sistema hipotalamo adenohipofisiario - … · sistema hipotalamo adenohipofisiario la hipófisis se...
TRANSCRIPT

SISTEMA HIPOTALAMO ADENOHIPOFISIARIO La hipófisis se origina por la diferenciación de dos brotes embrionarios. Uno de ellos es una evaginación del techo de la faringe, la bolsa de Rathke la cual crece dorsalmente hasta alcanzar la cavidad craneana y se ubica en lo que será la silla turca del esfenoides. El otro brote es una evaginación del piso del diencéfalo que crece ventralmente hasta ponerse en contacto con la bolsa de Rathke. La bolsa de Rathke da origen a las tres regiones de la hipófisis epitelial glandular o adenohipófisis. Ellas son la pars intermedia, la pars distalis y la pars tuberalis (Fig. 1). El brote diencefálico da origen a la hipófisis nerviosa o neurohipófisis, la cual se diferencia en tres regiones estructural y funcionalmente distintas: la eminencia media, el tallo y el lóbulo neural (Fig. 1). Adenohipófisis. Pars intermedia. Esta porción de la adenohipófisis está en íntimo contacto con el lóbulo neural. Está constituida por un tipo celular virtualmente único, organizado como una masa celular sólida pobre en vasos sanguíneos. Sería la glándula endocrina menos vascularizada. Sus células sintetizan pro-opiomelanocortina (POMC). Esta molécula precursora es procesada en estas células para dar origen a α-MSH y β-endorfina. La α-MSH (hormona melanóforo estimulante) se libera a la sangre y es considerada la hormona propia de la pars intermedia. No hay evidencias claras respecto de la liberación a la sangre de la β-endorfina de la pars intermedia. Las células secretorias de la pars intermedia están bajo control inhibitorio del hipotálamo. Este control se ejerce por medio de fibras nerviosas que establecen contactos directos (sinaptoídeos) con las células de la pars intermedia. Las fibras que ejercen este control inhibitorio se originan en neuronas del núcleo arcuato del hipotálamo, las cuales proyectan sus axones a través del lóbulo neural para alcanzar la pars intermedia. El neurotransmisor de estas neuronas es la dopamina. Esta amina biogénica es la que finalmente inhibe la secreción de α-MSH. En ciertas condiciones, tales como la oscuridad ambiental, las neuronas dopaminérgicas son inhibidas, lo cual resulta en una desinhibición de la pars intermedia y la consecuente liberación de α-MSH a la sangre. Pars tuberalis. Está localizada en íntima relación con la eminencia media y los vasos del plexo primario

del sistema porta hipofisiario (ver más adelante) (Figs. 1, 2, 5). En ella se pueden encontrar algunos de los tipos celulares propios de la pars distalis. Sin embargo, la mayor población de células son propias de la pars tuberalis, las cuales tienen características ultraestructurales de células glandulares. Sin embargo, hasta hoy se desconoce la naturaleza y función de la o las hormonas que secretaría la pars tuberalis. Pars distalis. Es la región más voluminosa de la hipófisis, está constituida por grupos y cordones de células epiteliales de tamaño variable. Entre esta red epitelial se encuentra un plexo vascular muy rico en capilares sanguíneos. Las células glandulares se pueden clasificar de acuerdo a su afinidad citoplasmática por colorantes básicos o ácidos. Las células acidófilas representan alrededor del 40% de las células secretorias de la pars distalis. Son las productoras de prolactina (PRL) y de hormona de crecimiento (GH). Ambas de naturaleza proteica. Estos tipos celulares son los que poseen los gránulos más grandes (PRL = 300-500 nm; GH = 150-400 nm). En condiciones fisiológicas basales las células GH son las más abundantes. Sin embargo, durante el embarazo y la lactancia las células PRL son las más numerosas. Las células basófilas incluyen a las productoras de tirotrofina (TSH), de hormona folículo estimulante (FSH) y hormona luteinizante (LH). Las tres son de naturaleza glicoproteica y por ello estas células son PAS-positivas. Las células TSH son escasas (1-2%) y tienen gránulos secretorios pequeños (40-150 nm). Las células FSH son abundantes y poseen una población de gránulos secretorios de tamaño heterogéneo (75-200 nm). Las células LH poseen una población más homogénea de gránulos. Las células que no muestran afinidad ni por colorantes básicos ni ácidos se denominan "cromófobas". A este grupo pertenecen las células intersticiales no secretorias (de reserva) y un sexto tipo secretor, el productor de la hormona adrenocorticotrofina (ACTH). Estas células poseen gránulos secretorios
pequeños (100-200 nm). A pesar de que el precursor de la ACTH (POMC) es una glicoproteína, los gránulos secretorios son cromófobos y no muestran afinidad por el PAS. Similares características tintoriales se encuentran en las células de la pars intermedia. Tanto las células ACTH de la pars distalis como las de la pars intermedia se caracterizan por sintetizar la misma pro-hormona (POMC). La diferencia está en el procesamiento de este precursor y por lo tanto en la expresión de los genes que codifican para las enzimas procesadoras. Mientras en la pars intermedia el resultado es la formación de α-MSH y β-endorfina; en las células ACTH el resultado es ACTH y β-endorfina, ambas liberadas a la sangre como hormonas. Sistema porta hipotálamo-hipofisiario. Un sistema vascular porta se caracteriza porque hay una capilarización extra respecto de la circulación sistémica clásica. En esta última la secuencia de vasos es: arteria arteriolas capilares venas aurícula derecha. En cambio, el orden en un sistema porta es: arteria arteriolas capilares venas capilares venas aurícula derecha. Por lo tanto, un sistema porta consiste de dos plexos capilares unidos por venas. El plexo capilar próximo a la arteria se denomina plexo primario y el que está próximo a la vena de drenaje se conoce como plexo secundario. El sentido de la circulación es desde el plexo primario al plexo secundario. Un sistema porta posibilita el transporte directo de sustancias desde un órgano "A", donde está el plexo primario, hasta un órgano "B", donde está el plexo secundario. Estas sustancias pueden llegar desde "A" hasta "B" en forma rápida y en altas concentraciones ya que no se diluyen en la sangre de todo el sistema circulatorio sino que sólo en el volumen de la sangre del sistema portal. En el sistema porta hipotálamo-hipofisiario el plexo primario se ubica en la eminencia media y parte del tallo neural (Fig. 2, Nº 4) y el plexo secundario en la pars distalis. Las venas portales (Fig. 2, Nº 3) corren a lo largo del tallo neural. El plexo primario se origina por la capilarización de la arteria hipofisiaria superior (Fig. 2, Nº 1). La mayor parte de la sangre que llega a la pars distalis es sangre portal. La arteria hipofisiaria inferior (Fig. 2, Nº 2), que provee la vascularización al lóbulo neural, envía algunos vasos que se comunican con los capilares de la pars distalis.

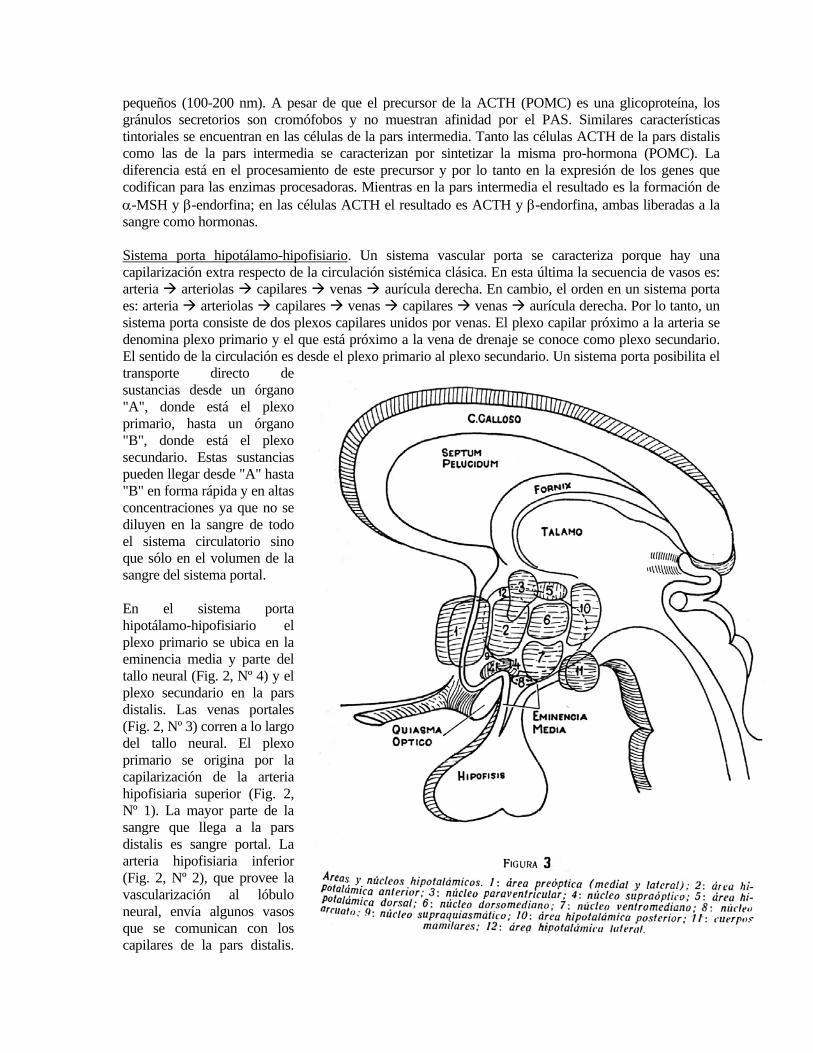
Este sería el único aporte de sangre arterial a la pars distalis. Los capilares de la pars distalis drenan finalmente en la vena hipofisiaria y ésta en la vena yugular. La sangre de la vena hipofisiaria es la que lleva al organismo todas las hormonas secretadas por la pars distalis. En el hipotálamo se encuentran varios grupos de neuronas formando núcleos o áreas (Fig. 3). Las áreas, formadas por neuronas laxamente dispuestas y sin límites definidos, son las siguientes: a) preóptica, b) hipotalámica anterior, c) hipotalámica dorsal, d) hipotalámica posterior, e) hipotalámica lateral. Los núcleos, de límites más definidos y formados por neuronas densamente agrupadas, son: 1) paraventricular (NPV); 2) periventricular (Nper); 3) supraóptico (NSO); 4) núcleo supraquiasmático (NSC); 5) arcuato (NAr); 6) ventromediano (NVM) y 7) dorsomediano (NDM). En gran parte de los núcleos hipotalámicos y en ciertas áreas hipotalámicas se encuentran neuronas secretoras de péptidos o aminas biogénicas, cuyos axones terminan en la eminencia media y liberan su contenido al plexo portal primario (Fig. 4, 5). Por lo tanto, pueden ser consideradas como neuronas neurosecretorias. Las neuronas neurosecretorias del sistema hipotálamo-neurohipofisiario y las del sistema hipotálamo-adenohipofisiario se diferencian en que las primeras liberan sus secreciones (OXY y AVP) a la circulación sistémica, las cuales llegan así a sus efectores que no son células glandulares, mientras que las segundas liberan su secreción a una circulación portal y sus efectores son células glandulares (pars distalis), las cuales a su vez responden liberando hormonas a la circulación sistémica (Fig. 8). Las neurohormonas peptídicas o aminas biogénicas liberadas por neuronas secretorias del sistema hipotálamo-adenohipofisiario controlan la liberación de cada una de las hormonas sintetizadas en la pars distalis (Fig. 6). La primera hormona hipotalámica reguladora de una función de la pars distalis que se descubrió es la (TRH) (thyrotropin-releasing hormone), un tripéptido que regula la liberación de tirotrofina (Fig. 7). El uso de anticuerpos contra TRH en técnicas de inmunocitoquímica permitió determinar que las neuronas que lo producen se localizan en el núcleo dorsomediano y sus axones terminan en el tercio medio de la eminencia media (Fig. 5). La segunda hormona hipotalámica en descubrirse fue la hormona liberadora de LH y FSH. Es un decapéptido y se denominó LH-RH

(luteinizing hormone-releasing hormone) o Gn-RH (gonadotropin releasing hormone). La LH-RH se sintetiza en neuronas ubicadas en el área preóptica y sus axones contactan los capilares portales de las zonas laterales de la eminencia media (Fig. 5). Según la cantidad y tiempo en el que se libere la Gn-RH a la sangre portal, se estimula la liberación de LH o de FSH a nivel de la pars distalis (Fig. 6). La liberación de la hormona de crecimiento (GH) de la pars distalis está bajo un doble control hormonal por parte del hipotálamo. La somatostatina (SRIF) que la inhibe y GH-RH (growth hormone-releasing hormone) que estimula la liberación de GH. La somatostatina tiene 14 aminoácidos (Fig. 7) y se sintetiza en neuronas de los núcleos periventriculares y arcuato, sus axones terminan en la región media de la eminencia media (Fig. 5). En condiciones basales el control inhibitorio que ejerce la somatostatina predomina sobre el control estimulatorio del GH-RH. Cuando la hipófisis se desconecta del control hipotalámico (accidentes, tumores o cirugía), las células GH se desinhiben e hipersecretan GH. La hormona hipotalámica que controla la liberación de ACTH es conocida como CRH (corticotropin-releasing hormone). La CRH posee 41 aminoácidos (Fig. 7) y se sintetiza en numerosos núcleos neuronales del SNC. Sin embargo, las únicas neuronas productoras de CRH cuyos axones proyectan a la eminencia, y liberan al sistema porta, están localizadas en la región parvocelular del NPV. En el NPV existen neuronas que son capaces de co-sintetizar dos péptidos derivados de genes diferentes: AVP y CRH. Estas neuronas productoras de AVP proyectan a la eminencia media y no al lóbulo neural. Si bien la CRH es la hormona específica para la liberación de ACTH, la acción de la CRH se potencia considerablemente si simultáneamente se libera AVP a la sangre portal. Por lo tanto, ambas hormonas pueden ser consideradas como liberadoras de ACTH.

En la Fig. 8 la neurona productora de CRH corresponde a la célula Nº 3. Ante un estímulo doloroso estresante las neuronas dibujadas en negro estimulan las neuronas CRH del NPV. Estas liberan CRH a la sangre portal (Nº 4 y 5). A nivel de la pars distalis (Nº 6) la CRH estimula a las células corticotropas para que liberen ACTH a la sangre sistémica (Nº 7). La ACTH al llegar a la glándula adrenal estimula las células de la corteza (Nº 8) para que liberen glucocorticoides a la sangre. Los glucocorticoides a su vez ejercerán su efecto a nivel del hígado (Nº 10) para que éste libere glucosa a la sangre. La hiperglicemia favorece la función muscular y con ello la respuesta ante el estímulo que causó el estrés. Pero los glucocorticoides en la sangre también pueden actuar sobre las neuronas productoras de CRH (Nº 11) ejerciendo un efecto inhibitorio y constituyendo el mecanismo de retroalimentación (feed-back) largo.
Los aumentos en la sangre periférica de los niveles de algunos de las hormonas de la pars distalis, ACTH en el ejemplo anterior, hacen que dicha hormona pueda actuar frenando la acción de la neurona hipotalámica correspondiente (en el ejemplo la neurona CRH). Esto constituye el mecanismo de retroalimentación (feed-back) corto (Fig. 8, Nº 13). La existencia de hormonas peptídicas hipotalámicas que estimulen (PRF) o inhiban (PIF), la secreción de prolactina (Fig. 6), no ha podido ser confirmada. El factor hipotalámico que sin duda ejerce un control sobre la secreción de PRL no es un péptido sino una catecolamina, la dopamina (DA). En el SNC existen

numerosos núcleos productores de DA. El que controla la PRL es el núcleo arcuato (Fig. 4). Las neuronas DA de este núcleo proyectan sus axones a la eminencia media donde la DA es liberada a la sangre portal (Fig. 5) para ejercer un control inhibitorio sobre la síntesis y liberación de PRL. Cualquier factor que interfiera con este control inhibitorio (disfunción o degeneración de neuronas DA, tumores que comprimen el tallo hipofisiario, accidentes, etc.) resultará en una desinhibición de las células prolactotropas y la consiguiente hipersecreción de prolactina. Una hiperprolactinemia crónica puede llevar a trastornos del ciclo menstrual (galactorrea y amenorrea) y probablemente también participe en la génesis de cáncer en la glándula mamaria.