seasonal succession of phototrophic biofilms in an … · the structural integrity and development...
TRANSCRIPT

AQUATIC MICROBIAL ECOLOGYAquat Microb Ecol
Vol. 45: 301–312, 2006 Published December 21
INTRODUCTION
Biomass attached at the interface between water andsubstrata interferes with many ecological, biogeochem-ical, and biotechnological and environmental engineer-ing processes (Battin et al. 2003, Larson & Passy 2005).This attached biomass is commonly called biofilm, isoften heterogeneous and multi-layered, and may con-tain minerals or abiotic materials that are trapped byextracellular products within the biofilm or releasedfrom cells as the biofilm ages (Sutherland 2001). Thestructure of the biofilm is maintained by ‘glue-like’
extracellular polymeric substances (EPS) secreted by avariety of organisms (Stal 2000). Cyanobacteria andeukaryotic microalgae such as diatoms and green algaeare the major phototrophic components of biofilms infreshwater ecosystems (Stevenson 1996).
Cyanobacteria secrete EPS as capsules and sheathsthat surround cells and filaments as amorphousmucilage in the medium and also as slime for glidingmotility (De Philippis & Vincenzini 1998). Diatom EPSproduction is commonly associated with aggregation(physical coagulation; Thornton 2002) and colony for-mation (mucilage pads, fibrils etc.) in pelagic species
© Inter-Research 2006 · www.int-res.com*Email: [email protected]
Seasonal succession of phototrophic biofilms in anItalian wastewater treatment plant: biovolume,
spatial structure and exopolysaccharides
Roberta Congestri1,*, Francesca Di Pippo1, Roberto De Philippis3, Isabella Buttino4, Gaio Paradossi2, Patrizia Albertano1
1Department of Biology, and 2Department of Chemical Sciences and Technologies, University of Rome ‘Tor Vergata’,Via della Ricerca Scientifica 1, 00133 Rome, Italy
3Department of Agricultural Biotechnology, University of Florence, Piazzale delle Cascine 24, 50144 Florence, Italy4Laboratory of Ecophysiology, Stazione Zoologica ‘A. Dohrn’, Villa Comunale, 80121 Naples, Italy
ABSTRACT: A multiphasic approach was applied to investigate the structural features of photo-trophic biofilms that grow in a wastewater treatment plant (WWTP) at Fiumicino Airport (Rome,Italy). Seasonal variations in species composition, biomass and exopolysaccharides produced wereanalyzed by light (LM) and electron microscopy (SEM), high performance liquid chromatography(HPLC) and circular dichroism (CD). Phototroph contribution to the 3-dimensional structure of thebiofilm and its development was assessed by confocal laser scanning microscopy. Analysis of biofilmsgrown on polypropylene slides showed a stable species composition; seasonal changes in biomasswere mostly due to changes of major cyanobacterial and algal taxonomic groups. Extensive growthwas evident on the range of artificial substrata that were implanted in the treatment plant. CD spec-tra and HPLC analyses of 2 operationally defined exopolysaccharide fractions extracted from samplesscraped off the tank walls revealed that negatively charged heteropolysaccharides comprised most ofthe matrix and capsular components of the biofilms. Cytochemical staining distinguished betweenacidic and sulphated residues in the samples observed by LM. The data provide a new insight intothe structural integrity and development of phototrophic biofilms in this hyper-eutrophic environ-ment, indicating a potential use of autochthonous consortia in an environmentally sound tertiarywater treatment alternative to conventional chemico-physical technologies.
KEY WORDS: Phototrophic biofilms · Exopolysaccharides · Cyanobacteria · Algae · Biovolume · Confocal laser scanning microscope · Wastewater treatment plant
Resale or republication not permitted without written consent of the publisher

Aquat Microb Ecol 45: 301–312, 2006
and with substrate adhesion (stalks, tubes) and loco-motion in benthic forms (Hoagland et al. 1993). Manyrepresentatives of green algae possess cellulose or gly-coprotein cell walls and capsules (van den Hoek et al.1995, Shubert 2003), several colonial forms are embed-ded in mucilage matrix, while mucilage is extruded bydesmids for gliding (Nultsch & Hader 1988). In addi-tion, many planktic forms release dissolved organicmaterial, which comprises a significant fraction ofpolysaccharides (Kaplan 1987, Lombardi & Vieria1999, Lombardi et al. 2005).
Overall, EPS are involved in adhesion to the sub-strata and cohesion between the cells of a biofilm; theyplay a role in surface-associated motility and provideprotection against desiccation and grazing by preda-tors, producing a matrix network that embeds cells anddetritus (Wingender et al. 1999, Decho 2000). Theseexopolymers provide a microenvironment where UV-absorbing pigments and proteins, including enzymes,are immobilized (Sutherland 2001). The major compo-nents of the matrix are polysaccharides, but proteinsand nucleic acids can comprise a significant part(Decho 2000). Extracellular polysaccharides may alsoserve in the immobilization and accumulation of nox-ious compounds, acting as a natural molecular sieve oran ion exchanger of xenobiotics and toxins; thus, theexploitation of biofilms may be particularly useful forbioremediation purposes (Craggs et al. 1996, Wingen-der et al. 1999). However, the potential application ofspontaneously developing phototrophic biofilm inwastewater treatment plants (WWTP) has been largelyneglected (Hoffmann 1998). In addition, there havebeen relatively few studies of the species compositionand biomass production of these photosynthetic micro-bial communities (Davis et al. 1990a,b, Sláde<ková1994, Sláde<ková & Matulová 1998). Overall, little isknown about the spatial organization, development orEPS characteristics of biofilms in out-door systems. Itmust be noted that EPS polysaccharide research in thefield is often hampered by many external processesthat interfere with their persistence inthe environment, such as scouring,irreversible adsorption to sedimentparticles and degradation by variousmicroorganisms. The situation is fur-ther complicated in that the EPSmatrix originates from different organ-isms, producing more than one type ofpolymer that differ in saccharide unitcomposition and sequence (Neu 1994).The synthesis of these polymers mayalso significantly vary in response toenvironmental factors and cell status(De Philippis & Vincenzini 1998, Staatset al. 1999, Smith & Underwood 2000,
de Brouwer et al. 2002, Otero & Vincenzini 2004,Underwood et al. 2004, Barranguet et al. 2005, Stal &Défarge 2005).
The aim of this study was to investigate the structuralintegrity, potential productivity and matrix polysac-charide characteristics of phototrophic biofilms colo-nizing a southern temperate WWTP (Fiumicino AirportWWTP, Rome, Italy). A variety of microscopic andchemico-physical techniques were used to determineseasonal changes in samples taken directly from theWWTP tank walls (providing high biomass for exo-polysaccharide studies) or from immersed polypropyl-ene slides (for biovolume estimates). Non-destructivesampling of biofilm material for confocal light scanningmicroscope (CLSM) was achieved by hanging artificialsubstrata in the overflow system of the tank.
MATERIALS AND METHODS
The Fiumicino WWTP (Leonardo da Vinci Airport) isan out-door system designed to process municipalwastewaters with an estimated inflow and outflow of6000 m3 d–1, and uses oxygen injections in the primarytank and constant stirring in the secondary tank toreduce biological oxygen demand (BOD).
Samples were collected seasonally (May, August,November 2001; February, May and August 2002;Table 1) by scraping biofilms from the concrete walls ofthe sedimentation tank (ST) and from immersedpolypropylene slides. Two slides (8 × 3 cm) wereplaced on the ledge of the tank (northwest unshadedside) below the weirs of the overflow system (waterdepth 1 to 2 cm) for 1 wk to allow colonization. ForCLSM analyses of biofilm structure, intact communi-ties were examined on 3 different artificial substrata,immersed as described above, for 1 or 2 wk in May,August, November 2001 and February 2002 (Table 1).
Phototrophic composition and biomass. Biofilmswere scraped off the polypropylene slides after incuba-
302
May 01 Aug 01 Nov 01 Feb 02 May 02 Aug 02
TW CD CD CD CDCytochem HPLC HPLC HPLC
Cytochem Cytochem Cytochem
AS Biovolume Biovolume Biovolume BiovolumeCLSM/LM CLSM/LM CLSM/LM CLSM/LM
SEM SEM SEM SEM
Table 1. Summary of sampling and analysis conducted during the study. TW:tank walls; AS: artificial substrata; CD: circular dichroism; Cytochem: cyto-chemical staining of exopolysaccharides; Biovolume: calculation of biovolume;CLSM: confocal light scanning microscopy; LM: light microscopy; SEM:
scanning electron microscopy

Congestri et al.: Phototrophic biofilms in a wastewater treatment plant
tion in the ST for 1 wk periods. The period of immer-sion was chosen to allow a similar period of coloniza-tion on the artificial substrata as on the tank walls. Thetank walls were routinely brushed to remove biofilmsat weekly intervals. Preliminary studies were made todetermine the area required to incorporate total varia-tion of the community comprising the biofilm. Macro-scopically, it was not possible to observe any variationand the samples appeared to be homogenous. Micro-scopic observations of intact biofilms using CLSMrevealed some patchiness; however, this was on amicro-scale. It was therefore decided to take 1 cm2
scrapings from the slides for 2 reasons: to safely incor-porate all possible variation within biofilms, and toreduce any error in obtaining a quantitatively accuratesample. The sampled biofilms were suspended in0.1 M phosphate buffer (pH 7.2), fixed in 2% formalde-hyde and 2.5% glutaraldehyde, and stored at 4°C.
Fixed samples were examined using a Zeiss Axio-skop light microscope equipped with differential inter-ference contrast (DIC) and 40 × and 100 × objectives.Further ultrastructural studies for diatom identificationwere conducted on acid-cleaned material (von Stosch1974) using scanning electron microscopy (SEM;Philips XL30 field emission scanning microscope) at5.0 kV.
To determine estimates of cell biovolume, the scrap-ings were sonicated twice for 3 min in a sonic waterbath in order to disaggregate and homogenize thesamples. Aliquots of sonicated suspensions wereappropriately diluted in phosphate buffer and left tosettle for 24 h in 25 ml counting chambers. Observa-tions were performed using a Nikon Eclipse Te200inverted microscope with a 40 × objective. Imageanalysis equipment (Nikon CoolSnap video-cameraand LUCIA software) was used to acquire optical fieldsand digital images. Semi-automated measurements ofselected morphometric parameters were used to esti-mate the biovolume of single cells using standardizedequations (Hillebrand et al. 1999).
Biofilm architecture. Polypropylene slides, What-man Anodisc filters and coverslips were used to visual-ize structure and seasonal development of intactbiofilms on different substrata. Two periods of immer-sion (1 or 2 wk) were chosen to analyze the temporalsuccession and development of biofilms. The substratawere retrieved and immediately fixed with 2%formaldehyde spreading fixative with a syringe in aPetri dish. Observations were conducted using a ZeissLSM 410 CLSM equipped with an argon ion laser and10, 25 and 40 × objectives. Visualization of spatial dis-tribution of phototrophs, biofilm development andarchitecture was achieved at the excitation wave-length of chl a (488 nm with emission at 575 to 650 nm).Optical sections, taken at different depth of the
biofilms, were acquired and combined using the Zeisssoftware package LSMDummy to obtain verticalreconstruction of samples.
Extraction of extracellular polysaccharides. The ex-traction of extracellular polysaccharides was con-ducted following Bellezza et al. (2003). Biofilm samplesof about 1000 g were scraped off the ST walls and cen-trifuged at 8000 rpm for 60 min (J2-21 Beckman cen-trifuge) to concentrate the sample. The resulting pellet,comprising the microorganisms and their envelopes(namely the ‘capsular’ [CPS] or ‘bound polysaccha-rides’), was then separated into 2 fractions. One frac-tion was re-suspended in phosphate buffer (pH 7, 1:10volume) and incubated at 4°C for 7 d. The suspensionwas then centrifuged (8000 rpm for 15 min) and the su-pernatant precipitated in absolute EtOH (1:1 volume).After further centrifugation (14 000 rpm for 60 min), thepellet was re-suspended in distilled water. Finally, thesupernatant was dialyzed against EDTA (0.01 M) andNaCl (0.5 M) for 2 h and then against distilled water for4 d. This procedure is referred to as ‘cold extraction’,and generated a cold extracted capsular polysaccha-ride fraction (CPSc). The second fraction of the initialpellet was at first re-suspended in distilled water (1:10)and incubated at 80°C for 60 min. Dialysis (as above)proceeded this ‘hot extraction’ and the obtained ex-tract was operationally referred to as CPSh.
Monosaccharide composition. Lyophilized CPScand CPSh samples were analyzed for their monosac-charide composition after hydrolysis with 2 N trifluo-roacetic acid (120°C for 45 min), according to theHPLC method described by Vincenzini et al. (1990a).
Circular dichroism analyses. CPSc and CPSh sam-ples were analyzed by circular dichroism (CD) in orderto investigate transition of polysaccharide molecularconformations as a function of pH and temperature,using a Jasco Spectropolarimeter J600 equipped withquartz cells of 0.1 cm, an optical path-length primarilyin the UV spectral region (200 to 300 nm) and between200 and 270 nm (20 nm min–1 scanning velocity), andthe original Jasco software. The pH of the 2 fractionswas first decreased with HClO4 (0.02 M) and thenincreased with NaOH (0.02 M). The pH values weremeasured using a calibrated digital combined pH-meter (Amel Instrumments, 334-B). Temperature of the2 fractions was increased from 25 to 60°C in singleincrements of 5 to 10°C, and was achieved using athermostat (Lauda M3)-controlled cell holder.
Exopolysaccharide cytochemistry. Biofilm sampleswere scraped off the tank walls for light microscopy(LM) observation (Zeiss Axioskop) after cytochemicalstaining using (1) Alcian Blue (AB) at pH 0.5 and 2.5 tovisualize the sulphated and carboxylated groups, re-spectively, and (2) Ruthenium Red (RR) to observe theacidic polysaccharides (Albertano & Bellezza 2001).
303
Aquat Microb Ecol 45: 301–312, 2006
RESULTS
Composition and seasonal distribution of phototrophic biomass
Phototrophs constituted the major components ofbiofilms grown seasonally on polypropylene slidesimmersed for 1 wk periods in the Fiumicino WWTP ST.These phototrophs were essentially composed ofcyanobacteria, diatoms and green algae (Table 2).Observations of the phototrophic community composi-tion were based on species identifications from previ-ous studies of biofilms in these tanks (Albertano et al.1999, Congestri et al. 2003, 2005). A variety ofChroococcalean cyanobacteria, either unicellular orcolonial forms, were distinguished along with 6 Oscil-latorialean taxa mostly belonging to Oscillatoria andPhormidium spp. In total, 15 diatom taxa were identi-fied. Raphid forms prevailed, with only 1 centric(Cyclotella meneghiniana Kützing) and 1 araphid
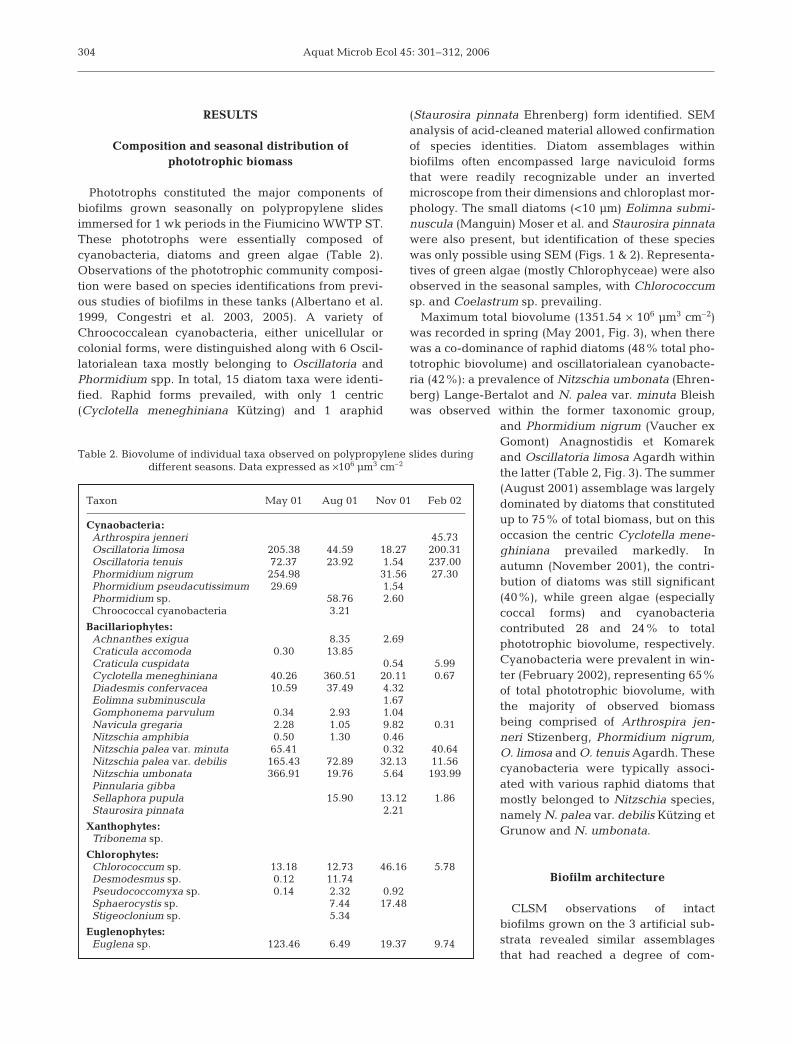
(Staurosira pinnata Ehrenberg) form identified. SEManalysis of acid-cleaned material allowed confirmationof species identities. Diatom assemblages withinbiofilms often encompassed large naviculoid formsthat were readily recognizable under an invertedmicroscope from their dimensions and chloroplast mor-phology. The small diatoms (<10 µm) Eolimna submi-nuscula (Manguin) Moser et al. and Staurosira pinnatawere also present, but identification of these specieswas only possible using SEM (Figs. 1 & 2). Representa-tives of green algae (mostly Chlorophyceae) were alsoobserved in the seasonal samples, with Chlorococcumsp. and Coelastrum sp. prevailing.
Maximum total biovolume (1351.54 × 106 µm3 cm–2)was recorded in spring (May 2001, Fig. 3), when therewas a co-dominance of raphid diatoms (48% total pho-totrophic biovolume) and oscillatorialean cyanobacte-ria (42%): a prevalence of Nitzschia umbonata (Ehren-berg) Lange-Bertalot and N. palea var. minuta Bleishwas observed within the former taxonomic group,
and Phormidium nigrum (Vaucher exGomont) Anagnostidis et Komarekand Oscillatoria limosa Agardh withinthe latter (Table 2, Fig. 3). The summer(August 2001) assemblage was largelydominated by diatoms that constitutedup to 75% of total biomass, but on thisoccasion the centric Cyclotella mene-ghiniana prevailed markedly. Inautumn (November 2001), the contri-bution of diatoms was still significant(40%), while green algae (especiallycoccal forms) and cyanobacteriacontributed 28 and 24% to totalphototrophic biovolume, respectively. Cyanobacteria were prevalent in win-ter (February 2002), representing 65%of total phototrophic biovolume, withthe majority of observed biomassbeing comprised of Arthrospira jen-neri Stizenberg, Phormidium nigrum,O. limosa and O. tenuis Agardh. Thesecyanobacteria were typically associ-ated with various raphid diatoms thatmostly belonged to Nitzschia species,namely N. palea var. debilis Kützing etGrunow and N. umbonata.
Biofilm architecture
CLSM observations of intactbiofilms grown on the 3 artificial sub-strata revealed similar assemblagesthat had reached a degree of com-
304
Taxon May 01 Aug 01 Nov 01 Feb 02
Cynaobacteria:Arthrospira jenneri 45.73Oscillatoria limosa 205.38 44.59 18.27 200.31Oscillatoria tenuis 72.37 23.92 1.54 237.00Phormidium nigrum 254.98 31.56 27.30Phormidium pseudacutissimum 29.69 1.54Phormidium sp. 58.76 2.60Chroococcal cyanobacteria 3.21
Bacillariophytes:Achnanthes exigua 8.35 2.69Craticula accomoda 0.30 13.85Craticula cuspidata 0.54 5.99Cyclotella meneghiniana 40.26 360.51 20.11 0.67Diadesmis confervacea 10.59 37.49 4.32Eolimna subminuscula 1.67Gomphonema parvulum 0.34 2.93 1.04Navicula gregaria 2.28 1.05 9.82 0.31Nitzschia amphibia 0.50 1.30 0.46Nitzschia palea var. minuta 65.41 0.32 40.64Nitzschia palea var. debilis 165.43 72.89 32.13 11.56Nitzschia umbonata 366.91 19.76 5.64 193.99Pinnularia gibbaSellaphora pupula 15.90 13.12 1.86Staurosira pinnata 2.21
Xanthophytes:Tribonema sp.
Chlorophytes:Chlorococcum sp. 13.18 12.73 46.16 5.78Desmodesmus sp. 0.12 11.74Pseudococcomyxa sp. 0.14 2.32 0.92Sphaerocystis sp. 7.44 17.48Stigeoclonium sp. 5.34
Euglenophytes:Euglena sp. 123.46 6.49 19.37 9.74
Table 2. Biovolume of individual taxa observed on polypropylene slides duringdifferent seasons. Data expressed as ×106 µm3 cm–2
Congestri et al.: Phototrophic biofilms in a wastewater treatment plant
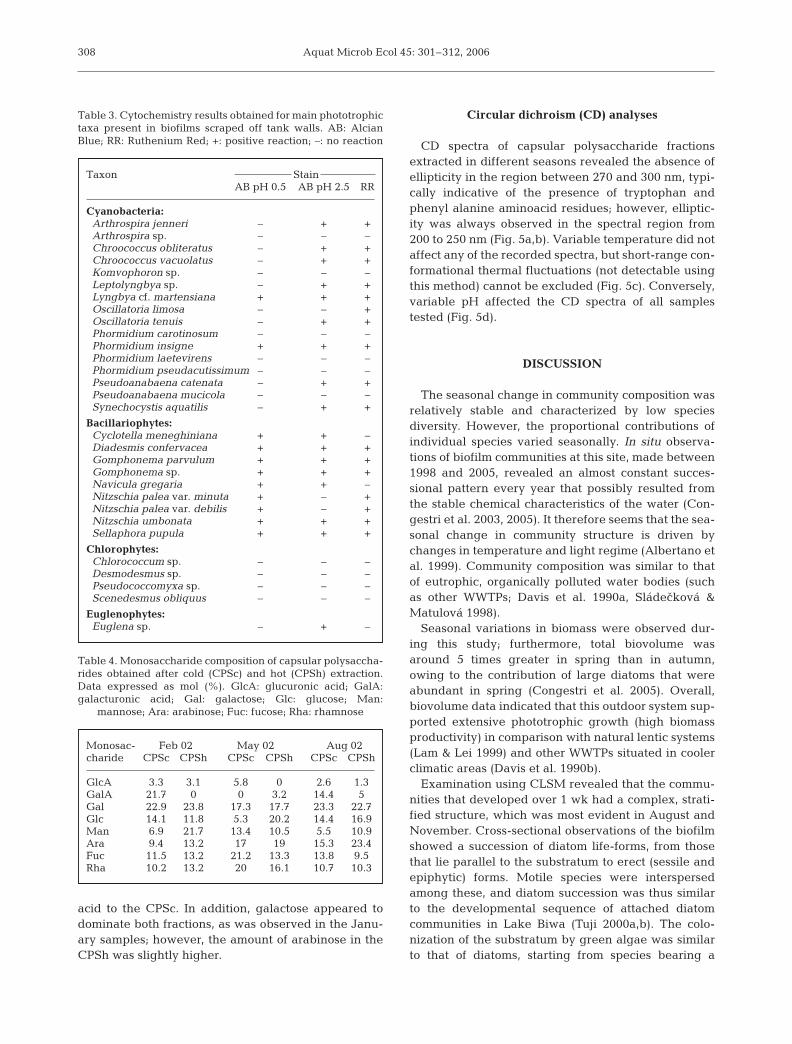
plexity within 7 d. Maximum biofilm thickness after a2 wk incubation period (~260 µm) was recorded inautumn (November 2001) on polypropylene slides.No obvious taxonomic or structural differences wereobserved among assemblages grown on the substratatested; however, spatial distribution of organisms wasrelatively more patchy on coverslips owing to thesloughing of filamentous and ramified forms. The col-onization of each substratum normally resulted inpatches of differing thickness, and combined epifluo-rescence and transmitted light observations alsorevealed the presence of voids. The mature, 1 wk-oldcommunities exhibited obvious developmental strati-fication of microorganisms (Fig. 4a,b). This exposuretime allowed development of forms sometimes adher-ent and parallel to the substratum (adnate diatomforms such as Achanthes spp., other diatoms,
Cyclotella meneghiniana, and the coccal green algaChlorococcum sp.), followed by erect, sessile taxa(stalked diatom species such as Gomphonema spp.and Encyonema minutum [Hilse] D. G. Mann, and fil-amentous and/or ramified green algae as Stigeoclo-nium sp. and Cladophora sp.). Tangled within thesefixed forms were the chain-forming diatomsDiadesmis confervacea Kützing and Staurosira pin-nata, in addition to filamentous oscillatorialeancyanobacteria. Motile diatoms such as Craticula acco-moda (Hustedt) D.G. Mann, Navicula gregariaDonkin and Nitzschia spp. were interspersed in thebiofilms, either occupying the very inner or the outer-most layers. The 2 wk-old communities showed fullcomplexity and horizontal heterogeneity. There weremore microorganisms colonizing the substrata thanwere recorded in the 1 wk-old community, and strati-
305
Fig. 1. SEM micrographs of (a) Cyclotella meneghiniana, external view; (b,c) Staurosira pinnata (b) outer and (c) inner valvesurface; (d) Achnanthes exigua, external view; and (e,f) Gomphonema sp. (e) outer surface and (f) detail of the apical pore field.
Scale bars: (a) 5 µm; (b,c,d,f) 2 µm; (e) 10 µm

Aquat Microb Ecol 45: 301–312, 2006
fication and successional stages were apparent (fromadherent forms to sessile, epiphytic taxa; Fig. 4c,d).However, this was not evident in winter biofilmsgrown on any type of substrate, which were charac-terized by very low diversity and low stratification(Fig. 4e,f). Seasonal variation in the architecture ofthe biofilm reflected changes in the compositionalpatterns: the highest degree of complexity, spatialheterogeneity and layering in phototroph distributionwas observed in November 2001.
Exopolysaccharide cytochemistry
Light microscopy after cytochemical staining of sam-ples scraped off tank walls showed that there were sul-phated (AB, pH 0.5) carboxylated (AB, pH 2.5) and
acidic groups (RR) present in the biofilm (Table 3). ABpH 0.5 treatment did not reveal sulphated residues inchroococcal cyanobacteria, whereas 2 oscillatorialeans(Lyngbya cf. martensiana Meneghini and Phormidiuminsigne [Skuja] Anagnostidis et Komarek) gave a posi-tive reaction. Sulphated groups were found in alldiatoms, but were absent from the green algae. AB pH2.5 staining revealed carboxyl groups in the envelopesof all chroococcalean and in about 50% of oscillatori-alean cyanobacteria. COOH residues were not presentin the green algae, but they were present in alldiatoms, except for the 2 varieties of Nitzschia palea.RR treatment revealed acidic polysaccharides for allchroococcalean cyanobacteria, in about 50% of oscilla-torialean forms, and in all the diatoms exceptCyclotella meneghiniana and Navicula gregaria.Green algae did not give a positive reaction to RR.
306
Fig. 2. SEM micrographs of (a,b) Diadesmis confervacea, (a) girdle view and (b) outer valve surface; (c) Eolimna subminuscula,external view; (d) Craticula accomoda, external view; (e) Nitzschia palea var. minuta apical areas in external and internal view;
and (f) Nitzschia umbonata larger valve, internal surface. Scale bars: (a) 5 µm; (b,c,d,e) 2 µm; (f) 10 µm

Congestri et al.: Phototrophic biofilms in a wastewater treatment plant
Extracellular monosaccharide composition
All fractions contained various proportions of 6 dif-ferent neutral monosaccharides and at least 1 uronicacid (Table 4). In February, galactose was the mainsugar in both fractions, followed by mannose in theCPSh and by galacturonic acid in the CPSc. The lattersugar was not found in the CPSh. In May, the contribu-tion of galactose decreased in both fractions, whereasconcentrations of glucose and of fucose and rhamnosewere prevalent in the CPSh and CPSc, respectively.The CPSc fraction contained the highest amount ofglucuronic acid in all the polymers analyzed, whereasthis sugar was not present in the CPSh. In contrast, theCPSh contained galacturonic acid, which was notdetected in the CPSc. Both uronic acids were presentin August, with a high contribution of galacturonic
307
1%
1%9% 18%
%
1%
%
8%
65%
33%1%
48%
1%9%42%
75
6%
24%
40%
28
8%
Cyanobacteria Diatoms Green algae Euglena sp.
Fig. 3. Contribution (%) of major taxonomic groups to totalphototroph biovolume in seasonal biofilm samples scraped off
polypropylene slides
Fig. 4. Confocal light scanning microscope images illustrating vertical reconstructions of biofilms on artificial substrata. Novembersamples grown for 1 wk on (a) polypropylene slides and (b) coverslips, with voids visible; 2 wk stratified biofilms from (c) summerand (d) autumn samples, and winter communities dominated by cyanobacteria on (e) polypropylene slides and (f) Anodisc
membrane filters. Scale bars: 30 µm
Aquat Microb Ecol 45: 301–312, 2006
acid to the CPSc. In addition, galactose appeared todominate both fractions, as was observed in the Janu-ary samples; however, the amount of arabinose in theCPSh was slightly higher.
Circular dichroism (CD) analyses
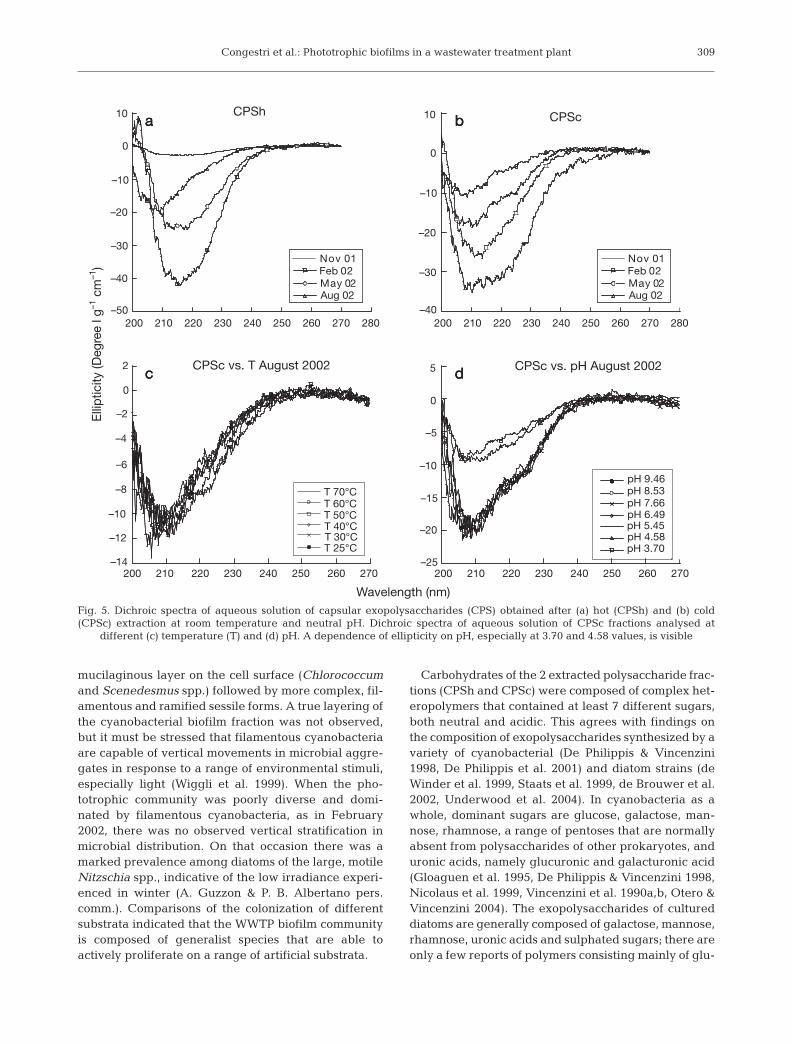
CD spectra of capsular polysaccharide fractionsextracted in different seasons revealed the absence ofellipticity in the region between 270 and 300 nm, typi-cally indicative of the presence of tryptophan andphenyl alanine aminoacid residues; however, elliptic-ity was always observed in the spectral region from200 to 250 nm (Fig. 5a,b). Variable temperature did notaffect any of the recorded spectra, but short-range con-formational thermal fluctuations (not detectable usingthis method) cannot be excluded (Fig. 5c). Conversely,variable pH affected the CD spectra of all samplestested (Fig. 5d).
DISCUSSION
The seasonal change in community composition wasrelatively stable and characterized by low speciesdiversity. However, the proportional contributions ofindividual species varied seasonally. In situ observa-tions of biofilm communities at this site, made between1998 and 2005, revealed an almost constant succes-sional pattern every year that possibly resulted fromthe stable chemical characteristics of the water (Con-gestri et al. 2003, 2005). It therefore seems that the sea-sonal change in community structure is driven bychanges in temperature and light regime (Albertano etal. 1999). Community composition was similar to thatof eutrophic, organically polluted water bodies (suchas other WWTPs; Davis et al. 1990a, Sláde<ková &Matulová 1998).
Seasonal variations in biomass were observed dur-ing this study; furthermore, total biovolume wasaround 5 times greater in spring than in autumn,owing to the contribution of large diatoms that wereabundant in spring (Congestri et al. 2005). Overall,biovolume data indicated that this outdoor system sup-ported extensive phototrophic growth (high biomassproductivity) in comparison with natural lentic systems(Lam & Lei 1999) and other WWTPs situated in coolerclimatic areas (Davis et al. 1990b).
Examination using CLSM revealed that the commu-nities that developed over 1 wk had a complex, strati-fied structure, which was most evident in August andNovember. Cross-sectional observations of the biofilmshowed a succession of diatom life-forms, from thosethat lie parallel to the substratum to erect (sessile andepiphytic) forms. Motile species were interspersedamong these, and diatom succession was thus similarto the developmental sequence of attached diatomcommunities in Lake Biwa (Tuji 2000a,b). The colo-nization of the substratum by green algae was similarto that of diatoms, starting from species bearing a
308
Taxon StainAB pH 0.5 AB pH 2.5 RR
Cyanobacteria:Arthrospira jenneri – + +Arthrospira sp. – – –Chroococcus obliteratus – + +Chroococcus vacuolatus – + +Komvophoron sp. – – –Leptolyngbya sp. – + +Lyngbya cf. martensiana + + +Oscillatoria limosa – – +Oscillatoria tenuis – + +Phormidium carotinosum – – –Phormidium insigne + + +Phormidium laetevirens – – –Phormidium pseudacutissimum – – –Pseudoanabaena catenata – + +Pseudoanabaena mucicola – – –Synechocystis aquatilis – + +
Bacillariophytes:Cyclotella meneghiniana + + –Diadesmis confervacea + + +Gomphonema parvulum + + +Gomphonema sp. + + +Navicula gregaria + + –Nitzschia palea var. minuta + – +Nitzschia palea var. debilis + – +Nitzschia umbonata + + +Sellaphora pupula + + +
Chlorophytes:Chlorococcum sp. – – –Desmodesmus sp. – – –Pseudococcomyxa sp. – – –Scenedesmus obliquus – – –
Euglenophytes:Euglena sp. – + –
Table 3. Cytochemistry results obtained for main phototrophictaxa present in biofilms scraped off tank walls. AB: Alcian Blue; RR: Ruthenium Red; +: positive reaction; –: no reaction
Monosac- Feb 02 May 02 Aug 02charide CPSc CPSh CPSc CPSh CPSc CPSh
GlcA 3.3 3.1 5.8 0 2.6 1.3GalA 21.7 0 0 3.2 14.4 5Gal 22.9 23.8 17.3 17.7 23.3 22.7Glc 14.1 11.8 5.3 20.2 14.4 16.9Man 6.9 21.7 13.4 10.5 5.5 10.9Ara 9.4 13.2 17 19 15.3 23.4Fuc 11.5 13.2 21.2 13.3 13.8 9.5Rha 10.2 13.2 20 16.1 10.7 10.3
Table 4. Monosaccharide composition of capsular polysaccha-rides obtained after cold (CPSc) and hot (CPSh) extraction.Data expressed as mol (%). GlcA: glucuronic acid; GalA:galacturonic acid; Gal: galactose; Glc: glucose; Man:
mannose; Ara: arabinose; Fuc: fucose; Rha: rhamnose
Congestri et al.: Phototrophic biofilms in a wastewater treatment plant
mucilaginous layer on the cell surface (Chlorococcumand Scenedesmus spp.) followed by more complex, fil-amentous and ramified sessile forms. A true layering ofthe cyanobacterial biofilm fraction was not observed,but it must be stressed that filamentous cyanobacteriaare capable of vertical movements in microbial aggre-gates in response to a range of environmental stimuli,especially light (Wiggli et al. 1999). When the pho-totrophic community was poorly diverse and domi-nated by filamentous cyanobacteria, as in February2002, there was no observed vertical stratification inmicrobial distribution. On that occasion there was amarked prevalence among diatoms of the large, motileNitzschia spp., indicative of the low irradiance experi-enced in winter (A. Guzzon & P. B. Albertano pers.comm.). Comparisons of the colonization of differentsubstrata indicated that the WWTP biofilm communityis composed of generalist species that are able toactively proliferate on a range of artificial substrata.
Carbohydrates of the 2 extracted polysaccharide frac-tions (CPSh and CPSc) were composed of complex het-eropolymers that contained at least 7 different sugars,both neutral and acidic. This agrees with findings onthe composition of exopolysaccharides synthesized by avariety of cyanobacterial (De Philippis & Vincenzini1998, De Philippis et al. 2001) and diatom strains (deWinder et al. 1999, Staats et al. 1999, de Brouwer et al.2002, Underwood et al. 2004). In cyanobacteria as awhole, dominant sugars are glucose, galactose, man-nose, rhamnose, a range of pentoses that are normallyabsent from polysaccharides of other prokaryotes, anduronic acids, namely glucuronic and galacturonic acid(Gloaguen et al. 1995, De Philippis & Vincenzini 1998,Nicolaus et al. 1999, Vincenzini et al. 1990a,b, Otero &Vincenzini 2004). The exopolysaccharides of cultureddiatoms are generally composed of galactose, mannose,rhamnose, uronic acids and sulphated sugars; there areonly a few reports of polymers consisting mainly of glu-
309
Aug 02
a b
c d
May 02
a
Nov 01Feb 02
a bb
cc dd
Elli
ptic
ity (D
egre
e l g
–1 c
m–1
)10
0
–10
–20
–30
–40
–50
CPSh
200 210 220 230 240 250 260 270 280 200 210 220 230 240 250 260 270 280
10
0
–10
–20
–30
–40
CPSc
CPSc vs. T August 2002 CPSc vs. pH August 20022
0
–2
–4
–6
–8
–10
–12
–14
5
0
–5
–10
–15
–20
–25200 210 220 230 240 250 260 270 200 210 220 230 240 250 260 270
Wavelength (nm)
T 70°CT 60°CT 50°CT 40°CT 30°CT 25°C
pH 8.53pH 7.66pH 6.49pH 5.45pH 4.58pH 3.70
pH 9.46
Aug 02May 02
Nov 01Feb 02
Fig. 5. Dichroic spectra of aqueous solution of capsular exopolysaccharides (CPS) obtained after (a) hot (CPSh) and (b) cold(CPSc) extraction at room temperature and neutral pH. Dichroic spectra of aqueous solution of CPSc fractions analysed at
different (c) temperature (T) and (d) pH. A dependence of ellipticity on pH, especially at 3.70 and 4.58 values, is visible

Aquat Microb Ecol 45: 301–312, 2006
cose (Hoagland et al. 1993, de Winder et al. 1999, Staatset al. 1999, de Brouwer & Stal 2002, Bellinger et al.2005). Mannose followed by glucuronic acid (Lombardiet al. 2005) and mannose associated with rhamnose(Lombardi & Vieria 1999) was found to comprise a largeproportion (>40%) of the exopolysaccharides of 2chlorococcalean green algae; in contrast, fucose andglucuronic acid predominated in strains of desmids(Domozych et al. 1993, Paulsen & Vieira 1994). Growthstatus and environmental conditions can affect thecomposition of polysaccharides; however, HPLC datawere in accordance with compositional patterns of sea-sonal samples. There were high concentrations ofgalacturonic acid in February and arabinose in August,which coincided with the dominance of cyanobacteriaand diatoms, respectively. Culture studies on the domi-nant cyanobacteria in the Fiumicino WWTP revealedthat production of arabinose is very low (Gloaguen etal. 1995, 1996); therefore, the high concentration of ara-binose in our samples was most likely a result of diatomdominance.
CD characterization of biofilm exopolysaccharides inaqueous solution indicated that proteic moieties wereabsent from all tested fractions, demonstrating thatsaccharide extraction did not damage the cells. Ellip-ticity observed in the region of 200 to 250 nm wasattributable to n → π* electronic transition of carbonylgroups, and confirmed the presence of uronic acidsembedded in the chiral polysaccharide moiety. Tem-perature did not affect the CD spectra obtained: noconformational variation in the polysaccharide back-bones was observed in any sample. This suggestedthat the overall polymeric conformation observed wasa ‘random coil’, agreeing with previous studies ofexopolysaccharides produced by some cyanobacteriaand by the green macroalga Ulva sp.. Furthermore,this also highlighted a disordered sequence of saccha-ride units in the polymer chain (Cesàro et al. 1990,Paradossi et al. 1999, 2002, Bellezza et al. 2003). Incontrast to temperature, pH did have an effect on thespectral qualities of biofilm exopolysaccharides. Thismay have resulted from the effect of the uronic moietyon the protonation equilibria in aqueous solutions dri-ven by pH changes. As far as uronic moiety determina-tion was concerned, CD data were qualitatively inagreement with the results from HPLC.
Cytochemical analysis distinguished among acidic,carboxylated and sulphated polysaccharides in thecapsules, sheaths and mucilage of around 30 biofilmspecies. No changes in this pattern were observedamong seasons. About 50% of cyanobacteria and 70%of diatoms reacted to staining for carboxylated poly-saccharide, whereas green algae did not. Accordingly,it can be hypothesized that the carboxyl moiety re-vealed by CD mainly resulted from the presence of
cyanobacteria and diatoms. Sulphated residues werepresent in all diatoms and only in Lyngbya marten-siana and Phormidium insigne of the cyanobacteria.The use of RR and AB dyes revealed the presence ofacidic groups in 6 epilithic cyanobacteria from Romanhypogea (Albertano & Bellezza 2001, Bellezza et al.2003) and in the sheath of Nostoc commune (Abdela-had & Bazzichelli 1989). In addition, carboxylated andsulphated sugars were also evidenced in the EPS pro-duced by Cylindrotheca closterium (de Brouwer et al.2002) and in the exudates excreted by a Pleurosigmasp. strain in culture (Sdrigotti & Talarico 1994).
Although investigation of the substratum coloniza-tion, biomass succession, and exopolysaccharide char-acteristics of phototrophic biofilms in WWTP watershas been seldom conducted to date, such studies arerelevant to the exploitation of attached communities inbioremoval applications and ultimately contribute torealistic assessments of biofilm interactions with theenvironment (Barranguet et al. 2004).
Our data revealed that biofilms growing in the Fiu-micino WWTP are rather common freshwater consor-tia, with widespread specific distribution and low bio-diversity. A high productivity potential and ability togrow on a variety of artificial substrata facilitates easyhandling of such communities in wastewater treatmentsystems, which can be based on cultures of selectedorganisms (Hoffman 1998). The negatively chargedgroups and conformational behavior of the exopolysac-charides of the biofilm matrix are important for theremoval of residual nutrients and noxious cations inthe treatment of wastewaters, and hence for the pro-tection of receiving water bodies and the developmentof sustainable waste treatment alternatives to conven-tional physico-chemical methods.
Acknowledgements. We thank Dr. N. T. W. Ellwood for con-structive criticism of the manuscript and the Agency for Envi-ronmental Protection of Aeroporti di Roma, Fiumicino (Italy)for fruitful collaboration and access to the plant. This researchwas partly conducted within the framework of EU projectPHOBIA (QLK3-CT2002-01938).
LITERATURE CITED
Abdelahad N, Bazzichelli G (1989) A cytochemical investiga-tion on the extracellular investment of Nostoc communeVauch. (Cyanophyta). Arch Hydrobiol Suppl Algol Stud57:437–447
Albertano P, Bellezza S (2001) Cytochemistry of cyanobacter-ial exopolymers in biofilms from Roman hypogea. NovaHedwigia 123:501–518
Albertano P, Congestri R, Shubert LE (1999) Cyanobacterialbiofilms in sewage treatment plants along the Tyrrheniancoast (Mediterranenan Sea), Italy. Arch Hydrobiol SupplAlgol Stud 94:13–24
Barranguet C, van Beusekom SAM, Veuger B, Neu TR, Man-
310

Congestri et al.: Phototrophic biofilms in a wastewater treatment plant
ders EMM, Sinke JJ, Admiraal W (2004) Studying un-disturbed autotrophic biofilms: still a technical challenge.Aquat Microb Ecol 34:1–9
Barranguet C, Veuger B, Van Beusekom SAM, Marian P,Sinke JJ, Admiral W (2005) Divergent composition ofalgal-bacterial biofilms developing under various externalfactors. Eur J Phycol 40:1–8
Battin TJ, Kaplan LA, Newbold JD, Hansen CME (2003)Contribution of microbial biofilms to ecosystem processesin stream mesocosm. Nature 426:439–441
Bellezza S, Paradossi G, De Philippis R, Albertano P (2003)Leptolyngbya strains from Roman hypogea: cytochemicaland physico-chemical characterisation of exopolysaccha-rides. J Appl Phycol 15:193–200
Bellinger BJ, Abdullahi AS, Gretz MR, Underwood GJC(2005) Biofilm polymers: relationship between carbo-hydrate biopolymers from estuarine mudflats and unialgalcultures of benthic diatoms. Aquat Microb Ecol 38:169–180
Cesàro A, Liut G, Bertocchi C, Navarini L, Urbani R (1990)Physico-chemical properties of the exocellular polysac-charide from Cyanospira capsulata. Int J Biol Macromol12:79–84
Congestri R, Sangiorgi VC, Albertano P (2003) Cytomorphol-ogy and distribution of periphytic cyanobacteria in oneItalian WWTP. Arch Hydrobiol Suppl Algol Stud 109:185–195
Congestri R, Cox EJ, Cavacini P, Albertano P (2005) Diatoms(Bacillariophyta) in phototrophic biofilms colonising anItalian wastewater treatment plant. Diatom Res 20:241–255
Craggs RJ, Adey WH, Jenson KR, St. John MS, Green FB, Os-wald WJ (1996) Phosphorus removal from wastewater us-ing an algal turf scrubber. Water Sci Technol 33:191–198
Davis LS, Hoffmann JP, Cook PW (1990a) Seasonal successionof algal periphyton from a wastewater treatment facility.J Phycol 26:611–617
Davis LS, Hoffmann JP, Cook PW (1990b) Production andnutrient accumulation by periphyton in a wastewatertreatment facility. J Phycol 26:617–623
de Brouwer JFC, Stal LJ (2002) Daily fluctuations of exopoly-mers in cultures of the benthic diatoms Cylindrothecaclosterium and Nitzschia sp. (Bacillariophyceae). J Phycol38:464–472
de Brouwer JFC, Wolfstein K, Stal LJ (2002) Physical charac-terization and diel dynamics of different fractions of extra-cellular polysaccharides in an axenic culture of a benthicdiatom. Eur J Phycol 37:37–44
Decho AW (2000) Microbial biofilms in intertidal systems: anoverview. Cont Shelf Res 20:1257–1273
De Philippis R, Vincenzini M (1998) Exocellular polysaccha-rides from cyanobacteria and their possible applications.FEMS Microbiol Rev 22:151–175
De Philippis R, Sili C, Paperi R, Vincenzini M (2001)Exopolysaccharide-producing cyanobacteria and theirpossible exploitation: a review. J Appl Phycol 13:293–299
de Winder B, Staats N, Stal LJ, Paterson DM (1999) Carbo-hydrate secretion by phototrophic communities in tidalsediments. J Sea Res 42:131–146
Domozych CR, Plante K, Blais P, Paliulis L, Domozych DS(1993) Mucilage processing and secretion in the greenalga Closterium. I. Cytology and biochemistry. J Phycol29:650–659
Gloaguen V, Morvan H, Hoffmann L (1995) Released andcapsular polysaccharides of Oscillatoriaceae (Cyano-phyceae, Cyanobacteria). Arch Hydrobiol Suppl AlgolStud 78:53–69
Gloaguen V, Vebret L, Morvan H (1996) Chemical composi-tion of the capsular polymers of the blue-green algal matsof the thermal spring of Neris-les-Bain (France). ArchHydrobiol Suppl Algol Stud 83:245–255
Hillebrand H, Dürselen CD, Kirschtel D, Pollingher U, ZoharyT (1999) Biovolume calculation for pelagic and benthicmicroalgae. J Phycol 35:403–424
Hoagland KD, Rosowsky JR, Gretz MR, Roemer SC (1993)Diatom extracellular polymeric substances: functions, finestructure, chemistry, and physiology. J Phycol 29:537–566
Hoffmann JP (1998) Wastewater treatment with suspendedand nonsuspended algae. J Phycol 34:757–763
Kaplan D, Christiaen D, Arad Malis S (1987) Chelating pro-perties of extracellular polysaccharides from Chlorellaspp. Appl Environ Microbiol 53:2953–2956
Lam PK, Lei A (1999) Colonization of periphytic algae onartificial substrates in a tropical stream. Diatom Res 14:307–322
Larson C, Passy SI (2005) Spectral fingerprinting of algalcommunities: a novel approach to biofilm analysis andbiomonitoring. J Phycol 42:439–446
Lombardi A, Vieira AAH (1999) Lead- and copper-complex-ing extracellular ligands released by Kirchneriella aperta(Chlorococcales, Chlorophyta). Phycologia 38:283–288
Lombardi AT, Hidalgo TMR, Vieira AAH (2005) Copper com-plexing properties of dissolved organic materials exudedby the freshwater microalgae Scenedesmus acuminatus(Chlorophyceae). Chemosphere 60:453–459
Neu TR (1994) Biofilms and microbial mats. In: Krumbein WE,Paterson DM, Stal LJ (eds) Biostabilization of sediments.BIS Verlag, Oldenburg, p 9–15
Nicolaus B, Panico A, Lama L, Romano I, Manca MC, DeGiulio A, Gambacorta A (1999) Chemical composition andproduction of exopolysaccharides from representativemembers of heterocystous and non-heterocystous cyano-bacteria. Phytochemistry 52:639–647
Nultsch W, Hader DP (1988) Photomovement in motilemicroorganisms II. Photochem Photobiol 47:837–869
Otero A, Vincenzini M (2004) Nostoc (Cyanophyceae) goesnude: extracellular polysaccharides serve as a sink forreducing power under unbalanced C/N metabolism.J Phycol 40:74–81
Paradossi G, Cavalieri F, Pizzoferrato L, Liquori AM (1999) Aphysico-chemical study on the polysaccharide ulvan fromhot extraction of the macroalga Ulva. Int J Biol Macromol25:309–315
Paradossi G, Cavalieri F, Chiessi E (2002) A conformationalstudy on the algal polysaccharide ulvan. Macromolecules35:6404–6411
Paulsen BS, Vieira AAH (1994) Structure of the capsular andextracellular polysaccharides producted by the desmidSpondylosium panduriforme (Chlorophyta). J Phycol 30:638–641
Sdrigotti E, Talarico L (1994) Excretion of exudated by adiatom under culture. In: Marino D, Montresor M (eds)Proc 13th Int Diatom Symp. Biopress, Bristol, p 47–61
Shubert LE (2003) Nonmotile coccoid and colonial greenalgae. In: Wehr JD, Sheath RS (eds) Freshwater algae ofNorth America: ecology and classification. Elsevier Sci-ence Publishers, New York, p 253–309
Sláde<ková A (1994) The role of periphyton in waste treat-ment technology. Verh Int Verein Theor Angew Limnol25:1929–1932
Sláde<ková A, Matulová D (1998) Periphyton as bioelimina-tor. Verh Int Verein Limnol 26:1777–1780
Smith DJ, Underwood GJC (2000) The production of extra-cellular carbohydrates by estuarine benthic diatoms, the
311

Aquat Microb Ecol 45: 301–312, 2006
effects of growth phase and light and dark treatment.J Phycol 36:321–333
Staats N, de Winder B, Stal LJ, Mur LR (1999) Isolation andcharacterization of extracellular polysaccharides from theepipelic diatoms Cylindrotheca closterium and Naviculasalinarum. Eur J Phycol 34:161–169
Stal LJ (2000) Cyanobacterial mats and stromatolites. In:Whitton BA, Potts M (eds) The ecology of cyanobacteria.Kluwer Academic, Dordrecht, p 61–120
Stal LJ, Defarge C (2005) Structure and dynamics of exopoly-mers in an intertidal diatom biofilm. Geomicrobiol J 22:341–352
Stevenson RJ (1996) An introduction to algal ecology in fresh-water benthic habitats. In: Stevenson RJ, Bothwell ML,Lowe RL (eds) Algal ecology. Freshwater benthic eco-systems. Academic Press, San Diego, CA, p 3–30
Sutherland IW (2001) Biofilm exopolysaccharides: a strongand sticky framework. Microbiology 147:3–9
Thornton DCO (2002) Diatom aggregation in the sea: mecha-nisms and ecological implications. Eur J Phycol 37:149–161
Tuji A (2000a) Observation of development processes inloosely attached diatom (Bacillariophyceae) communities.Phycol Res 48:75–84
Tuji A (2000b) The effect of irradiance on growth of differentforms of freshwater diatoms: implications for succession in
attached diatom communities. J Phycol 36:659–661Underwood GJC, Boulcott M, Raines CA, Waldron K (2004)
Enviromental effects on exopolymer production by marinebenthic diatoms: dynamics, changes in composition, andpathways of production. J Phycol 40:293–304
van den Hoek C, Mann DG, Jahns HM (1995) Algae: anintroduction to phycology. Cambridge University Press,Cambridge
Vincenzini M, De Philippis R, Sili C, Materassi R (1990a) Stud-ies on exopolysaccharide release by diazotrophic batchcultures of Cyanospira capsulata. Appl Microbiol Biotech-nol 34:392–396
Vincenzini M, De Philippis R, Sili C, Materassi R (1990b) Anovel exopolysaccharide from a filamentous cyanobac-terium: production, chemical characterisation and rheo-logical properties. In: Dawes EA (ed) Novel biodegrad-abile microbial polymers. Kluwer Academic, Dordrecht,p 295–310
Wiggli M, Smallcombe A, Bachofen R (1999) Reflectancespectroscopy and laser confocal microscopy as tools in anecophysiological study of microbial mats in an alpine bogpond. J Microbiol Methods 34:173–182
Wingender J, Neu TR, Flemming HC (1999) What are bacte-rial extracellular polymeric substances? In: Wingender J,Neu TR, Flemming HC (eds) Microbial extracellular poly-meric substances, Springer Verlag, Berlin, p 1–19
312
Editorial responsibility: Staffan Kjelleberg,Sydney, New South Wales, Australia
Submitted: May 26, 2006; Accepted: September 4, 2006Proofs received from author(s): December 11, 2006