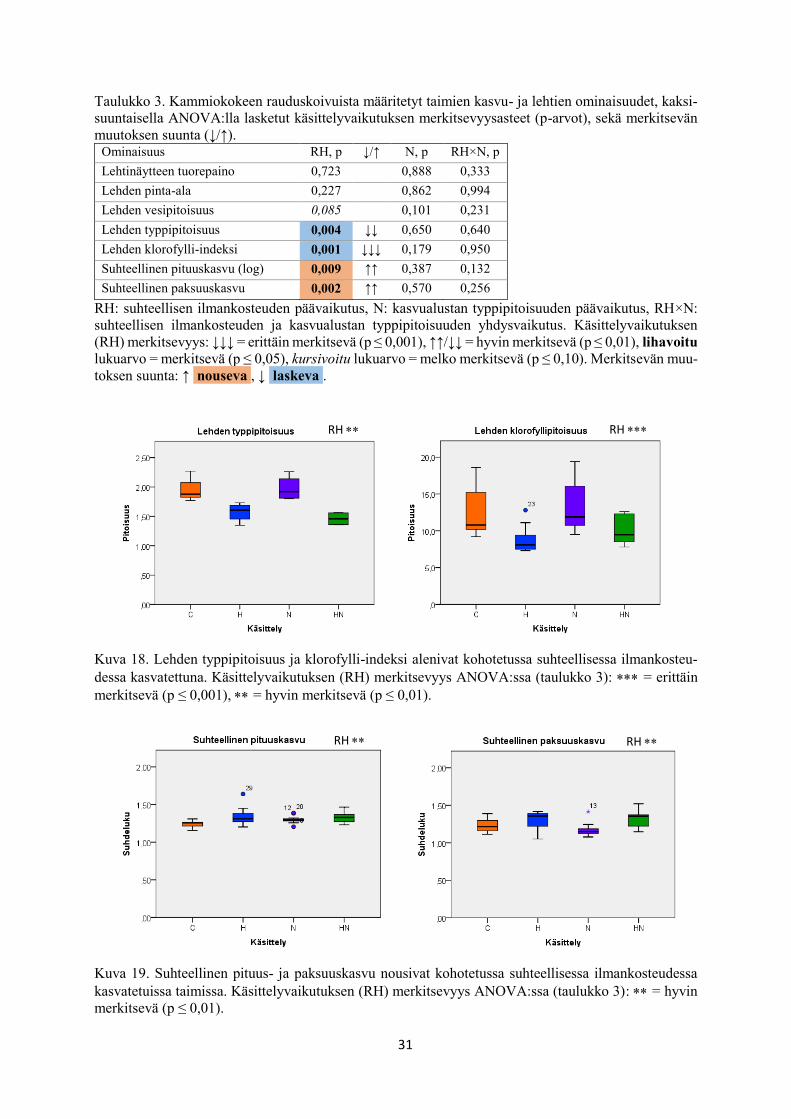
Taulukko 3. Kammiokokeen rauduskoivuista määritetyt taimien kasvu- ja lehtien ominaisuudet, kaksi-
suuntaisella ANOVA:lla lasketut käsittelyvaikutuksen merkitsevyysasteet (p-arvot), sekä merkitsevän
muutoksen suunta (↓/↑). Ominaisuus RH, p ↓/↑ N, p RH×N, p
Lehtinäytteen tuorepaino 0,723 0,888 0,333
Lehden pinta-ala 0,227 0,862 0,994
Lehden vesipitoisuus 0,085 0,101 0,231
Lehden typpipitoisuus 0,004 ↓↓ 0,650 0,640
Lehden klorofylli-indeksi 0,001 ↓↓↓ 0,179 0,950
Suhteellinen pituuskasvu (log) 0,009 ↑↑ 0,387 0,132
Suhteellinen paksuuskasvu 0,002 ↑↑ 0,570 0,256
RH: suhteellisen ilmankosteuden päävaikutus, N: kasvualustan typpipitoisuuden päävaikutus, RH×N:
suhteellisen ilmankosteuden ja kasvualustan typpipitoisuuden yhdysvaikutus. Käsittelyvaikutuksen
(RH) merkitsevyys: ↓↓↓ = erittäin merkitsevä (p ≤ 0,001), ↑↑/↓↓ = hyvin merkitsevä (p ≤ 0,01), lihavoitu
lukuarvo = merkitsevä (p ≤ 0,05), kursivoitu lukuarvo = melko merkitsevä (p ≤ 0,10). Merkitsevän muu-
toksen suunta: ↑ nouseva , ↓ laskeva .
Kuva 18. Lehden typpipitoisuus ja klorofylli-indeksi alenivat kohotetussa suhteellisessa ilmankosteu-
dessa kasvatettuna. Käsittelyvaikutuksen (RH) merkitsevyys ANOVA:ssa (taulukko 3): = erittäin
merkitsevä (p ≤ 0,001), = hyvin merkitsevä (p ≤ 0,01).
Kuva 19. Suhteellinen pituus- ja paksuuskasvu nousivat kohotetussa suhteellisessa ilmankosteudessa
kasvatetuissa taimissa. Käsittelyvaikutuksen (RH) merkitsevyys ANOVA:ssa (taulukko 3): = hyvin
merkitsevä (p ≤ 0,01).
RH RH
RH RH

32
Kuva 20. Lehtinäytteen tuorepaino, lehden pinta-ala ja vesipitoisuus eivät muuttuneet merkitsevästi
käsittelyjen vaikutuksesta.
5.5 Rauduskoivun taimien kasvu- ja lehtien ominaisuuksien sekä lehtinäytteistä määritettyjen
metaboliittipitoisuuksien välinen Pearson-korrelaatio
Tarkasteltaessa kammiokokeessa kasvatettujen rauduskoivujen kasvu- ja lehtien ominaisuuk-
sien sekä lehtinäytteistä määritettyjen metaboliittipitoisuuksien välistä lineaarista riippuvuutta
eli korrelaatioita, havaittiin lehtinäytteen tuorepainon korreloivan positiivisesti treonihapon
kanssa ja negatiivisesti galaktoosin, kiinihapon, alaniinin, epigallokatekiinin ja flavonoliglyko-
sidin kanssa (taulukko 4). Lehden pinta-alan kanssa positiivinen korrelaatio havaittiin
kiinihapolla, negatiivinen korrelaatio sorbitolilla (taulukko 4). Lehden vesipitoisuuden kanssa
positiivisesti korreloivat sorbitoli, malaatti, sitruuna-, sukkini- ja treonihappo, treonihappo-1,4-
laktoni ja treoniini, ja negatiivisesti korreloivat raffinoosi ja flavonoliglykosidi (taulukko 4).
Lehden typpipitoisuuden kanssa positiivisesti korreloivat galaktoosi, glukoosi, myo-inositoli,
ribitoli sorbitoli ja salidrosidi, negatiivista korrelaatiota lehden typpipitoisuuteen ei havaittu
(taulukko 4). Lehden klorofylli-indeksin kanssa positiivisesti korreloivat fruktoosi, sakkaroosi,
myo-inositoli, ribitoli, sorbitoli, malaatti, sitruuna-, sukkini- ja treonihappo, treonihappo-1-4-
laktoni, glutamiini, treoniini ja α-tokoferoli, ja negatiivisesti korreloivat raffinoosi, kiinihappo,
glutamiinihappo, kversetiini, epigallokatekiini, flavonoliglykosidi ja 3-trans-kumaryyli-kiini-
happo (taulukko 4). Suhteellisen pituuskasvun kanssa positiivinen korrelaatio havaittiin 3-cis-
kumaryyli-kiinihapolla ja negatiivinen korrelaatio galaktoosilla, glukoosilla, sedoheptuloosilla,
myo-inositolilla, kiinihapolla, sikimihapolla, β-sitosterolilla, epigallokatekiinilla ja salidrosi-
dilla (taulukko 4). Suhteellisen paksuuskasvun kanssa positiivinen korrelaatio havaittiin
glutamiinihapolla ja negatiivinen korrelaatio sakkaroosilla, myo-inositolilla, ribitolilla, malaa-
tilla, sitruuna-, sukkini- ja treonihapolla, glutamiinilla ja β-sitosterolilla (taulukko 4).
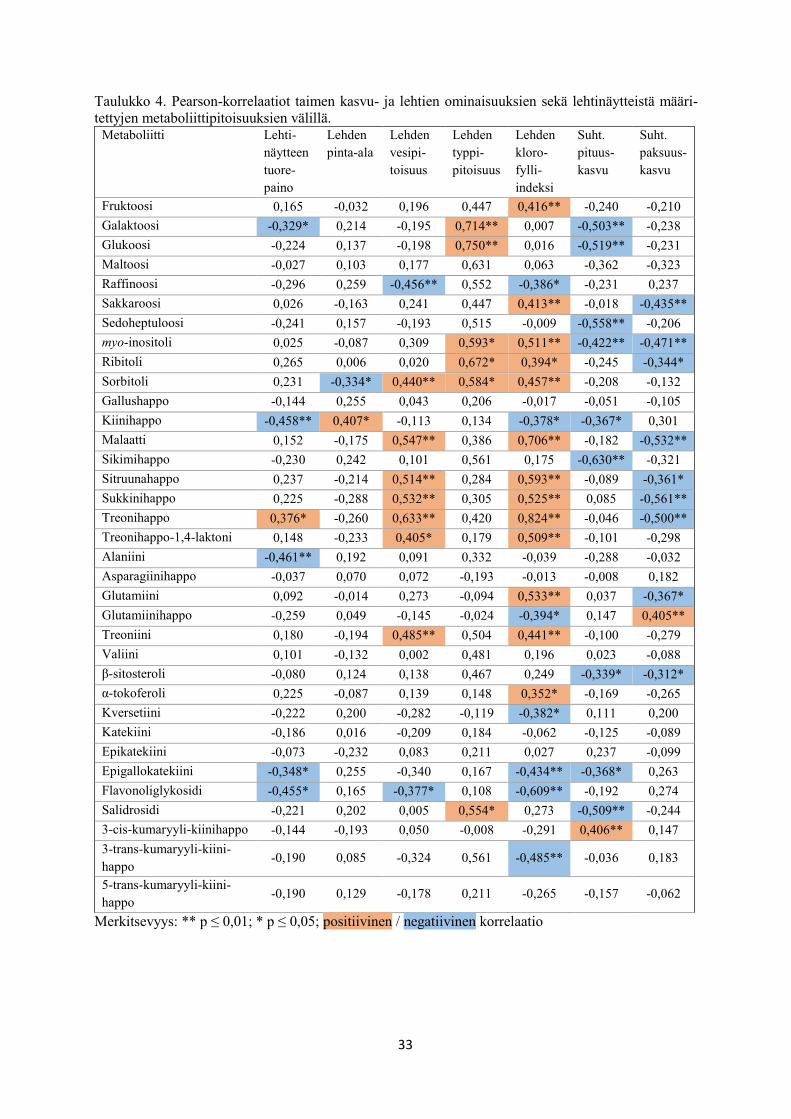
33
Taulukko 4. Pearson-korrelaatiot taimen kasvu- ja lehtien ominaisuuksien sekä lehtinäytteistä määri-
tettyjen metaboliittipitoisuuksien välillä. Metaboliitti Lehti-
näytteen
tuore-
paino
Lehden
pinta-ala
Lehden
vesipi-
toisuus
Lehden
typpi-
pitoisuus
Lehden
kloro-
fylli-
indeksi
Suht.
pituus-
kasvu
Suht.
paksuus-
kasvu
Fruktoosi 0,165 -0,032 0,196 0,447 0,416** -0,240 -0,210
Galaktoosi -0,329* 0,214 -0,195 0,714** 0,007 -0,503** -0,238
Glukoosi -0,224 0,137 -0,198 0,750** 0,016 -0,519** -0,231
Maltoosi -0,027 0,103 0,177 0,631 0,063 -0,362 -0,323
Raffinoosi -0,296 0,259 -0,456** 0,552 -0,386* -0,231 0,237
Sakkaroosi 0,026 -0,163 0,241 0,447 0,413** -0,018 -0,435**
Sedoheptuloosi -0,241 0,157 -0,193 0,515 -0,009 -0,558** -0,206
myo-inositoli 0,025 -0,087 0,309 0,593* 0,511** -0,422** -0,471**
Ribitoli 0,265 0,006 0,020 0,672* 0,394* -0,245 -0,344*
Sorbitoli 0,231 -0,334* 0,440** 0,584* 0,457** -0,208 -0,132
Gallushappo -0,144 0,255 0,043 0,206 -0,017 -0,051 -0,105
Kiinihappo -0,458** 0,407* -0,113 0,134 -0,378* -0,367* 0,301
Malaatti 0,152 -0,175 0,547** 0,386 0,706** -0,182 -0,532**
Sikimihappo -0,230 0,242 0,101 0,561 0,175 -0,630** -0,321
Sitruunahappo 0,237 -0,214 0,514** 0,284 0,593** -0,089 -0,361*
Sukkinihappo 0,225 -0,288 0,532** 0,305 0,525** 0,085 -0,561**
Treonihappo 0,376* -0,260 0,633** 0,420 0,824** -0,046 -0,500**
Treonihappo-1,4-laktoni 0,148 -0,233 0,405* 0,179 0,509** -0,101 -0,298
Alaniini -0,461** 0,192 0,091 0,332 -0,039 -0,288 -0,032
Asparagiinihappo -0,037 0,070 0,072 -0,193 -0,013 -0,008 0,182
Glutamiini 0,092 -0,014 0,273 -0,094 0,533** 0,037 -0,367*
Glutamiinihappo -0,259 0,049 -0,145 -0,024 -0,394* 0,147 0,405**
Treoniini 0,180 -0,194 0,485** 0,504 0,441** -0,100 -0,279
Valiini 0,101 -0,132 0,002 0,481 0,196 0,023 -0,088
β-sitosteroli -0,080 0,124 0,138 0,467 0,249 -0,339* -0,312*
α-tokoferoli 0,225 -0,087 0,139 0,148 0,352* -0,169 -0,265
Kversetiini -0,222 0,200 -0,282 -0,119 -0,382* 0,111 0,200
Katekiini -0,186 0,016 -0,209 0,184 -0,062 -0,125 -0,089
Epikatekiini -0,073 -0,232 0,083 0,211 0,027 0,237 -0,099
Epigallokatekiini -0,348* 0,255 -0,340 0,167 -0,434** -0,368* 0,263
Flavonoliglykosidi -0,455* 0,165 -0,377* 0,108 -0,609** -0,192 0,274
Salidrosidi -0,221 0,202 0,005 0,554* 0,273 -0,509** -0,244
3-cis-kumaryyli-kiinihappo -0,144 -0,193 0,050 -0,008 -0,291 0,406** 0,147
3-trans-kumaryyli-kiini-
happo -0,190 0,085 -0,324 0,561 -0,485** -0,036 0,183
5-trans-kumaryyli-kiini-
happo -0,190 0,129 -0,178 0,211 -0,265 -0,157 -0,062
Merkitsevyys: ** p ≤ 0,01; * p ≤ 0,05; positiivinen / negatiivinen korrelaatio

34
6 TULOSTEN TARKASTELU
Tämän pro gradu -tutkielman tulokset osoittavat, että kammiokokeen kohotettu suhteellinen
ilmankosteus vaikutti voimakkaasti rauduskoivun (Betula pendula Roth) lehtien metaboliaan.
Rauduskoivun lehtinäytteistä löytyi yhteensä 169 erilaista metaboliittia (Lihavainen ym.
2016a), joista tässä pro gradu -tutkielmassa tarkasteltaviksi valikoitui 36 metaboliittia. Useim-
pien primaarimetaboliittien tuotanto aleni kohotetussa suhteellisessa ilmankosteudessa kasva-
tetuissa rauduskoivun taimien lehdissä, ainoastaan raffinoosin pitoisuus nousi. Suhteellisen
ilmankosteuden kohottamisen vaikutus lehtien sekundaarimetaboliitteihin oli yhdisteestä
riippuvainen, nostaen tässä tutkielmassa tarkastelluista sekundaariyhdisteistä kversetiinin ja
flavonoliglykosidin pitoisuuksia ja alentaen salidrosidin pitoisuuksia. Kammiokokeessa
suhteellista ilmankosteutta on pystytty kontrolloidusti kasvattamaan hyvin suureksi, ollen tutki-
musasetelmassa 95 %. Ero alempana ilmankosteustasona käytettyyn luonnontasoiseen suhteel-
liseen ilmankosteuteen (RH 60 %) on suuri. Tällöin kammiokokeen kohotetussa suhteellisessa
ilmankosteudessa kasvattaminen on aiheuttanut rauduskoivun taimille voimakasta ympäristö-
stressiä, mikä näkyy lehtien metaboliavasteissa.
Kasvualustan kohotetussa typpipitoisuudessa kasvattaminen vaikutti rauduskoivun taimien
lehtien metaboliitteihin nostaen malaatin ja sukkinihapon sekä alentaen 3-trans-kumaryyli-
kiinihapon pitoisuuksia. Tällöin voidaan todeta, ettei kasvualustan typpipitoisuuden kohottami-
sella ole määrällisesti ollut yhtä merkittäviä vaikutuksia lehtien metaboliaan kuin mitä kohote-
tussa suhteellisessa ilmankosteudessa kasvatettujen taimien lehdissä havaittiin. Kasvualustan
alemmassa eli optimaalisessa typpitasossa kasvatetut taimet eivät luonnontasoisessa ilmankos-
teudessa osoittaneet metabolian osalta typpirajoitettua kasvua, jolloin peruslannoitus on ollut
riittävää metabolian normaaliin toimintaan ja taimien kasvuun. Kohotettu suhteellinen ilman-
kosteus on puolestaan rajoittanut kasvualustaan lisätyn typen hyödyntämistä kasvin meta-
boliassa sekä rajoittanut typen käyttöä myös optimaalisella typpitasolla kasvatetuilla kasveilla.
Suhteellisen ilmankosteuden ja kasvualustan typpipitoisuuden yhdysvaikutusta ei havaittu
metaboliittiaineistossa. Tällöin esimerkiksi kohotetun suhteellisen ilmankosteuden vaikutukset
lehtien metaboliaan ovat olleet samansuuntaisia riippumatta kasvualustan typpitasosta, sekä
kasvualustan kohotetun typpipitoisuuden vaikutukset lehtien metaboliaan ovat olleet saman-
suuntaisia riippumatta suhteellisen ilmankosteuden tasosta.

35
Kasvatus kohotetussa suhteellisessa ilmankosteudessa vaikutti rauduskoivun taimien
kasvuun lisäämällä suhteellista pituus- ja paksuuskasvua sekä lehtien ominaisuuksiin alenta-
malla lehden typpipitoisuutta ja klorofylli-indeksiä. Kasvualustan typpipitoisuuden kohotta-
misella ei havaittu vaikutusta kasvuparametreihin.
Suhteellisen ilmankosteuden ja kasvualustan typpipitoisuuden yhdysvaikutusta ei havaittu
kasvuparametriaineistossa. Tällöin esimerkiksi kohotetun suhteellisen ilmankosteuden vaiku-
tukset kasvuparametreihin ovat olleet samansuuntaisia riippumatta kasvualustan typpitasosta,
sekä kasvualustan kohotetun typpipitoisuuden vaikutukset kasvuparametreihin ovat olleet
samansuuntaisia riippumatta suhteellisen ilmankosteuden tasosta.
Korkea suhteellinen ilmankosteus voi häiritä ilmarakojen toiminnan säätelyä ja kaasujen
vaihtoa (Gislerød & Nelson 1989, Fordham ym. 2001, Lihavainen ym. 2016a,b). Vaikka
korkeassa suhteellisessa ilmankosteudessa suurin osa lehden alapinnalla olevista ilmaraoista on
auki, veden haihtuminen vähenee ympäröivän ilman vesihöyrypitoisuuden ollessa suuri.
Haihtumisen vähentyessä veden ja mineraalien imeytyminen maaperästä oletettavasti vähenee
johtojänteiden turgoripaineen alentuessa. Suhteellisen ilmankosteuden kohoamisen on havaittu
vähentävän rauduskoivujen ja hybridihaapojen (Populus tremula L.×Populus tremuloides
Michx.) kykyä ottaa typpeä maasta (Tullus ym. 2012, Sellin ym. 2013). Korkeassa suhteelli-
sessa ilmankosteudessa lehtipuut eivät välttämättä kykene hyödyntämään kasvualustan
suurempaa typpimäärää alentuneen haihdunnan ja rajoittuneen virtauksen vuoksi (Lihavainen
ym. 2016b, Sellin ym. 2016). Täten kohotetussa suhteellisessa ilmankosteudessa kasvin
voidaan odottaa allokoivan vähemmän typpeä kasvuun ja aineenvaihduntaan kuin alemmassa
suhteellisessa ilmankosteudessa.
Kammiokokeen aikana annettu typpilisä oli optimitasolla 112 mg ja kohotetulla typpitasolla
172 mg per taimi. Tämän pro gradu -tutkielman tuloksia voidaan tulkita käytettävissä olleen
typen hyödynnettävyyden suhteen niin, että vähiten typpeä oli hyödynnettävissä koivuilla, jotka
kasvoivat kohotetussa suhteellisessa ilmankosteudessa ja kasvualustan optimaalisessa typpi-
pitoisuudessa (näytteet H: RH95 N35). Nämä yksilöt edustivat voimakkaimmin typpirajoitettua
kasvua. Toiseksi vähiten typpeä oli hyödynnettävissä koivuilla, jotka kasvoivat kohotetussa
suhteellisessa ilmankosteudessa ja kasvualustan kohotetussa typpipitoisuudessa (näytteet HN:
RH95 N70). Toiseksi eniten typpeä oli hyödynnettävissä koivuilla, jotka kasvoivat alemmassa
suhteellisessa ilmankosteudessa ja kasvualustan optimaalisessa typpipitoisuudessa (näytteet C:
RH60 N35). Nämä yksilöt miellettiin kontrollinäytteiksi. Eniten typpeä oli hyödynnettävissä
koivuilla, jotka kasvoivat alemmassa suhteellisessa ilmankosteudessa ja kasvualustan kohote-
tussa typpipitoisuudessa (näytteet N: RH60 N70). Näiden yksilöiden tulisi teoriassa edustaa

36
typpirikasta kasvua, mutta tässä tutkielmassa tarkasteltuna kasvualustan kohotetulla typpipitoi-
suudella havaitut vaikutukset lehden metaboliaan olivat vähäisiä eikä vaikutusta rauduskoivun
taimien kasvuun ja lehden ominaisuuksiin havaittu.
6.1 Kohotetun suhteellisen ilmankosteuden vaikutus metaboliitteihin
Kasvatus kohotetussa suhteellisessa ilmankosteudessa vaikutti pitoisuutta alentavasti useilla
primaarimetaboliiteilla, jotka osallistuvat muiden muassa glykolyysiin, sitruunahappokiertoon
ja aminohappojen tuotantoon. Korkeassa suhteellisessa ilmankosteudessa veden ja ravinteiden
nilakuljetus vähenee lehtien ilmarakojen lisääntyneen aukiolon seurauksena (Gislerød &
Nelson 1989, Fordham ym. 2001, Lihavainen ym. 2016a), mikä aiheuttaa typen puutosta ja
edelleen hiilen sidonnassa keskeisen Rubisco-entsyymin määrän ja aktiivisuuden vähenemistä
kasvin lehdissä (Paul & Driscoll 1997). Tämä johtaa fotosynteesin inhibitioon (Lemoine ym.
2013), jolloin fotosynteesissä tuotettujen hiilirunkojen pitoisuudet alenevat kasvin lehdissä,
mikä puolestaan alentaa näistä hiilirungoista syntetisoitavien jatkotuotteiden, kuten hiilihyd-
raattien ja orgaanisten happojen, pitoisuuksia.
Tässä pro gradu -tutkielmassa tarkastelluista sokereista havaittiin glukoosin ja sakkaroosin
pitoisuuksien alenevan rauduskoivun lehdissä kohotetussa suhteellisessa ilmankosteudessa
kasvatettuna. Hiilihydraattien pitoisuuksien on useimmiten havaittu typen puutteessa kasvatet-
tujen kasvien lehdissä tämän tutkielman tuloksista poiketen nousevan, esimerkiksi tupakalla
(Nicotiana tabacum L.) (Paul & Driscoll 1997, Scheible ym. 1997) ja maissilla (Zea mays L.)
(Schlüter ym. 2012, 2013).
Raffinoosia käytetään kasvin lehdissä tärkkelyksen ohella hiilirunkojen varastoimismuotona
(Sprenger & Keller 2001). Raffinoosipitoisuuden nouseminen rauduskoivun lehdissä kohote-
tussa suhteellisessa ilmankosteudessa kasvatettuna osoittaisi hiilirunkojen varastoinnin
raffinoosiin lisääntyvän (Lihavainen ym. 2016a) samalla, kun muiden hiilihydraattien pitoisuus
alenee. Lehtien korkeampi raffinoosipitoisuus kohotetussa suhteellisessa ilmankosteudessa voi
lisäksi vaikuttaa ravinteiden kuljetukseen kasvisolukoiden välillä (Lihavainen ym. 2016a).
Pääasiallisesti kasvissa kuljetetaan sakkaroosia, mutta jotkin lajit kuljettavat myös raffinoosia
ja sokerialkoholeja (Lalonde ym. 2004). Kuljetettavien ravinteiden virtaaman vähentyessä
sakkaroosin nilaan lataaminen voi alentua, mikä johtaisi korkeampiin raffinoosipitoisuuksiin
lehdissä (Lemoine ym. 2013). Korkea raffinoosipitoisuus typen puutoksessa kasvatettujen
kasvien lehdissä on havaittu esimerkiksi maissilla (Schlüter ym. 2012, 2013). Raffinoosipitoi-

37
suuden nouseminen voi myös osaltaan kompensoida muiden osmolyyttien pitoisuuksien alene-
misesta mahdollisesti aiheutuvaa lehden osmoottisen säätelykyvyn heikentymistä (Barchet ym.
2013).
Tässä pro gradu -tutkielmassa havaittiin kaikkien tarkasteltujen sokerialkoholien, myo-inosi-
tolin, ribitolin ja sorbitolin, pitoisuuksien alenevan kohotetussa suhteellisessa ilmankosteudessa
kasvatetuissa rauduskoivun lehdissä. Sokerialkoholien on esitetty toimivan hiilivarantona sekä
osallistuvan muun muassa kasvin stressioireiden lieventämiseen ja lehden osmoottiseen sääte-
lyyn (Merchant & Richter 2011, Dumschott ym. 2017). Yleisenä stressivasteena sokerialko-
holien pitoisuuksien on havaittu nousevan, mutta tässä tutkielmassa tarkasteltuna suhteellisen
ilmankosteuden kohottaminen vaikutti sokerialkoholien pitoisuutta alentavasti.
Useiden sitruunahappokierron välituotteina toimivien orgaanisten happojen pitoisuuden
havaittiin tässä tutkielmassa alenevan koivunlehdissä, kun taimia kasvatettiin kohotetussa
suhteellisessa ilmankosteudessa. Tämä osoittaa typpirajoitettua kasvua, sillä sitruunahappo-
kierron välituotteiden pitoisuuksien on havaittu alenevan typen puutoksessa (Scheible ym.
1997). Esimerkiksi malaatti toimii kasvisolussa muun muassa osmoottisena säätelijänä ja on
keskeinen välituote useissa metaboliaprosesseissa, kuten fotosynteesissä ja sitruunahappokier-
rossa (Barchet ym. 2013). Tästä syystä malaatin pitoisuusvaihtelun vaikutusta kasvisolukossa
ei voida tarkoin kohdentaa yksittäiseen metaboliaprosessiin, koska sillä on moninaisia vaiku-
tuksia kasvin aineenvaihdunnassa.
Fenyylipropanoidien esiasteen sikimihapon pitoisuuden havaittiin alenevan rauduskoivun
lehdissä kohotetussa suhteellisessa ilmankosteudessa, mikä voi osoittaa sikimihapon jatkotuot-
teiden lisääntyneen biosynteesin (Lihavainen ym. 2016a). Sekundaarimetaboliittien tuotantoa
pidetään stressitilassa yllä kierrättämällä primaarisia lähtöaineita tehokkaammin kuin stressit-
tömässä tilassa, jolloin primaaristen lähtöaineiden alhaisetkin pitoisuudet riittävät sekundaari-
yhdisteiden tehokkaaseen tuotantoon (Kutchan ym. 2015).
Tässä pro gradu -tutkielmassa tarkastelluista aminohapoista vain treoniinin pitoisuuden
havaittiin alenevan merkitsevästi. Yleisenä vasteena suhteellisen ilmankosteuden kohoamiseen
ja erityisesti typen puutoksessa kasvattamiseen on useiden aminohappojen pitoisuuksien
havaittu alenevan kasvin lehdissä (Geiger ym. 1999, Scheible ym. 2004, Fritz ym. 2006).
Flavonoideista kversetiinin ja flavonoliglykosidin pitoisuus oli korkeampi kohotetussa kuin
alemmassa suhteellisessa ilmankosteudessa kasvatetuissa rauduskoivun taimien lehdissä.
Flavonoidien ja fenolisten glykosidien tuotannon lisääntyminen kasveissa on yleinen vaste
ravinteiden puutokseen (Koricheva ym. 1998, Scheible ym. 2004, Amtmann & Armengaud

38
2009, Schlüter ym. 2013). Myös rauduskoivulla on osoitettu flavonoliglykosidien pitoisuuksien
nousevan typen puutteessa kasvatettuna (de la Rosa ym. 2001, Keski-Saari ym. 2005).
6.2 Kasvualustan kohotetun typpipitoisuuden vaikutus metaboliitteihin
Verrattuna suhteellisen ilmankosteuden vaikutuksiin, kasvualustan typpipitoisuuden kohotta-
misen vaikutukset rauduskoivujen lehtimetaboliaan olivat tässä pro gradu -tutkielmassa
tarkasteltuna suhteellisen vähäisiä, rajoittuen merkitseviin muutoksiin vain kolmen yhdisteen
pitoisuudessa. Kasvualustan kohotetussa typpipitoisuudessa kasvaneiden rauduskoivujen
lehdissä malaatin ja sukkinihapon pitoisuudet nousivat ja 3-trans-kumaryyli-kiinihapon pitoi-
suudet alenivat. Kasvualustan typpipitoisuuden on osoitettu vaikuttavan koivujen lehtien
metaboliaan ja metaboliittipitoisuuksiin (Keinänen ym. 1999, Keski-Saari & Julkunen-Tiitto
2003, Keski-Saari ym. 2005). Typpilannoituskokeissa on havaittu merkittäviä eroavaisuuksia
hiilen allokoinnissa eri metaboliareiteille ja kohdistamiseen esimerkiksi yksittäisiin fenolisiin
yhdisteisiin (Haukioja ym. 1998, Koricheva ym. 1998, Keinänen ym. 1999). Keski-Saari &
Julkunen-Tiitto (2003) havaitsivat typpilannoituksen alentavan hieskoivun (B. pubescens)
alalajin tunturikoivun (B. pubescens Erhr. ssp. czerepanovii (Orlova) Hämetahti) lehtien
fenolien, kuten flavonoidien ja tanniinien, pitoisuuksia. Myös rauduskoivulla typpilannoituksen
on todettu vaikuttavan lehtien fenolipitoisuuksiin muun muassa alentamalla eräiden flavonoi-
dien pitoisuuksia (Keinänen ym. 1999, Keski-Saari ym. 2005). Keinänen ym. (1999)
havaitsivat, että muun muassa 3-kumaryyli-kiinihappojen pitoisuudet alenivat rauduskoivun
lehdissä genotyypistä riippumatta vasteena kasvualustan kohotettuun lannoitukseen, mikä on
yhtenevä tulos tässä pro gradu -tutkielmassa tehtyjen havaintojen kanssa.
6.3 Rauduskoivun kasvu kammiokokeessa
Kohotettu suhteellinen ilmankosteus vaikutti tässä pro gradu -tutkielmassa tarkasteltuihin
rauduskoivujen kasvu- ja lehtien ominaisuuksiin lisäämällä taimien suhteellista pituus- ja
paksuuskasvua, sekä alentamalla lehden typpipitoisuutta ja klorofylli-indeksiä. Havaittu
rauduskoivun taimien suhteellisen pituus- ja paksuuskasvun lisääntyminen kohotetussa suhteel-
lisessa ilmankosteudessa on samansuuntainen vaste, mitä kasvihuoneessa kasvatettujen
kukkakasvien kasvua eri ilmankosteustasoilla selvittäneissä tutkimuksissa on havaittu
(Mortensen 1986, Gislerød & Nelson 1989, Gislerød & Mortensen 1990). Gislerød & Nelson

39
(1989) esittivät, että kasvin suhteellisen kasvun lisääntyminen korkeassa suhteellisessa ilman-
kosteudessa olisi seurausta ilmarakojen lisääntyneen aukiolon hiilidioksidin sitomista
edistävästä vaikutuksesta. Sidottu lisähiili voidaan käyttää biomassan tuotantoon eli kasvin
kasvuun. Suhteellisen ilmankosteuden kohoamisen on osoitettu hidastavan nestevirtausta
kasvin versossa (Tullus ym. 2012, Sellin ym. 2013), jolloin ravinteiden otto maaperästä vähe-
nee ja siten myös typpeä kuljetetaan lehtiin vähemmän, mikä johtaa lehtien typpipitoisuuden
alenemiseen. Myös Gislerød & Mortensen (1990) havaitsivat lehtien typpipitoisuuden alenevan
kohotetussa suhteellisessa ilmankosteudessa. Keski-Saari & Julkunen-Tiitto (2003) havaitsivat,
että typpirajoitetulla kasvualustalla kasvatettuna koivunlehtien typpipitoisuus oli alhaisempi
kuin optimaalisella typpitasolla kasvatettuna. Lehtien klorofylli-indeksin aleneminen on yhtey-
dessä lehtien typpipitoisuuden alenemiseen (Geiger ym. 1999, Lu & Zhang 2000). Fotosyn-
teesin inhiboituminen typenpuutoksessa (Geiger ym. 1999, Lu & Zhang 2000, Lemoine ym.
2013) vähentää klorofyllisynteesiä, jolloin typpirajoitettua kasvua edustavissa kohotetussa
suhteellisessa ilmankosteudessa kasvatetuissa lehdissä klorofyllipitoisuus alenee.
Kun aiemmissa tutkimuksissa typenpuutetta on verrattu typen optimitasoon, on kasvualustan
korkeamman typpipitoisuuden havaittu lisäävän koivujen kasvua (Keski-Saari & Julkunen-
Tiitto 2003, Keski-Saari ym. 2005). Tässä tutkielmassa kasvualustan kohotetulla typpipitoisuu-
della ei havaittu merkitsevää vaikutusta tarkasteltuihin kasvuominaisuuksiin. Tämä toden-
näköisesti johtuu siitä, että nykyisessä koeasetelmassa kasvualustan typen optimitasoa verrat-
tiin typen ylimäärään (Lihavainen ym. 2016a). Myös kammiokokeen lyhyt kesto ja istutus-
aineksessa olleen typen lähtötaso ovat voineet vaikuttaa siihen, ettei kasvualustan typpi-
lisäyksen vaikutusta kasvuominaisuuksiin nähty.
6.4 Korrelaatio
Sokerialkoholeilla havaittiin positiivinen korrelaatio sekä lehden typpipitoisuuteen että kloro-
fylli-indeksiin, jolloin niillä koivuyksilöillä joilla sokerialkoholeja oli vähän, myös lehden
typpipitoisuus ja klorofylli-indeksi olivat alhaiset. Sitruunahappokierron välituotteiden positii-
vinen korrelaatio lehden klorofylli-indeksiin osoittaa, että niissä lehdissä joiden klorofylli-
indeksi oli alhainen, myös sitruunahappokierron välituotteita oli vähän. Negatiivinen korre-
laatio klorofylli-indeksin ja kversetiinin sekä flavonoliglykosidin välillä kertoo, että ne lehdet
joilla oli alhainen klorofylli-indeksi, sisälsivät paljon kyseisiä fenolisia yhdisteitä.

40
6.5 Kammiokoetulosten vertailu FAHM -kenttäkokeen tuloksiin
Kammiokokeen tulokset kertovat lyhyen ajan muutoksista nuoren rauduskoivun lehden meta-
boliassa ja taimen kasvussa vasteena käsittelyihin. FAHM (Free Air Humidity Manipulation) -
kenttäkokeessa rauduskoivuja ja hybridihaapoja oli istutettu sumutettuihin kenttiin, missä
suhteellista ilmankosteutta nostettiin keinotekoisesti sumuttamalla ilmaan vesihöyryä, sekä
sumuttamattomiin kenttiin jotka toimivat kontrollikenttinä (Kupper ym. 2011). Kammio- ja
kenttäkokeiden tulosten vertailussa tulee huomioida erot koeasetelmissa. Kenttäkokeesta on
kerätty kasvutilastoa useamman vuoden ajalta, ja rauduskoivut olivat jo kentälle istutettaessa
vanhempia (yhden vuoden, Kupper ym. 2011) kuin kammiokokeen rauduskoivut (kuusi
kuukautta, Lihavainen ym. 2016a). Kammiokokeessa rauduskoivut kasvoivat muun muassa
suhteellisen ilmankosteuden osalta hyvin vakioidussa ympäristössä, mutta kentällä herbivorian,
sääolojen ja muiden ympäristötekijöiden vaikutus lisää aineiston vaihtelua. Kammiokokeessa
on lisäksi mahdollista toteuttaa suurempi suhteellisen ilmankosteuden ero kuin kenttäkokeessa.
Koska suuri osa sumutetusta vedestä haihtuu avonaisella kentällä ympäristöön, ei suhteellista
ilmankosteutta voida kasvattaa kenttäolosuhteissa kuin rajallisesti. FAHM -kenttäkokeessa
suhteellinen ilmankosteus oli noin 7 prosenttiyksikköä suurempi sumutetuilla kuin sumuttamat-
tomilla kentillä (Kupper ym. 2011). Suljetussa kammiossa veden haihtuminen ympäristöön
voidaan puolestaan minimoida, ja kammiokokeessa käsittelyjen välinen suhteellisen ilmankos-
teuden ero oli 35 prosenttiyksikköä (Lihavainen ym. 2016a).
FAHM -kentällä kasvaneista neljännen kasvukauden rauduskoivuista ja hybridihaavoista on
tehty myös lehtinäytteiden metabolomiikka-analyysejä (Lihavainen ym. 2016b). Kenttä-
kokeessa sumutetuilla koekentillä eli kohotetussa suhteellisessa ilmankosteudessa kasvaneiden
rauduskoivujen lehdissä sikimihapon pitoisuudet olivat alhaisempia, ja sorbitolin, treonihapon
ja treonihappo-1,4-laktonin pitoisuudet olivat korkeampia kuin sumuttamattomilla koekentillä
eli luonnontasoisessa suhteellisessa ilmankosteudessa kasvaneiden rauduskoivujen lehdissä
(Lihavainen ym. 2016b). Tässä pro gradu -tutkielmassa tarkasteltuna kammiokokeen kohote-
tussa suhteellisessa ilmankosteudessa kasvaneiden rauduskoivujen lehdissä sikimihapon
pitoisuus aleni vastaavasti kuten kenttäkokeessa havaittiin, mutta sorbitolin, treonihapon ja
treonihappo-1,4-laktonin pitoisuudet alenivat. Lisäksi kammiokokeen kohotetussa suhteelli-
sessa ilmankosteudessa kasvattujen rauduskoivujen lehdissä havaittiin korkeampi
raffinoosipitoisuus ja matalammat glukoosin, myo-inositolin, sitruunahapon ja sukkinihapon
pitoisuudet verrattuna alhaisemmassa suhteellisessa ilmankosteudessa kasvettujen rauduskoi-
vujen lehtien metaboliittipitoisuuksiin. Puolestaan FAHM -kenttäkokeesta kerätyissä

41
lehtinäytteissä edellä mainittujen metaboliittien pitoisuudet eivät eronneet rauduskoivun
lehdissä ilmankosteuskäsittelyjen välillä (Lihavainen ym. 2016b). Koeasetelmien toisistaan
poikkeava tulos voi selittyä kammiokokeen suuremmalla suhteellisen ilmankosteuden taso-
erolla sekä kammiokokeen lyhyellä kestolla. FAHM -kentällä kasvaneista rauduskoivuista oli
otettu metaboliatutkimuksissa analysoitavat lehtinäytteet neljäntenä käsittelykautena, jolloin
rauduskoivut ovat jo ehtineet monin tavoin sopeutua ympäristön stressitekijöihin eli korkeam-
paan suhteelliseen ilmankosteuteen sumutetuilla kentillä. Poikkeavuutta kammiokokeeseen
tuovat kentällä myös muiden muassa erilainen maannos, sääolojen luontaiset vaihtelut ja
herbivoria.
FAHM -kenttäkokeessa lyhyen ajan vasteena maanpäällinen kasvu hidastui sekä rauduskoi-
vulla että hybridihaavalla (Tullus ym. 2012, Sellin ym. 2013). Kasvuvasteen osalta
kenttäkokeen tulokset eroavat tässä pro gradu -tutkielmassa tarkastelluista kammiokokeen
tuloksista, jossa rauduskoivujen taimien kasvu kiihtyi korkeammassa suhteellisessa ilmankos-
teudessa kasvatettuna. Kohotetusta suhteellisesta ilmankosteudesta johtuva vähentynyt
haihdunta lehdissä (Fordham ym. 2001) vaikutti typen ottoon maaperästä niin, että kenttä-
kokeen ensimmäisinä vuosina rauduskoivun lehtien typpipitoisuus oli alhaisempi sumutetuilla
kuin sumuttamattomilla kentillä (Sellin ym. 2013). Tältä osin tulos on samansuuntainen, mitä
tässä pro gradu -tutkielmassa kammiokokeen rauduskoivuilla havaittiin. Kenttäkokeen neljän-
tenä käsittelyvuotena rauduskoivujen lehtien typpipitoisuus ei kuitenkaan enää eronnut sumu-
tettujen ja sumuttamattomien kenttien välillä (Lihavainen ym. 2016b), toisin sanoen sumute-
tuilla kentillä lehtien typpipitoisuus oli palautunut lähtötasolle, eli sen osalta rauduskoivuilla
havaittiin sopeutumista korkeampaan suhteelliseen ilmankosteuteen.
7 JOHTOPÄÄTÖKSET
Lyhytkestoisessa kammiokokeessa Vehmersalmi 14 -genotyyppiä edustavien rauduskoivujen
(Betula pendula Roth) mikrolisäyksellä tuotetut kuuden kuukauden ikäiset taimet altistettiin
suhteellisen ilmankosteuden ja kasvualustan typpipitoisuuden kahdelle eri tasolle. Tässä pro
gradu -tutkielmassa tarkasteltiin, kuinka edellä mainitut muuttujat vaikuttivat kammiokokeen
aikana kasvaneiden rauduskoivun lehtien metaboliaan sekä koivuntaimien kasvuun ja lehtien
ominaisuuksiin. Suhteellisen ilmankosteuden kohottamisella havaittiin olevan voimakas
vaikutus lehtien metaboliaan. Rauduskoivulla esiintyvien primaari- ja sekundaarimetaboliittien
pitoisuusmuutosten vasteet suhteellisen ilmankosteuden muutokseen vaihtelivat yhdiste-
ryhmien sisällä yhdisteestä riippuen, eivätkä vasteet olleet aina samalla metaboliareitilläkään

42
yhdenmukaisia. Useiden sokereiden, sokerialkoholien ja orgaanisten happojen sekä β-sitoste-
rolin ja salidrosidin pitoisuuden alenivat, ja raffinoosin, kversetiinin ja flavonoliglykosidin
pitoisuudet nousivat kohotetussa suhteellisessa ilmankosteudessa kasvatettujen rauduskoivujen
lehdissä. Metaboliamuutokset kohotetussa suhteellisessa ilmankosteudessa osoittivat, ettei
typensidonta toiminut yhtä tehokkaasti kuin hiilensidonta. Useissa tutkimuksissa on osoitettu,
että korkean suhteellisen ilmankosteuden vaikutuksesta ravinteiden kuljetus kasvissa vähenee.
Myös tässä tutkimuksessa havaittiin, että kasvatus korkeassa suhteellisessa ilmankosteudessa
ilmensi kasveissa typpirajoitettua kasvua ja typen puutosta metaboliamuutosten suhteen.
Kasvualustan kohotetussa typpipitoisuudessa kasvatettujen rauduskoivujen lehdissä
malaatin ja sukkinihapon pitoisuudet nousivat ja 3-trans-kumaryyli-kiinihapon pitoisuus aleni
verrattuna alemmassa typpipitoisuudessa kasvatettujen rauduskoivujen lehtien metaboliitti-
pitoisuuksiin. Kasvualustan typpilisän vaikutukset lehtien metaboliaan olivat siten määrällisesti
vähäisempiä kuin mitä kohotetussa suhteellisessa ilmankosteudessa kasvaneiden raudus-
koivujen lehdissä havaittiin. On mahdollista, että kokeen lyhyt kesto ja istutusaineksen typen
lähtötaso ovat heikentäneet kasvualustan kohotetun typpilisän vaikutusta metaboliamuutoksiin.
Suhteellisen ilmankosteuden kohottamisella oli voimakas vaikutus myös tutkielmassa
tarkasteltuihin koivuntaimen kasvu- ja lehtien ominaisuuksiin. Koivuntaimien kasvu kiihtyi
korkeammassa suhteellisessa ilmankosteudessa, mikä voi johtua hiiliylimäärän lisääntyneestä
allokoinnista biomassan tuotantoon typpirajoitetuissa olosuhteissa. Lehden typpipitoisuuden ja
klorofylli-indeksin aleneminen kohotetussa suhteellisessa ilmankosteudessa osoitti kasvien
rajoittunutta typen ottoa. Tämä pro gradu -tutkimus osoittaa suhteellisen ilmankosteuden
olevan hyvin tärkeä tekijä, joka vaikuttaa hiilen ja typen allokaatioon sekä metaboliaan
kasveissa.
Tässä pro gradu -tutkimuksessa analysoitujen näytteiden ja tutkielmassa tarkasteltavien
metaboliittien lukumäärä oli rajattu vastaamaan tutkielman laajuutta. Tarkastelujoukkoa
kasvattamalla saataisiin kattavampia tuloksia ja voitaisiin keskustella laajemmin tutkimus-
asetelmassa käytettyjen muuttujien (suhteellinen ilmankosteus ja kasvualustan typpipitoisuus)
vaikutuksesta Vehmersalmi 14 -genotyyppiä edustavien rauduskoivujen metaboliaan. Laajen-
tamalla metabolian tutkimista esimerkiksi entsyymi- ja transkriptiotasolle saataisiin moni-
puolisempia, mutta myös monimutkaisempia, tuloksia sekä havaittaisiin selkeämpiä yhteyksiä
reaktioreittien välillä. Käyttämällä tutkimuksessa useampaa genotyyppiä, voitaisiin tässä tutki-
muksessa saatuja tuloksia paremmin yleistää koskemaan laajempaa koivupopulaatiota.
Kasveissa on lisäksi muitakin syntetisoivia osia kuin lehdet. Juuriston ja rungon sisältämien
yhdisteiden sekä fenologisten rakenteiden muutosten tutkimus vasteena ilmastonmuutoksen

43
stressitekijöihin on yhtä lailla tärkeää kuin lehtien metabolian tutkiminen, jotta voisimme
laajemmin ymmärtää ilmastonmuutoksen stressitekijöiden kokonaisvaikutuksia kasvien sopeu-
tumiseen muuttuvassa ympäristössä.
8 KIITOKSET
Suuret kiitokset ohjaajilleni Jenna Lihavaiselle GC-MS -laitteiston käytön opastuksesta, tove-
ruudesta laboratoriossa sekä monipuolisesta ohjauksesta tilastomenetelmien parissa kamp-
paillessani, Sarita Keski-Saarelle täsmällisistä vinkeistä ja kommenteista, joiden avulla tutkiel-
man sisältöä on useaan otteeseen paranneltu sekä Sari Kontunen-Soppelalle kärsivällisyydestä
ja tuesta henkilökohtaisten vaikeuksieni tiellä. Tutkimuksen rahoitus oli saatu Suomen Akate-
mialta ja se oli myönnetty Elina Oksasen tutkimusprojektiin (projekti nro 250636). Kiitos
Hannele Hakuliselle, joka toteutti kammiokoivujen mikrolisäyksen. Kiitos myös Metsätieteen
laitokselle, jossa kammiokokeessa käytetyt kasvukammiot sijaitsivat. Lisäksi haluan kiittää
opiskelutovereitani ja läheisiä ystäviäni vertaistuesta ja kanssakulkemisesta opintovuosieni
aikana.
KIRJALLISUUSLUETTELO
Akula, R., Ravishankar, G.A. 2011: Influence of abiotic stress signals on secondary
metabolites in plants. – Plant Signaling & Behavior 6: 1720-1731.
Amtmann, A., Armengaud, P. 2009: Effects of N, P, K and S on metabolism: new knowledge
gained from multi-level analysis. – Current Opinion in Plant Biology 12: 275-283.
Barchet, G.L.H., Dauwe, R., Guy, R.D., Schroeder, W.R., Soolanayakanahally, R.Y.,
Campbell, M.M., Mansfield, G.S. 2013: Investigating the drought stress response of hybrid
poplar genotypes by metabolite profiling. – Tree Physiology 34: 1203-1219.
Bi, J.L., Felton, G.W. 1995: Foliar oxidative stress and insect herbivory: Primary compounds,
secondary metabolites, and reactive oxygen species as components of induced resistance.
– Journal of Chemical Ecology 21: 1511-1530.
Buchanan, B.B., Gruissem, W. & Jones, R. 2015: Biochemistry and Molecular Biology of
Plants. – 1264 s. John Wiley & Sons, Singapore.
Bylesjö, M., Segura, V., Soolanayakanahally, R.Y., Rae, A.M., Trygg, J., Gustafsson, P.,
Jansson, S., Street, N.R. 2008: LAMINA: a tool for rapid quantification of leaf size and
shape parameters. – BMC Plant Biology 8: 82.
Chen, T.-I., Dai, R. 2015: Chapter 6: Metabolomics data processing based on mass
spectrometry platforms. – Teoksessa: Qi, X., Chen, X. & Wang, Y. (toim.): Plant
Metabolomics: Methods and Applications: 123-169. Springer Netherlands, Dortrecht.
de la Rosa, T.M., Julkunen-Tiitto, R., Lehto, T., Aphalo, P.J. 2001: Secondary metabolites
and nutrient concentrations in silver birch seedlings under five levels of daily UV-B
exposure and two relative nutrient addition rates. – New Phytologist 150: 121-131.
Disraeli, B. 1996: General summary statistics: Lies, damned lies and statistics. – Teoksessa:
Upton, G. & Cook, I. (toim.): Understanding Statistics: 36-83. Oxford University Press,
Lontoo.
Duan, L.-X., Qi, X. 2015: Chapter 7: Metabolite qualitative methods and the introduction of
metabolomics database. – Teoksessa: Qi, X., Chen, X. & Wang, Y. (toim.): Plant
Metabolomics: Methods and Applications: 171-193. Springer Netherlands, Dortrecht.

44
Dumschott, K., Richter, A., Loescher, W., Merchant, A. 2017: Post photosynthetic carbon
partitioning to sugar alcohols and consequences for plant growth. – Phytochemistry 144:
243-252.
Fiehn, O., Kopka, J., Dörmann, P., Altmann, T., Trethewey, R.N., Willmitzer, L. 2000:
Metabolite profiling for plant functional genomics. – Nature Biotechnology 18: 1157-1161.
Fordham, M.C., Harrison-Murray, R.S., Knight, L., Evered, C.E. 2001: Effects of leaf wetting
and high humidity on stomatal function in leafy cuttings and intact plants of Corylus
maxima. – Physiologia Plantarum 113: 233-240.
Fritz, C., Mueller, C., Matt, P., Feil, R., Stitt, M. 2006: Impact of the C-N status on the amino
acid profile in tobacco source leaves. – Plant, Cell and Environment 29: 2055-2076.
Geiger, M., Haake, V., Ludewig, F., Sonnewald, U., Stitt, M. 1999: The nitrate and
ammonium nitrate supply have a major influence on the response of photosynthesis,
carbon metabolism, nitrogen metabolism and growth to elevated carbon dioxide in
tobacco. – Plant, Cell and Environment 22: 1177-1199.
Gislerød, H.R., Nelson, P.V. 1989: The interaction of relative air humidity and carbon dioxide
enrichment in the growth of Chrysanthemum × morifolium Ramat. – Scientia Horticulturae
38: 305-313.
Gislerød, H.R., Mortensen, L.M. 1990: Relative Humidity and Nutrient Concentration Affect
Nutrient Uptake and Growth of Begonia × hiemalis. – HortScience 25: 524-526.
Godbold, D., Tullus, A., Kupper, P., Sõber, J., Ostonen, I., Godbold, J. A., Lukac, M.,
Ahmed, I.U., Smith, A.R. 2014: Elevated atmospheric CO2 and humidity delay leaf fall in
Betula pendula, but not in Alnus glutinosa or Populus tremula×tremuloides. – Annals of
Forest Science 71: 831-842.
Hartley, S.E., Firn, R.D. 1989: Phenolic biosynthesis, leaf damage, and insect herbivory in
birch (Betula pendula). – Journal of Chemical Ecology 15: 275-283. Haukioja, E., Ossipov, V., Koricheva, J., Honkanen, T., Larsson, S., Lempa, K. 1998:
Biosynthetic origin of carbon-based secondary compounds: cause of variable responses of
woody plants to fertilization? – Chemoecology 8: 133-139.
Hill, C. B., Roessner, U. 2013: Metabolic Profiling of Plants by GC-MS. – Teoksessa:
Weckwerth, W. & Kahl, G. (toim.): The Handbook of Plant Metabolomics: 3-23. Wiley-
VCH Verlag GmbH & Co. KGaA, Singapore.
Hiller, K., Hangebrauk, J., Jäger, C., Spura, J., Schreiber, K., Schomburg, D. 2009:
MetaboliteDetector: Comprehensive analysis tool for targeted and nontargeted GC/MS
based metabolome analysis. – Analytical Chemistry 81: 3429-3439.
Hummel. J., Strehmel, N., Selbig, J., Walther, D., Kopka, J. 2010: Decision tree supported
substructure prediction of metabolites from GC-MS profiles. – Metabolomics 6: 322-333.
Hynynen, J. Niemistö, P., Viherä-Aarnio, A., Brunner, A., Hein. S., Velling, P. 2010:
Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in northern
Europe. – Forestry 83: 103-119.
Ilmatieteenlaitos 2017: Teematietoa: Sääennuste: Tunne termit - ymmärrä säätiedotus: Ilman
kosteus: http://ilmatieteenlaitos.fi/ilman-kosteus 12.10.2017.
Jaagus, J., Mändla, K. 2014: Climate change scenarios for Estonia based on climate models
from the IPCC Fourth Assessment Report. – Estonian Journal of Earth Sciences 63: 166-
180.
Jylhä, K., Tuomenvirta, H., Ruosteenoja, K. 2004: Climate change projections for Finland
during the 21st century. – Boreal Environment Research 9: 127-152.
Keinänen, M., Julkunen-Tiitto, R., Mutikainen, P., Walls, M., Ovaska, J., Vapaavuori, E.
1999: Trade-offs in phenolic metabolism of silver birch: Effects of fertilization, defoliation
and genotype. – Ecology 80: 1970-1986.

45
Keski-Saari, S., Julkunen-Tiitto, R. 2003: Resource allocation in different parts of juvenile
mountain birch plants: effect of nitrogen supply on seedling phenolics and growth.
– Physiologia Plantarum 118: 114-126.
Keski-Saari, S., Pusenius, J., Julkunen-Tiitto, R. 2005: Phenolic compounds in seedlings of
Betula pubescens and B. pendula are affected by enhanced UVB radiation and different
nitrogen regimens during early ontogeny. – Global Change Biology 11: 1180-1194.
Kont, A., Jaagus, J., Aunap, R. 2003: Climate change scenarios and the effect of sea-level rise
for Estonia. – Global and Planetary Change 36: 1-15.
Kopka, J., Schauer, N., Krueger, S., Birkemeyer, C., Usadel, B., Bergmüller, E., Dörmann, P.,
Weckwerth, W., Gibon, Y., Stitt, M., Willmitzer, L., Fernie, A.R., Steinhauser, D. 2004: [email protected]: the Golm Metabolome Database. – Bioinformatics 21: 1635-1638.
Koricheva, J., Larsson, S., Haukioja, E., Keinänen, M. 1998: Regulation of woody plant
secondary metabolism by resource availability: hypothesis testing by means of meta-
analysis. – Oikos 83: 212-226.
Krapp, A. 2015: Plant nitrogen assimilation and its regulation: a complex puzzle with missing
pieces. – Current Opinion in Plant Biology 25: 115-122.
Kupper, P., Sõber, J., Sellin, A., Lõhmus, K., Tullus, A., Räim, O., Lubenets, K., Tulva, I.,
Uri, V., Zobel, M., Kull, O., Sõber, A. 2011: An experimental facility for free air humidity
manipulation (FAHM) can alter water flux through deciduous tree canopy.
– Environmental and Experimental Botany 72: 432-438.
Kurepa, J., Shull, T.E., Smalle, J.A 2016: Quercetin feeding protects plants against oxidative
stress. – F1000Research 5: 2430. DOI: 10.12688/f1000research.9659.1
Kutchan, T.M., Gershenzon, J., Møller, B.L, Gang, D.R. 2015: Chapter 24: Natural products.
– Teoksessa: Buchanan, B.B., Gruissem, W. & Jones, R. (toim): Biochemistry and
Molecular Biology of Plants: 1132-1206. John Wiley & Sons, Singapore.
Lalonde, S., Wipf, D., Frommer, W.B. 2004: Transport mechanisms for organic forms of
carbon and nitrogen between source and sink. – Annual Review of Plant Biology 55: 341-
372.
Lavola, A. 1998: Accumulation of flavonoids and related compounds in birch induced by
UV-B irradiance. – Tree Physiology 18: 53-58.
Lavola, A., Julkunen-Tiitto, R., de la Rosa, T.M., Lehto, T., Aphalo, P.J. 2000: Allocation of
carbon to growth and secondary metabolites in birch seedlings under UV-B radiation and
CO2 exposure. – Physiologia Plantarum 109: 260-267.
Lemoine, R., La Camera, S., Atanassova, R., Dédaldéchamp, F., Allario, T., Pourtau, N.,
Bonnemain, J.-L., Laloi, M., Coutos-Thévenot, P., Maurousset, L., Faucher, M., Girousse,
C., Lemonnier, P., Parrilla, J., Durand, M. 2013: Source-to-sink transport of sugar and
regulation by environmental factors. – Frontiers in Plant Science 4: 272.
DOI: 10.3389/fpls.2013.00272.
Lihavainen, J., Ahonen, V., Keski-Saari, S., Kontunen-Soppela, S., Oksanen, E., Keinänen,
M. 2016a: Low vapour pressure deficit affects nitrogen nutrition and foliar metabolites in
silver birch. – Journal of Experimental Botany 67: 4353-4365.
Lihavainen, J., Keinänen, M., Keski-Saari, S., Kontunen-Soppela, S., Sõber, A., Oksanen, E.
2016b: Artificially decreased vapour pressure deficit in field conditions modifies foliar
metabolite profiles of birch and aspen. – Journal of Experimental Botany 67: 4367-4378.
Lu, C., Zhang, J. 2000: Photosynthetic CO2 assimilation, chlorophyll fluorescence and
photoinhibition as affected by nitrogen deficiency in maize plants. – Plant Science 151:
135-143.
Maeda, H., Dudareva, N. 2012: The shikimate pathway and aromatic amino acid biosynthesis
in plants. – Annual Review of Plant Biology 63: 73-105.

46
Martemyanov, V.V., Pavlushin, S.V., Dubovskiy, I.M., Belousova, I.A., Yushkova, Y.V.,
Morosov, S.V., Glupov, V.V. 2015: Leaf surface lipophilic compounds as one of the
factors of silver birch chemical defense against larvae of gypsy Moth. – PloS one 10(3):
e0121917. DOI: 10.1371/journal.pone.0121917.
Merchant, A., Richter, A.A. 2011: Polyols as biomarkers and bioindicators for 21st century
plant breeding. – Functional Plant Biology 38: 934-940.
Mewis, I., Schreiner, M., Nguyen, C.N., Krumbein, A., Ulrichs, C., Lohse, M., Zrenner, R.
2012: UV-B Irradiation Changes Specifically the Secondary Metabolite Profile in Broccoli
Sprouts: Induced Signaling Overlaps with Defense Response to Biotic Stressors. – Plant
and Cell Physiology 53: 1546-1560.
Mortensen, L.M. 1986: Effect of relative humidity on growth and flowering of some
greenhouse plants. – Scientia Horticulturae 29: 301-307.
Paul, M.J., Driscoll, S.P. 1997: Sugar repression of photosynthesis: the role of carbohydrates
in signaling nitrogen deficiency through source:sink imbalance. – Plant, Cell and
Environment 20: 110-116.
Ranta, E., Rita, H. & Kouki, J. 1994: Biometria – tilastotiedettä ekologeille. – 569 s.
Yliopistopaino, Helsinki.
Richardson, A.D., Duigan, S.P., Berlyn, G.P. 2002: An evaluation of noninvasive methods to
estimate foliar chlorophyll content. – New Phytologist 153: 185-194.
Rosenvald, K., Tullus, A., Ostonen, I., Uri, V., Kupper, P., Aosaar, J., Varik, M., Sõber, J.,
Niglas, A., Hansen, R., Rohula, G., Kukk, M., Sõber, A., Lõhmus, K. 2014: The effect of
elevated air humidity on young silver birch and hybrid aspen biomass allocation and
accumulation – Acclimation mechanisms and capacity. – Forest Ecology and Management
330: 252-260.
Schauer, N., Steinhauser, D., Strelkov, S., Schomburg, D., Allison, G., Moritz, T., Lundgren,
K., Roessner-Tunali, U., Forbes, M.G., Willmitzer, L., Fernie, A.R., Kopka, J. 2005: GC-
MS libraries for the rapid identification of metabolites in complex biological samples.
– FEBS Letters 579: 1332-1337.
Scheible, W-R, Gonzalez-Fontes, A., Lauerer, M., Müller-Röber, B., Caboche, M., Stitt, M.
1997: Nitrate acts as a signal to induce organic acid metabolism and repress starch
metabolism in tobacco. – The Plant Cell 9: 783-798.
Scheible, W-R., Morcuende, R., Czechowski, T., Fritz, C., Osuna, D., Palacios-Rojas, N.,
Schindelasch, D., Thimm, O., Udvardi, M.K., Stitt, M. 2004: Genome-Wide
Reprogramming of Primary and Secondary Metabolism, Protein Synthesis, Cellular
Growth Processes, and the Regulatory Infrastructure of Arabidopsis in Response to
Nitrogen. – Plant Physiology 136: 2483-2499.
Schlüter, U., Mascher, M., Colmsee, C., Scholz, U., Bräutigam, A., Fahnenstich, H.,
Sonnewald, U. 2012: Maize source leaf adaptation to nitrogen deficiency affects not only
nitrogen and carbon metabolism but also control of phosphate homeostasis. – Plant
Physiology 160: 1384-1406.
Schlüter, U., Colmsee, C., Scholz, U., Bräutigam, A., Weber, A.P.M., Zellerhoff, N., Bucher,
M., Fahnenstich, H., Sonnewald, U. 2013: Adaptation of maize source leaf metabolism to
stress related disturbances in carbon, nitrogen and phosphorus balance. – BMC Genomics
14: 442. DOI: 10.1186/1471-2164-14-442.
Schreiner, M., Mewis, I., Huyskens-Keil, S., Jansen, M.A.K., Zrenner, R., Winkler, J.B.,
O’Brien, N., Krumbein, A. 2012: UV-B-Induced Secondary Plant Metabolites - Potential
Benefits for Plant and Human Health. – Critical Reviews in Plant Sciences 31: 229-240.
Sellin, A., Tullus, A., Niglas, A., Õunapuu, E., Karusion, A., Lõhmus, K. 2013: Humidity-
driven changes in growth rate, photosynthetic capacity, hydraulic properties and other
functional traits in silver birch (Betula pendula). – Ecological Research 28: 523-535.

47
Sellin, A., Rosenvald, K., Õunapuu-Pikas, E., Tullus, A., Ostonen, I., Lõhmus, K. 2015:
Elevated air humidity affects hydraulic traits and tree size but not biomass allocation in
young silver birches (Betula pendula). – Frontiers in Plant Science 6: 860-871.
Sellin, A., Alber, M., Keinänen, M., Kupper, P., Lihavainen, J., Lõhmus, K., Oksanen, E.,
Sõber, A., Sõber, J., Tullus, A. 2017: Growth of northern deciduous trees under increasing
atmospheric humidity: possible mechanisms behind the growth retardation. – Regional
Environment Change 17: 2135-2148.
Sprenger, N., Keller, F. 2001: Allocation of raffinose family oligosaccharides to transport and
storage pools in Ajuga reptans: The roles of two distinct galactinol synthases. – Plant
Journal 21: 249-258.
Stocker, T.F., Qin, D., Plattner, G-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia,
Y., Bex, V. & Midgley, P.M.: IPCC 2013: Climate Change 2013: The Physical Science
Basis. Contribution of Working Group I to the Fifth Assessment Report of the
Intergovernmental Panel on Climate Change. – 1535 s. Cambridge University Press,
Cambridge. DOI: 10.1017/CBO9781107415324.
Sweetlove, L.J., Fernie, A.R. 2013: The Spatial Organization of Metabolism within the Plant
Cell. – Annual Review of Plant Biology 64: 723-746.
Taiz, L. & Zeiger, E. 2012: Plant Physiology and Development. – 778 s. Sinauer Associates
Inc., Yhdysvallat.
Tullus, A., Kupper, P., Sellin, A., Parts, L., Sõber, J, Tullus, T., Lõhmus, K., Sõber, A.,
Tullus, H. 2012: Climate change at northern latitudes: Rising atmospheric humidity
decreases transpiration, N-uptake and growth rate of hybrid aspen. – PLoS One 7(8):
e42648. DOI: 10.1371/journal.pone.0042648.
Tullus, A., Sellin, A., Kupper, P., Lutter, R., Pärn, L., Jasińska, A.K., Alber, M., Kukk, M.,
Tullus, T., Tullus, H., Lõhmus, K., Sõber, A. 2014: Increasing air humidity – a climate
trend predicted for northern latitudes – alters the chemical composition of stemwood in
silver birch and hybrid aspen. – Silva Fennica 48: 1-16.
Viherä-Aarnio, A. 2008: Koivun biologiset ja metsänhoidolliset ominaisuudet. – Teoksessa:
Niemistö, P., Viherä-Aarnio, A., Velling, P., Heräjärvi, H. & Verkasalo, E. (toim): Koivun
kasvatus ja käyttö: 20-30. Metsäntutkimuslaitos, Metsäkustannus. Karisto Oy,
Hämeenlinna.
Worley, B., Powers, R. 2013: Multivariate analysis in metabolomics. – Current Metabolomics
1: 92-107.
Xue, Z., Duan, L.-X., Qi, X. 2015: Gas Chromatography Mass Spectrometry coupling
techniques. – Teoksessa: Qi, X., Chen, X. & Wang, Y. (toim.): Plant Metabolomics:
Methods and Applications: 25-44. Springer Netherlands, Dortrecht.
Zar, J.H. 1999: Biostatistical analysis. – 663 s. Prentice Hall, Upper Saddle River.
LIITTEET
LIITE 1, 3 sivua
Taulukko L1. Analysoidut ja identifioidut metaboliitit, kvantifiointi-ioni, retentioindeksi, retentioaika
(min), käsittely (C = RH60 N35, N = RH60 N70, H = RH95 N35, HN = RH95 N70), havaintoyksiköiden
lukumäärä (n) käsittelyittäin (C, H, HN: n ≤ 10, N: n ≤ 11), metaboliitin pitoisuuksien normalisoitujen
arvojen keskiarvo (ka.) ja keskiarvon keskivirhe (SE).
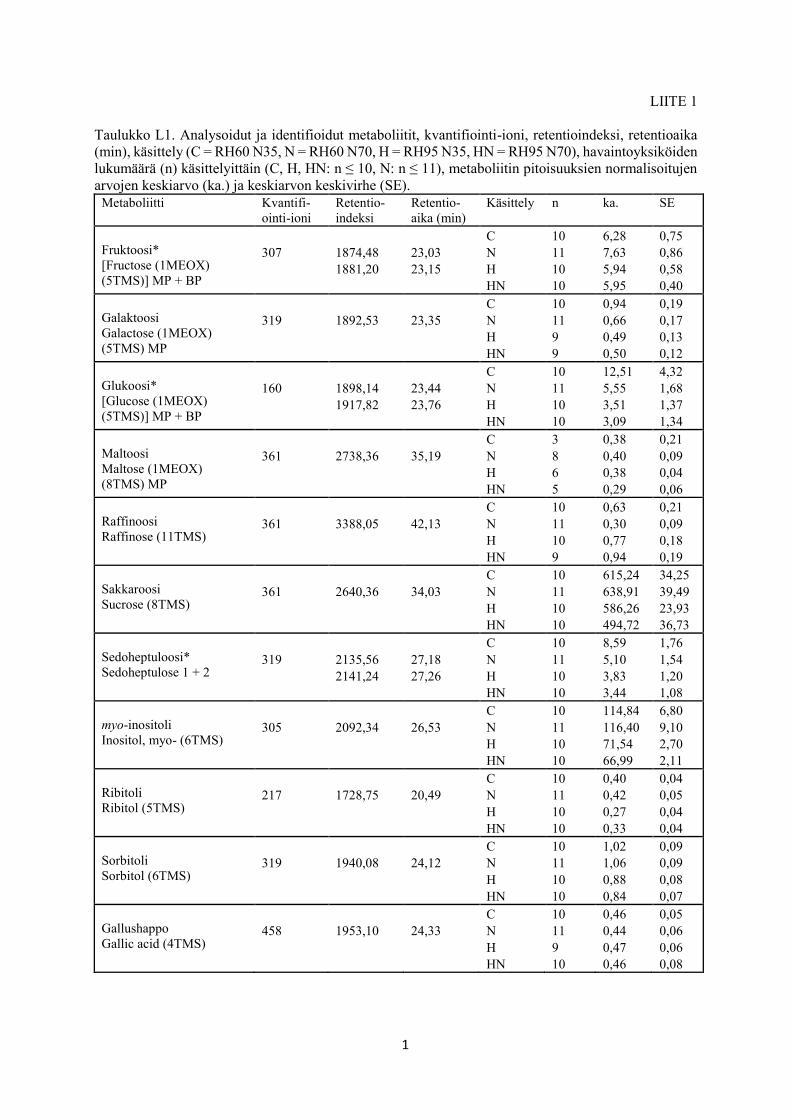
1
LIITE 1
Taulukko L1. Analysoidut ja identifioidut metaboliitit, kvantifiointi-ioni, retentioindeksi, retentioaika
(min), käsittely (C = RH60 N35, N = RH60 N70, H = RH95 N35, HN = RH95 N70), havaintoyksiköiden
lukumäärä (n) käsittelyittäin (C, H, HN: n ≤ 10, N: n ≤ 11), metaboliitin pitoisuuksien normalisoitujen
arvojen keskiarvo (ka.) ja keskiarvon keskivirhe (SE).
Metaboliitti Kvantifi-
ointi-ioni
Retentio-
indeksi
Retentio-
aika (min)
Käsittely n ka. SE
Fruktoosi*
[Fructose (1MEOX)
(5TMS)] MP + BP
307
1874,48
1881,20
23,03
23,15
C
N
H
HN
10
11
10
10
6,28
7,63
5,94
5,95
0,75
0,86
0,58
0,40
Galaktoosi
Galactose (1MEOX)
(5TMS) MP
319
1892,53
23,35
C
N
H
HN
10
11
9
9
0,94
0,66
0,49
0,50
0,19
0,17
0,13
0,12
Glukoosi*
[Glucose (1MEOX)
(5TMS)] MP + BP
160
1898,14
1917,82
23,44
23,76
C
N
H
HN
10
11
10
10
12,51
5,55
3,51
3,09
4,32
1,68
1,37
1,34
Maltoosi
Maltose (1MEOX)
(8TMS) MP
361
2738,36
35,19
C
N
H
HN
3
8
6
5
0,38
0,40
0,38
0,29
0,21
0,09
0,04
0,06
Raffinoosi
Raffinose (11TMS)
361
3388,05
42,13
C
N
H
HN
10
11
10
9
0,63
0,30
0,77
0,94
0,21
0,09
0,18
0,19
Sakkaroosi
Sucrose (8TMS)
361
2640,36
34,03
C
N
H
HN
10
11
10
10
615,24
638,91
586,26
494,72
34,25
39,49
23,93
36,73
Sedoheptuloosi*
Sedoheptulose 1 + 2
319
2135,56
2141,24
27,18
27,26
C
N
H
HN
10
11
10
10
8,59
5,10
3,83
3,44
1,76
1,54
1,20
1,08
myo-inositoli
Inositol, myo- (6TMS)
305
2092,34
26,53
C
N
H
HN
10
11
10
10
114,84
116,40
71,54
66,99
6,80
9,10
2,70
2,11
Ribitoli
Ribitol (5TMS)
217
1728,75
20,49
C
N
H
HN
10
11
10
10
0,40
0,42
0,27
0,33
0,04
0,05
0,04
0,04
Sorbitoli
Sorbitol (6TMS)
319
1940,08
24,12
C
N
H
HN
10
11
10
10
1,02
1,06
0,88
0,84
0,09
0,09
0,08
0,07
Gallushappo
Gallic acid (4TMS)
458
1953,10
24,33
C
N
H
HN
10
11
9
10
0,46
0,44
0,47
0,46
0,05
0,06
0,06
0,08
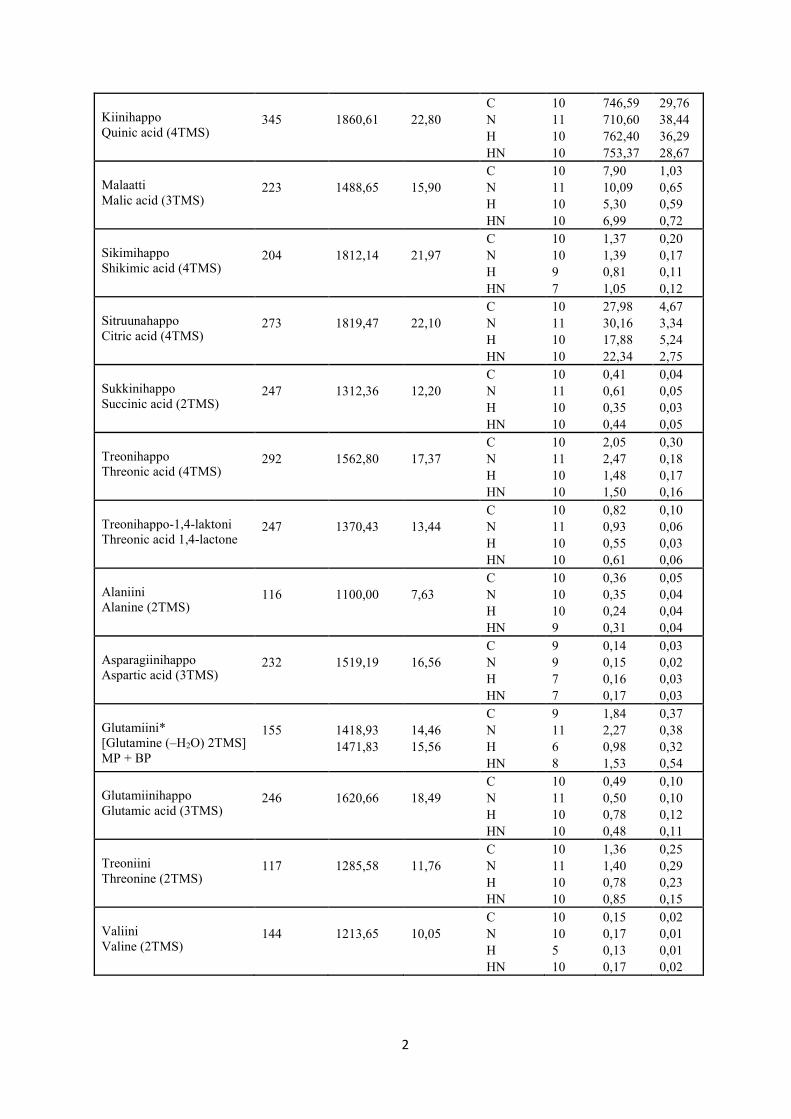
2
Kiinihappo
Quinic acid (4TMS)
345
1860,61
22,80
C
N
H
HN
10
11
10
10
746,59
710,60
762,40
753,37
29,76
38,44
36,29
28,67
Malaatti
Malic acid (3TMS)
223
1488,65
15,90
C
N
H
HN
10
11
10
10
7,90
10,09
5,30
6,99
1,03
0,65
0,59
0,72
Sikimihappo
Shikimic acid (4TMS)
204
1812,14
21,97
C
N
H
HN
10
10
9
7
1,37
1,39
0,81
1,05
0,20
0,17
0,11
0,12
Sitruunahappo
Citric acid (4TMS)
273
1819,47
22,10
C
N
H
HN
10
11
10
10
27,98
30,16
17,88
22,34
4,67
3,34
5,24
2,75
Sukkinihappo
Succinic acid (2TMS)
247
1312,36
12,20
C
N
H
HN
10
11
10
10
0,41
0,61
0,35
0,44
0,04
0,05
0,03
0,05
Treonihappo
Threonic acid (4TMS)
292
1562,80
17,37
C
N
H
HN
10
11
10
10
2,05
2,47
1,48
1,50
0,30
0,18
0,17
0,16
Treonihappo-1,4-laktoni
Threonic acid 1,4-lactone
247
1370,43
13,44
C
N
H
HN
10
11
10
10
0,82
0,93
0,55
0,61
0,10
0,06
0,03
0,06
Alaniini
Alanine (2TMS)
116
1100,00
7,63
C
N
H
HN
10
10
10
9
0,36
0,35
0,24
0,31
0,05
0,04
0,04
0,04
Asparagiinihappo
Aspartic acid (3TMS)
232
1519,19
16,56
C
N
H
HN
9
9
7
7
0,14
0,15
0,16
0,17
0,03
0,02
0,03
0,03
Glutamiini*
[Glutamine (–H2O) 2TMS]
MP + BP
155
1418,93
1471,83
14,46
15,56
C
N
H
HN
9
11
6
8
1,84
2,27
0,98
1,53
0,37
0,38
0,32
0,54
Glutamiinihappo
Glutamic acid (3TMS)
246
1620,66
18,49
C
N
H
HN
10
11
10
10
0,49
0,50
0,78
0,48
0,10
0,10
0,12
0,11
Treoniini
Threonine (2TMS)
117
1285,58
11,76
C
N
H
HN
10
11
10
10
1,36
1,40
0,78
0,85
0,25
0,29
0,23
0,15
Valiini
Valine (2TMS)
144
1213,65
10,05
C
N
H
HN
10
10
5
10
0,15
0,17
0,13
0,17
0,02
0,01
0,01
0,02
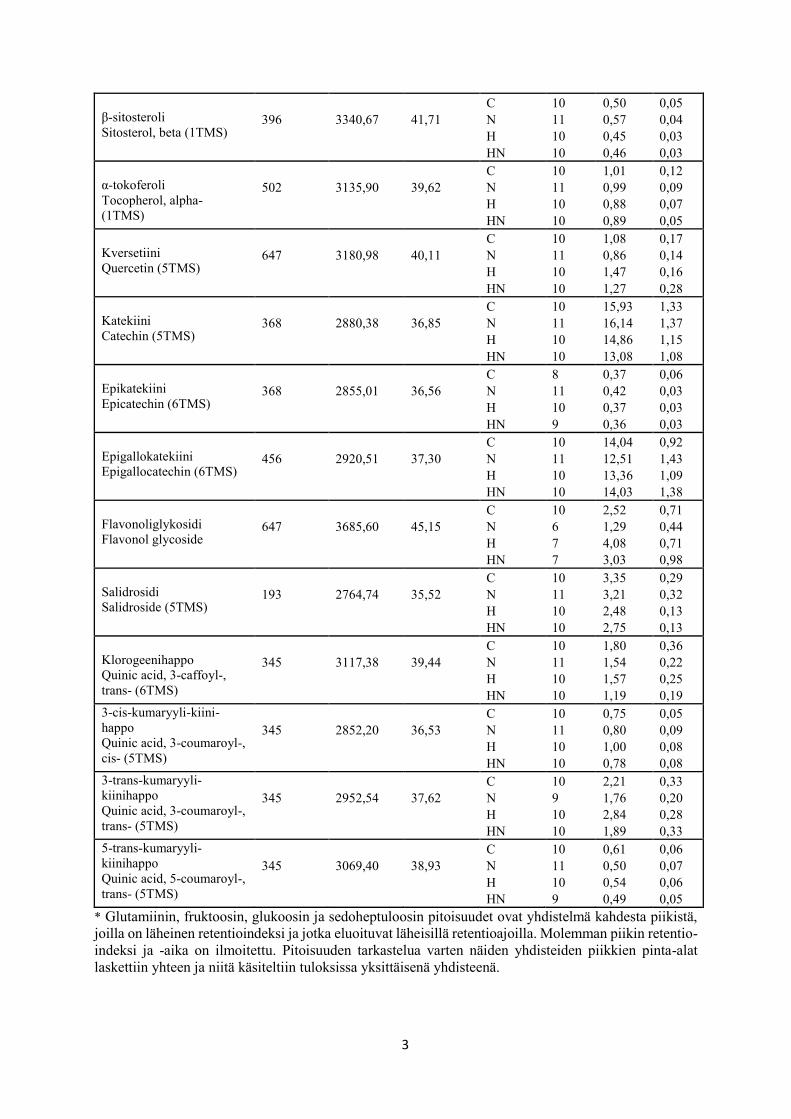
3
β-sitosteroli
Sitosterol, beta (1TMS)
396
3340,67
41,71
C
N
H
HN
10
11
10
10
0,50
0,57
0,45
0,46
0,05
0,04
0,03
0,03
α-tokoferoli
Tocopherol, alpha-
(1TMS)
502
3135,90
39,62
C
N
H
HN
10
11
10
10
1,01
0,99
0,88
0,89
0,12
0,09
0,07
0,05
Kversetiini
Quercetin (5TMS)
647
3180,98
40,11
C
N
H
HN
10
11
10
10
1,08
0,86
1,47
1,27
0,17
0,14
0,16
0,28
Katekiini
Catechin (5TMS)
368
2880,38
36,85
C
N
H
HN
10
11
10
10
15,93
16,14
14,86
13,08
1,33
1,37
1,15
1,08
Epikatekiini
Epicatechin (6TMS)
368
2855,01
36,56
C
N
H
HN
8
11
10
9
0,37
0,42
0,37
0,36
0,06
0,03
0,03
0,03
Epigallokatekiini
Epigallocatechin (6TMS)
456
2920,51
37,30
C
N
H
HN
10
11
10
10
14,04
12,51
13,36
14,03
0,92
1,43
1,09
1,38
Flavonoliglykosidi
Flavonol glycoside
647
3685,60
45,15
C
N
H
HN
10
6
7
7
2,52
1,29
4,08
3,03
0,71
0,44
0,71
0,98
Salidrosidi
Salidroside (5TMS)
193
2764,74
35,52
C
N
H
HN
10
11
10
10
3,35
3,21
2,48
2,75
0,29
0,32
0,13
0,13
Klorogeenihappo
Quinic acid, 3-caffoyl-,
trans- (6TMS)
345
3117,38
39,44
C
N
H
HN
10
11
10
10
1,80
1,54
1,57
1,19
0,36
0,22
0,25
0,19
3-cis-kumaryyli-kiini-
happo
Quinic acid, 3-coumaroyl-,
cis- (5TMS)
345
2852,20
36,53
C
N
H
HN
10
11
10
10
0,75
0,80
1,00
0,78
0,05
0,09
0,08
0,08
3-trans-kumaryyli-
kiinihappo
Quinic acid, 3-coumaroyl-,
trans- (5TMS)
345
2952,54
37,62
C
N
H
HN
10
9
10
10
2,21
1,76
2,84
1,89
0,33
0,20
0,28
0,33
5-trans-kumaryyli-
kiinihappo
Quinic acid, 5-coumaroyl-,
trans- (5TMS)
345
3069,40
38,93
C
N
H
HN
10
11
10
9
0,61
0,50
0,54
0,49
0,06
0,07
0,06
0,05
* Glutamiinin, fruktoosin, glukoosin ja sedoheptuloosin pitoisuudet ovat yhdistelmä kahdesta piikistä,
joilla on läheinen retentioindeksi ja jotka eluoituvat läheisillä retentioajoilla. Molemman piikin retentio-
indeksi ja -aika on ilmoitettu. Pitoisuuden tarkastelua varten näiden yhdisteiden piikkien pinta-alat
laskettiin yhteen ja niitä käsiteltiin tuloksissa yksittäisenä yhdisteenä.