specificity of baculovirus p6.9 basic dna-binding proteins and
TRANSCRIPT

JOURNAL OF VIROLOGY, Sept. 2010, p. 8821–8828 Vol. 84, No. 170022-538X/10/$12.00 doi:10.1128/JVI.00072-10Copyright © 2010, American Society for Microbiology. All Rights Reserved.
Specificity of Baculovirus P6.9 Basic DNA-Binding Proteins andCritical Role of the C Terminus in Virion Formation�†
Manli Wang,1,2 Era Tuladhar,1 Shu Shen,2 Hualin Wang,2 Monique M. van Oers,1Just M. Vlak,1* and Marcel Westenberg1,3
Laboratory of Virology, Wageningen University, Droevendaalsesteeg 1, 6708 PB Wageningen, Netherlands1; State Key Laboratory ofVirology, Key Laboratory of Molecular Virology and Joint Laboratory of Invertebrate Virology, Wuhan Institute of
Virology,Chinese Academy of Sciences, Wuhan 430071, People’s Republic of China2; and Pharmaceutical Science ResearchDivision, King’s College London, Franklin-Wilkins Building, 150 Stamford Street, London SE1 9NH, United Kingdom3
Received 13 January 2010/Accepted 21 May 2010
The majority of double-stranded DNA (dsDNA) viruses infecting eukaryotic organisms use host- or virus-expressed histones or protamine-like proteins to condense their genomes. In contrast, members of the Bacu-loviridae family use a protamine-like protein named P6.9. The dephosphorylated form of P6.9 binds to DNA ina non-sequence-specific manner. By using a p6.9-null mutant of Autographa californica multiple nucleopoly-hedrovirus (AcMNPV), we demonstrate that P6.9 is not required for viral DNA replication but is essential forthe production of infectious virus. Virion production was rescued by P6.9 homologs from a number ofAlphabaculovirus species and one Gammabaculovirus species but not from the genus Betabaculovirus, comprisingthe granuloviruses, or by the P6.9 homolog VP15 from the unrelated white spot syndrome virus of shrimp.Mutational analyses demonstrated that AcMNPV P6.9 with a conserved 11-residue deletion of the C terminuswas not capable of rescuing p6.9-null AcMNPV, while a chimeric Betabaculovirus P6.9 containing the P6.9C-terminal region of an Alphabaculovirus strain was able to do so. This implies that the C terminus ofbaculovirus P6.9 contains sequence elements essential for virion formation. Such elements may possiblyinteract with species- or genus-specific domains of other nucleocapsid proteins during virus assembly.
Condensation and packaging of viral DNA are integral andnecessary features of double-stranded (ds) viral assemblywithin infected eukaryotic host cells. To mediate DNA con-densation during viral assembly, dsDNA viruses may function-ally coopt host histone proteins, as do the Polyomaviridae (13,40) and Papillomaviridae (36), or express their own protamine-like protein with putative DNA condensation functions, as dothe Adenoviridae and Asfarviridae (6, 29). In contrast, membersof the Baculoviridae and Nimaviridae, which are exclusivelypathogenic for arthropods, predominantly within the ordersLepidoptera and Decapoda (1, 25), neither utilize host histonesnor express their own histone-like proteins but instead expressa single, functionally equivalent protamine-like protein, P6.9(41, 56). The p6.9 gene, which is present in all 50 baculovirusgenomes sequenced to date (46), encodes a small arginine- andserine-rich protein that, depending on the species, contains 49to 109 residues (see the supplemental material). Evidence in-dicates that P6.9 is prevented from binding to DNA as a resultof posttranslational phosphorylation of arginine and serineresidues (23, 53, 54). Upon viral assembly in the host nucleus,P6.9 is dephosphorylated, promoting DNA binding and en-abling condensation of the viral genome and packaging intothe viral nucleocapsid (14, 22, 28). In a newly infected cell, andfollowing rephosphorylation by a capsid-associated kinase,
P6.9 dissociates from the viral DNA, thereby releasing the viralgenome into the nucleus (14, 42, 54). At this stage, cellularhistones bind to the viral DNA, forming nucleosomes and anactive transcriptional complex (55).
The binding of P6.9 to DNA is apparently species indepen-dent, as evidenced by the facts that Heliothis zea single nucleo-polyhedrovirus (HezeNPV) P6.9 can bind �-DNA (22) and theP6.9 homolog VP15 of the unrelated white spot syndrome virus(WSSV) is able to bind Autographa californica multiple nucle-opolyhedrovirus (AcMNPV) DNA (56). Although DNA bind-ing by P6.9 exhibits a degree of species and sequence indepen-dence, we were interested in learning whether P6.9 possessesany species specificity for overall virion formation. We reporthere the results of experiments designed to examine whetherAcMNPV P6.9 can be functionally replaced either by P6.9-likeproteins from phylogenetically distinct baculovirus species orby WSSV VP15. Furthermore, by constructing AcMNPVgenomes encoding either P6.9 proteins with C-terminal deletionsor point mutations, or chimeric P6.9 fusion proteins, we dem-onstrate that the C-terminal domain of P6.9 appears to be anessential genus-specific feature required for infectious virionformation.
MATERIALS AND METHODS
General molecular methods. Classical restriction enzyme (RE)-based geneticengineering techniques, preparation of media, and related procedures wereperformed as described elsewhere (38a) unless otherwise stated. All PCRs wereperformed in volumes of 50 �l containing 15 pmol of each oligonucleotideprimer and 200 �M each deoxynucleoside triphosphate (dNTP) and were cata-lyzed with a high-fidelity DNA polymerase (Expand long-template PCR system[Roche] or Phusion [Finnzymes]). Recombineering, utilizing lambda phage re-combinases supplied via plasmid pBAD-��� (31), was performed essentially asdescribed previously (36). All plasmids and bacmid constructs generated during
* Corresponding author. Mailing address: Laboratory of Virology,Wageningen University, Droevendaalsesteeg 1, 6708 PB Wageningen,Netherlands. Phone: 31-317-483090. Fax: 31-317-484820. E-mail: [email protected].
† Supplemental material for this article may be found at http://jvi.asm.org/.
� Published ahead of print on 2 June 2010.
8821
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
the study were sequenced and/or analyzed by PCR to confirm constructionfidelity.
Cell culture and virus harvesting. The Spodoptera frugiperda cell line IPLB-Sf21 (47) was cultured at 27°C in plastic culture flasks (Nunc) in Grace’s insectmedium (pH 5.9 to 6.1; Invitrogen) supplemented with 10% fetal bovine serum(FBS). Viruses were harvested from culture supernatants following clarification(2,200 � g, 10 min) and filter sterilization (pore size, 0.45 �m).
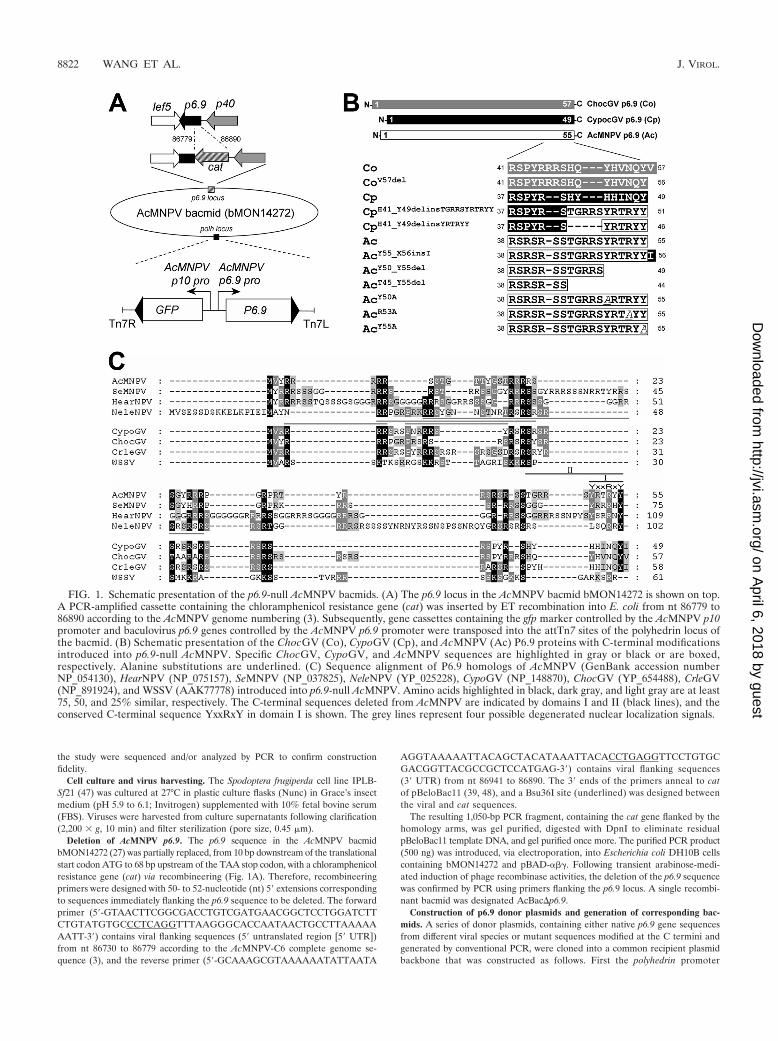
Deletion of AcMNPV p6.9. The p6.9 sequence in the AcMNPV bacmidbMON14272 (27) was partially replaced, from 10 bp downstream of the translationalstart codon ATG to 68 bp upstream of the TAA stop codon, with a chloramphenicolresistance gene (cat) via recombineering (Fig. 1A). Therefore, recombineeringprimers were designed with 50- to 52-nucleotide (nt) 5� extensions correspondingto sequences immediately flanking the p6.9 sequence to be deleted. The forwardprimer (5�-GTAACTTCGGCGACCTGTCGATGAACGGCTCCTGGATCTTCTGTATGTGCCCTCAGGTTTAAGGGCACCAATAACTGCCTTAAAAAAATT-3�) contains viral flanking sequences (5� untranslated region [5� UTR])from nt 86730 to 86779 according to the AcMNPV-C6 complete genome se-quence (3), and the reverse primer (5�-GCAAAGCGTAAAAAATATTAATA
AGGTAAAAATTACAGCTACATAAATTACACCTGAGGTTCCTGTGCGACGGTTACGCCGCTCCATGAG-3�) contains viral flanking sequences(3� UTR) from nt 86941 to 86890. The 3� ends of the primers anneal to catof pBeloBac11 (39, 48), and a Bsu36I site (underlined) was designed betweenthe viral and cat sequences.
The resulting 1,050-bp PCR fragment, containing the cat gene flanked by thehomology arms, was gel purified, digested with DpnI to eliminate residualpBeloBac11 template DNA, and gel purified once more. The purified PCR product(500 ng) was introduced, via electroporation, into Escherichia coli DH10B cellscontaining bMON14272 and pBAD-���. Following transient arabinose-medi-ated induction of phage recombinase activities, the deletion of the p6.9 sequencewas confirmed by PCR using primers flanking the p6.9 locus. A single recombi-nant bacmid was designated AcBac�p6.9.
Construction of p6.9 donor plasmids and generation of corresponding bac-mids. A series of donor plasmids, containing either native p6.9 gene sequencesfrom different viral species or mutant sequences modified at the C termini andgenerated by conventional PCR, were cloned into a common recipient plasmidbackbone that was constructed as follows. First the polyhedrin promoter
FIG. 1. Schematic presentation of the p6.9-null AcMNPV bacmids. (A) The p6.9 locus in the AcMNPV bacmid bMON14272 is shown on top.A PCR-amplified cassette containing the chloramphenicol resistance gene (cat) was inserted by ET recombination into E. coli from nt 86779 to86890 according to the AcMNPV genome numbering (3). Subsequently, gene cassettes containing the gfp marker controlled by the AcMNPV p10promoter and baculovirus p6.9 genes controlled by the AcMNPV p6.9 promoter were transposed into the attTn7 sites of the polyhedrin locus ofthe bacmid. (B) Schematic presentation of the ChocGV (Co), CypoGV (Cp), and AcMNPV (Ac) P6.9 proteins with C-terminal modificationsintroduced into p6.9-null AcMNPV. Specific ChocGV, CypoGV, and AcMNPV sequences are highlighted in gray or black or are boxed,respectively. Alanine substitutions are underlined. (C) Sequence alignment of P6.9 homologs of AcMNPV (GenBank accession numberNP_054130), HearNPV (NP_075157), SeMNPV (NP_037825), NeleNPV (YP_025228), CypoGV (NP_148870), ChocGV (YP_654488), CrleGV(NP_891924), and WSSV (AAK77778) introduced into p6.9-null AcMNPV. Amino acids highlighted in black, dark gray, and light gray are at least75, 50, and 25% similar, respectively. The C-terminal sequences deleted from AcMNPV are indicated by domains I and II (black lines), and theconserved C-terminal sequence YxxRxY in domain I is shown. The grey lines represent four possible degenerated nuclear localization signals.
8822 WANG ET AL. J. VIROL.
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

(polPROM) was deleted from pFastBac-1 (Invitrogen) by Bst1107I/StuI digestion,and the vector backbone was religated. The AcMNPV p6.9 promoter sequence,PCR amplified from pAcMP1 (18) with primers 5�-GGTCGACGTACCAAATTCCGTTTTGCGACG-3� and 5�-GGTCGACGGATCCGTTTAAATTGTGTAATTTATG-3� (underlined and italicized sequences are SalI and BamHI sites,respectively), was cloned, as a SalI fragment, into this polPROM minus vector.Subsequently, the p6.9 promoter sequence was removed as a SnaBI/BamHIfragment and was introduced, via the Bst1107I and BamHI sites, into pFastBacDUAL (Invitrogen), thereby deleting the vector’s polPROM sequence. Finally, agreen fluorescent protein (GFP) (smRS-GFP) (11) sequence was cloned, at aunique XmaI site, downstream of the p10 promoter in this pFastBacDUALderivative to yield the final base donor plasmid pFB-GFP-p6.9.
Native and mutant p6.9 sequences were generated by PCR amplification usingeither total or cloned fragments of the appropriate genomic DNA as templatesand forward and reverse primers (see Table S1 in the supplemental material)containing EcoRI and NotI sites at their respective 5� ends. p6.9 sequencescontaining modified C termini were generated by including the respective se-quence change within the 5� end of the reverse primer (see Table S1 in thesupplemental material). Initially, native p6.9 sequences of AcMNPV, Spodopteraexigua MNPV (SeMNPV), Helicoverpa armigera NPV (HearNPV), Neodiprionlecontei NPV (NeleNPV), Choristoneura occidentalis granulovirus (ChocGV),Cydia pomonella GV (CypoGV), and Cryptophlebia leucotreta GV (CrleGV), aswell as the vp15 sequence of WSSV, were generated. Subsequently, PCR ampli-fication using appropriate primer pairs (see Table S1 in the supplemental ma-terial) and either AcMNPV, ChocGV, or CypoGV DNA as a template generatedthe following series of modified p6.9 sequences: for AcMNPV, an isoleucineinsertion after tyrosine 55 (Y55_X56insI), deletion of residues 50 to 55(Y50_Y55del), deletion of residues 45 to 55 (T45_Y55del), and alanine substi-tution for residue 50 (Y50A), 53 (R53A), or 55 (Y55A); for ChocGV, deletionof valine 57 (V57del); for CypoGV, replacement of residues 41 to 47 by either 6(H41_Y49delinsYRTRYY) or 11 (H41_Y49delinsTGRRSYRTRYY) C-termi-nal residues of AcMNPV P6.9 (Fig. 1B). All amplicons were cloned into pFB-GFP-p6.9, and the resulting donor plasmids were introduced into AcBac�p6.9via Tn7-mediated transposition (Bac-to-Bac; Invitrogen) (Fig. 1A). The fidelityof the bacmids generated was confirmed by PCR as described previously (52).
Transfection-infection assay. DNA (approximately 1 �g) of each recombinantbacmid was transfected (10 �l Cellfectin; Invitrogen) into 1.5 � 106 Sf21 cells.Five days posttransfection (p.t.), cells were examined for GFP expression byfluorescence microscopy. An aliquot (500 �l) of the harvest supernatant was thenused to infect a new batch of Sf21 cells (1.5 � 106 in 2 ml medium). At 72 hpostinfection (p.i.), cells were again inspected for GFP expression.
One-step growth curves. For each recombinant virus, the production of bud-ded virus (BV) was monitored by constructing a viral growth curve as follows.Sf21 cells (1.5 � 105 per well; 24-well plates) were infected at a multiplicity ofinfection (MOI) of 10 50% tissue culture infective dose (TCID50) units/celland were then incubated (1 h, 27°C). After infection, the inoculum wasremoved, and the cells were washed three times (0.5 ml medium). At 0, 6, 12,18, 24, 48, and 72 h p.i the infected cell supernatants were collected. For eachtime point p.i. and each virus sample, triplicate samples were generated. Theconcentration of infectious BVs in each sample was determined by an end-point dilution assay on Sf21 cells (34).
Electron microscopy. Sf21 cells, collected at 72 h p.t. by centrifugation (2,000 �g, 10 min), were fixed (with 2.5% [wt/vol] glutaraldehyde in 0.1 M sodiumphosphate, pH 7.2 [NaPi], for 16 h at 4°C), washed (twice in NaPi for 15 min eachtime), further fixed (1% OsO4 in NaPi, 2 h, room temperature [RT]), washed (asdescribed above), and dehydrated (8 immersions [15 min each] in increasingconcentrations of ethanol [30 to 100%]). Specimens were embedded in capsulesand were polymerized (60°C, 48 h), and sections (thickness, 60 to 80 nm) werestained (2% [wt/vol] uranyl acetate, 15 min; lead citrate, 15 min) and observed bytransmission electron microscopy (FEI Tecnai G2 microscope at 200 kV).
qPCR analysis of BV release and viral DNA replication. To isolate BV DNAvirions, supernatants from harvested (0 and 48 h p.t.) cultures of Sf21 cellstransfected with different bacmid DNAs (1.0 �g, 1.0 � 106 cells) were treatedwith equal volumes of polyethylene glycol 8000 (PEG 8000) (20% [wt/vol] in1 M NaCl; 30 min, RT). Then 500-�l aliquots were collected by centrifugation(12,000 � g, 15 min), resuspended (20 �l H2O), and lysed by incubation(50°C, 1 h) in virus disruption buffer (80 �l; 10 mM Tris-HCl [pH 7.6], 10 mMEDTA, 0.25% sodium dodecyl sulfate [SDS]) and proteinase K (5 �l; 20 mg/ml).Lysed BVs were phenol extracted, and viral DNA was ethanol precipitated andresuspended (40 �l H2O). Total cellular DNA from pelleted and washed (threetimes, with 1 ml Grace’s medium) Sf21 cells was isolated with a commercialsystem (Genomic DNA rapid isolation kit; BioDev, China) according to themanufacturer’s instructions and was then incubated (4 h) with DpnI (10 U; New
England Biolabs) to digest residual bacmid DNA. BV DNA (5 �l) andDpnI-treated cellular DNA (5 �l) were used as templates in quantitative PCR(qPCR) analyses, performed as described previously (49), to determine viralcopy numbers.
Computer-assisted sequence analysis. Homologous sequences in theGenBank/EMBL databases were identified with the FASTA and BLAST programs(2, 35). Sequence alignments were performed with the ClustalW program(EMBL–European Bioinformatics Institute [http://www.ebi.ac.uk]) and were editedwith GeneDoc software (32). Putative nuclear localization signals were foundwith the PSORT II program (Human Genome Center, Institute for MedicalScience, University of Tokyo, Tokyo, Japan) (http://psort.ims.u-tokyo.ac.jp).
RESULTS
Disruption of p6.9 in an AcMNPV bacmid. Baculovirus P6.9plays an important role in condensing the viral genome in thenucleocapsid (14, 22, 28). The binding of these small, positivelycharged proteins to DNA seems not to be sequence specific(22). However, P6.9 might still have a species-specific role inthe viral assembly process. To investigate this possibility, theAcMNPV bacmid bMON14272 (27) was modified by deletingp6.9, generating AcBac�p6.9, followed by insertion of eitherintact homologous sequences from other species or p6.9 se-quences modified at their C termini.
Because p6.9 overlaps with late essential factor 5 (lef-5) inthe AcMNPV genome (3), the p6.9 open reading frame (ORF)of the AcMNPV bacmid bMON14272 was only partially de-leted to avoid potentially disrupting the function of lef-5. Toconfirm that this partial deletion had inactivated p6.9 and thatits replacement with cat had not functionally compromised theadjacent lef-5 and p40 genes, AcBac�p6.9 was fitted with cas-settes from either the GFP-expressing but empty donor plas-mid pFB-GFP-p6.9 or a derivative containing the completeAcMNPV p6.9 sequence under the control of the native p6.9promoter. Although Sf21 cells transfected with either bacmidexpressed GFP (Fig. 2A and B, upper panels), infectious viri-ons were produced only with the p6.9 rescue bacmid (Fig. 2Aand B, lower panels). Electron microscopic analysis of p6.9-null AcMNPV-transfected cells showed that no nucleocapsidswere formed (Fig. 3A). Instead, electron-lucent tubule-likestructures were present in the nucleus. These structures werenot present in cells transfected with the rescue bacmid, wherenormal nucleocapsid formation took place (Fig. 3B). Further-more, the kinetics of virion production of the rescue bacmid, asdetermined by one-step growth curve analysis, was similar(P, 0.05 by a two-tailed Student t test) to that of theparental AcMNPV bacmid provided with the GFP cassettealone (Fig. 4A). Taken together, these data indicate that p6.9was functionally inactivated by its partial deletion and that lef-5and p40 were not affected by the presence of cat.
P6.9 proteins from alpha- and gammabaculoviruses rescuep6.9-null AcMNPV. Recently, a new baculovirus classificationhas been proposed, consisting of four genera: Alphabaculovirus(lepidopteran NPVs), Betabaculovirus (lepidopteran granulo-viruses [GVs]), Gammabaculovirus (hymenopteran NPVs),and Deltabaculovirus (dipteran NPVs) (21). Based on phyloge-netic criteria and the type of envelope fusion protein (GP64 orF), the genus Alphabaculovirus can be phylogenetically subdi-vided further into groups I and II, respectively (8, 15–17). Toinvestigate whether p6.9 genes from phylogenetically distinctbaculoviruses could rescue p6.9-null AcMNPV (Alphabaculo-virus, group I), the p6.9 proteins of the group II Alphabaculo-
VOL. 84, 2010 ROLE OF THE BACULOVIRUS P6.9 C TERMINUS 8823
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
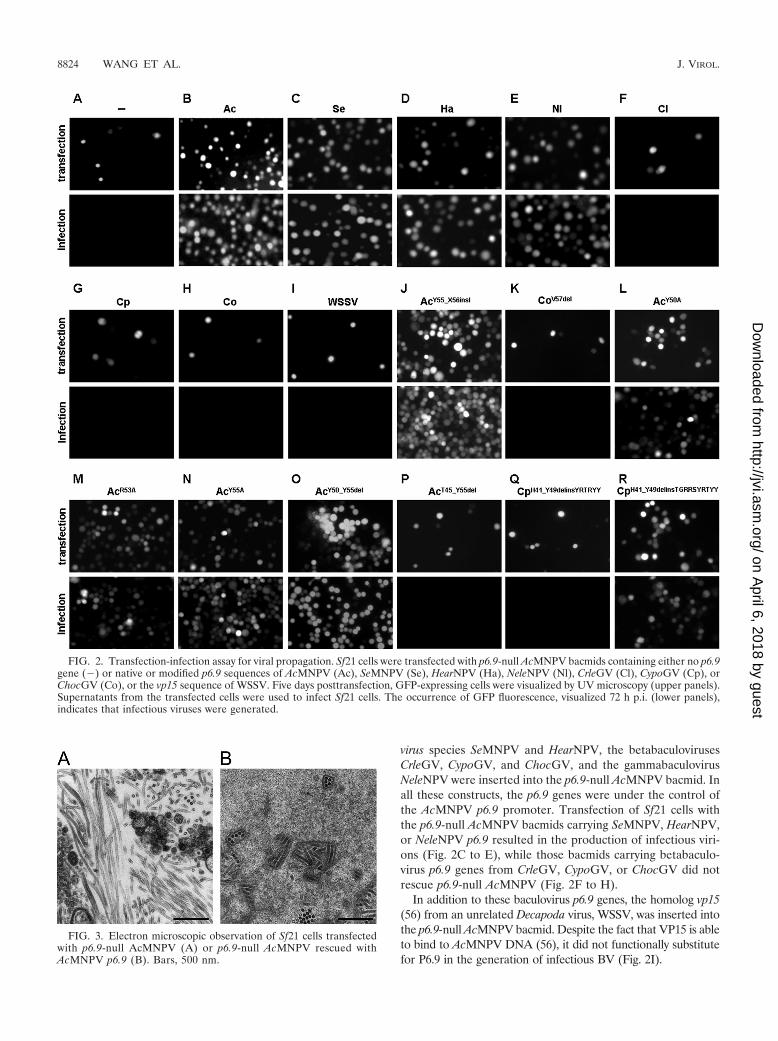
virus species SeMNPV and HearNPV, the betabaculovirusesCrleGV, CypoGV, and ChocGV, and the gammabaculovirusNeleNPV were inserted into the p6.9-null AcMNPV bacmid. Inall these constructs, the p6.9 genes were under the control ofthe AcMNPV p6.9 promoter. Transfection of Sf21 cells withthe p6.9-null AcMNPV bacmids carrying SeMNPV, HearNPV,or NeleNPV p6.9 resulted in the production of infectious viri-ons (Fig. 2C to E), while those bacmids carrying betabaculo-virus p6.9 genes from CrleGV, CypoGV, or ChocGV did notrescue p6.9-null AcMNPV (Fig. 2F to H).
In addition to these baculovirus p6.9 genes, the homolog vp15(56) from an unrelated Decapoda virus, WSSV, was inserted intothe p6.9-null AcMNPV bacmid. Despite the fact that VP15 is ableto bind to AcMNPV DNA (56), it did not functionally substitutefor P6.9 in the generation of infectious BV (Fig. 2I).
FIG. 3. Electron microscopic observation of Sf21 cells transfectedwith p6.9-null AcMNPV (A) or p6.9-null AcMNPV rescued withAcMNPV p6.9 (B). Bars, 500 nm.
FIG. 2. Transfection-infection assay for viral propagation. Sf21 cells were transfected with p6.9-null AcMNPV bacmids containing either no p6.9gene () or native or modified p6.9 sequences of AcMNPV (Ac), SeMNPV (Se), HearNPV (Ha), NeleNPV (Nl), CrleGV (Cl), CypoGV (Cp), orChocGV (Co), or the vp15 sequence of WSSV. Five days posttransfection, GFP-expressing cells were visualized by UV microscopy (upper panels).Supernatants from the transfected cells were used to infect Sf21 cells. The occurrence of GFP fluorescence, visualized 72 h p.i. (lower panels),indicates that infectious viruses were generated.
8824 WANG ET AL. J. VIROL.
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

For the rescued viruses, one-step growth curves were madeto determine whether the heterologous p6.9 genes had anyeffect on the kinetics of BV production (Fig. 4A). Surprisingly,only the virus carrying HearNPV p6.9 was significantly im-paired in BV production at 72 h p.i. (P, �0.05 by a two-tailedStudent t test), whereas virus production with p6.9 fromSeMNPV or the more phylogenetically distinct NeleNPV (Gam-mabaculovirus) was similar (P, 0.05 by a two-tailed Student ttest) to that of AcMNPV p6.9.
qPCR analysis of BV release and viral DNA replication. Thep6.9-null AcMNPV bacmids that either had no gene (�p6.9) orcarried Betabaculovirus p6.9 or WSSV vp15 did not produceinfectious BVs. However, to test whether noninfectious BVsare still produced, their respective viral copy numbers in cellculture supernatants at 0 and 48 h p.t. were determined byqPCR and compared with those for the p6.9 rescue bacmidand a previously constructed (49) gp64-null AcMNPV bacmid(�gp64) that is unable to produce BVs (Fig. 5A) (30). At 48 hp.t., a significant increase in the number of viral DNA copiesover that at 0 h p.t. was found only for the p6.9 rescue bacmid(P, �0.05 by a one-tailed Student t test). This demonstratesthat neither infectious nor noninfectious BVs were producedwith the p6.9-null AcMNPV bacmids carrying Betabaculovirusp6.9 or WSSV vp15. As a control, the numbers of viral DNAcopies in the transfected cells were also analyzed by qPCR(Fig. 5B). For all bacmids, transfected cells contained signifi-cantly increased copy numbers (P, �0.05 by a one-tailed Stu-dent t test), indicating that the presence of P6.9 per se is notessential for viral replication.
The AcMNPV P6.9 C-terminal domain is important forvirion production. Baculovirus P6.9 proteins, like protamines,are a rather heterogeneous protein class. Homology searchesand comparisons with FASTA and BLAST programs revealedsignificant amino acid identities only among P6.9 proteins fromclosely related baculoviruses. Consequently, our P6.9 align-
ments (see the supplemental material) are based more onaligning conserved arginine and serine residues than on globalsimilarities. However, the C-terminal domains of P6.9 proteinshave a greater degree of identity within the baculovirus genera.For example Alpha-, Gamma-, and Deltabaculovirus P6.9 pro-teins terminate at a tyrosine (Fig. 1C), while those of Betabacu-lovirus contain an extra valine or isoleucine (Fig. 1C; see alsothe supplemental material). To explore whether these addi-tional C-terminal hydrophobic amino acids in BetabaculovirusP6.9 proteins interfere with BV production by AcMNPV, twoAcMNPV mutants, one expressing native AcMNPV P6.9but with an additional isoleucine at the extreme C terminus(Y55_X56insI) and the other expressing ChocGV P6.9 with adeletion of the C-terminal valine (V57del), were constructed(Fig. 1B) and transfected into Sf21 cells (Fig. 2J and K, upperpanels). BVs were produced from Sf21 cells transfected withAcMNPV expressing P6.9 bearing the additional C-terminalisoleucine (Fig. 2J, lower panel) but not from those bearing theC-terminally modified ChocGV P6.9 (Fig. 2K, lower panel),demonstrating that, at least in these two cases, the presence orabsence of a C-terminal hydrophobic amino acid had littleeffect on P6.9 function.
In the Alpha- and Gammabaculovirus P6.9 C termini, theYxxRxY sequence is highly conserved (Fig. 1C; see also thesupplemental material). To investigate whether these conservedamino acids are important for BV production, AcMNPV p6.9sequences with single alanine substitutions at each conservedresidue (Y50A, R53A, and Y55A) were made (Fig. 1B) andinserted into the p6.9-null AcMNPV bacmid. None of the threesubstitutions disrupted the formation of infectious virions (Fig.2L to N). To take this one step further, two AcMNPV p6.9constructs were made, one with a deletion of the 6 C-terminalamino acids (Y50_Y55del) and one with a deletion of 11 C-terminal amino acids (Y45_Y55del) (Fig. 1B and C). Despitethe fact that the six deleted C-terminal amino acids included
FIG. 4. One-step growth curves. Sf21 cells were infected with the AcMNPV bacmid (parental) or with p6.9-null AcMNPV containing AcMNPV(Ac), SeMNPV (Se), HearNPV (Ha), or NeleNPV (Nl) p6.9 (A) or with p6.9-null AcMNPV containing either AcMNPV (Ac) p6.9 or CypoGV p6.9with the H41_Y49delinsTGRRSYRTRYY modification (B) at an MOI of 10 TCID50/cell. Supernatants were harvested at the indicated times p.i.,and viral titers were determined by endpoint dilution assays. Each data point represents the average for three independent infections. Error barsrepresent the standard errors of the means.
VOL. 84, 2010 ROLE OF THE BACULOVIRUS P6.9 C TERMINUS 8825
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

the three conserved amino acids Y50, R53, and Y55, the mod-ified P6.9 construct Y50_Y55del was able to rescue p6.9-nullAcMNPV (Fig. 2O). In contrast, deletion of an extra five, morepoorly conserved residues, compromising a TGRRS sequence,resulted in a complete block of BV production (Fig. 2P).
These results imply that this 11-residue stretch of aminoacids at the extreme C terminus of AcMNPV P6.9 is importantfor the formation of infectious AcMNPV virions. To furtherinvestigate the role of these conserved C-terminal residues,p6.9-null AcMNPV bacmids fitted with CypoGV p6.9 genesencoding P6.9 variants in which residues 41 to 47 were re-placed by either the 6 (H41_Y49delinsYRTRYY) or the 11(H41_Y49delinsTGRRSYRTRYY) C-terminal residues ofAcMNPV P6.9 were transfected into Sf21 cells (Fig. 2Q and R,upper panels). The H41_Y49delinsYRTRYY modification didnot rescue the loss of AcMNPV p6.9 (Fig. 2Q, lower panel),but interestingly, infectious AcMNPV virions were formedwith the H41_Y49delinsTGRRSYRTRYY C-terminal mod-ification (Fig. 2R, lower panel). However, a one-step growthcurve for this virus revealed that it produced 10 to 60 timesfewer infectious viruses over time (24 to 72 h) (P, �0.05 bya two-tailed Student t test) (Fig. 4B). These results indicatethat the C-terminal region of AcMPNV P6.9 contains resi-dues crucial for AcMNPV virion production that are presentin Alpha- and Gammabaculovirus but absent in Betabaculo-virus P6.9 proteins. Furthermore, the decreased virion pro-duction observed with the CypoGV P6.9 protein with theH41_Y49delinsTGRRSYRTRYY C-terminal modificationsuggests that Betabaculovirus P6.9 lacks additional impor-tant amino acid residues or domains associated with virionformation.
DISCUSSION
Protamines are typically short (50- to 110-amino-acid [aa])proteins containing a high density of positively charged aminoacids, most notably arginines (24). Although protamines lacksignificant secondary structure in solution, upon interactionwith DNA, they readily wrap themselves into the major grooveof the DNA helix (4, 5, 9, 19). Playing a functionally similarrole, baculovirus P6.9 is a small, heterogeneous arginine- andserine-rich protein, ranging from 49 to 109 aa (Fig. 1C; see alsothe supplemental material), that, in its dephosphorylated form,is believed to bind to and condense viral DNA during nucleo-capsid assembly and virion packaging (14, 22, 28, 54) in anapparently sequence independent manner (22). In this study,we investigated whether P6.9 might contain domains that in-teract with other components specific to the species or genusduring viral assembly.
We demonstrate, by deletion of p6.9 from the Alphabaculo-virus type species AcMNPV (21), that P6.9 is an essential genefor viral propagation (Fig. 2A and 4A). Electron microscopicobservations of p6.9-null AcMNPV-transfected cells showedthat viral nucleocapsids were not formed (Fig. 3A). Instead ofnucleocapsids, electron-lucent tubular structures were presentin the nuclei of p6.9-null AcMNPV-transfected cells. Thesetubular structures might be malformed nucleocapsids, in viewof the fact that similar structures have been found previouslywith a non-BV-producing orf103-null and 38k-null AcMNPV(44, 57). Furthermore, the genome copy number still increasesover time, indicating that P6.9 is not essential for viral repli-cation (Fig. 5B). The replication efficiency seems to be higherin the presence of AcMNPV P6.9 (Fig. 5B). This is probably
FIG. 5. Quantification of viral genome copies in supernatants (A) and Sf21 cells (B) at 0 and 48 h after transfection with p6.9-null AcMNPVbacmids containing either no p6.9 gene (), the p6.9 gene of AcMNPV (Ac), CrleGV (Cl), ChocGV (Co), or CypoGV (Cp), or the vp15 gene ofWSSV, or with a gp64-null AcMNPV bacmid (�GP64). Data are expressed as relative frequencies, calculated by dividing each data point by themean at 0 h. Error bars represent 1 standard deviation. Data were analyzed by a one-tailed Student t test. *, P � 0.05; **, P � 0.01; ***, P �0.001. Each data point represents the average for three experiments.
8826 WANG ET AL. J. VIROL.
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

due to the ability to have secondary infections within 48 h, asevidenced by the fact that the control gp64-null AcMNPVbacmids, which are not able to form BVs, have replicationefficiencies similar to that of the p6.9-null AcMNPV bacmid.
The defect in the production of infectious virions could berescued by reinsertion of p6.9 from baculoviruses belonging tothe genera Alpha- and Gammabaculovirus (Fig. 2B to D).Among P6.9 proteins from these two genera, only the insertionof the HearNPV p6.9 into p6.9-null AcMNPV gave a significantdecrease in the titer at 72 h p.i. (Fig. 4A). In contrast, althoughbetabaculoviruses are phylogenetically more closely related toalphabaculoviruses than are gammabaculoviruses, P6.9 pro-teins from betabaculoviruses were not capable of rescuingp6.9-null AcMNPV (Fig. 2E to G and 4A). This suggests that,in this particular case, Betabaculovirus P6.9 may have evolvedto interact specifically with a Betabaculovirus-specific nucleo-capsid protein during viral assembly.
The inability of Gammabaculovirus P6.9 to rescue p6.9-nullAcMNPV is presumably not due to dysfunctional translocationof the protein to the nucleus, since these proteins containmultiple nuclear localization signals. Most likely, P6.9 needs tointeract with one or more nucleocapsid proteins during viralassembly. The proteins VP39, VP91, VP1054, and Vlf-1 aregood candidates, since they are present in the nucleocapsids ofall baculoviruses (7, 46) and are important for viral assembly(26, 33, 38, 43). Baculovirus P6.9 proteins are a rather hetero-geneous group of proteins, but their C-terminal domains areconserved within the genera and therefore may contain do-mains designed to interact specifically with other viral pro-teins. Neither mutations in the conserved C-terminal do-main YxxRxY of AcMNPV P6.9 nor its complete deletionabolished the ability to rescue p6.9-null AcMNPV (Fig. 2J to O).However, deletion of 5 additional amino acids, which are lessconserved, eliminated BV production (Fig. 2P), whereas aCypoGV P6.9 protein whose C-terminal domain was replacedwith these 11 C-terminal amino acids of AcMNPV P6.9 rescuedthe p6.9-null AcMNPV phenotype (Fig. 2R). These results sup-port the view that the C terminus of P6.9 is important for viralassembly and, to a certain degree, acts in a sequence-specificmanner within the genera. The latter hypothesis could be testedby future protein-protein interaction studies using P6.9 as thebait. The amount of infectious virus produced with the hybridP6.9 was 60 times lower than that produced with the nativeAcMNPV P6.9 (Fig. 4B); this could imply that BetabaculovirusP6.9 lacks additional important crucial amino acids or domains orinteracts erratically with alpha- and gammabaculoviruses.
The majority of dsDNA viruses infecting Eukaryota use cel-lular histones or virally expressed protamine-like proteins tocondense their DNA (6, 13, 37, 38, 40), while baculoviruseshave evolved a different and more compact way to condensetheir genomes by expressing proteins homologous to prota-mines. P6.9 proteins, though, lack cysteine residues used tolink protamines together by multiple disulfide bridges in sper-matids of eutherian mammals (4). GenBank comparisons withFASTA and BLAST programs showed that P6.9 proteins are,however, more similar in amino acid composition to the pro-tamines of squids and marsupials, which also lack cysteineresidues (data not shown).
Baculoviruses are not the only dsDNA viruses using prota-mine-like proteins to condense their genomes. The white spot
syndrome virus (WSSV) uses a similar protein, named VP15(56), which is one of its major nucleocapsid proteins (45).VP15 is 31% identical to AcMNPV P6.9 and has been shownto bind both WSSV and AcMNPV DNA (56). Despite itshomology, VP15 was not able to rescue the p6.9-null AcMNPV(Fig. 2I), probably because it is similar to the betabaculovirusP6.9 proteins lacking the C-terminal domain needed forAcMNPV virion assembly. Construction and testing of a VP15hybrid with the C-terminal end of AcMNPV P6.9 would provethis point. Another striking difference that might explain theinability of VP15 to assemble AcMNPV BVs is the fact thatVP15 is rather lysine rich (21%), while the majority of P6.9proteins contain few or no lysines (maximum, 7%). VP15 isdetected only in a dephosphorylated form in WSSV-infectedshrimp tissue, as well as when overexpressed in insect cells (45,56). This could suggest that VP15 uses a different mode ofaction to bind to and be released from DNA.
The genomes of Heliothis zea virus 1 and Gryllus bimaculatusnudivirus also encode a protamine-like protein (10, 50). Theseviruses were previously called “nonoccluded baculoviruses.” Atthe moment they are unclassified (12), but since they share afew common genes with baculoviruses, including the P6.9 gene,it has been proposed that they be placed in the genus Nudiviruswithin the Baculoviridae (51). Chilo iridescent virus (CIV), alsoknown as invertebrate iridescent virus 6, contains two genesencoding protamine homologs (20). No experimental data asto whether CIV uses these proteins to condense its genome areavailable, but so far it looks as though the use of protamine-like proteins in viral DNA packaging is evolutionarily con-served and also includes those dsDNA viruses infecting inver-tebrates of the phylum Arthropoda.
ACKNOWLEDGMENTS
We thank D. Zuidema (Wageningen University, Netherlands) foradvice during the research, B. Arif (Great Lakes Forestry Centre, SaultSte. Marie, Canada) for kindly providing the ChocGV HindIII-E andNeleNPV HindIII-D constructs, J. A. Jehle (State Education and Re-search Center for Agriculture, Viticulture and Horticulture [SLFA],Neustadt an der Weinstraße, Germany) for kindly providing genomicCypoGV and CreleGV DNA, G. P. Pijlman for constructing pFB1-p6.9, M. F. Boyong for assistance as an undergraduate student, andC. T. Dolphin (King’s College, London, United Kingdom) for editingthe manuscript.
This research was supported by grants from the Royal NetherlandsAcademy of Arts and Sciences (KNAW) and the Chinese Academy ofScience (CAS) (Program Strategic Scientific Alliances projects 04-PSA-BD-02 and 2004CB720404) and by Intervet International BV,Boxmeer, Netherlands (WSSV).
REFERENCES
1. Adams, J. R., and J. T. McClintock. 1991. Nuclear polyhedrosis viruses ofinsects, p. 87–204. In J. R. Adams and J. R. Bonami (ed.), Atlas of inverte-brate viruses. CRC Press, Boca Raton, FL.
2. Altschul, S. F., T. L. Madden, A. A. Schaffer, J. Zhang, Z. Zhang, W. Miller,and D. J. Lipman. 1997. Gapped BLAST and PSI-BLAST: a new generationof protein database search programs. Nucleic Acids Res. 25:3389–3402.
3. Ayres, M. D., S. C. Howard, J. Kuzio, M. Lopez-Ferber, and R. D. Possee.1994. The complete DNA sequence of Autographa californica nuclear poly-hedrosis virus. Virology 202:586–605.
4. Balhorn, R. 2007. The protamine family of sperm nuclear proteins. GenomeBiol. 8:227.
5. Balhorn, R., M. Cosman, K. Thornton, V. V. Krishnan, M. Corzett, G.Bench, C. Kramer, J. Lee IV, N. V. Hud, M. Allen, M. Prieto, W. Meyer-Ilse,J. T. Brown, J. Kirz, X. Zhang, E. M. Bradbury, G. Maki, R. E. Braun, andW. G. Breed. 1999. Protamine mediated condensation of DNA in mamma-lian sperm, p. 55–70. In C. Gagnon (ed.), The male gamete: from basicscience to clinical applications. Cache River Press, Vienna, IL.
VOL. 84, 2010 ROLE OF THE BACULOVIRUS P6.9 C TERMINUS 8827
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

6. Borca, M. V., P. M. Irusta, G. F. Kutish, C. Carillo, C. L. Afonso, A. T.Burrage, J. G. Neilan, and D. L. Rock. 1996. A structural DNA bindingprotein of African swine fever virus with similarity to bacterial histone-likeproteins. Arch. Virol. 141:301–313.
7. Braunagel, S. C., W. K. Russell, G. Rosas-Acosta, D. H. Russell, and M. D.Summers. 2003. Determination of the protein composition of the occlusion-derived virus of Autographa californica nucleopolyhedrovirus. Proc. Natl.Acad. Sci. U. S. A. 100:9797–9802.
8. Bulach, D. M., C. A. Kumar, A. Zaia, B. Liang, and D. E. Tribe. 1999. GroupII nucleopolyhedrovirus subgroups revealed by phylogenetic analysis of poly-hedrin and DNA polymerase gene sequences. J. Invertebr. Pathol. 73:59–73.
9. Campos, E. I., and D. Reinberg. 2009. Histones: annotating chromatin.Annu. Rev. Genet. 43:559–599.
10. Cheng, C. H., S. M. Liu, T. Y. Chow, Y. Y. Hsiao, D. P. Wang, J. J. Huang,and H. H. Chen. 2002. Analysis of the complete genome sequence of theHz-1 virus suggests that it is related to members of the Baculoviridae. J. Vi-rol. 76:9024–9034.
11. Davis, S. J., and R. D. Vierstra. 1998. Soluble, highly fluorescent variants ofgreen fluorescent protein (GFP) for use in higher plants. Plant Mol. Biol.36:521–528.
12. Fauquet, C. M., M. A. Mayo, J. Maniloff, U. Desselberger, and L. A. Ball(ed.). 2005. Virus taxonomy. Eighth report of the International Committeeon Taxonomy of Viruses. Elsevier Academic Press, San Diego, CA. http://www.ncbi.nlm.nih.gov/ICTVdb/Ictv/index.htm.
13. Frost, E., and P. Bourgaux. 1978. Structures of polyoma virus: on the histonecomponent and virion core. J. Gen. Virol. 39:103–111.
14. Funk, C. J., and R. A. Consigli. 1993. Phosphate cycling on the basic proteinof Plodia interpunctella granulosis virus. Virology 193:396–402.
15. Hayakawa, T., G. F. Rohrmann, and Y. Hashimoto. 2000. Patterns of ge-nome organization and content in lepidopteran baculoviruses. Virology 278:1–12.
16. Herniou, E. A., T. Luque, X. Chen, J. M. Vlak, D. Winstanley, J. S. Cory, andD. R. O’Reilly. 2001. Use of whole genome sequence data to infer baculo-virus phylogeny. J. Virol. 75:8117–8126.
17. Herniou, E. A., J. A. Olszewski, J. S. Cory, and D. R. O’Reilly. 2003. Thegenome sequence and evolution of baculoviruses. Annu. Rev. Entomol.48:211–234.
18. Hill-Perkins, M. S., and R. D. Possee. 1990. A baculovirus expression vectorderived from the basic protein promoter of Autographa californica nuclearpolyhedrosis virus. J. Gen. Virol. 71:971–976.
19. Hud, N. V., F. P. Milanovich, and R. Balhorn. 1994. Evidence of novelsecondary structure in DNA-bound protamine is revealed by Raman spec-troscopy. Biochemistry 33:7528–7535.
20. Jakob, N. J., K. Muller, U. Bahr, and G. Darai. 2001. Analysis of the firstcomplete DNA sequence of an invertebrate iridovirus: coding strategy of thegenome of Chilo iridescent virus. Virology 286:182–196.
21. Jehle, J. A., G. W. Blissard, B. C. Bonning, J. S. Cory, E. A. Herniou, G. F.Rohrmann, D. A. Theilmann, S. M. Thiem, and J. M. Vlak. 2006. On theclassification and nomenclature of baculoviruses: a proposal for revision.Arch. Virol. 151:1257–1266.
22. Kelly, D. C., D. A. Brown, M. D. Ayres, C. J. Allen, and I. O. Walker. 1983.Properties of the major nucleocapsid protein of Heliothis zea singly envel-oped nuclear polyhedrosis virus. J. Gen. Virol. 64:399–408.
23. Kelly, D. C., and T. Lescott. 1984. Baculovirus replication: phosphorylationof polypeptides synthesized in Trichoplusia ni nuclear polyhedrosis virus-infected cells. J. Gen. Virol. 65:1183–1191.
24. Lewis, J. D., Y. Song, M. E. de Jong, S. M. Bagha, and J. Ausio. 2003. A walkthough vertebrate and invertebrate protamines. Chromosoma 111:473–482.
25. Lo, C. F., C. H. Ho, S. E. Peng, C. H. Chen, H. C. Shu, Y. L. Chiu, C. F.Chang, K. F. Liu, M. S. Su, C. H. Wang, and G. H. Kou. 1996. White spotsyndrome baculovirus (WSBV) detected in cultured and captured shrimp,crabs and other arthropods. Dis. Aquat. Org. 27:215–225.
26. Lu, S., G. Ge, and Y. Qi. 2004. Ha-VP39 binding to actin and the influenceof F-actin on assembly of progeny virions. Arch. Virol. 149:2187–2198.
27. Luckow, V. A., S. C. Lee, G. F. Barry, and P. O. Olins. 1993. Efficientgeneration of infectious recombinant baculoviruses by site-specific transpo-son-mediated insertion of foreign genes into a baculovirus genome propa-gated in Escherichia coli. J. Virol. 67:4566–4579.
28. Maeda, S., S. G. Kamita, and H. Kataoka. 1991. The basic DNA-bindingprotein of Bombyx mori nuclear polyhedrosis virus: the existence of anadditional arginine repeat. Virology 180:807–810.
29. Mirza, M. A., and J. Weber. 1982. Structure of adenovirus chromatin. Bio-chim. Biophys. Acta 696:76–86.
30. Monsma, S. A., A. G. Oomens, and G. W. Blissard. 1996. The GP64 envelopefusion protein is an essential baculovirus protein required for cell-to-celltransmission of infection. J. Virol. 70:4607–4616.
31. Muyrers, J. P., Y. Zhang, G. Testa, and A. F. Stewart. 1999. Rapid modifi-cation of bacterial artificial chromosomes by ET-recombination. NucleicAcids Res. 27:1555–1557.
32. Nicholas, K. B., H. B. Nicholas, Jr., and D. W. Deerfield II. 1997. GeneDoc:analysis and visualization of genetic variation. EMBnet News 4:1–4.
33. Olszewski, J., and L. K. Miller. 1997. Identification and characterization ofa baculovirus structural protein, VP1054, required for nucleocapsid forma-tion. J. Virol. 71:5040–5050.
34. O’Reilly, D. R., L. K. Miller, and V. A. Luckow. 1992. Baculovirus expressionvectors: a laboratory manual. W. H. Freeman and Co., New York, NY.
35. Pearson, W. R. 1990. Rapid and sensitive sequence comparison with FASTPand FASTA. Methods Enzymol. 183:63–98.
36. Pfister, H., and H. zur Hausen. 1978. Characterization of proteins of humanpapilloma viruses (HPV) and antibody response to HPV 1. Med. Microbiol.Immunol. 166:13–19.
37. Pijlman, G. P., J. C. Dortmans, A. M. Vermeesch, K. Yang, D. E. Martens,R. W. Goldbach, and J. M. Vlak. 2002. Pivotal role of the non-hr origin ofDNA replication in the genesis of defective interfering baculoviruses. J. Vi-rol. 76:5605–5611.
38. Russell, R. L., and G. F. Rohrmann. 1997. Characterization of P91, a proteinassociated with virions of an Orgyia pseudotsugata baculovirus. Virology233:210–223.
38a.Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, NY.
39. Shizuya, H., B. Birren, U. J. Kim, V. Mancino, T. Slepak, Y. Tachiiri, and M.Simon. 1992. Cloning and stable maintenance of 300-kilobase-pair fragmentsof human DNA in Escherichia coli using an F-factor-based vector. Proc. Natl.Acad. Sci. U. S. A. 89:8794–8797.
40. Tan, K. B. 1977. Histones: metabolism in simian virus 40-infected cells andincorporation into virions. Proc. Natl. Acad. Sci. U. S. A. 74:2805–2809.
41. Tweeten, K. A., L. A. Bulla, and R. A. Consigli. 1980. Characterization of anextremely basic protein derived from granulosis virus nucleocapsids. J. Virol.33:866–876.
42. Tweeten, K. A., L. A. Bulla, Jr., and R. A. Consigli. 1978. Characterization ofan alkaline protease associated with a granulosis virus of Plodia interpunc-tella. J. Virol. 26:703–711.
43. Vanarsdall, A. L., K. Okano, and G. F. Rohrmann. 2006. Characterization ofthe role of very late expression factor 1 in baculovirus capsid structure andDNA processing. J. Virol. 80:1724–1733.
44. Vanarsdall, A. L., M. N. Pearson, and G. F. Rohrmann. 2007. Characteriza-tion of baculovirus constructs lacking either the Ac 101, Ac 142, or the Ac144 open reading frame. Virology 367:187–195.
45. van Hulten, M. C., M. Reijns, A. M. Vermeesch, F. Zandbergen, and J. M.Vlak. 2002. Identification of VP19 and VP15 of white spot syndrome virus(WSSV) and glycosylation status of the WSSV major structural proteins.J. Gen. Virol. 83:257–265.
46. van Oers, M. M., and J. M. Vlak. 2007. Baculovirus genomics. Curr. DrugTargets 8:1051–1068.
47. Vaughn, J. L., R. H. Goodwin, G. J. Tompkins, and P. McCawley. 1977. Theestablishment of two cell lines from the insect Spodoptera frugiperda (Lepi-doptera; Noctuidae). In Vitro 13:213–217.
48. Wang, K., C. Boysen, H. Shizuya, M. I. Simon, and L. Hood. 1997. Completenucleotide sequence of two generations of a bacterial artificial chromosomecloning vector. Biotechniques 23:992–994.
49. Wang, M., Y. Tan, F. Yin, F. Deng, J. M. Vlak, Z. Hu, and H. Wang. 2008.The F-like protein Ac23 enhances the infectivity of the budded virus ofgp64-null Autographa californica multinucleocapsid nucleopolyhedroviruspseudotyped with baculovirus envelope fusion protein F. J. Virol. 82:9800–9804.
50. Wang, Y., R. G. Kleespies, A. M. Huger, and J. A. Jehle. 2007. The genomeof Gryllus bimaculatus nudivirus indicates an ancient diversification of bac-ulovirus-related nonoccluded nudiviruses of insects. J. Virol. 81:5395–5406.
51. Wang, Y., M. M. van Oers, A. M. Crawford, J. M. Vlak, and J. A. Jehle. 2007.Genomic analysis of Oryctes rhinoceros virus reveals genetic relatedness toHeliothis zea virus 1. Arch. Virol. 152:519–531.
52. Westenberg, M., F. Veenman, E. C. Roode, R. W. Goldbach, J. M. Vlak, andD. Zuidema. 2004. Functional analysis of the putative fusion domain of thebaculovirus envelope fusion protein F. J. Virol. 78:6946–6954.
53. Wilson, M. E., and R. A. Consigli. 1985. Characterization of a protein kinaseactivity associated with purified capsids of the granulosis virus infectingPlodia interpunctella. Virology 143:516–525.
54. Wilson, M. E., and R. A. Consigli. 1985. Functions of a protein kinase activityassociated with purified capsids of the granulosis virus infecting Plodia in-terpunctella. Virology 143:526–535.
55. Wilson, M. E., and L. K. Miller. 1986. Changes in the nucleoprotein com-plexes of a baculovirus DNA during infection. Virology 151:315–328.
56. Witteveldt, J., A. M. Vermeesch, M. Langenhof, A. de Lang, J. M. Vlak, andM. C. van Hulten. 2005. Nucleocapsid protein VP15 is the basic DNAbinding protein of white spot syndrome virus of shrimp. Arch. Virol. 150:1121–1133.
57. Wu, W., T. Lin, L. Pan, M. Yu, Z. Li, Y. Pang, and K. Yang. 2006. Autographacalifornica multiple nucleopolyhedrovirus nucleocapsid assembly is inter-rupted upon deletion of the 38K gene. J. Virol. 80:11475–11485.
8828 WANG ET AL. J. VIROL.
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from