species diversity concepts describing communities species
TRANSCRIPT

1
Species Diversity Concepts
Species RichnessSpecies-Area CurvesRarefactionDiversity Indices
- Simpson's Index- Shannon-Weiner Index- Brillouin Index
Species Abundance Models
Describing Communities
There are two important descriptors of a community:
1) its physiognomy(physical structure), as described in the previous lecture, and
2) the number of speciespresent and their relative abundances (species richness and diversity).
Species Richness
The simplest way to describe a community is to list the species in it.
Species richness (S) is the number of species on that list, and is most often used as the first pass estimate of diversity for acommunity.
How would one generate such a list? A simple and widely used method is to define the boundaries of the community and then walk through it seasonally, noting all the species you encounter. This is what we call a flora.
Species Richness
While simply finding and listing the species is useful, this method has many limitations.
If we wish to compare two or more communities, we need comparable samples, otherwise we might just find a difference because one was sampled more intensively than the other.
This begs the question, how much sampling should we do in order to be confident that we have found most of the species in each community?
Species-Area Curve
One way to make this interpretation is through the use of a species-area curve.
A graph of the total number of species found as the number of quadrats increases explains the relationship. We know that we have sampled sufficiently when the curve begins to "plateau".
0
5
10
15
20
25
30
1 3 5 7 9 11 13 15
No. of 1m2 quadrats
No
. of
Sp
eci
es
(S
)
Conclusion: 10-12 quadrats would be sufficient to describe the vegetation in this community.
Species-Area Curve
After many years of study, we now realize that the classical perception of the shape of a "typical" species-area curve is an artifact of the type of communities in which the relationship was first described.
Moreover, it is now apparent, that habitat grain, patch size, and equitability will have a substantial influence on the shape of the curve and must be evaluated in that context.
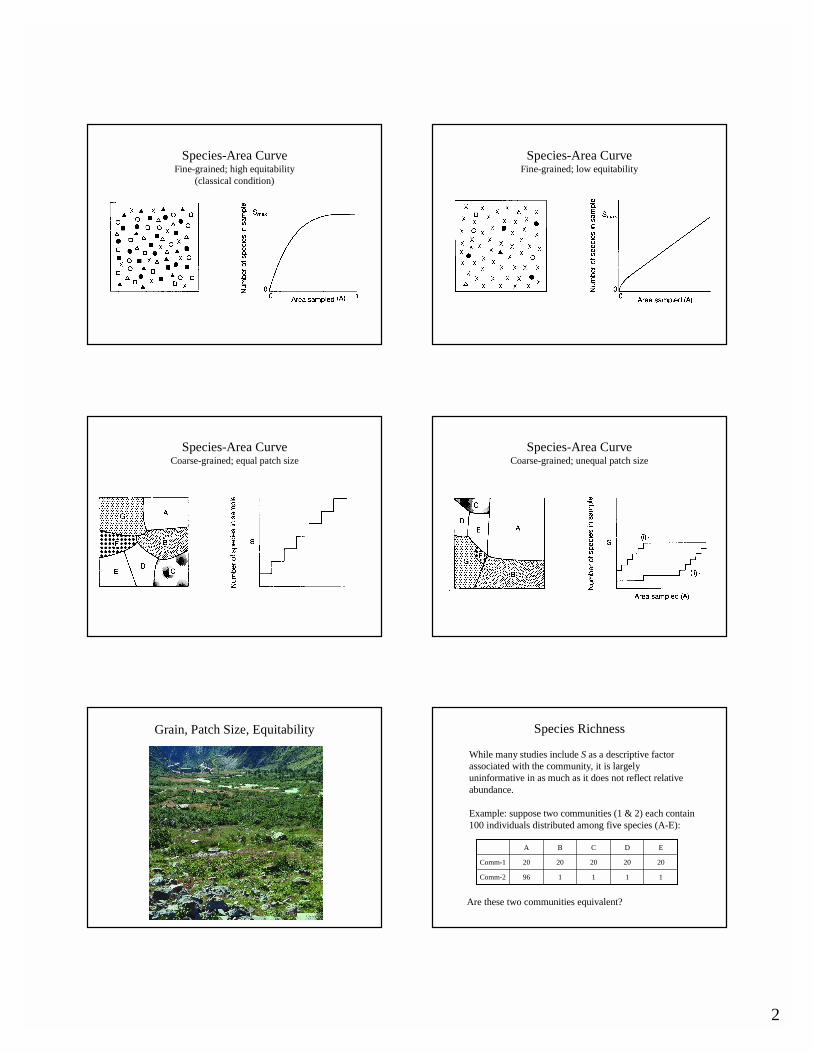
2
Species-Area CurveFine-grained; high equitability
(classical condition)
Species-Area CurveFine-grained; low equitability
Species-Area CurveCoarse-grained; equal patch size
Species-Area CurveCoarse-grained; unequal patch size
Grain, Patch Size, Equitability Species Richness
While many studies include S as a descriptive factor associated with the community, it is largely uninformative in as much as it does not reflect relative abundance.
Example: suppose two communities (1 & 2) each contain 100 individuals distributed among five species (A-E):
EDCBA
111196Comm-2
2020202020Comm-1
Are these two communities equivalent?

3
Species Richness
There are two ways to overcome this problem:
1) incorporate both species richness and abundance information in to one diversity index.
2) rely on species richness but control for the effects of sample size by a procedure called rarefaction.
We will examine both alternatives as they are widely used in ecology.
Rarefaction
One of the fundamental tenants of diversity is that the number of species found in a given sample is strongly dependent upon the size of that sample.
This makes good intuitive sense in that the more quadrats one samples in a plant community, the more likely you are to pick up more and more rare species.
One method of avoiding incompatibility of measurements resulting from samples of different sizes is called rarefaction.
Rarefaction
Si
i=1
N - N NE(S) = 1 - /
n n
∑
Where E(S) is the expected number of species in the rarefied sample, n is the standardized sample, N is the total number of in the sample to be rarefied, and Ni is the number of individuals in the ith species in the sample to be rarefied, summed over all species counted.
Rarefaction
The term N
n
is a "combination" that is calculated as:
N N! =
n n! N n !
where N! is a "factorial", e.g., 5! = 5×4×3×2×1 = 120
This combination is important, because it allows us to calculate all the possible numbers of unique species combinations...
Rarefaction
For example, if we have four species, A, B, C, D, then we have six species pairs: AB, AC, AD, BC, BD, CD.
Using the combinatorial equation:
4 4! 24 = = 6
2 2! 4 2 ! 4
=
Thus N
n
is the number of unique combinations of N
taken n at a time; i.e., the number of different ways of picking species pairs from four different species.
Rarefaction
Let's look at a fully worked real-world example (taken from Magurran 1988).
Imagine two moth traps that have been set out in a forest to monitor moth diversity. Trap-B was inadvertently left out for only about half the time as Trap-A.
We know this will be a problem because Trap-A "sampled" the environment more (longer period of time) and is likely to pick up more species. To compare S between traps would be misleading and inappropriate.

4
Moth light traps are typically suspended above the vegetation and contain a battery powered light. The trap is set to operate only during the evening hours. Moths are drawn to the light and become entrapped in the canister below.
Rarefaction
As expected, Trap-A has more species. The best way to correct for the difference in sampling time is to ask,
How many species would we expect to find in Trap-A if it too contained 13 individuals?
1323N
69S
0112
3111
5010
019
208
117
016
025
044
103
032
191
Trap-BTrap-ASpecies
No. of Individuals
Rarefaction
First, take the number of individuals of each species from Trap-A and insert them into the formula.
For species in Trap-A: N = 23, n = 13, Ni = 9, N-Ni = 14
( )
( )
[ ]{ }
i
N 23! =
n 13! 23 - 13 !
N-N 14! =
n 13! 14 - 13 !
therefore:
14! 23!1 - / 1 14 /1144066 1 0.00 1.00
13! 1! 13! 10!
= − = − = × ×
Rarefaction
Continue this same set of calculations for each species (to determine the expected number) and then sum the values (as per the Σ in the equation).
Zero values need not be included as they have no influence on the estimate.
6.58E(S)
0.571
0.571
0.571
0.571
0.571
0.822
0.984
0.933
1.009
ExpectedNi
Conclusion: If Trap-A contained 13 individuals, we would expect it to contain 6.58 species--about the same as Trap-B.
Diversity Indices
As already alluded to, the diversity of a community needs (in most instances) to account for both species richness and the evenness with which individuals are distributed among species.
One way to do this is through the use of a proportional abundance index. There are two major forms of these indices: dominance indices and information indices.
While more than 60 indices have been described, we will look at the three most widely used in the ecological literature: Simpson's, Shannon-Weiner, and Brillouin.
Simpson's Index
Simpson's Index is considered a dominance index because it weights towards the abundance of the most common species.
Simpson's Index gives the probability of any two individuals drawn at random from an infinitely large community belonging to different species.
For example, the probability of two trees, picked at random from a tropical rainforest being of the same species would be relatively low, whereas in boreal forest in Canada it would be relatively high.

5
Simpson's Index
( )( )( )( )
i
S1
n 1D
N N 1
Si
i
n
=
−=
−∑
The bias corrected form of Simpson's Index is:
where ni is the number of individuals in the ith species.
Since Ds and diversity are negatively related, Simpson's index is usually expressed as the reciprocal (1-D) so that as the index goes up, so does diversity.
Simpson's Index
A worked example for 201 trees of 5 species assessed in several quadrats:
201Total
1E
20D
30C
50B
100A
No. Individuals
Tree spp.
S
100 99 50 49 1 0D ... 0.338
201 200 201 200 201 200
Then 1/D = 1/0.388 = 2.96
× × × = + + = × × ×
Shannon-Weiner Index
The Shannon-Weiner Index belongs to a subset of indices that maintain that diversity can be measured much like the information contained in a code or message (hence the name information index).
The rationale is that if we know a letter in a message, we can know the uncertainty of the next letter in a coded message (i.e., the next species to be found in a community).
The uncertainty is measured as H', the Shannon Index. A message coded bbbbbb has low uncertainty (H' = 0).
Shannon-Weiner Index
The Shannon Index assumes that all species are represented in a sample and that the sample was obtained randomly:
S
i ii = 1
H' = - p ln p∑where pi is the proportion of individuals found in the ith species and ln is the natural logarithm.
Shannon-Weiner Index
A worked example from a community containing 100 trees distributed among 5 species:
-1.2011.001005Total
-0.0460.011E
-0.2170.099D
-0.2300.110C
-0.3610.330B
-0.3470.550A
pi ln pipiAbundSpecies
H' = 1.201
Shannon-Weiner Index
The most important source of error in this index is failing to include all species from the community in the sample.
This makes a species-area curve assessment very important at the beginning of a study.
Values of the Shannon diversity index for real communities typically fall between 1.5 and 3.5.

6
Shannon-Weiner Index
The Shannon index is affected by both the number of species and their equitability, or evenness.
A greater number of species and a more even distribution BOTH increase diversity as measured by H'.
The maximum diversity (Hmax) of a sample is found when all species are equally abundant. Hmax = ln S, where S is the total number of species.
Evenness
We can compare the actual diversity value to the maximum possible diversity by using a measure called evenness.
The evenness of the sample is obtained from the formula:
Evenness = H'/Hmax = H'/lnS
By definition, E is constrained between 0 and 1.0. As with H', evenness assumes that all species are represented within the sample.
Brillouin Index
When the randomness of a sample cannot be guaranteed, the Brillouin Index HB is preferable to the H':
iB
ln N! - ln n !H
N= ∑
where N is the total number of individuals and ni is the number of individuals in the ith species.
A worked example follows...
Brillouin Index
Σ = 23.95N = 25S = 5
4.7955
4.7954
4.7953
4.7952
4.7951
ln ni !No. IndividualsSpecies
iB
ln N! - ln n ! ln 25! - 23.95H 1.362
N 25= = =∑
Evenness
Evenness for the Brillouin Index is estimated as:
BHE =
Hwhere HBmax represents the maximum possible Brillouin diversity, that is, a completely equitable distribution of individuals between species.
In our example, we had complete equitability, therefore, HBmax = HB = 1.0.
Diversity Indices
As you have probably figured out, the choice of a particular index is chosen with respect to the goals of the study (emphasis on abundant vs rare species) and to what extent sampling can be assured to be random.
There are other factors that come in to play, but these are the 3 most widely used measures of diversity that incorporate both richness and evenness into the determination.
Note: There is generally NO relationship between one index and another.

7
Species Abundance Models
One of the earliest observations made by plant ecologists was that species are not equally common in a given community. Some were very abundant, other were uncommon.
A graphical way was sought to describe this pattern, and so arose species abundance models.
These models are strongly advocated among some ecologists because they emphasize abundance while utilizing species richness information and therefore provide the most complete mathematical description of the data.
Species Abundance Models
A species abundance model is generated by graphing the abundance of each species against its rank order abundance from 1 = highest to N = lowest.
One of four distributions usually arise:
Log normal distributionGeometric seriesLogarithmic seriesMcArthur's broken stick model
Species Abundance Models Species Abundance Models(Changes through succession - Bazzaz 1975)