structuralbasisfortheoxidationofprotein-boundsulfur ... ·...
TRANSCRIPT

Structural Basis for the Oxidation of Protein-bound Sulfurby the Sulfur Cycle Molybdohemo-Enzyme SulfaneDehydrogenase SoxCD*□S
Received for publication, October 12, 2010, and in revised form, November 26, 2010 Published, JBC Papers in Press, December 8, 2010, DOI 10.1074/jbc.M110.193631
Ulrich Zander‡§, Annette Faust‡, Bjorn U. Klink§, Daniele de Sanctis¶, Santosh Panjikar�, Armin Quentmeier**,Frank Bardischewsky**, Cornelius G. Friedrich**1, and Axel J. Scheidig‡§2
From the ‡Department of Structural Biology, Zoological Institute, Christian-Albrechts-University Kiel, 24118 Kiel, Germany, the**Fakultat Bio- und Chemieingenieurwesen, Technische Universitat Dortmund, Emil-Figge-Strasse 66, 44221 Dortmund, Germany,the §Department of Biophysics-Structural Biology, Saarland University, 66421 Homburg, Germany, the ¶Structural Biology Group,European Synchrotron Radiation Facility Grenoble, 6 Rue Jules Horowitz, B.P. 220, 38043 Grenoble Cedex 9, France, and the �EMBLHamburg Outstation, c/o DESY, Notkestrasse 85, D-22603 Hamburg, Germany
The sulfur cycle enzyme sulfane dehydrogenase SoxCD is anessential component of the sulfur oxidation (Sox) enzyme sys-tem of Paracoccus pantotrophus. SoxCD catalyzes a six-elec-tron oxidation reaction within the Sox cycle. SoxCD is an �2�2heterotetrameric complex of the molybdenum cofactor-con-taining SoxC protein and the diheme c-type cytochrome SoxDwith the heme domains D1 and D2. SoxCD1 misses the heme-2domain D2 and is catalytically as active as SoxCD. The crystalstructure of SoxCD1 was solved at 1.33 A. The substrate ofSoxCD is the outer (sulfane) sulfur of Cys-110-persulfide locatedat the C-terminal peptide swinging arm of SoxY of the SoxYZcarrier complex. The SoxCD1 substrate funnel toward themolyb-dopterin is narrow and partially shielded by side-chain residues ofSoxD1. For access of the sulfane-sulfur of SoxY-Cys-110 persul-fide we propose that (i) the blockage by SoxD-Arg-98 is openedvia interaction with the C terminus of SoxY and (ii) the C-termi-nal peptide VTIGGCGGof SoxY provides interactions with theentrance path such that the cysteine-bound persulfide is opti-mally positioned near themolybdenum atom. The subsequentoxidation reactions of the sulfane-sulfur are initiated by the nu-cleophilic attack of the persulfide anion on themolybdenumatom that is, in turn, reduced. The close proximity of heme-1 tothemolybdopterin allows easy acceptance of the electrons. Be-cause SoxYZ, SoxXA, and SoxB are already structurally charac-terized, with SoxCD1 the structures of all key enzymes of the Soxcycle are knownwith atomic resolution.
Reduced inorganic sulfur compounds like hydrogen sulfide,sulfur, or thiosulfate are attractive prokaryotic energy sources,
and their oxidation to sulfuric acid is one of the major reac-tions of the global sulfur cycle as shown for thiosulfate (Equa-tion 1).
S-SO32� � 5 H2O 3 2 SO4
2� � 8 e� � 10 H� (Eq. 1)
Oxidation of inorganic sulfur compounds to sulfate ismainly mediated by various specialized aerobic chemotrophicand anaerobic phototrophic prokaryotes, bacteria and archaea(1–3). Two different modes for bacteria have been proposedrecently; one as present in e.g. the anaerobic phototrophicsulfur oxidizing bacterium Allochromatium vinosum involvesthe reverse acting dissimilatory sulfite dehydrogenase(DsrAB) (Equation 2) which is with 13 other proteins encodedby the dsr operon (4). The product sulfite is subsequently oxi-dized to sulfate by adenosine 5�-phosphosulfate reductase orsulfite:acceptor oxidoreductase (5).
HS� � 3 H2O3 HSO3� � 6 e� � 6 H� (Eq. 2)
The other mode as present in e.g. the aerobic facultativechemotrophic bacterium Paracoccus pantotrophus (6) in-volves sulfane dehydrogenase SoxCD, which is together with14 other proteins encoded by the sox operon in this strain.SoxCD is an �2�2 heterotetrameric complex of the molyb-doprotein SoxC and the hybrid di-heme cytochrome c likeprotein SoxD. This sulfane dehydrogenase (formerly desig-nated sulfur dehydrogenase (7)) is a key enzyme of the sulfur-oxidizing (Sox)3 enzyme system and catalyzes the oxidation ofprotein-bound sulfane-sulfur (oxidation state �1) to sulfone(oxidation state �5) in a six-electron transfer reaction (8, 9)(Equation 3).
SoxZY-S� � 3 H2O � 6 Cyt cox3 SoxZY-SO3� � 6 Cyt cred
� 6 H� (Eq. 3)
The current model of the Sox reaction cycle involves se-quential activity of four different periplasmic proteins SoxXA,SoxB, SoxYZ, and SoxCD (Fig. 1) (2, 9). The four proteins oxi-
* This work was supported by Deutsche Forschungsgemeinschaft GrantsFr318/10-1 and Sche545/9-1.
□S The on-line version of this article (available at http://www.jbc.org) con-tains supplemental Table 1 and Fig. 1.
The atomic coordinates and structure factors (code 2XTS) have been depositedin the Protein Data Bank, Research Collaboratory for Structural Bioinformat-ics, Rutgers University, New Brunswick, NJ (http://www.rcsb.org/).
1 To whom correspondence may be addressed. E-mail: [email protected].
2 To whom correspondence may be addressed: Dept. of Structural Biology,Zoological Institute, Christian-Albrechts-University Kiel, Am BotanischenGarten 1-9, D-24118 Kiel, Germany. Fax: 49-431-880-4929; E-mail: [email protected].
3 The abbreviations used are: Sox, sulfur oxidation; SAD, single wavelengthanomalous dispersion; ESRF, European Synchrotron Radiation Facility;r.m.s.d., root mean square deviation.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 286, NO. 10, pp. 8349 –8360, March 11, 2011© 2011 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
MARCH 11, 2011 • VOLUME 286 • NUMBER 10 JOURNAL OF BIOLOGICAL CHEMISTRY 8349
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

dize thiosulfate in vitro to sulfate and release eight electronsto reduce the final electron acceptor horse cytochrome c. Theheme enzyme complex SoxXA covalently links thiosulfate tothe thiol of the active site SoxY-Cys-110 residue of the SoxYZcomplex to yield SoxY-Cys-110-S-thiosulfate, also designatedcysteine S-thiosulfonate (9–12). The dimanganese proteinSoxB hydrolyzes sulfate to yield SoxY-Cys-110-persulfide (9,13). The sulfane-sulfur of the Cys-110-persulfide is the sub-strate for SoxCD that transfers oxygen atoms from water mol-ecules to yield SoxY-Cys-110-S-sulfonate (Fig. 1). Finally, sul-fate is hydrolyzed again by SoxB to regenerate SoxY and to
complete the cycle. Besides thiosulfate, the Sox enzyme sys-tem also oxidizes hydrogen sulfide, sulfur, and sulfite. Withthiosulfate the core system SoxYZ, SoxXA, and SoxB, i.e. thesystem without SoxCD, yields just 2 mol of electrons/mol ofsulfur substrate (14). Also, the core system without SoxCDoxidizes thiosulfate at 25% the rate as compared with SoxCD.This material balance identified (i) the turnover number ofthe Sox system to be identical with and without SoxCD, and(ii) the molybdoprotein-cytochrome c complex SoxCD cata-lyzes a unique six-electron transfer reaction (Equation 2).Six electron-transfer reactions are rare in nature. The few
examples are the reverse-acting sulfite reductase rDsrAB con-taining various [4Fe4S] centers (15) and the pentaheme cyto-chrome c nitrite reductase (16).
The core Sox system without SoxCD oxidizes sulfite, andthis is the first example for a molybdoprotein-free sulfite oxi-dation (8). Thus, SoxCD is catalytically distinct from sulfitedehydrogenases. However, the 26.5% identity with respect tothe primary sequence of SoxC to the molybdoprotein SorA ofthe sulfite dehydrogenase SorAB of Starkeya novella (9) sug-gested a relation to sulfite dehydrogenase. Although theheme-1 domain of SoxD is conserved, the heme-2 domain ismissing in some chemo- and phototrophic bacteria. Deletionof the heme-2 domain leads to a SoxCD1, which is identicallyactive in the in vitro assay as SoxCD (7).
The proteins of the core Sox system from various sourceshave been structurally characterized earlier; SoxAX of thephototroph Rhodovulum sulfidophilum (10), SoxXA andSoxYZ of P. pantotrophus (11, 12), and a SoxB homologuefrom Thermus aquaticus (13). The structure of SoxCD pre-sented here delivers the missing link for a structural interpre-tation of the complete enzymatic reaction cycle. The structureof the sulfite dehydrogenase SorAB from S. novella (17) andassociated kinetic analysis (18–21) provided the first insightinto the molecular mechanism of the intramolecular electrontransfer between the molybdenum and the heme prostheticgroup of the bound c-type cytochrome subunit upon oxida-tion of a sulfur compound. Comparison of the SoxCD1 struc-ture with SorAB and further related enzymes like chickensulfite oxidase CSO (22) highlights structural similarities asthese molybdoenzymes have in common their catalysis of hy-droxylation reactions with a sulfur atom as substrate coupledwith dehydrogenation. SoxCD, however, differs significantlyfrom these enzymes as (i) SoxCD performs a six-electrontransfer reaction instead of a two-electron transfer reaction,and (ii) its sulfur substrate is covalently bound to a sulfur car-rier protein (SoxYZ).The structure enables further insight into the common fea-
tures and determinants for the electron transfer between themolybdenum and heme prosthetic group. Because a six-elec-tron transfer of a molybdoenzyme within one enzymatic con-version is so far unique, SoxCD will be of broader interest forenzymology and protein chemistry.
EXPERIMENTAL PROCEDURES
Production and Purification of SoxCD1—SoxCD fromP. pantotrophus is a complex of the molybdenum containingprotein SoxC, and the di-heme c-type cytochrome protein
FIGURE 1. Proposed steps for the Sox reaction cycle. A, the heterodimericSoxYZ protein complex does not contain any cofactor. The SoxYZ proteinbinds the sulfur substrate via a covalent link to the sulfhydryl moiety of thehighly conserved cysteine residue at the C-terminal end of SoxY (9, 12). Thislinkage reaction is catalyzed by SoxXA, a heterodimeric c-type cytochrome,with SoxX as a monoheme subunit and SoxA as a diheme subunit (10, 11).The dimanganese SoxB protein is proposed to hydrolyze off sulfate fromSoxY-cysteine-S-thiosulfate (9, 13). The SoxCD functions as sulfane dehydro-genase such that it catalyzes the oxidation of the SoxY-bound sulfane(SoxY-Cys-110-persulfide) to SoxY-bound sulfonate (SoxY-Cys-110-S-sulfon-ate), which is released as sulfate from SoxY by a SoxB-catalyzed reaction.B, shown are structure and name convention for sulfur compounds usedwithin the text. For some reaction intermediates, different names are usedwithin the literature; those used within the text are in bold within the figure.
Crystal Structure of Sulfane Dehydrogenase SoxCD
8350 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 10 • MARCH 11, 2011
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

SoxD. SoxCD1 is the construct in which the second heme-2domain of SoxD is deleted (7). The SoxCD1 complex was pu-rified from P. pantotrophus strain GB17 as described previ-ously (2, 23). SoxCD1 was enriched from the cell extract bydifferential centrifugation and ammonium sulfate fraction-ation. Soluble proteins precipitated between 44 and 65% am-monium sulfate saturation were redissolved and purified tohomogeneity involving anion exchange chromatography onQ-Sepharose, gel filtration on Sephadex G-200, and anionexchange on Resource Q. The activity of the SoxCD1 complexwas examined from the reconstituted thiosulfate-oxidizingenzyme system using horse heart cytochrome c as final elec-tron acceptor as described before (23).Crystallization—Crystallization experiments were carried
out using the hanging-drop method at room temperature.SoxCD1 crystals were prepared by mixing equal amounts ofprotein- and reservoir solution (4% (w/v) MPEG 5000, 40 mM
Tris-HCl (pH 8), 20 mM calcium acetate, and 10 mM Co(III)-hexamine chloride), and the drops were equilibrated againstthe reservoir solution. Rod-shaped crystals appeared within5–10 days and diffracted x-rays beyond 1.5 Å resolution. Thecrystals belonged to a trigonal space group with cell dimen-sions a � 123 Å and c � 76 Å. Before data collection the crys-tals were cryo-protected in a solution containing 4% (w/v)MPEG 5000, 40 mM Tris-HCl (pH 8), 20 mM calcium acetate,10 mM Co(III)-hexamine chloride, and 10% (v/v) glycerol andflash-cooled in liquid nitrogen.Data Collection and SAD Phasing—X-ray data were col-
lected on beamline ID23-1 (24) at the European SynchrotronRadiation Facility, Grenoble, France. To obtain experimentalphases, a data set was collected at the K-edge of iron. The dataset was indexed, integrated, and scaled using the XDS package(25). The structure was solved using the SAD protocol of Au-to-Rickshaw, the EMBL-Hamburg automated crystal struc-ture determination platform (26). The input diffraction datawere prepared and converted for use in Auto-Rickshaw usingprograms of the CCP4 suite (27). Structure factors were cal-culated using the program SHELXC (28). Based on an initialanalysis of the data, the maximum resolution for substructuredetermination and initial phase calculation was set to 3.2 Å.All of the heavy atoms requested were found using the pro-gram SHELXD (29). The correct enantiomorph space groupfor the substructure was determined using the programs ABS(30) and SHELXE (31). Initial phases were calculated afterdensity modification using the program SHELXE (31). The2-fold non-crystallographic symmetry operator was foundusing the program RESOLVE (32). Density modification,phase extension to 2.4 Å, and non-crystallographic symmetryaveraging was performed using the program DM (33). At thisstage, around one-third of the protein complex was built us-ing the program ARP/wARP (34, 35). This model togetherwith sequence information was used as a starting model forthe subsequent MRSAD (SAD combined with molecular re-placement) protocol of Auto-Rickshaw (36). The resultingelectron density map was improved further by density modifi-cation using DM (33). The resulting phases together with theSoxCD sequence were used as input for model building withPHENIX autobuild (37). Before further refinement with
REFMAC5 (38), the water molecules were manually removedfrom the Phenix-generated autobuild model. This resulted inthe first SoxCD1 model at 2.4 Å. A high resolution data setwas collected from a second crystal up to a resolution of 1.33Å. After integration using XDS, the data were scaled byXSCALE and converted into CCP4 format using XDSCONVkeeping the same reflections marked for Rfree assignment as inthe SAD dataset. The amplitudes from the high resolutiondata set and phases calculated from the 2.4 Å model wereused as input for ARP/wARP (35), also providing the SoxCDsequence. The resulting model was examined, cofactors werebuilt, and water molecules were added using Coot (39) fol-lowed by refinement using REFMAC5 until no further inter-pretable features in the electron density map were visible. Therefinement statistics are summarized in Table 1. Final modelanalysis, imaging, and ray tracing were performed using theprogram PyMOL (40). The search for structurally related pro-teins in the protein data bank (PDB (41)) was performed usingthe DALI server (42). The peptide model was manually builtusing PyMOL with structure idealization performed withREFMAC5.
TABLE 1Statistics for data collection, processing, and structure refinement
Data collection and processingData set SAD-peak High resolutionX-ray source ID23-1 (ESRF, Grenoble, France)Detector ADSCg quantum Q315rWavelength (Å) 1.74 0.98Temperature (K) 100 K 100 KCrystal-to-detector distance (mm) 180 180Oscillation range (degree) 1.0 0.5Total oscillation range (degree) 360 60Space group P3(1) P3(1)Cell dimensions (Å) a � 123.19, c � 76.42 a � 122.97, c � 76.39Resolution limit (Å)a 50-2.37 (3.0-2.37) 50-1.33 (1.4-1.33)Completeness (%)a 92.4 (88.2) 92 (72.9)No. observations (overall / unique) 550,606/48,615 507,433/273,185Average redundancy 11.3 (11.2) 1.8 (1.5)�I/�(I)�a 31.4 (19.5) 11.8 (2.6)Rp.i.m.
b 2.0 (5.9) 4.1 (25.2)B-factor fromWilson plot (Å2) 27.0 9.7
Refinement statisticsResolution limit (Å)a 50-1.33Number of unique reflectionsa 273,185Completeness of data (%)a 96.8Rcryst (%)
a,c 9.8Rfree (%)
a,d 11.2No. of non-H atoms 10,432Protein 9,473Solvent 959
Ramachandran plot (%)e 97.1/2.8/0.1Coordinate errorf 0.169r.m.s.d. from ideal valuesBond lengths (Å) 0.028Bond angles (degree) 2.368
Mean B-factor (Å2) per proteinchain A/B/C/D
10.7/13.2/10.9/12.8
Backbone 9.5/12.0/9.4/11.8Side chain 11.8/14.5/11.6/13.9Solvent 19.6
PDB entry code 2XTSa Values in parentheses are for the high resolution bin.b Where Ii(hkl) is the intensity of the ith individual measurement of the reflectionwith Miller indices hkl, and �Ii(hkl)� is the mean intensity of all measurements ofI(hkl), calculated for I � 3�(I). N is the redundancy or multiplicity of the ob-served reflection (67, 68).
c Rcryst � 100 � �(�F—-obs� � k�F—-calc�)/��F—-obs�, where Fobs and Fcalc are theobserved and calculated structure-factor amplitudes, respectively.
dRfree is equivalent to Rcryst but calculated from reflections (5%) omitted in therefinement process (69, 70).
e Calculated using the program PROCHECK (71); favored/allowed/generous.f Calculated based on a Luzzati plot using the program SFCHECK (56).g Area Detector Systems Corporation.
Crystal Structure of Sulfane Dehydrogenase SoxCD
MARCH 11, 2011 • VOLUME 286 • NUMBER 10 JOURNAL OF BIOLOGICAL CHEMISTRY 8351
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from
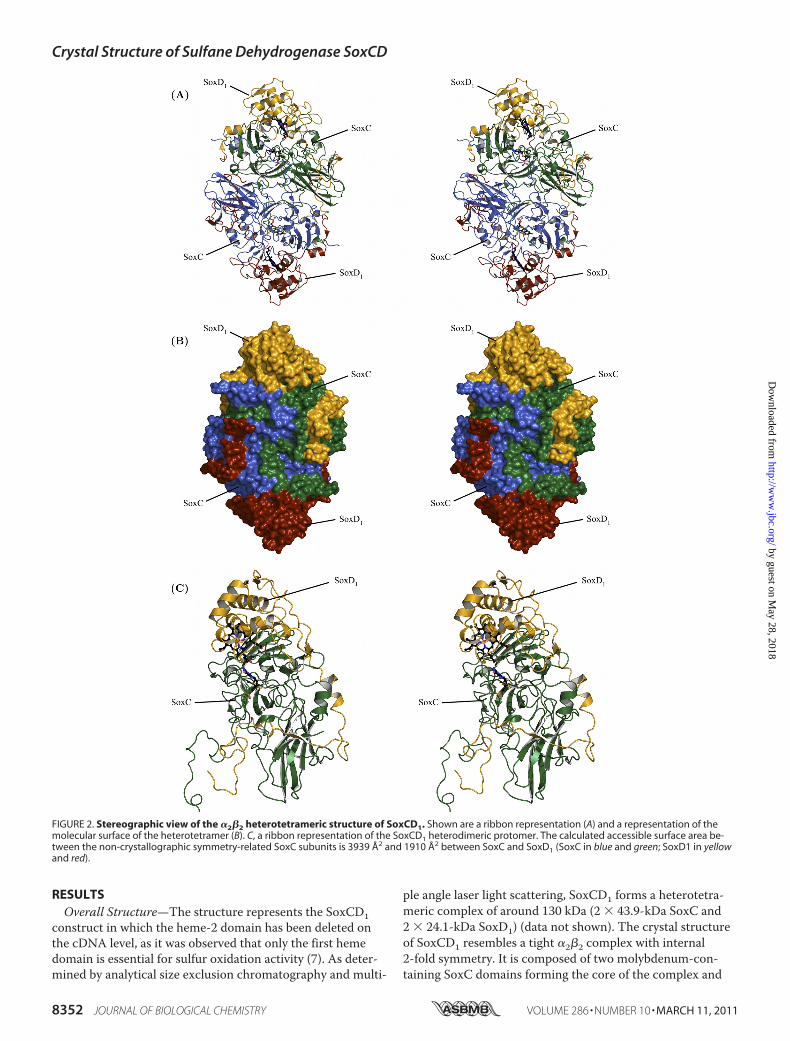
RESULTSOverall Structure—The structure represents the SoxCD1
construct in which the heme-2 domain has been deleted onthe cDNA level, as it was observed that only the first hemedomain is essential for sulfur oxidation activity (7). As deter-mined by analytical size exclusion chromatography and multi-
ple angle laser light scattering, SoxCD1 forms a heterotetra-meric complex of around 130 kDa (2 � 43.9-kDa SoxC and2 � 24.1-kDa SoxD1) (data not shown). The crystal structureof SoxCD1 resembles a tight �2�2 complex with internal2-fold symmetry. It is composed of two molybdenum-con-taining SoxC domains forming the core of the complex and
FIGURE 2. Stereographic view of the �2�2 heterotetrameric structure of SoxCD1. Shown are a ribbon representation (A) and a representation of themolecular surface of the heterotetramer (B). C, a ribbon representation of the SoxCD1 heterodimeric protomer. The calculated accessible surface area be-tween the non-crystallographic symmetry-related SoxC subunits is 3939 Å2 and 1910 Å2 between SoxC and SoxD1 (SoxC in blue and green; SoxD1 in yellowand red).
Crystal Structure of Sulfane Dehydrogenase SoxCD
8352 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 10 • MARCH 11, 2011
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

two heme-containing SoxD1 domains sitting at the exterior ofthe complex (Fig. 2). The two SoxD1 subunits are positionedat the edge of the SoxC dimer and do not contact each other.The C-terminal peptide of the SoxD1 subunit displays an ex-tended loop with two short �-helical and one short�-stranded conformation that embraces its closest SoxC sub-unit. SoxD1 represents a cytochrome-c-like-fold with 8 helicesand 5 strands. SoxC contains 6 �-helices and 23 �-strandsorganized in five �-sheets with the molybdenum binding sitein the center and a C-terminal immunoglobulin-like fold. Thetwo SoxC subunits of the tetramer form a large interior inter-face (3939.3 Å2 interface area, determined using SURFNET(43)) in a head-to-tail arrangement. The N-terminal part ofSoxC (residues 42 to 75) builds additional contacts with itsnon-crystallographic symmetry mate (Fig. 2).Molybdenum Binding Site—Due to the high resolution of
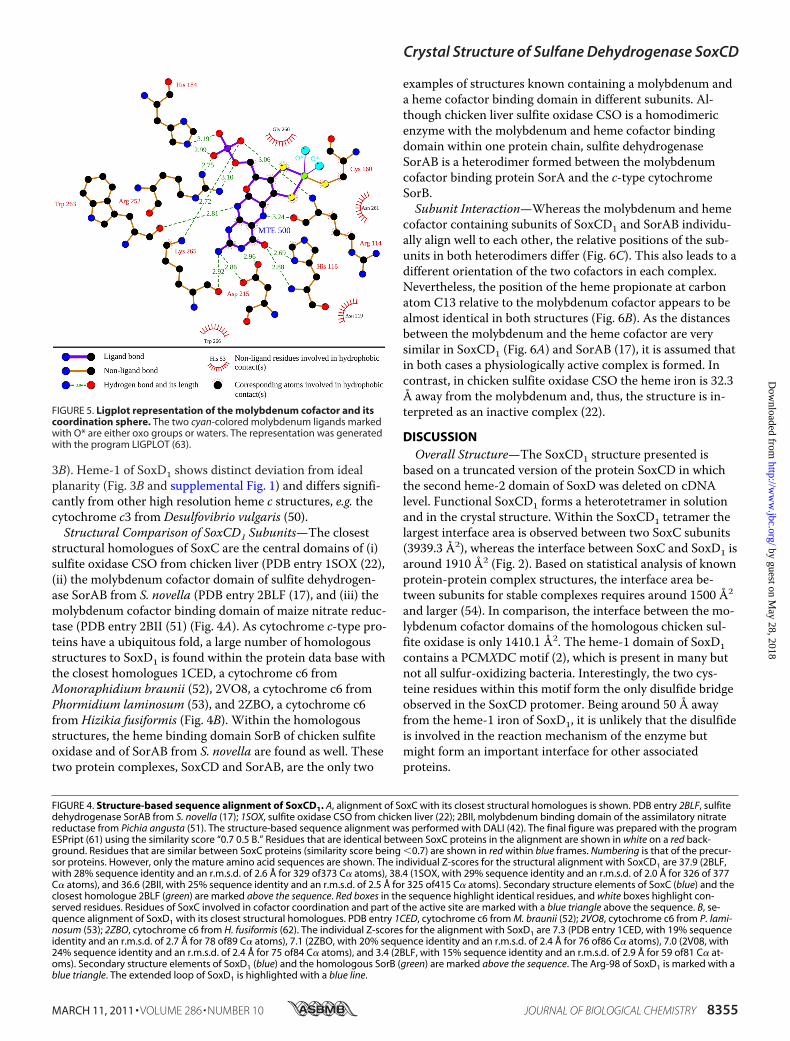
the final electron density map, the type of the molybdenum-containing prosthetic group (Fig. 3A) could be unambiguouslyidentified as the sulfite oxidase-type molybdenum cofactor(44, 45) as also reported for chicken sulfite oxidase CSO (22),the sulfur dehydrogenase SorAB from S. novella (17), the sul-fite oxidase from Arabidopsis thaliana (46), and the sulfiteoxidase from Escherichia coli (47) (Fig. 4A). As proposed in
the literature (45), the molybdenum-containing prostheticgroup will be subsequently referred to as molybdenum cofac-tor. Molybdenum cofactor binding domains are highly similarin their central architecture and form a distinct family (48).Eight residues are highly conserved and function as a coordi-nation sphere (Fig. 5 and supplemental Table 1). Among theseeight residues, one cysteine residue (Cys-160 in SoxC) isfound in all molybdenum cofactor-containing structures asone ligand for the molybdenum ion (Figs. 4A and 5). The mo-lybdenum ion in SoxC displays a distorted square pyramidalcoordination geometry where two thiolates from the molyb-dopterin heterocycle, the thiol group from cysteine residueCys-160, and one water/hydroxyl oxygen are the equatorialligands, and the terminal oxo group occupies the axial posi-tion (Fig. 3A and 5). The residue Arg-114 of SoxC is a highlyconserved residue in the vicinity of the molybdenum cofactor(Fig. 4A). Arg-114 is proposed to coordinate and thereby topolarize a water molecule before the concerted transfer of anoxygen atom onto the sulfur atom (presented to the active siteby the SoxYZ complex) and transfer of two electrons from thesulfur onto the molybdenum cofactor (20). This first oxida-tion step is completed by a stepwise transfer of the two elec-trons from the molybdenum cofactor to the heme group of
FIGURE 3. Stereographic view of the cofactors of SoxCD1 superimposed with the surrounding electron density distribution. The contour level of the2Fobs � Fcalc electron density map is 3.0�. Shown are molybdenum (A) and heme (B) cofactors with coordinating residues. The protein environment in pan-els A and B is displayed as tube representation in light gray.
Crystal Structure of Sulfane Dehydrogenase SoxCD
MARCH 11, 2011 • VOLUME 286 • NUMBER 10 JOURNAL OF BIOLOGICAL CHEMISTRY 8353
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

SoxD1. Subsequently these electrons are transferred to theiron of an interacting cytochrome subunit. His-116 of SoxC islocated between the molybdenum cofactor and heme-1 ofSoxD1 and might be involved in the transfer of the electronsfrom the molybdenum to the heme iron (Fig. 6A).
Heme-1 Site—Heme-1 displays typical properties of c-typecytochromes (49); the porphyrin ring system is covalently at-tached to two cysteine residues (Cys-70 and Cys-73). Its cen-tral iron is axially coordinated by the SD sulfur of methionine(Met-121) and the NE2 nitrogen of histidine (His-74) (Fig.
Crystal Structure of Sulfane Dehydrogenase SoxCD
8354 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 10 • MARCH 11, 2011
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from
3B). Heme-1 of SoxD1 shows distinct deviation from idealplanarity (Fig. 3B and supplemental Fig. 1) and differs signifi-cantly from other high resolution heme c structures, e.g. thecytochrome c3 from Desulfovibrio vulgaris (50).Structural Comparison of SoxCD1 Subunits—The closest
structural homologues of SoxC are the central domains of (i)sulfite oxidase CSO from chicken liver (PDB entry 1SOX (22),(ii) the molybdenum cofactor domain of sulfite dehydrogen-ase SorAB from S. novella (PDB entry 2BLF (17), and (iii) themolybdenum cofactor binding domain of maize nitrate reduc-tase (PDB entry 2BII (51) (Fig. 4A). As cytochrome c-type pro-teins have a ubiquitous fold, a large number of homologousstructures to SoxD1 is found within the protein data base withthe closest homologues 1CED, a cytochrome c6 fromMonoraphidium braunii (52), 2VO8, a cytochrome c6 fromPhormidium laminosum (53), and 2ZBO, a cytochrome c6from Hizikia fusiformis (Fig. 4B). Within the homologousstructures, the heme binding domain SorB of chicken sulfiteoxidase and of SorAB from S. novella are found as well. Thesetwo protein complexes, SoxCD and SorAB, are the only two
examples of structures known containing a molybdenum anda heme cofactor binding domain in different subunits. Al-though chicken liver sulfite oxidase CSO is a homodimericenzyme with the molybdenum and heme cofactor bindingdomain within one protein chain, sulfite dehydrogenaseSorAB is a heterodimer formed between the molybdenumcofactor binding protein SorA and the c-type cytochromeSorB.Subunit Interaction—Whereas the molybdenum and heme
cofactor containing subunits of SoxCD1 and SorAB individu-ally align well to each other, the relative positions of the sub-units in both heterodimers differ (Fig. 6C). This also leads to adifferent orientation of the two cofactors in each complex.Nevertheless, the position of the heme propionate at carbonatom C13 relative to the molybdenum cofactor appears to bealmost identical in both structures (Fig. 6B). As the distancesbetween the molybdenum and the heme cofactor are verysimilar in SoxCD1 (Fig. 6A) and SorAB (17), it is assumed thatin both cases a physiologically active complex is formed. Incontrast, in chicken sulfite oxidase CSO the heme iron is 32.3Å away from the molybdenum and, thus, the structure is in-terpreted as an inactive complex (22).
DISCUSSIONOverall Structure—The SoxCD1 structure presented is
based on a truncated version of the protein SoxCD in whichthe second heme-2 domain of SoxD was deleted on cDNAlevel. Functional SoxCD1 forms a heterotetramer in solutionand in the crystal structure. Within the SoxCD1 tetramer thelargest interface area is observed between two SoxC subunits(3939.3 Å2), whereas the interface between SoxC and SoxD1 isaround 1910 Å2 (Fig. 2). Based on statistical analysis of knownprotein-protein complex structures, the interface area be-tween subunits for stable complexes requires around 1500 Å2
and larger (54). In comparison, the interface between the mo-lybdenum cofactor domains of the homologous chicken sul-fite oxidase is only 1410.1 Å2. The heme-1 domain of SoxD1contains a PCMXDC motif (2), which is present in many butnot all sulfur-oxidizing bacteria. Interestingly, the two cys-teine residues within this motif form the only disulfide bridgeobserved in the SoxCD protomer. Being around 50 Å awayfrom the heme-1 iron of SoxD1, it is unlikely that the disulfideis involved in the reaction mechanism of the enzyme butmight form an important interface for other associatedproteins.
FIGURE 4. Structure-based sequence alignment of SoxCD1. A, alignment of SoxC with its closest structural homologues is shown. PDB entry 2BLF, sulfitedehydrogenase SorAB from S. novella (17); 1SOX, sulfite oxidase CSO from chicken liver (22); 2BII, molybdenum binding domain of the assimilatory nitratereductase from Pichia angusta (51). The structure-based sequence alignment was performed with DALI (42). The final figure was prepared with the programESPript (61) using the similarity score “0.7 0.5 B.” Residues that are identical between SoxC proteins in the alignment are shown in white on a red back-ground. Residues that are similar between SoxC proteins (similarity score being 0.7) are shown in red within blue frames. Numbering is that of the precur-sor proteins. However, only the mature amino acid sequences are shown. The individual Z-scores for the structural alignment with SoxCD1 are 37.9 (2BLF,with 28% sequence identity and an r.m.s.d. of 2.6 Å for 329 of373 C� atoms), 38.4 (1SOX, with 29% sequence identity and an r.m.s.d. of 2.0 Å for 326 of 377C� atoms), and 36.6 (2BII, with 25% sequence identity and an r.m.s.d. of 2.5 Å for 325 of415 C� atoms). Secondary structure elements of SoxC (blue) and theclosest homologue 2BLF (green) are marked above the sequence. Red boxes in the sequence highlight identical residues, and white boxes highlight con-served residues. Residues of SoxC involved in cofactor coordination and part of the active site are marked with a blue triangle above the sequence. B, se-quence alignment of SoxD1 with its closest structural homologues. PDB entry 1CED, cytochrome c6 from M. braunii (52); 2VO8, cytochrome c6 from P. lami-nosum (53); 2ZBO, cytochrome c6 from H. fusiformis (62). The individual Z-scores for the alignment with SoxD1 are 7.3 (PDB entry 1CED, with 19% sequenceidentity and an r.m.s.d. of 2.7 Å for 78 of89 C� atoms), 7.1 (2ZBO, with 20% sequence identity and an r.m.s.d. of 2.4 Å for 76 of86 C� atoms), 7.0 (2V08, with24% sequence identity and an r.m.s.d. of 2.4 Å for 75 of84 C� atoms), and 3.4 (2BLF, with 15% sequence identity and an r.m.s.d. of 2.9 Å for 59 of81 C� at-oms). Secondary structure elements of SoxD1 (blue) and the homologous SorB (green) are marked above the sequence. The Arg-98 of SoxD1 is marked with ablue triangle. The extended loop of SoxD1 is highlighted with a blue line.
FIGURE 5. Ligplot representation of the molybdenum cofactor and itscoordination sphere. The two cyan-colored molybdenum ligands markedwith O* are either oxo groups or waters. The representation was generatedwith the program LIGPLOT (63).
Crystal Structure of Sulfane Dehydrogenase SoxCD
MARCH 11, 2011 • VOLUME 286 • NUMBER 10 JOURNAL OF BIOLOGICAL CHEMISTRY 8355
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Cofactor Sites—The structures known so far of molybde-num-dependent sulfur-oxidizing enzymes harbor the samemolybdenum cofactor assigned as the sulfite oxidase-type (44,45). Based on the high resolution structure, the same type ofcofactor, molybdopterin, was identified for the sulfane dehy-drogenase SoxCD1. Heme-1 displays a nonplanar distortioninduced through the cysteine thioether linkages between theporphyrin pyrrole groups and the polypeptide. A similar,however weaker distortion is reported for the yeast iso-1-cy-tochrome c (55). It is proposed that the redox potential of theheme group can be modulated by such geometric variation(55).SoxYZ-bound Sulfur Compounds Are the Substrates for
SoxCD—The reaction of SoxCD differs significantly from thatof its close structural relative SorAB from S. novella. Whereas
SorAB solely oxidizes sulfite to sulfate, SoxCD cannot oxidizefree sulfite. To display catalytic activity, SoxCD requires thepresentation of the sulfur compound covalently bound toSoxYZ (7). SoxYZ carries the intermediates of thiosulfate-oxidation on the Cys-110 of the C-terminal peptide “swingingarm” 105VTIGGCGG112 of SoxY (12). Whereas the substratefor SoxCD, SoxZY-Cys-110-persulfide, and the final product,SoxZY-Cys-110-S-sulfonate, have been identified (7), the in-termediates SoxZY-Cys-110-S-sulfenate and SoxZY-Cys-110-S-sulfinate (Fig. 1B) are hypothetical. The protein-bound sub-strate for SoxCD is a fundamental difference to SorAB andcan be rationalized by analysis of the substrate binding pocketnext to the molybdenum cofactor.SoxCD Substrate Binding Pocket—The SoxCD1 structure
exhibits a positively charged entrance path leading like a fun-
FIGURE 6. Stereographic view of the relative orientation of the heme cofactor toward the molybdenum cofactor. A, shown is a stick representation of theheme and the molybdenum cofactor for one SoxCD1 protomer together with the intermediate positioned residue SoxC-His-116 and distance values. B, shown issuperposition of one SoxCD1 protomer (cofactors in black) onto SorAB (cofactors in orange). The superposition is based on the molybdenum cofactor containingsubunits (SoxC and SorA, respectively). C, a stereographic view of the structural alignment of SoxCD1 with the sulfite dehydrogenase SorAB from S. novella isshown. Molybdenum cofactor containing subunits (SoxC in blue and SorA in gray) are aligned to each other. The molybdenum cofactor-containing subunits aredisplayed as a main-chain trace; the heme cofactor-containing subunits (SoxD1 in red and SorB in gray) are displayed in ribbon representation.
Crystal Structure of Sulfane Dehydrogenase SoxCD
8356 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 10 • MARCH 11, 2011
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from
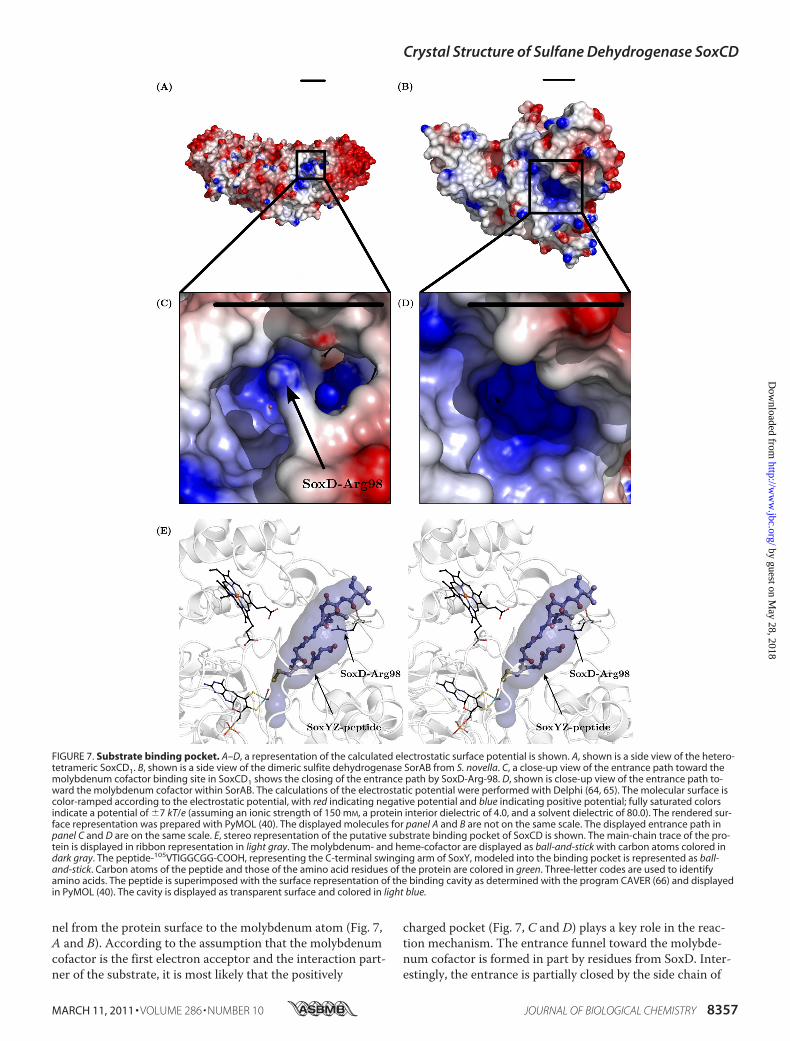
nel from the protein surface to the molybdenum atom (Fig. 7,A and B). According to the assumption that the molybdenumcofactor is the first electron acceptor and the interaction part-ner of the substrate, it is most likely that the positively
charged pocket (Fig. 7, C and D) plays a key role in the reac-tion mechanism. The entrance funnel toward the molybde-num cofactor is formed in part by residues from SoxD. Inter-estingly, the entrance is partially closed by the side chain of
FIGURE 7. Substrate binding pocket. A–D, a representation of the calculated electrostatic surface potential is shown. A, shown is a side view of the hetero-tetrameric SoxCD1. B, shown is a side view of the dimeric sulfite dehydrogenase SorAB from S. novella. C, a close-up view of the entrance path toward themolybdenum cofactor binding site in SoxCD1 shows the closing of the entrance path by SoxD-Arg-98. D, shown is close-up view of the entrance path to-ward the molybdenum cofactor within SorAB. The calculations of the electrostatic potential were performed with Delphi (64, 65). The molecular surface iscolor-ramped according to the electrostatic potential, with red indicating negative potential and blue indicating positive potential; fully saturated colorsindicate a potential of 7 kT/e (assuming an ionic strength of 150 mM, a protein interior dielectric of 4.0, and a solvent dielectric of 80.0). The rendered sur-face representation was prepared with PyMOL (40). The displayed molecules for panel A and B are not on the same scale. The displayed entrance path inpanel C and D are on the same scale. E, stereo representation of the putative substrate binding pocket of SoxCD is shown. The main-chain trace of the pro-tein is displayed in ribbon representation in light gray. The molybdenum- and heme-cofactor are displayed as ball-and-stick with carbon atoms colored indark gray. The peptide-105VTIGGCGG-COOH, representing the C-terminal swinging arm of SoxY, modeled into the binding pocket is represented as ball-and-stick. Carbon atoms of the peptide and those of the amino acid residues of the protein are colored in green. Three-letter codes are used to identifyamino acids. The peptide is superimposed with the surface representation of the binding cavity as determined with the program CAVER (66) and displayedin PyMOL (40). The cavity is displayed as transparent surface and colored in light blue.
Crystal Structure of Sulfane Dehydrogenase SoxCD
MARCH 11, 2011 • VOLUME 286 • NUMBER 10 JOURNAL OF BIOLOGICAL CHEMISTRY 8357
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from
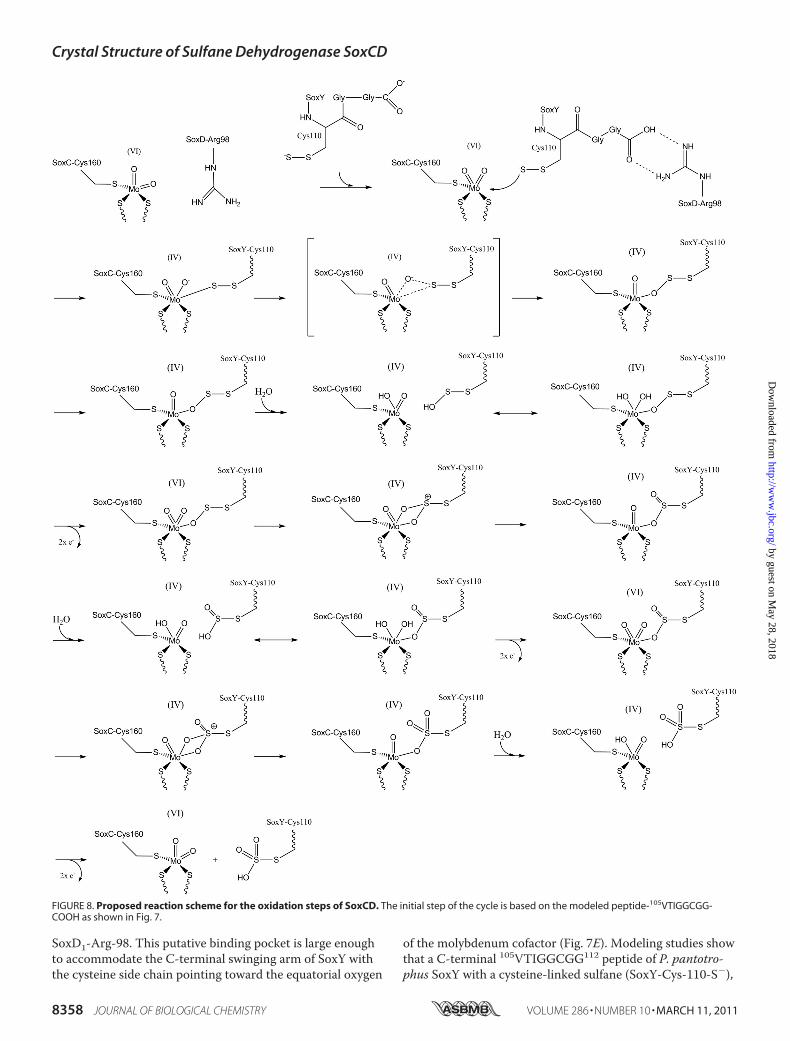
SoxD1-Arg-98. This putative binding pocket is large enoughto accommodate the C-terminal swinging arm of SoxY withthe cysteine side chain pointing toward the equatorial oxygen
of the molybdenum cofactor (Fig. 7E). Modeling studies showthat a C-terminal 105VTIGGCGG112 peptide of P. pantotro-phus SoxY with a cysteine-linked sulfane (SoxY-Cys-110-S�),
FIGURE 8. Proposed reaction scheme for the oxidation steps of SoxCD. The initial step of the cycle is based on the modeled peptide-105VTIGGCGG-COOH as shown in Fig. 7.
Crystal Structure of Sulfane Dehydrogenase SoxCD
8358 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 10 • MARCH 11, 2011
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

sulfenate (SoxY-Cys-110-SO�), sulfinate (SoxY-Cys-110-SO2
�), and sulfonate (SoxY-Cys-110-SO3�) (Fig. 1) can easily
be accommodated within the SoxC binding site next to themolybdenum cofactor (Fig. 7E). In the modeling studies theC-terminal peptide reaches into the active site such thatthe side chain of cysteine SoxY-Cys-110 is positioned closeto the axial oxo-group of the molybdenum atom. Interest-ingly, the C terminus of the C-terminal peptide is positionedsuch that it can form an ion pair with the side chain of SoxD-Arg-98. A similar stabilization of the C terminus is discussedfor the interaction of SoxB and SoxY (13). The side chain ofSoxD-Arg-98 is stretched over the entrance path such that itmight block smaller negatively charged molecules like sulfiteor thiosulfate from access to the active site. It is conceivablethat due to interaction with the substrate-loaded SoxYZ, thisblockage is released, and the sulfur-compound can be posi-tioned within the active site. This scenario might explain thefact that SoxCD can only catalyze the reaction of SoxY-Cyspersulfide but not free sulfite or thiosulfate (Fig. 8). Afterthree individual oxidation steps (transfer of each two elec-trons onto the molybdenum and one oxygen onto the sulfur),the final product SoxY-Cys-110-SO3
� is produced, and nofurther oxidation can take place. Due to its size, this finalproduct does not fit well in the active site any longer, and thetransient complex formed between SoxCD and SoxYZ is re-leased as SoxZY-Cys-110-SO3
� to enable another round ofreaction.Reaction Mechanism of SoxCD—A reaction mechanism for
bacterial sulfite oxidases has been proposed based on electronparamagnetic resonance spectroscopy (19, 57, 58). The initialstep in this mechanism is the coordination of one oxygenatom of sulfite to molybdenum followed by a nucleophilicattack of the free electron pair of the sulfur onto the axial oxo-group. For the initial step of oxidation in SoxCD, the elec-trons from the terminal sulfur atom of the SoxY-Cys-110-persulfide anion have to perform the nucleophilic attack ontothe molybdenum atom with subsequent transfer of oneMoAO oxygen onto the terminal sulfur atom. We proposethat the swinging arm sequence 105VTIGGCGG-COOHserves to localize the sulfane-sulfur of the SoxY-Cys-110-per-sulfide in close proximity to the equatorial MoAO oxygen(Fig. 8). For the second and third step of oxygen transfer, anintermediate cis-dioxomolybdenum(VI) (44) can be proposedsuch that the MoOOOS linkage is not released by the attackof hydroxide from the solvent. The MoOOOS linkage re-mains as long as further pairs of electrons can be trans-ferred from the terminal sulfur atom onto the molybdenumatom. After transfer of two electrons monooxomolybdenu-m(IV), Mo(IV)O, is subsequently reoxidized to Mo(VI) bytwo one-electron transfer steps onto heme-1 coupled withdeprotonation of a molybdenum-bound water molecule.After the addition of the third oxygen to the sulfur atomand electron transfer, the product SoxY-Cys-110-S-sulfo-nate is displaced by hydroxide, and the oxidized cofactorMo(VI)O2 is regenerated. As SoxY-Cys-110-S-thiosulfatecannot be further oxidized, it dissociates from the activesite binding pocket. Regarding individual rate constants, itcan be assumed that the transfer of electrons from the mo-
lybdenum cofactor to the heme cofactor, binding of hy-droxide ion to the molybdenum ion and subsequent attackof the molybdenum bound oxygen by the electron pairs ofthe terminal sulfur atom is much faster compared with therelease of the oxidized sulfur intermediate from the activesite binding pocket. Thus, as long as the sulfur compoundhas free electrons on the terminal sulfur atom, the oxida-tion takes place, and only after the third and last possibletransfer of an electron pair is the fully oxidized sulfur com-pound completely released.The distance of the reaction centers in SoxCD is very similar
to the distance in the active state of SorAB. Interestingly, therelative position of the molybdenum- and heme cofactor-harbor-ing subunits differs between SoxCD and SorAB (Fig. 6C). Thepropionate moiety at the carbon atomC13 of the heme groups,however, are almost identically positioned (Fig. 6B). This sug-gests that the propionate facilitates the electron transfer betweenthemolybdenum cofactor and the heme as it exhibits the closestdistance between the two (Fig. 6). It has been shown previ-ously that electrons can be tunneled in proteins betweenredox centers up to distances of 14 Å (59) at very fast rates.Between the molybdenum cofactor and the heme cofactor,the residue His-116 of SoxC is located with a distance of5.6 Å to the molybdenum, 2.7 Å to O4 of the aromatic ringsystem of the molybdenum cofactor, and 3.2 Å to the hemepropionate. These residues can serve as an alternative pathfor the electron transfer (Fig. 6).In 1982 Kelly (60) stated, “The reactions about which least
are known are those effecting thiosulfate cleavage and thoseconverting sulfur to sulfite.” The reaction for thiosulfatecleavage has been proposed earlier (9, 13). Considering theoxidation reaction of sulfur to sulfite, the structure of the mo-lybdoprotein-cytochrome c complex SoxCD1 provides de-tailed insight into the molecular basics for its unique six-elec-tron transfer that requires the substrate to be boundcovalently to its specific carrier protein. With SoxCD1, all en-zyme structures of the core reaction cycle for sulfur oxidationare determined.
Acknowledgments—We gratefully acknowledge access to the corefacilities of the Zentrum fur Biochemie und Molekularbiologie/La-bor fur Molekulare Biotechnologie der Christian-Albrechts-Univer-sitat zu Kiel. Additionally, we are grateful for access to beamlines atESRF (Grenoble, France), SLS (PSI Villigen, Switzerland), BESSY II(Berlin, Germany), and DESY (Hamburg, Germany) and thank thebeamline staff for technical support. We thank Renato Weisse forsize exclusion chromatography and multiple angle laser light scat-tering measurements.
REFERENCES1. Frigaard, N. U., and Dahl, C. (2009) Adv. Microb. Physiol. 54, 103–2002. Friedrich, C. G., Bardischewsky, F., Rother, D., Quentmeier, A., and Fis-
cher, J. (2005) Curr. Opin. Microbiol. 8, 253–2593. Kelly, D. P., Shergill, J. K., Lu, W. P., and Wood, A. P. (1997) Antonie
Van Leeuwenhoek 71, 95–1074. Dahl, C., Engels, S., Pott-Sperling, A. S., Schulte, A., Sander, J., Lubbe, Y.,
Deuster, O., and Brune, D. C. (2005) J. Bacteriol. 187, 1392–14045. Grimm, F., Dobler, N., and Dahl, C. (2010) Microbiology 156,
764–773
Crystal Structure of Sulfane Dehydrogenase SoxCD
MARCH 11, 2011 • VOLUME 286 • NUMBER 10 JOURNAL OF BIOLOGICAL CHEMISTRY 8359
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6. Rainey, F. A., Kelly, D. P., Stackebrandt, E., Burghardt, J., Hiraishi, A.,Katayama, Y., and Wood, A. P. (1999) Int. J. Syst. Bacteriol. 49, 645–651
7. Bardischewsky, F., Quentmeier, A., Rother, D., Hellwig, P., Kostka, S.,and Friedrich, C. G. (2005) Biochemistry 44, 7024–7034
8. Rother, D., Henrich, H. J., Quentmeier, A., Bardischewsky, F., andFriedrich, C. G. (2001) J. Bacteriol. 183, 4499–4508
9. Friedrich, C. G., Rother, D., Bardischewsky, F., Quentmeier, A., and Fis-cher, J. (2001) Appl. Environ. Microbiol. 67, 2873–2882
10. Bamford, V. A., Bruno, S., Rasmussen, T., Appia-Ayme, C., Cheesman,M. R., Berks, B. C., and Hemmings, A. M. (2002) EMBO J. 21,5599–5610
11. Dambe, T., Quentmeier, A., Rother, D., Friedrich, C., and Scheidig, A. J.(2005) J. Struct. Biol. 152, 229–234
12. Sauve, V., Bruno, S., Berks, B. C., and Hemmings, A. M. (2007) J. Biol.Chem. 282, 23194–23204
13. Sauve, V., Roversi, P., Leath, K. J., Garman, E. F., Antrobus, R., Lea,S. M., and Berks, B. C. (2009) J. Biol. Chem. 284, 21707–21718
14. Friedrich, C. G., Quentmeier, A., Bardischewsky, F., Rother, D., Kraft, R.,Kostka, S., and Prinz, H. (2000) J. Bacteriol. 182, 4677–4687
15. Oliveira, T. F., Vonrhein, C., Matias, P. M., Venceslau, S. S., Pereira,I. A., and Archer, M. (2008) J. Biol. Chem. 283, 34141–34149
16. Bamford, V. A., Angove, H. C., Seward, H. E., Thomson, A. J., Cole, J. A.,Butt, J. N., Hemmings, A. M., and Richardson, D. J. (2002) Biochemistry41, 2921–2931
17. Kappler, U., and Bailey, S. (2005) J. Biol. Chem. 280, 24999–2500718. Bailey, S., Rapson, T., Johnson-Winters, K., Astashkin, A. V., Enemark,
J. H., and Kappler, U. (2009) J. Biol. Chem. 284, 2053–206319. Rapson, T. D., Astashkin, A. V., Johnson-Winters, K., Bernhardt, P. V.,
Kappler, U., Raitsimring, A. M., and Enemark, J. H. (2010) J. Biol. Inorg.Chem. 15, 505–514
20. Emesh, S., Rapson, T. D., Rajapakshe, A., Kappler, U., Bernhardt, P. V.,Tollin, G., and Enemark, J. H. (2009) Biochemistry 48, 2156–2163
21. Raitsimring, A. M., Kappler, U., Feng, C., Astashkin, A. V., and Enemark,J. H. (2005) Inorg. Chem. 44, 7283–7285
22. Kisker, C., Schindelin, H., Pacheco, A., Wehbi, W. A., Garrett, R. M.,Rajagopalan, K. V., Enemark, J. H., and Rees, D. C. (1997) Cell 91,973–983
23. Quentmeier, A., Kraft, R., Kostka, S., Klockenkamper, R., and Friedrich,C. G. (2000) Arch. Microbiol. 173, 117–125
24. Nurizzo, D., Mairs, T., Guijarro, M., Rey, V., Meyer, J., Fajardo, P., Cha-vanne, J., Biasci, J. C., McSweeney, S., and Mitchell, E. (2006) J. Synchro-tron Radiat. 13, 227–238
25. Kabsch, W. (2010) Acta Crystallogr. D 66, 133–14426. Panjikar, S., Parthasarathy, V., Lamzin, V. S., Weiss, M. S., and Tucker,
P. A. (2005) Acta Crystallogr. D 61, 449–45727. CCP4. (1994) Acta Crystallogr. D 50, 760–76328. Sheldrick, G. M., Hauptman, H. A., Weeks, C. M., Miller, R., and Uson,
I. (2001) International Tables for Crystallography, pp. 333–345, KluwerAcademic Publishers Group, Dordrecht, The Netherlands
29. Schneider, T. R., and Sheldrick, G. M. (2002) Acta Crystallogr. D 58,1772–1779
30. Hao, Q. (2004) J. Appl. Crystallogr. 37, 498–49931. Sheldrick, G. M. (2002) Z. Kristallogr. 217, 644–65032. Terwilliger, T. C. (2000) Acta Crystallogr. D 56, 965–97233. Cowtan, K. D., and Main, P. (1996) Acta Crystallogr. D 52, 43–4834. Perrakis, A., Morris, R., and Lamzin, V. S. (1999) Nat. Struct. Biol. 6,
458–46335. Morris, R. J., Perrakis, A., and Lamzin, V. S. (2003)Methods Enzymol.
374, 229–24436. Panjikar, S., Parthasarathy, V., Lamzin, V. S., Weiss, M. S., and Tucker,
P. A. (2009) Acta Crystallogr. D 65, 1089–109737. Adams, P. D., Afonine, P. V., Bunkoczi, G., Chen, V. B., Davis, I. W.,
Echols, N., Headd, J. J., Hung, L. W., Kapral, G. J., Grosse-Kunstleve,R. W., McCoy, A. J., Moriarty, N. W., Oeffner, R., Read, R. J., Richard-
son, D. C., Richardson, J. S., Terwilliger, T. C., and Zwart, P. H. (2010)Acta Crystallogr. D 66, 213–221
38. Murshudov, G. N., Vagin, A. A., and Dodson, E. J. (1997) Acta Crystal-logr. D 53, 240–255
39. Emsley, P., and Cowtan, K. (2004) Acta Crystallogr. D 60, 2126–213240. DeLano, W. L. (2005) Drug Discov. Today 10, 213–21741. Berman, H. M., Westbrook, J., Feng, Z., Gilliland, G., Bhat, T. N., Weis-
sig, H., Shindyalov, I. N., and Bourne, P. E. (2000) Nucleic Acids Res. 28,235–242
42. Holm, L., and Park, J. (2000) Bioinformatics 16, 566–56743. Laskowski, R. A. (1995) J. Mol. Graph 13, 323–33044. Hille, R. (2005) Arch. Biochem. Biophys. 433, 107–11645. Schwarz, G. (2005) Cell. Mol. Life Sci. 62, 2792–281046. Schrader, N., Fischer, K., Theis, K., Mendel, R. R., Schwarz, G., and
Kisker, C. (2003) Structure 11, 1251–126347. Bertero, M. G., Rothery, R. A., Palak, M., Hou, C., Lim, D., Blasco, F.,
Weiner, J. H., and Strynadka, N. C. J. (2003) Nat. Struct. Biol. 10,681–687
48. Wootton, J. C., Nicolson, R. E., Cock, J. M., Walters, D. E., Burke, J. F.,Doyle, W. A., and Bray, R. C. (1991) Biochim. Biophys. Acta 1057,157–185
49. Bowman, S. E., and Bren, K. L. (2008) Nat. Prod. Rep. 25, 1118–113050. Ozawa, K., Takayama, Y., Yasukawa, F., Ohmura, T., Cusanovich, M. A.,
Tomimoto, Y., Ogata, H., Higuchi, Y., and Akutsu, H. (2003) Biophys. J.85, 3367–3374
51. Fischer, K., Barbier, G. G., Hecht, H. J., Mendel, R. R., Campbell, W. H.,and Schwarz, G. (2005) Plant Cell 17, 1167–1179
52. Banci, L., Bertini, I., Quacquarini, G., Walter, O., Diaz, A., Hervas, M.,and De la Rosa, M. A. (1996) J. Biol. Inorg. Chem. 1, 330–340
53. Worrall, J. A., Schlarb-Ridley, B. G., Reda, T., Marcaida, M. J., Moorlen,R. J., Wastl, J., Hirst, J., Bendall, D. S., Luisi, B. F., and Howe, C. J. (2007)J. Am. Chem. Soc. 129, 9468–9475
54. Jones, S., Marin, A., and Thornton, J. M. (2000) Protein Eng. 13, 77–8255. Hobbs, J. D., and Shelnutt, J. A. (1995) J. Protein Chem. 14, 19–2556. Vaguine, A. A., Richelle, J., and Wodak, S. J. (1999) Acta Crystallogr. D
55, 191–20557. Hille, R. (1994) Biochim. Biophys. Acta 1184, 143–16958. Sezer, D., Prandolini, M. J., and Prisner, T. F. (2009) Phys. Chem. Chem.
Phys. 11, 6626–663759. Page, C. C., Moser, C. C., Chen, X., and Dutton, P. L. (1999) Nature 402,
47–5260. Kelly, D. P. (1982) Phil. Trans. R. Soc. B 298, 499–52861. Gouet, P., Courcelle, E., Stuart, D. I., and Metoz, F. (1999) Bioinformatics
15, 305–30862. Akazaki, H., Kawai, F., Chida, H., Matsumoto, Y., Hirayama, M.,
Hoshikawa, K., Unzai, S., Hakamata, W., Nishio, T., Park, S. Y., and Oku,T. (2008) Acta Crystallogr. F 64, 674–680
63. Wallace, A. C., Laskowski, R. A., and Thornton, J. M. (1995) Protein Eng.8, 127–134
64. Rocchia, W., Alexov, E., and Honig, B. (2001) J. Phys. Chem. B 105,6507–6514
65. Rocchia, W., Sridharan, S., Nicholls, A., Alexov, E., Chiabrera, A., andHonig, B. (2002) J. Comput. Chem. 23, 128–137
66. Petrek, M., Otyepka, M., Banas, P., Kosinova, P., Koca, J., and Dambor-sky, J. (2006) BMC Bioinformatics 7, 316
67. Weiss, M. (2001) J. Appl. Crystallogr. 34, 130–13568. Diederichs, K., and Karplus, P. A. (1997) Nat. Struct. Biol. 4,
269–27569. Brunger, A. T. (1992) Nature 355, 472–47570. Tickle, I. J., Laskowski, R. A., and Moss, D. S. (2000) Acta Crystallogr. D
56, 442–45071. Laskowski, R. A., MacArthur, M. W., Moss, D. S., and Thornton, J. M.
(1993) J. Appl. Crystallogr. 26, 283–291
Crystal Structure of Sulfane Dehydrogenase SoxCD
8360 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 10 • MARCH 11, 2011
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Armin Quentmeier, Frank Bardischewsky, Cornelius G. Friedrich and Axel J. ScheidigUlrich Zander, Annette Faust, Björn U. Klink, Daniele de Sanctis, Santosh Panjikar,
Molybdohemo-Enzyme Sulfane Dehydrogenase SoxCDStructural Basis for the Oxidation of Protein-bound Sulfur by the Sulfur Cycle
doi: 10.1074/jbc.M110.193631 originally published online December 8, 20102011, 286:8349-8360.J. Biol. Chem.
10.1074/jbc.M110.193631Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2010/12/13/M110.193631.DC1
http://www.jbc.org/content/286/10/8349.full.html#ref-list-1
This article cites 70 references, 11 of which can be accessed free at
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from