role of igfbps in the morphogenesis of lingual papillae
TRANSCRIPT

Role of IGFBPs in the Morphogenesisof Lingual Papillae
YUKO SUZUKI*Department of Oral Anatomy, School of Dentistry, Health Sciences University of
Hokkaido, Ishikari-Tobetsu, Japan
ABSTRACTThe expression of insulin-like growth factor binding proteins (IGFBPs)
during the morphogenesis of lingual papillae of mice was examined by insitu hybridization. Among seven mouse IGFBPs, IGFBP-1, -6, and -7 mR-NAs were not expressed in the tongue tissue. At E12, though no papillaehave formed yet, IGFBP-2, -4, and -5 were expressed in the entire tongueepithelium. At E14, fungiform papillae appeared in the anterior region andcircumvallate papillae were distinguished in the posterior region. Strongexpression of IGFBP-5 was observed in the apical region of both fungiformand circumvallate papillae. At this stage, the epithelial elevation of filiformpapillae was not clear; but IGFBP-5 was expressed in the apex. At E15,foliate papillae were distinguished and IGFBP-5 was expressed in thedorsal epithelium of ridges. In filiform papillae, IGFBP-3 was expressed inthe core of the connective tissue. At E17, the expression of IGFBP-5 disap-peared from the apical region of fungiform, filiform, foliate, and circumval-late papillae, whereas that of IGFBP-2 remained. This finding suggests thatIGFBP-5 and -2 function to cause evagination of the epithelium into a raisedstructure. In the epithelium of trenches of foliate and circumvallate papil-lae, strong expression of IGFBP-4 was observed at E15 and E17. As previ-ously suggested from a study on postnatal mice (Suzuki et al. J CompNeurol 2005;482:74–84), IGFBP-4 acts in the epithelial invagination toform the trenches, grooves, or furrows of lingual papillae during develop-ment. © 2005 Wiley-Liss, Inc.
Key words: insulin-like growth factor binding protein; in situhybridization; fungiform papillae; circumvallatepapillae; filiform papillae
The dorsal surface of the mammalian tongue is coveredwith four kinds of papillae: fungiform, circumvallate, foli-ate, and filiform. These papillae are distributed in a spe-cific pattern over the tongue in mice: fungiform and fili-form papillae are located on the anterior two-thirds of thetongue. The fungiform papillae have a mushroom-shapedstructure, composed of a multilayered epithelium andmesenchymal core (Farbman and Mbiene, 1991). Filiformpapillae are small conical surface projections found ingreat numbers among the fungiform papillae (Baratz andFarbman, 1975). Foliate papillae are located on the pos-terior lateral margins of the tongue, and circumvallatepapillae on the posterior midline. Foliate papillae consistof several ridges that alternate with deep grooves in themucosa. In mice and rats, single circumvallate papilla ispresent in the middle part of the terminal sulcus and it issurrounded by a deep circular groove into which open the
von Ebner’s glands. Except for the filiform papillae, thesepapillae contain taste buds (Nosrat et al., 1997).
Grant sponsor: Academic Frontier Project for Private Univer-sities; Grant sponsor: mathcing fund pubsidy from MEXT (Min-istry of Education, Culture, Sports, Science, and Technology),2002-2006.
*Correspondence to: Yuko Suzuki, Department of Oral Anat-omy, School of Dentistry, Health Sciences University of Hok-kaido, Ishikari-Tobetsu 061-0293, Japan. Fax: 81-1332-3-1236.E-mail: [email protected]
Received 21 February 2005; Accepted 11 April 2005DOI 10.1002/ar.a.20219Published online 27 July 2005 in Wiley InterScience(www.interscience.wiley.com).
THE ANATOMICAL RECORD PART A 286A:841–847 (2005)
© 2005 WILEY-LISS, INC.

Several signaling molecules regulate the morphogenesisof the papillae. For example, sonic hedgehog (Shh) is ex-pressed only in the dorsal surface of fungiform papillaeduring embryonic days 13–18 (Hall et al., 1999, 2003;Jung et al., 2004), and its signal disruption alters thenumber and location of fungiform papillae (Hall et al.,2003; Liu et al., 2004). In pax 9-deficient mice, a cornifiedlayer is absent, thus filiform papillae lack anterior-poste-rior polarity (Jonker et al., 2004). In mice knocked out forbrain-derived neurotrophic factor (BDNF) and its receptorTrkB, fungiform, foliate, and circumvallate papillae aresmaller or have an aberrant morphology or both (Nosrat etal., 1997; Oakley et al., 1998; Mistretta et al., 1999).
In our previous study, insulin-like growth factor bindingprotein (IGFBP)-4, a member of the family of insulin-likegrowth factor (IGF) and related molecules, was expressedin the bottom of the trenches of circumvallate papillae ofpostnatal day 2 mice, suggesting that it regulates thedown-growth of grooves (Suzuki et al., 2005). The IGFfamily comprises IGF-I, IGF-II, their receptors (IGF-IR,IGF-IIR), and seven high-affinity IGFBPs. The actions ofthese IGFs appear to be regulated and coordinated by afamily of IGFBPs. The IGFBPs are thought to have fourmajor functions that are essential to the regulation andcoordination of the biological activities of IGFs. That is,IGFBPs are considered to act as transport proteins inplasma and to control the efflux of IGFs from the vascularspace; to prevent IGFs from being degraded and to prolongthe half-lives of IGFs; to provide a means of tissue and celltype-specific localization; and to modulate directly the in-teraction of the IGFs with their receptors and therebyindirectly control their biological actions (Duan, 2002).Recent evidence suggests that the IGFBPs can also havedirect IGF-independent actions on cellular functions(Zhou et al., 2003). All lingual papillae begin as epithelialthickenings and then evaginate to form raised papillaewith a mesenchymal core during embryonic development(cf. Farbman and Mbiene, 1991). Because IGF-I and -IIare known to be expressed in the embryonic tongue in rats(Ayer-LeLievre et al., 1991) and in mice (Ferguson et al.,1992; Yamane et al., 2000), the IGF family has been sug-gested to regulate the morphogenesis of the lingual papil-lae. The aim of this article was to examine the expressionof IGFBPs during embryonic development of lingual pa-pillae.
MATERIALS AND METHODSAnimals
Timed pregnant ddY mice, were obtained from SankyoLaboratories (Tokyo, Japan). They were maintained in aheat- and humidity-controlled vivarium with food and wa-ter provided ad libitum. Experimental protocols concern-ing animal handling were reviewed and approved by theInstitutional Animal Care Committee of the Health Sci-ences University of Hokkaido.
Tissue PreparationThe day of appearance of vaginal plug was designated
as E0. The study was conducted on 12 pregnant femalemice. To obtain embryos, mice were killed by cervicaldislocation and their uteri with fetuses (E12–17) carefullydissected out. For in situ hybridization, the tongues werefixed with 4% paraformaldehyde in 0.1 M phosphatebuffer (pH 7.4) overnight at 4°C. For immunohistochem-
istry, the tongues were fixed with periodate-lysine-para-formaldehyde (PLP) solution. Each specimen was washedin phosphate-buffered saline (PBS) solution, cryoprotectedwith 25% sucrose, and embedded in OCT compound(Oken, Tokyo, Japan). The tissues were sectioned coro-nally at a thickness of 8–10 �m. Sections were collectedand placed on silane-coated slides.
RNA Probes and In Situ HybridizationcDNA fragments of IGF-IR, IGFBP-2, IGFBP-3, IG-
FBP-4, IGFBP-5, IGFBP-6, and IGFBP-7 were generatedby reverse transcription-polymerase chain reaction (RT-PCR) using total RNA extracted from the tongue and thenused for the synthesis of cRNA probes. The sequences ofthe primers were as follows: 5�-TCTCTCTCTGGCCGAC-GAGT-3� and 5�-GAGCAGAAGTCACCGAATCG-3� (977bp; AF056187) for IGF-IR; 5�-GACGCTACGCTGCTATC-CCA-3� and 5�-GTCTCCTGCTGCTCGTTGTA-3� (614 bp;NM008342) for IGFBP-2; 5�-GGAAACATCAGTGAGTC-CGA-3� and 5�-GCTGAGGCAATGTACGTCGT-3� (458 bp;X8158.1) for IGFBP-3; 5�-GGAGAAGCCCCTGCGTA-CAT-3� and 5�-ACCCCTGTCTTCCGATCCAC-3� (434 bp;X76066) for IGFBP-4; 5�-AGTAACGTTGAGT-GACGCGT-3� and 5�-CAGTGTTGGGGGTGCGTACT-3�(750 bp; L12447) for IGFBP-5; 5�-TAATGCTGTTGT-TCGCTGCG-3� and 5�-CACTGCTGCTTGCGGTAGAA-3�(552 bp; NM008344) for IGFBP-6; and 5�-AAGGTCCTTC-CATAGTGACG-3� and 5�-CAGGGTTATAGCTGTCG-GCT-3� (439 bp; NM008048) for IGFBP-7. The PCR wascarried out for 35 cycles. Each resulting fragment wascloned into HindIII/EcoRI sites of pT7/T3 DH5�-FT (In-vitrogen, Tokyo, Japan) and sequenced. Digoxigenin(DIG)-labeled antisense and sense probes were producedby use of an RNA transcription kit (Roche Diagnostics,Mannheim, Germany). Sections were washed in PBS andtreated for 20 min with 0.2 N HCl and for 15–20 min withproteinase K (1 �g/ml in PBS; Takara, Kyoto, Japan) at37°C. Next, the sections were washed in PBS and refixedwith 4% paraformaldehyde in 0.1 M phosphate buffer for20 min. After having been washed twice in PBS, the sec-tions were air-dried and hybridized. Hybridization wasperformed at 47°C for 16 hr with an RNA probe in ahybridization solution containing 50% formamide, 0.3 MNaCl, 0.02 M Tris-HCl, 1 mM EDTA, 10% dextran sulfate,1 � Denhardt’s solution, 1 mg/ml yeast tRNA, and 0.02%SDS. Hybridized sections were washed at 47°C in a solu-tion containing 50% formamide and 2 � SSC for 1 hr, andthereafter twice in 2 � SSC for 5 min each time. Then theywere treated with 20 �g/ml of RNase (TypeII-A; Sigma) at37°C for 30 min and washed at 47°C in 50% formamide/2XSSC followed by 50% formamide/1 � SSC for 1 hr foreach. After having been washed three times in PBS, thesections were incubated with 1% blocking reagent(Boeringer Mannheim, Mannheim, Germany) in maleicacid buffer (pH 7.5) for 1 hr at room temperature. Subse-quently, they were incubated overnight at 4°C with alka-line phosphatase-conjugated anti-DIG Fab fragments di-luted 1:500 in PBS. After three washes in PBS,chromogenic reactions were carried out by using NBT/BCIP (Boeringer).
ImmunohistochemistryThe sections were incubated in a blocking solution
(Dako Protein Block Serum-Free; Dako, Carpinteria, CA)
842 SUZUKI

for 10 min at room temperature. Then the sections wereincubated overnight at 4°C with antirabbit PGP9.5 anti-body (Ultraclone, Isle of Wight, U.K.), diluted 1:100 inPBS. After having been washed in PBS, the sections weretreated with Alexa Fluor 594-conjugated donkey antirab-bit IgG (Molecular Probes, Eugene, OR) for 2 hr at roomtemperature. Sections were viewed with a Leica confocallaser scanning microscope.
RESULTSAmong IGFBPs, it was reported that IGFBP-1 mRNA
was detected only in adult liver tissue (Suzuki et al., 2005)and so it was not examined in the present study. IGFBP-6and -7 mRNAs were not expressed in tongue tissue by insitu hybridization in the present study. Immunohisto-chemically IGFBP-6 was detected in the nerve fibers in-nervating the taste buds of circumvallate papillae (Suzukiet al., 2005). IGFBP-7 was specifically expressed in devel-oping lens (not shown). Therefore, IGFBP-2, -3, -4, and -5mRNAs were detected in the developing tongue.
At embryonic day (E) 12, the early fetal tongue has arelatively homogeneous epithelium with no obvious pap-illary structure. Among IGFBPs, IGFBP-2 was expressedin the entire lingual epithelium (Fig. 1A); IGFBP-5 wasexpressed in the lingual epithelium and in myoblast cells(Fig. 1B). IGFBP-4 was expressed in the entire lingualepithelium and mesenchyme (not shown). IGFBP-3 wasnot detected in the tongue epithelium but was found insome mesenchymal cells (Fig. 1C).
Fungiform and Filiform PapillaeAt E14, in the anterior region of tongue, the developing
fungiform papillae were small and slightly elevated intothe oral cavity. They also contained a small amount ofconnective tissue in their medullary core. IGFBP-2 and -4were expressed in the epithelium of both fungiform papil-lae and nonpapilla tongue tissue. IGFBP-4 was expressedalso in the mesenchyme (Fig. 1D). IGF-IR expression wasobserved in the tongue epithelium throughout the embry-onic stages (Fig. 1E). IGFBP-5 was intensely expressed inthe fungiform papillae (Fig. 1F). Although the surface ofthe tongue remained flat, IGFBP-5 was expressed also inthe presumed filiform papillae, which were interspersedwith fungiform papillae (Fig. 1F). At E15, regularlyspaced dermal papillae appeared in the presumptive fili-form papillae. Expression of IGFBP-5 was observed in theapical to middle region of the filiform epithelium (Fig. 1G),and IGFBP-3 was expressed in mesenchymal core (Fig.1H). At E17, the filiform papillae start to form a cornifiedlayer in the epithelium, which exhibit anterior-posteriorpolarity. IGFBP-3 was expressed in the core of these pa-pillae (Fig. 1I). In fungiform papillae, expression of IG-FBP-3 was weak (Fig. 1I, inset). IGFBP-2 was expressedin the basal layer of the epithelium in the anterior regionof each filiform papilla (Fig. 1J) and in the apex of thefungiform papillae (Fig. 1K). The expression of IGFBP-4and IGFBP-5 had mostly disappeared from the anteriortongue epithelium by E17 (Fig. 1L and M).
Circumvallate PapillaeAt E14 in the posterior region of the tongue, circumval-
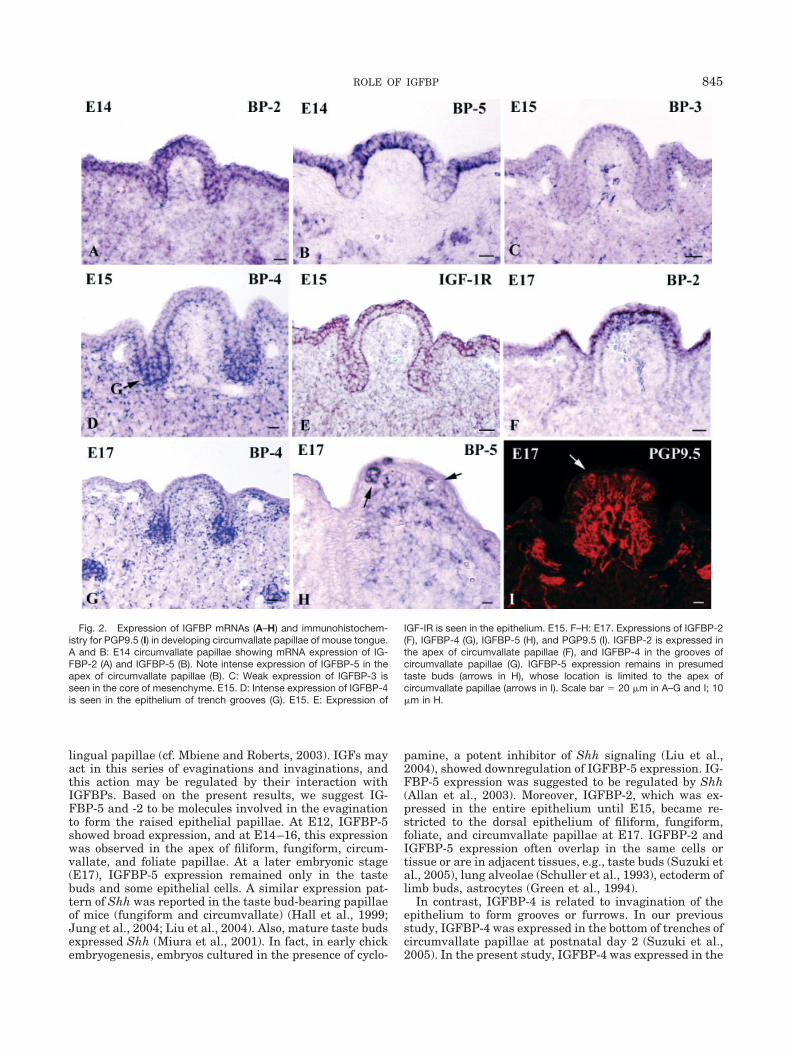
late papillae were observed to have epithelial elevationsand shallow grooves. IGFBP-2 was expressed in the entireepithelium of these papillae (Fig. 2A). IGFBP-5 was ex-
pressed in the elevated epithelium of papillae but not inthe grooves (Fig. 2B). At E15, IGFBP-3 was expressed in afew cells of mesenchymal core (Fig. 2C). IGFBP-4 wasintensely expressed in the bottom of the grooves (Fig. 2D).IGF-IR expression was observed in the epitheliumthroughout the embryonic stages (Fig. 2E). At E17, thecircumvallate papillae were more developed. The trenchgrooves had deepened further into the underlying mesen-chyme, and IGFBP-2 expression had become restricted tothe elevated epithelium of the papillae (Fig. 2F). Intenseexpression of IGFBP-4 was still observed in the epithe-lium of the bottom of the trenches, and IGFBP-4 wasexpressed also in minor salivary glands, von Ebner’sglands (Fig. 2G). IGFBP-5 expression had mostly disap-peared from the epithelium of the circumvallate papillae,but it was expressed in presumptive taste buds and someepithelial cells at the boundary with the connective tissue(Fig. 2H). At this stage, numerous PGP9.5-positive nervefibers entered the circumvallate papillae, and a few tastebuds were located at the top of the papillae (Fig. 2I).
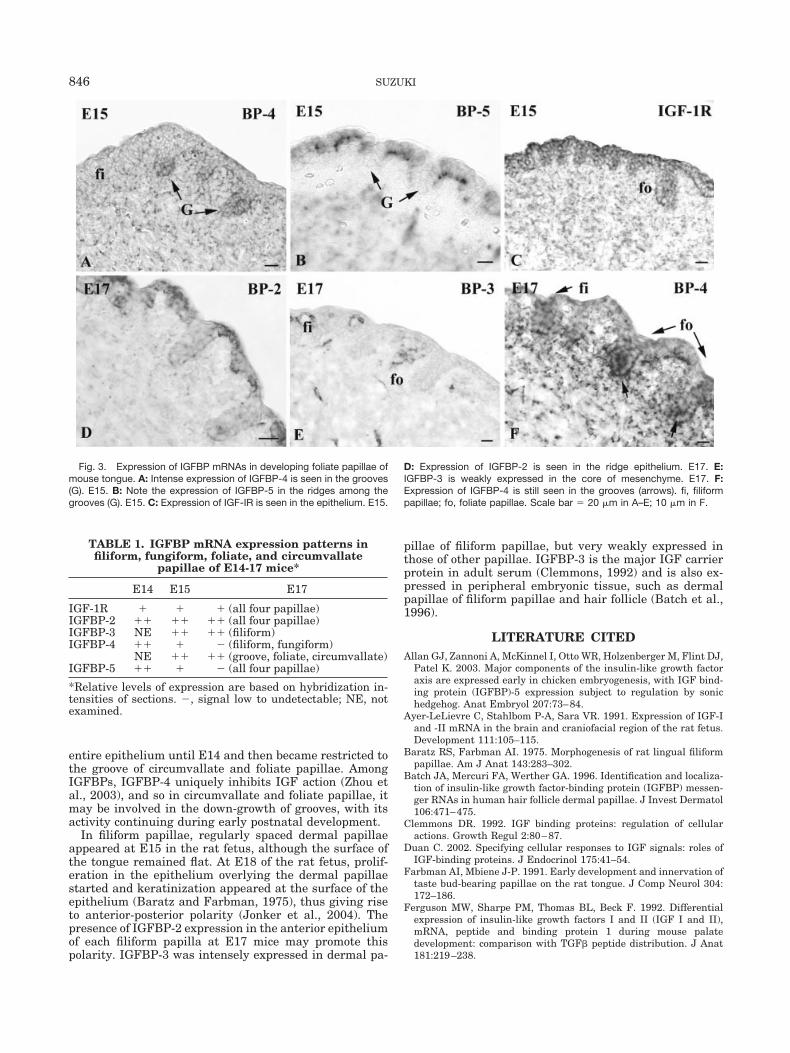
Foliate PapillaeAt E15, foliate papillae were observed in the margin of
the posterior region of tongue. At this stage, IGFBP-4 wasintensely expressed in the shallow grooves among theridges (Fig. 3A). IGFBP-5 was expressed in the dorsalepithelium of papillae but not in the grooves (Fig. 3B).IGF-IR expression was observed in the epitheliumthroughout the embryonic stages (Fig. 3C). The expressionof IGFBP-2 was observed in the entire epithelium of pa-pillae at E15 and had restricted to the dorsal epithelium ofthe papillae at E17 (Fig. 3D). At E17, IGFBP-3 wasweakly expressed in the mesenchyme (Fig. 3E). At thisstage, intense expression of IGFBP-4 was still observed inthe bottom of grooves (Fig. 3F). These results were sum-marized in Table 1.
DISCUSSIONIn the embryonic tongue of mice, IGF-I and -II were
earlier found by immunohistochemical means to be ex-pressed in the dorsal epithelium and muscles (Yamane etal., 2000). By in situ hybridization, high levels of IGF-IImRNA were found in the mesenchyme, muscles, and con-nective tissue of the embryonic tongues of mice (Fergusonet al., 1992) and rats (Ayer-LeLievre et al., 1991). Theexpression of IGF-I mRNA in the embryonic rat tonguewas weak, and thus its localization was not clear (Ayer-LeLievre et al., 1991). IGF-1 and -II bind to the receptorIGF-IR. IGF-IIR has no known growth-mediating effectsand may simply act as a cell-surface depot for storage ofligand (cf. Zhou et al., 2003). In the present study, IGF-IRmRNA was expressed weakly in lingual epithelium, in-cluding the papillae. Therefore, IGFs may function tocause the morphogenesis of the lingual papillae throughIGF-IR, acting in an autocrine or paracrine manner. How-ever, the expression of these molecules was not restrictedto the lingual papillae. The present study revealed specificexpression patterns of IGFBP-2, -3, -4, and -5 in the lin-gual papillae at E14–17. Morphogenesis of lingual papil-lae includes a series of evaginations and invaginations,which are coordinated interactions between the epithe-lium and underlying connective tissue. In fact, proliferat-ing cells were observed in the apex of epithelium, trenchgrooves, and core of connective tissue of the developing
843ROLE OF IGFBP

Fig. 1. Expression of IGFBP mRNAs in developing fungiform andfiliform papillae of the mouse tongue detected by in situ hybridizationwith RNA probes. A–C: E12 tongue showing IGFBP-2 (A), IGFBP-5 (B),and IGFBP-3 (C) expressions. D–F: E14 tongue showing IGFBP-4 (D),IGF-IR (E), and IGFBP-5 (F) expressions. Expressions of IGFBP-4 andIGF-IR are seen in the epithelium and mesenchyme including fungiformpapillae (fu in D and E). Note the intense expression of IGFBP-5 in theapex of fungiform (fu) and filiform (fi) papillae (F). G and H: E15 tongueshowing mRNA expression of IGFBP-5 (G) and IGFBP-3 (H). IGFBP-3 is
expressed in the mesenchymal core of presumptive filiform papillae (H).I–M: E17 tongue showing IGFBP-3 (I), IGFBP-2 (J, K), IGFBP-4 (L), andIGFBP-5 m RNA (M) expression. Cornified surface of each filiform papilla(fi) is seen (I). IGFBP-3 is expressed in the mesenchymal core of filiformpapillae (I), but very weak in fungiform papillae (I, inset). IGFBP-2 isexpressed in the basal layer of the epithelium of filiform papillae (J) andat the apex of fungiform papillae (K). Expression of IGFBP-4 is absent inthe epithelium (L). Expression of IGFBP-5 mostly disappears from fili-form and fungiform papillae (M). Scale bar � 20 �m.
844 SUZUKI
lingual papillae (cf. Mbiene and Roberts, 2003). IGFs mayact in this series of evaginations and invaginations, andthis action may be regulated by their interaction withIGFBPs. Based on the present results, we suggest IG-FBP-5 and -2 to be molecules involved in the evaginationto form the raised epithelial papillae. At E12, IGFBP-5showed broad expression, and at E14–16, this expressionwas observed in the apex of filiform, fungiform, circum-vallate, and foliate papillae. At a later embryonic stage(E17), IGFBP-5 expression remained only in the tastebuds and some epithelial cells. A similar expression pat-tern of Shh was reported in the taste bud-bearing papillaeof mice (fungiform and circumvallate) (Hall et al., 1999;Jung et al., 2004; Liu et al., 2004). Also, mature taste budsexpressed Shh (Miura et al., 2001). In fact, in early chickembryogenesis, embryos cultured in the presence of cyclo-
pamine, a potent inhibitor of Shh signaling (Liu et al.,2004), showed downregulation of IGFBP-5 expression. IG-FBP-5 expression was suggested to be regulated by Shh(Allan et al., 2003). Moreover, IGFBP-2, which was ex-pressed in the entire epithelium until E15, became re-stricted to the dorsal epithelium of filiform, fungiform,foliate, and circumvallate papillae at E17. IGFBP-2 andIGFBP-5 expression often overlap in the same cells ortissue or are in adjacent tissues, e.g., taste buds (Suzuki etal., 2005), lung alveolae (Schuller et al., 1993), ectoderm oflimb buds, astrocytes (Green et al., 1994).
In contrast, IGFBP-4 is related to invagination of theepithelium to form grooves or furrows. In our previousstudy, IGFBP-4 was expressed in the bottom of trenches ofcircumvallate papillae at postnatal day 2 (Suzuki et al.,2005). In the present study, IGFBP-4 was expressed in the
Fig. 2. Expression of IGFBP mRNAs (A–H) and immunohistochem-istry for PGP9.5 (I) in developing circumvallate papillae of mouse tongue.A and B: E14 circumvallate papillae showing mRNA expression of IG-FBP-2 (A) and IGFBP-5 (B). Note intense expression of IGFBP-5 in theapex of circumvallate papillae (B). C: Weak expression of IGFBP-3 isseen in the core of mesenchyme. E15. D: Intense expression of IGFBP-4is seen in the epithelium of trench grooves (G). E15. E: Expression of
IGF-IR is seen in the epithelium. E15. F–H: E17. Expressions of IGFBP-2(F), IGFBP-4 (G), IGFBP-5 (H), and PGP9.5 (I). IGFBP-2 is expressed inthe apex of circumvallate papillae (F), and IGFBP-4 in the grooves ofcircumvallate papillae (G). IGFBP-5 expression remains in presumedtaste buds (arrows in H), whose location is limited to the apex ofcircumvallate papillae (arrows in I). Scale bar � 20 �m in A–G and I; 10�m in H.
845ROLE OF IGFBP
entire epithelium until E14 and then became restricted tothe groove of circumvallate and foliate papillae. AmongIGFBPs, IGFBP-4 uniquely inhibits IGF action (Zhou etal., 2003), and so in circumvallate and foliate papillae, itmay be involved in the down-growth of grooves, with itsactivity continuing during early postnatal development.
In filiform papillae, regularly spaced dermal papillaeappeared at E15 in the rat fetus, although the surface ofthe tongue remained flat. At E18 of the rat fetus, prolif-eration in the epithelium overlying the dermal papillaestarted and keratinization appeared at the surface of theepithelium (Baratz and Farbman, 1975), thus giving riseto anterior-posterior polarity (Jonker et al., 2004). Thepresence of IGFBP-2 expression in the anterior epitheliumof each filiform papilla at E17 mice may promote thispolarity. IGFBP-3 was intensely expressed in dermal pa-
pillae of filiform papillae, but very weakly expressed inthose of other papillae. IGFBP-3 is the major IGF carrierprotein in adult serum (Clemmons, 1992) and is also ex-pressed in peripheral embryonic tissue, such as dermalpapillae of filiform papillae and hair follicle (Batch et al.,1996).
LITERATURE CITEDAllan GJ, Zannoni A, McKinnel I, Otto WR, Holzenberger M, Flint DJ,
Patel K. 2003. Major components of the insulin-like growth factoraxis are expressed early in chicken embryogenesis, with IGF bind-ing protein (IGFBP)-5 expression subject to regulation by sonichedgehog. Anat Embryol 207:73–84.
Ayer-LeLievre C, Stahlbom P-A, Sara VR. 1991. Expression of IGF-Iand -II mRNA in the brain and craniofacial region of the rat fetus.Development 111:105–115.
Baratz RS, Farbman AI. 1975. Morphogenesis of rat lingual filiformpapillae. Am J Anat 143:283–302.
Batch JA, Mercuri FA, Werther GA. 1996. Identification and localiza-tion of insulin-like growth factor-binding protein (IGFBP) messen-ger RNAs in human hair follicle dermal papillae. J Invest Dermatol106:471–475.
Clemmons DR. 1992. IGF binding proteins: regulation of cellularactions. Growth Regul 2:80–87.
Duan C. 2002. Specifying cellular responses to IGF signals: roles ofIGF-binding proteins. J Endocrinol 175:41–54.
Farbman AI, Mbiene J-P. 1991. Early development and innervation oftaste bud-bearing papillae on the rat tongue. J Comp Neurol 304:172–186.
Ferguson MW, Sharpe PM, Thomas BL, Beck F. 1992. Differentialexpression of insulin-like growth factors I and II (IGF I and II),mRNA, peptide and binding protein 1 during mouse palatedevelopment: comparison with TGF� peptide distribution. J Anat181:219–238.
TABLE 1. IGFBP mRNA expression patterns infiliform, fungiform, foliate, and circumvallate
papillae of E14-17 mice*
E14 E15 E17
IGF-1R � � � (all four papillae)IGFBP-2 �� �� �� (all four papillae)IGFBP-3 NE �� �� (filiform)IGFBP-4 �� � � (filiform, fungiform)
NE �� �� (groove, foliate, circumvallate)IGFBP-5 �� � � (all four papillae)
*Relative levels of expression are based on hybridization in-tensities of sections. �, signal low to undetectable; NE, notexamined.
Fig. 3. Expression of IGFBP mRNAs in developing foliate papillae ofmouse tongue. A: Intense expression of IGFBP-4 is seen in the grooves(G). E15. B: Note the expression of IGFBP-5 in the ridges among thegrooves (G). E15. C: Expression of IGF-IR is seen in the epithelium. E15.
D: Expression of IGFBP-2 is seen in the ridge epithelium. E17. E:IGFBP-3 is weakly expressed in the core of mesenchyme. E17. F:Expression of IGFBP-4 is still seen in the grooves (arrows). fi, filiformpapillae; fo, foliate papillae. Scale bar � 20 �m in A–E; 10 �m in F.
846 SUZUKI

Green BN, Jones SB, Streck RD, Wood TL, Rotwein P, Pinter JE.1994. Distinct expression patterns of insulin-like growth factorbinding protein 2 and 5 during fetal and postnatal development.Endocrinology 134:954–962.
Hall JM, Hooper JE, Finger TE. 1999. Expression of Sonic hedgehog,Patched, and Gli1 in developing taste papillae of the mouse. J CompNeurol 406:143–155.
Hall JM, Bell ML, Finger TE. 2003. Disruption of sonic hedgehogsignaling alters growth and patterning of lingual taste papillae.Dev Biol 255:263–277.
Jonker L, Kist R, Aw A, Wappler I, Peters H. 2004. Pax9 is requiredfor filiform papilla development and suppresses skin-specific differ-entiation of the mammalian tongue epithelium. Mech Dev 121:1313–1322.
Jung H-S, Akita K, Kim J-Y. 2004. Spacing patterns on tongue sur-face-gustatory papilla. Int J Dev Biol 48:157–161.
Liu H-X, MacCallum DK, Edwards C, Gaffield W, Mistretta CM. 2004.Sonic hedgehog exerts distinct, stage-specific effects on tongue andtaste papilla development. Dev Biol 276:280–300.
Mbiene J-P, Roberts JD. 2003. Distribution of keratin 8-containingcell clusters in mouse embryonic tongue: evidence for a prepatternfor taste bud development. J Comp Neurol 457:111–122.
Mistretta CM, Goosens KA, Farinas I, Reichardt LF. 1999. Alterna-tions in size, number, and morphology of gustatory papillae and
taste buds in BDNF null mutant mice demonstrate neural depen-dence of developing taste organs. J Comp Neurol 409:13–24.
Miura H, Kusakabe Y, Sugiyama C, Kawamatsu M, Ninomiya Y,Motoyama J, Hino A. 2001. Shh and Ptc are associated with tastebud maintenance in adult mouse. Mech Dev 106:143–145.
Nosrat CA, Blomlf J, Elshamy WM, Ernfors P, Olson L. 1997. Lingualdeficits in BDNF and NT3 mutant mice leading to gustatory andsomato-sensory disturbances, respectively. Development 124:1333–1342.
Oakley B, Brandemihl A, Cooper D, Lau D, Lawton A, Zhang C. 1998.The morphogenesis of mouse gustatory vallate epithelium and tastebuds requires BDNF-dependent taste neurons. Dev Brain Res 105:85–96.
Schuller AGP, Zwarthoff EC, Drop SLS. 1993. Gene expression of thesix insulin-like growth factor binding proteins in the mouse concep-tus during mid- and late gestation. Endocrinology 132:2544–2550.
Suzuki Y, Takeda M, Sakakura Y, Suzuki N. 2005. Distinct expres-sion pattern of insulin-like growth factor family in rodent tastebuds. J Comp Neurol 482:74–84.
Yamane A, Mayo ML, Shuler C. 2000. The expression of insulin-likegrowth factor-I, II and their cognate receptor 1 and 2 during mousetongue embryonic and neonatal development. Zool Sci 17:935–945.
Zhou R, Diehl D, Hoeflich A, Lahm H, Wolf E. 2003. IGF-bindingprotein-4: biochemical characteristics and functional consequences.J Endocrinol 178:177–193.
847ROLE OF IGFBP