sensibilidad de especies leñosas mediterráneas a la ... · seguimos extrañando por su ánimo y...
TRANSCRIPT

Sensibilidad de especies leñosas Mediterráneas a la presencia de zinc en el
medio. Interacción con disponibilidad hídrica
Karen Disante Bertone



Sensibilidad de especies leñosas Mediterráneas a la presencia de zinc en el
medio. Interacción con disponibilidad hídrica
Memoria presentada por Dña. Karen Disante Bertone
para optar al grado de Doctor por la Universidad de Alicante
Director Jordi Cortina i Segarra
Alicante, 21 de octubre de 2013


Jordi Cortina i Segarra, Catedrático de Ecología de la Universidad de Alicante, HACE CONSTAR: Que el trabajo descrito en la presente memoria, titulada: “Sensibilidad de especies leñosas Mediterráneas a la presencia de zinc en el medio. Interacción con disponibilidad hídrica” ha sido realizado bajo su dirección por Dña. Karen Disante Bertone en el Departamentos de Agroquímica y Bioquímica y en el Departamento de Ecología, ambos de la Universidad de Alicante, y reúne todos los requisitos necesarios para su aprobación como Tesis Doctoral.
Alicante, 21 de octubre de 2013
Jordi Cortina i Segarra DIRECTOR DE LA TESIS


A mi familia,
la de siempre y la de ahora…


Agradecimientos
El 2 de abril del año 2002, pisé España por primera vez gracias a una beca de la Agencia Española de Cooperación Internacional. Sólo venía por tres meses. Y 11 años después aquí sigo, pero ahora con la vida compartida a ambos lados del charco. Ahora, que cierro un capítulo de mi vida, quiero aprovechar a agradecer a todas esas personas que me han ayudado a mí y a esta tesis.
En primer lugar quiero agradecer a Jordi Cortina Segarra. Como director de tesis me ha ayudado profesionalmente desde el mismo día que llegué al departamento de Ecología. Al cabo de estos años me ha enseñado más de lo que él cree. Me ha motivado a orientar mi interés profesional hacia la ecología, y también, ha orientado muchos de sus proyectos atendiendo mi formación en biotecnología. Gracias por confiar en mí, por las discusiones de ciencia, por valorar mi opinión, no siempre acertada, por mantener la calma en los deadlines y por aceptar el ritmo de esta tesis con buen humor. Pero especialmente quiero darle las gracias por las muchas horas que dejó a un lado cuestiones científicas, para hablar, y para compartir alegrías. Gracias Jordi por la amistad y el compañerismo, que tengo la certeza que no dependen de los presupuestos en ciencia.
Quisiera darle las gracias al tribunal de esta tesis y a los revisores externos, por buscar tiempo para dedicarle a este trabajo.
Quiero agradecer las facilidades del departamento de Ecología y a su personal. A Fina, Emilio, Rosa, Juanfra, por aceptar cuestiones de todo tipo siempre a último momento, a los técnicos de laboratorio Fran, Jose, Silvia por su ayuda y a Clara y Mónica, por no quejarse de cómo dejamos la mesa del despacho ni el suelo del laboratorio.
A Susana Bautista por preocuparse de mi trabajo y de mí. Gracias Su! Al personal del CEAM, Esteban, Joanet y especialmente a Jaime y Alberto, por las veces que me han prestado ayuda, y no han sido pocas. A Jaime Puértolas quiero darle las gracias por atenderme siempre, y revisar esta tesis.
A Beatriz Amat por colaborar en esta tesis, por respetar tiempos y espacios y estar siempre cuando la necesitas. A Juame Tormo quiero agradecer su buen humor, su ayuda en la revisión de trabajos y sus conocimientos de R! A María, que aunque compartí con ella trabajo ajeno a esta tesis, he disfrutado mucho de las horas de laboratorio y otras muchas horas lejos de la universidad.

A todos los que han pasado por el departamento de Ecología y CEAM como becarios, contratados o estudiantes en prácticas. Algunos han colaborado en esta tesis de una manera muy directa y desinteresada, y otros, que colaboraron a formar un ambiente de trabajo más agradable, y para muestra, las paellas de San Alberto Magno! Al inagotable Santi, que me ha animado y ayudado, no sólo con cuestiones estadísticas. A Marian, Amina e Isa, porque quienes las conocimos las seguimos extrañando por su ánimo y ayuda. A Carolina y Juanjo, estudiantes que vinieron a hacer prácticas y acabaron preparando soluciones de Zn y que aún hoy me sonrío al recordarlos. A Azucena, Diana, Fran, Luna, Anna por los buenas horas de despacho y comida. A Noelia, Fede, Roberta, Valeria Bortolotti, Erdwin, Diego, Mirian Gobbi, Tadas Bitinas, Pere, Bea, Estrella, Soraya, Encarni, Hassam, Silvana, Nuria, Victor…Especialmente, quiero recordar a Juan Peña, una de las primeras personas que conocí al llegar y que ha dejado un hueco enorme.
Quiero agradecer a todos los becarios que han formado poco o mucho tiempo parte de la pecera, especialmente a tres pececitos que ya no están aquí: David Ramírez, Jorge Monerris y Anahí con quienes compartí mucho más que despacho. Es un enorme placer haberlos conocido.
No me cansaré de agradecer a Thanos y Olga, que a pesar de no seguir aquí y haber sido de los últimos en llegar, se convirtieron en amigos con los que sé que siempre podré contar. Al incansable Cholo, por ayudar, reír y estar siempre cuando alguien lo necesita.
A Alberto y Marina, por compartir los momentos importantes de mi vida. A Antonio, que vino con Rosario y se ha convertido en una amigo increíble. A Javi y Vane, quien como yo, vino sólo por tres meses y con quien he pasado de compartir café malo de máquina a tardes de parque con los bebés.
Y a todos esos locos bajitos que han venido al mundo en este tiempo para cambiar nuestras conversaciones y prioridades pero cuyas sonrisas nos siguen dando fuerza: Naiara, Irati, Álex, Héctor, Jimena, Blai, Bet, Garoé.
Quiero agradecer especialmente a Román, fue uno de mis primeros amigos aquí y junto a él y David he compartido mucho, casa, papeleos de extranjería y varias cervezas en el uruguayo. A Adela, por los buenos ratos en la universidad y fuera de ella, a Rosario, porque me enseña cada día a ser mejor, y a Marina, por ser simplemente Marina, y a las tres, por la terapia de grupo en la máquina de café!

Alejandro, Toñi, Pedro, si pienso en familia alicantina, pienso en ustedes tres. No sólo por los aportes de Alejandro a esta tesis, que han sido muy enriquecedores. Si tuviera que ponerle cara a ciertos momentos, vería las caritas de los tres en infinidad de ocasiones, cuando pienso en la nevera llena luego de un regreso triste desde Argentina, la primera visita de mi hijo, el ánimo en los momentos más difíciles, los calditos cuando Iván se enferma…...a los tres, Gracias.
La línea que separa lo profesional y lo personal es muy delgada y a menudo se desdibuja. Por eso quiero agradecer el cariño recibido de tanta gente que me ha ayudado a avanzar. Dani, Ale, Belén, Mica, Zarita, Mariano. Aunque nos vemos una vez al año, es como si siguiéramos viéndonos todas las semanas mezclando risas con cosas serias y porque sé, Be y Ale, que si estuvieran aquí, estarían ayudándome a maquetar esta tesis.
A toda esa gran familia que me encontré al llegar aquí y me recibió con los brazos abiertos. Manuela, Topi, Javi, Irma, Nacho y Pablo, Gracias por cuidar de mí.
No quiero dejar de agradecer a Juanan y Esther, nunca pensé que la ayuda prestada para aprender a usar el IRGA desencadenara en una amistad tan auténtica e incondicional. A Jorge y Raquel, por tantos años de amistad y de buenos momentos compartidos. A Ángeles, porque siempre está para echar una mano y una cerveza, y ahora la extraño.
Quiero agradecer desde aquí a toda mi familia de Pergamino, a mi tía Norma, a mis queridísimos primos, a sus parejas y a sus preciosísimos hijos. Todos han estado conmigo incondicionalmente. A mis tíos, Yuky y Martha, que me han ayudado en cada etapa. Especialmente quiero recordar a mi Abuela y tío Héctor, a los dos, muchas, muchas gracias.
Sé que mis padres no esperan un agradecimiento, todo lo que han hecho por mí ha sido desde el amor más profundo, pero de verdad, es un lujo tenerlos de padres. Les quiero agradecer que hayan entendido, respetado y compartido mi vida desde 12000km de distancia. Ale, si es verdad lo que me dijiste, que los hermanos son los seres con los que compartimos más tiempo, entonces yo soy la persona más afortunada del mundo. Gracias por ayudarme y regalarme a Juli, y también a Pablo, que siempre está para ayudarnos.
Por último quiero darles las gracias a Iván y David. Esta tesis no sería posible sin los dos. David más que mi pareja, es mi amigo y compañero en cada proyecto que

emprendo. He llegado hasta aquí y sigo aquí porque vos estás siempre a mi lado. Ahora que el futuro se ha vuelto más incierto, saber que estamos juntos es el mejor augurio de que vendrán buenos tiempos. Gracias por ayudarme desde el principio y por regalarme a Iván, que es el sol de mi vida, y quien nos empuja a seguir avanzando. Los tres formamos el mejor equipo que pueda encontrar.

i
Síntesis
El uso de metales pesados ha posibilitado el desarrollo de la sociedad industrial. Por ello, la entrada de estos elementos en suelos y agua continúa produciéndose, a pesar de las graves consecuencias que su acumulación puede conllevar. No en vano, la contaminación con metales pesados representa uno de los problemas ambientales más graves a escala mundial. Entre los metales pesados, el Zn es un micronutriente esencial para el desarrollo vegetal. Sin embargo, a altas concentraciones afecta al metabolismo, la morfología, y el desarrollo de las plantas.
El desarrollo industrial también es responsable de cambios drásticos en otros componentes del medio, como el clima. En regiones de clima seco, incluidas las mediterráneas, el agua es el principal factor limitante de la producción vegetal. En las próximas décadas, en estas zonas se producirá un aumento de la temperatura media, una disminución de las precipitaciones y una concentración de las mismas, con lo que la frecuencia e intensidad del estrés hídrico probablemente se acentuará.
Ante esta situación, es previsible que la combinación de contaminación por metales pesados y estrés hídrico afecte a áreas más extensas y de manera más intensa. No obstante, pocos estudios han abordado este tema hasta la fecha.
Para abordar estas cuestiones se han desarrollado diversos estudios que conforman esta Tesis doctoral. Su principal objetivo es evaluar los efectos de la aplicación de Zn sobre especies forestales mediterráneas, así como su interacción con la disponibilidad hídrica.
Se ha pretendido alcanzar este objetivo general a partir de los siguientes objetivos parciales:
1) Determinar índices toxicológicos para especies leñosas mediterráneas abundantes y especies clave, y por ello especies importantes para el funcionamiento de los ecosistemas y la restauración ecológica. Evaluar la relevancia de la adaptación a ambientes áridos en la respuesta al aumento de Zn.

ii
2) Evaluar la respuesta morfológica y fisiológica de plántulas de Q. suber a concentraciones crecientes de Zn. Determinar si la exposición a este elemento puede mitigar o agravar los efectos de la sequía.
3) Estudiar cambios hormonales y en la conductividad hidráulica de plántulas de Q. suber en respuesta al aumento de Zn. Determinar la existencia de sinergias y antagonismos en la respuesta morfológica y fisiológica de plántulas de esta especie a la combinación de baja disponibilidad hídrica y Zn.
4) Analizar el perfil metabólico de plántulas de Q. suber tratadas con Zn y expuestas a niveles de riego contrastados. Identificar respuestas comunes y específicas a los dos factores de estrés evaluados.
Los objetivos 1, 2 y 3 se abordan en los trabajos que conforman los Capítulos 2-4 de esta memoria, y han sido publicados en revistas cuyo alcance científico abarca los temas tratados en cada trabajo. El último objetivo se desarrolla en el trabajo incluido en la memoria como Apéndice 1 y ha sido enviado para su publicación.
En el Capítulo 2 de la memoria se ha partido de la hipótesis de que los rasgos morfológicos de especies adaptadas a la sequía podrían favorecer la tolerancia al Zn y, como resultado, las especies de ambientes xéricos podrían exhibir mayor resistencia a este elemento que las especies de ambientes más mésicos. Un objetivo derivado de este capítulo es la identificación de especies útiles para la restauración de sitios contaminados con este elemento.
Nuestra hipótesis parte de observaciones previas que sugieren que las especies que colonizan suelos con altos contenidos de metales poseen características morfológicas similares a aquellas adaptadas a la sequía. Además, debido a las propiedades físicas del suelo, las especies que crecen en suelos con altos contenidos de metales de origen natural con frecuencia están expuestas a baja disponibilidad hídrica.
En base a la hipótesis y a los objetivos planteados, plántulas de especies representativas de la cuenca Mediterránea, fueron expuestas a concentraciones crecientes de Zn. Los resultados obtenidos mostraron que las respuestas morfo-fisiológicas de las especies leñosas mediterráneas a Zn fueron específicas de la especie. Contrario a nuestra hipótesis, la resistencia de las especies al Zn

iii
(evaluada mediante índices de toxicidad) se relacionó positivamente con la precipitación anual mínima de los hábitats de las especies analizadas. Quercus suber resultó la especie más resistente, probablemente debido a su capacidad para excluir Zn en el estadío de plántula, mientras que Tetraclinis articulata fue la especie más sensible.
Entre las especies evaluadas, las más adaptadas a ambientes más mésicos, Quercus suber y Pinus pinea fueron también las más resistentes al Zn y podrían tenerse en consideración en la restauración de zonas contaminadas por Zn que presenten este tipo de clima. De un modo similar, Pinus halepensis fue la especie más resistente al Zn entre las especies más adaptadas a zonas xéricas. Aunque nuestros resultados surgen de estudios realizados en condiciones controladas, suponen un punto de partida para futuras pruebas de campo. Además, de acuerdo a los patrones de acumulación y a la capacidad de acumular metales de las especies analizadas, es importante destacar que ninguna de las especies estudiadas mostró potencial para la fitorremediación.
Por un lado, los resultados del Capítulo 2 mostraron que dosis altas de Zn pueden alterar negativamente el desarrollo radicular, y por lo tanto afectar la capacidad de las plantas de captar agua y nutrientes. Por otro lado, además de ser un micronutriente esencial para el desarrollo vegetal, hay estudios que sugieren que, en dosis adecuadas, el Zn podría favorecer la resistencia de las plantas al estrés hídrico. Por ello, como hipótesis del Capítulo 3 planteamos que la capacidad de Q. suber para resistir un periodo de estrés hídrico sería favorecida por pequeñas dosis de Zn y por el contrario, una alta disponibilidad de Zn en la solución del suelo podría intensificar los efectos negativos del estrés hídrico. En este estudio expusimos plántulas de Q. suber a concentraciones crecientes de Zn (3, 10, 50 y 150 µM) y luego de un tiempo, suspendimos el riego y realizamos una caracterización morfológica. Durante el periodo de sequía, evaluamos el estado fisiológico de las plantas.
Los resultados obtenidos mostraron que la morfología y fisiología de plántulas de Q. suber bien regadas fueron negativamente afectadas por el aumento de Zn en el medio. El efecto negativo del Zn fue especialmente evidente a partir de concentraciones externas superiores a 50 µM. La conductancia estomática disminuyó notablemente con el aumento de Zn y la eficiencia fotoquímica mostró

iv
un patrón similar, sugiriendo la existencia de fotoinhibición cuando la disponibilidad de Zn fue alta. Contrariamente a lo esperado, la adición de pequeñas cantidades de Zn no tuvo un efecto positivo sobre la morfología y la fisiología de las plántulas y, por lo tanto, sobre su capacidad para responder al estrés hídrico. Sin embargo, cuando el riego cesó, las plantas control (expuestas a 3 µM de Zn), con alta conductancia estomática, mostraron una rápida pérdida de humedad y un descenso brusco del intercambio de gases. En contraste, el cierre de estomas inducido por elevadas dosis de Zn, pudo favorecer la conservación del contenido hídrico y el mantenimiento de la asimilación de CO2 durante un corto periodo de sequía. Sin embargo, debemos ser cautos al extrapolar nuestros resultados a periodos más largos de sequía. En estas condiciones, los efectos negativos del Zn sobre la fisiología y, especialmente, sobre la morfología radicular, podrían limitar el acceso a los recursos y por lo tanto, limitar el crecimiento de la planta.
La ausencia de efectos negativos de Zn que agravasen las consecuencias de un periodo corto de estrés hídrico, nos condujo a preguntarnos sobre la respuesta de las plántulas cuando los dos factores de estrés, Zn y escasez hídrica, actúan simultáneamente. Así, la hipótesis del Capítulo 4, fue que las respuestas morfo-fisiológicas a una baja disponibilidad hídrica podrían mitigar los efectos de la alta disponibilidad de Zn.
Para validar esta hipótesis, expusimos plántulas de Q. suber a concentraciones crecientes de Zn (3, 10, 50 y 150 µM) y a dos regímenes de riego bien diferenciados (HW y LW, alta y baja frecuencia de riego, respectivamente). El análisis de las plantas mostró que la concentración de Zn en las hojas y las raíces aumentó con el incremento Zn en la solución nutriente, independientemente del régimen de riego. Sin embargo, la dosis externa más alta de Zn, indujo concentraciones tisulares casi dos veces superiores en plantas HW que en plantas LW. La respuesta de la mayoría de variables evaluadas dependió de la interacción entre los dos factores de estrés. El aumento de Zn en plantas HW se tradujo en una disminución de la conductancia hídrica del sistema radicular, una pérdida de funcionalidad de las hojas y un sistema radicular menos desarrollado, en comparación con las plantas control (3µM Zn, HW). Además de estos efectos negativos, la concentración de Zn más alta indujo un aumento de la acumulación de auxina IAA. Por el contrario, en plantas LW la acumulación de biomasa

v
radicular se vió favorecida por altas dosis de Zn, y la conductancia estomática aumentó a dosis intermedias de este metal. Otras variables mostraron tendencias similares.
En resumen, estos resultados indican la ausencia de sinergia entre ambos factores de estrés, aunque la combinación de Zn y baja disponibilidad hídrica no revirtió el estado de las plantas hacia un estado semejante al de las plantas control. Además, los resultados sugieren que la disminución de disponibilidad hídrica podría reducir la toxicidad de Zn al limitar su acumulación en tejido.
En los Capítulos 2-4 evaluamos distintos aspectos de la respuesta de plántulas al Zn y a la interacción de Zn con baja disponibilidad hídrica. Un aspecto poco estudiado de esta interacción lo constituye la respuesta del metaboloma como un todo y de los metabolitos de forma individual. Por ello, en el trabajo que se presenta como Apéndice 1, utilizamos el análisis metabolómico como herramienta de estudio, con el fin de analizar cambios inducidos por Zn y niveles de riego contrastados sobre plántulas de Q. suber.
El metaboloma de Q. suber se vió afectado por el Zn y por la sequía, aunque no observamos interacción entre ambos factores de estrés. A escala de metabolito, el estudio permitió identificar tres grupos de compuestos: (i) metabolitos específicos de cada tipo de estrés (ácido málico, aspartato, ácido α-cetoglutárico, etc., en respuesta al Zn, y ácido quínico, manosa, fructosa, entre otros, en respuesta a la sequía), (ii) metabolitos cuya acumulación varió en respuesta a los dos factores de estrés actuando de manera aislada (glucosa, ramnosa, ácido cítrico y una hexosa), y (iii) metabolitos cuya respuesta dependió de la combinación de ambos factores de estrés (α- tocoferol, glicerol, xilitol). El primer grupo de metabolitos constituirían indicadores de un tipo de estrés específico, mientras que el segundo grupo podría formar parte de un mecanismo de respuesta general al estrés de Q. suber.
En conjunto, los datos presentados en esta Tesis muestran que las especies leñosas Mediterráneas son sensibles al aumento de Zn en el medio. El aumento de la concentración de Zn indujo cambios notables en la morfología, fisiología y bioquímica de las plantas expuestas, aunque estas respuestas se produjeron a diferentes valores umbrales. Nuestros resultados nos permiten identificar

vi
variables de respuesta sensibles a pequeños incrementos de Zn en el medio y otras variables cuyo valor sólo cambia significativamente ante concentraciones foliares de Zn muy elevadas (Apéndice 2). Esta información se integró para elaborar el modelo general de respuesta al Zn que se muestra en el Capítulo 5. El modelo presentado también contempla datos nutricionales, que aunque no se han incluido en ningún de los trabajos publicado hasta la fecha, han sido incorporados a esta Tesis en forma de Apéndice (Apéndice 3), pues consideramos que son relevantes para explicar los cambios observados y pueden ser de utilidad para futuros estudios.
La combinación de Zn y escasez hídrica, de forma secuencial o simultánea, no generó efectos sinérgicos. Por el contrario, nuestros datos sugieren que algunas respuestas morfo-funcionales inducidas por cada factor podrían mitigar los efectos negativos del otro. En base a estos resultados, en el Apéndice 4 revisamos de manera sucinta la actual legislación sobre contaminación de metales pesados, intentando señalar sus carencias.
Por último, mencionar que en este trabajo hemos identificado respuestas comunes a ambos tipos de estrés, pero nuestros resultados refuerzan la idea de que, paralelamente, existen mecanismos de respuesta específicos de cada tipo de estrés.

vii
Índice
Capítulo 1. Introducción.……………………………………………………………………… 1
1. Metales pesados…………………………………………………………………….. 3
1.1. La problemática de los metales pesados …………………… 4
1.2. Zinc: caso de estudio …………………….………………………... 7
2. Zn en el sistema suelo-planta …………………….…………………………… 10
2.1. Zn en suelo …………………….…………………….………………… 10
2.2. Biodisponibilidad ………….…………………….………………….. 10
2.3. Absorción y transporte ….…………………….…………………… 12
2.4. Acumulación ….…………………….…………………………………. 13
3. Zn en las plantas ….…………………….…………………………………………. 14
3.1. Efectos sobre la fisiología y la morfología vegetal……... 14
3.2. Sensibilidad de las especies leñosas al Zn …………………. 16
4. Estrés múltiple ….…………………….…………………………………………….. 17
4.1. Teoría del estrés: respuesta general vs. respuesta específica ……..……………….……………………………………………… 17
4.2. Respuesta al estrés múltiple..…….………………………………. 18
4.3. Interacción entre estrés por metales y sequía …………… 20
5. Umbrales de fitoxicidad. El contexto de la restauración ecológica de sitios contaminados. ……………………………………………………………. 21
6. Contexto del trabajo realizado y objetivos ……………………………….. 22
Capítulo 2. Sensitivity to zinc of Mediterranean woody species important for restoration……………………………………………………………………………………… 27
Capítulo 3. Response to drought of Zn-stressed Quercus suber L. seedlings. 53
Capítulo 4. Alleviation of Zn toxicity by low water availability……………….. 75
Capítulo 5. Discusión general ………………………………………………………………. 105
1. Zinc en especies Mediterráneas ………………………………………………. 107

viii
1.1. Aproximación experimental y sensibilidad de las variables morfo-funcionales al aumento de la biodisponibilidad de Zn ………………………….……………………… 107
1.2. Modelo conceptual: morfología y fisiología de plántulas expuestas al Zn ……………………………………………………………… 113
2. Zn y disponibilidad hídrica ……………………………………………………. 118
2.1. La combinación de Zn y escasez hídrica. Relaciones causa-efecto. ………………………………………………………………… 118
2.2. Respuestas comunes a dos factores de estrés actuando de forma aislada ……………………………………………………………. 120
3. Caracterización experimental de la tolerancia al Zn ………………….. 123
3.1 Consideraciones sobre la relación entre adaptación a zonas áridas y umbrales de toxicidad al Zn……………………… 123
Conclusiones …………………………………………………………………………………….. 127
Referencias ……………………………………………………………………………………….. 131
Apéndices ………………………………………………………………………………………… 155
Apéndice 1. Metabolome responses of Quercus suber seedlings to zinc concentration and water availability……………………………………. 155
Apéndice 2. Relación entre concentración de Zn foliar y variables de respuesta Ecofisiológica ………………………………………………………. 178
Apéndice 3. Concentración de nutrientes en hojas y raíces de plantas de Quercus suber de 5 meses de edad expuestas a cuatro niveles de Zn y frecuencias de riego contrastadas…………………………. 179
Apéndice 4. Normativa sobre metales pesados …………………………….. 181

Introducción


Introducción
3
1. Metales pesados
La explotación y utilización de los metales ha sido fundamental en el desarrollo de la sociedad moderna (Wilson, 1996). La gama de aplicaciones de los metales es muy amplia, y el uso de algunos de ellos (generalmente denominados metales pesados) conlleva un precio ambiental significativo. Sin embargo, nuestra dependencia a los metales sigue dando lugar a grandes aportes al medio, a pesar de las graves consecuencias que pueden ocasionar (Han et al., 2002).
En las últimas décadas, la contaminación de suelos naturales y agrícolas con metales pesados ha aumentado notablemente, y actualmente se considera un problema grave en todo el mundo (Boyd, 2010). Aunque algunos metales pesados, como Zn, Fe y Cu, desempeñan un papel importante en el metabolismo de la planta como micronutrientes esenciales, elevadas concentraciones de los mismos pueden interferir con una amplia gama de procesos fisiológicos y bioquímicos, provocando inhibición del crecimiento vegetal y baja productividad (Kabata-Pendias y Pendias, 1992; Nagajyoti et al., 2010). Del mismo modo, la continuada industrialización a escala mundial ha inducido cambios drásticos en la biología, física y química del medioambiente, incluyendo cambios en la composición y diversidad de especies vegetales, el aumento de las concentraciones de CO2 atmosférico, el incremento de la temperatura media anual, y cambios en los patrones de precipitación, entre otros (IPCC, 2007). Además, factores como la temperatura y la precipitación, susceptibles de sufrir grandes modificaciones debido al cambio climático, pueden influenciar la distribución de químicos tóxicos (Noyes et al., 2009). Por ejemplo, el calentamiento del suelo podría favorecer la descomposición de la materia orgánica, la lisis de la célula microbiana y la destrucción de los agregados del suelo, aumentando así la biodisponibilidad de metales (Antoniadis y Alloway, 2001; Sardans et al., 2008). De un modo similar, una acidificación del suelo, podría reducir la adsorción de metales pesados en la fracción orgánica y mineral del suelo (Ryan et al., 2001; Yanai et al., 2006; Rajkumar et al., 2013), dando como resultado la liberación de metales (Balsberg Påhlsson, 1989). Por otro lado, la disminución de la disponibilidad hídrica podría reducir la movilidad de los metales (Pascual et al., 2004).
Es decir, diversos aspectos relacionados con el cambio climático y sus consecuencias podrían modificar indirectamente la biodisponibilidad de ciertos

Capítulo I
4
contaminantes, incluyendo metales pesados. Además, estas condiciones estresantes podrían actuar de manera combinada con los efectos tóxicos de los metales y ejercer efectos sinérgicos negativos sobre el crecimiento y desarrollo de las plantas que viven en suelos contaminados y sobre aquellas especies que, por actividades antrópicas, son expuestas a aumentos graduales de los niveles de metales en su entorno.
1.1. La problemática de los metales pesados
El término metal pesado se refiere a cualquier metal o metaloide cuya densidad atómica es mayor que 5 g/cm3 (Hawkes, 1997; Adriano, 2001). Dentro de los metales pesados encontramos plomo (Pb), cadmio (Cd), níquel (Ni), cobalto (Co), hierro (Fe), zinc (Zn), cromo (Cr), arsénico (As), plata (Ag) y elementos del grupo del platino.
Varios autores han discutido el uso del término metal pesado para referirse a un grupo de elementos con distintas propiedades físico- químicas y con funciones biológicas contrastadas (Woolhouse, 1983; Alloway, 1995). Así mismo, utilizan el término elemento traza para referirse a elementos que normalmente se encuentran en ambientes naturales y perturbados en cantidades pequeñas y que a elevadas concentraciones pueden resultar tóxicos para los seres vivos (Adriano, 2001). En este trabajo utilizaremos el término metal pesado, de manera general, para referirnos a todos aquellos metales que pueden resultar potencialmente tóxicos para los seres vivos (Poschenrieder y Barceló, 2004a).
Entre los metales pesados se encuentran elementos que son esenciales para las plantas y animales, elementos que pueden resultar beneficiosos para el crecimiento y desarrollo de algunas especies, bajo determinadas circunstancias, y otros cuyo efecto positivo para los seres vivos no ha sido descrito hasta el momento. En determinadas cantidades, cualquier metal puede resultar tóxico para los seres vivos (Kabata-Pendias y Pendias, 1992). Diferentes trabajos muestran un incremento de la presencia de metales pesados en zonas cercanas a grandes urbes, zonas industriales, minas metalíferas y autovías, probablemente debido al aumento de la producción y demanda de metales en países desarrollados (Alloway, 1995; Adriano, 2001). En España y en concreto en la Comunidad Valenciana, diversos estudios han mostrado el incremento de metales pesados en suelo como consecuencia de actividades mineras, industriales y urbanas (Roca-Perez et al., 2010; Oyarzun et al., 2011; Micó et al., 2006)

Introducción
5
Los metales pesados como contaminantes ambientales constituyen un problema ecológico, evolutivo, nutricional y ambiental que se ha ido agravando en los últimos años (Nagajyoti et al., 2010). Pero la presencia de metales pesados en el medio no sólo tiene un origen antrópico.
Fuentes naturales de metales pesados
La fuente natural más importante de metales pesados es la roca madre. La composición y concentración de metales pesados depende del tipo de roca y de las condiciones ambientales que activan los procesos de meteorización (Nagajyoti et al., 2010). Los metales procedentes de la roca madre entran en el medio al ser disueltos y particulados, o al ser inyectados directamente en la atmósfera en el caso de emisiones procedentes de erupciones volcánicas. Estas dos entradas de metales suponen el 80% del metal total procedente de fuentes naturales (Callender, 2004). El polvo arrastrado por el viento, aerosoles marinos e incendios forestales contribuyen al transporte de metales entre distintos ambientes aunque es escasa la bibliografía que cuantifique estos aportes (Callender, 2004; Nagajyoti et al., 2010).
Fuentes antrópicas de metales pesados
Existen numerosas fuentes antrópicas de metales. Estas fuentes pueden agruparse según su procedencia. Las fuentes industriales incluyen: minería, fundición, refinería y cerámica. Las dos primeras actividades suponen la mayor aportación antropogénica de metales al medio (Callender, 2004; González et al., 2011). La contaminación del suelo a partir de sedimentos provenientes de minas metalíferas es una fuente común de contaminación de metales en todo el mundo. Ejemplo de esto es lo ocurrido en Aznalcóllar en el año 1998, donde una balsa de residuos de de metales pesados procedentes de una mina de la zona, se rompió vertiendo gran cantidad de metales al río Guadiamar y a las tierras circundantes (Clemente et al., 2003; Foto 1).
Las actividades industriales basadas en la combustión de petróleo, así como la producción de cemento, las plantas nucleares, líneas de alta tensión, estaciones eléctricas, el procesamiento de plástico, papel, textiles, microelectrónica y

Capítulo I
6
conservación de madera también liberan a la atmósfera metales como Se, B, Cd, Cu, Zn, Cs y Ni (Callender, 2004).
Foto 1. Ejemplo catastrófico de contaminación del medio por metales pesados resultante de los procesos de minería. En este caso se muestra la rotura de la balsa de residuos en Aznalcóllar en el año 1998 (Fuente: http://xurl.es/npg9s)
Las aguas residuales no depuradas constituyen probablemente la principal fuente de metales pesados en ríos y lagos y la escorrentía urbana se considera una fuente importante de contaminación de aguas superficiales También se ha observado que los detergentes enzimáticos contienen a menudo trazas de Fe, Mn, Cr, Co, Zn, Sr and B (Nagajyoti et al., 2010).
Los fertilizantes orgánicos e inorgánicos son la principal fuente de metales pesados en suelos agrícolas. Los fungicidas y fertilizantes fosforados contienen cantidades variables de Cd, Cr, Ni, Pb y Zn, dependiendo de sus orígenes. Su uso prolongado puede conducir a la acumulación de cantidades peligrosas de metales (Verkleij, 1993). Otras fuentes de contaminación asociada a la agricultura son herbicidas, insecticidas, enmiendas orgánicas procedentes de residuos animales y lodos de depuradora (Nagajyoti et al., 2010).
El uso de biosólidos (subproductos generados durante el proceso de depuración de aguas residuales) en la restauración de zonas degradadas y en la formulación de substratos de cultivo para la producción viverística, representan nuevos escenarios de introducción de contaminantes en suelos naturales (Knight et al., 1997; Adriano, 2001; Fuentes et al., 2007a). Los lodos de depuradora o biosólidos, presentan altos contenidos de materia orgánica y nutrientes que, a priori, les confieren un alto potencial para su utilización en la restauración de zonas

Introducción
7
degradadas. El uso de lodos en restauración forestal ha aumentado en las últimas décadas debido al incremento de producción de los mismos, que hizo necesaria la diversificación frente al uso predominantemente agrícola, y a su valor fertilizante (Valdecantos et al. 2001, 2006; Querejeta 2001, Fuentes et al 2007a). Sin embargo, los biosólidos pueden contener cantidades elevadas de metales pesados, lo que, unido a la presencia de elevadas cantidades de sales solubles y patógenos, puede comprometer su efecto beneficioso (Adriano, 2001; Fuentes et al., 2010).
En concreto, según datos de la Entidad Pública de Saneamiento de Aguas Residuales (EPSAR, 2009), un 10% de los lodos potencialmente utilizables en agricultura eran descartados para este uso debido a los altos niveles de Cu, Ni y Zn. Si bien los tres metales están descritos como micronutrientes esenciales, a altas dosis, pueden afectar el desarrollo y crecimiento vegetal (Marschner, 1995). Ciertos trabajos sugieren que el Zn, además, puede estimular la resistencia de las plantas a la sequía y a altas temperaturas (Kabata-Pendias y Pendias, 1992; Gadallah, 2000). Su carácter de micronutriente, la toxicidad en concentraciones elevadas y su interacción con la economía hídrica de las plantas, unido a su abundancia en los lodos de depuradora de la Comunidad Valenciana, justifica la elección de este elemento para el presente estudio.
1.2. Zinc: caso de estudio
El Zn ocupa el puesto 23 entre los elementos más abundantes de la tierra, y es el segundo metal de transición más abundante en los organismos, después del Fe (Broadley et al., 2007). Este metal es un constituyente natural de los suelos y un micronutriente esencial para humanos, animales y plantas (Marschner, 1995; Hagemeyer, 2004). Las fuentes de Zn naturales y antropogénicas son similares a las de otros metales. En las últimas décadas, la concentración de Zn en los suelos de todo el mundo ha incrementado como consecuencia de actividades humanas (Alloway, 1995; Pacyna y Pacyna, 2001).
Si bien existen numerosas evidencias del impacto humano en la biogeoquímica global del Zn, no existen datos recientes sobre emisiones globales de Zn. El inventario más reciente de metales pesados y metaloides, según nuestro conocimiento, incluye estimaciones totales de emisión para los diferentes continentes y diversas categorías de fuentes, y se refiere a 1995 (Pacyna y Pacyna,

Capítulo I
8
2001). La Tabla 1 muestra datos de emisión de Zn agrupados según las principales categorías de fuentes antropogénicas.
Las emisiones globales de Zn supusieron 57.010 toneladas por año, valor superado sólo por V (240.255 Mg a-1), Pb (119.259 Mg a-1) y Ni (95.287 Mg a-1). Las emisiones del resto de metales (Mn, Mo, As, Cd, Cr, Cu y Hg entre otros) fueron muy inferiores al valor del Zn. De acuerdo a este trabajo, la producción de metales no ferrosos constituye la mayor fuente de emisiones de Zn a la atmósfera (72% del total). El mismo estudio muestra además, que las emisiones procedentes del continente asiático representaron el 60% del valor total. Las fuentes antrópicas de Zn procedentes de Europa fueron responsables del 13,5% de las emisiones mundiales (Pacyna y Pacyna, 2001). Los autores indican en el trabajo que, en general, los datos de Zn son a menudo subestimados debido a la falta de conocimiento de las fuentes de emisión de este elemento.
Tabla 1. Emisiones mundiales de Zn a la atmósfera en la década de 1990 – 1999 según las principales fuentes antrópicas. Fuente: Pacyna y Pacyna (2001)
Fuente Zn (Mg año-1) Combustión estacionaria de combustibles fósiles 9.417 Producción de metales no-ferrosos 40.872 Producción de hierro y acero 2.118 Producción de cemento 2.670 Eliminación de residuos 1.933 Total 57.010
Respecto a la Unión Europea (UE), las emisiones de Zn en 2011 en el conjunto de los 27 países miembros se redujeron en un 42% respecto al valor de 1990, aunque, debido a la ausencia de datos en algunos países, este número podría estar subestimado. Sólo tres Estados miembros contribuyeron a más del 10% de las emisiones de Zn en 2011: Alemania, Polonia e Italia. España fue el quinto país que mayor emisión de Zn tuvo en el año 2011 (393 Mg) y además, mientras que en la mayoría de países la emisión de Zn en 2011 se redujo respecto a años anteriores, en España aumentó un 52% respecto al valor de 1990.
En el ámbito español, la bibliografía muestra un aumento generalizado de concentración de Zn en suelos y vegetación. Por ejemplo, el impacto humano

Introducción
9
sobre el incremento de la concentración de Zn en suelos Mediterráneos se evidenció mediante el análisis de suelos industriales y agrícolas, y su posterior comparación con suelos naturales no contaminados (Roca-Perez et al., 2010). En cuanto a concentraciones de Zn en tejido vegetal, Peñuelas y Filella (2002) analizaron la concentración mineral en 24 especies de plantas vasculares y 3 de briófitas de un herbario procedente del noreste de España. El objetivo del trabajo fue realizar un estudio retrospectivo de la evolución de la contaminación por metales y otros elementos durante el pasado siglo (Peñuelas y Filella, 2002). En el caso del Zn, su concentración comenzó a incrementar a comienzos de siglo, y entre los años 1985 y 1995 su valor fue un 68% y un 24% superior al valor determinado en los años 1929-1930 en briófitas y en plantas vasculares, respectivamente. De acuerdo con los autores del trabajo, estos datos proporcionan, en primer lugar, evidencia biológica sobre la alteración de la biogeoquímica global durante el siglo XX como consecuencia de las actividades humanas y, en segundo lugar, indican que los metales que alcanzan el medio son retenidos, al menos en parte, en ecosistemas terrestres aunque éstos no se encuentren cerca de áreas industriales o urbanizadas. En el sudeste de España, minas localizadas en distintos puntos como las que se encuentran en las zonas de Cartagena-La Unión, en Mazarrón, o en Rodalquilar han contribuido al incremento de Zn en ecosistemas terrestres y costeros (Oyarzun et al., 2009, 2011; Riba et al., 2010; Conesa y Faz, 2011).
Actualmente, la actividad minera en los países más desarrollados se realiza en el marco de leyes ambientales y, normalmente, se acompaña de trabajos de restauración. Sin embargo, operaciones mineras que finalizaron hace décadas y que se realizaron en ausencia de regulaciones ambientales, han dejado residuos en el suelo que todavía suponen una fuente de contaminación y de riesgo ambiental para el entorno (Conesa et al., 2006).
En la Comunidad Valenciana, como se mencionó anteriormente, el Zn es uno de los tres metales más abundantes en los residuos generados durante el proceso de depuración de aguas domésticas e industriales. Aunque el promedio de concentración de Zn en lodos de depuradora domésticos se encuentra en niveles considerablemente menores a los marcados por la legislación vigente para su aplicación agrícola, la aplicación masiva y sistemática de estos residuos en agricultura puede suponer ciertos riesgos, debido a su acumulación en el suelo a largo plazo y a aplicaciones poco controladas.

Capítulo I
10
2. Zn en el sistema suelo-planta
2.1. Zn en suelo
La concentración de Zn en suelo suele venir definida en la bibliografía como concentración total, es decir mg Kg-1, µg g-1, etc. Sin embargo, este valor puede resultar un valor confuso si se pretende comparar resultados, ya que la biodisponibilidad de Zn puede variar dependiendo del tipo y determinadas propiedades del suelo como pH, contenido de materia orgánica y arcillas, etc. Por este motivo se considera que la concentración de Zn en la solución del suelo refleja con mayor precisión la cantidad de Zn disponible para la planta (Knight et al., 1997). En general, se utiliza cada vez más la biodisponibilidad de contaminantes en lugar de contenido total, como un indicador clave de los posibles riesgos para el medio (Adriano et al., 2004; González et al., 2011).
Según Kabata-Pendias y Pendias (1992) la concentración de Zn en solución en un amplio rango de suelos naturales se encuentra entre 1,5 y 262 µM. Knight et al. (1997) analizaron la concentración de metales pesados en diferentes tipos de suelo de toda Europa que habían sido expuestos a metales pesados, y la máxima concentración de Zn en solución que encontraron fue 20 µM. En contraste, otros estudios encontraron valores de Zn más elevados en sitios también expuestos a metales pesados. Por ejemplo, en un estudio realizado en Almería en una zona donde hasta hace pocas décadas se explotó una mina de azufre, se tomaron muestras de suelo de un vertedero de residuos de la mina, de sedimentos fluviales y de puntos no ocupados por la minería. La concentración de Zn determinada en las soluciones de suelo de estos tres sitios fue respectivamente 1.468 ± 887; 90 ± 157; 3,7 ± 4,0 µM (Media y desviación estándar; González et al., 2011).
2.2. Biodisponibilidad
El Zn presente en el suelo se encuentra distribuido en distintas fracciones: (i) soluble, (ii) intercambiable, (iii) adsorbido, quelado o complejado (unido a ligandos orgánicos), (iv) contenido en minerales secundarios arcillosos y óxidos metálicos insolubles, y (v) contenido en minerales primarios. El número de grupos de esta clasificación puede variar levemente, dependiendo del autor (Alloway 1995; Reichman, 2002, Broadley et al., 2007).

Introducción
11
Sólo las fracciones de Zn soluble y aquéllas que pueden ser solubilizadas se consideran biodisponibles para las plantas (Alloway, 1995). Por lo tanto, los factores que afecten a la concentración y especiación del Zn en la solución del suelo también afectarán a su biodisponibilidad (Reichman, 2002). La distribución de metales entre las fases sólidas y líquidas del suelo, así como su solubilidad y disponibilidad depende en gran medida de las características químicas del metal y su especiación, y de las características físico-químicas del suelo como pH, contenido de materia orgánica, fracciones finas y óxidos de Fe, Al y Mn, fuerza iónica de la solución del suelo, potencial redox y capacidad de intercambio catiónico, entre otras (Alloway, 1995; Rieuwerts et al., 1998; Reichman, 2002). Además de las condiciones físico-químicas, los flujos de metales entre las distintas fases varían con el tiempo (Rieuwerts et al., 1998). La Figura 1, muestra de manera simplificada las formas de los metales en el suelo y la velocidad de cambio entre las distintas formas.
Figura 1. Esquema simplificado de las formas de Zn edáfico y escalas temporales de las transformaciones entre las diferentes formas.
De los diversos factores que influyen en la distribución de Zn entre las distintas fracciones, el principal es el pH (Rieuwerts et al., 1998; Martínez y Motto, 2000; Broadley et al., 2007). En la revisión de Alloway (1995) podemos encontrar la siguiente ecuación que da una idea de la importancia de esta relación:
–log[Zn2+] = pZn = 2 pH – 5.8
El pH afecta también a la solubilidad de otros metales. En general, se observa una mayor retención y menor solubilidad de los cationes metálicos cuando los valores de pH son altos (García-Miragaya, 1984; Stahl y James, 1991; Basta et al., 1993). Diversos estudios muestran la relación inversamente proporcional y lineal entre el pH y la solubilidad de Zn en suelos ácidos, y no lineal en suelos calcáreos
No disponible
Minerales primarios
Intermedio
Minerales secundarios
Lábil
Compuestos e iones
adsorbidos
Zn Soluble
Iones y complejos solubles
Años Meses Días

Capítulo I
12
(Martínez y Motto, 2000). Una eventual bajada de pH da paso a la liberación de los metales pesados unidos a distintas fracciones, incrementando así su disponibilidad y absorción por las plantas (Balsberg Påhlsson, 1989), o su lixiviación hacia capas de suelo más profundas (Karathanasis et al., 2005; Toribio y Romanyà, 2006).
Por otra parte, el Zn puede ser adsorbido por la materia orgánica, los filosilicatos y los carbonatos del suelo (Alloway, 1995; Kabata-Pendias y Pendias, 2001). A la formación de carbonatos metálicos en el suelo se ha atribuido parte de la retención de Zn (Illera et al., 2000). La disponibilidad de metales pesados se puede ver notablemente reducida en zonas secas y semiáridas (Pascual et al., 2004), con frecuencia asociadas a suelos carbonatados (Fuller y Tucker, 1977). La presencia o aplicación de materia orgánica al suelo normalmente conlleva una reducción de la disponibilidad de metales (Pascual et al., 2004; Sánchez-Monedero et al., 2004), aunque en el caso del Zn también puede acarrear un aumento de su movilidad, ya que puede estar asociado a las fracciones solubles de la materia orgánica.
2.3. Absorción y transporte
La interacción con otros elementos presentes en la solución del suelo puede afectar a la absorción de metales por las plantas (Greger, 2004). Las plantas han desarrollado estrategias para regular la absorción de metales, para maximizarla, cuando son limitantes, o para minimizarla cuando se encuentran en concentraciones elevadas. Las raíces son capaces de alterar la solubilidad y especiación de metales en la rizosfera mediante la liberación de compuestos quelantes y la alteración del pH (Reichman, 2002). En general, la absorción de metales aumenta con el incremento de la concentración de metal en el medio pero este aumento no es lineal (Greger, 2004). La eficiencia de la acumulación es mayor cuando la concentración de metal por superficie de absorción es inferior al nivel de saturación, disminuyendo para concentraciones más altas.
El movimiento de metales desde la solución externa hacia la pared celular es un proceso pasivo, dirigido por difusión o flujo de masas (Marschner, 1995; Greger, 2004). Los metales se unen a la pared celular mediante un intercambiador iónico de baja afinidad y baja selectividad. Luego, los sistemas de transporte y los sitios de unión de alta afinidad median y dirigen la absorción a través de la membrana plasmática (Marschner, 1995; Clemens et al., 2002). Diversos estudios indican que

Introducción
13
la absorción de Zn se realiza mediante transportadores específicos (Lasat et al., 2000; Pence et al., 2000).
Existe consenso sobre la forma de Zn predominantemente absorbida por las raíces: el Zn+2, aunque también puede ser absorbido como formas hidratadas, complejos y quelatos orgánicos de Zn (Alloway, 1995; Kabata Pendias y Pendias, 2001; Broadley et al., 2007). Una vez dentro de la planta, la translocación de los metales a la parte aérea se produce a través de dos mecanismos principales, vía xilema y vía floema (Reichman, 2002). En el caso del Zn, su transporte se realiza predominantemente dentro del xilema en forma de quelato de ácidos orgánicos promovido por la transpiración (Kabata Pendias y Pendias, 2001; Reichman, 2002; Greger, 2004).
2.4. Acumulación
La tarea de la membrana celular es concentrar los elementos nutrientes y, aunque no totalmente, excluir los elementos tóxicos (Greger, 2004). Debido a que algunos metales actúan como micronutrientes esenciales, la regulación estricta de la acumulación de estos metales dentro de unos límites fisiológicos estrechos es de vital importancia, no sólo a nivel celular, sino para el organismo completo. Por ejemplo, los iones de Zn son componentes estructurales o catalíticos clave en las proteínas de unión al ADN y enzimas hidrolíticas, respectivamente. Sin embargo, un exceso de Zn podría inducir una inactivación de las proteínas (Clemens et al., 2002). Un fenómeno común a varios metales es su acumulación e inmovilización en raíces. En la bibliografía se menciona la existencia de barreras fisiológicas con respecto al transporte de metales como Zn desde la raíz hacia las partes aéreas (Kabata-Pendias y Pendias, 2001). Este tipo de plantas, llamadas excluidoras, poseen mecanismos especializados para restringir la absorción de metales por las raíces (Baker, 1981). Sin embargo, el concepto de exclusión de metal no se comprende totalmente (Lasat et al, 2000). Se ha propuesto que la retención de metales en raíces podría producirse debido a algún mecanismo desconocido que evita que un exceso de metales acceda al xilema (Reichman, 2002).

Capítulo I
14
3. Zn en las plantas
3.1. Efectos sobre la fisiología y la morfología vegetal
Mantener la concentración de metales dentro de los rangos fisiológicos requiere la existencia de una buena coordinación entre absorción, transporte y acumulación (Clemens et al., 2002). La respuesta de las plantas a los metales es dependiente de la dosis, vía de entrada y tiempo de contacto con el metal (Broadley et al., 2007). En el caso de los metales esenciales, la curva de respuesta muestra tres regiones bien diferenciadas (Fig. 2): (i) en la primera fase de la curva, el crecimiento incrementa con el incremento del elemento en el medio y, por lo tanto, en la planta (rango de deficiencia), (ii) en la etapa siguiente, el crecimiento alcanza un máximo y permanece independiente del incremento del elemento en el medio (rango óptimo), y (iii) en la última fase de la curva, el crecimiento disminuye con el incremento del metal en el medio (rango de toxicidad) (Marschner, 1995; Reichman, 2002; Hagemeyer, 2004).
Figura 2. Desarrollo vegetal en función de la concentración foliar de Zn
La concentración de Zn en plantas varía considerablemente dependiendo de la especie, del genotipo y del medio. Por ello, los valores de Zn considerados como normales y críticos en la presente tesis, basados en diferentes revisiones bibliográficas, son orientativos (Balsberg Påhlsson, 1989; Kabata Pendias y Pendias, 2001; Hagemeyer, 2004).
A concentraciones foliares óptimas (20-100 µ g-1 por peso seco de hoja), el Zn desempeña un papel metabólico esencial. Las funciones básicas de Zn están

Introducción
15
relacionadas con el mantenimiento de la integridad de membranas, metabolismo de carbohidratos, auxinas y RNA, formación de ribosomas y síntesis proteica (Marschner, 1995; Cakmak, 2000). Además, hay evidencias de que el Zn puede estimular la resistencia de las plantas a la sequía y a altas temperaturas (Gadallah, 2000; Kabata Pendias y Pendias, 2001). La relevancia del Zn como micronutriente se basa en su papel como un componente integral de la estructura de un gran número de enzimas. En estas proteínas, su función puede ser catalítica (por ejemplo, anhidrasa carbónica y carboxipeptidasa) o co-catalítica o estructural (alcohol deshidrogenasa, proteínas que participan en la replicación del ADN) Balsberg Påhlsson, 1989; Marschner, 1995).
Concentraciones de Zn inferiores a las consideradas normales u óptimas inducen en las plantas síntomas de deficiencia, que incluyen clorosis, tamaño de hoja reducido, acortamiento de la longitud de los internudos, inhibición del crecimiento aéreo y radicular, y aumento del daño oxidativo y foto-oxidación. Los problemas de deficiencia de Zn en cultivos se extienden a numerosos países y suponen elevadas pérdidas económicas (Cakmak, 2000; Hagemeyer, 2004; Broadley et al., 2007). Por el contrario, a concentraciones tisulares elevadas, el Zn resulta tóxico para muchas especies (Balsberg Påhlsson, 1989; Adriano, 2001). Niveles altos de este elemento pueden dañar severamente el desarrollo de las plantas. Los efectos de altas concentraciones de Zn incluyen cambios en la morfología, fisiología y bioquímica vegetal (Woolhouse, 1983; Marschner, 1995). Algunos de estos efectos serán evaluados a lo largo de este trabajo, centrándonos en especies leñosas y discutiremos además, sus causas y posibles efectos sobre el funcionamiento general de las plantas y, especialmente, sobre la economía hídrica. Brevemente, altos niveles tisulares de Zn pueden dañar el desarrollo y la elongación radicular (Ruano et al., 1998, Fuentes et al., 2007b), alterar la anatomía del xilema (Robb et al., 1980), inducir alteraciones nutricionales (Marschner, 1995; Monnet et al., 2001), modificar los patrones de acumulación de biomasa (Ruano et al., 1998; Reichman et al., 2001), reducir la tasa fotosintética y la fluorescencia de clorofilas (Van Assche y Clijsters, 1986; Monnet et al., 2001; Vaillant et al., 2005) y disminuir el contenido de pigmentos fotosintéticos (Vaillant et al., 2005). La alteración de la morfología y arquitectura radicular podría comprometer la capacidad de las raíces para explorar el suelo, y capturar agua y nutrientes.

Capítulo I
16
3.2. Sensibilidad de las especies leñosas al Zn
Los estudios sobre efectos del Zn en especies leñosas son relativamente escasos en comparación con los estudios realizados sobre especies agrícolas y herbáceas, aunque en los últimos años ha aumentado notablemente el número de trabajos sobre este tema (Balsberg Påhlsson, 1989; Fuentes et al., 2007a, 2007b; Domínguez et al., 2008). Existen numerosas razones que justifican la necesidad de estudiar la respuesta de especies leñosas a metales pesados.
(i) De acuerdo con Chapin (1980), las especies con crecimiento rápido son más sensibles a los metales pesados que aquellas de crecimiento lento. Esto implicaría que los niveles de toxicidad variarán dependiendo de la estrategia de la planta.
(ii) Las especies leñosas son menos eficaces que las especies herbáceas en cuanto a la capacidad de captar metales pesados. Sin embargo, los árboles podrían ser más eficientes porque tienen una mayor superficie radicular para capturar estos elementos (Pulford y Watson, 2003).
(iii) La longevidad de los árboles dificulta la selección de genotipos tolerantes, los cuales se generan al azar o inducidos por contaminantes (Pulford y Watson, 2003).
(iv) Según Hütermann (2004), la diferencia en la sensibilidad a metales podría ser importante para la regeneración natural y la competencia entre especies coexistentes, pudiendo alterar la composición de las comunidades y el funcionamiento de los ecosistemas.
(v) Más información sobre la respuesta de estas especies a metales pesados permitiría optimizar su uso en la restauración de zonas contaminadas y establecer los riesgos potenciales de la incorporación de estos elementos en la cadena alimenticia (Domínguez et al., 2008).
Resumiendo, la vegetación leñosa es diferente a nivel taxonómico y en cuanto a sus estrategias morfo-funcionales, por lo que los conocimientos obtenidos con especies de interés agrícola sobre los límites de toxicidad, bioacumulación, etc. son difícilmente extrapolables de una manera directa.
Las especies leñosas pueden ser muy sensibles a los metales pesados incluyendo el Zn, particularmente en el estadio de plántula, cuando dependen de las capas superiores de suelo, usualmente las más contaminadas (Adriano, 2001; Reichman

Introducción
17
et al., 2001; Hüttermann et al., 2004). Si bien la mayoría de estudios sobre el efecto de los metales pesados se han centrado en los aspectos bioquímicos y fisiológicos, las características morfo-funcionales podrían ser relevantes en la tolerancia a los metales. Por ejemplo, especies adaptadas a suelos desarrollados sobre serpentinas frecuentemente presentan menor área específica foliar y tamaño de hojas, hojas más gruesas y raíces más profundas que la misma especie creciendo en un suelo no-serpentínico (Wierzbicka y Panufnik, 1998; Brady et al., 2005). Del mismo modo, estas modificaciones podrían afectar a la capacidad de las plantas de enfrentarse a otras fuentes de estrés.
4. Estrés múltiple
4.1. Teoría del estrés: respuesta general vs. respuesta específica
Las plantas tienen un nivel de plasticidad que les permite sobrevivir a un amplio rango de situaciones de estrés presente en el medio natural. Esto implica la evolución de mecanismos que les permiten disminuir el impacto del factor o los factores de estrés sobre el desarrollo y crecimiento vegetal (Lusk et al., 2008; Schmidt, 2008). Algunas de esas respuestas son específicas, y se manifiestan sólo ante un tipo de estrés concreto. Por ejemplo, las enzimas fotoliasas reparan el ADN dañado tras la exposición a radiación ultravioleta B (Britt y Fiscus, 2003) y no se ha descrito otras funciones de las mismas. Las xantofilas, por otra parte, protegen el fotosistema II del exceso de luz y temperatura (Davison, et al., 2002).
Sin embargo, bioquímicos, fisiólogos y ecólogos han sugerido la existencia de mecanismos de respuesta comunes a diversos tipos de estrés (Grime, 1977, Chapin, 1991; Leshem et al., 1998; Potters et al., 2009). Chapin (1991) propuso que las plantas exhiben un sistema centralizado de respuesta que puede ser iniciado por diferentes factores de estrés. Este sistema de respuesta está mediado por hormonas e incluye cambios en los balances nutricional, hídrico, de carbono y hormonal de la planta.
Leshem y Kuiper (1996) y Leshem et al. (1998) aplicaron la hipótesis del Síndrome de Adaptación General, abreviado como SAG, a plantas. Este término fue propuesto originalmente por Selye (1936), quien describió las fases de respuesta al estrés en humanos. En sus trabajos, Leshem y colaboradores discutieron los mecanismos de respuesta comunes a varios tipos de estrés, aunque éstos pudieran

Capítulo I
18
ser totalmente opuestos; por ejemplo, estrés por elevadas temperaturas y la congelación, la desecación y el encharcamiento (Leshem y Kuiper, 1996; Leshem et al., 1998). Estos mecanismos incluyen procesos fisiológicos y morfológicos. Los primeros incluyen síntesis de poliaminas, prolina, ácido abscísico (ABA), proteínas de choque térmico (Hsp en inglés), metil jasmonato y derivados, antioxidantes, óxido nítrico, citocromo P450, chaperona y ubiquitina, osmorregulación, así como cambios en el metabolismo de especies reactivas del oxígeno (abreviadas ROS, por sus siglas en inglés) (Leshem et al., 1998; Potters et al., 2007). El segundo tipo de respuesta general al estrés, incluye cambios morfológicos. Sin embargo, este tipo de respuesta ha recibido relativamente menos atención que la respuesta fisiológica (Potters et al., 2007). Cambios en la pubescencia foliar, en el grosor o curvatura de los tilacoides, en el tamaño y movimiento foliar y en la morfología radicular son ejemplos de este tipo de respuesta (Leshem et al., 1998; Potters et al., 2009).
Potters y colaboradores estudiaron la respuesta morfogénica a varios estreses, concluyendo que esta respuesta comprende: (i) inhibición de la elongación celular, (ii) estimulación localizada de la división celular, y (iii) alteraciones en el estatus de diferenciación celular. Estos cambios ocurrirían de manera coordinada para disminuir los efectos de la exposición al estrés y estarían “mediados” por la acción conjunta de ROS, auxinas y etileno (Potters et al., 2007, 2009).
4.2. Respuesta al estrés múltiple
La teoría de SAG implica la existencia de co-estrés, es decir, la estrategia de una planta frente a un tipo de estrés puede conferir resistencia a otro tipo. Este mecanismo podría resultar beneficioso para las plantas ya que, en la naturaleza, los factores de estrés no suelen actuar de manera aislada, por lo que éstas suelen estar sometidas a una combinación de estreses (Larcher, 2003). En áreas sometidas a sequía, por ejemplo, las plantas deben enfrentarse a menudo a otros factores estresantes como la salinidad o las altas temperaturas (Valladares y Pearcy, 1997).
La respuesta de la planta a la combinación de diferentes factores estresantes no puede predecirse a partir del estudio de los factores individuales. La respuesta a un único factor podría verse influida por la variación de otros factores ambientales. Es decir, la interacción entre distintas fuentes de estrés podría

Introducción
19
reforzar, revertir o enmascarar la respuesta de la planta a un único factor de estrés (Larcher, 2003). Así, la interacción es sinérgica cuando la combinación de estreses induce un efecto de mayor impacto en las plantas que el observado cuando los diferentes tipos de estrés actúan aisladamente. Por ejemplo, el efecto negativo de las altas temperaturas sobre la productividad de plantas de cebada se ve agravado cuando se combina con sequía. El peso del grano se redujo un 20 y un 5% tras la aplicación de forma separada de sequía y alta temperatura, respectivamente. Cuando se combinaron ambos estreses, la reducción en el peso del grano individual fue alrededor del 30% (Savin y Nicolas, 1996).
En otros casos, la aclimatación o adaptación a un tipo de estrés puede conferir protección frente a otro factor simultáneo de estrés. En este caso, la interacción es antagónica. Por ejemplo, durante el estrés por elevadas temperaturas, las plantas abren sus estomas para enfriar sus hojas por transpiración. Sin embargo, si las altas temperaturas se combinan con sequía, la apertura estomática se inhibe y la temperatura de las hojas incrementa (Rizhsky et al., 2002). Las plantas se verían sometidas a una situación similar si son expuestas a la combinación de estrés por metales pesados o salinidad y altas temperaturas. En este caso, el incremento de transpiración podría conducir a una mayor absorción de sales y metales (Mittler, 2006). El ajuste de las plantas a una combinación de diferentes tipos de estrés, por lo tanto, requiere una respuesta apropiada y “personalizada” a cada una de las condiciones de estrés individuales implicadas. Además, esta respuesta debe adaptarse a la necesidad de compensar o ajustar las posibles interacciones antagónicas (Mitler, 2006).
Existen diversas razones que justifican estudiar la respuesta de las plantas a una combinación de factores de estrés. Nilsen y Orcutt (1996) las han resumido en cuatro puntos. (i) El crecimiento de las plantas está regulado por múltiples factores limitantes más que por un único factor, por lo que, para entender la regulación del crecimiento vegetal, es necesario comprender cómo interaccionan los diferentes factores de estrés. (ii) La adaptación genética se produce en el contexto de múltiples factores de estrés; por lo tanto, para entender la adaptación de las plantas es necesario analizar la influencia de la combinación de estos factores. (iii) Las limitaciones y sobre-abundancia de algunos recursos podría restringir la respuesta de la planta a factores de estrés. (iv) Cambios en la disponibilidad de recursos y en la intensidad de los diferentes factores de estrés podrían influir en la capacidad competitiva de especies co-existentes.

Capítulo I
20
Otros motivos para estudiar las interacciones entre múltiples fuentes de estrés se relacionan con cambios antropogénicos de las condiciones ambientales globales. Por un lado, la disponibilidad de recursos naturales necesarios para el establecimiento y desarrollo vegetal se ha visto drásticamente alterada por actividades humanas. Por ejemplo, se ha estimado que la disponibilidad de agua para las plantas disminuirá durante el siglo XXI debido al incremento de evapotranspiración consecuencia del incremento de temperatura global (IPCC 2001). Así mismo, la transferencia de nitrógeno entre la atmósfera y la biosfera se ha duplicado en los últimos 100 años (Gruber y Galloway, 2008). Por otro lado, factores estresantes de origen antropogénico originan nuevos escenarios que deben ser considerados. Concentraciones crecientes de gases potencialmente tóxicos (CO, SO2, O3, etc.) y deposición de metales pesados son algunos de los factores que, en interacción con cambios climáticos y de otra índole, pueden alterar el crecimiento y la ecología vegetal.
4.3. Interacción entre estrés por metales y sequía
Los efectos morfológicos y fisiológicos del Zn sobre las plantas podrían afectar a su capacidad de respuesta a otras fuentes de estrés. Por ejemplo, algunas modificaciones morfológicas y fisiológicas (como alteración del tamaño radicular y cierre estomático) generadas por elevadas concentraciones de Zn, podrían afectar a la capacidad de las plantas de capturar y transportar agua y nutrientes (Barceló y Poschenrieder, 1990; Kasim, 2007). Esto sería especialmente perjudicial en plántulas, el estadio más vulnerable en la regeneración de las plantas leñosas (Villar et al., 2004). Pero la bibliografía sobre este tema es escasa. Más aún, no existe un consenso en la literatura especializada sobre el efecto neto que tiene el Zn sobre la capacidad de las plantas para resistir el estrés hídrico (Balsberg Påhlsson, 1989; Poschenrieder y Barceló, 2004b). Los estudios sobre la interacción entre Zn y disponibilidad hídrica resultan particularmente importantes debido a que la combinación de estos factores podría incrementar en un futuro próximo (IPCC, 2001; Peñuelas y Filella, 2002).

Introducción
21
5. Umbrales de fitotoxicidad. El contexto de la restauración ecológica de sitios contaminados
En la actualidad se emplean diferentes metodologías para la recuperación de un sitio contaminado, aunque algunos de los términos que se utilizan en la bibliografía pueden derivar en diferentes interpretaciones. En este trabajo, se hace referencia a la restauración ecológica, aceptando como tal la definición de la Sociedad de Restauración Ecológica: “proceso de ayudar a la recuperación de un ecosistema que ha sido degradado, dañado o destruido” (Society for Ecological Restoration Science and Policy Working Group 2002). En otras palabras, recuperar las condiciones históricas de un sitio.
Macnair (1987) sugirió que la colonización de áreas como las minas es un reto para las plantas, ya que deben desarrollar tolerancia a niveles tóxicos de metales pesados, así como adaptarse a otras restricciones edáficas, como baja disponibilidad de nutrientes o condiciones de sequía. Por esta razón, se ha sugerido que las plantas que estén pre-adaptadas para las duras condiciones edáficas existentes en las minas podrían colonizar estas regiones más fácilmente (Antonovics et al., 1971; Macnair, 1987). El principio de pre-adaptación es análogo al de tolerancia cruzada.
La restauración de ecosistemas contaminados con metales requiere especial atención. Con el fin de lograr una vegetación sostenible en tierras contaminadas con metales, es esencial la elección de especies que sean capaces de establecerse, crecer, reproducirse y colonizar el sitio. Es decir, las especies utilizadas deben ser tolerantes a los metales presentes y tolerantes a las posibles condiciones desfavorables características de un sitio en particular, como pueden ser la sequía y la falta de nutrientes (Li, 2006). En este sentido, el conocimiento de la tolerancia de las especies nativas a uno o varios metales constituye una clara ventaja a la hora de escoger las especies para la restauración (Wong, 2003).
Los valores umbral de toxicidad de metales para las plantas pueden ser utilizados para estimar la capacidad de una planta para establecerse y sobrevivir en un sitio contaminado (Paschke et al., 2005). Estos valores umbral podrían utilizarse para estimar con mayor precisión los riesgos asociados a la contaminación por metales del suelo y para establecer qué especies son las más apropiadas para la revegetación de un sitio contaminado. Sin embargo, hay escasez de datos sobre los umbrales de toxicidad para especies de plantas nativas y para algunas especies

Capítulo I
22
utilizadas para la restauración de sitios contaminados con metales pesados (Li, 2006; Paschke et al., 2005).
Para la restauración de un sitio, no sólo la evaluación de la tolerancia resulta importante sino también el patrón de acumulación del metal que presenta una determinada especie. Por ejemplo, las plantas que restringen el transporte de metales hacia la parte aérea son una buena opción para la revegetación de un sitio contaminado evitando que los metales alcancen la cadena alimenticia (Wong, 2003).
6. Contexto del trabajo realizado y objetivos
La investigación contenida en esta memoria se enmarca dentro de dos proyectos de investigación realizados en el Departamento de Ecología de la Universidad de Alicante. En primer lugar, el proyecto BIOMON (REN2000-0181-P4-03 Biosólidos en Montes Mediterráneos) fruto de la colaboración de distintas entidades (Fundación CEAM, Departamento de Ecología de la Universidad de Alicante, Universidad de Barcelona y la EPSAR de la Comunidad Valenciana). El objetivo del proyecto fue determinar en qué medida ciertos tipos de biosólidos, que se desechaban para su uso agrícola por el alto contenido de metales pesados, podían ser aprovechados para la fertilización orgánica de repoblaciones forestales de una manera segura y controlada.
En el marco de este proyecto se realizaron diferentes experimentos con el fin de evaluar la sensibilidad de especies leñosas Mediterráneas a metales pesados. En concreto, en el Capítulo II de esta memoria se describe el experimento llevado a cabo para establecer umbrales de fitotoxicidad al Zn en diversas especies Mediterráneas leñosas (Foto 2).
En este trabajo evaluamos la respuesta morfológica de las plántulas a este metal, y discutimos los criterios de selección de especies para su posible uso en restauración ecológica de zonas contaminadas Mediterráneas. Para más información sobre este proyecto, se pueden consultar los trabajos de Fuentes et al. (2007b, 2007c).
En segundo lugar, dentro del proyecto multidisciplinar CREOAK (Conservation and restoration of European cork oak woodlands: a unique ecosystem in the balance QLRT-2001-01594) nos centramos en la interacción entre disponibilidad

Introducción
23
hídrica y niveles de Zn sobre la especie Quercus suber L. Esta especie, además de haber presentado un mayor nivel de tolerancia al metal que otras especies Mediterráneas en el capítulo anterior, es un árbol ampliamente distribuido en la cuenca mediterránea occidental. Los bosques de alcornoques producen bienes y servicios valiosos, como el corcho, y están protegidos por la Unión Europea (Directiva Hábitat 92/43/CEE; Pausas et al., 2009).
Foto 2 Detalle del cultivo hidropónico utilizado en el capítulo II e imagen de las plántulas de Pinus pinea utilizadas.
Así, desarrollamos distintos experimentos que nos permitieron, por un lado, caracterizar la respuesta de esta especie al Zn desde distintos enfoques y, por otro lado, aportar más información sobre la respuesta de las plantas al estrés múltiple. En el Capítulo III analizamos el efecto secuencial de la aplicación de Zn y un período de estrés hídrico breve en Q. suber mediante la respuesta morfo-fisiológica de las plántulas (Foto 3). Posteriormente, llevamos a cabo un experimento donde expusimos plántulas de esta especie a la aplicación simultánea de Zn y niveles de riego contrastados.

Capítulo I
24
Foto 3. Detalle del montaje experimental y de dos sistemas radiculares correspondientes al Capítulo III.

Introducción
25
Foto 4. Plántulas de Quercus suber (arriba) y cámara de cultivo empleada para realizar el experimento del Capítulo III (abajo, izquierda). Vista en detalle de la medición de intercambio de gases (abajo, derecha).
En el Capítulo IV analizamos la respuesta morfo-fisiológica de estas plantas (Foto 4) y en el Anexo I de esta memoria mostramos el análisis metabolómico para evaluar la respuesta de las plántulas a nivel de metaboloma y metabolito. Este trabajo se realizó durante una estancia en el Umeå Plant Science Center en Suecia (Foto 5).
Foto 5. Ejemplo de espectros de masas resultantes del análisis metabolómico (izquierda) e imagen del equipo utilizado (derecha; Web de Umeå Plant Science Center).

Capítulo I
26
Estructura del trabajo y objetivo
Esta tesis doctoral tiene, como objetivo general, evaluar los efectos de la aplicación de Zn sobre especies forestales mediterráneas, así como su interacción con la disponibilidad hídrica. Se ha pretendido alcanzar este objetivo general a partir de los siguientes objetivos parciales:
1. Determinar índices toxicológicos para especies leñosas Mediterráneas importantes para restauración ecológica. Evaluar la relevancia de la adaptación a ambientes áridos en la respuesta al aumento de Zn.
2. Evaluar la respuesta morfológica y fisiológica de plántulas de Quercus suber a concentraciones crecientes de Zn. Determinar si la exposición de plántulas de Q. suber a Zn puede mitigar o incrementar los efectos de un periodo de sequía.
3. Estudiar posibles cambios hormonales y alteración de la conductividad hidráulica en plántulas de Q. suber en respuesta al aumento de Zn. Determinar la existencia de sinergias o antagonismos en la respuesta morfológica y fisiológica de plántulas de Q. suber a la combinación de baja disponibilidad hídrica y aumento de Zn.
4. Analizar el perfil metabólico de plántulas de Q. suber tratadas con Zn y expuestas a niveles de riego contrastados. Determinar respuestas comunes y específicas a los dos factores evaluados.

Sensitivity to zinc of Mediterranean
woody species important for restoration
Sensibilidad a Zn de especies leñosas Mediterráneas importantes para restauración
Karen B. Disante, David Fuentes, Jordi Cortina
Science of the Total Environment 408: 2216-2225 (2010)


Sensitivity to zinc of Mediterranean woody species important for restoration
29
Abstract
Heavy metals have increased in natural woodlands and shrublands over the last several decades as a consequence of anthropogenic activities. However, our knowledge of the effects of these elements on woody species is scarce. In this study, we examined the responses of six Mediterranean woody species to increasing levels of zinc in hydroponic culture and discussed the possible implications for the restoration of contaminated sites. The species used, Pinus pinea L., Pinus pinaster Ait., Pinus halepensis Mill., Tetraclinis articulata (Vahl) Mast., Rhamnus alaternus L. and Quercus suber L. represent a climatic gradient from dry sub-humid to semi-arid conditions. Zinc concentrations in shoots ranged from 53 μg g-1 in Q. suber to 382 μg g-1 in T. articulata and were well below the levels found in roots. Zinc inhibited root elongation and root biomass and changed the root length distribution per diameter class, but the magnitude of the effects was species-specific. Only P. halepensis and Q. suber showed toxicity symptoms in aboveground parts. Species more characteristic from xeric environments (T. articulata, R. alaternus and P. halepensis) were more sensitive to zinc than species from mesic environments (Q. suber, P. pinaster and P. pinea). According to the Zn responses and bioaccumulation, Q. suber P. pinea and P. halepensis are the best candidates for field trials to test the value of woody species to restore contaminated sites. None of the species tested seemed suitable for phytoremediation.
Keywords: toxicity thresholds; ecological restoration; heavy metal; EC50; Zinc translocation.

Capítulo II
30
Resumen
La concentración de metales pesados en bosques naturales y zonas de matorral ha incrementado durante las últimas décadas como consecuencia de las actividades antrópicas. A pesar de ello, nuestro conocimiento de los efectos de estos elementos sobre las especies leñosas es escaso. En este estudio, examinamos la respuesta de seis especies leñosas Mediterráneas a niveles crecientes de Zinc en cultivo hidropónico y discutimos su posible implicación en la restauración de zonas contaminadas. Las especies utilizadas fueron Pinus pinea L., Pinus pinaster Ait., Pinus halepensis Mill., Tetraclinis articulata (Vahl) Mast., Rhamnus alaternus L. y Quercus suber L. que representan un gradiente climático desde seco- subhúmedo hasta semiárido. Las concentraciones de zinc en la biomasa aérea varió entre 53 μg g-1 en Q. suber hasta 382 μg g-1 en T. articulata y se mantuvieron muy por debajo de los valores encontrados en las raíces. La presencia de Zn inhibió la elongación y biomasa radicular y cambió la distribución de longitud radicular por clases diamétricas, aunque la magnitud del efecto dependió de la especie. Únicamente P. halepensis y Q. suber mostraron síntomas de toxicidad en la parte aérea. Las especies más características de ambientes xéricos (T. articulata, R. alaternus y P. halepensis) fueron más sensibles al zinc que especies de ambientes mésicos (Q. suber, P. pinaster y P. pinea). En función de su respuesta al Zn y su bioacumulación Q. suber, P. pinea y P. halepensis son los mejores candidatos para evaluar en campo la validez de especies leñosas en la restauración de zonas contaminadas.
Palabras clave: umbrales de toxicidad; restauración ecológica; metales pesados; EC50; translocación de Zn.

Sensitivity to zinc of Mediterranean woody species important for restoration
31
1. Introduction
Human activities are contributing to a worldwide rise in the concentrations and bioavailability of zinc (Zn) in natural ecosystems (Kabata-Pendias and Pendias, 1992). Zinc is required at low levels (e.g., 20-100 μg g-1 dry weight in leaves) for plant enzymatic activity, maintenance of biomembrane integrity, carbohydrate metabolism and protein synthesis, and it participates in the metabolism of indoleacetic acid (Marschner, 1995; Cakmak, 2000). But at high concentrations (e.g., above 200 μg g-1 d.w. in leaves), Zn has toxic effects on plants (Balsberg Påhlsson, 1989; Adriano, 2001). High levels of this element can impair root development and elongation rate (Reichman et al, 2001; Fuentes et al., 2007b), alter xylem anatomy (Rosolem et al., 2005), unbalance mineral nutrition (Kabata-Pendias and Pendias, 1992), modify biomass accumulation patterns (Menon et al., 2005) and reduce photosynthetic rate and plant growth (Balsberg Påhlsson, 1989).
As compared to crop species, studies on the responses of woody species to Zn are relatively scarce. But woody species can be very sensitive to this element, particularly at the seedling stage when they strongly depend on upper, usually more contaminated, soil layers (Adriano, 2001; Reichman et al., 2001). In addition, information on species response to heavy metals is needed to optimize their use in the restoration of contaminated areas and to assess the potential risks to wildlife upon incorporation of these elements into the food chain (Domínguez et al., 2008).
Metal toxicity thresholds for species used in ecological restoration have great ecological significance, because they can be used to estimate the ability of plants to establish themselves on contaminated sites (Paschke et al., 2000). Studies on the effects of heavy metals on woody species may help to explain current and potential species distributions (Lottermoser et al., 2008) and plant community responses to pollution (Hütermann et al., 2004).
Habitats where heavy metal availability is high are frequently subject to other sources of stress such as low water or nutrient availability (Larcher, 2003; Brady et al., 2005), precluding the identification of specific morpho-functional traits of heavy-metal-tolerant plants. On the other hand, most studies on heavy metal tolerance have focused on biochemical and physiological mechanisms that specifically involve heavy metal uptake, transport and storage (Prasad, 2004). In comparison, morpho-functional strategies to withstand heavy metals have

Capítulo II
32
received much less attention. However, some evidence suggests that plant responses to heavy metals may be similar to those deployed against other sources of stress. For example, plant species adapted to serpentine soils often show lower specific leaf area and leaf size, thicker leaves and deeper roots when grown on serpentine soils than on non-serpentine soils (Wierzbicka and Panufnik, 1998; Brady et al., 2005). These traits have also been related to resistance to drought and extreme temperatures (Larcher, 2003). Furthermore, it has been suggested that plant adaptation to limited water and nutrient availability may contribute to the ability to colonize contaminated areas (Poschenrieder and Barceló, 2004b; Brady et al., 2005).
To study the sensitivity of Mediterranean woody species to heavy metals, we have quantified morphological responses to increasing doses of Zn and identified thresholds in these responses. The species selected cover a climatic gradient from dry sub-humid to semi-arid Mediterranean conditions. Our hypothesis was that morphological traits that increase species resistance to drought may buffer the toxic effects of Zn and, as a result, xeric species may exhibit more resistance to this element than mesic species.
2. Materials and methods
2.1 Plant material
We used seeds of Pinus pinea L. (Stone pine), Pinus pinaster Ait. (Maritime pine), Pinus halepensis Mill. (Aleppo pine), Tetraclinis articulata (Vahl) Mast. (Barbary thuja), Rhamnus alaternus L. (Italian buckthorn) and Quercus suber L (Cork oak). Seeds were provided by the Regional Forest Seed Bank (Banc de Llavors Forestals, Generalitat Valenciana). These materials are collected within the Region of Valencia and comply with local regulations (Conselleria de Medio Ambiente, DOCV 2946).
These species cover a wide climatic gradient, from semi-arid to dry sub-humid conditions, with mean annual precipitation to mean annual potential evapotranspiration ratios of 0.2 – 0.5 and 0.5 – 0.65, respectively (UNEP, 1997). Tetraclinis articulata is a widely distributed tree in N Africa where it covers a total area of 10 000 km2 (Nicolás et al., 2004). Its widespread presence in the

Sensitivity to zinc of Mediterranean woody species important for restoration
33
Iberian Peninsula in pre-historic times has been reduced to one single location. It is more thermophilic and xerophilic than the other species tested and it occurs in semi-arid regions of Spain with average annual precipitation between 150 to 400 mm (Bartolomé et al., 2005). Pinus halepensis covers 25 000 km2 in its native range and, together with T. articulata, is one of the few European tree species capable of withstanding semi-arid conditions. It outcompetes T. articulata on north-facing slopes where they coexist (Nicolás et al., 2004). It has been used extensively for forestation over the last century (Maestre and Cortina, 2004) and grows on a wide range of soils (Richardson, 1998). Rhamnus alaternus is a shrub to small tree common in open areas and in the understory of Quercus ilex L. forests (Masclans, 1981). It is sensitive to freezing and grows on a wide range of soil types, with preference for basic soils. Pinus pinea and Pinus pinaster are common tree species under dry sub-humid and humid Mediterranean conditions. Pinus pinaster covers 13 000 km2 in the Mediterranean basin on various types of soil. Pinus pinea covers 3 200 km2 mostly on sandstone and sandy soils, although it can also be found on dolomite (Richardson, 1998). Quercus suber is found in sub-humid regions of the Mediterranean, with average annual precipitation above 600 mm and average annual temperature near 15 ºC. It commonly grows on neutral to slightly acidic well-drained soils (Serrasolses et al., 2009).
We used minimum annual precipitation as an indicator of species resistance to drought. Extreme climatic events, such as unusually dry years, have intense effects on plant performance and may control species distribution (Easterling et al., 2000). To our knowledge, climate envelopes have not been defined for the species studied. Thus, we estimated the minimum average annual precipitation for each species by using two different approaches. First, we used ANTHOS database (Anthos, 2009), to randomly select 50 locations per species, except for T. articulata as only 4 geographical references were available for this species. Then, we used the New LocClim 1.10 software (FAO Agroclimatic databases, http://www.fao.org/sd/2002/EN1203a_en.htm) to simulate precipitation dynamics and estimate mean minimum precipitation for each location. This software provides a rough estimate of climatic conditions, so we validated these results by comparing the outputs of this exercise with minimum average annual precipitation values obtained from the distribution range of each species as described in Martín et al. (1998) and Bartolomé et al. (2005). This information was not available for R. alaternus. The estimates obtained with both approaches were

Capítulo II
34
highly correlated (r=0.98, F1,3=124, P=0.002), suggesting that they provide reasonable estimates of species climatic constraints.
2.2. Growing conditions
Seeds from local provenances were sown on silica sand and kept at 25 ± 2 ˚C until germination. Seedlings with a mean radicle length of 5 cm and visible aboveground parts were transferred to 1 L pots filled with a nutrient solution containing 170 μM NH4NO3, 42 μM KH2PO4, 88 μM KNO3, 3.1 μM MgSO4·7H2O, 0.27 μM Fe-EDTA, 0.19 μM H3BO3, 0.14 μM Mn-EDTA, 0.078 μM Zn-EDTA, 0.047 μM Cu-EDTA and 0.0004 μM (NH4)6Mo7O24 (as described in Arduini et al., 1994). Pots were kept in a growth chamber under controlled conditions of temperature (23 ± 1 ˚C/ 18 ± 1 ˚C, day/night) and photoperiod (12 h, 300 μmol m-2 s-1 photosynthetic photon flux density during the day). The nutrient solution was constantly aerated and renewed three times a week. Seven days after transplanting, the pots received one of five Zn doses with the nutrient solution: 0.078 (hereafter control), 5, 10, 20 and 90 μM zinc as ZnSO4.
Ion speciation estimates with MINTEQA2 (Allison Geoscience Consultants, Inc. Flowery Branch, Georgia, USA and HydroGeoLogic, Inc. Herndon, Virginia, USA) indicated that Fe and Zn were highly available, as more than 90% of the Fe in all nutrient solutions was present as chelated Fe(III) and 99% of the Zn present was as a free ion (i.e., Zn precipitation and complexation with EDTA was negligible).
We used a randomized block design which included 3 successive blocks in time (starting dates: November 15th 2003 and January 20th 2004 and March 26th 2004). Each block consisted of one pot per zinc dose and species. Each pot contained 4-5 seedlings of the same species that were supported by a perforated polystyrene top.
2.3. Seedling growth and morphology
The second day after seedling transplant, roots were marked 10 mm above the root tip with a permanent marker. Root growth was measured every 2 days and

Sensitivity to zinc of Mediterranean woody species important for restoration
35
relative growth rate (in week-1) calculated as RGR=[ln(L2)-ln(L1)]/[t2-t1], where L1 and L2 correspond to the root length measured on successive dates t1 and t2.
Three weeks after the onset of Zn application, seedlings were harvested, stems cut above the root collar and the rooting system carefully washed. Roots were digitized by scanning on an A3 flatbed scanner (Epson Expression 1680 Pro, Long Beach, California, USA) fitted with a transparency adaptor at 300 dpi, using an 8-bit greyscale. Images were analyzed with specific software (WinRhizo, Regent Instruments, Québec, Canada) to evaluate average root diameter, total length and length per diameter class. We defined 20 uniform diameter classes from 0 to 4 mm. The root length in each diameter class was normalized to the total root length. Finally, all biomass fractions were dried at 65 ºC for 48 h and weighed. Specific root length was estimated as the ratio between total root length and root biomass.
2.4. Root topology
The topology of rooting systems projected onto a two-dimensional plane can be described by three variables: magnitude (µ), total exterior pathlength (Pe) and altitude (a) (Fitter, 1991, see Figure 1, Supplementary material).
Magnitude is the number of root tips. Total exterior pathlength is calculated as the sum of all links (i.e, root segments between successive branchings) counted from each of the root tips back to the base of the rooting system. Altitude is the number of links in the longest unique path from the base link to an exterior link (Fitter, 1991).
Using digitised images and the software WinRhizo, we counted the number of root tips and calculated topological indices for the entire root system of each seedling. Then, we estimated expected altitude (E(a) = 2.65µ0.5) and expected total exterior pathlength (E(Pe) = 2.17µ1.42), that is, the value of a and Pe for a randomly branched rooting system of a given µ, following the method suggested by Werner and Smart (1973), as modified by Berntson (1994). Increases in the ratios a/E(a) and Pe/E(Pe) indicate more herringbone-like root systems, whereas decreases in those ratios correspond to dichotomous branching patterns.

Capítulo II
36
2.5. Zinc concentration in plant tissues
Roots and shoots were digested in a heating block at 250 ºC with a mixture of concentrated sulphuric acid and hydrogen peroxide (Jones and Case, 1990). Digests were analyzed for Zn by ICP-OES (Perkin Elmer Optima 4300 Inductively Coupled Plasma with Optical Emission Spectrometry). Olea europaea leaf standard reference materials (BCR: CRM 062, Commission of the European Communities Bureau of Reference, Brussels) were included in the analyses for quality control. Coefficient of variation ranged from 3.5 to 7.1%.
2.6. Toxicity thresholds
EC is the effective concentration of a toxin that induces a negative effect in the organism under study (Köhl and Lösch, 2004). In our experiment, we estimated EC50-L, EC50-RB and EC50-RGR as the concentration of Zn producing a 50% reduction in root length, root biomass and relative growth rate, respectively. To estimate EC50, average root length, relative growth rate and root biomass per pot were plotted against the external concentration of zinc for each species. The resulting dose-response curves were fitted to the model y = y0 + ae-bx, where y corresponds to the response variable (root length, root biomass and relative growth rate, in cm, mg and week-1, respectively), y0 = asymptotic value of the response variable at increasing Zn concentrations (same units as y), y0+a = value of the response variable when no Zn is available (same units as y), b = the decrease rate in the response variable for a given increase in Zn concentration (in μM-1), and x = Zn concentration (in μM). We tested different models to fit the curves (hyperbolic decay with 2 and 3 parameters, y = ae-bx and y = y0 + ae-bx, respectively, and exponential decay with 2 and 3 parameters, y = ab/(b+x) and y = y0 + ab/(b+x), respectively). The negative exponential with 3 parameters provided the best fit for all species (R2>0.81 and P < 0.05). Finally, we calculated the effective concentration (EC) in (μM) for the three variables (see Figure 2, Supplementary material for an example on EC estimation).

Sensitivity to zinc of Mediterranean woody species important for restoration
37
2.7. Statistical analyses
Seedlings growing in a single container for a given block and Zn dose were not independent, and thus they could not be treated as true replicates. We used an unreplicated randomized block design to analyze our data. Differences in morphological and topological traits between seedlings subjected to various zinc doses were analysed using univariate analysis of variance (ANOVA) with zinc dose and block as fixed factors.
Treatment effects on EC50 indices were evaluated using univariate ANOVA with species and block as fixed factors. Root biomass was transformed using base-10 logarithm prior to analysis to homogenize the variance. When significant differences between treatments were found, means were compared pairwise by using Tukey’s HSD test. All analyses were performed using the SPSS v.15.0 statistical package (SPSS Inc., Chicago, USA).
3. Results
3.1. Zinc bioaccumulation
At the highest application rate, zinc accumulation in shoots ranged from 53 μg g-1 in Q. suber to 382 μg g-1 in T. articulata (Fig. 1). Zinc concentration was, on average, one order of magnitude higher in roots than in shoots. Quercus suber showed the highest contrast between root and shoot concentration of zinc and T. articulata the lowest. The shape of the curve describing the relationship between Zn availability and plant tissue concentration was dependent on the species and the tissue. Rhamnus alaternus and P. pinaster showed a sharp increase in Zn in roots and shoots at low application rates, approaching a saturation level at 5 μM. In contrast, Zn concentration above and belowground in P. pinea and Q. suber increased gradually with increasing application rate. Quercus suber was the only species showing gradual increases in Zn concentration over the whole range of Zn availability tested, with no apparent signs of saturation. This observation, together with the low levels of Zn found in roots suggest that this species was the best Zn excluder of the whole set of species. In contrast to the other species, Zn accumulation followed different patterns for shoots and roots in P. halepensis and T. articulata.

Capítulo II
38
Zinc concentration in nutrient solution (μM)
0 10 20 30 40 50 60 70 80 90 100
Roo
t zn
conc
entra
tion
( μg
g-1)
0
1000
2000
3000
4000
5000
6000
0 10 20 30 40 50 60 70 80 90 100
Shoo
t zn
conc
entra
tion
(μg
g-1)
0
50
100
150
200
250
300
350
400
450
500
0 10 20 30 40 50 60 70 80 90 100
Shoo
t zn
conc
entra
tion
(μg
g-1)
0
50
100
150
200
250
300
350
400
450
500
0 10 20 30 40 50 60 70 80 90 100
Sho
ot z
n co
ncen
tratio
n (μ
g g-1
)
0
50
100
150
200
250
300
350
400
450
500
0 10 20 30 40 50 60 70 80 90 100
Sho
ot z
n co
ncen
tratio
n (μ
g g-1
)
0
50
100
150
200
250
300
350
400
450
500
0 10 20 30 40 50 60 70 80 90 100
Roo
t zn
conc
entra
tion
( μg
g-1)
0
1000
2000
3000
4000
5000
6000
0 10 20 30 40 50 60 70 80 90 100
Roo
t zn
conc
entra
tion
( μg
g-1)
0
1000
2000
3000
4000
5000
6000
0 10 20 30 40 50 60 70 80 90 100
Roo
t zn
conc
entra
tion
( μg
g-1)
0
1000
2000
3000
4000
5000
6000
P. pinea P. pinea
P. pinaster P. pinaster
P. halepensis P. halepensis
T. articulata T. articulata
y = 13.74 x 0.05
F = 14.71 P = 0.031
R2 = 0.83
y = 250.45 +( 3385.48x) / (17.33 + x)
F = 33.84 P = 0.029
R2 = 0.97
y = ( 236.03x) / (0.18 + x)
F = 19.74 P = 0.021
R2 = 0.87
F = 43.74 P = 0.007
R2 = 0.93
y = ( 3537.88x) / (15.19 + x)
y = 53.07 +(139.64 x) / (1.90 + x)
F = 20.26 P = 0.047
R2 = 0.95
y = 166.20 +(3342.56 x) / (5.58 + x)
F = 184.21 P = 0.005
R2 = 0.99
y = 81.77 +(400.50 x) / (30.49 + x)
F = 1002.08 P = 0.001
R2 = 0.99
y = 66.00 +(3342.56 x) / (5.58 + x)
F = 19.87 P = 0.048
R2 = 0.95
A B
C D
E F
G H

Sensitivity to zinc of Mediterranean woody species important for restoration
39
Figure 1. Effects of Zn application on Zn concentration in shoots (A, C, E, G, I, K) and roots (B, D, F, H, J, L) of Pinus pinea (A, B), Pinus pinaster (C, D), Pinus halepensis (E, F), Tetraclinis articulata (G, H), Rhamnus alaternus (I, J) and Quercus suber (K, L). Note the different scales used for root and shoot. Models describing the relationships between Zn concentration and Zn availability are shown.
3.2. Biomass accumulation and root morphology
Foliar biomass accumulation decreased in response to Zn in P. halepensis and Q. suber (Table 1). Roots of all species exposed to Zn doses above 5 μM became brown soon after the onset of Zn application, and root biomass accumulation decreased significantly with increasing Zn dose in P. pinea, P. halepensis, T. articulata, R. alaternus and Q. suber at relatively low Zn levels; in contrast, root biomass accumulation was not affected by Zn in P. pinaster . We found no differences in specific root length (SRL) in T. articulata and Q. suber (Table 1). In contrast, Rhamnus alaternus and all pine species showed a decrease in SRL between 30 and 50% as compared to control seedlings at application rates higher than 5 μM. Relative growth rate (RGR) of the main root was reduced in response
0 10 20 30 40 50 60 70 80 90 100
Roo
t zn
conc
entra
tion
(μg
g-1)
0
1000
2000
3000
4000
5000
6000
Zn concentration in nutrient solution (μM)
0 10 20 30 40 50 60 70 80 90 100
Shoo
t zn
conc
entra
tion
(μg
g-1)
0
50
100
150
200
250
300
350
400
450
5000 10 20 30 40 50 60 70 80 90 100
Shoo
t zn
conc
entra
tion
(μg
g-1)
0
50
100
150
200
250
300
350
400
450
500
0 10 20 30 40 50 60 70 80 90 100
Roo
t zn
conc
entra
tion
(μg
g-1)
0
1000
2000
3000
4000
5000
6000
R. alaternusR. alaternus
Q. suber Q. suber
y = (234.35) x / (0.23 + x)
F = 63.97 P= 0.004
R2 = 0.95
y = (4039.27 x) / (1.46+ x)
F = 41.22 P= 0.008
R2 = 0.93
y = 15.01 x0.26
F = 8.15 P= 0.06
R2 = 0.73
y = 91.02 + (588.50 x) / (168.82 + x)
F = 152.01 P= 0.006
R2 = 0.99
I J
K L

Capítulo II
40
to Zn, and it was virtually zero in all species, at application rates above 10-20 μM Zn (Table 1). At low Zn doses root elongation rate decreased sharply in R. alaternus and T. articulata, whereas the decrease was gradual in the other species. Despite that block had a significant effect on several morphological traits: foliar biomass (P. pinaster, P. halepensis, T. articulata and Q. suber), root biomass (P. pinaster, P. halepensis and Q. suber), SRL (P. pinaster and Q. suber) and RGR (P. pinea), the effect of Zn was similar across all blocks (see Figure 3, Supplementary material for an example on main root RGR).
Root length per diameter class was dependent on species and Zn dose (Fig. 2). Roots in the 0.4-0.6 mm diameter class accounted for more than 40, 50 and 70% of the total root length in control seedlings of P. pinaster, P. halepensis and T. articulata, respectively. A similar pattern was found for R. alaternus, but in this species maximum relative root length corresponded to smaller root diameter classes. Unlike the other species, root length in P. pinea was evenly distributed between four diameter classes ranging from 0.2 to 1 mm. Finally, in Q. suber, root length per diameter class gradually decreased with increasing diameter, and no class showed relative root length accumulation above 25%. Zinc increased the relative abundance of thick roots in P. pinea and, especially, in R. alaternus and P. halepensis. In contrast, T. articulata showed the opposite trend. Finally, root length distribution was not affected by increasing Zn in P. pinaster, and Q. suber. Average root diameter was not significantly affected by Zn but plants growing in consecutive blocks showed differences in average root diameter (P. pinea, P. halepensis and Q. suber) (Fig. 3A, Table 2).
3.3. Root topology
Root system magnitude ranged from 4 to 145 in control seedlings of T. articulata and Q. suber, respectively. This variable decreased significantly in P. pinaster, R. alaternus and Q. suber at application rates above 10-20 μM Zn and was not significantly affected by Zn in the other species tested (Fig. 3B and Appendix 1). Plants growing in consecutive blocks showed differences in and root system magnitude (P. pinaster, R. alaternus and Q. suber, Table 2). All species showed herringbone-like root systems that did not change after exposure to increasing Zn doses (data not shown).

Sensitivity to zinc of Mediterranean woody species important for restoration
41
Table 1. Morphological traits of woody seedlings exposed to increasing doses of Zn in the growing solution. SRL: Specific Root Length. Means, standard deviation of n = 3 blocks and results of an univariate ANOVA are shown. Different letters indicate significant differences at P < 0.05 (Tukey-HSD test). In title row t: treatment; b: block.
Zinc application rate (μM)
0.078 5 10 20 90 Ft4,8 (P)
Fb2,8 (P)
Foliar biomass (mg) Pinus pinea
147±10 158±5 158±28 139±23 142±13 0.72
(0.603) 0.44
(0.657) Pinus pinaster
37±8 40±4 45±17 49±13 41±12 1.55
(0.280) 14.60
(0.002) Pinus halepensis
22±3a 19±4ab 17±5ab 19±6ab 4±6b 5.06 (0.025)
23.62 (<0.001)
Tetraclinis articulata
9±2 10±3 9±1 8±1 8±1 1.02
(0.451) 8.99
(0.009) Rhamnus alaternus
7±2 7±1 5±1 6±0 5±0 2.76 (0.103)
2.33 (0.159)
Quercus suber
249±91ab 231±100ab 297±111a 199±104ab 178±122b 4.17 (0.041)
32.93 (<0.001)
Root biomass(mg) Pinus pinea
45±6a 40±7ab 36±6ab 28±2b 28±6b 6.26
(0.014) 2.06
(0.190) Pinus pinaster
13±3 12±2 11±3 10±4 8±2 3.50 (0.062)
7.12 (0.017)
Pinus halepensis
9±4a 7±2ab 5±2ab 5±2ab 3±1b 6.60 (0.012)
8.23 (0.009)
Tetraclinis articulata
5±1a 3±1ab 2±0ab 2±0b 1±0b 6.89
(0.011) 2.04
(0.192) Rhamnus alaternus
3±1a 2±0ab 2±0ab 1±0b 1±0b 6.69 (0.011)
1.20 (0.350)
Quercus suber
763±245a 545±322bc 682±184ab 541±280bc 385±266c 10.69 (0.003)
58.16 (<0.001)
Specific root length (cm mg-1) Pinus pinea
1.3±0.3a 0.8±0.2b 0.9±0.2b 0.7±0.0b 0.6±0.1b 8.56
(0.005) 1.39
(0.302) Pinus pinaster
2.5±0.4a 2.0±0.2b 1.7±0.3b 1.2±0.2c 1.3±0.2c 49.57 (<0.001)
20.40 (0.001)
Pinus halepensis
2.9±0.4a 2.2±0.2ab 1.4±0.3b 1.6±0.3b 2.0±0.3b 12.11 (0.002)
1.21 (0.346)
Tetraclinis articulata
3.1±0.1 2.5±0.8 2.8±0.7 3.5±0.3 5.1±1.9 2.06
(0.178) 0.076
(0.927) Rhamnus alaternus
5.8±0.7a 4.2±1.0ab 3.0±1.1b 3.8±0.8ab 4.4±0.3ab 4.44 (0.035)
1.19 (0.354)

Capítulo II
42
Zinc application rate (μM)
0.078 5 10 20 90 Ft4,8 (P)
Fb2,8 (P)
Quercus suber
0.2±0.0 0.3±0.0 0.3±0.1 0.2±0.0 0.2±0.0 1.63 (0.258)
5.44 (0.032)
Relative growth rate in main root length (week-1)x10 Pinus pinea
5.3±0.5a 3.5±0.9b 1.7±0.5c 0.5±0.0d 0.1±0.1d 87.73 (<0.001)
5.24 (0.035)
Pinus pinaster
4.9±1.7a 4.0±0.7a 1.0±0.2b 0.3±0.1b 0.2±0.1b 18.69
(<0.001) 0.57
(0.589) Pinus halepensis
4.6±0.5a 2.4±1.1b 0.2±0.1c 0.2±0.1c 0.0±0.0c 36.83 (<0.001)
0.52 (0.615)
Tetraclinis articulata
5.4±0.4a 0.8±0.4b 0.7±0.1b 0.2±0.1b 0.1±0.1b 216.14 (<0.001)
1.97 (0.202)
Rhamnus alaternus
4.0±1.2a 0.6±0.4b 0.2±0.1b 0.1±0.1b 0.2±0.1b 19.19
(<0.001) 0.13
(0.877) Quercus suber
3.0±0.7a 3.2±0.7a 2.0±0.7a 1.7±0.2ab 0.1±0.1b 10.70 (0.003)
0.10 (0.901)
3.4. Toxicity thresholds
We calculated the external Zn concentration that produces a 50% reduction in root biomass accumulation (EC50-RB) for those species and blocks where the response curve leveled off (Table 3). Thus, EC50-RB was not calculated for any block of P. pinea, P. pinaster, and one block of Q. suber and R. alaternus. Toxicity thresholds estimated using the index EC50-RB were higher than those estimated with indices based on root length and relative growth rate. Both EC50-L and EC50-RGR identified R. alaternus and T. articulata as the most sensitive species to Zn and Q. suber as the most tolerant.
We found a positive relationship between EC50-L index and the mean minimum annual precipitation in the geographical distribution range of each species (Fig. 4). Results were similar when mean maximum annual precipitation and EC50-RGR were used (data not shown).

Sensitivity to zinc of Mediterranean woody species important for restoration
43
Q. suber
0.00-0.20
0.20-0.40
0.40-0.60
0.60-0.80
0.80-1.00
1.00-1.20
1.20-1.40
1.40-1.60
1.60-1.80
1.80-2.00
2.00-2.20
2.20-2.40
2.40-2.60
2.60-2.80
2.80-3.00
3.00-3.20
3.20-3.40
3.40-3.60
3.60-3.80
Roo
t len
gth
(%)
0
10
20
30
40
50
60
70
80
0.078 μM5 μM10 μM20 μM90 μM
R. alaternus
0.00-0.20
0.20-0.40
0.40-0.60
0.60-0.80
0.80-1.00
1.00-1.20
1.20-1.40
1.40-1.60
1.60-1.80
1.80-2.00
2.00-2.20
2.20-2.40
2.40-2.60
Roo
t len
gth
(%)
0
10
20
30
40
50
60
70
80
0.078 μM5 μM10 μM20 μM90 μΜ
T. articulata
0.00-0.200.20-0.40
0.40-0.600.60-0.80
0.80-1.001.00-1.20
1.20-1.401.40-1.60
1.60-1.801.80-2.00
2.00-2.202.20-2.40
2.40-2.60
Roo
t len
gth
(%)
0
10
20
30
40
50
60
70
80
0.078 μM5 μM10 μM20 μM90 μM
P. halepensis
0.00-0.20
0.20-0.40
0.40-0.60
0.60-0.80
0.80-1.00
1.00-1.20
1.20-1.40
1.40-1.60
1.60-1.80
1.80-2.00
2.00-2.20
2.20-2.40
2.40-2.60
Roo
t len
gth
(%)
0
10
20
30
40
50
60
70
80
0.078 μM5 μM10 μM20 μM90 μM
P. pinea
0.00-0.20
0.20-0.40
0.40-0.60
0.60-0.80
0.80-1.00
1.00-1.20
1.20-1.40
1.40-1.60
1.60-1.80
1.80-2.00
2.00-2.20
2.20-2.40
2.40-2.60
Roo
t len
gth
(%)
0
10
20
30
40
50
60
70
80
P. pinaster
0.00-0.20
0.20-0.40
0.40-0.60
0.60-0.80
0.80-1.00
1.00-1.20
1.20-1.40
1.40-1.60
1.60-1.80
1.80-2.00
2.00-2.20
2.20-2.40
2.40-2.60
Roo
t len
gth
(%)
0
10
20
30
40
50
60
70
80
0.078 μM5 μM10 μM20 μM90 μM
A
Diameter class (mm)
aababbb
0.078 μM5 μM10 μM20 μM90 μM
E
B
C
aabababb
D
abbbb
a
aa
ab
b
a
bbbc
c
F
a
b
bcbcc
aabb
bb
Figure 2. Root length distribution per diameter class. Means ± 0.5 standard deviation of n= 3 blocks are shown. Different letters indicate significant differences at P < 0.05 (Tukey-HSD test). Note the different X-scale used for Q. suber.

Capítulo II
44
Table 2. Results of an univariate ANOVA for average root diameter and root system magnitude. Different letters indicate significant differences at P < 0.05 (Tukey-HSD test). In title row t: treatment; b: block.
Ft4,8 Pt Fb2,8 Pb
Average root diameter
Pinus pinea 2.53 0.123 11.89 0.004
Pinus pinaster 0.60 0.693 1.68 0.245
Pinus halepensis 1.79 0.224 5.75 0.028
Tetraclinis articulata 2.97 0.099 1.903 0.218
Rhamnus alaternus 2.42 0.134 0.835 0.468
Quercus suber 2.08 0.175 15.22 0.002
Root system magnitude
Pinus pinea 1.82 0.218 1.40 0.300
Pinus pinaster 4.93 0.027 30.38 <0.001
Pinus halepensis 3.71 0.058 3.17 0.097
Tetraclinis articulata 1.02 0.458 0.915 0.481
Rhamnus alaternus 5.30 0.022 13.69 0.003
Quercus suber 7.03 0.010 28.87 <0.001
4. Discussion
4.1. Zinc uptake and bioaccumulation
Zinc accumulation in shoots and roots increased with increasing Zn concentration in the growing medium. However, Zn was strongly retained belowground, as the concentrations of this element were substantially higher in roots than in shoots. High Zn accumulation in roots has been observed in other studies (Paschke et al., 2000; Reichman et al., 2001; Fuentes et al., 2007b), and may result from metal binding to negatively charged binding sites in the cell wall (Greger, 2004) and restricted transport to shoots (Utriainen et al., 1997; Pulford and Watson, 2003).
Maximum Zn concentrations belowground were observed in P. halepensis and R. alaternus (4358 and 4559 µg g-1, respectively) and were similar to those found in

Sensitivity to zinc of Mediterranean woody species important for restoration
45
woody species grown in hydroponic culture (e.g., 3444 µg g-1 to 5700 µg g-1; Reichman et al., 2001; Fuentes et al., 2007b). Zinc concentrations in roots showed a non-linear response to Zn availability, reaching an asymptote within the range of Zn concentrations used in this study in all species but Q. suber. This type of curve has been described in the literature (Reichman et al., 2001; Greger, 2004) and may reflect the saturation of metal uptake (Nyquist and Greger, 2007). Indeed, Brunner et al. (2008) found that Zn moves from the cell walls into the lumen after reaching an equilibrium or saturation. When metal concentration is low, ionic competition may be negligible, and uptake efficiency may be high. But, as the Zn concentration in the growing medium increases, competition among Zn ions may follow, thus decreasing uptake efficiency (Greger, 2004). In addition, kinetic studies have found gradual decreases in metal absorption efficiency owing to induced saturation of metal transporters (Hart et al., 1998). Finally, the direct toxic effect of metal may contribute to hinder its uptake (Nyquist and Greger, 2007). Quercus suber was the only species showing an almost linear increase in belowground Zn concentrations over the range of Zn doses tested. In addition, maximum Zn concentrations in roots of Q. suber (2141µg g-1) were relatively low compared to
0 10 20 30 40 50 60 70 80 90 100A
vera
ge d
iam
eter
(mm
)
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
2,0
P. pineaP. pinasterP. halepensisT. articulataR. alaternusQ. suber
A
Zinc application rate (mM)
0 10 20 30 40 50 60 70 80 90 100
Mag
nitu
de
0
30
60
90
120
150
180
210
240
270
300
P. pineaP. pinaster P. halepensisT. articulataR. alaternusQ. suber
B
abab
a
b
b
ab a aab
ba abb ab b
Figure 3. Average root diameter (A) and number of root tips (magnitude) of the rooting system (B) of seedlings of six Mediterranean woody species grown under different Zn application rates. Symbols and bars represent means ± 0.5 standard deviation of n = 3 blocks. Different letters indicate significant differences at P < 0.05 (Tukey-HSD test).

Capítulo II
46
the other species tested and other studies using hydroponics. For example, Reichman et al. (2001) found that Zn concentration in roots of Eucalyptus camaldulensis and Acacia holosericea ranged from 4000 µg g-1 to 6000 µg g-1 when external Zn concentration was 100 µM. Four weeks after exposing Salix fragilis to 76.5 μM of Zn, the Zn concentration in roots was 3370 µg g-1 (Dos Santos Utmazian et al., 2007). Thus, results suggest that at the early seedling stage, Q. suber may behave as a powerful Zn excluder.
Table 3. Toxicity threshold for seedlings of six Mediterranean woody species subjected to increasing Zn concentration in the nutrient solution. Dose-response curves were fitted to the model: y = y0 + ae-bx. EC50-RB, EC50-length and EC50-RGR were calculated as the Zn concentration in the growing media resulting in a 50% reduction in seedling root biomass, root length and relative root grow rate, respectively. Means, standard deviation of n = 3 blocks and results of univariate ANOVA are shown. Different letters indicate significant differences between species at P < 0.05 (Tukey-HSD test).
Species EC50-RB (μM) EC50-L (μM) EC50-RGR (μM)
Pinus pinea >90 10.0 ± 4.1ab 6.3 ± 1.7a
Pinus pinaster >90 13.6 ± 3.6ab 5.3 ± 0.2a
Pinus halepensis 36.3 ± 32.5 4.2 ± 2.1a 4.3 ± 1.2a
Tetraclinis articulata 11.8 ± 3.0 4.0 ± 3.2a 1.8 ± 1.0a
Rhamnus alaternus 15.7 ± 12.1 3.7 ± 1.1a 1.8 ± 0.7a
Quercus suber 30.0 ± 16.5 19.8 ± 9.7b 19.2 ± 7.0b
Ft F3,4 = 0.76 F5,10 = 5.05 F5,10 = 13.45
P 0.573 0.014 <0.001
Fb F2,4 = 0.94 F2,10 = 0.56 F2,10 = 1.52
P 0.463 0.586 0.265
Zinc concentrations in shoots remained relatively constant for Zn concentrations above 5 μM in all species but T. articulata. The range of maximum Zn translocation aboveground was relatively narrow, ranging from 5% in Q. suber to 20% in P. pinea, P. pinaster, P. halepensis and R. alaternus. Weak increases in Zn concentrations in shoots in response to an increased external concentration of Zn have been attributed to controlled Zn transport, i.e., regulation of metal

Sensitivity to zinc of Mediterranean woody species important for restoration
47
translocation and they may reflect the outcome of a suite of mechanisms allowing tolerance to this heavy metal (Denny and Wilkins, 1987; Utriainen et al., 1997). Tetraclinis articulata differed from the others species in that Zn concentrations in shoots reached no asymptote with increasing Zn dose. This may reflect uncontrolled or unrestricted Zn transport, as has been observed in other species, and may result in severe toxicity and death (Denny and Wilkins, 1987).
Minimun annual precipitation (mm)0 50 100 150 200 250 300 350 400 450 500 550 600
EC50
-L (μ
M z
inc)
0
5
10
15
20
25
30
T. articulata P. halepensis
P. pinea
Q. suber
P. pinaster
R. alaternus
Figure 4. Relationship between the EC50-L and mean minimum annual precipitation within the geographical distribution of 6 woody Mediterranean species. Bars represent 0.5 standard deviations of n = 3 blocks (for EC50-L data), and 50 replicated sites chosen randomly from the Anthos database (for rainfall data), except for Tetraclinis articulata, where only 4 populations are available in Spain. EC50-L (in μM) was estimated as the zinc concentration resulting in a 50% reduction in seedling root length.
Toxicity limits for Zn depend on plant species, genotype and growth stage (Kabata- Pendias and Pendias, 1992). Most species may be sensitive to Zn when its concentration in aboveground parts ranges between 200 and 300 μg g-1 (Balsberg Påhlsson, 1989). In our study, three species (T. articulata, P. pinaster and R. alaternus) reached Zn concentration values in leaves and needles that can be considered critical (382, 235 and 217 μg g-1, respectively). Nevertheless, none of the six species studied showed visual symptoms of Zn toxicity, such as chlorosis or necrosis.

Capítulo II
48
4.2. Biomass accumulation and root morphology
Shoot growth was not sensitive to Zn in most species, save for P. halepensis and Q. suber. This finding contrasts with longer-term studies showing decreases in aboveground biomass accumulation in response to this element. But we may note that our experiment lasted for only three weeks and thus it was shorter than other experiments reported in the literature (Balsberg Påhlsson, 1989; Reichman et al., 2001; Fuentes et al., 2007b). Root growth was more sensitive to Zn than shoot growth, as observed in other studies (Paschke et al., 2000). Elongation of the main root was completely arrested at the highest Zn doses in all species. Inhibition of root elongation is a common early indicator of heavy metal toxicity, which may be a result of impaired cell elongation (Woolhouse, 1983). This may also have favoured the increase in the proportion of roots of higher diameter observed in P. pinea, P. halepensis and R. alaternus, a phenomenon described elsewhere (Zobel et al., 2007). Changes in root growth patterns may impair the capacity of seedlings to explore the soil for water and nutrients, as fast root elongation after germination is a key factor in the survival and early growth of Mediterranean seedlings (Padilla and Pugnaire, 2007).
Seedlings from different blocks differed in various morphological and topological variables for some species. The causes of such variability are difficult to unveil. Seedlings in different blocks germinated in different times and may have experimented slightly different growing conditions, despite that they were grown in a controlled chamber. Unfortunately, it is not possible to analyse the interaction between Treatment and Block in an unreplicated randomized block design.
The control seedlings showed a different pattern of root length distribution per diameter class. In P. pinea and Q. suber the proportions of thick roots were higher than in the other species. For example, roots thicker than 2 mm represented 15% of the total root length in Q. suber. According to our results, Zn can increase, decrease or have no significant effect on root length distribution per diameter class depending on the species. Changes in root length distribution per diameter class have not been evaluated in studies on heavy metal toxicity. Our results emphasize the importance of evaluating this trait, as species that did not differ in their sensitivity to heavy metal availability in terms of root and shoot biomass, and other commonly measured response variables did, however, show strongly

Sensitivity to zinc of Mediterranean woody species important for restoration
49
contrasted responses in root abundance per diameter class. Changes in root length per diameter class may affect seedling performance because this trait is related to root longevity, carbon balances and seedling ability to capture and transport soil resources (Pregitzer et al., 1998; Eissenstat et al., 2000).
Other morphological changes in response to heavy metals such as decreased diameter of xylem elements and root elongation may impair species ability to withstand drought (Barceló and Poschenrieder, 1990). In turn, adaptive traits to withstand drought could affect seedling resistance to heavy metal toxicity. Some authors have pointed out the importance of pre-adaptation to drought for colonizing metal-rich soils (Poschenrieder and Barceló, 2004b). However, to our knowledge this topic has received little attention in the literature.
4.3. Toxicity thresholds
Under the experimental conditions of the current study, EC50-RB was not suitable as a predictor of Zn toxicity, as this value could not be estimated for all species. In addition, Zn may induce root suberization and lignification (Barceló and Poschenrieder, 1990), eventually outbalancing the changes in root biomass resulting from lower growth rates. Increased suberization would lead to over-estimations of EC50-RB. Both EC50-L and EC50-RGR showed similar results and may be recommended as important metrics to assess toxicity when the response of entire rooting system to Zn does not differ from that of the main root, as was the case in the current experiment. In those cases where Zn may have contrasted effects on different parts of the rooting system, we would recommend the use of EC50-L as it provides an evaluation of the whole rooting system. In addition, EC50-L and EC50-RGR do not reflect changes in root diameter and root tissue density.
The most sensitive species to zinc, T. articulata, differed from the most resistant, Q. suber, in several trait responses: higher Zn accumulation and translocation, sharp decrease in root elongation rate at relatively low doses of Zn and increased root length in small diameter roots. In addition, by the end of the experiment, T. articulata seedlings were substantially smaller than Q. suber seedlings. Thus, effective concentrations, such as those provided by the indices EC50-L and EC50-RGR, suggest that T. articulata, R. alaternus and P. halepensis, species with lower requirements of rainfall, are more sensitive to Zn than species from more mesic

Capítulo II
50
areas, such as P. pinea, P. pinaster and Q. suber. We note that our results are based on a small set of species, and we did not control for phylogenetic factors, seed size or seed provenance. But the strength of the relationship suggests further studies on this topic are needed.
4.4. Zinc toxicity and ecological restoration
The use of plants to remove, destroy or sequester heavy metals has gained considerable attention in the last decades. Revegetation of contaminated sites must be based on the selection of appropriate species both to prevent the risk of heavy metals entering the food chain and to optimise seedling survival and growth (Pulford and Watson, 2003). In this context, target species should be metal-tolerant and restrict metal uptake and translocation aboveground, particularly to edible parts. Under the experimental conditions of the present study, Q. suber and P. pinea can be recommended for restoring plant cover in contaminated sites in mesic areas. It is worth noting that these species can thrive in moderately acidic soils (Richardson, 1998; Serrasolses et al., 2009), but may contribute to decrease soil pH (Richardson, 1998), thereby increasing the mobilization of heavy metals. This must be monitored because increasing leaching could increase the heavy metal content in groundwater (Domínguez et al., 2008; Tack and Vandecasteele, 2008). In more xeric environments, P. halepensis may be the best choice, owing to the low aboveground Zn accumulation observed. In contrast, T. articulata was very sensitive to Zn and accumulated substantial quantities of this element aboveground increasing the risk of entering Zn the food chain. Thus, this species may be unsuitable for the restoration of contaminated sites. On the other hand, plants may be used for harvesting metals from contaminated sites (i.e., phytoremediation). In this case, high aboveground uptake is desired. Pinus pinaster in mesic areas and T. articulata in xeric areas would be the two species selected among the ones tested in the current study. However, phytoaccumulation ranged from 0.08 to 0.09% of dry weight in T. articulata and P. pinaster, respectively. These concentrations were low compared with 1%, the threshold for defining a plant as Zn hyperaccumulator (Prasad, 2004).

Sensitivity to zinc of Mediterranean woody species important for restoration
51
5. Conclusions
The morpho-physiological responses of Mediterranean woody species to Zn were strongly species-specific. Quercus suber was the more resistant species, probably due to its ability to exclude Zn at the early seedlings stage, whereas Tetraclinis articulata was the most sensitive. Species showing higher Zn uptake and translocation rates also showed stronger morphological responses to this heavy metal. Surprisingly, species resistance to Zn was positively related to minimum annual precipitation of the species habitats providing a compelling stimulus for future research.
Of the species tested, Quercus suber and Pinus pinea in mesic environments and Pinus halepensis in xeric environments are the best suited to withstand Zn, and can be recommended for further field trials; but none of the species tested showed a strong potential for phytoremediation.
Acknowledgements
We thank Juanjo Torrecillas, Santiago Soliveres and Jose Huesca for their assistance in the lab, Banc de Llavors Forestals (Generalitat Valenciana) for the seeds used in this experiment and the University of Alicante for a FPU (Formación del Profesorado Universitario) grant awarded to K. Disante. This research was funded by the Spanish Ministry of Science and Innovation (Projects BIOMON, REN REN2000-0181P4-03 and GRACCIE, Programa Consolider-Ingenio 2010). CEAM (Centro de Estudios Ambientales del Mediterráneo) is funded by Generalitat Valenciana and Fundación Bancaja.

Capítulo II
52
Appendix 1
Example of scanned images of rooting systems of Pinus halepensis (A, B), Pinus pinaster (C, D) and Rhamnus alaternus (E, F). Images A, C and E correspond to control seedlings. Seedlings B, D and F were exposed to a nutrient solution containing 90 μM Zn. Vertical bars are 1 cm long.

Response to drought of Zn-stressed Quercus suber L. seedlings
Respuesta a la sequía de plántulas de Quercus suber L. expuestas a Zn
Karen B. Disante, David Fuentes, Jordi Cortina
Environmental and Experimental Botany 70: 96-103 (2011)


Response to drought of Zn-stressed Quercus suber L. seedlings
55
Abstract
Zinc is an essential nutrient for higher plants but it becomes toxic as its availability increases. In nature, different stress factors commonly occur concurrently, challenging our ability to predict their impacts. Information on zinc (Zn) effect on plant ability to withstand other sources of stress is scarce. This study examines the effect of zinc supply rate on the response of Quercus suber L. seedlings to water stress. Seedlings were treated with four levels of zinc from 3 to 150 μM, and then exposed to a short severe drought. Zinc concentration in leaves and roots increased with zinc availability. Maximum photosynthetic rate, photochemical efficiency, root length and specific root length decreased as Zn availability increased. The decrease was particularly intense between 50 and 150 µM Zn. The relative effects of drought were less intense in seedlings receiving higher doses of Zn than in those receiving 3 μM. Thus, at severe drought, relative water content of detached leaves decreased to 52% in seedlings receiving the lowest level of Zn. In contrast, relative water content remained above 70% in seedlings exposed to high concentrations of this metal. The pattern was similar for photosynthetic rate and stomatal conductance, as the decrease in these variables resulting from severe drought was 100% and 90% in seedlings receiving 3 μM and 65% and 56% in seedlings receiving the highest Zn dose. Our results suggest that morpho-physiological responses to zinc may foster water conservation strategies and alleviate the effects of drought in the short-term, but they may impair seedling ability to root and grow in a longer term.
Keywords: heavy metal, growth response, water use efficiency, relative water content

Capítulo III
56
Resumen
El zinc es un nutriente esencial para las plantas superiores, pero puede resultar tóxico si su disponibilidad aumenta. En la naturaleza existen distintos factores de estrés que actúan simultáneamente lo que pone a prueba nuestra habilidad para predecir sus impactos. La información del efecto de Zn sobre la capacidad de las plantas para enfrentarse a otros factores de estrés es escasa. Este estudio examina el efecto de distintas dosis de Zn en plántulas de Quercus suber L. y su respuesta al estrés hídrico. Tras su exposición a cuatro niveles de Zn, desde 3 hasta 150 μM, las plántulas fueron sometidas a un periodo corto de sequía intensa. La concentración de Zn en hojas y raíces incrementó con la disponibilidad de este metal. La tasa fotosintética máxima, la eficiencia fotoquímica, la longitud radicular y la longitud específica radicular disminuyeron a medida que incrementó la disponibilidad de Zn. El descenso fue particularmente intenso entre 50 y 150 μM de Zn. Comparativamente, los efectos de la sequía fueron menos severos en las plántulas que recibieron mayores dosis de Zn en relación con las que recibieron 3 μM. De este modo, durante la sequía intensa, el contenido hídrico relativo de las hojas decreció hasta el 52% en plántulas que recibieron los menores niveles de Zn mientras que este valor se mantuvo por encima del 70% en las plántulas expuestas a la concentración más alta del metal. El comportamiento de la tasa fotosinténtica y la conductancia estomática fue similar, ya que el descenso en estas variables tras la sequía fue respectivamente, de 100% y 90% en las plántulas que recibieron 3 μM y de 65% y 56% en las que recibieron la mayor dosis de Zn. Nuestros resultados sugieren que las respuestas morfo-fisiológicas al Zn podrían promover estrategias de conservación hídrica y aliviar los efectos de la sequía a corto plazo, pero a largo plazo, podrían perjudicar el crecimiento radicular y el desarrollo de las plántulas.
Palabras clave: metal pesado, crecimiento vegetal, eficiencia en el uso del agua, contenido hídrico relativo.

Response to drought of Zn-stressed Quercus suber L. seedlings
57
1. Introduction
Zinc (Zn) concentration in soils has gradually increased in the last decades as a consequence of human activities (Alloway, 1995). Anthropogenic sources include industrial processes, agriculture and metal mining (Escrig and Morell, 1997; Kabata-Pendias and Pendias, 1992). The increasing use of biosolids has further contributed to Zn incorporation in soils (Smith, 1996; Fuentes et al., 2007a).
Zinc is a natural constituent of soils and an essential micronutrient for humans, animals, and higher plants. In plants, Zn modulates the activity of a large number of enzymes, is involved in the maintenance of biomembrane integrity, participates in carbohydrate metabolism and protein synthesis, and plays an important role in the metabolism of indoleacetic acid (Balsberg Påhlsson, 1989; Marschner, 1995). Further, Zn protects cells from damage from reactive oxygen species (Cakmak, 2000).
Beyond certain concentrations, Zn is toxic to vascular plants (Woolhouse, 1983; Balsberg Påhlsson, 1989). Zinc interferes with photosynthesis, mineral nutrition and root development (Monet et al., 2001; Vaillant et al., 2005; Fuentes et al., 2007b). Symptoms of Zn toxicity and their intensity depend on plant species, genotypes and development stage (Kabata-Pendias and Pendias, 1992).
Woody species can be very sensitive to the presence of Zn (Reichman et al., 2001; Fuentes et al., 2007b). Zn concentration in leaves usually ranges between 15-100 μg g-1 (Hagemeyer, 2004). Zn concentrations above 200 μg g-1 may impair plant performance (Balsberg Påhlsson, 1989). Zinc may induce changes in their morphology and physiology, including the alteration of the root system (Disante et al., 2010), modifications in xylem anatomy (Kasim, 2007; Barceló and Poschenrieder, 1990) decreases in the supply of essential nutrients (Balsberg Påhlsson, 1989; Kabata-Pendias and Pendias, 1992), and reductions in photosynthetic rate and plant growth (Menon et al., 2005; Di Baccio et al., 2009).
Morphological and physiological effects of Zn may affect plant capacity to capture and transport water (Barceló and Poschenrieder, 1990; Kasim, 2007). But information on this topic is scarce. Indeed, there is no agreement on whether high Zn concentrations in soil solution increase or decrease plant resistance to water stress (Balsberg Påhlsson, 1989; Poschenrieder and Barceló, 2004b). On the one hand, Zn availability may promote water conservation by means of decreased

Capítulo III
58
transpiration rate and increased root lignification (Barceló and Poschenrieder, 1990; Päivöke, 2003). In addition, Zn may stimulate the accumulation of osmotically active solutes facilitating water uptake (Gadallah, 2000). On the other hand, arrested root growth induced by high Zn levels may limit seedling capacity to acquire water and nutrients (Poschenrieder and Barceló, 2004b; Kasim, 2007). The lack of information on the interaction between heavy metals and drought is particularly upsetting as the combination of both stress factors may be more widespread in the future (IPCC, 2001; Peñuelas and Filella, 2002).
We carried out a study to evaluate the effect of previous exposition to Zn on the response of seedlings of a common eastern Mediterranean tree species, Quercus suber L., to low water availability. Quercus suber is a calcifuge species (Serrasolses et al., 2009) which is sensitive to high Zn availability, although it may be more resistant to this heavy metal than other woody Mediterranean species (Disante et al., 2010). Following our previous discussion on the effects of Zn on plant water economy, we hypothesized that Q. suber ability to withstand drought would show a unimodal response to Zn availability, small supply rates increasing tolerance to low water availability and high Zn availability in soil solution intensifying the negative effect of water stress. Our study provides new insights on the potential indirect effects of Zn pollution on plant performance in areas subjected to periodic drought.
2. Material and methods
We grew Q. suber seedlings at four levels of Zn availability for 2 months, characterized seedling morphology and physiology at the end of this period, and finally evaluated their response to drought. Quercus suber is an evergreen tree widely distributed in the eastern Mediterranean basin, and it has great socio-economic and cultural value (Pausas et al., 2009). Quercus suber forests are protected by the European Union (Habitat Directive 1992). They may experiment high zinc availability after wildfire (Pereira and Úbeda, 2010). In addition, Q. suber has been recently exposed to zinc as a consequence of human activities such as mining (Grimalt et al., 1999) and biosolids application (Nèble et al., 2007).

Response to drought of Zn-stressed Quercus suber L. seedlings
59
2.1. Growing conditions and treatments
We used Q. suber acorns provided by the Regional Forest Seed Bank (Banc de Llavors Forestals, Generalitat Valenciana). Acorns were sown in silica sand at 25 ± 2˚C until germination in a growing chamber. After 2 weeks, when the radicle was approximately 2 cm long, acorns were transplanted into 5 cm diameter and 40 cm length PVC tubes filled with silica sand. Tubes were placed in a growing chamber at 22 ± 2˚C, 50% relative humidity and 12 h photoperiod (600 μmol m-2 s-
1 of photosynthetic photon flux density).
Growing medium was irrigated to saturation three times per week with a nutrient solution containing 500 μM KCl, 435 μM NaNO3, 300 μM MgSO4·7H2O, 45 μM NaH2PO4, 300 μM CaCl2, 0.37 μM FeSO4·7H2O, 0.012 μM CuSO4·5H2O, 1.6 μM H3BO3, 0.006 μM KI, 0.6 μM MnSO4·7H2O and 0.0004 μM (NH4)6Mo7O24 (modified from Arduini et al., 1994). We added ZnSO4 to the nutrient solution to achieve a Zn concentration of 3 (Control), 10, 50 and 150 μM corresponding to those commonly found in the soil solution of uncontaminated and contaminated soils (Kabata-Pendias and Pendias, 1992). The pH of the nutrient solution was adjusted to 6.0 to mimic natural conditions in Q. suber forests (Serrasolses et al., 2009).
Each Zn supply rate was replicated 10 times. Ion speciation calculations for solutions containing different amounts of Zn were conducted using MINTEQA2 software (Allison Geoscience Consultants, Inc. and HydroGeoLogic, Inc.). Calculations showed that more than 91% of Fe was present as Fe (III) chelators and 99% of Zn as free ion in all treatments, suggesting that Zn precipitation and complexation was negligible.
2.2. Seedling characterization
We measured photosynthesis response to decreasing light using a Li-Cor 6400 portable infrared gas analyzer system (Li-Cor Inc. Nebraska, US) on N= 4 - 5 seedlings two months after the onset of the experiment. Photosynthetic photon flux density was adjusted from 1200 to 0 μmol m-2 s-1 in seven steps. The maximum photosynthetic rate (Am) and maximum apparent quantum yield (Φm) at decreasing light were estimated from the light response curve of each seedling

Capítulo III
60
by adjusting the data to a non-rectangular hyperbola (Photosyn Assistant 1.1 software. Dundee Scientific, Dundee, UK).
After that, we measured changes in photochemical efficiency at decreasing levels of leaf hydration. One leaf per seedling and treatment was fully hydrated in distilled water (24 h, 4ºC in darkness) and allowed to dry under the same conditions. We weighed the leaves every 30 minutes, calculated the relative water content (RWC) and measured chlorophyll fluorescence of detached leaves (Vilagrosa et al., 2003a). The RWC was calculated as RWC= (FW-DW)/(TW-DW) x 100, where FW is the fresh weight measured every 30 minutes, DW is the dry weight (at 65ºC for 48h) and TW is leaf weight at full turgor (24h, 4ºC in darkness).
Chlorophyll fluorescence was measured by means of a portable fluorometer (pulse-amplitude modulated photosynthesis yield analyzer, PAM-2001 Walz, Effeltrich, Germany). We measured minimum fluorescence yield (F0) under weak red light and maximum fluorescence (Fm) after a saturating pulse of white light. Maximal PSII photochemical efficiency (Fv:Fm) was calculated as Fv:Fm= (Fm - F0)/ Fm (Genty et al., 1989). Maximal PSII photochemical efficiency and RWC data for each leaf was then fitted to a sigmoid curve (Fv:Fm = a/1+exp-((RWC - RWC0) / b)), where a, b and RWC0 are parameters. Adjustment between this model and observed data was highly significant in all cases (R2 > 0.96, P<0.0014). From every curve, we analyzed the value of Fv:Fm at three cell hydration levels (100%, 75% and 60% RWC), which correspond to maximum turgor, and approximate values of onset of the reduction of electron transport rate, and permanent photoinhibition, respectively (Medrano and Flexas, 2004).
Root systems were carefully washed and biomass fractions separated. Leaves and roots were digitized by scanning on an A3 flatbed scanner (HP Deskscan, HP Co., Palo Alto, USA) fitted with a transparency adaptor at 300 dpi, using an 8-bit greyscale. The images were analyzed with specific software (WinRhizo, Regent Instruments, Québec, Canada) to evaluate projected foliar surface, total root surface and root length. All biomass fractions were then dried at 65ºC for 48h and weighed. Specific root length (SRL) was calculated as the ratio between total root length and root biomass.
Finally, roots and leaves were digested in a heating block at 250ºC with a mixture of sulphuric acid and hydrogen peroxide (Jones and Case, 1990). Digests were

Response to drought of Zn-stressed Quercus suber L. seedlings
61
analyzed for Zn by ICP-OES (Perkin Elmer Optima 4300 Inductively Coupled Plasma Optical Emission Spectrometry). Olea europaea L. leaf standard reference materials (BCR: CRM 062, Commission of the European Communities Bureaus of Reference, Brussels) were digested and analyzed for quality control.
2.3. Seedling response to drought
Two months after the beginning of the experiment we stop watering 5 randomly selected seedlings per treatment for 1 week. Physiological measures were performed as seedlings dried out. Water potential soil (Ψs) was estimated by periodic measurements of soil water content and the characteristic moisture curve for this substrate (dew-point psychrometer; WP4 Dewpoint PotentiaMeter; Decagon Device, Inc. Pullman, USA). Seedling performance throughout the imposed drought was evaluated 0, 3 and 6 days after watering was discontinued by measuring chlorophyll fluorescence and RWC on dark-adapted leaves, following the procedures described above.
The Ψs estimated 0, 3 and 6 days after watering was discontinued ranged from –0.96 to –1.01, from –2.78 to -4.21 and from –4.63 to –9.50 respectively. These Ψs correspond to well watering conditions, mild drought and severe drought respectively (Valladares et al., 2005). A Li-Cor 6400 portable infrared gas analyzer system (Li-Cor Inc. Nebraska, US) was used to measure net photosynthetic rate (A), stomatal conductance (gs) and transpiration rate on the same dates on N= 4 - 5 seedlings. From these data we calculated the instantaneous water use efficiency (iWUE). Flow rate was 350 ml min-1, CO2 concentration was 400 ppm and the fixed photosynthetic photon flux density 600 μmol m-2 s-1. Measurements were conducted in early morning (09:00–11:00 h solar time).
2.4. Data analysis
Oneway analyses of variance were conducted to evaluate the effects of Zn on morpho-physiological traits and light-response curve parameters by using Zn dose as fixed factor.
We used repeated-measures ANOVA analysis over time to evaluate the response to decreased water availability on physiological trait of Q. suber seedlings

Capítulo III
62
receiving different Zn supply. Time (0, 3 and 6 days since the onset of drought) was used as within-subject factor and Zn as fixed between-subjects factor. As repeated-measures ANOVA showed significant interactions between Zn and drought, we used separate oneway analysis of variance to evaluate the effects of Zn under different intensities of drought. Zinc dose was used as fixed factor in these analyses. Stomatal conductance and Fv:Fm were raised to a power of 0.5 and 3, respectively, to homogenize the variance before analysis. Factor levels were compared pairwise (Tukey’s HSD test) when ANOVA showed a significant treatment effect.
The relationships RWC vs. maximal PSII photochemical efficiency (Fv:Fm) and Zn dose vs. Zn concentration in leaves were assessed using regression analysis with RWC and Zn dose as independent variables. All statistical analyses were performed with SPSS v.15.0 statistical package (SPSS Inc., Chicago, USA).

Response to drought of Zn-stressed Quercus suber L. seedlings
63
3. Results
3.1. Morphological and physiological response to Zn
Foliar biomass did not respond to Zn (Table 1). Belowground biomass showed similar results (F3,16=1.14; P=0.361, data not shown). Root length was significantly reduced as Zn availability increased, particularly when Zn concentrations were higher than 50 μM. Specific root length decreased at the highest Zn application rate compared to 10 µM Zn.
Zinc concentration was higher in roots than in leaves. It increased in both biomass fractions with the increase in Zn availability, with no evidence of saturation (Fig. 1). The maximum apparent quantum yield (Φm) estimated from the slope of light response curves was ca. 56 % lower in seedlings treated with 150 μM Zn than in control seedlings (Table 2). Maximum net photosynthetic rate (Am)
gradually decreased as Zn concentration increased, but only seedlings receiving 150 μM significantly differed from seedlings receiving 3 and 10 µM.
Maximal photochemical efficiency in fully hydrated leaves was not affected by Zn application rates equal or lower than 50 μM (Table 3, Appendix). Above this concentration, it showed a small but statistically significant reduction. The effect of Zn application was gradually reduced as leaves dried. We may note however that the coefficient of variation increased as RWC decreased.
Table 1. Morphological traits of two month-old seedlings of Quercus suber growing in hydroponic culture with increasing Zn availability. SRL: specific root length. Means, standard errors of n = 5 replicates and results of oneway ANOVA are shown. Different letters indicate significant differences at P < 0.05 (Tukey’s HSD test).
Zinc concentration in solution (μM)
3 10 50 150 F3,16 (P)
Foliar biomass (mg)
507 ± 40 405 ± 43 409 ± 79 366 ± 42 1.25 (0.325)
Root length (m) 1.28 ± 0.12a 1.17 ± 0.15ab 1.12 ± 0.10ab 0.75 ± 0.12b 3.45
(0.042)
SRL (cm mg-1) 0.53 ± 0.05ab 0.65 ± 0.07a 0.48 ± 0.05ab 0.35 ± 0.06b 4.30 (0.021)

Capítulo III
64
Zn concentration (μM)
0 20 40 60 80 100 120 140 160
Folia
r Zn
conc
entra
tion
(μg
g-1)
25
50
75
100
125
150
175
200
225
250
A
y= 17.73 + 1.25 x
R2= 0.99
F1,2= 985.61 P= 0.0010
Zn concentration (μM)
0 20 40 60 80 100 120 140 160
Roo
t Zn
conc
entra
tion
(μg
g-1)
0
500
1000
1500
2000
2500
3000
B
y= 14475.93 x / (772.61 + x)
R2= 0.99
F1,2= 347.69 P= 0.0029
Figure 1. Zinc concentration in leaves (A) and roots (B) of 2-month-old seedlings of Quercus suber growing in hydroponic culture containing different levels of zinc. Models describing the relationships between Zn concentration and Zn availability are shown in each graph.

Response to drought of Zn-stressed Quercus suber L. seedlings
65
Table 2. Light-response curve parameters determined in 2-month-old seedlings of Quercus suber growing in hydroponic culture containing different doses of zinc. Means, standard error of n= 5 replicates and results of oneway ANOVA are shown. Different letters indicate significant differences at P<0.05 (Tukey’s HSD test). Φm: maximum apparent quantum yield. Am: maximum assimilation rate at light saturation.
Zn concentration in solution (μM)
Φm (relative units) Am (µmol m-2s-
1) 3 0.043 ± 0.003a 8.70 ± 1.31a 10 0.032 ± 0.005ab 8.16 ± 0.41a 50 0.032± 0.005ab 7.04 ± 0.75ab 150 0.019 ± 0.055b 4.04 ± 1.09b
F3,16 3.12 4.29 P 0.055 0.021
Table 3. Maximal photochemical efficiency (Fv:Fm) in drying leaves of Quercus suber affected by increasing Zn availability. Means, standard error of n=5 replicates and results of oneway ANOVA per RWC level are shown. Different letters indicate significant differences at P < 0.05 (Tukey’s HSD test).
RWC (%) Zinc concentration in solution (μM)
3 10 50 150 F3,16 P
100 0.69 ± 0.01a 0.70 ± 0.01a 0.68 ± 0.01a 0.64 ± 0.01b 8.43 0.001
75 0.63 ± 0.03ab 0.67 ± 0.01a 0.64 ± 0.01ab 0.59 ± 0.02b 2.81 0.073
60 0.42 ± 0.10 0.39 ± 0.13 0.54 ± 0.04 0.37 ± 0.10 0.63 0.603
3.2. Effects of drought
The response of Q. suber seedlings to the combined effect of Zn and water stress depended on the physiological trait measured (Table 4). We found no effect of Zn on RWC at the onset of drought. Relative water content decreased with decreasing water availability (Table 5; Fig. 2A). At the end of the drought period, seedlings treated with the highest Zn dose showed significantly higher RWC than seedlings exposed to lower doses.

Capítulo III
66
Maximum photochemical efficiency decreased in all seedlings as water stress progressed (Table 4, Fig. 2B). We found no overall effect of Zn or the interaction Zn x water stress on this variable. One-way ANOVA showed that Fv:Fm was lower in seedlings receiving 150 μM of Zn than in seedlings receiving lower doses when water was not limiting. At the end of the drought period, however, the effect of Zn on Fv:Fm was no longer significant (Table 5, Fig. 2B).
Repeated-measures ANOVA revealed a significant Zn x drought interaction on gas exchange variables (Table 4). At the onset of drought, gs was more than two-fold lower in seedlings exposed to 150 μM of Zn than in the other seedlings. In contrast, under severe drought, differences in stomatal conductance resulting from Zn dose were no longer significant (Table 5, Fig 3A). When water was not limiting, differences in A in seedlings receiving increasing doses of Zn were not significant (Table 5). At the end of the drought period, seedlings receiving 50 and 150 μM of Zn showed a marginally significant trend towards higher A than seedlings receiving 3 and 10 μM of Zn (Fig 3B). Instantaneous water use efficiency was not affected by Zn in well watered seedlings (Table 5). Three days after irrigation was discontinued iWUE increased in control seedlings and then decreased in all but in seedlings receiving 150 μM of Zn. After six days, iWUE was higher in seedlings receiving the highest Zn dose than in control seedlings (Table 4, Fig. 3C).
Table 4. Results of repeated measures analysis of variance to evaluate the effect of time and Zn dose on Quercus suber seedlings along a drought period.
Time Zn dose Zn x Time
FT P FZn P FZn x T P
Fv:Fm F3,16 = 15.49 <0.001 F3,16 = 1.55 0.239 F3,16 = 0.36 0.818
RWC F3,16 = 17.40 <0.001 F3,16 = 3.38 0.044 F3,16 = 1.26 0.314
A F3,15 = 60.28 <0.001 F3,15 = 0.71 0.560 F3,15 = 2.40 0.052
gs F3,15 = 40.49 <0.001 F3,15 = 3.44 0.044 F3,15 = 2.98 0.047
iWUE F3,16 = 7.64 0.002 F3,16 = 0.73 0.549 F3,16 = 3.31 0.012

Response to drought of Zn-stressed Quercus suber L. seedlings
67
Time since the onset of drought (days)
0 3 6
RW
C (%
)
0
20
40
60
80
100
3 μM 10 μM 50 μM 150 μMA
aab
abb
Time since the onset of drought (days)
0 3 6
F v:F
m (R
elat
ive
units
)
0.0
0.2
0.4
0.6
0.8 3 μM 10 μM 50 μM 150 μMBa a a
b
Figure 2. Relative water content of detached leaves (RWC; A) and predawn maximum photochemical efficiency (Fv:Fm; B) of seedlings of Quercus suber treated with different doses of Zn under a drought cycle. Different letters indicate significant differences at P < 0.05 (Tukey’s HSD test).

Capítulo III
68
4. Discussion
4.1. Effects of Zn on seedling performance
Exposition to heavy metals may affect plant ability to cope with water stress. The outcome of this effect is not simple as it may depend on the chemical species present and their availability, and the type and magnitude of induced responses in plant morpho-functional traits (Barceló and Poschenrieder, 1990; Kasim, 2007). In our study, Zn accumulated in all biomass fractions, particularly in roots. This pattern has been attributed to metal binding on the cell wall (Greger, 2004) restricted Zn transport aboveground (Utriainen et al., 1997) and vacuolar storage (Ernst et al., 1992). Zinc concentration in the leaves of control seedlings was similar to concentrations found in one-year-old Q. suber (Robert et al., 1996) and in other species (Marschner, 1995), and did not reach critical levels (i.e., 200 µg g-
1d.w. in leaves; Balsberg Påhlsson, 1989) when Zn concentration was 50 μM or lower.
Table 5. Results of oneway analyses of variance to evaluate the effects of water availability on the physiological status of Quercus suber seedlings subjected to increasing Zn availability.
Time since the onset of drought (days)
0 3 6
F P F P F P
FvFm F3,16 = 9.40 0.001 F3,16 = 1.14 0.363 F3,16 = 0.41 0.751
RWC F3,16 = 1.45 0.265 F3,16 = 1.33 0.299 F3,16 = 3.34 0.046
A F3,16 = 1.82 0.185 F3,16 = 1.02 0.409 F3,15 = 2.86 0.072
gs F3,16 = 5.38 0.009 F3,15 = 0.61 0.618 F3,16 = 0.63 0.604
iWUE F3,16 = 1.87 0.175 F3,16 = 1.931 0.165 F3,16 = 3.14 0.054

Response to drought of Zn-stressed Quercus suber L. seedlings
69
Zinc had a weak effect on biomass accumulation and allocation patterns. This may reflect the duration of the experiment and the relatively high resistance of Q. suber to this metal compared to other Mediterranean woody species (Disante et al., 2010).
Time since the onset of drought (days)
0 3 6
g s (m
mol
H2O
m-2
s-1
)
0
20
40
60
80
100
120
140
160
3 μM10 μM50 μM150 μM
A aa
a
b
Time since the onset of drought (days)
0 3 6
A (μ
mol
CO
2 m
-2 s
-1)
-2
0
2
4
6
8
10
12
14
16
3 μM10 μM50 μM150 μM
B
Time since the onset of drought (days)
0 3 6
iWU
E (μ
mol
CO
2 m
mol
H2O
-1)
0
2
4
6
8
10
12
14
3 μM10 μM50 μM150 μM
C
a
ab ab
b
Figure 3. Stomatal conductance (A), photosynthesis rate at 600 µmol m-2 s-1 (B) and instantaneous water use efficiency (C) in seedlings of Quercus suber treated with different doses of Zn and exposed to a drought cycle. Different letters indicate significant differences at P < 0.05 (Tukey’s HSD test).

Capítulo III
70
A trend towards lower biomass accumulation in seedlings receiving the highest levels of Zn, and a significant decrease in C fixation rates in those seedlings suggest that biomass accumulation would be affected by Zn in a longer experiment (Ruano et al., 1988).
Root length and specific root length showed a significant effect of Zn. The reduction in SRL resulted from changes in root geometry, as average root diameter increased with Zn dose whereas root tissue density did not change (data not shown). These results suggest that Zn did not increase lignification, a process that is commonly associated with higher root tissue density (Päivöke, 2003). Lower SRL has been related with increased root longevity (Eissenstat and Yanai, 1997), but these observations are based on data from unstressed plants and may not be applicable to roots exposed to Zn. Conversely, reductions in root length and SRL may constrain plant access to soil resources (Comas and Eissenstat, 2004), and compromise seedling chances of establishment.
4.2. Zinc availability and water stress
The degree of exposition to Zn affected Q. suber response to water stress, but the magnitude of the response depended on the trait measured. Well watered seedlings exposed to 150 µM of Zn showed lower stomatal conductance (as well as lower transpiration rates; data not shown) than seedlings receiving lower doses of this metal. Thus, the rate of soil water depletion was probably lower in these seedlings. Relative water content decreased in control seedlings to 52% in response to severe drought. When a RWC below 50% is reached, cells may not recover when rehydrated (Lawlor and Cornic, 2002). In contrast, RWC of seedlings receiving 50 and 150 μM Zn was always above 70%. Highest doses of Zn may thus contribute to reduce short-term water losses and attenuate the decrease in RWC. Observed values of Fv:Fm and A support this explanation.
At the end of drought, predawn Fv:Fm in intact leaves decreased by 8-14% in all seedlings and was not affected by Zn dose. In detached leaves Fv:Fm was unimpaired until RWC decreased below 70%. These results support previous observations of threshold responses in maximal photochemical efficiency under drought (Methy et al., 1996; Lawlor and Cornic, 2002; Vilagrosa et al., 2003a).

Response to drought of Zn-stressed Quercus suber L. seedlings
71
In well watered seedlings, increasing Zn accumulation resulted in a decrease in gs
and a trend towards lower CO2 assimilation. These changes occurred in parallel, suggesting that stomatal closure was a major driver of the reduction in C fixation. Decreased transpiratory losses have been associated with water stress induced by Zn toxicity (Poschenrieder and Barceló, 2004b; Menon et al., 2005). However, in our study RWC, which is considered a robust indicator of water status (Lawlor and Cornic, 2002) was not affected by Zn application in well watered plants, suggesting that Zn effect on gs did not result from reduced cell hydration. Zinc may also affect stomatal dynamics by inhibiting water channels (Yang et al., 2004) and reducing K+ uptake, thus altering the ratio between K+ and other ions controlling stomata movement (Tsalandzono et al., 1994; Vaillant et al., 2005).
The reduction in stomatal conductance was probably not the only cause of the observed trend towards lower C fixation rates as Zn availability increased. Zinc may decrease the activity of enzymes involved in C fixation (Myśliwa-Kurdziel et al., 2004), reduce the content of chlorophyll by competition with Fe (Balsberg Påhlsson 1989; Myśliwa-Kurdziel et al., 2004), and affect the electron transport chain (Ralph and Burchett, 1998; Myśliwa-Kurdziel et al., 2004). In addition, heavy metals may increase plant sensitivity to PSII photoinhibition (Pätsikkä et al., 2002). The decrease in Fv:Fm, Φm and Am observed in our experiment and described in other studies may be a consequence of this (Long et al., 1994). Photoinhibition commonly has minor direct effects on the performance of woody Mediterranean species, even at high stress levels (Valladares et al., 2005). However, under prolonged unfavourable climatic conditions, photoinhibition may reduce carbon gain and affect the carbon balance (Werner et al., 2001).
As drought progressed, all seedlings showed a significant decrease in CO2 exchange. But the effects of increasing drought on A and gs depended on Zn dose. While drought decreased A in seedlings receiving 150 µM Zn by 65%, A was completely inhibited in control seedlings. In a similar way, gs showed the highest reduction in control seedlings. These results can be partly explained by differences in water conservation and status among seedlings receiving different Zn dose.
Changes in net photosynthetic rate and stomatal conductance in control seedlings resulted in strong variations in iWUE. Instantaneous water use efficiency increased in seedlings subjected to mild drought, as has been widely described in

Capítulo III
72
the literature (Peñuelas et al., 2000; Gimeno et al., 2008). At severe drought, the large reduction in net photosynthetic rates in these plants favoured an intense decrease in iWUE (Valladares and Sánchez-Gómez, 2006). In contrast, changes in A and gs in seedlings receiving 150 �M Zn were less intense, as were changes in iWUE.
Our results show that Zn toxicity may elicit water conservation strategies in Q. suber seedlings. We have shown that these responses may reduce water losses and attenuate the effects of drought. However, in our study drought only lasted for one week. Strategies to withstand long-term drought include changes in leaf area and biomass allocation patterns (van Hees, 1997; Jiménez et al., 2009). Arrested root growth has dramatic effects on seedling ability to establish in areas subjected to seasonal drought (Padilla et al., 2009). Thus, zinc toxicity will likely impair seedling capacity to root and significantly reduce its changes to survive in the long term.
5. Conclusions
To what extent these data confirm our hypotheses? The addition of small quantities of Zn had no significant effect on the morpho-physiological traits of Q. suber seedlings and thus on seedling ability to respond to water stress. On the other hand, the negative effects of high Zn supply rates on seedling traits may have opposing effects on plant performance under drought. In the short term, stomatal closure, decreased transpiratory losses and slow decrease of C fixation may promote water conservation and delay the harmful effects of drought. But the decrease in root length caused by Zn may limit root capacity to obtain soil resources. Our results suggest that in the log term, Zn may have a synergistic interaction with water stress and aggravate the effects of drought.
Acknowledgements
We thank Jose Huesca for their assistance in the lab, David Ramírez for his comments on an earlier draft, Banc de Llavors Forestals (Generalitat Valenciana) for the seeds used in this experiment and the University of Alicante for a FPU (Formación del Profesorado Universitario) grant awarded to K. Disante. This research was funded by the Spanish Ministry of Science and Innovation (Project

Response to drought of Zn-stressed Quercus suber L. seedlings
73
GRACCIE, Programa Consolider-Ingenio 2010 (CSD 2007-00067)). CEAM (Centro de Estudios Ambientales del Mediterráneo) is funded by Generalitat Valenciana and Fundación Bancaja.
Appendix
RWC (%)
0102030405060708090100110
Fv:F
m (r
elat
ive
units
)
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
3 μM10 μM 50 μM 150 μM
Changes in maximal photochemical efficiency and relative water content in detached leaves of Quercus suber seedlings growing in hydroponic culture containing different doses of zinc. Bars correspond to standard errors of 5 leaves per treatment.


Alleviation of Zn toxicity by low water availability
Disminución de la toxicidad por Zn en
condiciones de baja disponibilidad hídrica
Karen B. Disante, Jordi Cortina, Alberto Vilagrosa, David
Fuentes, Encarni I. Hernández, Karin Ljung
Physiologia Plantarum, in press (2013)


Capítulo IV
77
Abstract
Heavy metal contamination and drought are expected to increase in large areas worldwide. However, their combined effect on plant performance has been scantly analyzed. This study examines the effect of Zn supply at different water availabilities on morpho-physiological traits of Quercus suber L in order to analyze the combined effects of both stresses. Seedlings were treated with four levels of zinc from 3 to 150 µM and exposed to low (LW) or high (HW) frequency of watering in hydroponic culture, using a growth chamber. Under both watering regimes, Zn concentration in leaves and roots increased with Zn increment in nutrient solution. Nevertheless, at the highest Zn doses, Zn tissue concentrations were almost twice in HW than in LW seedlings. Functional traits as leaf photosynthetic rate and root hydraulic conductivity, and morphological traits as root length and root biomass decreased significantly in response to Zn supply. Auxin levels increased with Zn concentrations, suggesting the involvement of this phytohormone in the seedling response to this element. LW seedlings exposed to 150 µM Zn showed higher root length and root biomass than HW seedlings exposed to the same Zn dose. Our results suggest that low water availability could mitigate Zn toxicity by limiting internal accumulation. Morphological traits involved in the response to both stresses probably contributed to this response.
Key words: heavy metal, drought, Zn application, auxin, Quercus suber, multiple stresses

Alleviation of Zn toxicity by low water availability
78
Resumen
A escala global se espera un incremento tanto de la contaminación por metales pesados, como de la duración de los periodos de sequía. Aun así, el efecto de la acción simultánea de estos factores sobre el desarrollo de las plantas ha sido escasamente analizado. Este estudio examina el efecto de la aplicación de Zn y diferentes niveles de disponibilidad hídrica sobre las características morfo-funcionales de Quercus suber L. con el fin de analizar el efecto combinado de ambos factores de estrés. Las plántulas fueron tratadas con cuatro niveles de Zn desde 3 hasta 150 µM y expuestas a baja (LW) o alta (HW) frecuencia de riego en cultivo hidropónico, utilizando una cámara de crecimiento. La concentración de Zn en hojas y raíces incrementó con el incremento de Zn en la solución nutriente independientemente del nivel de riego. Sin embargo, para la dosis más alta de Zn, la concentración de Zn en los tejidos fue casi dos veces mayor en las plántulas HW respecto a las LW. Algunas características funcionales, como la tasa fotosintética y la conductividad hidráulica de la raíz, y morfológicas como la longitud y biomasa radicular decrecieron significativamente en respuesta a la aplicación de Zn. Los niveles de auxina incrementaron con el aumento de Zn, sugiriendo la implicación de esta fitohormona en la respuesta de la planta a este elemento. Las plántulas LW tratadas con 150 µM Zn mostraron mayor longitud y biomasa radicular que las HW expuestas a la misma dosis de metal. Nuestros resultados sugieren que la baja disponibilidad hídrica podría mitigar la toxicidad de Zn limitando su acumulación interna. Las caracterísiticas morfológicas involucradas en la respuesta a ambos estreses probablemente contribuyeron a esta respuesta.
Palabras clave: metal pesado, sequía, aplicación de Zn, auxina, Quercus suber, estrés múltiple.

Capítulo IV
79
1. Introduction
In nature, plants are commonly exposed to a combination of stress factors. Multiple stresses may affect plants in ways that are hardly predictable from studies of single stress factors, as synergistic and antagonistic interactions are widespread (Nilsen and Orcutt, 1996; Larcher, 2003). Water is the most limiting factor for plant productivity worldwide and several studies have evaluated the combined effect of drought and other sources of stress such as extreme temperature and radiation (Valladares and Pearcy, 1997; Aranda et al., 2005, Gimeno et al., 2008). In contrast, the combined effect of drought and heavy metal contamination has received less attention, despite the potential interaction between both stress factors (Poschenrieder and Barceló 2004b; Brady et al., 2005; Sardans and Peñuelas, 2007). Several studies have shown that the effect of heavy metals on plant performance may be modulated by other sources of stress as sulphur and proton concentration in rainfall, heat and frost (Kukkola et al., 1997; Öncel et al., 2000).
Among heavy metals, Zn is an essential micronutrient for higher plants and it is required for several physiological processes (Marschner, 1995; Cakmak, 2000; Broadley et al., 2007). Zinc abundance is increasing in natural systems as a consequence of human activities (Kabata-Pendias and Pendias 1992; Fuentes et al., 2007a; Dominguez et al., 2008). When Zn concentration exceeds a critical level, it becomes toxic (Balsberg Påhlsson, 1989; Woolhouse, 1983). Symptoms of Zn toxicity include alteration on biomass partitioning (Ruano et al., 1988), morphological changes in root system (Fuentes et al., 2007b; Disante et al., 2010), reductions in photosynthetic rate and chlorophyll measurements (Di Baccio et al., 2009; Disante et al., 2011), unbalance mineral nutrition (Kabata-Pendias and Pendias, 1992; Monet et al., 2001) and modifications in xylem anatomy (Kasim, 2007). These changes may affect plant capacity to uptake and transport water to above ground parts (Barceló and Poschenrieder, 1990; Kasim, 2007).
High levels of Zn and low water availability may induce similar responses in vascular plants, including stomatal closure and decreased CO2 uptake, reduced chlorophyll fluorescence and foliar area, and activation of defence mechanisms against oxidative damage (Vaillant et al., 2005; Valladares et al., 2005). Conversely, plants may exhibit contrasted responses to Zn toxicity and drought. While reduced root length and root biomass have been described in seedlings

Alleviation of Zn toxicity by low water availability
80
exposed to Zn (Wolhouse, 1983), adaptive mechanisms in response to water limitation include deeper root systems and increased proportion of biomass allocated belowground (Nicotra et al., 2002).
The interaction between Zn and drought may result in synergistic or antagonistic responses. Thus, small increases in Zn availability (20 mg l-1 of Zn) promoted growth in water-stressed soybeans (Gadallah, 2000). Similarly, heavy metal-enriched sewage sludge induced a positive effect on morpho-physiological traits of seedlings subjected to drought (Pascual et al., 2004; Fuentes et al., 2007a; Santala and Ryser, 2009). Finally, in a previous study we observed that exposition of Quercus suber seedlings to Zn delayed the effects of a short drought (Disante et al., 2011). High Zn concentrations could hamper seedling capacity to absorb and transport water and compromise seedling ability to withstand drought due to reduction of root length and biomass (Disante et al., 2010). Limited water availability may also predispose leaves to cellular damage and photoinhibition, altering the carbon balance (Werner et al., 2001; Vilagrosa et al., 2010). Finally, drought may impair uptake and acropetal translocation of nutrients, including Zn (Hu and Schmidhalter, 2005).
Despite the potential interaction between Zn and drought, studies dealing with the simultaneous effect of both stress factors are scarce, and they frequently show conflictive results (Balsberg Påhlsson, 1989). The objective of this study is to evaluate the combined effect of Zn supply and water availability on plant performance, and contribute to unveil the interactions between both sources of stress. The underlying hypothesis is that morpho-physiological responses to drought may mitigate the effects of high Zn availability. For this study we used cork oak (Quercus suber L.) seedlings, a drought-resistant tree widely distributed in the western Mediterranean basin. Cork oak woodlands produce valuable goods and services, including cork, and they are protected by the European Union (Habitat directive 92/43/EEC; (Pausas et al., 2009)). In addition, we performed this experiment with young seedlings because this stage is one of the most sensitive for plant capacity to resist stress conditions (Vallejo et al., 2000; Vilagrosa et al., 2003a). To account for this objective, we analyzed plant physiological status, root capacity to supply water to leaves, and phytohormonal response in several Zn supply rates under two levels of water availability.

Capítulo IV
81
2. Materials and methods
2.1. Plant material and growing conditions
We cultivated Q. suber seedlings from acorns. Seeds from local origin were supplied by the Forest Service Seed Bank (Banco de Semillas El Serranillo, Ministerio de Medio Ambiente y Medio Rural y Marino). Quercus suber acorns were sown in silica sand and kept at 24 ± 2˚C. Two weeks after germination, when the radicle was approximately 2 cm long, acorns were transplanted into 5 cm diameter and 30 cm length tubes filled with fine, medium and coarse silica sand (the size of sand ranged from 0.5 to 2.2 mm) in a proportion corresponding to 1:2:3 by volume, respectively. The substrate was tested in the lab before the onset of the experiment to ensure proper drainage. The bottom of the tubes was closed with a 1 x 1 mm mesh, which prevented substrate loss. Tubes were placed in a growth chamber under controlled conditions of temperature (22±1°C/18±1°C, day/night), 12 h photoperiod (600 μmol m-2 s-1 of photosynthetic photon flux density during the day) and 50% relative humidity. During this period, seedlings were irrigated with a nutrient solution containing 500 μM KCl, 435 μM NaNO3, 300 μM MgSO4·7H2O, 45 μM NaH2PO4, 300 μM CaCl2, 0.37 μM FeSO4·7H2O, 0.012 μM CuSO4·5H2O, 1.6 μM H3BO3, 0.006 μM KI, 0.6 μM MnSO4·7H2O and 0.0004 μM (NH4)6Mo7O24. This solution was a modification of the one used by Arduini et al., (1994) to simulate forest soil conditions.
2.2. Experimental design
Eight weeks after germination, we added ZnSO4 to the nutrient solution to achieve a Zn concentration of 3 (Control), 10, 50 and 150 μM, corresponding to those commonly found in uncontaminated and contaminated soils (Knight et al., 1997; Kabata-Pendias and Pendias, 1992; González et al., 2011).
We applied two irrigation regimes using this solution: high watering (HW) and low watering (LW). HW seedlings were irrigated three times a week (Monday, Wednesday and Friday), whereas LW seedlings were irrigated every 5 days. During the experiment, seedlings were irrigated with 60 ml of nutrient solution. Substrate moisture content (SMC) was estimated by measurements of water content by gravimetric method. SMC for HW and LW treatments ranged from 2.0

Alleviation of Zn toxicity by low water availability
82
± 0.1% to 7.4 ± 0.1% and from 0.3 ± 0.1% to 6.40 ± 0.2% respectively. The moisture content (SMC) for the substrate during 18 consecutive days is shown in Fig. 1 as example of SMC dynamics.
Days0 2 4 6 8 10 12 14 16 18 20
SMC
(%)
0
2
4
6
8
10
SMC LWSMC HW
Figure 1. Change in substrate moisture content (SMC) for HW and LW seedlings during 18 consecutive days. HW and LW correspond to high and low watering frequency, respectively. Means and standard errors of N= 6-7 tubes per treatment are shown.
Each Zn level (hereafter treatments) was replicated 10 times, i.e., 10 seedlings per Zn treatments and irrigation level. Ion speciation calculations for solutions containing different amounts of Zn were conducted using MINTEQA2 software (Allison Geoscience Consultants, Inc. Flowery Branch, Georgia, USA and HydroGeoLogic, Inc. Herndon, Virginia, USA). The concentration in mol/L of all anions and cations present in the nutrient solution (described above) were entered in MINTEQA2 software to estimate the equilibrium composition of the experimental solution. pH was fixed at 5.5 (the pH of the nutrient solution was adjusted to this value) and quartz was added as the mineral present in the equilibrium.
Calculations showed that 98% of Fe was present as Fe (II) and 81% of Zn was present as free ion and 18% as ZnNO3+. Both elements were highly available for

Capítulo IV
83
plants, as only small precipitations of PO4-3 (0.2%) and Mn+2 (16.2%) were found
by chemical equilibrium estimations using MINTEQ2A (see Appendix S1 in Supporting information). Leachates obtained immediately after watering showed that Zn concentration in the solution was always close to the intended concentration, independently of water availability and plant uptake. For 3 (Control), 10, 50 and 150 μM Zn were: 4.2 ± 0.4, 10.3 ± 3.7, 47.6 ± 8.7; 150.2 ± 12.9 μM Zn and 2.8 ± 0.4, 7.8 ± 1.3, 46.8 ± 6.8, 162.9 ± 16.3 μM Zn for HW and LW seedlings, respectively. Twelve weeks after the onset of the treatments, when seedlings were 5-months-old, we assessed their physiological status, and measured hydraulic conductivity and IAA concentration. Finally, we quantified their morphological traits.
2.3. Gas exchange and chlorophyll fluorescence
Physiological status was assessed 5 months after the onset of the experiment on 5 seedlings per treatment level (i.e. 40 seedlings in total). Gas exchange was analysed with a Li-Cor 6400 portable infrared gas analyser system (Li-Cor Inc. Lincoln, Nebraska, USA). We determined photosynthetic rate (A), stomatal conductance (gs) and transpiration rate (E) in one fully expanded leaf per seedling. From these data, we calculated the instantaneous water use efficiency (IWUE). We maintained gas flow rate at 350 mL min-1, CO2 concentration at 400 ppm and a photon flux density of 600 μmol m-2 s-1 during the course of the measurements. Previous determinations of light response curves showed that a value of 600 μmol m-2 s-1 was sufficient to saturate the PSII photosystem in seedlings of this species. Measurements were conducted early in the morning (09:00–11:00 h solar time).
A portable fluorometer (pulse-amplitude modulated photosynthesis yield analyzer, PAM-2010 Walz, Effeltrich, Germany) was used to measure chlorophyll fluorescence from intact leaves. In dark adapted leaves, we measured the minimum fluorescence yield (F0) under weak red light and the maximum fluorescence (Fm) after a saturating pulse of white light. Maximal PSII photochemical efficiency (Fv:Fm) was calculated as Fv:Fm= (Fm - F0)/ Fm according to Genty et al. (1989)

Alleviation of Zn toxicity by low water availability
84
2.4. Root hydraulic conductance
Four to five seedlings per Zn dose and irrigation regime were used to measure water transport capacity of complete and intact root systems by means of a High Pressure Flowmeter (HPFM, Dynamax Inc., Houston, USA). Measures were carried out at the same time that gas exchange variables were measured. The night before the measurements, seedlings were watered to pot capacity, which is the amount of water remaining in a pot after irrigation and visible drainage had ceased. Root hydraulic conductance (KR) measurements were carried out early in the morning. Root systems were kept in their substrate and perfused with distilled and de-gassed ultra-pure water filtered through a 0.2 µm water filtration membrane using the high pressure flow meter method (HPFM, Dynamax, USA) connected to the stump. KR was measured in a transient way within a range of 0–0.5 MPa pressure at a constant increment rate of 3–5 kPa s−1 and the flow rate (F) was recorded every 3 s as described in Tyree et al., (1995). KR (Kg m s-1 MPa-1) was calculated as the slope of ΔF (Kg s-1) versus ΔP (MPa) as KR = dF/dP. Kr was corrected for variations in water temperature. Morphological variables were used to normalize root hydraulic conductance (KR) on root surface area basis (KR-RA; Kg m-2 s-1 MPa-1).
2.5. Seedling morphology
Five months after the onset of the experiment, seedlings were harvested, stems cut above the root collar and the rooting system carefully washed. Roots and leaves were digitized by scanning on an A3 flatbed scanner (Epson Expression 1680 Pro, Long Beach, California, USA) fitted with a transparency adaptor at 300 dpi, using an 8-bit greyscale. Images were analyzed with specific software (WinRhizo, Regent Instruments, Québec, Canada) to evaluate projected leaf area, total root length and total root surface area. All biomass fractions were dried at 65ºC for 48 h, and then weighed. Specific root length (SRL) was calculated as the ratio between total root length and root biomass.

Capítulo IV
85
2.6. Zn concentration in plant tissues
Dry roots and leaves were digested in a heating block at 250ºC with a mixture of sulphuric acid and hydrogen peroxide (Jones and Case 1990). Digests were analyzed for Zn by ICP-OES (Perkin Elmer Optima 4300 Inductively Coupled Plasma Optical Emission Spectrometry). Olea europaea L. leaf standard reference material (BCR: CRM 062, Commission of the European Communities Bureau of Reference, Brussels) was digested and analysed for quality control.
2.7. Quantification of indole-3-acetic-acid (IAA)
Frozen leaves (10 mg) were homogenised in 0.5 ml of 0.05 M sodium phosphate buffer (pH 7.0) containing 0.02% diethyldithiocarbamic acid as an antioxidant and 500 pg13C6-IAA as an internal standard, using the Retsch vibration mill (Retsch MM 200 mixer mill, Retsch GmbH, Haan, Germany) and a 3-mm tungstencarbide bead at a frequency of 30 Hz for 3 min.The pH was adjusted to 2.7 with 1 M HCl, and the samples were purified by solid phase extraction (SPE) and derivatized as described Andersen et al., (2008). Finally, samples were dissolved in 30 µl heptane and analyzed by GC-SRM-MS (gas chromatography - selected reaction monitoring - mass spectrometry) as described in Edlund et al. (1995).
2.8. Data analysis
Treatment effects on physiological and morphological variables were evaluated using two-way ANOVA. When the interaction term was significant, one-way ANOVA and Student-t were used to evaluate the effect of Zn at each watering level, and the effect of increased water availability at each Zn level, respectively. Root biomass and instantaneous water use efficiency were raised to a power of 0.5 and root Zn concentration was transformed using natural logarithm prior to analysis, to homogenize the variance. Factor levels were compared pairwise (Tukey’s HSD test) when ANOVA showed a significant treatment effect.
We used regression analysis to evaluate the significance of the relationship between KR and A and gs. All analyses were performed using the SPSS v.15.0 statistical package (SPSS Inc., Chicago, Illinois, USA).

Alleviation of Zn toxicity by low water availability
86
3. Results
3.1. Zn bioaccumulation
Foliar Zn concentration increased in HW and LW seedlings with increasing Zn concentration in the growing medium (Fig. 2A). The magnitude of the increase was dependent on water availability (Table 1, Table 2). Thus, at the highest Zn dose, its concentration was two-fold higher in leaves of HW seedlings than in leaves of LW seedlings. The Zn bioaccumulation in roots was affected by the increase in Zn load and by the interaction between both factors (Fig 2B, Table 1).
3 10 50 150
Folia
r Zn
conc
entr
atio
n (m
g g-1
)
0
50
100
150
200
250
300
350HWLW
A
Zn dose (μM)3 10 50 150
Root
Zn
conc
entr
atio
n (m
g g-
1 )
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
HWLW
B
AB
C
D
a a
b
c
*
*
*
AB
C
D
ab
c
c
*
Figure 2. Foliar (A) and root (B) Zn concentration of Quercus suberseedlings growing at four Zn doses and two contrasting water availability levels (HW and LW for high and low watering frequency, respectively). Bars are means and standard errors of N=5 seedlings. Note the different scale used for leaves and roots. Note that data distribution along the X-axis is not proportional. Different capital letters and lowercase letters indicate significant differences at P < 0.05 for HW seedlings and LW seedling respectively (Tukey’s HSD test). Asterisks indicate significant differences at P < 0.05 between seedlings HW and LW exposed to the same Zn dose (t-Student).

Capítulo IV
87
3.2. Morphological response
Foliar biomass was not affected by Zn application, water availability or the interaction between both (Table 1and 3). On the contrary, root biomass showed a significant effect of the increase in Zn load, and the interaction between Zn application and water availability. While root biomass in HW seedlings decreased with increasing Zn dose, LW seedlings showed the opposite trend (Table 2). At the highest Zn dose, root biomass accumulation was 33% lower in HW seedlings than in LW seedlings. This response was similar for root length, but changes in root length in LW seedlings with increasing Zn concentration were not statistically significant (Table 2). We found a marginally significant trend towards lower specific root length as Zn dose increased at any watering level (Table 1 and 3).
3.3. Physiological response
Maximum PSII photochemical efficiency (Fv:Fm) ranged between 0.65 and 0.72. Fv:Fm was significantly decreased by Zn but not by water availability (Table 1). The interaction between Zn and watering regime was marginally significant as the effect of drought on this variable decreased as Zn dose increased above 3 μM Zn (Table 1, Appendix S2 in Supporting Information). In HW seedlings, Fv:Fm decreased with the increase of Zn while Fo showed the opposite trend (Table 2).We found no significant effect of Zn application in Fv:Fm and Fo in LW seedlings.
Zinc application, water availability and their interaction significantly affected gas exchange (Table 1, Fig. 3A and 3B). HW seedlings receiving 3μM Zn, had 2.8 and 3.9 times higher net photosynthetic rate (A) and stomatal conductance (gs) than LW seedlings, respectively. Stomatal conductance and A decreased with increasing Zn dose in HW (Table 2). In contrast, A in LW seedlings was weakly affected by Zn application, and gs showed a unimodal response to increase in Zn load (Table 2). Instantaneous water-use-efficiency (IWUE) increased as Zn dose increased and, in contrast to other variables, this effect was independent of the irrigation regime (see Appendix S3 in Supporting Information).

Alleviation of Zn toxicity by low water availability
88
Table 1. Results of two-way ANOVA to evaluate the effects of Zn and water availability (w) on morpho-physiological traits of Quercus suber seedlings. Foliar Zn: foliar Zn concentration, Root Zn: root Zn concentration, SRL: specific root length, Fv:Fm: predawn maximum photochemical efficiency, Fo: minimum fluorescence, A: photosynthesis; gs: stomatal conductance, IWUE: instantaneous water use efficiency, KR-RA: root hydraulic conductance per root area, IAA: auxin concentration in leaves.
Variables FZn PZn Fw Pw FZnxw PZnxw
Foliar Zn F3,26 =105.34 <0.001 F1,26 =12.78 0.001 F3,26 =3.34 0.034
Root Zn F3,26 =46.67 <0.001 F1,26 =0.01 0.920 F3,26 =2.86 0.056
Foliar
biomass
F3,48 =1.95 0.135 F1,48 =0.01 0.905 F1,48 =2.00 0.127
Root
biomass
F3,50 =2.95 0.041 F1,50 =0.94 0.337 F3,50 =5.11 0.004
Root length F3,48 =0.25 0.858 F1,48 =0.02 0.870 F3,48 =3.29 0.028
SRL F3,45 =2.44 0.076 F1,45 =0.93 0.339 F3,45 =0.13 0.943
Fv:Fm F3,30 =5.09 0.006 F1,30 =2.14 0.154 F3,30 =2.36 0.091
Fo F3,30 =2.66 0.066 F1,30 =1.18 0.287 F3,30 =2.85 0.054
A F3,30 =3.74 0.021 F1,30 =32.52 <0.001 F3,30 =3.97 0.017
gs F3,30 =4.04 0.016 F1,30 =31.35 <0.001 F3,30 =4.00 0.017
IWUE F3,30 =2.86 0.054 F1,30 =2.71 0.111 F3,30 =0.40 0.756
KR-RA F3,32 =2.96 0.047 F1,32 =22.76 <0.001 F3,32 =6.96 0.001
IAA F3,23 =3.58 0.029 F1,23 =0.08 0.773 F3,23= 2.84 0.060
3.4. Hydraulic conductance
Root hydraulic conductance in a root area basis (KR-RA) was sensitive to Zn doses, watering regime and their interaction (Fig. 4, Table 1). KR-RA gradually decreased as Zn availability increased in HW seedlings, whereas KR-RA was not affected by Zn doses in LW seedlings (Fig. 4, Table 2). KR-RA was significantly higher in HW seedlings than in LW seedlings at low Zn availability, but this difference

Capítulo IV
89
disappeared at Zn concentrations of 50 μM and higher (Appendix S4 in Supporting Information). Root hydraulic conductance per unit of leaf area and root hydraulic conductance per unit of root length showed similar trends (data not shown).
Table 2. Results of one-way ANOVA to evaluate the effects of Zn on morpho-physiological traits of Quercus suber seedlings subjected to high and low watering frequency (HW and LW, respectively). Foliar Zn: foliar Zn concentration, Root Zn: root Zn concentration, Fv:Fm: predawn maximum photochemical efficiency, Fo: minimum fluorescence; A: photosynthesis; gs: stomatal conductance, KR-RA: root hydraulic conductance per root area, IAA: auxin concentration in leaves.
Variables FHW PHW FLW PLW
Foliar Zn F3,13 = 92.33 <0.001 F3,13 = 31.51 <0.001
Root Zn F3,14 = 99.27 <0.001 F3,14 = 35.55 <0.001
Root biomass F3,23 = 4.23 0.016 F3,27 = 2.93 0.052
Root length F3,22 = 2.57 0.080 F3,26 = 1.35 0.281
Fv:Fm F3,15 = 6.66 0.004 F3,15 = 0.86 0.485
Fo F3,15 = 4.88 0.015 F3,15 = 0.31 0.819
A F3,16 = 5.02 0.012 F3,15 = 2.46 0.106
gs F3,16 = 4.61 0.017 F3,15 = 3.34 0.050
KR-RA F3,15 = 7.30 0.003 F3, 18 = 0.63 0.602
IAA F3,11 = 4.39 0.029 F3,12 = 0.90 0.468

Alleviation of Zn toxicity by low water availability
90
3 10 50 150
A (m
mol
m-2
s-1)
0
2
4
6
8
10
12HWLW
g s (m
ol m
-2 s-1
)
A
Zn dose (μM)3 10 50 150
0,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18HWLW
B
A
ABAB
B* *
A
AB AB
B* *
Figure 3. Photosynthesis (A) and stomatal conductance (B) of Quercus suber seedlings exposed to different levels of Zn availability and contrasting watering frequency. Bars are means and standard errors of N=5 seedlings. Note that data distribution along the X-axis is not proportional. Different capital letters and lowercase letters indicate significant differences at P < 0.05 for HW seedlings and LW seedling respectively (Tukey’s HSD test). Asterisks indicate significant differences at P < 0.05 between seedlings HW and LW exposed to the same Zn dose (t-Student).
3.5. IAA concentration
Foliar IAA concentration was significantly affected by the increase in Zn load, and marginally affected by the interaction between Zn application and water availability (Fig. 5, Table 1). This was the result of a substantial increase in IAA in HW seedlings receiving 150 μM of Zn compared to HW seedlings receiving lower Zn doses and LW seedlings receiving any Zn dose (Table 2).

Capítulo IV
91
4. Discussion
In Mediterranean type ecosystems, water stress is the main constraint to plant performance (Di Castri, 1973; Rhizopoulou and Mitrakos, 1990). In these areas, prolonged summer drought, low water storage capacity of forest soils and high summer temperatures could amplify the effects of other sources of stress such as heavy metals, high temperatures or high radiation levels. In addition, the deleterious impact of both sources of stress may be enlarged in the future as a consequence of climate change and increased inputs of heavy metals (Sardans and Peñuelas, 2007; Micó et al., 2006; IPCC 2001). Results of the present study show that water availability and heavy metal contamination may indeed interact in complex ways, and they suggest that the combined effects of both sources of stress may not be additive simply.
4.1. Zn effects on seedling performance
Zinc concentration in leaves of seedlings exposed to 3 μM Zn was similar to concentrations found in 1-year-old Q. suber (Robert et al., 1996). Foliar Zn concentration reached critical levels (e.g., 200 μg g−1d.w.; Balsberg Pahlsson, 1989) only after exposition to 150 μM Zn. Maximum Zn concentration belowground in HW seedlings was 1.7 higher than in LW seedlings. Similar values of Zn bioaccumulation have been found in other woody species grown in hydroponic culture (e.g., 3444 μg g−1 to 5700 μg g−1; Reichman et al., 2001; Fuentes et al., 2007b).
Aboveground biomass accumulation showed no evidence of Zn toxicity. Similar response had been observed previously (Disante et al., 2011). This could be a consequence of the time span of the experiment and low Zn concentration in shoots, as belowground biomass accumulation and ecophysiological status (as leaf photosynthetic rate) were affected by Zn application. In addition, stem elongation in oaks follows successive flushes (Johnson et al., 2001). Thus, we might observe changes in shoot biomass in response to the increase in Zn load after a new flush of growth. Reductions in aboveground biomass accumulation and chlorosis in seedlings exposed to high levels of Zn have been reported in other woody species in experiments of different time span (Reichman et al., 2001; Fuentes et al., 2007b; Disante et al., 2010).

Alleviation of Zn toxicity by low water availability
92
Zn dose (μM)3 10 50 150
K R-R
A (K
g s-1
MPa
-1 m
-2 x
10-4
)
0
2
4
6
8
10
12
14
16
18
HW LW
A
B
B
B*
*
Zn dose (μM)3 10 50 150
IAA
(pg
mg- 1
)
0
2
4
6
8
10
12
14
HWLW *
1 Different capital letters and lowercase letters indicate significant differences at P < 0.05 for HW seedlings and LW seedling respectively (Tukey’s HSD test). Asterisks indicate significant differences at P < 0.05 between seedlings HW and LW exposed to the same Zn dose (t-Student).
On the contrary, belowground parts were more sensitive to increase in Zn load than aboveground parts as reflected by the reduction in root biomass and the marginally significant decrease in root length of well watered seedlings receiving increasing doses of heavy metal. The reduction in root biomass accumulation is a common and quick response to high Zn availability (Ruano et al., 1988). Indeed
Figure 4. Specific hydraulic conductance per root surface area in Quercus suber seedlings exposed to different levels of Zn availability and contrasting watering frequency. Bars are means and standard errors of N=4-5 seedlings. Note that data distribution along the X-axis is not proportional (1).
Figure 5. Indole-3-acetic acid (IAA) concentration in leaves of Quercus suber seedlings exposed to different levels of Zn availability and contrasting watering frequency. Bars are means and standard errors of N=3-4 seedlings. Note that data distribution along the X-axis is not proportional (1).

Capítulo IV
93
Zn concentration in roots was up to ten times higher than in shoots. This may be a result of the strong control of Zn translocation of Q. suber, possibly excluder species (Disante et al., 2010).
Functionality of Q. suber leaves was negatively affected by the increase in Zn load. Previous studies suggested that Zn increases plant sensitivity to photoinhibition (Joshi and Mohanty, 2004; Mateos-Naranjo et al., 2008). The decline observed in Fv:Fm may be due to damage in the reaction centre and down-regulation processes (Osmond et al., 1999). In our experiment, this reduction was caused by a weak increase in Fo that can be induced by damages to the PSII reaction centre (Moustakas et al., 1997), including reductions in the energy transfer from the antennae to the reaction centre associated with chronic photoinhibition (Ralph and Burchett, 1998; Popovic et al., 2003). Similarly, we observed a reduction in A and gs in seedlings exposed to Zn application, as described in previous works (Myśliwa-Kurdziel et al., 2004; Disante et al., 2011). Impaired carbon fixation may partly reflect photoinhibition and photon damage, but also the reduction in gs.
Another direct effect over gs may be associated with the decrease in water transport capacity in roots affected by increment of this heavy metal. Supporting this hypothesis, we found a reduction in hydraulic conductance of roots and a positive relationship between KR and gas exchange rates (R2= 0.64, F= 10.66, P= 0.0171, and R2= 0.73, F= 15.99, P= 0.0071 for A and gs respectively). These results are in agreement with observations on stomata sensitivity to decreasing KR, and the prevention of xylem cavitation (Brodribb and Holbrook, 2003; Vilagrosa et al., 2003b; Otieno et al., 2007). The negative effect of the increase in Zn load altering root morphology supports our observations in root functionality (i.e. decreases of KR). Heavy metals may modify root hydraulic conductance by different mechanisms, but studies on this topic are relatively scarce (Poschenrieder and Barceló, 2004b).

Alleviation of Zn toxicity by low water availability
94
Tabl
e 3.
Mor
phol
ogic
al a
ttrib
utes
of 5
-mon
ths-
old
Que
rcus
sub
er s
eedl
ings
gro
wn
unde
r a
rang
e of
Zn
supp
ly
rate
s an
d co
ntra
sted
irr
igat
ion
regi
mes
. H
W a
nd L
W c
orre
spon
d to
hig
h an
d lo
w w
ater
ing
freq
uenc
y,
resp
ectiv
ely.
Mea
ns a
nd s
tand
ard
erro
rs o
f N=6
-7 s
eedl
ings
are
sho
wed
. Diff
eren
t cap
ital l
ette
rs a
nd lo
wer
case
le
tters
indi
cate
sig
nific
ant d
iffer
ence
s at
P <
0.0
5 fo
r H
W s
eedl
ings
and
LW
see
dlin
g re
spec
tivel
y (T
ukey
’s H
SD
test
). A
ster
isks
indi
cate
sig
nific
ant d
iffer
ence
s at
P <
0.0
5 be
twee
n se
edlin
gs H
W a
nd L
W e
xpos
ed to
the
sam
e Zn
dos
e (t-
Stud
ent).

Capítulo IV
95
Thus, previous studies found that Hg can block aquaporins in cell membranes (Lee et al., 2005), and Zn and Cd may foster the deposition of phenolic compounds and the reduction in vessel size (Fuhrer, 1982; Poschenrieder et al., 1989). Independently of the mechanisms involved, the reduction of KR and the concomitant decrease in A and gs, together with changes in root biomass accumulation and root morphology, suggest that Zn may hamper the ability of Q. suber seedlings to access and transport soil resources to aboveground parts. The development of extended rooting system is important to ensure plant survival and early growth in Mediterranean conditions, and especially for seedling establishment in areas subjected to intense droughts (Padilla and Pugnaire, 2007; León et al., 2011).
Hormonal response to heavy metals is another response poorly studied, especially in woody species. We found a substantial increase in IAA in leaves of well watered seedlings as Zn concentration increased. Auxins are mainly synthesized in shoots, and then translocated and accumulated in roots (Marschner, 1995; Ljung et al., 2005) This hormone is involved in several process related to the coordination of root and shoot growth and development, participating in processes as cell elongation, formation and differentiation of vascular tissue, and in the shaping of root architecture (Mattsson et al., 2003; Aloni et al., 2006; Robert and Friml, 2009). Our results show that the increase in Zn load may elicit alterations in endogenous auxin levels and suggest that this could promote the observed alterations in root morphology and functionality. Indeed, other studies have found changes in auxin accumulation after Cu exposition, which was related to morphological changes as the formation of lateral roots (Lequeux et al., 2010; Pető el at., 2011).
4.2. Combined effects of Zn and water availability
We found no significant effect of the increase in Zn load on aboveground biomass accumulation in HW seedlings, whereas aboveground biomass showed a marginally significant increase with Zn application in LW seedlings. Several studies have found increases in plant growth in response to small amounts of available Zn (Gadallah, 2000; Fuentes et al., 2007b). In contrast, belowground biomass increased and decreased in LW and HW seedlings, respectively, as Zn availability increased. The marginal increase in root biomass in LW seedlings was

Alleviation of Zn toxicity by low water availability
96
probably due to the combination of small changes in root diameter (data not shown) and root length (Table 3): 5% and 12% increase, respectively, in seedlings exposed to 150 μM Zn compared to seedlings exposed to 3 μM Zn. We may note that root biomass and root length were 1.5 times higher in LW seedlings than in HW seedlings when both sets received 150 μM Zn. A study combining Zn and salinity found similar results, as Zn application reduced root length and root biomass accumulation in unstressed seedlings, but it increased root growth in seedlings subjected to saline stress (Gadallah and Ramadan, 1997).
Quercus suber displays intermediate tolerance to drought (Ramírez-Valiente et al., 2009). In our experiment, we did not test intense and extended drought that could probably affect root anatomy. Our objective was to study how the reduction in the frequency of irrigation influences seedling response to different Zn doses. This may explain why contrasted watering frequency did not induce changes in root length or root biomass in seedlings receiving 3 μM Zn. Data on Table 1 show that the effects of Zn and watering frequency on several seedling traits depend on the levels of the other factors. The interaction between drought and Zn points out at a synergistic or antagonistic effect of the two factors. Table 2 and Table 3 show that Zn had opposite effects on seedlings depending on the watering level. Despite that the effect of Zn on root architecture, (i.e., root biomass) was marginally significant in LW seedlings (P= 0.052), we consider this effect to be important and worth discussing. Besides, low watering induced a marginal decrease in root average diameter compared to HW (Appendix S5 in Supporting information). Results of a two-way ANOVA showed the lack of a significant interaction between Zn and drought, and indicate that roots of seedlings receiving low irrigation were thinner than HW seedlings. In other words, LW induced changes in root morphology.
Physiological response to increasing Zn availability also differed in LW and HW seedlings. Low irrigation frequency decreased gas exchange rates, and probably limited Zn uptake and accumulation. This may explain the lower foliar and root Zn concentrations found in LW seedlings compared with HW seedlings, and may be partially responsible for the smaller effect of Zn doses observed in the former seedlings. Zinc is incorporated by specific transporters (Lasat et al., 2000, Takahashi et al., 2012) and to our knowledge there is no report about the effect of water availability on these transporters. But drought generally reduces nutrient

Capítulo IV
97
transport because it impairs transpiratory flow and active transport (Hu and Schmidhalter, 2005).
All physiological traits measured in Q. suber decreased as Zn concentration increased in HW seedlings, whereas most traits showed a unimodal response, albeit not statistically significant, to increasing Zn in LW seedlings. Stomatal conductance and carbon assimilation rates of LW seedlings receiving the lowest Zn corresponded to a values obtained in Q. suber seedling exposed to mild drought (Disante et al., 2011), but they were not further reduced by the increase in Zn load. As a consequence, physiological status of LW and HW seedlings strongly differed in seedlings receiving the lowest doses of Zn, but differences gradually disappeared as Zn availability increased, or Zn concentration in roots increased above ca. 1000 µg g-1 (Appendix S2). When water was limiting, the addition of 50 μM Zn increased A and gs in a 67% and 73% as compared with 3 μM Zn, respectively. Moreover, in the last case this increment was significant. These results are in agreement with observed trends in morphological variables.
In the same way, KR-RA showed a similar behaviour as gas exchange rates. KR-RA decreased with increasing Zn concentration in LW seedlings and increases a 30% after 50 μM Zn. This may be a consequence of the morphological changes occurring in LW plants subjected to increased Zn availability. As a result, KR-RA values converged in HW and LW seedlings at root Zn concentration above ca. 1000 µg-1 (see Appendix S4 in Supporting Information). Similar results were observed when we analysed KR on a leaf area and a root length basis. Limitations to water flow through xylem vessels can increase the risk of cavitation by increasing water tension from roots to leaves (Sperry et al., 2002). However, some limitation to water flow may increase IWUE of plants by regulating water losses. This response would be beneficial to maintain a positive water balance, especially for plants living in water limited ecosystems (Vilagrosa et al., 2003a; Hernández et al., 2010).
The effect of heavy metals on IWUE has been scarcely studied, but an increase in IWUE can alleviate the effects of drought. We found that the increase in Zn load enhanced carbon assimilation per unit of water loss independently of the irrigation regime. This is remarkable, as A and gs behaved in quite different ways in both groups of seedlings.

Alleviation of Zn toxicity by low water availability
98
To what extent was the differential morpho-physiological response of LW seedlings to Zn a simple consequence of reduced Zn uptake? The relationships between root Zn concentration, on the one hand, and gs and KR, on the other, did not overlap in LW and HW seedlings (see Appendix S4 in Supporting Information). These results suggest that the response of seedlings to low water availability influence the response of Q. suber seedlings to Zn application.
5. Conclusion
Our results demonstrate that the combination of the increase in Zn load and moderate water stress did not induce a synergistic response in Q. suber seedlings, supporting our initial hypothesis. The combination of both stress factors, may contribute to alleviate their negative impact. On the one hand, low water availability prevented Zn accumulation, and probably buffered its negative effects. On the other hand, when water was limiting, intermediate Zn concentrations may have contributed to the improvement of traits as water use efficiency and root growth, buffering the effects of water limitation. Low water availability expanded the range of Zn concentrations where this heavy metal may have a positive effect on plant performance, shifting the toxicity level up. Our results suggest that legal and practical regulations of Zn levels (Appendix 4 of this Thesis) should take into account the water regime of each zone.
Acknowledgements
We thank Jose Huesca for their assistance in the lab, Banco de Semillas El Serranillo for the seeds used in this experiment and the University of Alicante for an FPU (Formacion del Profesorado Universitario) grant awarded to K. Disante. This research was funded by the Spanish Ministry of Science and Innovation (Project GRACCIE, Programa Consolider-Ingenio 2010 (CSD 2007-00067) and SURVIVE (CGL-2011-30531-CO2-02)) and Generalitat Valenciana (FEEDBACKS-PROMETEO/2009/006). E.I. Hernández thanks the University of Alicante for her FPU research fellowship. CEAM is supported by Generalitat Valenciana.

Capítulo IV
99
Supporting information
Appendix S1. Results obtained from MINTEQ2A. Equilibrated mass distribution between the dissolved, adsorbed, and multiple solid phases is shown. Data used to feed MINTEQ2A consists of total dissolved concentrations in mol/L of all anions and cations present in the nutrient solution (as described in the Materials and Methods section).
Ion DISSOLVED SORBED PRECIPITATED
mol/L % mol/L % mol/L %
Fe+2 3.70E-07 100 0.00E+00 0 0.00E+00 0
NH4+1 2.40E-09 100 0.00E+00 0 0.00E+00 0
NO3-1 2.58E-01 100 0.00E+00 0 0.00E+00 0
K+1 4.80E-04 100 0.00E+00 0 0.00E+00 0
PO4-3 4.49E-05 99.8 0.00E+00 0 9.74E-08 0.2
Mg+2 3.00E-04 100 0.00E+00 0 0.00E+00 0
SO4-2 3.00E-04 100 0.00E+00 0 0.00E+00 0
H3BO3 1.60E-06 100 0.00E+00 0 0.00E+00 0
MoO4-2 2.80E-09 100 0.00E+00 0 0.00E+00 0
Zn+2 3.00E-06 100 0.00E+00 0 0.00E+00 0
Cu+2 1.20E-08 100 0.00E+00 0 0.00E+00 0
Na+1 4.80E-04 100 0.00E+00 0 0.00E+00 0
Cl-1 8.00E-04 100 0.00E+00 0 0.00E+00 0
Ca+2 3.00E-04 100 0.00E+00 0 0.00E+00 0
I-1 6.00E-09 100 0.00E+00 0 0.00E+00 0
H+1 9.20E-05 100 0.00E+00 0 0.00E+00 0
H4SiO4 9.62E-05 100 0.00E+00 0 0.00E+00 0
Mn+2 5.03E-07 83.8 0.00E+00 0 9.74E-08 16.2
Percentage distribution of components among Type I and Type II (dissolved and
adsorbed) species:
Fe+2: 98.4% bound in species Fe+2
Zn+2: 80.8% bound in species Zn+2; 18.0% bound in species ZnNO3+

Alleviation of Zn toxicity by low water availability
100
Appendix S2. Results of Student-t test to evaluate the effects of drought on morpho-physiological traits of Quercus suber seedlings subjected to increasing Zn concentration. Foliar Zn:foliar Zn concentration, Root Zn:root Zn concentration, Fv/Fm:predawn maximum photochemical efficiency, Fo: minimum fluorescence, A: net photosynthesis; gs: stomatal conductance, KR-RA: root hydraulic conductance per root area, IAA: auxin concentration in leaves.
Zn dose (μM)
3 10 50 150
Variables t P t P t P t P
Foliar Zn 0.82 0.436 2.56 0.038 2.40 0.061 2.48 0.042
Root Zn 1.61 0.145 0.76 0.471 1.83 0.126 2.67 0.032
Root
biomass
1.27 0.228 1.37 0.196 0.31 0.125 4.10 0.001
Root
length
0.42 0.681 0.86 0.407 0.92 0.381 3.00 0.009
Fv:Fm 2.11 0.072 1.42 0.224 0.96 0.370 1.55 0.183
Fo 1.57 0.160 2.05 0.074 0.90 0.400 1.67 0.152
A 4.67 0.002 1.81 0.112 3.26 0.012 1.40 0.200
gs 4.92 0.002 1.62 0.150 2.67 0.028 2.11 0.068
KR-RA 5.49 0.001 2.36 0.043 0.03 0.974 0.80 0.444
IAA 0.49 0.642 0.71 0.506 1.17 0.306 2.73 0.026

Capítulo IV
101
Zn dose (μM)3 10 50 150
IWU
E (m
mol
CO
2 m
mol
H2O
-1)
0
1
2
3
4
5
6
HW LW
Appendix S3. Instantaneous water use efficiency of Quercus suber seedlings exposed to different levels of Zn availability and contrasting watering frequency. Bars are means and standard errors of N=5 seedlings.

Alleviation of Zn toxicity by low water availability
102
Root Zn concentration (μg g-1)
0 1000 2000 3000 4000 5000
g s (m
mol
m-2
s-1)
0,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
Root Zn concentration (μg g-1)
0 1000 2000 3000 4000 5000
K R-R
A (K
g s-1
MPa
-1 m
-2 x1
0-4)
2
4
6
8
10
12
14
16
18
HWLW
R2= 0.86F = 12.61P = 0.071
R2= 0.99F = 68.85P = 0.085
HWLW
R2= 0.94F = 32.28P = 0.030
R2= 0.99F = 1407.85P = 0.0188
y = (0.17x / (472.63 + x)) - 4.20x10-5x
y = 6.55 + (633.45 / x)
y = (9.52x /(179.19 + x)) - 0.0014x
y = (0.14 x 2460.64) / (2460.64 + x)
Appendix S4. Relationship between gs and root Zn concentration (A), and between KR_RA and root Zn concentration (B) in Quercus suber seedlings exposed to different levels of Zn availability and contrasting watering frequency. Models describing the relationship between variables and values of R2, F and P of each regression curve are shown.

Capítulo IV
103
Appendix S5. Average root diameter of 5-month-old Quercus suber seedlings grown under a range of Zn concentrations and contrasted irrigation regimes. HW and LW correspond to high and low watering frequency, respectively. Means and standard errors of N = 6-7 seedlings per treatment and results of a two-way ANOVA are shown.
Zn HW LW
3 0.85 ± 0.02 0.82 ± 0.02
10 0.95 ± 0.07 0.87± 0.04
50 0.91 ± 0.09 0.85 ± 0.02
150 0.93 ± 0.04 0.87 ± 0.05
FZn= 1.00; P= 0.401
FW= 3.00; P= 0.092
FZnxW= 0.13; P= 0.940


Discusión general


Discusión general
107
1. Zinc en especies Mediterráneas
1.1. Aproximación experimental y sensibilidad de las variables morfo-funcionales al aumento de la biodisponibilidad de Zn.
La exposición a niveles tóxicos de Zinc, promueve un complejo sistema de respuesta en los vegetales. Este sistema se complica cuando esta fuente de estrés se combina con otras, como la sequía, y especialmente, cuando la contaminación con Zn y la sequía se manifiestan en el marco de la infinidad de interacciones ecológicas de los sistemas naturales. Ante la dificultad de analizar sistemas complejos de forma holística, se hace necesario simplificarlos y, con ello, alcanzar un compromiso entre realismo y simplicidad constructiva. Una de las primeras decisiones que debimos tomar al plantear esta Tesis fue la relativa a la aproximación experimental. Se trataba de escoger el tipo de estudios que pudiera informarnos sobre la respuesta de los vegetales, manteniendo un nivel de realismo adecuado y evitando un exceso de ruido que pudiera dificultar el análisis de los procesos. Por esta razón, optamos por utilizar cultivos hidropónicos sin soporte y en arena. Por otra parte, otra decisión clave en este tipo de estudios es la selección de variables de respuesta. No todas las variables son igualmente sensibles, ni tampoco resulta deseable analizar todas y cada una de las respuestas morfo-funcionales a cada uno de los niveles de organización. En este apartado discutimos ambas cuestiones.
Consideraciones sobre la aproximación experimental
Los estudios hidropónicos con solución nutriente diluida resultan un buen método de imitación de la solución del suelo (Reichman, 2002) aunque suponen una simplificación de este sistema. Existe una serie de diferencias destacables entre ambos medios que podrían influir en la respuesta de la planta a un elemento tóxico: 1- ciertos factores ausentes en cultivo en solución, pero presentes en el suelo podrían aliviar el estrés por metal. Estos factores incluyen organismos de la rizosfera como micorrizas (Köhl y Lösch, 2004), y materia orgánica y arcillas que pueden retener el metal (Alloway, 1995). 2- En suelo, las concentraciones de metal pueden variar en función del espacio y tiempo. El suelo constituye un medio heterogéneo con posibles micro-sitios donde las raíces pueden estar en contacto o no con un determinado elemento. En un hidropónico donde la solución es

Capítulo V
108
renovada periódicamente, la concentración de un metal es relativamente constante en espacio y tiempo, exceptuando pequeñas diferencias generadas entre la rizosfera y el grueso de la solución. Entonces, por un lado las plantas en solución suelen ser más sensibles al metal al estar sus raíces en pleno contacto con él. Por otro lado, en el suelo las raíces deben explorar el medio continuamente en busca de agua y nutrientes, por lo tanto, es probable que en el suelo una mínima reducción en el crecimiento radicular se traduzca en un efecto amplificado sobre el estado global de la planta (Reichman, 2002). 3- Factores presentes en la naturaleza como herbivoría, competencia, patógenos y diferentes tipos de estrés podrían interaccionar de manera sinérgica o antagónica con el metal, modificando la capacidad de la planta para responder al mismo. 4- La concentración de nutrientes en la solución de un cultivo hidropónico también puede influir sobre la respuesta de la planta al metal. La absorción del metal puede ser menor en una solución concentrada, debido a la competencia entre los elementos presentes en el medio (Greger, 2004). Soluciones diluidas como las empleadas en los capítulos II, III y IV se asemejan más a la solución del suelo que soluciones nutrientes altamente fertilizadas. Además, soluciones diluidas disminuyen la posibilidad de interacción entre elementos como por ejemplo, la precipitación, que disminuiría la disponibilidad del elemento precipitado (Kabata-Pendias y Pendias, 2001).
Debido a lo anteriormente discutido, la gran heterogeneidad presente en condiciones de campo, hace que el cultivo hidropónico de plantas resulte un procedimiento adecuado para evaluar la respuesta de plántulas a la toxicidad por metal e identificar concentraciones umbrales. Si bien estos resultados no son directamente extrapolables al campo, ofrecen una primera visión de la respuesta de diferentes especies a un metal, permitiendo establecer comparaciones, identificar variables de respuesta adecuadas y establecer concentraciones tisulares umbrales.
El cultivo hidropónico en arena, es un medio intermedio entre el cultivo hidropónico en solución y el suelo. Factores presentes en este último como micorrizas, rizosfera, materia orgánica debido a la descomposición de raíces, disponibilidad hídrica variable, diferencias de pH (por ejemplo entre la rizosfera y el medio circundante) que pueden modificar la toxicidad de un metal, también pueden estar presentes en el hidropónico con arena (Köhl y Lösch, 2004).

Discusión general
109
En nuestro trabajo, para una misma concentración de Zn se observaron mayores efectos en las plántulas de Q. suber creciendo en solución respecto a las plántulas de la misma especie que crecieron en arena, lo cual es lógico si tenemos en consideración lo discutido anteriormente. Diversos estudios han evaluado la respuesta de la planta creciendo en diferentes medios, encontrando tanto diferencias como similitudes (Watson et al., 2003; Bernhard et al., 2005; Zornoza et al., 2010). Por ejemplo, en un estudio donde se evaluó la respuesta de los mismos clones de Salix al Cu y Ni en campo y en un sistema hidropónico, se observó una correlación significativa entre los resultados obtenidos en ambos medios (Watson et al., 2003).
Consideraciones sobre las variables de respuesta
No todas las variables morfo-funcionales muestran idéntico nivel de sensibilidad a la presencia de elevadas concentraciones de Zn. La figura 1 muestra la concentración de Zn acumulado en tejido foliar que induce una reducción del 35% en las diferentes variables morfo-fisiológicas evaluadas en Q. suber respecto al máximo valor observado (ver Apéndice II). No utilizamos una reducción del 50%, como es más habitual, ya que hubiera sido necesario extrapolar los valores en las curvas de respuesta. Como se puede observar, la concentración de Zn que indujo esta reducción varió entre los 30 y 384 µg g-1 de Zn en hoja, en función de la variable considerada.
La conductividad hidráulica expresada en función del área radicular (KR-RA) fue la variable más sensible a la aplicación de Zn. Este resultado resulta lógico ya que la KR-RA representa la conductividad hidráulica en raíz, órgano que entra en contacto con el metal en primer lugar y de forma más íntima. La reducción del 35% de esta variable pudo observarse a partir de 31 µg g-1 de Zn acumulado en tejido foliar. La reducción de la capacidad de transporte de agua por el sistema radical en respuesta al Zn podría deberse al bloqueo de los canales de agua (acuaporinas) y a la disminución del tamaño de vasos del xilema (Barceló y Poschenrieder, 1990; Poschenrieder y Barceló, 2004). A partir de 111 µg g-1 de Zn foliar, la conductancia estomática (gs) se redujo en un 35% respecto al valor observado en plantas no sometidas a este metal. Los mecanismos por los cuales el aumento de Zn puede inducir el cierre estomático incluyen inhibición de los canales transportadores de agua, una alteración en la proporción K/Ca en las células de guarda, o un aumento en la concentración de ácido abscísico (Tsalandzono et al., 1994;

Capítulo V
110
Marschner, 1995; Yang et al., 2004). Una reducción similar en la tasa fotosintética (A) se obtuvo a mayores concentraciones de Zn en hoja, cuando la concentración promedio de Zn alcanzó los 200 µg g-1, y probablemente se debió en parte a la disminución de gs. Cambios en A pueden estar también relacionados con efectos directos del Zn sobre la biosíntesis de la clorofila, sobre diversos componentes de la cadena de transporte de electrones, y sobre enzimas que participan en la asimilación de carbono (Balsberg-Påhlsson, 1989; Monnet et al., 2001; Myśliwa-Kurdziel et al., 2004).
A partir de 200 µg g-1se produjo un aumento del 35% en la concentración de la auxina IAA, y a una concentración de Zn foliar superior (252 µg g-1), observamos una reducción de la misma magnitud en la longitud radicular, lo que supone el primer cambio morfológico significativo. El ácido indol-3-acético (IAA) es la auxina natural más abundante y controla una amplia gama de procesos fisiológicos en plantas, incluyendo la dominancia apical, el tropismo, elongación de los brotes, formación de raíces laterales y diferenciación del tejido vascular (Aloni et al., 2006; Vanneste y Friml, 2009). Esta hormona participa también en la respuesta a factores de estrés abiótico, como sequía, metales pesados o temperatura (Elobied y Polle, 2012). Es más, otros estudios han relacionado la acumulación de esta auxina con un aumento de la densidad de raíces laterales tras la exposición a elevadas concentraciones de Cu en el medio (Lequeux et al., 2010; Pető el at., 2011).
Diversos trabajos describen cambios en la acumulación de biomasa radicular en respuesta a un aumento de Zn en la solución externa (Ruano et al., 1998; Reichman et al., 2001). En el caso de Q. suber, una reducción del 35% de la biomasa subterránea se obtendría a valores de Zn foliares superiores a los registrados en los experimentos descritos previamente (308 µg g-1 de Zn en hoja). Por lo tanto, la acumulación de biomasa radicular resultó menos sensible al aumento de Zn en solución que la mayoría de variables evaluadas. Un pequeño aumento del diámetro promedio radicular podría haber contribuido a la menor sensibilidad de la biomasa radicular de Q. suber en respuesta al Zn (Capítulo IV).
Asimismo, muchos trabajos han identificado a la fluorescencia de las clorofilas como un posible indicador fisiológico de toxicidad por metales, así como de otros tipos de estrés (Joshi y Mohanty, 2004). Sin embargo, en este trabajo fue la variable menos sensible a la aplicación del metal dentro del rango de

Discusión general
111
concentraciones evaluado. Una reducción de Fv:Fm equiparable al de las otras variables se produciría a concentraciones de Zn mayores a las evaluadas (384 µg g-1 de Zn en hoja), cuando procesos como la fotosíntesis serían prácticamente nulos. Esto sugiere una gran estabilidad del fotosistema II (PSII) a la exposición al Zn. La resistencia del PSII no es exclusiva de Q. suber ni de la respuesta al estrés por Zn, sino que parece ser una característica intrínseca del fotosistema, independientemente de la especie y factor de estrés evaluados (Methy et al., 1996; Vilagrosa et al., 2010). El efecto negativo del aumento de Zn sobre la eficiencia fotosintética pueden deberse al efecto del Zn sobre el centro de reacción del PSII o a una reducción en la transferencia de energía desde el complejo antena al centro de reacción (Ralph y Burchett, 1998).
CT35 (μgg-1)
0 50 100 150 200 250 300 350 400 450
a
b
c
d
e
f
g
KR-RA
gs
A
IAA
Longitud radicular
Biomasa radicular
Fv:Fm
Figura 1. Concentración de Zn en tejido foliar (expresado en µg g-1) que induce una reducción del 35% en el valor de diversas variables morfo-fisiológicas respecto al valor máximo observado. Las barras representan el promedio y el error estándar. Fv:Fm: eficiencia fotoquímica máxima al alba, A: fotosíntesis; gs: conductancia estomática; KR-RA: conductancia hidráulica por área radicular; IAA: concentración de la auxina ácido indolacético en hojas.
Sintetizando, el incremento en la concentración foliar de Zn desencadenó en primer lugar una respuesta fisiológica, que incluyó la disminución del transporte de agua y del intercambio de gases. A medida que la concentración interna de Zn

Capítulo V
112
aumentó, se produjeron cambios en la morfología radicular y en la acumulación de biomasa. Por último, se redujo la eficiencia fotosintética, a concentraciones internas de Zn más elevadas, en lo que se podría considerar un colapso casi completo de las funciones vegetales. La sensibilidad de las variables fisiológicas al Zn sugiere la importancia de su evaluación en estudios sobre toxicidad por metales.
En los diferentes trabajos realizados en esta tesis hemos considerado a priori que el umbral de toxicidad se había superado si la concentración foliar de Zn superaba los 200 µg g-1. Este valor se fijó en base a las referencias bibliográficas disponibles para diferentes especies (Balsberg Påhlsson, 1989; Kabata-Pendias and Pendias, 1992; Hagemeyer et al., 2004). Sin embargo, los valores umbral tienen carácter orientativo ya que pueden variar para una misma especie dependiendo de la variable empleada para evaluar la respuesta de la planta, como demuestran nuestros estudios.
Esta diferenciación entre variables de respuesta se debe analizar con precaución. Por un lado, la escala de respuesta de las diferentes variables no es la misma y por otro, la respuesta de estas variables no siempre es lineal. Por ello, una disminución de un 35% de una de las variables, no es directamente trasladable, en términos funcionales, con una disminución de la misma magnitud en otra. Por ejemplo, la reducción del cociente Fv:Fm de un 35%, hasta un valor de 0.46, sería indicador de un estrés muy intenso, puesto que esta variable se mantiene en valores relativamente altos hasta que se supera un umbral de estrés elevado, momento a partir del cual desciende abruptamente (Vilagrosa et al., 2010). Por el contrario, una reducción de 35% de la conductancia estomática o la tasa fotosintética ocurren de forma mucho más frecuente en la vida de algunas plantas, especialmente en ecosistemas Mediterráneos, y no se pueden considerar indicadoras de un elevado nivel de estrés. No obstante, esta diferenciación entre variables de respuesta pone de manifiesto la necesidad de llevar a cabo una evaluación integral del estado de la planta a la hora de evaluar el impacto de la contaminación con metales (y, probablemente, de otras fuentes de estrés), y la necesidad de seleccionar adecuadamente las variables de respuesta en este tipo de estudios. En otras palabras, cuando se pretenda establecer niveles de toxicidad para Zn, habría que referirse a la concentración foliar de Zn a partir de la cual se induce una respuesta rápida (cierre estomático, por ejemplo), a la que se inducen cambios morfológicos, o a partir de la cual se generan daños más severos.

Discusión general
113
1.2. Modelo conceptual: morfología y fisiología de plántulas expuestas al Zn
Ante un factor estresante externo las plantas responden de una manera global, con el fin de adaptarse a la nueva condición (Larcher, 1995). En este contexto, cada cambio inducido afecta a numerosos procesos desde la raíz hasta las hojas. A partir de los resultados obtenidos en los diferentes experimentos, y de las consideraciones presentadas en el apartado anterior, hemos generado un modelo que integra los múltiples efectos del incremento de Zn sobre las diferentes variables morfo-fisiológicas evaluadas (Fig. 2). El modelo destaca la relación entre las distintas variables consideradas, y pone especial énfasis en describir cómo los cambios producidos afectan a la capacidad de la planta de captar y hacer uso del agua. La mayor parte de las variables ilustradas han sido discutidas a lo largo de esta tesis y por este motivo las mencionaremos haciendo referencia a los capítulos donde se analizaron.
De acuerdo a la Fig. 1, la conductancia de agua por las raíces es muy sensible a pequeños incrementos en la concentración externa de Zn (Capítulo IV). Si bien muchos trabajos han sugerido que el Zn puede afectar la capacidad de las plantas de transportar agua, según nuestro conocimiento, la relación entre concentración de Zn y la KR-RA no ha sido cuantificada. Es importante destacar que aunque en el apartado anterior nos centramos en el análisis de KR-RA , los datos de KR-LA, KR-RL (conductancia hidráulica expresada en función del área foliar o longitud radicular, respectivamente) muestran un comportamiento similar en respuesta al Zn. Por lo tanto, en el modelo presentado en la Fig. 2 hablamos de Kh para referirnos de un modo general, a la capacidad de transporte de agua por el sistema radical.
Una menor conductividad hidráulica supone una menor capacidad de transporte de agua, lo cual podría no tener consecuencias negativas para la planta en condiciones de alta disponibilidad hídrica. Pero bajo condiciones de alta demanda evaporativa o estrés hídrico podría conducir a potenciales hídricos más negativos para mantener la conductividad y el suministro de agua a las hojas y así aumentar el riesgo de cavitar (Vilagrosa et al., 2003b). Otros trabajos han sugerido que la eficiencia del xilema para el transporte de agua puede limitar la transpiración y la fotosíntesis (Hubbard et al., 2001). Varios trabajos han relacionado alteraciones en la conductancia hidráulica con cambios en la regulación estomática para prevenir la desecación, manteniendo el potencial hídrico del xilema por encima del umbral mínimo (Sperry et al., 2002; Vilagrosa et al., 2003b). Como se observó en la Fig. 1 y

Capítulo V
114
en los Capítulos III y IV, gs resultó ser la segunda variable más sensible, disminuyendo bruscamente con el incremento de Zn el medio.
Figura 2. Modelo conceptual describiendo los efectos del Zn sobre las plantas y su posible interacción con la economía hídrica. Los cuadros en color azul y lila muestran respectivamente, efectos inducidos por Zn y compuestos que participan en la coordinación de la planta completa en la respuesta al estrés. En línea continua se muestran los efectos encontrados en esta tesis y en línea discontinua efectos descritos en la bibliografía pero no evaluados en este trabajo. Los recuadros de color verde y naranja representan consecuencias de la acción del Zn que favorecen o disminuyen la economía hídrica de las plantas. SRL: longitud radicular específica, Fv:Fm: eficiencia fotoquímica máxima al alba, Fo: fluorescencia mínima, A: fotosíntesis; gs: conductancia estomática, Kh: conductancia hidráulica por área radicular, IAA: concentración de auxina IAA en hojas.
La asimilación neta de CO2 y la transpiración (E) también fueron afectadas por Zn debido, en parte, a la disminución de gs. La magnitud de la disminución de E fue menor que la de la fotosíntesis. Por lo tanto, la WUEi (que juega un papel importante en la respuesta de la planta a la sequía) aumentó en repuesta al

Discusión general
115
aumento del metal en el medio (Capítulos III y IV). A su vez, la reducción de la asimilación foliar de carbono podría limitar el crecimiento vegetal y la acumulación de sustancias de reserva para sobrevivir periodos de balance negativo. Bajo el epígrafe ‘daños en componentes fotosintéticos y cambios en la nutrición mineral’ se incluyen una serie de cambios inducidos por Zn que tendrían como resultado final un efecto negativo sobre los procesos fotosintéticos.
En el Apéndice III presentamos resultados sobre el efecto del Zn sobre la nutrición mineral en Q. suber. El aumento de Zn indujo una reducción significativa en la concentración foliar de Mn y Mg. Otros trabajos han mostrado que el Zn interfiere con la captación de diversos nutrientes (Ruano et al., 1987; Kabata-Pendias y Pendias, 2001). El Mn es necesario para la fotólisis del agua durante la fotosíntesis. La disminución de la concentración foliar de Mn de manera paralela al aumento de Zn podrían favorecer la substitución del Mn por Zn en el PSII, impidiendo la fotólisis del agua y por ende inhibiendo la fotosíntesis (Van Asche y Clijsters, 1986, Marschner, 1995). De un modo similar, un aumento en la proporción Zn/Mg favorecería al primero en la competencia entre ambos por formar complejo con el enzima Rubisco, lo cual, disminuiría la afinidad del enzima por el CO2 (Marschner, 1995; Mateos-Naranjo et al., 2008).
En conjunto, los datos presentados en los Capítulos III y IV ilustran la reducción de la eficiencia fotosintética, y sugieren fotoinhibición permanente inducida por altas dosis de Zn (ver medidas de Fv:Fm, Fo, Am, фm; Long et al., 1994; Galmes et al., 2007; Mateos-Naranjo et al., 2008). Otros trabajos han establecido una correlación entre fotoinhibición y niveles de compuestos derivados del alfa-tocoferol (Munné-Bosch, 2005). Este compuesto es la principal forma de la vitamina E en cloroplastos, donde contribuye a la preservación de un estado redox adecuado en los cloroplastos y a mantener la estructura y función de la membrana tilacoide durante el desarrollo de la planta y en la respuesta al estrés (Munné-Bosch y Alegre, 2002; Munné-Bosch, 2005). Estos datos podrían explicar el aumento de alfa-tocoferol observado (Apéndice I).
El apartado del modelo conceptual relativo a ‘metabolitos secundarios’ engloba los compuestos descritos y discutidos en detalle en el Apéndice I (aminoácidos, azúcares, ácidos orgánicos, lípidos, etc). Es importante destacar que, al igual que este epígrafe incluye compuestos pertenecientes a distintos grupos de compuestos, las funciones que éstos desempeñan son muy variadas. Por ejemplo,

Capítulo V
116
algunos de ellos podrían actuar como osmolitos y el incremento de su concentración podría favorecer la conservación del contenido hídrico tisular (azúcares, ácidos orgánicos; Nilsen y Orcutt, 1996; Romero et al., 2004). Otros actuarían en funciones de defensa contra el estrés oxidativo (alfa-tocoferol), o podrían participar en rutas de señalamiento. Así, cambios en la concentración de azúcares pueden activar diferentes rutas de transducción de señales que en último término se integran para regular la expresión génica (Chaves y Pereira, 2004). La auxina IAA, además de actuar como regulador del crecimiento y desarrollo vegetal, es un componente esencial de las vías que median respuestas de las plantas al estrés abiótico (Elobied y Polle, 2012). El aumento observado en su concentración tras la exposición de las plántulas al Zn (Capítulo IV) sugiere un papel importante de la IAA en la coordinación de la respuesta de la planta a este metal.
Cambios en la morfología de la planta y en la acumulación de biomasa inducidos por Zn inciden directamente en su capacidad de captar recursos. En los Capítulos II, III y IV observamos que la disminución de la longitud radicular fue un síntoma generalizado de toxicidad por Zn en distintas especies Mediterráneas, como se ha observado en otras especies (Woolhouse, 1983; Ruano et al., 1988, Reichman et al., 2001).
En los ecosistemas Mediterráneos, la sequía y la desecación del suelo son los principales limitantes para el establecimiento de las plántulas. Sistemas radiculares profundos podrían mejorar la captura de agua e incrementar de este modo, su supervivencia (Padilla y Pugnaire, 2007). Esto significa que un sistema radicular superficial, debido a la inhibición de la elongación radicular inducida por Zn podría restringir el acceso al agua y nutrientes, limitando así, su capacidad de establecimiento y supervivencia en medios xéricos y poco fértiles.
Además de la longitud radicular, en el Capítulo II comprobamos que el aumento de Zn puede alterar la distribución de la longitud radicular por clase diamétrica así como el número de ápices radiculares. Aunque la magnitud de los cambios fue dependiente de la especie, estos cambios sugieren que la arquitectura radicular global se ve alterada por el aumento de Zn en el medio, y que en especies como Q. suber, P. pinaster y P. halepensis, la disminución de la longitud radicular se ve acompañada por un descenso del número de ápices radiculares, lo que podría dificultar aún más la exploración del suelo y la captación de recursos.

Discusión general
117
Foto 1. Plántulas de Pinus pinea sometidas a dosis crecientes de Zn (0.078, 5, 10, 20 y 90 µM) de izquierda a derecha y de arriba abajo, respectivamente (Capítulo II).
En los experimentos aquí presentados, (Capítulos II, III y IV) la magnitud del efecto del Zn sobre la acumulación de biomasa fue dependiente de la especie, e incluso de la edad de la planta. Procesos como la suberización o la lignificación no sólo podrían inducir cambios en la acumulación de biomasa, sino que además influirían en la entrada y en la salida de agua de las raíces (Barceló y Poschenrieder, 1990). Aunque no observamos indicios de cambios en la suberización o la lignificación en Q. suber, no podemos descartarlos en el resto de especies.
Los cambios producidos en la longitud radicular y en la biomasa radicular se tradujeron en cambios en la longitud radicular específica (SRL; Capítulos II, III y

Capítulo V
118
IV). La relación negativa entre Zn y SRL podría explicarse debido a la mayor sensibilidad al metal de la longitud radicular que de la biomasa. Esto no es extraño ya que numerosos estudios han demostrado que la elongación celular es uno de los principales procesos afectados por metales pesados (Woolhouse, 1983). Se ha propuesto el cociente entre longitud radicular y la biomasa radicular como un índice de beneficio y coste radicular (Fitter, 1991). La longitud radicular sería proporcional a la adquisición de recursos (beneficio) y la biomasa radicular sería proporcional a la construcción y mantenimiento (coste) (Eissenstat y Yanai, 1997). Es decir, la disminución de SRL inducida por una alta concentración de Zn implicaría una disminución del volumen de suelo explorado por unidad de biomasa invertida. O dicho de otra manera, la relación entre beneficio obtenido en relación a los recursos invertidos, disminuiría. Además, se ha sugerido que cambios en SRL podrían alterar la conductancia hidráulica (Eissenstat, 1991). Este es un ejemplo más de cómo cambios puntuales inducidos por Zn pueden influenciar otras variables, reajustando finalmente, la respuesta general de la planta.
2. Zn y disponibilidad hídrica
2.1. La combinación de Zn y escasez hídrica. Relaciones causa-efecto.
Como se ha mencionado a lo largo de esta tesis, en la naturaleza las plantas están expuestas a diversos factores de estrés que pueden actuar de manera combinada, como por ejemplo, disponibilidad hídrica, alta radiación, temperatura, etc. (Larcher, 2003). La respuesta de la planta a la combinación de dos o más factores de estrés dependerá de los factores involucrados y de la severidad del estrés. En esta tesis hemos evaluado dos situaciones posibles en las que interaccionan diferentes concentraciones de Zn y disponibilidades hídricas contrastadas en el medio de cultivo.
Presencia consecutiva de Zn y estrés hídrico
Como se ha descrito a lo largo de esta memoria, la conductancia estomática resultó ser una variable muy sensible al incremento de Zn en la solución del suelo y, como consecuencia del cierre estomático, se produjo la disminución de la transpiración. Ante un periodo corto de sequía, la reducción de la pérdida de agua

Discusión general
119
por transpiración tuvo un efecto positivo sobre la capacidad de conservación del contenido hídrico, como demostraron los valores de RWC.
Sin embargo, a largo plazo la disminución de la longitud radicular y de SRL por aumento de Zn conllevaría una reducción del volumen de suelo explorado por las raíces (Ostonen et al., 2007). Además, concentraciones altas de Zn suelen tener un impacto negativo en la asimilación de carbono, que es el soporte del crecimiento vegetal.
Por lo tanto, a largo plazo la mayor capacidad de conservación de agua posiblemente no compensaría los efectos negativos del aumento de Zn y la plántula no podría mantener su crecimiento. Hay que tener en cuenta que la verificación de esta hipótesis requeriría el establecimiento de condiciones que eviten el confinamiento de las raíces a largo plazo.
Presencia simultánea de Zn y estrés hídrico moderado
En este caso, el aumento de Zn en los tejidos y el descenso de la transpiración ejercieron una regulación mutua: como consecuencia de la reducción de la transpiración por efecto del Zn, la absorción de este elemento fue limitada, al ser el flujo de masas el principal mecanismo de transporte de este elemento desde el suelo hacia la raíz. Por lo tanto, los efectos nocivos del metal fueron amortiguados cuando la frecuencia de riego se redujo. Cuando el agua fue limitante, el aumento de Zn en el medio y en los tejidos supuso un aumento significativo en gs y en la biomasa radicular, acompañados por una tendencia no significativa similar en otras variables. Es importante destacar que, en primer lugar, estos efectos no pueden ser explicados únicamente por una menor concentración de Zn tisular (como consecuencia de la menor frecuencia de riego). En segundo lugar, en ningún caso la combinación de ambos factores indujo una mejora en el estado general de la planta (evaluado en función de medidas ecofisiológicas), respecto a la situación de plantas bien regadas y con una concentración de Zn en el medio baja.
Aunque la aplicación de los factores de estrés se haya realizado de manera consecutiva o simultánea, no encontramos un efecto sinérgico de ambos estreses, excepto a escala de metabolito (glucosa, ácido cítrico, ramnosa). Por el contrario, la combinación de niveles crecientes de Zn y disponibilidades hídricas contrastadas representó una situación compleja, que no es simplemente la suma

Capítulo V
120
de dos factores estresantes. A nivel metabólico, cada factor desencadenó una respuesta independiente del otro factor, mientras que a escalas superiores (fisiología y morfología), para la mayoría de las variables evaluadas, la respuesta a un factor estuvo condicionada por la presencia del otro factor estresante.
2.2. Respuestas comunes a dos factores de estrés actuando de forma aislada
Si bien el carácter y magnitud de la respuesta de las plantas a un factor de estrés, dependerá del tipo de estrés, y de la intensidad y duración del mismo, es posible establecer correspondencias entre los distintos tipos de respuesta. Para esto, es necesario considerar los efectos de los factores de estrés (Zn y disponibilidad hídrica) actuando individualmente. El efecto de la disminución de disponibilidad hídrica lo evaluamos en los Capítulos III, IV y en el Apéndice I. En el Capítulo III la exposición de plántulas Control de Q. suber (tratadas 3 µM Zn) expuestas a un periodo corto de estrés hídrico, nos permitió establecer la respuesta a un estrés hídrico severo, mientras que en los Capítulos IV y en el Apéndice I la comparación entre plantas HW y LW expuestas a 3 µM Zn, mostró la respuesta de plántulas de Q. suber a un estrés hídrico moderado, en ausencia de otro factor de estrés. De un modo similar, evaluamos los efectos del Zn en ausencia de otros factores en los Capítulos III y IV y en el Apéndice I al considerar la respuesta de plántulas expuestas a concentraciones crecientes de Zn antes del periodo de sequía (Capítulo III) y de las plántulas HW (Capítulos IV y Apéndice I).
Un periodo corto de estrés hídrico indujo un descenso significativo en el intercambio de gases (A, gs y E) y en Fv:Fm (Capítulo III). Sin embargo, en el primer grupo de variables la reducción fue casi inmediata, mientras que los valores de Fv:Fm se mantuvieron prácticamente sin cambios hasta el fin del ciclo de sequía. Como se ha comentado anteriormente, los valores de Fv:Fm suelen disminuir a partir de un potencial hídrico umbral, cuando el estrés hídrico es muy severo y el intercambio de gases prácticamente nulo (Mhety el at., 1996; Lawlor y Tezara 2009; Vilagrosa et al., 2010). Igualmente, los valores de A y gs disminuyeron significativamente en plantas con bajo riego (3 µM Zn LW) mientras que el descenso de Fv:Fm respecto a las plantas control fue sólo marginal. Estos datos concuerdan con un estrés hídrico moderado más que con un estrés hídrico severo, el cual no fue el objeto de estudio en el Capítulo IV.

Discusión general
121
El aumento de Zn tuvo un efecto negativo en las variables citadas (A, gs, E y Fv:Fm; Capítulos III y IV). Es decir, el intercambio de gases y la eficiencia fotosintética muestran una respuesta similar a la disminución de disponibilidad hídrica y al incremento de Zn en el medio.
Del mismo modo, la conductividad hidráulica descendió significativamente en ambos casos, tanto tras la aplicación de Zn como por la disminución de la frecuencia de riego (Capítulo IV). La pérdida de conductividad hidráulica en respuesta a una menor disponibilidad hídrica es un proceso descrito a menudo en la bibliografía (Lo Gullo et al., 1998; Steudle, 2000; Valladares et al., 2004).
En cuanto a la morfología, mientras que el aumento de Zn provocó una disminución notable de la longitud radicular (Capítulos III y IV), un descenso en la frecuencia de riego no redujo esta variable de forma significativa (Capítulo IV). El déficit hídrico puede inducir un aumento transitorio en la longitud radicular total, una respuesta que permitiría a la planta aumentar la captación de agua (Medrano y Flexas, 2004; Chiatante et al., 2005). Según otros estudios, en caso de estrés hídrico severo o de larga duración, el crecimiento radicular puede disminuir significativamente debido a una alteración del balance hormonal, a la reducción de la fotosíntesis, a un menor potencial de turgencia y a la muerte de raíces finas (Chapin, 1991; Chiatante et al., 2005, Nielsen y Orcutt, 1996).
La inhibición de la longitud radicular ha sido reconocida como una respuesta morfológica inducida por estrés o SIMR por sus siglas en inglés (Potters et al., 2007 y 2009). Según estos autores, este cambio constituye una respuesta morfológica genérica a condiciones de estrés crónico o moderado. Entre los factores abióticos que inducen inhibición de la longitud radicular se encuentran diversos metales pesados, el herbicida paraquat, peróxido de hidrógeno y bajas concentraciones de fosfato en la rizosfera, entre otros.
A diferencia de lo ocurrido con la longitud radicular, el diámetro medio de las raíces fue sensible a la disponibilidad hídrica, pero no varió significativamente con el aumento de Zn. Las raíces de las plantas LW fueron más finas que las raíces de las que crecieron a un potencial hídrico mayor (Capítulo IV).
La biomasa foliar es habitualmente muy sensible a la presencia de metales pesados y a la disponibilidad hídrica (Reichman et al., 2001; Vilagrosa et al., 2003; Valladares et al., 2004). Cambios en la frecuencia de riego no indujeron cambios

Capítulo V
122
sustanciales en esta variable (Capítulo IV), mientras que la magnitud del efecto del Zn sobre la misma dependió de la edad de la planta (Capítulos II, III y IV). La biomasa radicular mostró un comportamiento similar, aunque en este caso la diferencia de disponibilidad hídrica se tradujo en una reducción, no significativa, del 17% de esta variable (Capítulo IV).
El estudio de la respuesta metabólica al incremento de Zn y a la frecuencia de riego, permitieron diferenciar metabolitos comunes a ambos tipos de estrés (glucosa, ramnosa, ácido cítrico, hexosa), compuestos cuya respuesta fue específica a la disponibilidad hídrica (ácido quínico, varios hidratos de carbono), y finalmente metabolitos sensibles exclusivamente a la aplicación de Zn (ácido málico, aspartato, ácido láurico, beta-D-methylfructofuranosido).
Los cambios en el metabolismo, fisiología y desarrollo comentados anteriormente son desencadenados por muchas señales primarias, pero también por señales secundarias. En este punto las fitohormonas desempeñan un papel fundamental (Chaves y Pereira, 2004). El aumento de concentración foliar de IAA con el aumento de Zn (plantas HW) contrasta con la ausencia de cambios entre plantas con riegos bien diferenciados y expuestas a 3 µM Zn (Capítulo IV). Esto sugiere que IAA podría mediar la respuesta al Zn, pero no parece relevante en la transducción de señales en respuesta a un descenso de la disponibilidad hídrica en plántulas de Q. suber.
Entonces, ¿podemos extraer conclusiones sobre respuestas comunes y específicas a la reducción de la disponibilidad hídrica y al aumento de Zn en el medio? A modo de síntesis, a nivel bioquímico se produjeron tanto cambios similares como diferentes a ambos factores de estrés. Mientras que los cambios fisiológicos fueron similares, los cambios morfológicos dependieron de la variable evaluada.
El estudio de un posible sistema centralizado de respuesta al estrés como sugirió Chapin (1991) debería contemplar posibles condiciones de referencia. Por ejemplo, el cierre de estomas es presumiblemente una variable sensible, mientras que la fluorescencia de clorofilas probablemente se mantiene hasta un valor umbral donde disminuye abruptamente. Entonces, ¿qué ocurre con la fisiología, bioquímica y morfología de las plántulas sometidas a diferentes tipos de estrés en el momento en que se produce el cierre de estomas y en el momento en que la fluorescencia de clorofilas disminuye?

Discusión general
123
Además, es lógico pensar que bajo condiciones de estrés muy severo, independientemente del tipo de estrés, las plantas responderán de manera similar: reducirán su crecimiento. En casos extremos, podrían no sobrevivir. Por el contrario, si las condiciones de estrés son moderadas o leves, la magnitud de la respuesta variará dentro de un rango más amplio que incluye la no-respuesta, efectos positivos y efectos negativos.
Por lo tanto, la existencia de una respuesta general a ambos factores, Zn y escasez hídrica, sería cierta para diferentes variables y diferentes niveles de organización. Por ello, de acuerdo a los resultados obtenidos en esta Tesis, no se puede hablar de un patrón de respuesta común (en el sentido estricto de la palabra) al aumento de Zn y a una baja disponibilidad hídrica.
Si consideramos nuestros resultados y otros trabajos previos podríamos concluir que los procesos moleculares y bioquímicos desencadenados por una condición de estrés específica pueden diferir de aquéllos que se activan por una señal ambiental ligera o marcadamente diferente. Sin embargo, las distintas vías iniciadas por un factor de estrés podrían conectarse entre ellas para desarrollar fenotipos en esencia diferentes pero con características similares (Mittler, 2006; Potters et al., 2009).
3. Caracterización experimental de la tolerancia al Zn
3.1 Consideraciones sobre la relación entre adaptación a zonas áridas y umbrales de toxicidad al Zn.
En el Capítulo II evaluamos si las especies de zonas áridas serían más resistentes al Zn que especies típicas de zonas húmedas. Por su elevado grado de ajuste, la relación obtenida, mayor tolerancia al metal en especies de zonas húmedas, resulta particularmente interesante. Un experimento realizado en el marco de uno de los proyectos de investigación en el que se encuadra esta tesis, evaluó la tolerancia de especies Mediterráneas a tres metales diferentes (Fuentes et al., 2007b). En este trabajo se estimaron las concentraciones internas que inducen una reducción del 10% en la biomasa. Respecto al Zn, aunque no pudieron calcularse los índices para todas las especies, no se observó una relación similar a la hallada en este trabajo entre la distribución de cada especie y la reducción en biomasa.

Capítulo V
124
Hay que destacar que existen varias diferencias entre estos dos estudios. En el trabajo de Fuentes et al. (2007b) podemos ver que: (i) el rango de distribución de las especies utilizadas es más reducido, (ii) la variable respuesta utilizada fue la biomasa, mientras que en esta tesis se descartó esta variable por no ser idónea (ver Capítulo II), y (iii) los efectos negativos del Zn fueron en general, más leves. Esto nos lleva a preguntarnos cuál o cuáles son las causas de la relación presentada en el Capítulo II ¿Es posible que características vegetales asociadas a la adaptación a zonas secas puedan reducir la capacidad de las plantas de soportar la presencia de Zn? Si fuera así, estas adaptaciones deberían ser genéticas ya que en un cultivo hidropónico la disponibilidad hídrica no es un limitante. ¿Qué mecanismos podrían entonces explicar este trade-off ecológico a escala interespecífica? Para responder esto habría que realizar un ensayo similar incluyendo mayor diversidad de especies de procedencias contrastadas y con diferentes estrategias de resistencia al déficit hídrico y evaluar si distintas características asociadas con la adaptación a la sequía, por ejemplo, eficiencia del uso del agua, conductancia estomática máxima, conductividad hidráulica, área foliar específica, tamaño de los vasos del xilema, etc. podrían explicar la tolerancia al Zn.
Además, cabe la posibilidad de que las plantas adaptadas a zonas secas lo estén también a suelos predominantemente carbonatados (Fuller y Tucker, 1977), en los que la disponibilidad de metales es comúnmente baja (Martinez y Motto, 2000). Por ello, el contacto con elevadas concentraciones de Zn tendría un efecto particularmente pernicioso en estas especies.
Asimismo, hay que considerar que la especie más mesófila de las estudiadas (Q. suber), es una reconocida calcífuga (Serrasolses et al., 2009), que podría estar adaptada a suelos que suelen presentar mayor disponibilidad de metales que el resto de especies. Ello podría suponer la presencia de una serie de mecanismos de exclusión o de tolerancia a diversos elementos, entre ellos el Zn.
Nuestros resultados, basados en un número limitado de especies, no permiten interpretar las causas de este aparente trade-off evolutivo entre respuesta a la sequía y respuesta a Zn sin entrar en el terreno de la especulación. Sin embargo, la consistencia de la relación sugiere la necesidad de profundizar más en esta cuestión. Por otra parte, habría que considerar otras posibles explicaciones a la relación presentada en el Capítulo II (Fig. 3).

Discusión general
125
Peso semilla (g)
0 1 3 4 5
EC50
-RG
R (μ
M Z
n)
0
5
10
15
20
25 Tamaño medioTamano mínimoTamano máximo
P. pineaP. pinaster
P. halepensis
R. alaternus
Q. suber
EC50
-L (μ
M Z
n)
0
5
10
15
20
25
30
Tamaño medioTamano mínimoTamano máximo
P. pineaP. pinaster
P. halepensisR. alaternus
Q. suber
Figura 3. Relación entre el peso medio de la semilla y el EC50 calculado en función de la longitud radicular total (izquierda) y en función del crecimiento relativo de la raíz principal (derecha). La especie Tetraclinis articulata no se incluye en las gráficas ya que no se pudo obtener datos sobre peso mínimo, medio y máximo de semilla de la misma.
En el momento de comenzar el tratamiento con Zn, las plántulas de todas las especies habían desplegado sus cotiledones y ya poseían hojas. Si bien el tiempo necesario para alcanzar la total independencia de las reservas de los cotiledones se desconoce para la mayoría de las especies (Bonfil, 1998), a partir del momento en que salen las primeras hojas aumenta la dependencia nutricional del medio externo, el cual contenía concentraciones crecientes de Zn en una solución

Capítulo V
126
nutriente diluida (Capítulo II). Si aceptamos como válida la relación entre tamaño de semilla y tolerancia al Zn sugerida por la Fig. 3, esto podría ser consecuencia de la presencia de reservas nutricionales aportadas por las semillas antes de la exposición al metal. Un contenido menor de reservas nutricionales podría inducir una mayor dependencia del medio circundante, el cual contenía Zn.
Esta hipótesis no puede ser aceptada o rechazada en base a los experimentos realizados en este trabajo. Muchos estudios han intentado relacionar el tamaño de la semilla con diferentes aspectos relacionados al establecimiento, competencia, adaptación a nichos umbríos etc., de las plántulas emergentes (Coomes y Grubb, 2003; Matilla, 2004). Sin embargo, a nuestro conocimiento, ningún estudio ha evaluado la importancia del tamaño de la semilla en la respuesta de las plántulas a metales pesados. Al analizar la Fig. 3 hay que considerar el peso de la especie Q. suber en esta relación, ya que este punto parece dominar la tendencia. Si eliminamos esta especie de las gráficas, en el caso de la gráfica de la derecha (EC50-RGR vs. peso de la semilla), podríamos ajustar los puntos a una función exponencial (R2= 0,71; F= 7,32; P= 0,073). Por el contrario, en el caso de EC50-L en función de peso de la semilla, no existe una relación significativa entre estas variables. Sin embargo, al igual que ocurre con la relación obtenida entre el área de distribución y la respuesta a Zn, la relación obtenida entre tamaño de semilla y sensibilidad a Zn sugiere la necesidad de más estudios en este sentido.

Conclusiones


Conclusiones
129
Conclusiones generales
En los diferentes Capítulos de esta tesis he ido desgranando las conclusiones parciales de cada estudio. Atendiendo al conjunto de experimentos, presento de forma sucinta las Conclusiones generales de la Tesis.
• Las especies leñosas Mediterráneas resultaron muy sensibles al aumento de Zn en el medio, y esta sensibilidad podría estar relacionada con la vulnerabilidad a la sequía.
• Las respuestas morfo-fisiológicas de las especies leñosas mediterráneas a Zn fueron características de cada especie. Quercus suber fue la especie más resistente, probablemente debido a su capacidad para excluir Zn en la etapa de plántula, mientras que Tetraclinis articulata fue la más sensible. Altas tasas de translocación se correlacionaron con respuestas morfológicas más marcadas a este metal pesado.
• Ninguna de las especies estudiadas mostró un fuerte potencial para la fitorremediación, aunque algunas de estas especies por su resistencia, podrían ser utilizadas para la restauración ecológica de sitios contaminados.
• La adición de pequeñas cantidades de Zn no tuvo un efecto positivo significativo en la respuesta morfo-fisiológica de las plántulas de Quercus suber a la sequía El aumento de la concentración de Zn afectó negativamente al desarrollo de las plantas. Las respuestas se produjeron a diferentes valores umbral para las diferentes variables. Nuestro estudio ha permitido identificar variables de respuesta sensibles a pequeños incrementos de Zn en el medio, como la conductancia estomática, y otras variables de respuesta cuyo valor sólo se vio modificado apreciablemente ante concentraciones foliares de Zn elevadas, como la eficiencia fotosintética.
• En conjunto, nuestros datos sugieren la necesidad de revisar la normativa actual sobre metales pesados en suelo, atendiendo a posibles factores

Conclusiones
130
ambientales que puedan provocar una disminución o un aumento de la sensibilidad de las plantas a la presencia de metales pesados en suelos.
• El presente trabajo analiza mediante distintos enfoques la respuesta integral de la planta al Zn y la interacción con la disponibilidad hídrica. Los efectos observados incluyen respuestas resilientes, susceptibles de recuperar su estado inicial y otras respuestas, cuya reversibilidad debería analizarse en experimentos a largo plazo. La interacción entre Zn y escasez hídrica no mostró efectos sinérgicos (i.e., intensificación de la respuesta como consecuencia de la combinación de ambos tipos de estrés), independientemente de si la aplicación de los dos factores se realizó de manera secuencial o simultánea. Por el contrario, la compleja respuesta de la planta a un factor de estrés podría contribuir a mitigar los efectos negativos del otro. En este sentido la conductancia estomática desempeña un papel fundamental. Se trata de una variable muy sensible al estrés hídrico y a altas dosis de Zn. La disminución de la conductancia estomática, por un lado, limita la absorción de Zn y sus efectos tóxicos y por otro, limita la pérdida de agua por transpiración, favoreciendo la conservación del contenido hídrico.
• Nuestros datos destacan la importancia de emplear herramientas de estudio, experimentales y estadísticas, que posibiliten un análisis integral de la respuesta de la planta. En este trabajo identificamos respuestas comunes a ambos tipos de estrés, operando a diferentes niveles. La respuesta al estrés es compleja: algunos mecanismos tienen un carácter general, independiente del factor de estrés, y por ello estarían de acuerdo con una teoría de respuesta común al estrés. Sin embargo, otros son idiosincráticos, y no necesariamente aumentarían la capacidad de las plantas de sobrellevar factores de estrés adicionales.

Referencias


Referencias
133
Adriano DC, Wenzel WW, Vangronsveld J, Bolan NS. 2004. Role of assisted natural remediation in environmental cleanup. Geoderma 122, 121-142.
Adriano DC. 2001. Trace Elements in Terrestrial Environments: Biogeochemistry, Bioavailability and Risk of Metals. New York: Springer-Verlag.
Ain-Lhout F, Zunzunegui M, Barradas MD, Tirado R, Clavijo A, Novo FG. 2001. Comparison of proline accumulation in two Mediterranean shrubs subjected to natural and experimental water deficit. Plant Soil 230, 175-183.
Alia, Pardha Saradhi P, Mohanty P. 1997. Involvement of proline in protecting thylakoid membranes against free radical-induced photodamage. J Photochem Photobiol B: Biology 38, 253-257.
Alloway BJ. 1995. Heavy Metals in Soils, 2nd edition. London: Blackie Academic & Professional.
Aloni R, Aloni E, Langhans M, Ullrich CI. 2006. Role of cytokinin and auxin in shaping root architecture: regulating vascular differentiation, lateral root initiation, root apical dominance and root gravitropism. Ann Bot 97: 883-893
Andersen SU, Buechel S, Zhao Z, Ljung K, Novák O, Busch W, Schuster C, Lohmann JU. 2008. Requirement of B2-type cyclin-dependent kinases for meristem integrity in Arabidopsis thaliana. Plant Cell 20: 88 - 100
Anderson MJ, Willis TJ. 2003. Canonical analysis of principal coordinates: a useful method of constrained ordination for ecology. Ecology 84, 511-525.
Anderson MJ. 2001. A new method for non-parametric multivariate analysis of variance. Austral Ecol 26, 32-46.
Anderson MJ. 2003. PCO: a FORTRAN computer program for principal coordinate analysis. Department of Statistics, University of Auckland, New Zealand.
Andreu V, Gimeno E. 1996. Total content and extractable fraction of cadmium, cobalt, copper, nickel, lead, and zinc in calcareous orchard soils. Commun Soil Sci Plant, 27, 2633-2648.
Anthos Project. 2009. Sistema de información de las plantas de España. Real Jardín Botánico, CSIC Fundación Biodiversidad, www.anthos.es. The site was visited on January 2009.
Antoniadis V, Alloway BJ. 2001. Availability of Cd, Ni and Zn to ryegrass in sewage sludge-treated soil at different temperatures. Water Air Soil Poll 132, 201-14.
Antonovics J, Bradshaw AD, Turner RG. 1971. Heavy metal tolerance in plants. Adv Ecol Res 7, 1-85.

Referencias
134
Aranda I, Castro L, Alia R, Pardos JA, Gil L. 2005. Low temperature during winter elicits differential responses among populations of the Mediterranean evergreen cork oak (Quercus suber). Tree Physiol 25, 1085-1090.
Arbona V, Argamasilla R, Gómez-Cadenas A. 2010. Common and divergent physiological, hormonal and metabolic responses of Arabidopsis thaliana and Thellungiella halophila to water and salt stress. J Plant Physiol 167, 1342-1350.
Arduini I, Godbold DL, Onnis A. 1994. Cadmium and copper change root growth and morphology of Pinus pinea and Pinus pinaster seedlings. Physiol Plantarum 92, 675-680.
Baker, AJ. 1981. Accumulators and excluders‐strategies in the response of plants to heavy metals. J Plant Nut 3, 643-654.
Balsberg Påhlsson AM. 1989. Toxicity of heavy metals (Zn, Cu, Cd, Pb) to vascular plants. A literature review. Water Air Soil Pollut 47, 287-319.
Barceló J, Poschenrieder Ch. 1990. Plant water relations as affected by heavy metal stress: a review. J Plant Nutr 13, 1-37.
Bartolomé C, Álvarez J, Vaquero J, Costa M, Casermeiro MA, Giraldo J, et al. 2005. Bosques de Tetraclinis articulata. In: Ministerio del Medio Ambiente. D.G. Biodiversidad, eds. Los Tipos de Hábitats de Interés Comunitario en España. Madrid: Ministerio del Medio Ambiente. p. 266-267.
Basta NT, Pantone DJ, Tabatabai MA. 1993. Path analysis of heavy metal adsorption by soil. Agron J 85, 1054-1057.
Bernhard R, Verkleij JA, Nelissen HJ, Vink JP. 2005. Plant-specific responses to zinc contamination in a semi-field lysimeter and on hydroponics. Environmental Pollution 138, 100-108.
Berntson GM. 1994. Modelling root architecture: are there tradeoffs between efficiency and potential of resource acquisition? New Phytol 127, 483-493.
Bohnert HJ, Nelson DE, Jensen RG. 1995. Adaptations to environmental stresses. Plant Cell 7, 1099-1111.
Bonfil C. 1998. The effects of seed size, cotyledon reserves, and herbivory on seedling survival and growth in Quercus rugosa and Q. laurina (Fagaceae). Am J Bot 85, 69-79.
Boyd RS. 2010. Heavy Metal Pollutants and Chemical Ecology: Exploring New Frontiers. J Chem Ecol 36, 46-58.
Brady KU, Kruckeberg AR, Bradshaw Jr HD. 2005. Evolutionary ecology of plant adaptation to serpentine soils. Ann Rev Ecol Evol Syst 36, 243-66

Referencias
135
Britt A, Fiscus EL. 2003. Growth responses of Arabidopsis DNA repair mutants to solar irradiation. Physiol Plantarum 118, 183-192.
Broadley MR, White PJ, Hammond JP, Zelko I, Lux, A. 2007. Zinc in plants. New Phytol 173, 677-702.
Brodribb TJ, Holbrook NM. 2003. Stomatal closure during leaf dehydration, correlation with other leaf physiological traits. Plant Physiol 132, 2166-2173.
Brunner I, Luster J, Günthardt-Goerg MS, Frey B. 2008. Heavy metal accumulation and phytostabilisation potential of tree fine roots in a contaminated soil. Environ Pollut 152, 559-568.
Bundy JG, Davey MP, Viant MR. 2009. Environmental metabolomics: a critical review and future perspectives. Metabolomics 5, 3-21.
Cakmak I. 2000. Possible roles of zinc in protecting plant cells from damage by reactive oxygen species. New Phytol 146, 185-205
Callender E. 2003. Heavy metals in the environment-historical trends. Treatise on Geochem 9, 67-105.
Chapin FS. 1980. The mineral nutrition of wild plants. Ann Rev Ecol Syst 11, 233-260.
Chapin FS. 1991. Integrated responses of plants to stress. BioScience 41, 29-36.
Chaves MM, Pereira JS. 2004. Respuestas de la estabilidad de las plantas al estrés múltiple y la habilidad de enfrentarse a un ambiente cambiante. In. Reigosa MJ, Pedrol N, Sánchez A, eds. La Ecofisiologıa Vegetal. Una ciencia de síntesis. Spain, Thomson. pp. 577-602.
Chaves MM, Pereira JS, Maroco J, Rodrigues ML, Ricardo CPP, Osorio ML, Carvalho I, Faria T, Pinheiro C. 2002. How plants cope with water stress in the field? Photosynthesis and growth. Ann Bot 89, 907-916.
Chiatante D, Di Iorio A, Scippa GS. 2005. Root responses of Quercus ilex L. seedlings to drought and fire. Plant Biosys Int J Deal Asp Plant Biol 139, 198-208.
Clemens S, Palmgren MG, Krämer U. 2002. A long way ahead: understanding and engineering plant metal accumulation. Trends Plant Sci 7, 309-315.
Clemente R, Walker DJ, Roig A, Bernal MP. 2003. Heavy metal bioavailability in a soil affected by mineral sulphides contamination following the mine spillage at Aznalcóllar (Spain). Biodegradation 14, 199-205.
Comas LH, Eissenstat DM. 2004. Linking fine root traits to maximum potential growth rate among 11 mature temperate tree species. Funct Ecol 18, 388-397.

Referencias
136
Conesa HM, Faz Á, Arnaldos R. 2006. Heavy metal accumulation and tolerance in plants from mine tailings of the semiarid Cartagena-La Unión mining district (SE Spain). Sci Tot Environ 366, 1-11.
Conesa HM, Faz Á. 2011. Metal uptake by spontaneous vegetation in acidic mine tailings from a semiarid area in South Spain: Implications for revegetation and land management. Water Air Soil Poll 215, 221-227.
Coomes DA, Grubb PJ. 2003. Colonization, tolerance, competition and seed-size variation within functional groups. Trends Ecol Evol 18, 283-291.
Davison PA, Hunter CN, Horton P. 2002. Overexpression of β-carotene hydroxylase enhances stress tolerance in Arabidopsis. Nature 418, 203-206.
Denny HJ, Wilkins DA. Zinc tolerance in Betula spp. I. Effect of external concentration of zinc on growth and uptake. New Phytol 106, 517-524.
Derome J, Nieminen T. 1998. Metal and macronutrient fluxes in heavy-metal polluted Scots pine ecosystems in SW Finland. Environ Pollut 103, 219-228.
Di Baccio D, Tognetti R, Minnocci A, Sebastiani L. 2009. Responses of the Populus×euramericana clone I-214 to excess zinc: Carbon assimilation, structural modifications, metal distribution and cellular localization. Environ Exp Bot 67, 153-163.
Di Castri F. 1973. Climatographical comparisons between Chile and the western coast of North America. In: Castri F, Mooney HA, eds. Mediterranean type-ecosystems, Springer-Verlag, Berlin, Germany, pp 21 – 36.
Disante KB, Cortina J, Vilagrosa A, Fuentes D, Hernández EI, Ljung K. 2013. Alleviation of Zn toxicity by low water availability. Physiol Plantarum, in press.
Disante KB, Fuentes D, Cortina J. 2010. Sensitivity to zinc of Mediterranean woody species important for restoration. Sci Tot Environ 408, 2216-2225.
Disante KB, Fuentes D, Cortina J. 2011. Response to drought of Zn-stressed Quercus suber L. seedlings. Environ Exp Bot 70, 96-103.
Domínguez MT, Marañón T, Murillo JM, Schulin R, Robinson BH. 2008. Trace element accumulation in woody plants of the Guadiamar Valley, SW Spain: a large-scale phytomanagement case study. Environ Pollut 152, 50-59.
Dos Santos Utmazian MN, Wieshammer G, Vega R, Wenzel WW. 2007. Hydroponic screening for metal resistance and accumulation of cadmium and zinc in twenty clones of willows and poplars. Environ Pollut, 148, 155-165.
Easterling DR, Meehl GA, Parmesan C, Changnon SA, Karl TR, Mearns LO. 2000. Climate Extremes: Observations, Modelling, and Impacts. Science 289, 2068-2074.

Referencias
137
Edlund A, Eklof S, Sundberg B, Moritz T, Sandberg G. 1995. A microscale technique for gas chromatography-mass spectrometry measurements of picogram amounts of indole-3-acetic acid in plant tissues. Plant Physiol 108, 1043-1047.
Eissenstat DM. 1991. On the relationship between specific root length and the rate of root proliferation: a field study using citrus rootstocks. New Phytol 118, 63-68.
Eissenstat DM, Wells CE, Yanai RD, Whitbeck JL. 2000. Building roots in a changing environment: Implications for root longevity. New Phytol 147, 33-42.
Eissenstat DM, Yanai RD. 1997. The ecology of root lifespan. Adv Ecol Res 27, 1-60.
Ellobied M, Polle A. 2012. Interference of Heavy Metal Toxicity with Auxin Physiology. In: Gupta DK, Sandalio LM, eds. Metal Toxicity in Plants: Perception, Signaling and Remediation, Springer-Verlag, Berlin, Heidelberg.
EPSAR. 2009. Entidad Pública de Saneamiento de Aguas Residuales de la Comunidad Valenciana. Memoria de Gestión. Generalitat Valenciana. Consellería de Medi Ambient, Aigua, Urbanismo i Habitatge.
Ernst WHO, Verkleij JAV, Schat H. 1992. Metal tolerance in plants. Acta Bot Neerl 41, 229-248.
Escrig I, Morell I. 1998. Effect of calcium on the soil adsorption of cadmium and zinc in some Spanish sandy soils. Water Air Soil Pollut 105, 507-520.
Fitter AH. 1991. Characteristic and functions of root systems. In: Waisel Y, Eshel A, Kafkafi U, eds. Plant Roots: The Hidden Half. New-York: Marcel Dekker Inc. p. 3- 24.
Fuentes D. 2011. Uso de biosolidos para el establecimiento de especies leñosas en medios Mediterráneos degradados. Tesis Doctoral. Universidad de Alicante.
Fuentes D, Disante KB, Valdecantos A, Cortina J, Vallejo VR. 2007a. Response of Pinus halepensis Mill. seedlings to biosolids enriched with Cu, Ni and Zn in three Mediterranean forest soils. Environ Pollut 145, 316-323.
Fuentes D, Disante KB, Valdecantos A, Cortina J, Vallejo VR. 2007b. Sensitivity of Mediterranean woody seedlings to copper, nickel and zinc. Chemosphere 66, 412-420.

Referencias
138
Fuentes D, Valdecantos A, Cortina J, Vallejo VR. 2007c. Seedling performance in sewage sludge-amended degraded Mediterranean woodlands. Ecol Engin 31, 281-291.
Fuentes D, Valdecantos A, Llovet J, Cortina J, Vallejo VR. 2010. Fine-tuning of sewage sludge application to promote the establishment of Pinus halepensis seedlings. Ecol Engin 36, 1213-1221.
Fuhrer J. 1982. Early effects of excess cadmium uptake in Phaseolus vulgaris. Plant Cell Environ 5, 263-270.
Fuller WH, Tucker TC. 1977. Land Utilization and disposal of Organic Wastes in Arid Regions. In: Elliot and Stevenson, eds. Soils for management of Organic Wastes and Waste Waters. American society of Agronomy, Inc.
Gadallah MAA, Ramadan T. 1997. Effects of zinc and salinity on growth and anatomical structure of Carthamus tinctorius L. Biol Plantarum 39, 411-418
Gadallah MAA. 2000. Effects of indole-3-acetic acid and zinc on the growth, osmotic potential and soluble carbon and nitrogen components of soybean plants growing under water deficit. J Arid Environ 44, 451-467.
Galmés J, Medrano H, Flexas J. 2007. Photosynthesis and photoinhibition in response to drought in a pubescent (var. minor) and a glabrous (var. palaui) variety of Digitalis minor. Environ Exp Bot, 60, 105-111.
García LV. 2004. Escaping the Bonferroni iron claw in ecological studies. Oikos 105, 657-663.
Garcia-Miragaya J. 1984. Levels, chemical fractionation, and solubility of lead in roadside soils of Caracas, Venezuela. Soil Sci 138, 147-152.
Gebre GM, Tschaplinski TJ. 2002. Solute accumulation of chestnut oak and dogwood leaves in response to throughfall manipulation of an upland oak forest. Tree Physiol 22, 251-260.
Genty B, Briantais JM, Baker NR. 1989. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim Biophys Acta 990, 87-92.
Ghnaya AB, Hourmant A, Cerantola S, Kervarec N, Cabon JY, Branchard M, Charles G. 2010. Influence of zinc on soluble carbohydrate and free amino acid levels in rapeseed plants regenerated in vitro in the presence of zinc. Plant Cell Tiss Org 102, 191-197.
Gibson SI. 2000. Plant sugar-response pathways. Part of a complex regulatory web. Plant Physiol 124, 1532-1539.

Referencias
139
Gimeno TE, Pas B, Lemos-Filho JP, Valladares F. 2008. Plasticity and stress tolerance override local adaptation in the responses of Mediterranean holm oak seedlings to drought and cold. Tree Physiol 29, 87-98.
González V, García I, del Moral F, de Haro S, Sánchez JA, Simón M. 2011. Impact of unconfined sulphur-mine waste on a semi-arid environment (Almería, SE Spain). J Environ Manag 92, 1509-1519.
Greger M. 2004. Metal availability, uptake, transport and accumulation in plants. In: Prasad MNV ed. Heavy Metal Stress in Plants. From Molecule to Ecosystems. Springer, Berlin, pp. 1-27.
Grimalt JO, Ferrer M, Macpherson E. 1999. The mine tailing accident in Aznalcóllar. Sci Total Environ 242, 3-11.
Grime JP. 1977. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am Nat, 1169-1194.
Gruber N, Galloway JN. 2008. An Earth-system perspective of the global nitrogen cycle. Nature 451, 293-296.
Gullberg J, Jonsson P, Nordström A, Sjöström M, Moritz T. 2004. Design of experiments: an efficient strategy to identify factors influencing extraction and derivatization of Arabidopsis thaliana samples in metabolomic studies with gas chromatography/mass spectrometry. Anal Biochem 331, 283-295.
Habitat Directive. 1992. Habitat Directive 92/43/EEC of 21 of May on the Conservation of natural habitats and of wild fauna and flora. Brussels, Belgium.
Hagemeyer J. 2004. Ecophysiology of plant growth under heavy metal stress. In: Prasad MNV, ed. Heavy Metal Stress in Plants. From Molecule to Ecosystems. Springer, Berlin, pp. 201-222.
Hall RD. 2006. Plant metabolomics: from holistic hope, to hype, to hot topic. New Phytol 169, 453-468.
Han FX, Banin A, Su Y, Monts DL, Plodinec MJ, Kingery WL, Triplett GE. 2002. Industrial age anthropogenic inputs of heavy metals into the pedosphere. Naturwissenschaften 89, 497-504.
Harmens H, Koevoets PL, Verkleij JA, Ernst WH. 1994. The role of low molecular weight organic acids in the mechanism of increased zinc tolerance in Silene vulgaris (Moench) Garcke. New Phytol 126, 615-621.
Hart JJ, Norvell WA, Welch RM, Sullivan LA, Kochian LV. 1998. Characterization of zinc uptake, binding, and translocation in intact seedlings of Bread and Durum wheat cultivars. Plant Physiol 118, 219-226.

Referencias
140
Havaux M, Eymery F, Porfirova S, Rey P, Dörmann P. 2005. Vitamin E protects against photoinhibition and photooxidative stress in Arabidopsis thaliana. Plant Cell Online 17, 3451-3469.
Hawkes, SJ. 1997. What Is a" Heavy Metal"? Journal of Chemical Education, ACS Publications
Hernández EI, Vilagrosa A, Pausas JG, Bellot J. 2010. Morphological traits and water use strategies in seedlings of Mediterranean coexisting species. Plant Ecol 207, 233-244.
Hubbard RM, Ryan MG, Stiller V, Sperry JS. 2001. Stomatal conductance and photosynthesis vary linearly with plant hydraulic conductance in ponderosa pine. Plant Cell Environ 24, 113-121.
Hu Y, Schmidhalter U. 2005. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J Plant Nutr Soil Sc 168, 541-549.
Hüttermann A, Arduini I, Godbold DL. 2004. Metal pollution and forest decline. In: Prasad MNV, ed. Heavy Metal Stress in Plants. From Molecule to Ecosystems. Berlin, Springer. p. 295-312.
IPCC. 2001. Climate Change 2001: The Scientific Basis. Contribution of Working Group I. In: Hougton JT, Dung Y, Griggs DJ, Noguer M, Van der Linden PJ, Dui X, Maskell K, Johson CA, eds. Third Assessment Report of Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge.
Jha AB, Dubey RS. 2004. Carbohydrate metabolism in growing rice seedlings under arsenic toxicity. J Plant Physiol 161, 867-872.
Jiménez MD, Pardos M, Puértolas J, Kleczkowski LA, Pardos JA. 2009. Deep shade alters the acclimation response to moderate water stress in Quercus suber L. Forestry 82, 285-298.
Johnson PS, Shifley SR, Rogers R. 2001. The ecology and silviculture of oaks. CABI Publishing. Oxon, UK
Jones Jr JB, Case VW. 1990. Sampling, handling, and analyzing plant tissue samples. In: Westerman RL, ed. Soil Testing and Plant Analysis, SSSA Book Series 3, Madison, Wisconsin, p. 389-427.
Jonsson P, Johansson AI, Gullberg J, Trygg J, Grung B, Marklund S, Sjöström M, Antti H, Moritz T. 2005. High-throughput data analysis for detecting and identifying differences between samples in GC/MS-based metabolomic analyses. Anal Chem 77, 5635-5642.
Joshi MK, Mohanty P. 2004. Chlorophyll a fluorescence as a probe of heavy metal ion toxicity in plants. In: Papageorgiou GC, Govindjee, eds. Chlorophylla fluorescence. A signature of photosynthesis, Springer Netherlands, p. 637-661.

Referencias
141
Kabata-Pendias A, Pendias H. 1992. Trace elements in soils and plants. Boca Raton, FL: CRC Press.
Karathanasis AD, Johnson DMC, Matocha CJ. 2005. Biosolid colloid-mediated transport of copper, zinc, and lead in waste-amended soils. J Environ Qual 34, 1153-1164.
Kasim WA. 2007. Physiological consequences of structural and ultra-structural changes induced by Zn stress in Phaseolus vulgaris. I. Growth and photosynthetic apparatus. Int J Bot 3, 15-22.
Knight B, Zhao FJ, McGrath SP, Shen ZG. 1997. Zinc and cadmium uptake by the hyperaccumulator Thlaspi caerulescens in contaminated soils and its effects on the concentration and chemical speciation of metals in soil solution. Plant Soil 197, 71-78.
Köhl K, Lösch R. 2004. Experimental characterization of metal tolerance. In: Prasad MNV, ed. Heavy Metal Stress in Plants. From Molecule to Ecosystems. Berlin, Springer. p. 434-455.
Kukkola E, Huttunen S, Bäck J, Rautio P. 1997. Scots pine needle injuries at subarctic industrial sites. Trees 11, 378-387.
Larcher W. 2003. Physiological plant ecology: Ecophysiology and stress physiology of functional groups, 4th edition. Berlin, Springer.
Lasat MM, Pence NS, Garvin DF, Ebbs SD, Kochian LV. 2000. Molecular physiology of zinc transport in the Zn hyperaccumulator Thlaspi caerulescens. J Exp Bot 51, 71-79.
Lawlor DW, Cornic G. 2002. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ 25, 275-294.
Lawlor DW, Tezara W. 2009. Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells: a critical evaluation of mechanisms and integration of processes. Ann Bot 103, 561-579.
Lee SH, Chung GC, Steudle E. 2005. Low temperature and mechanical stresses differently gate aquaporins of root cortical cells of chilling-sensitive cucumber and resistant figleaf gourd. Plant Cell Environ 28, 1191-1202.
León MF, Squeo FA, Gutiérrez JR, Holmgren M. 2011. Rapid root extension during water pulses enhances establishment of shrub seedlings in the Atacama Desert. J Veg Sci 22, 120-129.
Lepš J, Šmilauer P. 2003. Multivariate analysis of ecological data using CANOCO. Cambridge University Press.

Referencias
142
Lequeux H, Hermans C, Lutts S, Verbruggen N. 2010. Response to copper excess in Arabidopsis thaliana: Impact on the root system architecture, hormone distribution, lignin accumulation and mineral profile. Plant Physiol Biochem 48, 673-682.
Leshem YY, Kuiper PJC, Erdei L, Lurie S, Perl-Treves R. 1998. Do Selye's mammalian “GAS” concept and “Co-stress” response exist in plants? Ann NY Acad Sci 851, 199-208.
Leshem YY, Kuiper PJC. 1996. Is there a GAS (general adaptation syndrome) response to various types of environmental stress? Biol Plantarum 38, 1-18.
Li MS. 2006. Ecological restoration of mineland with particular reference to the metalliferous mine wasteland in China: A review of research and practice. Sci Tot Environ, 357, 38-53.
Ljung K, Hull AK, Celenza J, Yamada M, Estelle M, Normanly J, Sandberg, G. 2005. Sites and regulation of auxin biosynthesis in Arabidopsis roots. Plant Cell 17, 1090-1104.
Lo Gullo MA, Nardini A, Salleo S, Tyree MT. 1998. Changes in root hydraulic conductance (KR) of Olea oleaster seedlings following drought stress and irrigation. New Phytol, 140, 25-31.
Long SP, Humphries S, Falkowski PG. 1994. Photoinhibition of photosynthesis in nature. Annu Rev Plant Physiol Plant Mol Biol 45, 633-62.
Lottermoser BG, Ashley PM, Munksgaard NC. 2008. Biogeochemistry of Pb-Zn gossans, northwest Queensland, Australia: Implications for mineral exploration and mine site rehabilitation. Appl Geochem 23, 723-742.
Lusk CH, Reich PB, Montgomery RA, Ackerly DD, Cavender-Bares J. 2008. Why are evergreen leaves so contrary about shade? Trends Ecol Evol 23, 299-303.
Ma JF, Furukawa J. 2003. Recent progress in the research of external Al detoxification in higher plants, a minireview. J Inorg Biochem 97, 46-51.
Macnair MR. 1987. Heavy metal tolerance in plants: a model evolutionary system. Trends Ecol Evol 2, 354-359.
Maestre FT, Cortina J. 2004. Are Pinus halepensis plantations useful as a restoration tool in semiarid Mediterranean areas? For Ecol Manag 198, 303-317.
Marschner H. 1995. Mineral Nutrition of Higher Plants. 2nd edition. London: Academic Press.
Martín JAR, Arias ML, Corbí JMG .2009. Metales pesados, materia orgánica y otros parámetros de los suelos agrícolas y pastos de España. Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria.

Referencias
143
Martín S, Diaz-Fernández P, De Miguel J. 1998. Regiones de procedencia de especies forestales españolas. Generos Abies, Fagus, Pinus, y Quercus. Madrid: O.A. Parques Nacionales.
Martínez CE, Motto HL. 2000. Solubility of lead, zinc and copper added to mineral soils. Environ Pollut 107, 153-158.
Masclans F. 1981. Guia per a conèixer els arbres. Barcelona, Montblanc-CEC.
Mateos-Naranjo E, Redondo-Gómez S, Cambrollé J, Luque T, Figueroa ME. 2008. Growth and photosynthetic responses to zinc stress of an invasive cordgrass, Spartina densiflora. Plant Biol 10, 754-762.
Matilla A. 2004. Ecofisiología de la germinación de semillas. In: Reigosa MJ, Pedrol N, Sánchez A eds. La Ecofisiologıa Vegetal. Una ciencia de síntesis. Spain, Thomson. pp. 901-923.
Mattsson J, Ckurshumova W, Berleth T. 2003. Auxin signaling in Arabidopsis leaf vascular development. Plant Physiol 131, 1327-1339.
Medrano H, Flexas J. 2004. Relaciones hídricas de las plantas. In: Reigosa MJ, Pedrol N, Sanchez A, eds. La ecofisiología vegetal. Una ciencia de síntesis. Madrid, Thomson, 253-286.
Menon M, Hermle S, Abbaspour KC, Günthardt-Goerg MS, Oswald SE, Schulin R. 2005. Water regime of metal-contaminated soil under juvenile forest vegetation. Plant Soil 271, 227-241.
Méthy M, Damesin C, Rambal S. 1996. Drought and photosystem II activity in two Mediterranean oaks. Ann Sci For 53, 255-262.
Micó C, Recatala L, Peris M, Sánchez J. 2006. Assessing heavy metal sources in agricultural soils of an European Mediterranean area by multivariate analysis. Chemosphere 65, 863-872.
Mico Llopis C, Peris M, Sanchez J, Recatala L. 2006. Heavy metal content of agricultural soils in a Mediterranean semiarid area: the Segura River Valley (Alicante, Spain). Spanish J Agric Res 4, 363-372.
Mittler R. 2006. Abiotic stress, the field environment and stress combination. Trends Plant Sci 11, 15-19.
Monnet F, Vaillant N, Vernay P, Coudret A, Sallanon H, Hitmi A. 2001. Relationship between PSII activity, CO2 fixation, and Zn, Mn and Mg contents of Lolium perenne under zinc stress. J Plant Physiol 158, 1137-1144.

Referencias
144
Monni S, Uhlig C, Hansen E, Magel E. 2001. Ecophysiological responses of Empetrum nigrum to heavy metal pollution. Environ Pollut 112, 121-129.
Morgan JM. 1984. Osmoregulation and water stress in higher plants. Ann Rev Plant Physiol 35, 299-319
Moustakas M, Eleftheriou EP, Ouzounidou G. 1997. Short-term effects of aluminium at alkaline pH on the structure and function of the photosynthetic apparatus. Photosynthetica 34, 169-177.
Munné-Bosch S, Alegre L. 2003. Drought-induced changes in the redox state of α-tocopherol, ascorbate, and the diterpene carnosic acid in chloroplasts of Labiatae species differing in carnosic acid contents. Plant Physiol 131, 1816-1825.
Munne-Bosch S. 2005. The role of α-tocopherol in plant stress tolerance. J Plant Physiol 162, 743-748.
Myśliwa-Kurdziel B, Prasad MNV, Strzalka K. 2004. Photosynthesis in heavy metal stressed plants. In: Prasad MNV, ed. Heavy Metal Stress in Plants. From Molecule to Ecosystems. Springer, Berlin, pp. 146-181.
Nagajyoti PC, Lee KD, Sreekanth TVM. 2010. Heavy metals, occurrence and toxicity for plants: a review. Environ Chem Lett 8, 199-216.
Nèble S, Calvert V, Le Petit J, Criquet S. 2007. Dynamics of phosphatase activities in a cork oak litter (Quercus suber L.) following sewage sludge application. Soil Biol Biochem 39, 2735-2742.
Nicolás MJ, Esteve MA, Palazón JA, López Hernández JJ. 2004. Modelo sobre las preferencias de hábitat a escala local de Tetraclinis articulata (Vahl) Masters en una población del límite septentrional de su área de distribución. Anales de Biología 26,157-167.
Nicotra AB, Babicka N, Westoby M. 2002. Seedling root anatomy and morphology: an examination of ecological differentiation with rainfall using phylogenetically independent contrasts. Oecologia 130, 136-145.
Nilsen ET, Orcutt DM. 1996. The physiology of plants under stress: abiotic factors. New York, John Wiley and Sons.
Noyes PD, McElwee MK, Miller HD, Clark BW, Van Tiem LA, Walcott KC, et al. 2009. The toxicology of climate change: environmental contaminants in a warming world. Environ Int 35, 971-86.
Nyquist J, Greger M. 2007. Uptake of Zn, Cu, and Cd in metal loaded Elodea canadensis. Environ Exp Bot 60, 219-226.

Referencias
145
Öncel I, Keles Y, Üstün AS. 2000. Interactive effects of temperature and heavy metal stress on the growth and some biochemical compounds in wheat seedlings. Environ Pollut 107, 315-320.
Orgeas J, Ourcival JM, Bonin G. 2003. Seasonal and spatial patterns of foliar nutrients in cork oak (Quercus suber L.) growing on siliceous soils in Provence (France). Plant Ecol 164, 201-211.
Osmond B, Schwartz O, Gunning B. 1999. Photoinhibitory printing on leaves, visualised by chlorophyll fluorescence imaging and confocal microscopy, is due to diminished fluorescence from grana. Aust J Plant Physiol 26, 717-724.
Otieno DO, Schmidt MWT, Kurz-Besson C, Vale RLD, Pereira JS, Tenhunen JD. 2007. Regulation of transpirational water loss in Quercus suber trees in a Mediterranean-type ecosystem. Tree Physiol 27, 1179-1187.
Ostonen I, Püttsepp Ü, Biel C, Alberton O, Bakker MR, Lõhmus K, Brunner I. 2007. Specific root length as an indicator of environmental change. Plant Biosyst, 141, 426-442.
Oyarzun R, Cubas P, Higueras P, Lillo J, Llanos W. 2009. Environmental assessment of the arsenic-rich, Rodalquilar gold-(copper-lead-zinc) mining district, SE Spain: data from soils and vegetation. Environ Geol 58, 761-777.
Oyarzun R, Lillo J, Lopez-Garcia JA, Esbrí JM, Cubas P, Llanos W, Higueras P. 2011. The Mazarrón Pb-(Ag)-Zn mining district (SE Spain) as a source of heavy metal contamination in a semiarid realm: Geochemical data from mine wastes, soils, and stream sediments. J Geochem Explor 109, 113-124.
Pachauri RK, Reisinger A. 2007. Climate Change 2007: Synthesis Report. Contribution of Working Groups I, II and III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change (IPCC). Intergovernmental Panel on Climate Change, 1. Cambridge University Press.
Pacyna JM, Pacyna EG. 2001. An assessment of global and regional emissions of trace metals to the atmosphere from anthropogenic sources worldwide. Environ Rev 9, 269-298.
Padilla FM, Miranda JDD, Pugnaire FI. 2009. Early root growth plasticity in seedlings of three Mediterranean woody species. Plant Soil 296, 103-113.
Padilla FM, Pugnaire FI. 2007. Rooting depth and soil moisture control Mediterranean woody seedling survival during drought. Funct Ecol 21, 489-495.
Päivöke AEA. 2003. Responses of Pisum Sativum to soil arsenate, lead and zinc: A greenhouse study of mineral elements, phytase activity, ATP and chlorophylls. Academic dissertation. Faculty of Science of the University of Helsinki.

Referencias
146
Paschke MW, Redente EF, Levy DB. 2000. Zinc toxicity thresholds for important reclamation grass species of the Western United States. Environ Toxicol Chem 19, 2751-2756.
Paschke MW, Valdecantos A, Redente EF. 2005. Manganese toxicity thresholds for restoration grass species. Environ Pollut 135, 313-322.
Pascual I, Antolín MC, García C, Polo A, Sánchez-Díaz M. 2004. Plant availability of heavy metals in a soil amended with a high dose of sewage sludge under drought conditions. Biol Fert Soils 40, 291-299.
Passarinho JA, Lamosa P, Baeta JP, Santos H, Ricardo CP. 2006. Annual changes in the concentration of minerals and organic compounds of Quercus suber leaves. Physiol Plantarum 127, 100-110.
Pätsikka E, Kairavuo M, Šeršen F, Aro EM, Tyystjärvi E. 2002. Excess copper predisposes photosystem II to photoinhibition in vivo by outcompeting iron and causing decrease in leaf chlorophyll. Plant Phsysiol 129, 1359-1367.
Pausas J, Pereira J, Aronson J. 2009. The tree. In: Aronson J, Pereira JS, Pausas JG, eds. Cork Oak Woodlands on the Edge. Ecology, Adaptive Management and Restoration, Island Press, Washington DC, pp 11-25.
Pence NS, Larsen PB, Ebbs SD, Letham DL, Lasat MM, Garvin DF, Kochian LV. 2000. The molecular physiology of heavy metal transport in the Zn/Cd hyperaccumulator Thlaspi caerulescens. P Nat Acad Sci 97, 4956-4960.
Peñuelas J, Filella I, Lloret F, Piñol J, Siscart D. 2000. Effects of a severe rain exclusion on water and nitrogen use by Quercus ilex and Phillyrea latifolia. Biol Plant 43, 47-53.
Peñuelas J, Filella I. 2002. Metal pollution in Spanish terrestrial ecosystems during the twentieth century. Chemosphere 46, 501-505.
Pereira P, Úbeda X. 2010. Spatial distribution of heavy metals released from ashes after a wildfire. J Environ Eng Landsc Manag 18, 13-22.
Peris M, Recatalá L, Micó C, Sánchez R, Sánchez J. 2008. Increasing the knowledge of heavy metal contents and sources in agricultural soils of the European Mediterranean region. Water Air Soil Poll 192, 25-37.
Pető A, Lehotai N, Lozano-Juste J, León J, Tari I, Erdei L, Kolbert Z. 2011. Involvement of nitric oxide and auxin in signal transduction of copper-induced morphological responses in Arabidopsis seedlings. Ann Bot 108, 449-457.
Pohlmeier A. 2004. Metal speciation, chelation and complexing ligands in plants. In: Prasad MNV, ed. Heavy Metal Stress in Plants. From Molecule to Ecosystems. 2nd edition. Berlin, Springer, 28-46.

Referencias
147
Popovic R, Dewez D, Juneau P. 2003. Application of chlorophyll fluorescence in ecotoxicology: heavy metals, herbicides, and air pollutants. In: Dell JR, Toivinen PMA, eds. Practical applications of chlorophyll fluorescence in plant biology. Kluwer Academic Publishers, Boston, pp 151-184.
Poschenrieder C, Barceló J. 2004b. Water relations in heavy metal stressed plants. In: Prasad MNV, ed. Heavy Metal Stress in Plants. From Molecule to Ecosystems, Springer, Berlin, pp. 249-271.
Poschenrieder C, Gunse B, Barceló J. 1989. Influence of cadmium on water relations, stomatal resistance, and abscisic acid content in expanding bean leaves. Plant Physiol 90, 1365-1371.
Poschenrieder Ch, Barceló J. 2004a. Estrés por metales pesados. Relaciones hídricas de las plantas. In: Reigosa MJ, Pedrol N, Sánchez A, eds. La ecofisiología vegetal. Una ciencia de síntesis. Madrid. Spain. pp. 413-442.
Potters G, Pasternak TP, Guisez Y, Jansen MA. 2009. Different stresses, similar morphogenic responses: integrating a plethora of pathways. Plant Cell Environ 32, 158-169.
Potters G, Pasternak TP, Guisez Y, Palme KJ, Jansen MA. 2007. Stress-induced morphogenic responses: growing out of trouble? Trends Plant Sci 12, 98-105.
Prasad MNV. 2004. Metallothioneins, metal binding complexes and metal sequestration in plants. In: Prasad MNV, ed. Heavy Metal Stress in Plants. From Molecule to Ecosystems. Berlin, Springer, p. 47-83.
Pregitzer KS, Laskowski MJ, Burton AJ, Lessard VC, Zak DR. 1998. Variation in sugar maple root respiration with root diameter and soil depth. Tree Physiol 18, 665-670.
Pulford ID, Watson C. 2003. Phytoremediation of heavy metal-contaminated land by trees—a review. Environ Int 29, 529-540.
Rajkumar M, Prasad MNV, Swaminathan S, Freitas H. 2013. Climate change driven plant–metal–microbe interactions. Environ Int 53, 74-86.
Ralph PJ, Burchett MD. 1998. Photosynthetic response of Halophila ovalis to heavy metal stress. Environ Pollut 103, 91-101.
Ramírez-Valiente JA, Valladares F, Gil L, Aranda I. 2009. Population differences in juvenile survival under increasing drought are mediated by seed size in cork oak (Quercus suber L.). For Ecol Manag 257, 1676-1683.
Reichman SM, Asher CJ, Mulligan DR, Menzies NW. 2001. Seedling responses of three Australian tree species to toxic concentrations of zinc in solution culture. Plant Soil 235, 151-158.

Referencias
148
Reichman SM. 2002. The Responses of Plants to Metal Toxicity: A Review Focusing on Copper, Manganese & Zinc. Melbourne, Australia: Australian Minerals & Energy Environment Foundation, pp. 22-26.
Rhizopoulou S, Mitrakos K. 1990. Water relations of evergreen sclerophylls. I. Seasonal changes in the water relations of eleven species from the same environment. Ann Bot 65, 171-178.
Riba I, García-Luque E, Maz-Courrau A, de Canales MLG, DelValls TÁ. 2010. Influence of salinity in the bioavailability of Zn in sediments of the gulf of Cádiz (Spain). Water Air Soil Poll 212, 329-336.
Richardson DM. 1998. Ecology and biogeography of Pinus. Cambridge, University Press.
Rieuwerts JS, Thornton I, Farago ME, Ashmore MR. 1998. Factors influencing metal bioavailability in soils: preliminary investigations for the development of a critical loads approach for metals. Chem Spec Bioavailab 10, 61-75.
Rizhsky L, Liang H, Mittler R. 2002. The combined effect of drought stress and heat shock on gene expression in tobacco. Plant Physiol 130, 1143-1151.
Rizhsky L, Liang H, Shuman J, Shulaev V, Davletova S, Mittler R. 2004. When defense pathways collide. The response of Arabidopsis to a combination of drought and heat stress. Plant Physiol 134, 1683-1696.
Robb J, Busch L, Rauser WE. 1980. Zinc toxicity and xylem vessel wall alterations in white beans. Ann Bot 46, 43-50.
Robert B, Berton G, Sayag D, Masson P. 1996. Assessment of mineral nutrition of cork oak through foliar analysis. Commun Soil Sci Plant Anal 27, 2091-2109.
Robert HS, Friml J. 2009. Auxin and other signals on the move in plants. Nat Chem Biol 5, 325-332.
Roca-Perez L, Gil C, Cervera ML, Gonzálvez A, Ramos-Miras J, Pons V, Boluda R. 2010. Selenium and heavy metals content in some Mediterranean soils. J Geochem Explor 107, 110-116.
Romero C, Espinosa A, Cutanda M, Cortina C, Hernández P, Culiáñez A. 2004. La osmorregulación: mecanismos y significado. In: Reigosa MJ, Pedrol N, Sánchez A, eds. La Ecofisiologıa Vegetal. Una ciencia de síntesis. Spain, Thomson. pp 603-620.
Rosolem CA, Sacramento LVS, Oliveira DMT. 2005. Kinetics of zinc uptake and anatomy of roots and leaves of coffee trees as affected by zinc nutrition. J Plant Nutr 28, 2101-2112.

Referencias
149
Ruano A, Barcelo J, Poschenrieder C. 1987. Zinc toxicity‐induced variation of mineral element composition in hydroponically grown bush bean plants. J Plant nut 10, 373-384.
Ruano A, Poschenrieder Ch, Barceló J. 1988. Growth and biomass partitioning in zinc-toxic bush beans. J Plant Nutr 11, 577-588.
Ryan PR, Delhaize E, Jones DL. 2001. Function and mechanism of organic anion exudation from plant roots. Annu Rev Plant Physiol Plant Mol Biol 52, 527-560.
Sagardoy R, Morales F, Rellán-Álvarez R, Abadía A, Abadía J, López-Millán AF. 2011. Carboxylate metabolism in sugar beet plants grown with excess Zn. J Plant Physiol 168, 730-733.
Samarakoon AB, Rauser WE. 1979. Carbohydrate levels and photoassimilate export from leaves of Phaseolus vulgaris exposed to excess cobalt, nickel, and zinc. Plant Physiol 63, 1165-1169.
Sánchez-Monedero MA, Mondini C, De Nobili M, Leita L, Roig A. 2004. Land application of biosolids. Soil response to different stabilization degree of the treated organic matter. Waste Manag 24, 325-332.
Santala KR, Ryser P. 2009. Influence of heavy-metal contamination on plant response to water availability in white birch, Betula papyrifera. Environ Exp Bot 66, 334-340.
Sardans J, Penuelas J, Prieto P, Estiarte M. 2008. Changes in Ca, Fe, Mg, Mo, Na, and S content in a Mediterranean shrubland under warming and drought. J Geophys Res 13, 1-11.
Sardans J, Penuelas J. 2005. Trace element accumulation in the moss Hypnum cupressiforme Hedw. and the trees Quercus ilex L. and Pinus halepensis Mill. in Catalonia. Chemosphere 60, 1293-1307.
Sardans J, Peñuelas J. 2007. Drought changes the dynamics of trace element accumulation in a Mediterranean Quercus ilex forest. Environ Pollut 147, 567-583.
Savin R, Nicolas ME. 1996. Effects of short periods of drought and high temperature on grain growth and starch accumulation of two malting barley cultivars. Funct Plant Biol 23, 201-210.
Schat H, Sharma SS, Vooijs R. 1997. Heavy metal-induced accumulation of free proline in a metal-tolerant and a nontolerant ecotype of Silene vulgaris. Physiol Plantarum 101, 477-482.
Schmidt W. 2008 Inner voices meet outer signals: the plasticity of rhizodermic cells. Plant Sci 174, 239-245.

Referencias
150
Selye H. 1936. The alarm reaction. Can Med Assoc J, vol. 34, p. 706.
Serrasolses I, Pérez-Devesa M, Vilagrosa A, Pausas JG, Sauras T, Cortina J, et al. 2009. Soil properties constraining cork oak distribution. In: Aronson J, Pereira JS, Pausas JG, eds. Cork Oak Woodlands on the Edge. Ecology, Adaptive Management and Restoration. Washington DC, Island Press. p. 89-99.
Sharma SS, Dietz KJ. 2006. The significance of amino acids and amino acid-derived molecules in plant responses and adaptation to heavy metal stress. J Exp Bot 57, 711-726.
Shulaev V, Cortes D, Miller G, Mittler R. 2008. Metabolomics for plant stress response. Physiol Plantarum 132, 199-208.
Smith SR, 1996. Agricultural Recycling of Sewage Sludge and the Environment. CAB International, Wallingford, UK.
Sperry JS, Hacke UG, Oren R, Comstock JP. 2002. Water deficits and hydraulic limits to leaf water supply. Plant Cell Environ 25, 251-263.
Stahl RS, James BR. 1991. Zinc sorption by iron-oxide-coated sand as a function of pH. Soil Sci Soc Am J 55, 1287-1290.
Steudle E. 2000. Water uptake by roots: effects of water deficit. J Exp Bot 51, 1531-1542.
Sun X, Zhang J, Zhang H, Ni Y, Zhang Q, Chen J, Guan Y. 2010. The responses of Arabidopsis thaliana to cadmium exposure explored via metabolite profiling. Chemosphere 78, 840-845.
Tack FM, Vandecasteele B. 2008. Cycling and ecosystem impact of metals in contaminated calcareous dredged sediment-derived soils (Flanders, Belgium). Sci Total Environ 400, 283-289.
Takahashi R, Ishimaru Y, Shimo H, Ogo Y, Senoura T, Nishizawa NK, Nakanishi H. 2012. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice. Plant Cell Environ 35, 1948-1957.
Tolrà RP, Poschenrieder C, Barceló J. 1996. Zinc hyperaccumulation in Thlaspi caerulescens. II Influence on organic acids. J Plant Nut 19, 1541-1550.
Toribio M, Romanyà J. 2006. Leaching of heavy metals (Cu, Ni and Zn) and organic matter after sewage sludge application to Mediterranean forest soils. Sci Tot Environ 363, 11-21.
Trubat R, Cortina J, Vilagrosa A. 2010. Nursery fertilization affects seedling traits but not field performance in Quercus suber L. J Arid Environ 74, 491-497.
Tsalandzono NG, Omokolo ND, Tita AM. 1994. Effect of Fe2+, Mn2+, Zn2+, and Pb2+on H+/K+ fluxes in excised Pistia stratiotes roots. Biol Plant 36, 591-597.

Referencias
151
Tyree MT, Patiño S, Bennink J, Alexander J. 1995. Dynamic measurements of roots hydraulic conductance using a high-pressure flowmeter in the laboratory and field. J Exp Bot 46, 83-94.
UNEP (United Nations Environment Program). 1997. World atlas of desertification, 2nd ed. Oxford, Oxford University Press for the UNEP.
Utriainen MA, Kärenlampi LV, Kärenlampi SO, Schat H. 1997. Differential tolerance to copper and zinc of micropropagated birches tested in hydroponics. New Phytol 137, 543-549.
Vaillant N, Monnet F, Hitmi A, Sallanon H, Coudret A. 2005 Comparative study of responses in four Datura species to a zinc stress. Chemosphere 59, 1005-1013.
Valladares F, Dobarro I, Sánchez-Gómez D, Pearcy RW. 2005. Photoinhibition and drought in Mediterranean woody saplings: scaling effects and interactions in sun and shade phenotypes. J Exp Bot 56, 483-494.
Valladares F, Pearcy RW. 1997. Interactions between water stress, sun-shade acclimation, heat tolerance, and photoinhibition in the sclerophyll Heteromeles arbutifolia. Plant Cell Environ 20, 25-36.
Valladares F, Sánchez-Gómez D. 2006. Ecophysiological traits associated with drought in Mediterranean tree seedlings: individual responses versus interspecific trends in eleven species. Plant Biol 8, 688-697.
Valladares F, Vilagrosa A, Peñuelas J, Ogaya R, Camarero JJ, Corcuera L, Gil-Pelegrín E. 2004. Estrés hídrico: ecofisiología y escalas de la sequía. Ecología del bosque mediterráneo en un mundo cambiante. Organismo Autónomo de Parques Nacionales. Ministerio de Medio Ambiente, Madrid.
Vallejo VR, Bautista S, Cortina J. 2000. Restoration for soil protection after disturbances. In: Trabaud L, ed. Life and Environment in the Mediterranean, WIT Press, Southampton, pp. 199-208.
Van Assche F, Clijsters H. 1986. Inhibition of Photosynthesis in Phaseolus vulgaris by Treatment with Toxic Concentration of Zinc: Effect on Ribulose-1, 5-bisphosphate Carboxylase/Oxygenase. J Plant Physiol 125, 355-360.
van Hees A. 1997. Growth and morphology of pedunculate oak Quercus robur L. and beech Fagus sylvatica L. seedling in relation to shading and drought. Ann Sci For 54, 9-18.
Vanneste S, Friml J. 2009. Auxin: a trigger for change in plant development. Cell 136, 1005-1016.
Verkleij JA. 1993. The effects of heavy metal stress on higher plants and their use as biomonitors. Plant as bioindicators: indicators of heavy metals in the terrestrial environment. New York, VCH, pp. 415-424.

Referencias
152
Vilagrosa A, Cortina J, Gil-Pelegrin E, Bellot J. 2003a. Suitability of drought preconditioning techniques in Mediterranean climate. Restor Ecol 11, 208-216.
Vilagrosa A, Morales F, Abadía A, Bellot J, Cochard H, Gil-Pelegrin E. 2010. Are symplast tolerance to intense drought conditions and xylem vulnerability to cavitation coordinated? An integrated analysis of photosynthetic, hydraulic and leaf level processes in two Mediterranean drought-resistant species. Environ Exp Bot 69, 233-242.
Vilagrosa A, Vallejo VR, Bellot J, Gil-Pelegrín E. 2003b. Cavitation, stomatal conductance, and leaf dieback in seedlings of two co-occurring Mediterranean shrubs during an intense drought. J Exp Bot 54, 2015-2024.
Watson C, Pulford ID, Riddell‐Black D. 2003. Screening of willow species for resistance to heavy metals: comparison of performance in a hydroponics system and field trials. Int J Phytoremediation 5, 351-365.
Werner C, Ryel RJ, Correia O, Beyschlag W. 2001. Effects of photoinhibition on whole-plant carbon gain assessed with a photosynthesis model. Plant Cell Environ 24, 27-40.
Werner C, Smart JS. 1973. Some new methods of topologic classification of channel networks. Geogr Anal 5, 271-295.
Wierzbicka M, Panufnik D. 1998. The adaptation of Silene vulgaris to growth on a calamine waste heap (S. Poland). Environ Pollut 101, 415-426.
Wilson AJ. 1996. The Living Rock: The Story of Metals Since Earliest Time and Their Impact on Civilization, p. 272. Whitehurst & Clark, Flemington
Wong MH. 2003. Ecological restoration of mine degraded soils, with emphasis on metal contaminated soils. Chemosphere 50, 775-780.
Woolhouse HW. 1983. Toxicity and tolerance in the responses of plants to metals. In: Lange OL, ed. Encyclopedia of Plant Physiology. Berlin, Springer-Verlag. p. 245-300.
Yanai J, Zhao FJ, McGrath SP, Kosaki T. 2006. Effect of soil characteristics on Cd uptake by the hyperaccumulator Thlaspi caerulescens. Environ Pollut 139, 167-75.
Yang HM, Zhang XY, Wang GX, 2004. Effects of heavy metals on stomatal movements in broad bean leaves. Russ J Plant Physiol 51, 464-468.
Yang X, Li T, Yang J, He Z, Lu L, Meng F. 2006. Zinc compartimentation in root, transport into xylem, and absorption into leaf cells in the hyperaccumulating species of Sedum alfredii Hance. Planta 224, 185-195.

Referencias
153
Zobel RW, Kinraide TB, Baligar VC. 2007. Fine root diameters can change in response to changes in nutrient concentrations. Plant Soil 297, 243-254.
Zornoza P, Millán R, Sierra MJ, Seco A, Esteban E. 2010. Efficiency of white lupin in the removal of mercury from contaminated soils: Soil and hydroponic experiments. Journal of Environmental Sciences 22, 421-427.
Zunzunegui M, Barradas MCD, Ain-Lhout F, Alvarez-Cansino L, Esquivias MP, Novo FG. 2011. Seasonal physiological plasticity and recovery capacity after summer stress in Mediterranean scrub communities. Plant Ecol 212, 127-142.


Apéndice I
Metabolome responses of Quercus suber
seedlings to zinc concentration and water availability
Respuestas del metaboloma de plántulas de Quercus suber a Zinc y a distintos niveles de
disponibilidad hídrica
Karen B. Disante, Jordi Cortina, Karin Ljung
Artículo enviado a Journal of Experimental Botany


Metabolome responses of Quercus suber seedlings to zinc concentration and water availability
157
Abstract
In nature, plants are usually subjected to a combination of different sources of stress. The identification of common and specific responses to stress factors, synergies and antagonisms is still controversial. Metabolomics offer a unique framework to explore the interaction between different sources of stress. This study valuates the effect of zinc (Zn) concentration and water availability on the metabolic profile of Quercus suber L. seedlings with the aim to contribute to this ongoing discussion. Seedlings were treated with four levels of Zn from 3 to 150 µM, and exposed to low (LW) and high (HW) watering frequency. The metabolome of Q. suber was affected by Zn supply and water availability. Metabolic profiling revealed the existence of common and specific metabolites. The carbohydrate response to the combination of Zn supply and water availability was metabolite-specific. Aspartate was the only amino acid that was sensitive to the treatments, as proline unexpectedly failed to respond to Zn and water availability. α-tocopherol and polyols were the only metabolites showing a significant Zn x watering interaction. In contrast, the metabolome as a whole was not affected by the interaction between Zn and drought. Results indicate that metabolic response to Zn and watering is highly metabolite-specific. Seedlings response to Zn and water availability involved specific and common metabolites, but a common general response to both types of stress was not evident.
Key words: heavy metal, drought, metabolite profile, abiotic stress, zinc, multiple stress

Apéndice 1
158
Resumen
En la naturaleza, las plantas usualmente están expuestas a una combinación de diferentes factores de estrés. La identificación de respuestas comunes y específicas a diferentes factores, y la existencia de sinergias y antagonismos, continúa siendo discutida. La metabolómica ofrece un marco único para explorar la interacción entre diferentes fuentes de estrés. Este estudio evalúa el efecto del zinc (Zn) y la disponibilidad hídrica sobre el perfil metabólico de plántulas de Quercus suber L. con el objetivo de contribuir a la discusión anterior. Las plántulas fueron tratadas con dosis de Zn entre 3 y 150 µM, y expuesta a baja (LW) y alta (HW) frecuencia de riego. El metaboloma of Q. suber fue sensible al aumento de Zn y a la reducción de la disponibilidad hídrica. El análisis del perfil metabólico mostró la existencia de compuestos comunes y específicos. La respuesta de los carbohidratos a la combinación de Zn y nivel de riego fue específica de cada compuesto. Entre los aminoácidos, el aspartato fue el único sensible a los tratamientos y prolina sorpresivamente no mostró cambios en su acumulación en respuesta a Zn y disponibilidad hídrica. α-tocoferol and polioles fueron los únicos metabolitos que mostraron una interacción significativa entre Zn y disponibilidad hídrica. Al contrario, el metaboloma como un todo, no fue afectado por la interacción de ambos factores. Los resultados indican que la respuesta metabólica a Zn y nivel de riego es característica de cada metabolito. La respuesta de las plántulas a estos factores incluye compuestos comunes y específicos, pero no encontramos evidencias de una respuesta general a ambos factores de estrés.
Palabras clave: metal pesado, sequía, perfil metabólico, estrés abiótico, estrés múltiple

Metabolome responses of Quercus suber seedlings to zinc concentration and water availability
159
1. Introduction
The combination of drought and heavy metal pollution occurs frequently in nature (Derome and Nieminen, 1998; Brady et al., 2005), and it will probably become more widespread as a result of climate change and a worldwide increase in heavy metal mobilization (Peñuelas and Filella, 2002; Sardans and Peñuelas, 2005; IPCC, 2007).
Human activities have contributed to a raise the concentration and bioavailability of zinc (Zn) in natural ecosystems (Kabata-Pendias and Pendias, 1992; Alloway, 1995). As other micronutrients, Zn is required for plants (Marschner, 1995; Cakmak, 2000), but at high concentrations, Zn has toxic effects (Adriano, 2001). Zinc toxicity impairs root growth, photosynthesis, hydraulic conductivity and mineral nutrition (Ruano et al., 1988; Disante et al., 2011; Monet et al., 2001). Most studies on Zn effects on plants have focused on their physiology and morphology. In contrast, information on metabolite response to heavy metal stress is scarce (Sun et al., 2010).
Plant response to drought is complex, involving deleterious effects and adaptive changes. In drylands, drought is frequently accompanied by light, nutrient, heat and salinity stress (Valladares and Percy, 1997; Mittler, 2006; Valdecantos et al., 2006). Under water limitation, turgor pressure is reduced and cell expansion is small (Nielsen and Orcutt, 1996). Stomatal closure is induced and photosynthesis declines (Medrano and Flexas, 2004). Drought disturbs nutrient relations through its effect on nutrient uptake, nutrient translocation and belowground biomass allocation patterns (Chaves et al., 2002; Hu and Schmidhalter, 2005). In addition, several works have showed changes in the metabolic profile of seedlings exposed to drought (Rizhsky et al., 2004; Arbona et al., 2010).
Important attention should be paid to common and specific physiological and molecular responses to abiotic stress as they may provide insights into tolerance mechanisms (Arbona et al., 2010). Thus, organic acids take part in heavy metal chelation and exclusion and have been implied in metal detoxification (Tolrà et al., 1996; Pohlmeier, 2004), and they accumulate as metabolically cheap sources of solutes for osmotic adjustment and cation balance (Nilsen and Orcutt, 1996). Therefore, the concentration of organic acids may increase with the increase in Zn availability and the decrease in water availability. In contrast, soluble carbohydrates as glucose and fructose act as osmoregulators (Nilsen and Orcutt,

Apéndice 1
160
1996), and they frequently increase in water-limited seedlings, whereas they may decrease in seedlings subjected to high Zn availability as photoshyntesis declines (Disante et al., 2011).
Plants are commonly subjected to multiple stresses acting simultaneously (Larcher 2003). Some works have suggested that plants have a centralized response system to different sources of stress, promoting generic morpho-physiological modifications that mitigate their impact. For example, arrested growth and resource acquisition is a common response caused by various sources of stress, such as water scarcity, excess radiation, salinity and heavy metal toxicity (Chapin, 1991; Nilsen and Orcutt, 1996; Larcher, 2003). Similarly, several studies reveal similarities in accumulated metabolites and transcripts under different types of stress (Bohnert et al, 1995; Rizhsky et al., 2004; Arbona et al., 2010). However, plants also develop specific responses to different sources of stress. For example, photolyases specifically restore DNA damage resulting from the exposure to UV-radiation (Britt and Fiscus, 2003), and xanthophylls protect photosystem II from excessive radiation and heat (Davison et al., 2002). The integrated molecular and metabolic response to combined sources of stress is usually unique, and it cannot be easily inferred from plant response to the isolated sources (Valladares and Percy, 1997; Mittler, 2006). Unfortunately, integrative analyses of the consequences of multiple stresses are scarce.
Metabolomics provide a comprehensive and quantitative overview of the metabolites present in an organism, cell or tissue under specific environmental conditions (Hall, 2006; Arbona et al., 2010). Thus, they are particularly suited to study the plasticity of metabolic pathways, and perform integrated assessments of plant response to multiple stresses (Hall, 2006; Bundy et al., 2009).
We performed an experiment to study the metabolic response of Quercus suber L. seedlings to contrasted frequency of irrigation and different Zn concentrations. The objective is to increase our knowledge of the physiological response of Q. suber to both sources of stress, and contribute to the development of an integrative theory of plant response to multiple stresses. To our knowledge, this is the first metabolomic study performed on this species.

Metabolome responses of Quercus suber seedlings to zinc concentration and water availability
161
2. Material and methods
2.1. Plant material and growing conditions
Quercus suber seedlings were cultivated from acorns as described (Disante et al., 2013). Seedlings were grown in 30 cm length PVC tubes filled with silica sand under controlled temperature (22±1°C/18±1°C, day/night), 12 h photoperiod (600 μmol m-2 s-1 of photosynthetic photon flux density during the day) and 50% relative humidity. During this period, seedlings were irrigated with a nutrient solution modified from Arduini et al. (1994) to mimic forest soil conditions (Disante et al., 2013).
2.2. Experimental setup Eight weeks after germination, ZnSO4 was added to the nutrient solution to achieve a Zn concentration of 3 (Control), 10, 50 and 150 μM, corresponding to those commonly found in uncontaminated and contaminated soils (Kabata-Pendias and Pendias, 1992). Then, two irrigation regimes were applied, high and low watering (HW and LW, respectively). HW seedlings were irrigated three times a week (Monday, Wednesday and Friday), whereas LW seedlings were irrigated every 5 days. During the experiment, seedlings were irrigated with 60 ml of nutrient solution. Each Zn level (hereafter treatments) was replicated 10 times, i.e., 10 seedlings per Zn treatments and irrigation level. pH was fixed at 5.5 (the pH of the nutrient solution was adjusted to this value) and quartz was added as the mineral present in the equilibrium. Twelve weeks after the onset of the treatments, when seedlings were 5-months-old, the metabolic profile was assessed (Disante et al., 2013).
2.3. Metabolomics
From each combination of irrigation regime and Zn concentration, 6 leaf samples of 10 mg fresh weight from 6 different seedlings were analysed. Extraction of metabolites, GC/MS analysis, and data processing followed the procedures described in Gullberg et al. (2004) and Jonsson et al. (2005). Metabolites were identified by comparing retention index and mass spectra with those found in the

Apéndice 1
162
Golm metabolome database (http://csbdb.mpimp-golm.mpg.de/gmd.html) and commercially available mass spectral libraries.
2.4. Statistical analysis
We identified 295 metabolites. To reduce the dimensionality of our database and avoid multi-collinearity, we checked the correlation between different metabolites using Spearman correlation coefficients. For highly correlated metabolites (Spearman coefficients above 0.8), only one of them was considered for further analyses (García, 2004).
Treatment effects were evaluated using permutational multivariate analysis of variance (PERMANOVA; Anderson, 2001). This method allows including experimental designs in the analysis of multivariate datasets on the basis of any distance measured using permutation methods. For these analyses, we used the Euclidean distance and 9999 permutations of raw data. The p-values were obtained using a random Monte Carlo sample from the asymptotic distribution of the pseudo F-statistic under permutation (Anderson and Willis, 2003). Additionally, square root transformations were used to increase the influence of rare species (Lepš and Šmilauer, 2003). Whenever differences were detected among treatments, these were examined using pair-wise tests using PERMANOVA test statistic (a multivariate pseudo t-test) and 9999 permutations (see http://goo.gl/0IUh0Q).
To visualize patterns in multivariate data, we used principal coordinates analyses (hereafter PCO) with the same distance measure (see Anderson, 2003 for a detailed description of the method and http://goo.gl/0IUh0Q).
To identify which metabolites contributed to the patterns found, the values on the first two axes of each plot were correlated with the relative intensities of each metabolite using Spearman correlation analysis. Spearman correlation coefficient ranges from -1 to +1, indicating negative or positive associations, respectively. Some metabolites that were significantly correlated with the axes were represented by vectors as examples. The length and direction of each vector indicate the strength and sign, respectively, of the relationship between a given metabolite and the PCO axes.

Metabolome responses of Quercus suber seedlings to zinc concentration and water availability
163
Treatment effects on metabolites that were significantly correlated with PCO axes and those with relevant physiological function were evaluated by using two-way ANOVA.
When the interaction term was significant, one-way ANOVA was used to evaluate the effect of Zn at each watering level. Factor levels were compared pairwise (Tukey’s HSD test) when ANOVA showed a significant treatment effect. Spearman correlation analysis and two-way ANOVA were performed using the SPSS v.15.0 statistical package (SPSS Inc., Chicago, USA).
3. Results
3.1. Metabolic profile
The metabolic profile was affected by Zn and watering (Table 1). In contrast, the interaction between both factors was not significant. Control seedlings (3 μM Zn) showed different metabolic profile than seedlings exposed to the highest Zn dose (150 μM) (Table 2).
Table 1. Results of PERMANOVA analysis to evaluate the effects of Zn and water availability on the metabolome of 5-months-old Quercus suber seedlings Quercus suber seedlings.
Source DF Pseudo-F p
Zn 3 1.58 0.0430
Watering 1 9.42 0.0001
Zn x Watering 1 0.86 0.6626
PCO analysis showed large intra-group dispersion (Fig. 1). The first and the second PCO axes (henceforth PCO1 and PCO2) explained 51% of the total variation. PCO1 accounted for 40% of the variance, while PCO2 explained 11% of the total variation.

Apéndice 1
164
Table 2. Results of pairwise tests comparing the metabolome of 5-months-old Quercus suber seedlings receiving different Zn doses.
Zn concentration (μM)
Pseudo-t test p
3-10 1.10 0.2413-50 1.36 0.0633-150 1.49 0.03050-10 1.13 0.21450-150 0.11 0.24910-150 1.27 0.099
Figure 1. Principal coordinate analysis of the metabolome of Quercus suber seedlings exposed to increasing levels of Zn and water stress. PCO1 and PCO2 indicate the first and the second PCO axes, respectively. Figures in the legend correspond to Zn doses (in µM) and letters to high (HW) and low (LW) frequency of watering. The inserted Figure shows the centroid for each combination of water and Zn availability. Percentage of variance explained by each PCO axis is shown in brackets.

Metabolome responses of Quercus suber seedlings to zinc concentration and water availability
165
Results of the various ANOVA supported what the PCO analysis suggested. Among the amino acids, ornithine showed a significant negative correlation with PCO1. Other amino acids showed a weak correlation with both, PCO1 and PCO2 (Table 3). Most carbohydrates showed a strong negative correlation with PCO1. In contrast, sucrose and fructose were negatively correlated with PCO2. D-xylofuranose correlation with both axes showed opposite sign. Lauric acid was negatively correlated with PCO1 and PCO2, whereas citric, quinic and pentonic acid showed the opposite response. Other acids showed no significant correlation with PCO axes. The response of polyols and lipids was strongly specific. The phytohormone α-tocopherol, was negatively correlated with PCO2.
Threonine showed a trend to increase with the increase in Zn (Supplementary data, Table S1). In contrast, aspartate decreased with the increase in Zn, and it was the only amino acid significantly affected by the treatments (Table 4). There was not a common carbohydrate response to Zn, as their behaviour was highly idiosyncratic (Table 4, Fig 2 A, B y E). Thus, beta-D-methylfructofuranoside increased in response to Zn. In contrast, Zn application decreased the accumulation of the compound identified as a hexose (Fig. 2E, Table 4). In general, other sugars were only affected by water availability (Supplementary data, Table S1).
The compound α-ketoglutaric acid increased with the increase in Zn and was unaffected by watering frequency (Supplementary data, Table S1). Organic acids showed contrasted behavior in response to the treatments. The concentration of citric acid was modified by both stress factors, whereas malic acid decreased with increasing Zn concentration (Table 4), and quinic acid increased under low water availability (Fig. 2D). The lipid, lauric acid, accumulated in leaves in response to Zn, but no to watering frequency (Table 4, Fig. 2J).
The compounds α-tocopherol and xylitol were only affected by the interaction between Zn and water availability but not for the action of the individual factors.
The highest accumulation of α-tocopherol was observed in seedlings receiving 150 µM Zn and exposed to high watering conditions (Tables 4 and 5, Fig. 2F). In contrast, maximum xylitol accumulation was observed in seedlings exposed to 50 µM Zn when water was limiting but the increase was not significant. The

Apéndice 1
166
behaviour of glycerol was modified by watering and by the interaction between both stress factors (Tables 4 and 5, Fig. 2H).
The concentrations of pentonic acid and octadecanoic acid methyl ester were negatively affected by low water availability and remained unchanged under the Zn concentrations evaluated (Supplementary data, Table S1).Metabolites affected by watering frequency, by Zn, or by their combination are showed in a Venn diagram (Fig. 3).

Metabolome responses of Quercus suber seedlings to zinc concentration and water availability
167
PZn= 0.072
PW < 0.001
PZnW= 0.746
PZn= 0.761
PW= 0.010
PZnW= 0.737
PZn= 0.033
PW= 0.003
PZnW= 0.851
PZn= 0.767
PW< 0.001
PZnW= 0.801
PZn= 0.022
PW< 0.001
PZnW= 0.312
PZn= 0.118
PW= 0.748
PZnW= 0.017
Quinic acid
Zn dose (μM)3 10 50 150
Rel
ativ
e co
nten
t (10
4 )
0
100
200
300
400
HWLW
α-Tocopherol
Zn dose (μM)
3 10 50 150
Rel
ativ
e co
nten
t (10
4 )
0
2
4
6
8
10
HWLW
Glucose
Zn dose (μM)3 10 50 150
Rel
ativ
e co
nten
t (10
4 )
0
10
20
30
40
50
HW LW
Citric acid
Zn dose (μM)3 10 50 150
Rel
ativ
e co
nten
t (10
4 )
0
1
2
3
4
HWLW
Zn dose (μM)3 10 50 150
Rel
ativ
e co
nten
t (10
4 )
0
5
10
15
20
HW LW
Fructose
Zn dose (μM)3 10 50 150
Rel
ativ
e co
nten
t (10
4 )
0
1
2
3
4
HW LW
Hexose
A B
DC
E F
Figure 2. Relative content of representative metabolites in leaves of Quercus suber seedlings growing at four Zn concentrations and two contrasting water availability levels (HW and LW for high and low watering frequency, respectively). Bars are means and standard errors of N=5 seedlings. Note the different scales used for different compounds. Different letters indicate significant differences at P < 0.05 (Tukey’s HSD test). (Figure continued on next page).
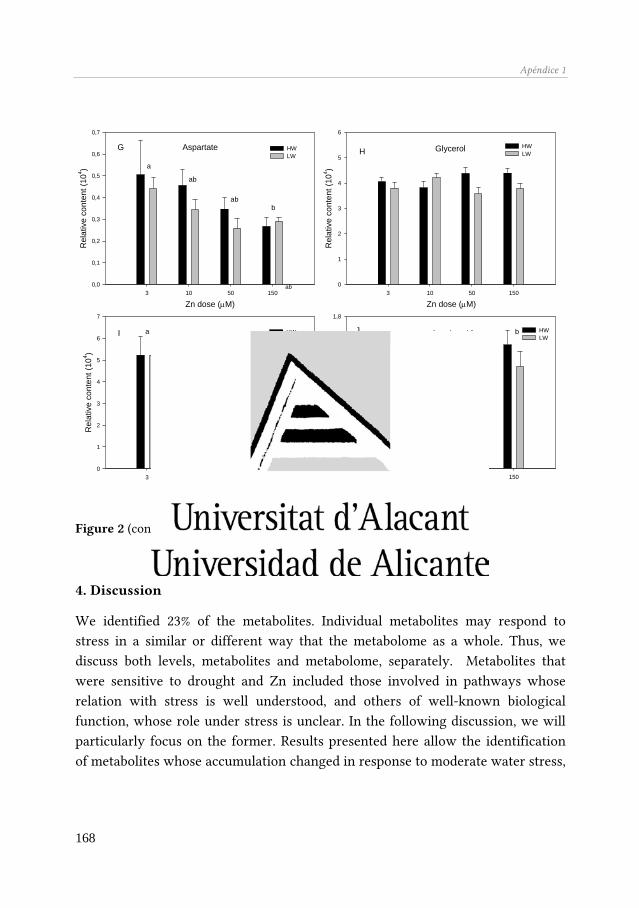
Apéndice 1
168
Aspartate
Zn dose (μM)3 10 50 150
Rel
ativ
e co
nten
t (10
4 )
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
HW LW
Glycerol
Zn dose (μM)3 10 50 150
Rel
ativ
e co
nten
t (10
4 )
0
1
2
3
4
5
6
HW LW
Malic acid
Zn dose (μM)3 10 50 150
Rel
ativ
e co
nten
t (10
4 )
0
1
2
3
4
5
6
7
HW LW
Lauric acid
Zn dose (μM)3 10 50 150
Rel
ativ
e co
nten
t (10
4 )
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
HW LW
a
ab
abb
a
ab abb
ab a
ab
b
ab
G H
I J
Figure 2 (continuation)
4. Discussion
We identified 23% of the metabolites. Individual metabolites may respond to stress in a similar or different way that the metabolome as a whole. Thus, we discuss both levels, metabolites and metabolome, separately. Metabolites that were sensitive to drought and Zn included those involved in pathways whose relation with stress is well understood, and others of well-known biological function, whose role under stress is unclear. In the following discussion, we will particularly focus on the former. Results presented here allow the identification of metabolites whose accumulation changed in response to moderate water stress,

Metabolome responses of Quercus suber seedlings to zinc concentration and water availability
169
Table 3. Spearman correlation coefficient between the first two axes of the PCO analysis, and identified compounds or unidentified compounds whose correlation coefficient was above 0.5. (*) P < 0.05 and (**) P < 0.01. UI: unidentified compound.
Compounds PCO1 PCO2 Compounds PCO1 PCO2 Amino acids Acids and hidroxi-acids
Aspartate 0.11 -0.16 Citric acid 0.59* -0.29**
Glycine 0.06 0.16 Malic acid -0.05 -0.20
Ornithine -0.49* -0.06 Quinic acid -0.27** 0.58*
Proline -0.03 0.04 Succinic acid 0.10 0.17
Serine 0.15 0.03 Pentonic acid 0.35* -0.32*
Threonine 0.18 0.14 Polyols
Pyroglutamic acid -0.02 -0.04 Xylitol 0.29** -0.41*
Carbohydrates Glycerol 0.29* -0.14
Glucose -0.99* 0.06 Glycosides
Fructose -0.84* -0.30* Methyl-β-D-G2 0.16 -0.28*
Sucrose -0.21 -0.57* Lipids
Talose -0.98* 0.10 Hexadecanoic acid 0.14 0.00
Mannose -0.87* 0.15 Lauric acid -0.17 -0.40*
Rhamnose -0.79* 0.08 O-acid methyl ester3 0.12 -0.29*
D-Xylofuranose -0.54* 0.40* Phytohormones
D-Xylopyranose -0.23 0.02 α-Tocopherol -0.19 -0.35*
β-D-M1 -0.29** -0.23 Unidentified
Putative hexose -0.62* 0.03 UI 1 0.80** -0.03
Isomaltose 0.18* -0.15 UI 2 0.30* -0.51**
Myoinositol -0.09 -0.22 UI 3 -0.19 -0.29**
Ketone UI 4 0.17 -0.11
α-Ketoglutaric acid 0.20 -0.02 UI 5 0.04 -0.18
UI 6 -0.56** -0.12 1β-D-Methylfructofuranoside
2Methyl-β-D-galactopyranoside 3Octadecanoic acid methyl ester

Apéndice 1
170
to Zn, or to their interaction. The different behaviour are summarized in Fig. 3 and discussed below. Metabolomics represent a powerful tool to perform integrative evaluations of plant response to multiple stresses that occur in the field. It is important to note that metabolite identification is hampered by the lack of reference standards for many of the secondary metabolites, and the use of model species to construct databases (Hall, 2006).
4.1. Metabolic response to contrasted watering frequency
In contrast to our expectations, amino acid concentration barely changed in water stressed plants compared to well irrigated plants. Proline accumulation in response to drought has been described for several species (Zunzunegui et al. 2011). Various studies suggest that proline could lessen dehydration damage. In addition, proline accumulation may contribute to increase nitrogen and carbon reserves to be used once water deficit is relieved (Alia et al., 1997; Larcher, 2003). In our experiment, proline concentration was not affected by low frequency of irrigation, probably because seedlings were exposed to moderate water stress and not severe drought. Proline accumulation may only occur below a threshold in leaf water potential (Ain-Lhout et al., 2001).
Most monosaccharides were correlated with PCO1, and they were sensitive to water availability. Indeed, the correlation coefficient between the relative content of glucose and PCO1 was very close to -1. The accumulation of water soluble carbohydrates and organic nitrogen compounds via synthesis, conversion of starch and translocation has been described in plants under drought and other sources of stress (Morgan, 1984; Larcher, 2003). This process delays turgor loss and enhances carbon assimilation, especially in arid regions (Larcher, 2003). Soluble sugars may act as source of carbon for maintenance and re-growth during recovery of the photosynthetic apparatus following short periods of drought (Chaves et al., 2002).
Organic acids as malate and citrate may also contribute to osmoregulation (Morgan, 1984). In our experiment, the former was not affected by changes in watering frequency. Citric acid, in contrast, decreased in seedlings exposed to low frequency of watering. Similarly, a study in Q. suber showed that citric acid concentration was very low during a hot dry summer (Passarinho et al., 2006).

Metabolome responses of Quercus suber seedlings to zinc concentration and water availability
171
Table 4. Results of two-way ANOVA to evaluate the effects of Zn and water availability (W) on the metabolic profile of 5-months-old Quercus suber seedlings.
Compounds Zn W ZnW
Amino acids
Aspartate F= 3.00, P= 0.043 F= 0.53, P= 0.470 F= 1.25, P= 0.305
Proline F= 0.70, P= 0.559 F= 0.03, P=0.861 F = 0.71, P=0.554
Carbohydrates
Glucose F= 2.52, P=0.072 F= 35.27, P< 0.001 F= 0.41, P= 0.746
Rhamnose F= 2.73, P=0.058 F= 25.35, P< 0.001 F= 0.12, P= 0.949
Fructose F= 0.39, P= 0.761 F= 13.88, P=0.001 F= 0.42, P= 0.737
Sucrose F=0.63, P= 0.598 F= 1.77, P= 0.191 F = 0.57, P= 0.641
β-D-M1 F=7.88 P< 0.001 F= 0.31, P= 0.583 F = 1.90, P= 0.147
Putative hexose F=3.60 P= 0.022 F= 24.81, P< 0.001 F= 1.23, P= 0.312
Acids and hidroxi-acids
Citric acid F=3.02, P= 0.033 F= 9.88, P= 0.003 F= 0.26, P= 0.851
Malic acid F= 3.02, P=0.041 F= 0.00, P= 0.988 F= 0.33, P= 0.805
Quinic acid F= 0.38, P= 0.767 F= 21.91, P< 0.001 F= 0.33, P= 0.801
Polyols
Xylitol F= 1.78, P= 0.167 F= 0.16, P= 0.690 F= 3.15, P= 0.035
Glycerol F= 0.22, P= 0.884 F= 4.60, P= 0.038 F= 2.89, P= 0.048
Lipids
Lauric acid F= 3.21, P= 0.034 F= 0.02, P= 0.901 F= 1.17, P= 0.333
Phytohormones
α-tocopherol F= 2.08, P= 0.118 F= 0.10, P= 0.748 F= 3.824, P= 0.017 1 β-D-Methylfructofuranoside
We must take into account that other acids, as quinic acid, may dominate the metabolome and play a stronger protective role in Quercus species, than malate or citrate (Gebre and Tschaplinski, 2002; Passarinho et al., 2006).

Apéndice 1
172
Figure 3. Venn diagram showing the metabolites of Quercus suber seedlings affected by contrasted watering frequency (left), Zn doses (right) or by the interaction between both factors (intersection) (P < 0.05; Tukey’s HSD test). Metabolites affected by both factors but not for their interaction are showed above of the dotted line. Asterisks indicate significant differences at P < 0.1 (Tukey’s HSD test).
4.2. Metabolic response to Zn
Fructose and, particularly, sucrose showed a negative relationship with PCO2. In our experiment, carbohydrate response to Zn was weak and metabolite-specific. Indeed, the concentration of most carbohydrates showed no significant changes when seedlings were exposed to Zn. Conversely, the monosaccharides glucose and rhamnose showed a trend towards an increase in concentration with the increase in Zn availability. The increase in foliar carbohydrate concentration after exposure to heavy metals has been widely described in the literature (Samarakoon and Rauser, 1979; Jha and Dubey, 2004; Ghnaya et al., 2010), and has been related to alterations in the behaviour of starch and sugar metabolising enzymes, and restricted transport of photoassimilates (Samarakoon and Rauser, 1979; Jha and Dubey, 2004).

Metabolome responses of Quercus suber seedlings to zinc concentration and water availability
173
Other authors have observed changes in foliar amino acid concentration in response to Zn, especially proline (Schat et al., 1997; Sharma and Dietz, 2006). However, proline accumulation in these studies was associated to water deficit induced by heavy metals (Schat et al., 1997; Sharma and Dietz, 2006). In our study, we found no significant proline accumulation in response to low water availability (see above). Besides, in a previous experiment, we observed no signal of water stress in Q. suber, when Zn was applied at the same experimental doses as in the current experiment (Disante et al., 2011).
Table 5. Results of one-way ANOVA to evaluate the effects of Zn on metabolites of 5-months-old Quercus suber seedlings subjected to high and low watering frequency (HW and LW, respectively).
Compounds FHW PHW FLW PLW
Xylitol F= 0.65 0.590 F= 0.60 0.621
Glycerol F= 1.68 0.206 F= 1.46 0.254
α-tocopherol F= 6.66 0.003 F= 0.29 0.829
The metabolite α-ketoglutaric acid showed a positive response to Zn supply. Although several amino acids (Glu, Gln, Asp, Asn) are derived from this compounds, to our knowledge, a possible role for α-ketoglutaric acid in heavy metal stress response remain unknown.
Organic acids as citric and malic acid may act as chelators inside the cell, converting heavy metals into their non-toxic forms (Pohlmeier, 2004). In addition, organic acids exudation could play a role in alleviating Al toxicity in soil (Ma and Furukawa, 2003) Increases in the accumulation of citric and malic acid after exposition to Zn have been observed in Zn-tolerant species, and species and ecotypes with a strong capacity to accumulate heavy metals (Harmens et al., 1994; Yang et al., 2006; Sagardoy et al., 2011). However, several studies have found no changes or a reduction in the concentration of organic acids in plants exposed to heavy metals (Harmens et al., 1994; Monni et al., 2001, Sun et al., 2010).We found that the concentration of organic acids was indeed reduced in plants exposed to Zn. This reduction could be due to the decreased photosynthetic activity in seedling exposed to Zn (Disante et al., 2013).

Apéndice 1
174
α-tocopherol is the major vitamin E compound found in leaf chloroplasts. This compound has antioxidant properties and, in cooperation with other antioxidants, plays a key role in defending chloroplasts against oxidative stress (Munné-Bosch and Alegre, 2003; Munné-Bosch 2005). Under environmental stress, tocopherol levels first increase, preventing oxidative damage, and then decrease, when stress is severe and degradation exceeds synthesis (Munné-Bosch 2005). Others works have proposed that tocopherols may protect PSII from photoinhibition (Havaux et al., 2005). This agrees when previous results were we found symptoms of photoinhibition only in seedlings exposed to 150 µM Zn and HW conditions and suggesting that photosynthesis-derived reactive oxygen species (ROS) were induced (Disante et al., 2011).
4.3. Metabolome response to Zn and watering frequency
Many stress-induced metabolic responses confer protection against a single stress. However, several studies suggest the existence of a general adaptation syndrome by which different types of stress evoke identical coping mechanisms (Leshem et al., 1998). The general response of the metabolome to a combination of stress may coincide or not with the response to each type of stress acting separately (Rizhsky et al., 2004; Arbona et al., 2010). The metabolite profile of Q. suber was dependent on Zn dose and water availability. Decreased watering frequency was the principal factor inducing alterations in the metabolic profile. The influence of water availability on the metabolite profile has been previously documented (Rizhsky et al., 2004; Shulaev et al., 2008). Conversely, studies analyzing changes in the metabolome in response to heavy metals are scarce, especially in woody species.
Seedling separation following watering regimes was observed along PCO1 axis, with HW and LW samples occupying opposite sides of the graph. PCO2 axis explained slightly more than 10% of the total variation. This axis was associated with Zn. However, seedling distribution based on Zn levels showed high dispersion. It is interesting to note that the centroid of samples exposed to 50 µM Zn LW was closer to the centroid of samples exposed to low Zn than to those exposed to the highest Zn dose.

Metabolome responses of Quercus suber seedlings to zinc concentration and water availability
175
In contrast to our expectations, we found no overall significant interaction between both stress factors, thus revealing the absence of global synergistic or antagonistic relations. In a previous work, we found that Zn effects on morpho-physiological traits of Q. suber seedlings were controlled by water availability (Disante et al., 2013). Disagreement between morpho-physiological and metabolic responses is not surprising, as plant response to stress initiates at sub-cellular levels and progress upwards (Hall, 2006). Morpho-physiological changes result from complex alterations in the metabolism and the signalling processes, and interactions may come out as emergent properties (Gibson, 2000; Potters et al., 2009).
The lack of interaction between Zn and drought suggests that metabolome responses to Zn and drought were independent at this scale. However, when examined in detail, we found that some metabolites responded differently when both sources of stress acted together. In some cases, the combination of Zn and watering strengthened the response of the individual metabolite to a single stress factor in a synergistic way. Thus, the accumulation of glucose, citric acid and rhamnose increased when both factors were combined, compared to the independent effect of Zn and drought. We may note that glucose concentration in seedlings subjected to Zn and drought was 28 times that of control seedlings. In other cases, Zn and drought had opposite effects, and the interaction was antagonistic. This was the case of the metabolite identified as hexose. Furthermore, glycerol and, particularly, the antioxidant α-tocopherol increased when seedlings were subjected to the highest Zn dose, but this only happened when water was not limiting. Finally, maximum xylitol accumulation occurred in low watered seedlings exposed to 50 µM Zn.
As we mention previously, the results presented and the discussion are based in a subset of metabolites of Q. suber. Therefore, other relevant relationships than those showed here could exist between other compounds not included in these analysis and the PCO axes.

Apéndice 1
176
5. Conclusions
The metabolome as a whole responded differentially to water and heavy metal stress, showing no clear evidence of synergistic or antagonistic responses. At a lower level, some metabolites responded to a single stress factor, and may be considered as potential indicators of specific stress. In contrast, other metabolites were affected by both stress factors. In some cases, the interaction between both sources of stress was synergistic, and may be part of a common stress-coping strategy in Q. suber. In other cases, the interaction was antagonistic. More else, the response of a third group of metabolites was conditioned by the combined levels of both stress factors. Thus, results partly support the existence of a general strategy to cope with abiotic stress, but they also show that this is only part of a complex array of common and specific responses that are activated by Zn and drought. Our study suggests that conclusions on global responses to stress based on a limited set of metabolites should be drawn with care, and emphasize the need to use descriptive statistics and statistical inference on large metabolomic data sets. Metabolomics are a valuable tool to investigate the pathways involved in specific and general responses to multiple stress, and will contribute to the refinement of multiple stress theory.
Acknowledgements
We thank Jose Huesca for their assistance in the lab, David Fuentes and Santiago Soliveres for their comments on an earlier draft, Banco de Semillas El Serranillo for the seeds used in this experiment and the University of Alicante for an FPU (Formacion del Profesorado Universitario) grant awarded to K. Disante. This research was funded by the Spanish Ministry of Science and Innovation (Project GRACCIE, Programa Consolider-Ingenio 2010 (CSD 2007-00067).

Metabolome responses of Quercus suber seedlings to zinc concentration and water availability
177
Supplementary material
Results of two-way ANOVA of metabolites not included in Table 4.
Table S1. Results of two-way ANOVA to evaluate the effects of Zn and water availability (W) on the metabolic profile of 5-months-old Quercus suber seedlings.
Compounds Zn W ZnW
Amino acids
Glycine F=1.20, P=0.324 F=0.01, P=0.955 F =0.29, P=0.833
Ornithine F=1.13, P=0.348 F=1.74, P=0.194 F=1.10, P=0.361
Serine F=1.33, P=0.279 F=1.08, P=0.304 F =1.62, P=0.200
Threonine F=2.26, P=0.096 F=0.26, P=0.610 F =1.56, P=0.214
Pyroglutamic acid F=1.47, P=0.239 F=0.38, P=0.539 F =0.23, P=0.873
Carbohydrates
Talose F=0.93, P=0.434 F=37.62, P<0.001 F =0.328, P=0.805
Mannose F=1.83, P=0.157 F=28.23, P<0.001 F =0.76, P=0.521
D-Xylofuranose F=2.03 P=0.125 F=8.54, P=0.006 F =1.43, P=0.248
D-Xylopyranose F=1.22, P=0.314 F=0.52, P=0.476 F =0.50, P=0.686
Isomaltose F=0.22, P=0.883 F=4.01, P=0.052 F =0.52, P=0.672
Myoinositol F=1.56, P=0.215 F=0.39, P=0.534 F =1.31, P=0.286
Ketone
α-Ketoglutaric acid F=3.20, P=0.034 F=0.28, P=0.599 F =0.88, P=0.460
Acids and hidroxi-acids
Succinic acid F=0.53, P=0.666 F=0.81, P=0.374 F =0.39, P=0.761
Pentonic acid F=1.27, P=0.296 F=6.96, P=0.012 F =1.25, P=0.304
Lipids
Hexadecanoic acid F=1.79, P=0.165 F=2.55, P=0.118 F =1.33, P=0.277
O-acid methyl ester1 F=0.54, P=0.658 F=3.88, P=0.056 F =0.77, P=0.515 1Octadecanoic acid methyl ester

Apéndice 2
178
Apéndice 2. Relación entre concentración de Zn foliar y variables de respuesta ecofisiológica
Para estimar la reducción del 35% de cada variable ecofisiológica, graficamos los valores obtenidos para cada variable en función de la concentración foliar de Zn determinada (Capítulos III y IV). Las curvas resultantes fueron ajustadas a los modelos presentados en la Tabla 1, donde “y” representa el 35% del valor máximo obtenido para cada variable (Capítulos III y IV).
Tabla 1. Modelos que describen la relación entre la concentración de Zn en hojas de Quercus suber y diferentes variables ecofisiológicas evaluadas. CT35: Concentración de Zn en tejido foliar que induce una reducción del 35% en el valor de diversas variables ecofisiológicas respecto al valor máximo observado.
Variable CT35 R2 F P
A ((11,2x314,6)/y)-314,6 0,57 8,15 0,029
gs ((14,1x191,1)/y)-191,1 0,70 17,61 0,006
Fv:Fm (89,1 ln((0,7/y)-1)+437,7 0,96 63,53 <0,001
IAA (ln(y/6,5))/0,002 0,92 22,79 0,041
KR-RA -1,8/((1/y)-0,2) 0,99 58,39 0,092
Longitud radicular (ln(y/1,4))/-0,002 0,54 6,95 0,038
Biomasa radicular
91,0+251,9(Raíz(2,7/y-1)) 0,69 4,57 0,093
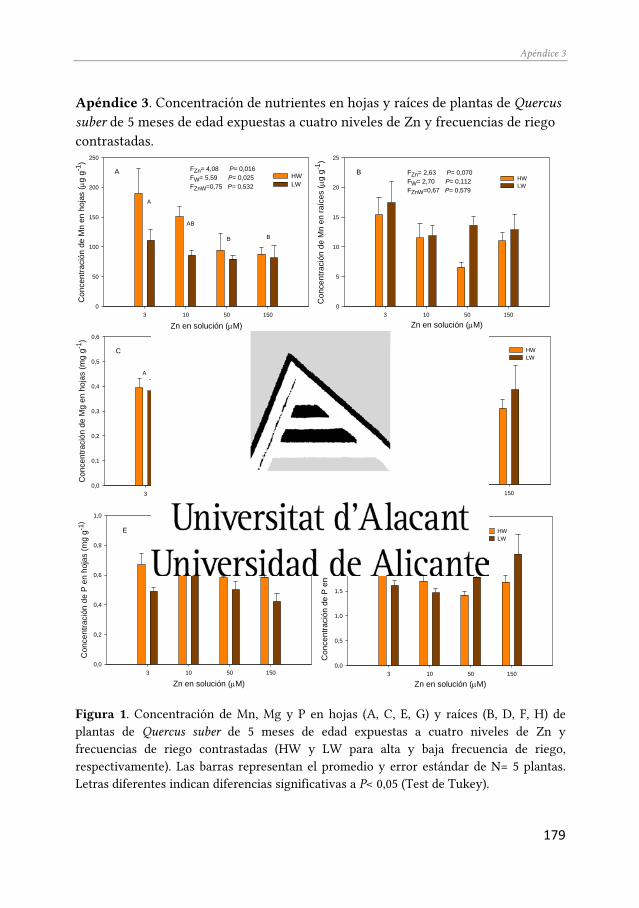
Apéndice 3
179
Apéndice 3. Concentración de nutrientes en hojas y raíces de plantas de Quercus suber de 5 meses de edad expuestas a cuatro niveles de Zn y frecuencias de riego contrastadas.
Zn en solución (μM)3 10 50 150
Con
cent
raci
ón d
e M
n en
hoj
as (μ
g g-1
)
0
50
100
150
200
250
HWLW
Zn en solución (μM)3 10 50 150
Con
cent
raci
ón d
e M
n en
raíc
es (μ
g g-1
)
0
5
10
15
20
25
HWLW
FZn= 2,63 P= 0,070FW= 2,70 P= 0,112FZnW=0,67 P= 0,579
FZn= 4,08 P= 0,016FW= 5,59 P= 0,025FZnW=0,75 P= 0,532
A
AB
B B
Zn en solución (μM)3 10 50 150
Con
cent
raci
ón d
e M
g en
raíc
es (m
g g-1
)
0,00
0,05
0,10
0,15
0,20
0,25
0,30
HWLW
Zn en solución (μM)3 10 50 150
Con
cent
raci
ón d
e M
g en
hoj
as (m
g g-1
)
0,0
0,1
0,2
0,3
0,4
0,5
0,6
HWLW
FZn= 3,30 P= 0,035FW= 0,45 P= 0,505FZnW=0,39 P= 0,764
AAB
AB
B
FZn= 1,78 P= 0,174FW= 0,01 P= 0,933FZnW=0,74 P= 0,536
A B
C D
Zn en solución (μM)3 10 50 150
Con
cent
raci
ón d
e P
en
hoja
s (m
g g-1
)
0,0
0,2
0,4
0,6
0,8
1,0
HWLW
Zn en solución (μM)3 10 50 150
Con
cent
raci
ón d
e P
en
raíc
es (m
g g-1
)
0,0
0,5
1,0
1,5
2,0
2,5
3,0
HWLWFZn= 1,78 P= 0,173
FW= 0,71 P= 0,405FZnW= 1,61 P= 0,209
FZn= 2,17 P= 0,114FW= 0,13 P= 0,725FZnW= 3,85 P= 0,020
E F
Figura 1. Concentración de Mn, Mg y P en hojas (A, C, E, G) y raíces (B, D, F, H) de plantas de Quercus suber de 5 meses de edad expuestas a cuatro niveles de Zn y frecuencias de riego contrastadas (HW y LW para alta y baja frecuencia de riego, respectivamente). Las barras representan el promedio y error estándar de N= 5 plantas. Letras diferentes indican diferencias significativas a P< 0,05 (Test de Tukey).

180
Zn en solución (μM)3 10 50 150
Con
cent
raci
ón d
e K
en
hoja
s (m
g g-1
)
0
2
4
6
8
HWLW
Zn en solución (μM)3 10 50 150
Con
cent
raci
ón d
e K
en ra
íces
(mg
g-1)
0
1
2
3
4
5
6
HWLW
(μ ) (μ )
G HFZn= 0,53 P= 0,662FW= 0,03 P= 0,868FZnW= 2,03 P= 0,132
FZn= 0,37 P= 0,771FW= 2,83 P= 0,104FZnW= 0,48 P= 0,699
Figura 1. (Continuación)
Dado el efecto significativo de la interacción Zn x nivel de riego sobre la concentración de fósforo en raíces (Fig 1F), realizamos un ANOVA de un factor para cada nivel de riego con el fin de evaluar el efecto del Zn sobre esta variable. La concentración radicular de P aumentó con el aumento de Zn en raíces de plantas LW (F= 3,58 P= 0,041) y mostró una tendencia a disminuir en plantas HW (F= 2,59, P= 0,095).

Apéndice 4
181
Apéndice 4. Normativa sobre metales pesados
Debido al incremento de la concentración de metales en suelo, muchos países han establecido umbrales de toxicidad y de intervención para la recuperación de suelos contaminados. Por ejemplo, Holanda ha desarrollado una legislación rigurosa donde quedan especificados valores objetivo y valores umbrales a partir de los cuales se ha de intervenir para la restauración. Los valores objetivo indican el nivel en el que hay una calidad del suelo o agua sostenible. En otras palabras, establecen el nivel que tiene que ser logrado para recuperar completamente las propiedades de un suelo funcional para los seres humanos, y la vida vegetal y animal. La ley holandesa también establece valores de fondo y niveles de intervención para suelos y aguas subterráneas, diferenciando además, aguas superficiales y profundas (Tabla 1).
La normativa sobre suelos contaminados en España se establece en la Ley 22/2011, de 28 de julio, de Residuos y suelos contaminados y en el Real Decreto 9/2005, de 14 de enero (BOE Nº 15 del 14 de enero de 2005).
La Ley 22/2011, de 28 de julio, de Residuos y Suelos Contaminados contempla la siguiente definición de “Suelo contaminado”: aquel cuyas características han sido alteradas negativamente por la presencia de componentes de carácter peligroso procedentes de la actividad humana, en concentración tal que comporte un riesgo inaceptable para la salud humana o el medio ambiente, de acuerdo con los criterios y estándares que se determinen por el Gobierno, y así se haya declarado mediante resolución expresa.
Los aspectos esenciales recogidos en ambas normas son:
1. La definición de un conjunto de actividades económicas consideradas como potencialmente contaminantes del suelo.
2. El establecimiento de un régimen de información en materia de suelos contaminados entre los titulares de las actividades potencialmente contaminantes y las administraciones regionales competentes.
3. Los criterios por los que un suelo puede ser declarado como contaminado.
4. La obligación, con carácter general, para los causantes de la contaminación o propietarios de los suelos afectados de acometer tareas de descontaminación.

Apéndice 4
182
Las Comunidades Autónomas, conforme a dicha norma, tienen la obligación de elaborar un inventario de los suelos contaminados existentes en su ámbito territorial así como una relación priorizada de éstos. A partir de esta información, el Ministerio de Agricultura, Alimentación y Medio Ambiente elaborará el inventario estatal de suelos contaminados.
El nivel genérico de referencia (NGR) es la concentración de una sustancia contaminante en el suelo que no conlleva un riesgo superior al máximo aceptable para la salud humana (en función del uso del suelo) o los ecosistemas. Es decir, es la concentración máxima de dicha sustancia en el suelo que nos permite asegurar que el riesgo que supone su presencia es aceptable.
El Real Decreto 9/2005, de 14 de enero no establece valores NGR para metales aplicables al ámbito nacional, aunque sí fija los procedimientos para calcular los valores NGR para la protección de la salud humana y los valores NGR para la protección de los ecosistemas. Dichos valores deben establecerse a escala regional en base a los niveles de fondo geoquímicos correspondientes (BOE Nº 15 del 14 de enero de 2005). Únicamente algunas Comunidades Autónomas han elaborado el inventario de estos niveles. En aquellas Comunidades Autónomas que aún no hayan calculado los valores NGR para metales, se propone un método para estimarlos mediante el valor correspondiente a la concentración media de una sustancia en suelos sobre zonas no contaminadas próximas y con sustratos geológicos de similares características, más dos veces la desviación estándar.
Una fuente común de entrada de metales pesados en suelos agrícolas y forestales (bien sea por acciones de restauración o por cambios de uso) son las aplicaciones de residuos orgánicos. En este sentido la legislación es menos difusa y de aplicación obligada. La Directiva comunitaria 86/278/CEE, regula las condiciones en que se pueden aplicar los lodos de depuradora a los suelos agrícolas, y su transposición a la legislación española se efectuó mediante el Real Decreto 1310/1990, donde se estipula un nivel máximo de metales pesados en los suelos receptores (en base a dos niveles de pH) y en los lodos (siempre con un cierto nivel de pre-tratamiento). Para otros productos orgánicos estos límites se recogen de manera general en el Real Decreto 824/2005, de 8 de julio, sobre productos fertilizantes, modificado posteriormente en la Orden PRE/630/2011, de 23 de marzo. En estos dos Decretos quedan excluidos los sustratos o soportes de cultivos, que se incluyen en un nuevo Real Decreto 865/2010, de 2 de julio donde,

Apéndice 4
183
entre otros aspectos, se establece un nivel máximo de metales pesados (Fuentes, 2011).
Tabla 1. Valores límites de Zn para diferentes compartimentos y ámbitos de aplicación
Valor determinado
Ámbito de aplicación Compartimento Concentración de Zn
Valor de fondo Nacional (Holanda) Suelo / sedimento 140 mg/Kg
Valor objetivo Nacional (Holanda) Suelo / sedimento 140 mg/kg
Valor de intervención
Nacional (Holanda) Suelo / sedimento 720 mg/Kg
Valor objetivo Nacional (Holanda) Aguas superficiales1 65 µg/l
Valor de fondo Nacional (Holanda) Aguas superficiales1 24 µg/l
Valor objetivo Nacional (Holanda) Aguas subterráneas2 24 µg/l
Valor de intervención
Nacional (Holanda) Aguas subterráneas2 800 µg/l
NGR para ecosistemas
Autonómico (Cataluña)
Suelo (zona agropecuaria y forestal) 170 mg/ kg
NGR para ecosistemas
Autonómico (Galicia)
Suelo 200 mg/ Kg
NGR para ecosistemas
Autonómico (Aragón)
Suelo (otros usos3) 420 mg/ Kg
RD. 1310/1990 Nacional (España) Suelo pH<74 150
RD. 1310/1990 Nacional (España) Suelo pH>74 450
1 < 10 metros. 2 > 10 metros. 3 Suelos que, no siendo ni urbano ni industrial, son aptos para el desarrollo de actividades agrícolas, forestales y ganaderas. 4 Concentración máxima en los suelos sobre los que podrán aplicarse lodos de depuradora.

Apéndice 4
184
El incremento de metales pesados en suelos Mediterráneos es un hecho probado y descrito en la bibliografía (Andreu y Gimeno, 1996; Peris et al., 2008). En los últimos años se está haciendo un esfuerzo considerable en distintos ámbitos para lograr la protección de los suelos. Prueba de ello son las diferentes medidas adoptadas y el establecimiento de niveles límites (Tabla 1).
Aún así quedan muchas cuestiones pendientes. Es importante destacar que, a nivel práctico, la ley española establece normas para establecer un suelo como contaminado, pero no distingue entre valores de referencia de suelo y valores en la solución del suelo o valores biodisponibles. En general, para establecer un valor de referencia se utiliza el contenido total de metales en suelo, el cual proporciona una información incompleta sobre la peligrosidad de la contaminación por metales pesados (Adriano et al., 2004; González et al., 2011). Dependiendo de las propiedades físico-químicas del mismo, la concentración biodisponible de un metal, que en definitiva es la que determinará su riesgo potencial, puede variar dentro de un rango relativamente amplio. Así mismo, la climatología local debería ser tenida en consideración ya que, como hemos visto en esta tesis, la disponibilidad hídrica puede alterar los valores umbrales de toxicidad. En este sentido, cualquier factor (textura, propiedades edáficas, régimen hídrico, etc.) que pueda potenciar o amortiguar los efectos nocivos de los metales, debería ser considerado en la estimación de los valores genéricos de referencia correspondientes.
Leyes sobre metales pesados citadas en la Tabla 1
Real Decreto 1310/1990. http://xurl.es/bowod
Ley 22/2011, de 28 de julio, de residuos y suelos contaminados. http://xurl.es/ceflg
Real Decreto 9/2005, de 14 de enero. http://xurl.es/zenb2
Niveles genéricos de referencia (NGR). Catalunya http://xurl.es/ey4ml
Niveles genéricos de referencia de metales pesados y otros elementos traza en suelos de Galicia. http://xurl.es/xjwzx
Determinación de niveles de fondo y niveles de referencia para metales en suelos de la Comunidad Autónoma de Aragón. http://xurl.es/t2s2k
Dutch Target and Intervention Values, 2000. http://xurl.es/9lx8n