scarless healing of oral mucosa is characterized by faster resolution of inflammation and control of...
TRANSCRIPT

Journal of Dermatological Science 56 (2009) 168–180
Scarless healing of oral mucosa is characterized by faster resolution ofinflammation and control of myofibroblast action compared to skin woundsin the red Duroc pig model
Karen Mak a, Aleem Manji a,b, Corrie Gallant-Behm c,d, Colin Wiebe a, David A. Hart c,Hannu Larjava a, Lari Hakkinen a,*a Faculty of Dentistry, University of British Columbia, Vancouver, BC, Canadab Schulich School of Dentistry, University of Western Ontario, London, ON, Canadac Department of Surgery, McCaig Institute of Bone and Joint Health, University of Calgary, Calgary, AB, Canadad Department of Surgery, Feinberg School of Medicine, Northwestern University, Chicago, IL, USA
A R T I C L E I N F O
Article history:
Received 8 April 2009
Received in revised form 21 July 2009
Accepted 11 September 2009
Keywords:
Wound healing
Scar
Oral mucosa
Skin
Transforming growth factor-beta
A B S T R A C T
Background: Scar formation following skin trauma can have devastating consequences causing
physiological and psychosocial concerns. Currently, there are no accepted predictable treatments to
prevent scarring which emphasizes a need for a better understanding of the wound healing and scar
formation process.
Objectives: Previously it was shown that healing of small experimental wounds in the oral mucosa of red
Duroc pigs results in significantly reduced scar formation as compared with equivalent full-thickness
skin wounds. In the present study, scar formation was assessed in 17 times larger experimental wounds
in both oral mucosa and skin of the red Duroc pigs.
Methods: Equivalent experimental wounds were created in the oral mucosa and dorsal skin of red Duroc
pigs, and scar formation, localization and abundance of key wound healing cells, transforming growth
factor-b (TGF-b) and phosphorylated Smad3 (pSmad3) were assessed.
Results: Oral mucosal wounds displayed significantly less clinical and histological scar formation than
did the corresponding skin wounds. The number of macrophages, mast cells, TGF-b and pSmad3
immunopositive cells was significantly reduced in the oral mucosal wounds as compared with skin
wounds during the maturation stage of the healing process. Although the number of myofibroblasts was
significantly elevated, the oral mucosal wounds showed significantly less contraction than did the skin
wounds over time.
Conclusions: Earlier resolution of the inflammatory reaction and reduced wound contraction may
promote scarless oral mucosal wound healing. In addition, scar formation likely depends not only on the
number of myofibroblasts but also on the extracellular environment which regulates their function.
� 2009 Japanese Society for Investigative Dermatology. Published by Elsevier Ireland Ltd. All rights
reserved.
Contents lists available at ScienceDirect
Journal of Dermatological Science
journal homepage: www. int l .e lsev ierhea l th .com/ journa ls / jods
1. Introduction
Scar formation following trauma to skin is a common outcomeof the wound healing process. Clinically, scars can range from finelines to expansive hypertrophic or keloid scars. However,regardless of the type of scars that individuals suffer, they leadto morbidity that includes psychosocial impacts or physical
* Corresponding author at: Faculty of Dentistry, University of British Columbia,
Department of Oral Biological and Medical Sciences, Laboratory of Periodontal
Biology, 2199 Wesbrook Mall, Vancouver, BC, Canada V6T 1Z3.
Tel.: +1 604 822 0096; fax: +1 604 822 3562.
E-mail address: [email protected] (L. Hakkinen).
0923-1811/$36.00 � 2009 Japanese Society for Investigative Dermatology. Published b
doi:10.1016/j.jdermsci.2009.09.005
complications or both [1,2]. Recently, it was reported that at least45 million patients in the US alone undergo procedures each yearthat could benefit from therapies that reduce scar formation [2]. Inthe past few decades, use of various animal models has lead tosteady progress in the understanding of the wound healingprocess, and this has helped to identify factors that may promotescar-free wound regeneration. For example, novel information hasbeen obtained from studies where wound regeneration in newt,zebrafish, hydra and planaria has been compared to scar formingwound healing in mammals [3]. These studies have suggested thatan altered inflammatory response, expressions of transforminggrowth factor-b (TGF-b) isoforms and a distinct balance betweenproteolytic enzymes and their inhibitors may contribute to the scarformation [3–6]. Despite these advances in knowledge, currently
y Elsevier Ireland Ltd. All rights reserved.

K. Mak et al. / Journal of Dermatological Science 56 (2009) 168–180 169
no effective therapy to prevent scar formation exists, indicatingthat further understanding regarding the biological processes thatregulate wound healing and scar formation is needed.
A lack of suitable animal models in mammals has hampered theunderstanding of the biology of scarring since ethical reasons havelimited systematical studies in humans. Although several rodentmodels such as rats, mice, hamsters and rabbits have been used tostudy wound healing and scar formation, these loose-skinnedanimals have significant differences in the wound healing processand scar formation compared to humans [7]. On the other hand,porcine skin is similar to human skin in that both have a relativelythick epidermis with distinct rete pegs, a comparable collagenstructure, and they heal by similar mechanisms as human skinwounds resulting in scar formation [7–9]. We have recentlystudied healing of relatively small (2 mm � 12 mm) experimentalexcisional wounds in the oral palatal mucosa in pigs and similarwounds in humans. The results have shown that wound healing inthe oral mucosa results in significantly reduced scar formation ascompared to similar wounds in the skin [10]. In addition, both pigand human oral mucosal wounds showed similar changes in themolecular composition of the wounds overtime that were distinctfrom the corresponding skin wounds [10]. Thus, comparingscarless wound healing in the oral mucosa to comparable woundsin scar forming skin of these animals provides a novel model tosystematically study the mechanisms influencing scar formationoutcomes. Previous findings have indicated that wound size affectsscar formation in that larger wounds result in more frequent andsevere scarring [8,11–14]. Therefore, in the present study, wecompared wound healing and scar formation of larger wounds(15 mm � 27 mm) in the oral mucosa and skin of the red Durocpigs over time post-injury. Increased abundance of macrophages,mast cells, myofibroblasts, vasculature and signaling molecules ofthe TGF-b pathway have also been associated with scar formation[5,15–17]. To this end, we also compared their abundance betweenthe skin and oral mucosal wounds. The findings indicate that thepattern of results with these larger wounds (17 times larger)between the skin and oral mucosa is maintained, and thus, theprevious results were not due to the small wound sizes evaluated.Furthermore, novel information about the cellular and moleculardifferences at the two wound sites was identified.
2. Materials and methods
2.1. Animal experimentation
Twelve juvenile, female red Duroc pigs, weighing approxi-mately 30 kg, were obtained from the Neufeld Farm (Acme, AB,Canada). The pigs were housed at the University of Calgary AnimalCare Facility in accordance with Canadian Council of Animal Careguidelines with 12-h light/dark cycles and fed antibiotic-free foodand water ad libitum. The procedures were approved by the AnimalCare Committee of the Faculty of Medicine (University of Calgary)and were performed as described previously [18,19]. Briefly, theanimals were premedicated with ketamine (15 mg/kg i.m.) andacepromazine (0.4 mg/kg), and general anesthesia was induced bythe administration of 1–2% isofluorane by mask. In each pig, twoidentical full-thickness excisional wounds (15 mm � 27 mm) were
Table 1Modified Manchester Scar Proforma for clinical scar assessment.
Score/parameters 0
Color Perfect match with the surrounding tissue
Contour Flush with surrounding tissue
Distortion None
Total score 0–6
created in the left and right side of the oral mucosal gingiva of thehard palate using a scalpel. This is the maximum size of thewounds that can be created in this tissue site. In addition, each pigreceived ten dorsal skin wounds identical in size to those in the oralmucosa on the left and right side of the spine. The pigs weredivided to the experimental groups so that 6 pigs received the 3-and 28-day wounds on the left and right side, respectively, whileanother set of 6 pigs received, 14- and 60-day wounds,respectively. Thus, in total 6 parallel oral mucosal wounds and60 skin wounds were created in 6 different animals at each timepoint. The skin wounds were placed a minimum of 20 mm apart. Inthe oral mucosa, full-thickness tissue near to the depth of thepalatal bone was removed leaving some soft tissue above theperiosteum. In the skin, the epidermis, dermis and subcutaneousfat were removed exposing the underlying fascia. The oral mucosalwounds were left untreated to heal while the skin wounds weresprayed with gentamycin containing wound-spray and bandagedfor 3 days. In order to assess wound healing and scar formationclinically and to quantify wound contraction, standardized digitalimages were taken of the unwounded tissue and from the woundsimmediately after wounding and at days 3, 11, 14, 25, 28, 32, 46, 57and 60 after wounding. All the animals were sacrificed at the sametime and the wound samples collected at that time. Tissue biopsieswere collected from unwounded tissue (day 0 samples) and fromthe wounds including 3 mm of the surrounding tissue at days 3, 14,28 and 60 after wounding. The tissue samples were embedded inOptimal Cutting Temperature Compound (Tissue-Tek1, SakuraFinetek Inc., Torrance, CA, USA) and immediately frozen in liquidnitrogen. Tissue sections (6 mm thick) were prepared using a 2800Frigocut Cryostat Microtome (Leica, Nussloch, Germany) andplaced on 3-amino-opropyltriethoxysilane-coated slides andstored at �86 8C until use.
2.2. Assessment of wound contraction and clinical scar formation
Wound contraction was quantified as described previously[19]. Briefly, clinical digital images were taken from six parallelanimals at each time (n = 6 oral mucosal wounds and 60 skinwounds at each time point). The images were calibrated by using astandard ruler placed in the frame. The original wound margin(edge of unwounded tissue) was easily identified based onpigmentation, texture, and lack of hair in the skin, and byanatomical location and lack of rugae in the palate. The totalwound surface area was measured as the area within the originalwound margin (irrespective of the presence or absence of a de novoepithelium) using ImageJ software (http://rsb.info.nih.gov/ij/; USNational Institutes of Health, Bethesda, MD, USA). The results arepresented as mean area � SEM. Statistical significance of thedifference between skin and oral mucosal scores at the given timepoint was determined using Student’s t-test and multiple compar-isons were performed by Anova (*p � 0.05; **p � 0.01; ***p � 0.001).
To assess clinical scar development, the standardized digitalimages from days 32, 46 and 60 wounds were assessed by twoindependent, calibrated investigators using a modified version ofthe Manchester Scar Proforma (Table 1) [20]. In total 6 oralmucosal wounds and 60 skin wounds were analyzed at the giventime point. Scar formation was assessed by comparing the color,
1 2
Slight mismatch Obvious mismatch
Slightly proud or indented Hypertrophic
Moderate Severe

Table 2Histological scar assessment scale.
Score/epithelium 0 1 2
Rete ridges Normal Some restoration A lot more/less restoration
Thickness Normal Slightly thicker/thinner A lot thicker/thinner
Score/connective tissue 0 1 2
Collagen fiber bundle orientation Normal basket-weave Some parallel/no orientation A lot parallel/no orientation
Collagen fiber bundle density Normal Slightly abnormal A lot abnormal
Collagen fiber bundle maturity Normal Slightly abnormal A lot abnormal
Fibroblast density Normal Slightly more/less A lot more/less
Total score 0–12
K. Mak et al. / Journal of Dermatological Science 56 (2009) 168–180170
contour and distortion of the wound site to the unwounded normaltissue. A value of 0, 1 or 2 was given for each parameter, with nodifference to normal tissue given a score of 0 and gross mismatchscoring 2. The values for each parameter were summed for eachsample for a total scar score. The results are presented as meanscores � SEM of wounds at the given group, and statisticalsignificance between palatal mucosal and skin wounds werecompared at a given time point using the nonparametric Mann–Whitney test (*p < 0.05; **p < 0.01; ***p < 0.001).
2.3. Histological scar assessment
A modified version of the Histological scar assessment scale wasused to assess scar development histologically (Table 2) [20]. In theepithelium, presence or absence of rete ridges and epidermalthickness were evaluated. In the connective tissue, collagen fiberbundle orientation (basket-weave to parallel or no orientation),density (inter-fiber bundle space), maturity (fiber bundle thick-ness) and fibroblast density (number of spindle-shaped connectivetissue cells per unit area) were assessed with a score of 0, 1 or 2given for each parameter (score of 0 indicated no difference fromunwounded, normal tissue while a value of 2 indicated grossmismatch). For the assessment, two independent, calibratedexaminers evaluated tissue sections from the unwounded tissueand from the mid-wound area of 28- and 60-day wounds from fourparallel animals at each time point. For the evaluation, the sectionswere stained with hematoxylin and eosin (H&E) and viewed usingan Axiolab E light microscope (Carl Zeiss, Jena, Germany). UV light-induced eosin autofluorescence emitted from collagen fiberbundles was examined using a rhodamine filter in a Laser ConfocalScanning Microscope (Carl Zeiss) to assess collagen organization.The results are presented as mean scores � SEM of four parallelwounds from different animals at each time point. Statisticalsignificance between palatal mucosal and skin wounds werecompared at a given time point using the nonparametric Mann–Whitney test (*p < 0.05; **p < 0.01; ***p < 0.001).
2.4. Primary antibodies
Mouse anti-CD163 (MCA2311) and anti-mast cell tryptase(MCA1438T) antibodies were purchased from AbD Serotec(Raleigh, NC, USA). Mouse anti-a-smooth muscle actin (a-SMA;A2547) and rabbit anti-laminin-1 antibodies were purchased fromSigma (Saint Louis, MO, USA). Rabbit anti-TGF-b1, 2, 3 antibodywas purchased from Santa Cruz Biotechnology (Santa Cruz, CA,USA) and rabbit anti-phosphorylated Smad3 (pSmad3) antibodywas purchased from Abcam (Cambridge, MA, USA).
2.5. Immunostaining
Three to six parallel frozen tissue sections from unwoundedtissue (day 0) and from the mid-wound area of 3-, 14-, 28- and 60-day wounds from four parallel animals at each time point were
used for immunohistochemical staining. To this end, tissuesections were fixed with ice-cold acetone at room temperaturefor 5 min, washed and rehydrated in phosphate buffered saline(PBS; pH 7.4) for 5 min. Nonspecific antibody binding sites wereblocked by incubation with appropriate normal blocking serum(Vectastain Kit, Vector Laboratories, Burlingame, USA) at roomtemperature for 30 min. For immunofluorescence staining ofmacrophages (anti-CD163 antibody; 1:200 dilution), blood vessels(anti-laminin-1 antibody; 1:50 dilution) and myofibroblasts (anti-aSMA antibody; 1:100 dilution), the primary antibodies werediluted in PBS containing bovine serum albumin (BSA; 1 mg/ml)and Triton X-100 (0.01%) and incubated on the samples in ahumidified chamber at 4 8C for 18 h followed by an incubationwith appropriate Alexa-488 conjugated secondary antibody (1:100dilution; Invitrogen, Eugene, OR, USA) at room temperature for 1 h.The sections were mounted with Immu-Mount solution (ThermoShandon, Pittsburg, PA, USA).
The immunostaining for mast cell tryptase (1:500 dilution ofthe antibody), pSmad3 (1:50 dilution) and TGF-b1,2,3 (1:50dilution) was performed using the Vectastain ABC Elite Kit andVector DAB Substrate Kit (Vector Laboratories). To this end,samples were fixed, the nonspecific binding sites were blocked andsamples were incubated with the primary antibody as above. Thiswas followed by incubation with the appropriate biotinylatedsecondary antibody (Vectastain ABC Elite Kit) at room temperaturefor 1 h. The sections were then incubated with freshly preparedVectastain ABC reagent at room temperature for 30 min. DABsubstrate was prepared according to manufacturer’s instructionsand allowed to react with the samples until suitable colordevelopment was noted. Color development was terminated bya wash with distilled water. All samples were stained at the sametime with the given antibody and a standardized reaction time forDAB color development was used for each antibody. The sectionswere mounted using Vecta-Mount solution (Vector Laboratories).Control stainings were performed by omitting the primaryantibody or by replacing the primary antibody with thecorresponding non-immune serum and gave only a very weaknonspecific background staining (data not shown).
2.6. Quantitative image analysis
Representative, standardized images were taken from three tosix parallel immunostained sections from each unwounded tissueor mid-wound sample using the Zeiss Laser Confocal ScanningMicroscope (Carl Zeiss). Images were captured using NorthernEclipse software (Empix, Imaging, Mississauga, ON, Canada) usinga standard offset (background darkness) and exposure time(overall brightness) individually standardized for each antibody.For quantification, the grey scale images were converted to binaryimages and immunopositive cells were separated using thewatershed function and quantified using the particle analysisfunction in the ImageJ software (NIH). The subepithelial (0.28 mm2
area from the basement membrane zone or from the surface of a

K. Mak et al. / Journal of Dermatological Science 56 (2009) 168–180 171
non-epithelialized wound to the depth of 1 mm) and deep (similararea located deeper than the superficial area) connective tissueareas were quantified separately. Myofibroblasts were identifiedbased on their positive immunoreactivity for a-SMA and theirfibroblast-like, spindle-shaped morphology. a-SMA positive cellsrepresenting pericytes that formed brightly stained cellularclusters that resembled morphologically blood vessels wereexcluded from the calculations of myofibroblast density. Theresults are presented as mean scores � SEM of four parallel woundsfrom different animals at each time point and statistical significancebetween skin and oral mucosal scores at the given time point orrelative to day 0 sample in a given tissue was determined usingStudent’s t-test and multiple comparisons were performed by usingAnova (*p < 0.05; **p < 0.01; ***p < 0.001). In cases where there wereno statistically significant differences in the quantitative results for agiven molecule in the subepithelial and deep tissue, the results fromthe two areas were pooled together.
3. Results
3.1. Large oral mucosal wounds show significantly reduced
clinical scar formation and wound contraction as compared
to similar size skin wounds
Healing of all wounds occurred without complications orwound infections. Clinically, both oral mucosal (n = 6) and skinwounds (n = 60) were open and covered by a blood clot at day 3after wounding. A mild inflammatory erythema was notedsurrounding the wounds. At day 14, all oral mucosal and skinwounds were still open, but the inflammation was clearly reducedas compared to day 3. At day 14, no crust was present andmigrating epithelium had started to partially cover the oralmucosal wounds. In contrast, the skin wounds were completelycovered with a crust which was not removed by the investigators.At day 28, 5 out of the 6 oral mucosal wounds were completelyclosed, and in those wounds no clinical signs of inflammation werenoted. In the wound that was partially open, a mild inflammationwas noted at the wound margins. At this point, all skin wounds(n = 60) were still partially covered with a crust, but the uncoveredwound area appeared relatively free of inflammation and wascovered with epithelium. At day 32, all of the 6 oral mucosalwounds were closed. At day 32 and 46 after wounding, 76% (46 outof 60) and 38% (23 out of 60) of the skin wounds still had a crustpartially covering the wound site, respectively, although theuncovered area appeared healed At day 60, the skin woundsappeared free of crust. During the late stages of wound healing,scar formation was evident in skin while oral mucosal woundsresembled unwounded mucosa except that the prominent palatalrugae had not reformed (Fig. 1a). Scar formation was assessedclinically using the modified Manchester Scar Proforma (Table 1) atday 32, 46 and 60 after wounding. The oral mucosal wounds hadsignificantly lower scores in color, contour and distortion resultingin significantly lower total scar score by day 60 post-woundingcompared to skin wounds (Fig. 1b).
Wound contraction was studied over time by measuring thewound area based on the location of the original wound margins. Asignificant difference was noted between the area of oral mucosaland skin wounds around day 3 where the wounds began tocontract in skin, while the oral mucosal wounds increased slightlyin size (Fig. 1c). After that, the contraction occurred in parallel inboth tissues until day 25. This was followed by significantdifferences in wound contraction as the contraction rate increasedin skin at days 28 and 32 while the oral mucosal wounds relaxedback to their original size over time. Wound areas in skin remainedrelatively constant from days 32 to 60, and showed contraction toabout 30% of the original size.
3.2. Large oral mucosal wounds show significantly better
regeneration of connective tissue organization and reduced
histological scar formation as compared to similar size skin wounds
Histological analysis showed that at day 3 after wounding, ablood clot with numerous inflammatory cells filled the woundspace, and the wound epithelium had migrated for a short distancealong the wound edge between the connective tissue and the bloodclot in both oral mucosal and skin wounds (Fig. 2a, E–H). At day 14,in both tissues the migration of the wound epithelium hadprogressed across the wound bed, but the middle two thirds of thewound space were still open. In the wound connective tissue atboth sites, a highly cellular granulation tissue had formed. A denseinflammatory cell infiltrate was noted on the most superficial areaof the open wound surface, while the other areas showed amoderate inflammatory cell infiltrate (Fig. 2a, I–L). At day 28, bothoral mucosal and skin wounds were fully re-epithelialized with athickened epithelium and varying degrees of rete ridge formation.The number of inflammatory cells in the wound area was stronglyreduced as compared to earlier time points. Unlike oral mucosalwounds, skin wounds also showed wound contraction that hadbrought the wound edges closer together (Fig. 2a, M–P). In the oralmucosal wound connective tissue, the density of the cells withtypical spindle-shaped fibroblast morphology was significantlylower as compared to skin wounds (Fig. 2c). In addition, fibroblastswere aligned perpendicularly to the wound edges, while in the skinwounds they appeared more round and more randomly arranged(Fig. 2a, inserts in N and P, respectively). At day 60, the woundconnective tissue in the oral mucosa and skin was less cellular thanat the earlier time points, but the oral mucosal wounds still showedsignificantly lower fibroblast density as compared to skin wounds(Fig. 2a, Q–T and Fig. 2c). In the oral mucosal wounds, epithelialrete ridge formation and epidermal thickness were comparable tothe unwounded tissue, while in the skin the epithelial rete ridgeswere not normalized by day 60 after wounding (Fig. 2a, Q and S).
In order to analyze the organization of the collagen-rich woundconnective tissue extracellular matrix, eosin stained tissuesections were viewed under fluorescent light (Fig. 2b). Inunwounded skin, the collagen fiber bundles were relatively thickand formed a typical basket-weave organization. In the oralmucosa, the collagen fiber bundles appeared slightly thinner andmore randomly organized as compared to skin. At day 14 afterwounding, the extracellular matrix in the mid-wound connectivetissue was composed of very thin, randomly organized fibers(Fig. 2b, B and F). At day 28, somewhat thicker fiber bundles werepresent in both oral mucosal and skin wound connective tissue.However, the bundles were much thinner, showed larger spacingand were more randomly organized compared to the unwoundedtissue (Fig. 2b, C and G and Fig. 2c). At day 60, the organization(orientation, density and maturity) of the collagen fiber bundleswere significantly closer to unwounded tissue in oral mucosalwounds as compared to skin (Fig. 2b, D and H and Fig. 2c). Thehistological scar assessment showed significantly lower scores forthe wound connective tissue parameters as compared with skin,resulting in a significantly lower total histological scar score in oralmucosa as compared to skin 60-day post-wounding (Fig. 2c).
3.3. Skin wounds show an increased number of macrophages as
compared to oral mucosal wounds
It has been demonstrated that macrophages may promote scarformation [17]. To this end, macrophages in the wound sub-epithelial and deep connective tissue were identified by immu-nostaining for the macrophage cell surface protein CD163 followedby quantification of the cell numbers by image analysis (Fig. 3a andb). No statistically significant differences were noted between
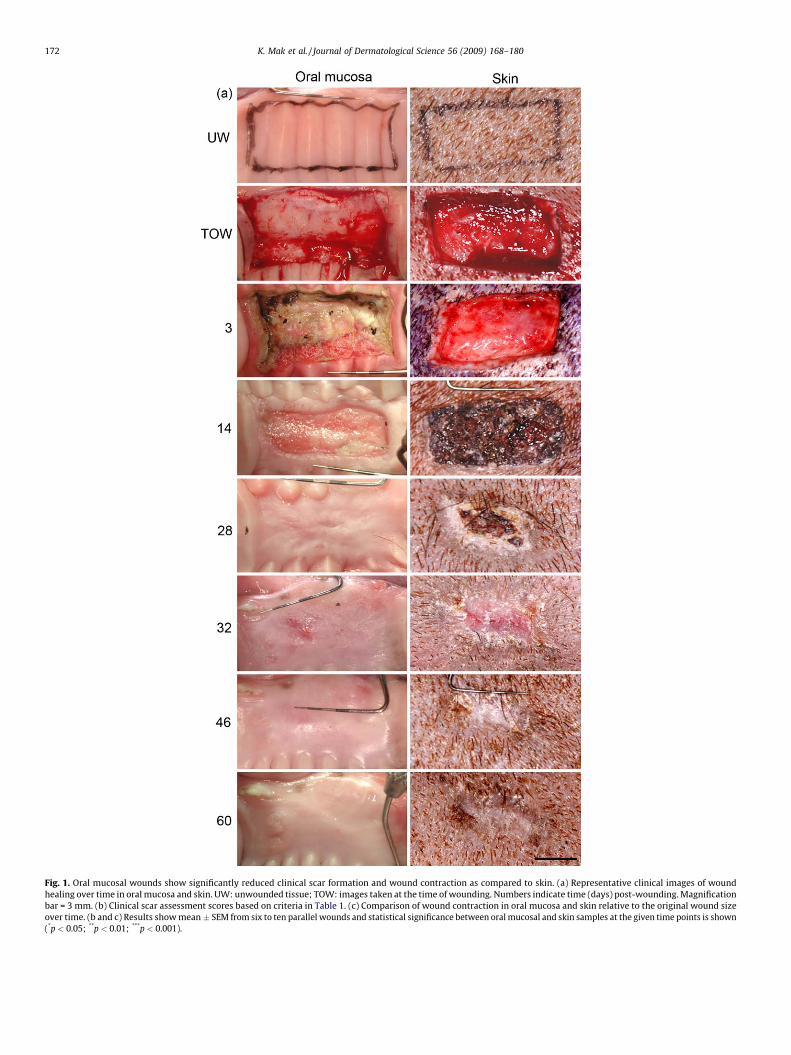
Fig. 1. Oral mucosal wounds show significantly reduced clinical scar formation and wound contraction as compared to skin. (a) Representative clinical images of wound
healing over time in oral mucosa and skin. UW: unwounded tissue; TOW: images taken at the time of wounding. Numbers indicate time (days) post-wounding. Magnification
bar = 3 mm. (b) Clinical scar assessment scores based on criteria in Table 1. (c) Comparison of wound contraction in oral mucosa and skin relative to the original wound size
over time. (b and c) Results show mean � SEM from six to ten parallel wounds and statistical significance between oral mucosal and skin samples at the given time points is shown
(*p < 0.05; **p < 0.01; ***p < 0.001).
K. Mak et al. / Journal of Dermatological Science 56 (2009) 168–180172
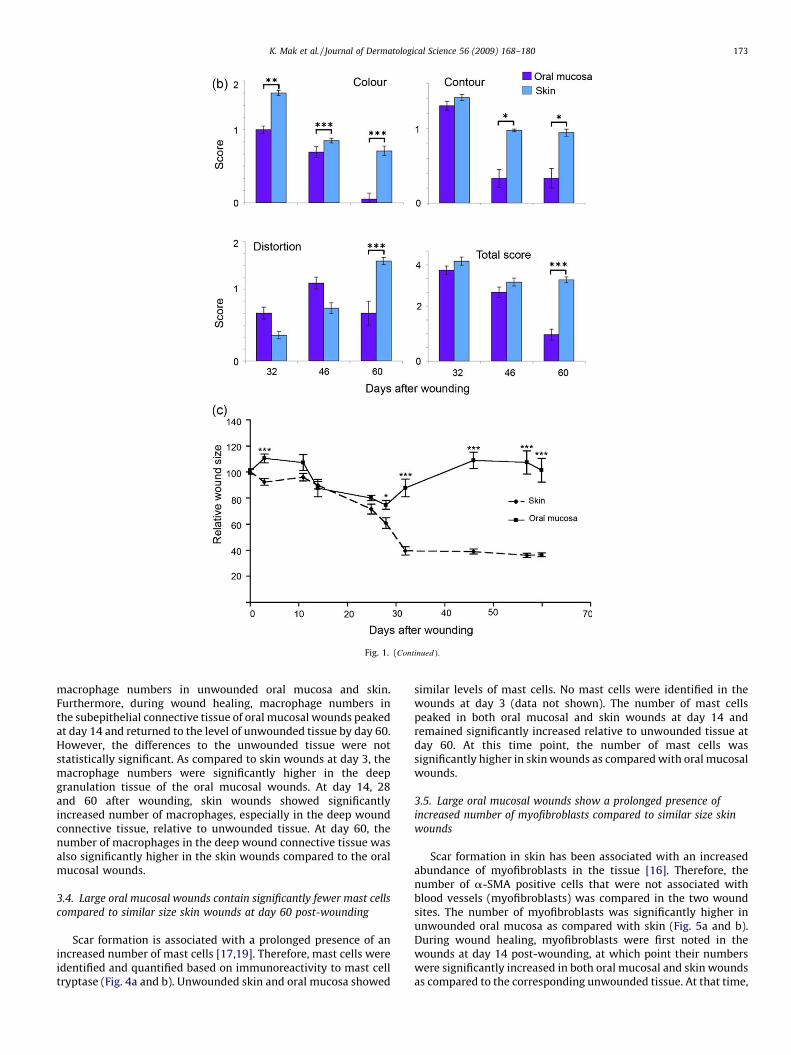
Fig. 1. (Continued ).
K. Mak et al. / Journal of Dermatological Science 56 (2009) 168–180 173
macrophage numbers in unwounded oral mucosa and skin.Furthermore, during wound healing, macrophage numbers inthe subepithelial connective tissue of oral mucosal wounds peakedat day 14 and returned to the level of unwounded tissue by day 60.However, the differences to the unwounded tissue were notstatistically significant. As compared to skin wounds at day 3, themacrophage numbers were significantly higher in the deepgranulation tissue of the oral mucosal wounds. At day 14, 28and 60 after wounding, skin wounds showed significantlyincreased number of macrophages, especially in the deep woundconnective tissue, relative to unwounded tissue. At day 60, thenumber of macrophages in the deep wound connective tissue wasalso significantly higher in the skin wounds compared to the oralmucosal wounds.
3.4. Large oral mucosal wounds contain significantly fewer mast cells
compared to similar size skin wounds at day 60 post-wounding
Scar formation is associated with a prolonged presence of anincreased number of mast cells [17,19]. Therefore, mast cells wereidentified and quantified based on immunoreactivity to mast celltryptase (Fig. 4a and b). Unwounded skin and oral mucosa showed
similar levels of mast cells. No mast cells were identified in thewounds at day 3 (data not shown). The number of mast cellspeaked in both oral mucosal and skin wounds at day 14 andremained significantly increased relative to unwounded tissue atday 60. At this time point, the number of mast cells wassignificantly higher in skin wounds as compared with oral mucosalwounds.
3.5. Large oral mucosal wounds show a prolonged presence of
increased number of myofibroblasts compared to similar size skin
wounds
Scar formation in skin has been associated with an increasedabundance of myofibroblasts in the tissue [16]. Therefore, thenumber of a-SMA positive cells that were not associated withblood vessels (myofibroblasts) was compared in the two woundsites. The number of myofibroblasts was significantly higher inunwounded oral mucosa as compared with skin (Fig. 5a and b).During wound healing, myofibroblasts were first noted in thewounds at day 14 post-wounding, at which point their numberswere significantly increased in both oral mucosal and skin woundsas compared to the corresponding unwounded tissue. At that time,
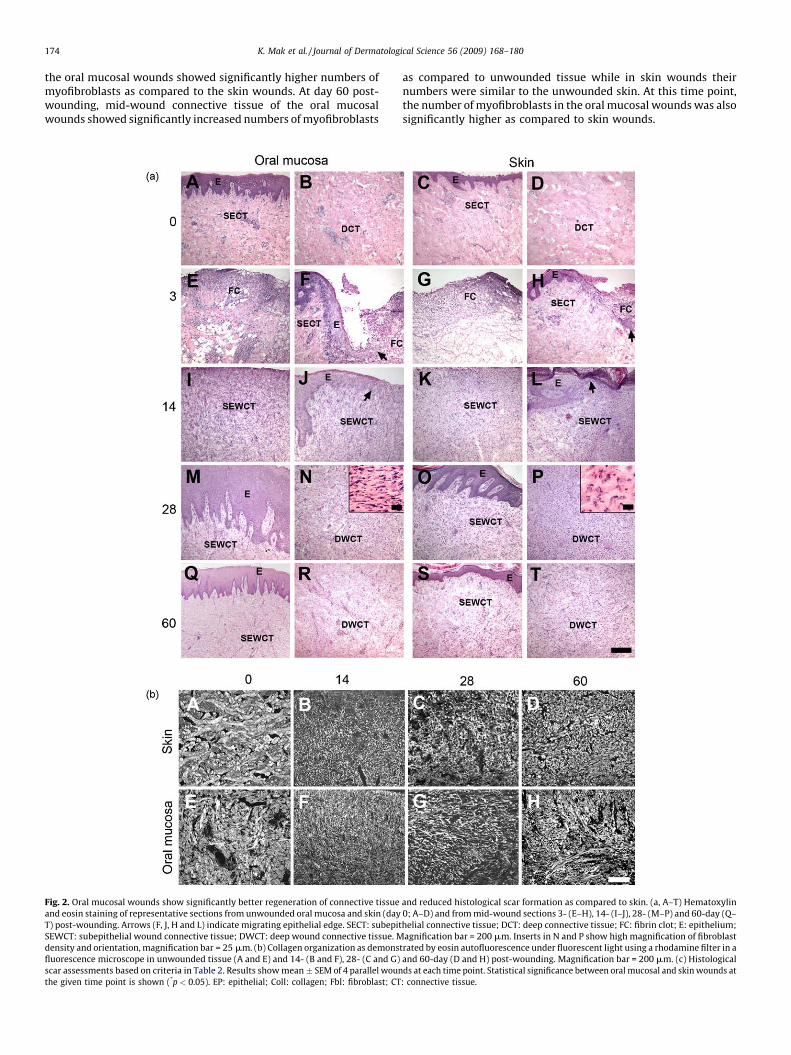
K. Mak et al. / Journal of Dermatological Science 56 (2009) 168–180174
the oral mucosal wounds showed significantly higher numbers ofmyofibroblasts as compared to the skin wounds. At day 60 post-wounding, mid-wound connective tissue of the oral mucosalwounds showed significantly increased numbers of myofibroblasts
Fig. 2. Oral mucosal wounds show significantly better regeneration of connective tissue
and eosin staining of representative sections from unwounded oral mucosa and skin (day
T) post-wounding. Arrows (F, J, H and L) indicate migrating epithelial edge. SECT: subepit
SEWCT: subepithelial wound connective tissue; DWCT: deep wound connective tissue. M
density and orientation, magnification bar = 25 mm. (b) Collagen organization as demons
fluorescence microscope in unwounded tissue (A and E) and 14- (B and F), 28- (C and G)
scar assessments based on criteria in Table 2. Results show mean � SEM of 4 parallel woun
the given time point is shown (*p < 0.05). EP: epithelial; Coll: collagen; Fbl: fibroblast; CT
as compared to unwounded tissue while in skin wounds theirnumbers were similar to the unwounded skin. At this time point,the number of myofibroblasts in the oral mucosal wounds was alsosignificantly higher as compared to skin wounds.
and reduced histological scar formation as compared to skin. (a, A–T) Hematoxylin
0; A–D) and from mid-wound sections 3- (E–H), 14- (I–J), 28- (M–P) and 60-day (Q–
helial connective tissue; DCT: deep connective tissue; FC: fibrin clot; E: epithelium;
agnification bar = 200 mm. Inserts in N and P show high magnification of fibroblast
trated by eosin autofluorescence under fluorescent light using a rhodamine filter in a
and 60-day (D and H) post-wounding. Magnification bar = 200 mm. (c) Histological
ds at each time point. Statistical significance between oral mucosal and skin wounds at
: connective tissue.
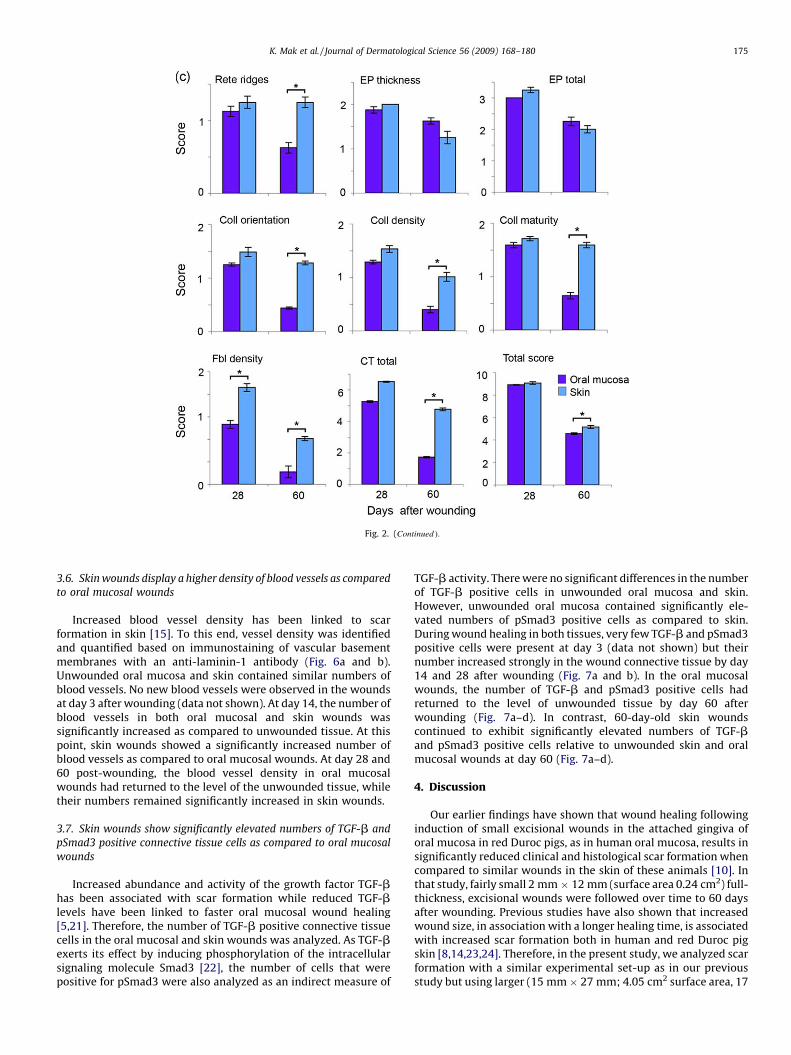
Fig. 2. (Continued ).
K. Mak et al. / Journal of Dermatological Science 56 (2009) 168–180 175
3.6. Skin wounds display a higher density of blood vessels as compared
to oral mucosal wounds
Increased blood vessel density has been linked to scarformation in skin [15]. To this end, vessel density was identifiedand quantified based on immunostaining of vascular basementmembranes with an anti-laminin-1 antibody (Fig. 6a and b).Unwounded oral mucosa and skin contained similar numbers ofblood vessels. No new blood vessels were observed in the woundsat day 3 after wounding (data not shown). At day 14, the number ofblood vessels in both oral mucosal and skin wounds wassignificantly increased as compared to unwounded tissue. At thispoint, skin wounds showed a significantly increased number ofblood vessels as compared to oral mucosal wounds. At day 28 and60 post-wounding, the blood vessel density in oral mucosalwounds had returned to the level of the unwounded tissue, whiletheir numbers remained significantly increased in skin wounds.
3.7. Skin wounds show significantly elevated numbers of TGF-b and
pSmad3 positive connective tissue cells as compared to oral mucosal
wounds
Increased abundance and activity of the growth factor TGF-bhas been associated with scar formation while reduced TGF-blevels have been linked to faster oral mucosal wound healing[5,21]. Therefore, the number of TGF-b positive connective tissuecells in the oral mucosal and skin wounds was analyzed. As TGF-bexerts its effect by inducing phosphorylation of the intracellularsignaling molecule Smad3 [22], the number of cells that werepositive for pSmad3 were also analyzed as an indirect measure of
TGF-b activity. There were no significant differences in the numberof TGF-b positive cells in unwounded oral mucosa and skin.However, unwounded oral mucosa contained significantly ele-vated numbers of pSmad3 positive cells as compared to skin.During wound healing in both tissues, very few TGF-b and pSmad3positive cells were present at day 3 (data not shown) but theirnumber increased strongly in the wound connective tissue by day14 and 28 after wounding (Fig. 7a and b). In the oral mucosalwounds, the number of TGF-b and pSmad3 positive cells hadreturned to the level of unwounded tissue by day 60 afterwounding (Fig. 7a–d). In contrast, 60-day-old skin woundscontinued to exhibit significantly elevated numbers of TGF-band pSmad3 positive cells relative to unwounded skin and oralmucosal wounds at day 60 (Fig. 7a–d).
4. Discussion
Our earlier findings have shown that wound healing followinginduction of small excisional wounds in the attached gingiva oforal mucosa in red Duroc pigs, as in human oral mucosa, results insignificantly reduced clinical and histological scar formation whencompared to similar wounds in the skin of these animals [10]. Inthat study, fairly small 2 mm � 12 mm (surface area 0.24 cm2) full-thickness, excisional wounds were followed over time to 60 daysafter wounding. Previous studies have also shown that increasedwound size, in association with a longer healing time, is associatedwith increased scar formation both in human and red Duroc pigskin [8,14,23,24]. Therefore, in the present study, we analyzed scarformation with a similar experimental set-up as in our previousstudy but using larger (15 mm � 27 mm; 4.05 cm2 surface area, 17
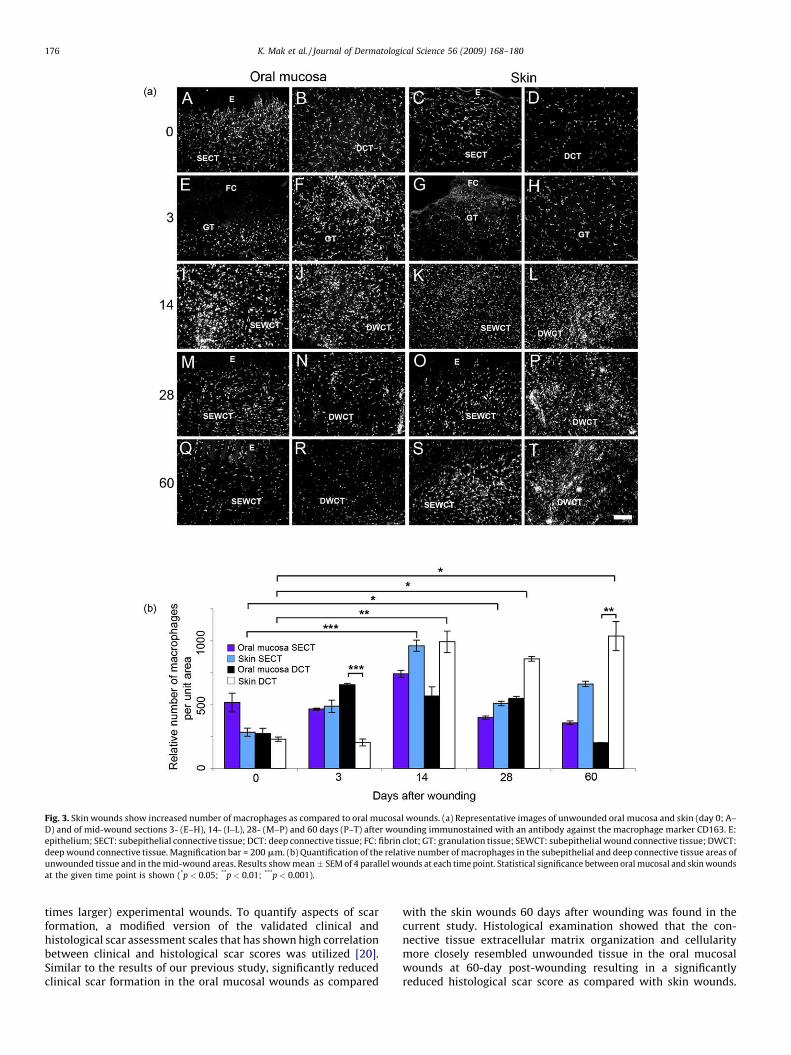
Fig. 3. Skin wounds show increased number of macrophages as compared to oral mucosal wounds. (a) Representative images of unwounded oral mucosa and skin (day 0; A–
D) and of mid-wound sections 3- (E–H), 14- (I–L), 28- (M–P) and 60 days (P–T) after wounding immunostained with an antibody against the macrophage marker CD163. E:
epithelium; SECT: subepithelial connective tissue; DCT: deep connective tissue; FC: fibrin clot; GT: granulation tissue; SEWCT: subepithelial wound connective tissue; DWCT:
deep wound connective tissue. Magnification bar = 200 mm. (b) Quantification of the relative number of macrophages in the subepithelial and deep connective tissue areas of
unwounded tissue and in the mid-wound areas. Results show mean � SEM of 4 parallel wounds at each time point. Statistical significance between oral mucosal and skin wounds
at the given time point is shown (*p < 0.05; **p < 0.01; ***p < 0.001).
K. Mak et al. / Journal of Dermatological Science 56 (2009) 168–180176
times larger) experimental wounds. To quantify aspects of scarformation, a modified version of the validated clinical andhistological scar assessment scales that has shown high correlationbetween clinical and histological scar scores was utilized [20].Similar to the results of our previous study, significantly reducedclinical scar formation in the oral mucosal wounds as compared
with the skin wounds 60 days after wounding was found in thecurrent study. Histological examination showed that the con-nective tissue extracellular matrix organization and cellularitymore closely resembled unwounded tissue in the oral mucosalwounds at 60-day post-wounding resulting in a significantlyreduced histological scar score as compared with skin wounds.
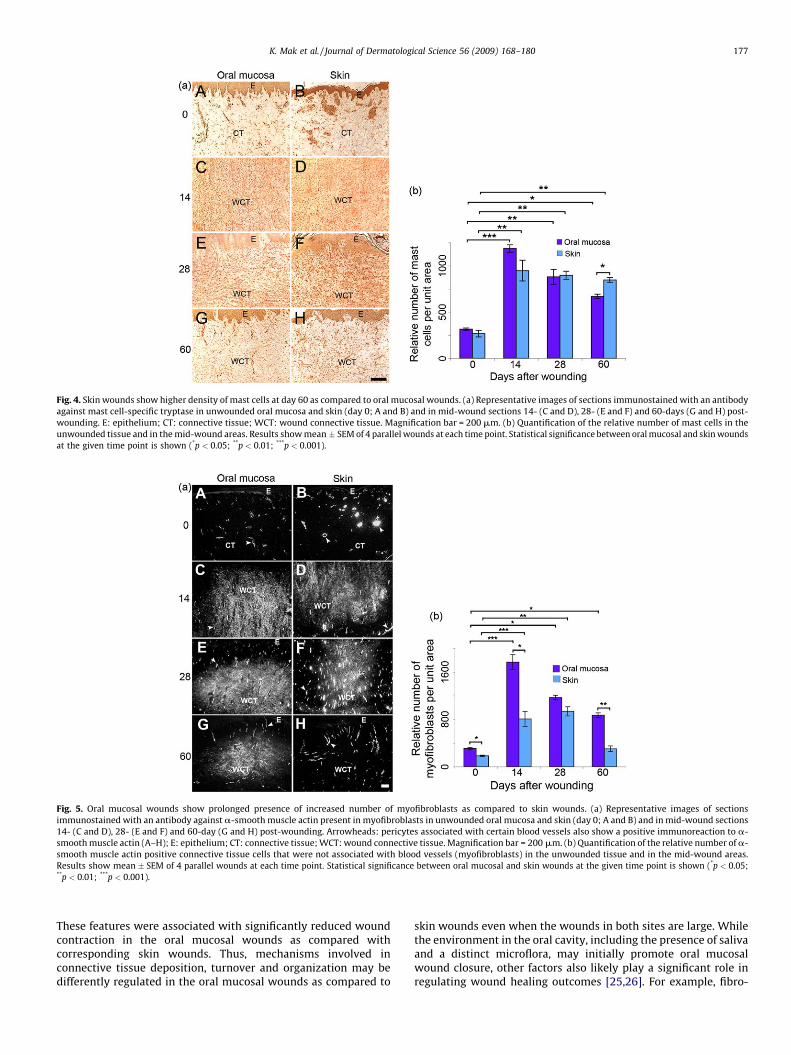
Fig. 4. Skin wounds show higher density of mast cells at day 60 as compared to oral mucosal wounds. (a) Representative images of sections immunostained with an antibody
against mast cell-specific tryptase in unwounded oral mucosa and skin (day 0; A and B) and in mid-wound sections 14- (C and D), 28- (E and F) and 60-days (G and H) post-
wounding. E: epithelium; CT: connective tissue; WCT: wound connective tissue. Magnification bar = 200 mm. (b) Quantification of the relative number of mast cells in the
unwounded tissue and in the mid-wound areas. Results show mean � SEM of 4 parallel wounds at each time point. Statistical significance between oral mucosal and skin wounds
at the given time point is shown (*p < 0.05; **p < 0.01; ***p < 0.001).
Fig. 5. Oral mucosal wounds show prolonged presence of increased number of myofibroblasts as compared to skin wounds. (a) Representative images of sections
immunostained with an antibody against a-smooth muscle actin present in myofibroblasts in unwounded oral mucosa and skin (day 0; A and B) and in mid-wound sections
14- (C and D), 28- (E and F) and 60-day (G and H) post-wounding. Arrowheads: pericytes associated with certain blood vessels also show a positive immunoreaction to a-
smooth muscle actin (A–H); E: epithelium; CT: connective tissue; WCT: wound connective tissue. Magnification bar = 200 mm. (b) Quantification of the relative number of a-
smooth muscle actin positive connective tissue cells that were not associated with blood vessels (myofibroblasts) in the unwounded tissue and in the mid-wound areas.
Results show mean � SEM of 4 parallel wounds at each time point. Statistical significance between oral mucosal and skin wounds at the given time point is shown (*p < 0.05;**p < 0.01; ***p < 0.001).
K. Mak et al. / Journal of Dermatological Science 56 (2009) 168–180 177
These features were associated with significantly reduced woundcontraction in the oral mucosal wounds as compared withcorresponding skin wounds. Thus, mechanisms involved inconnective tissue deposition, turnover and organization may bedifferently regulated in the oral mucosal wounds as compared to
skin wounds even when the wounds in both sites are large. Whilethe environment in the oral cavity, including the presence of salivaand a distinct microflora, may initially promote oral mucosalwound closure, other factors also likely play a significant role inregulating wound healing outcomes [25,26]. For example, fibro-
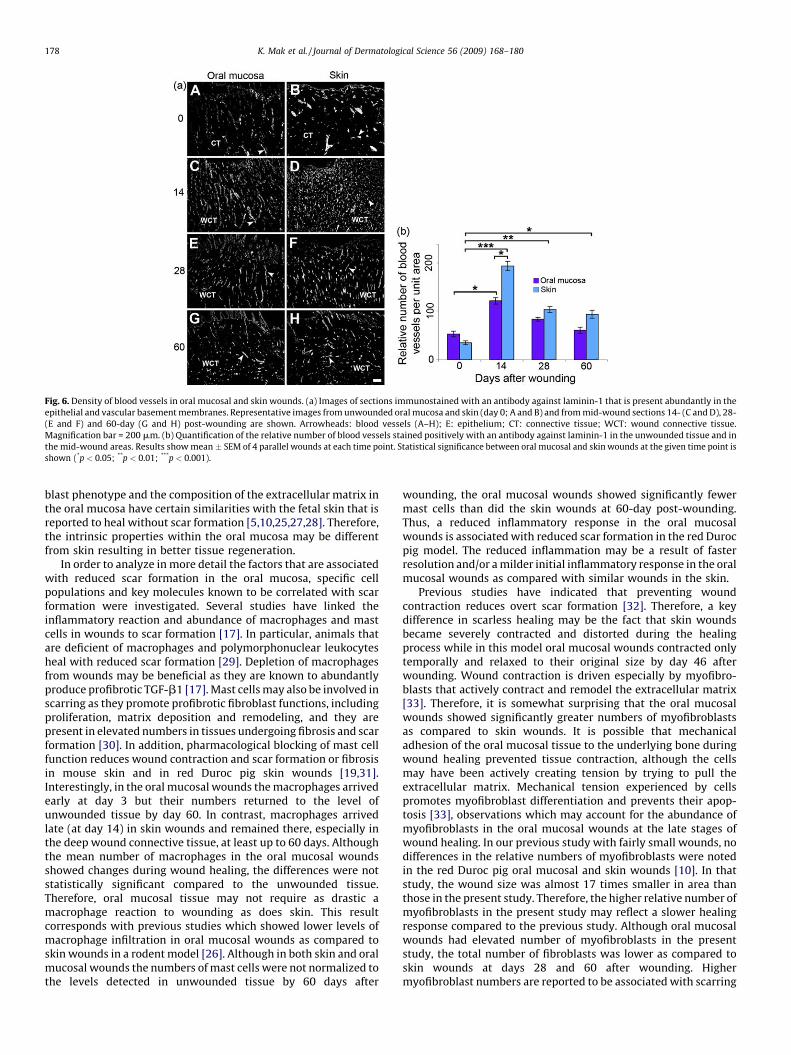
Fig. 6. Density of blood vessels in oral mucosal and skin wounds. (a) Images of sections immunostained with an antibody against laminin-1 that is present abundantly in the
epithelial and vascular basement membranes. Representative images from unwounded oral mucosa and skin (day 0; A and B) and from mid-wound sections 14- (C and D), 28-
(E and F) and 60-day (G and H) post-wounding are shown. Arrowheads: blood vessels (A–H); E: epithelium; CT: connective tissue; WCT: wound connective tissue.
Magnification bar = 200 mm. (b) Quantification of the relative number of blood vessels stained positively with an antibody against laminin-1 in the unwounded tissue and in
the mid-wound areas. Results show mean � SEM of 4 parallel wounds at each time point. Statistical significance between oral mucosal and skin wounds at the given time point is
shown (*p < 0.05; **p < 0.01; ***p < 0.001).
K. Mak et al. / Journal of Dermatological Science 56 (2009) 168–180178
blast phenotype and the composition of the extracellular matrix inthe oral mucosa have certain similarities with the fetal skin that isreported to heal without scar formation [5,10,25,27,28]. Therefore,the intrinsic properties within the oral mucosa may be differentfrom skin resulting in better tissue regeneration.
In order to analyze in more detail the factors that are associatedwith reduced scar formation in the oral mucosa, specific cellpopulations and key molecules known to be correlated with scarformation were investigated. Several studies have linked theinflammatory reaction and abundance of macrophages and mastcells in wounds to scar formation [17]. In particular, animals thatare deficient of macrophages and polymorphonuclear leukocytesheal with reduced scar formation [29]. Depletion of macrophagesfrom wounds may be beneficial as they are known to abundantlyproduce profibrotic TGF-b1 [17]. Mast cells may also be involved inscarring as they promote profibrotic fibroblast functions, includingproliferation, matrix deposition and remodeling, and they arepresent in elevated numbers in tissues undergoing fibrosis and scarformation [30]. In addition, pharmacological blocking of mast cellfunction reduces wound contraction and scar formation or fibrosisin mouse skin and in red Duroc pig skin wounds [19,31].Interestingly, in the oral mucosal wounds the macrophages arrivedearly at day 3 but their numbers returned to the level ofunwounded tissue by day 60. In contrast, macrophages arrivedlate (at day 14) in skin wounds and remained there, especially inthe deep wound connective tissue, at least up to 60 days. Althoughthe mean number of macrophages in the oral mucosal woundsshowed changes during wound healing, the differences were notstatistically significant compared to the unwounded tissue.Therefore, oral mucosal tissue may not require as drastic amacrophage reaction to wounding as does skin. This resultcorresponds with previous studies which showed lower levels ofmacrophage infiltration in oral mucosal wounds as compared toskin wounds in a rodent model [26]. Although in both skin and oralmucosal wounds the numbers of mast cells were not normalized tothe levels detected in unwounded tissue by 60 days after
wounding, the oral mucosal wounds showed significantly fewermast cells than did the skin wounds at 60-day post-wounding.Thus, a reduced inflammatory response in the oral mucosalwounds is associated with reduced scar formation in the red Durocpig model. The reduced inflammation may be a result of fasterresolution and/or a milder initial inflammatory response in the oralmucosal wounds as compared with similar wounds in the skin.
Previous studies have indicated that preventing woundcontraction reduces overt scar formation [32]. Therefore, a keydifference in scarless healing may be the fact that skin woundsbecame severely contracted and distorted during the healingprocess while in this model oral mucosal wounds contracted onlytemporally and relaxed to their original size by day 46 afterwounding. Wound contraction is driven especially by myofibro-blasts that actively contract and remodel the extracellular matrix[33]. Therefore, it is somewhat surprising that the oral mucosalwounds showed significantly greater numbers of myofibroblastsas compared to skin wounds. It is possible that mechanicaladhesion of the oral mucosal tissue to the underlying bone duringwound healing prevented tissue contraction, although the cellsmay have been actively creating tension by trying to pull theextracellular matrix. Mechanical tension experienced by cellspromotes myofibroblast differentiation and prevents their apop-tosis [33], observations which may account for the abundance ofmyofibroblasts in the oral mucosal wounds at the late stages ofwound healing. In our previous study with fairly small wounds, nodifferences in the relative numbers of myofibroblasts were notedin the red Duroc pig oral mucosal and skin wounds [10]. In thatstudy, the wound size was almost 17 times smaller in area thanthose in the present study. Therefore, the higher relative number ofmyofibroblasts in the present study may reflect a slower healingresponse compared to the previous study. Although oral mucosalwounds had elevated number of myofibroblasts in the presentstudy, the total number of fibroblasts was lower as compared toskin wounds at days 28 and 60 after wounding. Highermyofibroblast numbers are reported to be associated with scarring
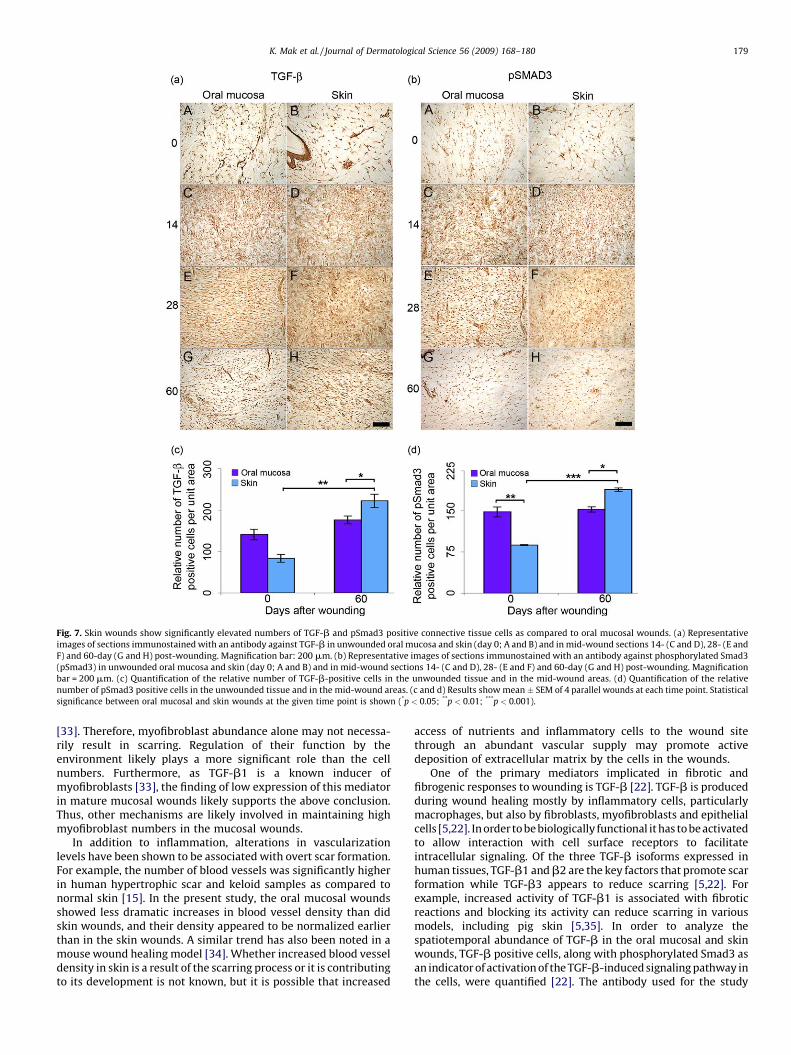
Fig. 7. Skin wounds show significantly elevated numbers of TGF-b and pSmad3 positive connective tissue cells as compared to oral mucosal wounds. (a) Representative
images of sections immunostained with an antibody against TGF-b in unwounded oral mucosa and skin (day 0; A and B) and in mid-wound sections 14- (C and D), 28- (E and
F) and 60-day (G and H) post-wounding. Magnification bar: 200 mm. (b) Representative images of sections immunostained with an antibody against phosphorylated Smad3
(pSmad3) in unwounded oral mucosa and skin (day 0; A and B) and in mid-wound sections 14- (C and D), 28- (E and F) and 60-day (G and H) post-wounding. Magnification
bar = 200 mm. (c) Quantification of the relative number of TGF-b-positive cells in the unwounded tissue and in the mid-wound areas. (d) Quantification of the relative
number of pSmad3 positive cells in the unwounded tissue and in the mid-wound areas. (c and d) Results show mean � SEM of 4 parallel wounds at each time point. Statistical
significance between oral mucosal and skin wounds at the given time point is shown (*p < 0.05; **p < 0.01; ***p < 0.001).
K. Mak et al. / Journal of Dermatological Science 56 (2009) 168–180 179
[33]. Therefore, myofibroblast abundance alone may not necessa-rily result in scarring. Regulation of their function by theenvironment likely plays a more significant role than the cellnumbers. Furthermore, as TGF-b1 is a known inducer ofmyofibroblasts [33], the finding of low expression of this mediatorin mature mucosal wounds likely supports the above conclusion.Thus, other mechanisms are likely involved in maintaining highmyofibroblast numbers in the mucosal wounds.
In addition to inflammation, alterations in vascularizationlevels have been shown to be associated with overt scar formation.For example, the number of blood vessels was significantly higherin human hypertrophic scar and keloid samples as compared tonormal skin [15]. In the present study, the oral mucosal woundsshowed less dramatic increases in blood vessel density than didskin wounds, and their density appeared to be normalized earlierthan in the skin wounds. A similar trend has also been noted in amouse wound healing model [34]. Whether increased blood vesseldensity in skin is a result of the scarring process or it is contributingto its development is not known, but it is possible that increased
access of nutrients and inflammatory cells to the wound sitethrough an abundant vascular supply may promote activedeposition of extracellular matrix by the cells in the wounds.
One of the primary mediators implicated in fibrotic andfibrogenic responses to wounding is TGF-b [22]. TGF-b is producedduring wound healing mostly by inflammatory cells, particularlymacrophages, but also by fibroblasts, myofibroblasts and epithelialcells [5,22]. In order to be biologically functional it has to be activatedto allow interaction with cell surface receptors to facilitateintracellular signaling. Of the three TGF-b isoforms expressed inhuman tissues, TGF-b1 and b2 are the key factors that promote scarformation while TGF-b3 appears to reduce scarring [5,22]. Forexample, increased activity of TGF-b1 is associated with fibroticreactions and blocking its activity can reduce scarring in variousmodels, including pig skin [5,35]. In order to analyze thespatiotemporal abundance of TGF-b in the oral mucosal and skinwounds, TGF-b positive cells, along with phosphorylated Smad3 asan indicator of activation of the TGF-b-induced signaling pathway inthe cells, were quantified [22]. The antibody used for the study

K. Mak et al. / Journal of Dermatological Science 56 (2009) 168–180180
recognized the TGF-b1, -b2 and -b3 isoforms and does not,therefore, allow distinction of the relative abundance of theindividual isoforms. Wound healing responses led to a strongincrease in both TGF-b and pSmad3 positive cells in both oralmucosal and skin wounds. However, in oral mucosal wounds, thisresponse was only temporary while a significantly elevated numberof positive cells persisted in skin wounds until at least 60 days.Therefore, the cellular and molecular mechanisms that regulateTGF-b abundance and signaling are differentially regulated in oralmucosal as compared to skin wounds, and such regulation maycontribute to reduced scar formation in the oral mucosa.
Taken together, wound healing in the oral mucosa of red Durocpigs resulted in significantly reduced clinical and histological scarformation and wound contraction, and these outcomes are linkedwith earlier resolution of the inflammatory response, reduceddensity of blood vessel and TGF-b and pSmad3 positive cells in thewounds. However, the number of myofibroblasts was uniquely andpersistently elevated in the oral mucosal wounds. These findingssuggest that scar formation is a complex process in either of thetwo sites investigated, and likely depends not only on the numberof myofibroblasts, but also on the environment which regulatestheir function. Thus, the red Duroc pig model can be used to furtherelucidate the cellular and molecular pathways that lead to reducedscar formation and the promotion of functional wound healing.
Acknowledgements
Supported by the CIHR Institute of Musculoskeletal Health andArthritis (LH), the CIHR Institute for Gender and Health, the CalgaryFoundation-Grace Glaum Professorship (DAH) and the Network ofOral Research Training and Health (KM, AM).
References
[1] Durani P, Occleston N, O’Kane S, Ferguson MW. Avotermin: a novel antiscar-ring agent. Int J Low Extrem Wounds 2008;7:160–8.
[2] Occleston NL, O’Kane S, Goldspink N, Ferguson MW. New therapeutics for theprevention and reduction of scarring. Drug Discov Today 2008;13:973–81.
[3] Tanaka E, Galliot B. Triggering the regeneration and tissue repair programs.Development 2009;136:349–53.
[4] Bullard KM, Longaker MT, Lorenz HP. Fetal wound healing: current biology.World J Surg 2003;27:54–61.
[5] Ferguson MW, O’Kane S. Scar-free healing: from embryonic mechanisms toadult therapeutic intervention. Philos Trans R Soc Lond B Biol Sci 2004;359:839–50.
[6] Greenhalgh DG. Consequences of excessive scar formation: dealing with theproblem and aiming for the future. Wound Repair Regen 2007;15(Suppl 1):S2–5.
[7] Ramos ML, Gragnani A, Ferreira LM. Is there an ideal animal model to studyhypertrophic scarring? J Burn Care Res 2008;29:363–8.
[8] Zhu KQ, Carrougher GJ, Gibran NS, Isik FF, Engrav LH. Review of the femaleDuroc/Yorkshire pig model of human fibroproliferative scarring. WoundRepair Regen 2007;15(Suppl 1):S32–9.
[9] de Hemptinne I, Gallant-Behm CL, Noack CL, Parreno J, Hart DA. Dermalfibroblasts from red Duroc and Yorkshire pigs exhibit intrinsic differencesin the contraction of collagen gels. Wound Repair Regen 2008;16:132–42.
[10] Wong JW, Gallant-Behm C, Wiebe C, Mak K, Hart DA, Larjava H, et al. Woundhealing in oral mucosa results in reduced scar formation as compared to skin:
evidence from the red Duroc pig model and humans. Wound Repair Regen2009;17:717–29.
[11] Cass DL, Bullard KM, Sylvester KG, Yang EY, Longaker MT, Adzick NS. Woundsize and gestational age modulate scar formation in fetal wound repair. JPediatr Surg 1997;32:411–5.
[12] Harunari N, Zhu KQ, Armendariz RT, Deubner H, Muangman P, Carrougher GJ,et al. Histology of the thick scar on the female, red Duroc pig: final similaritiesto human hypertrophic scar. Burns 2006;32:669–77.
[13] Dunkin CS, Pleat JM, Gillespie PH, Tyler MP, Roberts AH, McGrouther DA.Scarring occurs at a critical depth of skin injury: precise measurement in agraduated dermal scratch in human volunteers. Plast Reconstr Sur 2007;119:1722–32.
[14] Gallant-Behm CL. The molecular biology and genetics of skin wound healing inthe red Duroc pig. PhD Thesis, University of Calgary; 2007.
[15] Amadeu T, Braune A, Mandarim-de-Lacerda C, Porto LC, Desmouliere A, CostaA. Vascularization pattern in hypertrophic scars and keloids: a stereologicalanalysis. Pathol Res Pract 2003;199:469–73.
[16] Darby IA, Hewitson TD. Fibroblast differentiation in wound healing andfibrosis. Int Rev Cytol 2007;257:143–79.
[17] Wilgus TA. Immune cells in the healing skin wound: influential players at eachstage of repair. Pharmacol Res 2008;58:112–6.
[18] Wang JF, Olson ME, Reno CR, Wright JB, Hart DA. The pig as a model forexcisional skin wound healing: characterization of the molecular and cellularbiology, and bacteriology of the healing process. Comp Med 2001;51:341–8.
[19] Gallant-Behm CL, Hildebrand KA, Hart DA. The mast cell stabilizer ketotifenprevents development of excessive skin wound contraction and fibrosis in redDuroc pigs. Wound Repair Regen 2008;16:226–33.
[20] Beausang E, Floyd H, Dunn KW, Orton CI, Ferguson MW. A new quantitativescale for clinical scar assessment. Plast Reconstr Surg 1998;102:1954–61.
[21] Schrementi ME, Ferreira AM, Zender C, DiPietro LA. Site-specific production ofTGF-beta in oral mucosal and cutaneous wounds. Wound Repair Regen2008;16:80–6.
[22] Verrecchia F, Mauviel A. Transforming growth factor-beta and fibrosis. World JGastroenterol 2007;13:3056–62.
[23] Cubison TC, Pape SA, Parkhouse N. Evidence for the link between healing timeand the development of hypertrophic scars (HTS) in paediatric burns due toscald injury. Burns 2006;32:992–9.
[24] Wang J, Dodd C, Shankowsky HA, Scott PG, Tredget EE. Deep dermal fibroblastscontribute to hypertrophic scarring. Lab Invest 2008;88:1278–90.
[25] Hakkinen L, Uitto VJ, Larjava H. Cell biology of gingival wound healing.Periodontology 2000;24:127–52.
[26] Szpaderska AM, Zuckerman JD, DiPietro LA. Differential injury responses inoral mucosal and cutaneous wounds. J Dent Res 2003;82:621–6.
[27] Schor SL, Ellis I, Irwin CR, Banyard J, Seneviratne K, Dolman C, et al. Sub-populations of fetal-like gingival fibroblasts: characterisation and potentialsignificance for wound healing and the progression of periodontal disease.Oral Dis 1996;2:155–66.
[28] van Beurden HE, Von den Hoff JW, Torensma R, Maltha JC, Kuijpers-JagtmanAM. Myofibroblasts in palatal wound healing: prospects for the reduction ofwound contraction after cleft palate repair. J Dent Res 2005;84:871–80.
[29] Martin P, D’Souza D, Martin J, Grose R, Cooper L, Maki R, et al. Wound healing inthe PU.1 null mouse–tissue repair is not dependent on inflammatory cells.Curr Biol 2003;13:1122–8.
[30] Navi D, Saegusa J, Liu FT. Mast cells and immunological skin diseases. Clin RevAllergy Immunol 2007;33:144–55.
[31] Levi-Schaffer F, Segal V, Barak V, Rubinchik E, Nagler A. Regulation of thefunctional activity of mast cells and fibroblasts by mononuclear cells inmurine and human chronic graft-vs.-host disease. Exp Hematol 1997;25:238–45.
[32] Yannas IV, Kwan MD, Longaker MT. Early fetal healing as a model for adultorgan regeneration. Tissue Eng 2007;13:1789–98.
[33] Hinz B. Formation and function of the myofibroblast during tissue repair. JInvest Dermatol 2007;127:526–37.
[34] Szpaderska AM, Walsh CG, Steinberg MJ, DiPietro LA. Distinct patterns ofangiogenesis in oral and skin wounds. J Dent Res 2005;84:309–14.
[35] Singer AJ, Huang SS, Huang JS, McClain SA, Romanov A, Rooney J, et al. A novelTGF-beta antagonist speeds reepithelialization and reduces scarring of partialthickness porcine burns. J Burn Care Res 2009;30:329–34.