rights / license: research collection in copyright - …31839/... · erneute...
TRANSCRIPT

Research Collection
Doctoral Thesis
Untersuchungen über einen aus Melolontha melolonthaisolierten Stamm von Bacillus cereus
Author(s): Kabay, Antal
Publication Date: 1965
Permanent Link: https://doi.org/10.3929/ethz-a-000087786
Rights / License: In Copyright - Non-Commercial Use Permitted
This page was generated automatically upon download from the ETH Zurich Research Collection. For moreinformation please consult the Terms of use.
ETH Library

Prom. Nr. 3593
Untersuchungen über einen aus
Melolontha melolontha
isolierten Stamm von Bacillus cereus
Von der
EIDGENÖSSISCHEN TECHNISCHEN
HOCHSCHULE IN ZÜRICH
Zur Erlangung der Würde eines Doktors der
technischen Wissenschaften genehmigte
PROMOTIONSARBEIT
Vorgelegt von
Antal Kabay
dipl. Ing.-Agr. ETH
von Ungarn
Referent: Herr Professor Dr. L. Ettlinger
Korreferent: Herr Professor Dr. H. Kern
Zürich 1965
Ed. Trunlnger, Urania. Zürich

Leer - Vide - Empty

Inhaltsverzeichnis
Seite
Einleitung 7
A) BESCHREIBUNG DER ISOLIERTEN STÄMME 9
I. MORPHOLOGISCHE EIGENSCHAFTEN 9
1. Material und Methoden
2. ErgebnisseII. BIOCHEMISCHE UND PHYSIOLOGISCHE EIGENSCHAFTEN 21
1. Material und Methoden
2. VersuchsergebnisseIII. IMMUNOBIOLOGISCHE EIGENSCHAFTEN 25
1. Material und Methoden
2. VersuchsergebnisseIV. SYSTEMATIK 28
B) ZÜCHTUNG 29
1. Problemstellung2. Material und Methoden
3. Versuchsergebnisse
C) WIRKSAMKEIT VON SPORENPRÄPARATEN 35
I. IM LABORATORIUM DURCHGEFÜHRTE INFEKTIONSVERSUCHE 35
1. Material und Methoden
2. VersuchsergebnisseIL FREILANDVERSUCH 37
1. Problemstellung2. Material und Methoden
3. Versuchsergebnisse
D) UNTERSUCHUNGEN DER HAEMOLYMPHE VON INFIZIERTEN UND
GESUNDEN MELOLONTHA-LARVEN 39
I. BESTIMMUNGEN DES GEHALTES AN FREIEN AMINOSÄUREN IN DER
HAEMOLYMPHE GESUNDER UND KRANKER MELOLONTHA-LARVEN 39
1. Material und Methoden
2. VersuchsergebnisseII. BESTIMMUNGEN DES PROTEINGEHALTES IN DER HAEMOLYMPHE
GESUNDER UND KRANKER MELOLONTHA-LARVEN UND UNTERSUCHUNGEN
ZUM NACHWEIS HUMORALER IMMUNITÄTSFAKTOREN 41
1. Material und Methoden
2. VersuchsergebnisseIII. PASSIVE IMMUNISIERUNG 46
1. Material und Methoden
2. Versuchsergebnisse
Zusammenfassung 46
Literaturverzeichnis 49
Dank
Lebenslauf

Leer - Vide - Empty

MEINEN LIEBEN ELTERN

Leer - Vide - Empty

Einleitung
Da chemisch-synthetische Insektizide infolge Nebenwirkungen oft zu Bedenken Anlass geben, hat
die Schädlingsbekämpfung auf mikrobiologischem Weg in letzter Zeit an Bedeutung zugenommen.
Ein Zweig davon, nämlich die bakteriologische Schädlingsbekämpfung, hat mit Bacillus thurin¬
giensis Berliner (gegen Lepidopteren) und Bacillus popilliae Dutky (gegen Popillia japonica) be¬
reits in der Praxis Eingang gefunden.
In der Schweiz wurden die ersten Untersuchungen zur bakteriologischen Engerlingsbekämpfungim Jahre 1949 am Entomologischen Institut und am Institut fUr landwirtschaftliche Bakteriologieund Gärungsbiologie (heute : Mikrobiologisches Institut) der Eidgenössischen Technischen Hoch¬
schule aufgenommen.
Im Rahmen dieser Untersuchungen isolierte KERN (1950) aus Melolontha-Larven einen sporen¬
bildenden Stamm, der als "Bacillus X" bezeichnet wurde. Dieser Stamm zeigte in Laboratoriums¬
versuchen gute Resultate, kam jedoch nicht zur praktischen Anwendung in Freilandversuchen.
Die Arbeit wurde von WIKEN und WILLE (1953) an den gleichen Instituten weitergeführt, und
zwar durch Isolationen von ca. 150 Bakterienstämmen. In den im Laboratorium durchgeführtenPathogenitätsversuchen erwiesen sich ein sporenbildender Bacillus - von den Autoren "Ai" ge¬nannt - und ein "Kokkenbazillus Stamm 175" als besonders aussichtsreich. In Freilandversuchen
wurde mit Bacillus AY in den Jahren 1953, 1954 und 1955 (WIKEN, BOVEY, WILLE und WILD¬
BOLZ, 1954, und WILLE, WIKEN und BOVEY, 1962) eine 50%ige Reduktion der Engerlingspopu¬lationen erzielt. Der "Kokkenstamm 175" erzielte in Freilandversuchen im Jahre 1955 (WILLE,WIKEN und BOVEY, 1962) ebenfalls eine 50%ige Mortalitätsrate. Leider war es uns nicht mög¬lich, Proben dieses Materials zu erhalten.
Etwa gleichzeitig im Jahre 1955 beobachteten HURPIN (1955) in Frankreich und WILLE (1956) in
der Schweiz eine Krankheit, die bis dahin bei Engerlingen des Maikäfers noch nicht beschrieben
worden war. Krankheitsbild und Erreger entsprachen der von DUTKY (1940) entdeckten "MilkyDisease" des Japankäfers, Popillia japonica Newm., hervorgerufen durch Bacillus popilliae Dut¬
ky. Während HURPIN (1. c. ) den Krankheitserreger von Engerlingen nur als eine auf Melolontha
spezialisierte europäische Rasse von Bacillus popilliae ansah, glaubte sich WILLE (I.e.) zur
Aufstellung einer neuen Species, Bacillus fribourgensis Wille, berechtigt. Welche Auffassungrichtig ist, blieb unentschieden, da es bis heute noch nicht gelungen ist, Bacillus popilliae von
Popillia oder von Melolontha bzw. Bacillus fribourgensis in künstlicher Kultur zu einem vollstän¬
digen Wachstumszyklus mit Sporenbildung zu bringen. Aus dem gleichen Grund kam dieser er¬
folgversprechende Erreger in Europa noch nicht zur praktischen Anwendung. (Der erste Frei¬
landversuch mit Bacillus fribourgensis wird in der vorliegenden Arbeit beschrieben. )
KRIEG (1957) untersuchte die Pathogenität von Bacillus thuringiensis auf Melolontha-Larven durch
perorale Verabreichung des Sporen-Endotoxin-Komplexes, fand aber keine biologische Wirkung.Dieses Resultat wird in der vorliegenden Arbeit durch Untersuchungen mit verschiedenen Bacil¬
lus thuringiensis-Stämmen - wovon einer von LEISINGER (ETTLINGER, LEISINGER und KABAY,1964) aus Melolontha-Larven isoliert wurde - bestätigt.
Erneute Untersuchungen im Hinblick auf eine mikrobiologische Engerlingsbekämpfung wurden in
der Schweiz im Jahre 1961 an unserem Institut aufgenommen. Wie schon berichtet (ETTLINGER,LEISINGER und KABAY, 1964), wurden in Altdorf (Kt. Uri) aus kranken Melolontha-Larven neue
Isolationen vorgenommen. Diese Stämme zeigten in Laboratoriumsversuchen vorerst gute Wirk¬
samkeit gegen Melolontha-Larven, und in der Hoffnung, dass sie sich für die bakteriologischeSchädlingsbekämpfung eignen könnten, wurden in der vorliegenden Arbeit *
Untersuchungen zur
Erforschung ihres Charakters und ihrer Wirksamkeit durchgeführt.
* Diese Arbeit wurde durch einen Kredit der Eidgenössischen Volkswirtschafts-Stiftung finanziert, wo¬
für an dieser Stelle bestens gedankt wird.
7

Hatten wir anfänglich geglaubt, bei diesen Stämmen handle es sich um Vertreter eines neuen,
kristallbildenden Bacillus aus der Gruppe des Bacillus thuringiensis, so beruhte dies in erster
Linie auf einer fehlerhaften Ausführung der Fettfärbung mit Sudanschwarz. Später erwiesen
sich die vermeintlichen 3 Kristallkörper im Sporangium als aus Poly-beta-hydroxybuttersäurebestehende Fettropfen. Damit ordnete sich Stamm Altdorf, wie er im folgenden genannt wird,
zwanglos in die Art Bacillus cereus Frankland et Frankland ein. Die in unserer vorläufigen Mit¬
teilung (ETTLINGER, LEISINGER und KABAY, 1964) in Aussicht gestellte Beschreibung einer
neuen Species konnte noch rechtzeitig unterdrückt werden und der dort gebrauchte Name "Ba¬
cillus polystictus" ist ein nomen nudum geblieben.
8

A) BESCHREIBUNG DER ISOLIERTEN STÄMME
Problemstellung
Das Hauptanliegen der nachfolgenden Untersuchungen war die Erforschung der morphologi¬schen, physiologischen und serologischen Eigenschaften der isolierten Stämme. Die serolo¬
gischen Untersuchungen betrafen ausschliesslich die H-antigenen Eigenschaften der begeis-selten Formen und strebten die Abklärung folgender Fragen an :
a) Haben alle Bacillus er 3us Altdorf-Stämme untereinander die gleiche Antigen-Struktur ?
b) Haben die Bacillus ceieus Altdorf-Stämme gemeinsame antigène Eigenschaften mit ande¬
ren Bacillus cereus Stämmen oder Vertretern anderer Arten der Bacillus cereus Gruppeund erlauben die vorhandenen Unterschiede in der Partialantigenstruktur die Aufstellungverschiedener serologischer Gruppen ?
I. MORPHOLOGISCHE EIGENSCHAFTEN
1. Material und Methoden
a) Prüfung der Beweglichkeit
Die Beweglichkeit der Bacillus cereus AltdorfrStämme wurde auf folgenden Medien
studiert :
Nährbouillon : 10 g Pepton Difco, 10 g Oxo Lab Lemco, 4 g NaCl, H2O dest. ad 1 1,
pH 7.
Halbstarres Medium : Nährbouillon +0,4% Agar.Kombination von halbstarrem und flüssigem Medium : wie unter Kapitel "Immuno-
biologische Eigenschaften" beschrieben.
Modifiziertes Fischer's Medium nach NORDBERG (1951 und 1953)
b) Färbungen
Geisseifärbung nach LEIFSON (1960)
Die zu untersuchenden Mikroorganismen wurden in Nährbouillon in SchUttelkultur
(100 Ausschläge/Min. ) während 6 Std. bei 30° C bebrütet, mit 5 % Formalin ver¬
setzt, mit H2O dest. verdünnt und während 6 Minuten bei 600 g zentrifugiert.
Herstellung der Farblösungen und angewandte Färbungstechnik s. LEIFSON (1960).
Die folgenden drei Färbungen wurden nach den "Staining Procedures used by the
Biol. Stain Commission (I960)" durchgeführt :
Gramfärbung nach HUCKER
Sporenfärbung nach DORNER und WIRTZ
Kapselfärbung nach ANTHONY
Darstellung von Einschlüssen lipider Natur mit Sudan Schwarz B nach BURDON (1946)
Das Material wurde auf Medium 1) (s. Kapitel "Züchtung") mit 8, 16 und 24 Std. Be-
brütung in Schüttelkulturen mit 100 Ausschlägen/Min. gezüchtet. Zum Vergleichwurden die Färbungen auch nach vorheriger Lipidextraktion durchgeführt. Als Ex¬
traktionsmittel dienten 96%iger Alkohol und Aceton mit Einwirkungszeiten von 24,
48, 72 und 195 Std. Günstige Färbungszeit : 18-20 Min., Gegenfärbungszeit:10-15 Min.

c) Beobachtungen im Licht-Mikroskop
Die Entwicklung von vegetativen Zellen und Sporen wurde mit einem LEITZ-Mikro¬
skop (Typ Ortholux II) mit eingebauter Beleuchtung durch einen Mikroansatz "Mikas"und das dazu gehörende LEICA-Gehäuse Typ LEICA M3 aufgenommen. Als Licht¬
quelle diente ein Mikroblitz-Gerät (Mikroblitz 300) mit einstellbarer Lichtintensitätvon 150 und 300 Wattsekunden. FUr Phasenkontrast- und Dunkelfeld-Einstellung wur¬de ein Objektiv Pv. Ap. Oel 90/1,15 n (apochromatisches Oelimmersionssystem) ver¬
wendet.
d) Elektronenmikroskopi sehe Untersuchungen
Es wurden drei Verfahren angewendet :
Verfahren I. Methode nach KELLENBERGER ET AL (1958)
Bakteriensuspensionen wurden auf Medium 1) (Kartoffelextrakt + 0,5% Glukose) ge¬züchtet und in verschiedenen Altersstadien geprüft. Je 10 cc Zellmaterial wurden
abzentrifugiert, das Sediment mit 1%-igem OSO4 fixiert, während 2 Std. im Kühl¬
schrank bei + 4° C aufbewahrt und anschliessend entwässert mit : 25 %igem und 50 %igem Aceton je 30 Minuten ; 75%-igem Aceton + 1 %Uranylacetat Über Nacht ; 95 %-igem und 100 %igem Aceton je 30 Minuten und 2mal Aceton Über CuSO^ je 30 Minu¬
ten. Eingebettet wurde in Araldit und Aceton im Mischungsverhältnis 1:32 Std. -
1:12 Std. - 3 : 1 über Nacht. Anschliessend wurde das Präparat in eine Gelatine-
Kapsel gelegt und in Araldit-Gemisch 6 Std. bei 80° C polymerisiert. Die DUnn-
schnitte wurden mit einem PORTER-BLUM Ultramikrotom hergestellt. Die Schnitt¬dicke betrug ca. 500 - 700 Â. Verwendetes Elektronen-Mikroskop: PHILIPS 100.
Verfahren II.
Die Bakterien wurden ebenfalls auf Medium 1) gezüchtet. Das Bakterien-Material
wurde mit dem Gefrierätzverfahren behandelt. FUr die detaillierte Beschreibung der
Präparierung s. MOOR und MÜHLETHALER (1963). Verwendetes Elektronen-
Mikroskop: SIEMENS Elmiskop I. *
Verfahren III.
Zur Darstellung der freien Sporen wurde das Material auf Formvar-beschichtete
Trägernetzchen direkt aufgetragen. Formvar-Filme wurden nach der Eintauchme¬
thode von SCHAEFFER und HARKER (1942) zubereitet. Elektronen-Mikroskop :
PHILIPS 100.
e) Prüfung der Hitze- und UV-Resistenz
Das Sporenmaterial wurde auf Medium 11) (s. Kapitel "Züchtung") hergestellt, drei¬
mal mit H2O dest. gewaschen und bei Zimmertemperatur getrocknet. FUr die Prü¬
fung der Trockenhitze- und UV-Resistenz wurden die Sporen auf Petrischalendeckel
in ca. 0,2 mm dicker Schicht gleichmässig verteilt und im Trockenschrank der Hit¬
ze ausgesetzt, resp. in einem geschlossenen Kasten (ca. 120 x 50 x 50 cm) mit
einer 30 Watt UV-Lampe (G30T8 Gen. Electr. Germ. Lamp ; Wellenlänge 2537 Â)von 20 cm Entfernung bestrahlt. Als weiterer Versuch wurde eine Oese voll Sporenin Nährbouillon über der Heizplatte bei 100° C gekocht, bzw. bei 120° C und 1 atu
autoklaviert.
Die vegetativen Zellen wurden nach 20 Std. BebrUtung (in Nährbouillon) auf Agar ab¬
geimpft und bestrahlt (ohne Deckel), bzw. auf Nährbouillon abgeimpft und im Wasser¬
bad erhitzt.
Die Keimungsfähigkeit der behandelten Sporen und vegetativen Zellen wurde jeweilsin Nährbouillon geprüft.
* FUr diese Aufnahmen sei Herrn PD Dr. H. Moor, Elektronenmikroskopisches Laboratorium der
E.T.H., recht herzlich gedankt.
10

2. Ergebnisse
a) Allgemeines
Der Organismus Bacillus cereus Stamm Altdorf zeigt als Vertreter des Genus Bacil¬
lus einen morphologischen Phasenwechsel von vegetativer Phase und Sporenphase.Nach dem Austritt der Spore aus dem Sporangium hat dieses keine Funktion mehr
und verschwindet nach und nach durch Lysis. Zur Sporenbildung sind alle vegetati¬ven Zellen fähig, wobei eine Zelle jeweils eine einzige Spore bildet. Die Entwick¬
lung der vegetativen Zelle zur freien Spore ist von bestimmten Umweltsbedingungenabhängig (s. Kapitel "Züchtung")
b) Einzelzelle
Die Einzelzelle ist stäbchenförmig mit abgerundeten Zellenden und einer Grösse
von durchschnittlich 4,3 x 1,6 u mit Streuungswerten von s = + 1,5 x 0,4 u. Die
Zellen treten einzeln oder in kettenförmigen Verbänden auf. Sie sind im Blut der
Engerlinge, sowie in allen unter Material und Methoden beschriebenen Medien be¬
weglich. Als Begrenzungsfaktor der Beweglichkeit gilt auf allen Prüfmedien das Al¬
tersstadium : mit zunehmendem Alter nimmt die Beweglichkeit ab. Die Geissein
sind peritrich angeordnet. Die vegetativen Zellen sind gram-positiv. Die Sporensind ellipsoid bis zylindrisch, liegen parazentral bis subterminal und weisen eine
Grösse von durchschnittlich 1,6 x 1,2 u auf. Sie lassen sich mit gewöhnlichen Ani¬
linfarbstoffen gut anfärben PORNER-WIRTZ). Die Sporenmutterzelle (Sporangium)wird nicht aufgetrieben. Eine Kapselbildung konnte nicht nachgewiesen werden
(ANTHONY'S Methode 1931).
c) Kolonien
Auf Agar gezüchtet bildet Bacillus cereus Stamm Altdorf unregelmässige, glatte,flache, lappige, bei auffallendem Licht weisse, in durchfallendem Licht leicht
bräunliche Kolonien. Im Gelatinestich erfolgt sackartige Verflüssigung. Auf Nähr¬
bouillon gezüchtet, bildet sich eine flockige Oberflächenhaut bei sonst klarer Brühe.
In der Schüttelkultur bilden sich in Schollen zerfallende Kolonien ; es tritt keine
gleichmässige Trübung auf.
d) Licht- und elektronenmikroskopische Untersuchungen über die Entwicklung der Spo¬ren und Einschlusskörper
Neben der Endospore bildet Bacillus cereus Stamm Altdorf in der gleichen Sporen¬mutterzelle in der Regel 3 Einschlusskörper, die bei der Auflösung des Sporangiumsvon der Spore getrennt freigesetzt werden. Die Grösse beträgt im Durchschnitt ca.
0,4-0,6 u. Im Lichtmikroskop bei Phasenkontrast erscheinen die in Wasser sus¬
pendierten vegetativen Zellen dunkel auf hellerem Grund (Abb. 1). Im Dunkelfeld
(Abb. 2) erkennt man körnige Einschlüsse des Zytoplasmas (bei Bacillus thuringien-sis var. thuringiensis von YOUNG and FITZ-JAMES, 1959, "Grana" genannt). Auf
einem Sporulationsmedium (s. Kapitel "Züchtung") erscheinen nach 12-13 Std. Be-
brütung die Vorsporen bei Phasenkontrast als helle, bzw. später (nach 16-18 Std. )die Sporen als glänzend weisse Zonen in den Sporenmutterzellen. Die Einschluss¬
körper sind als helle, lichtbrechende Körperchen erkennbar (Abb. 3 und 4). Im
Dunkelfeld sind Sporen und Einschlusskörper strahlend weiss (Abb. 5 und 6).
Die Entwicklung der Vorspore zur Spore und deren Feinstruktur, sowie der Ein¬
schlusskörper lässt sich im Elektronenmikroskop anhand von Ultradünnschnitten
gut verfolgen. Nach ca. 12 Std. Bebrütung erscheint die Vorspore in eher rundlicher
Form, umgeben von den zwei Vorsporenmembranen, sowie vom Exosporium, das
sich jedoch von der äusseren Vorsporenmembran noch nicht deutlich abhebt (Abb. 9)Nach 14-16 Std. Bebrütung wird die Vorspore länglich und das Exosporium hebt sich
klar von den Vorsporenmembranen ab. Ausserhalb des Exosporiums; d. h. im WinkelL
11

zwischen Zellwand und Exosporium werden die kugeligen Einschlusskörper ausge¬bildet (Abb. 10 und 11). Nach ca. 18 Std. Bebrtltung ist die Spore vollständig ausge¬bildet ; die Sporenwand, die innere und äussere Sporenmembran und das Exosporiumsind deutlich erkennbar (Abb. 12). Nach ca. 20 Std. Bebrlitung, also kurz vor dem
Austritt der Sporen und Einschlusskörper, nehmen die Einschlusskörper unregel¬mässige und teilweise kantige Formen an (Abb. 13 und 14). Die Sporen behalten
nach der Freisetzung das Endosporium (Abb. 7 und 8).
e) Hitze- und UV-Resistenz
Die Sporen sind in getrocknetem Zustand nach 2 Std. Behandlung mit 190° C Trok-
kenhitze und nach 50 Min. Kochen bei 100° C noch keimungsfähig. Eine 100 %ige Ab-
tötung der Sporen wurde durch Trockenhitze-Behandlung während 30 Min. bei 220° C,
resp. 20 Min. Autoklavieren bei 120° C und 1 attl erreicht. Diese relativ hohe Hitze¬
resistenz der Sporen erleichtert die Isolation des Bacillus.
Die vegetativen Zellen wurden durch 4-5 Minuten UV-Bestrahlung total abgetötet.Die Sporen waren nach 24 Std. noch keimungsfähig. (BONNEFOI & BEGUIN (1959)berichten über einen UV-resistenten Bacillus thuringiensis Stamm "Anduze", der
bis 18 Std. Bestrahlung aushielt).
12

Abb. 1 Vegetative Zellen von Bacillus cereus Stamm
Altdorf im Lichtmikroskop mit Phasenkontrast-
Einstellung.X 3'250
Abb. 2 Vegetative Zellen von Bacillus cereus Stamm
Altdorf im Lichtmikroskop mit Dunkelfeld-Ein¬
stellung. Zellwand und Grana deutlich erkennbar.
X 3*250
13
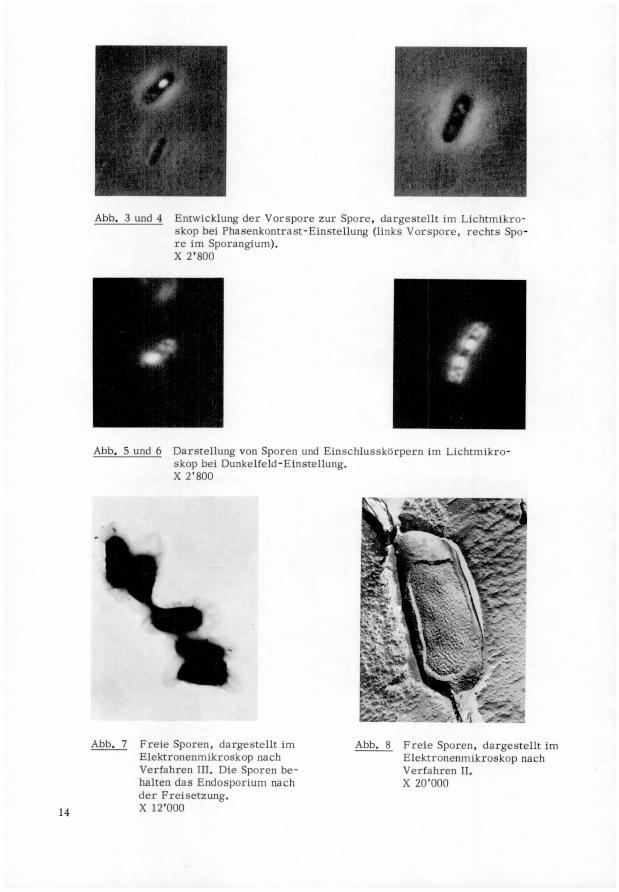
Abb. 3 und 4 Entwicklung der Vorspore zur Spore, dargestellt im Lichtmikro-
skop bei Phasenkontrast-Einstellung (links Vorspore, rechts Spo¬re im Sporangium).X 2'800
Abb. 5 und 6 Darstellung von Sporen und Einschlusskorpern im Lichtmikro-
skop bei Dunkelfeld-Einstellung.X 2'800
14
Abb. 7 Freie Sporen, dargestellt im
Elektronenmikroskop nach
Verfahren III. Die Sporen be¬
halten das Endosponum nach
der Freisetzung.X 12'OÜO
Abb. 8 Fieie Sporen, dargestellt im
Elektronenmikroskop nach
Verfahren II.
X 20'000

9 Zelle nach 12 Std. Bebrutung. Die Vorspore (VSP) ist von der inneren Vor¬
sporenmembrane (IVSPM), der äusseren Vorsporenmembrane (AVSPM) und
vom Exosporium (EX) umgeben, das sich von der äusseren Vorsporenmem¬brane noch nicht überall deutlich abhebt. (ZW = Zellwand)
Elektronenmikroskopische Darstellung nach Verfahren II.
X 40'000
15

-v »,<•
Abb. 10 Zelle nach 14 Std. Bebrutung. Die Vorspore
(VSP) ist länglich, und das Exosporium (EX)hebt sich klar von den beiden Vorsporenmem¬branen ab. Im Winkel zwischen Zellwand
(ZW) und Exosporium (EX) werden die kuge¬
ligen Einschlusskorper (E) ausgebildet.
Elektronenmikroskopische Darstellung nach
Verfahren II.
X 32'000
16

Abb. 11 Zelle nach 15 Std. Bebrutung. Längliche Vor¬
spore (VSP) umgeben von Exosponum (EX).Im Winkel zwischen Zellwand und Exosponum
(EX) die Einschlusskorper (E).
Elektronenmikroskopische Darstellung nach
Verfahren I.
X 30"000
17

Abb. 12 Zelle nach 18 Std. Bebrutung. Die Spore ist
vollständig ausgebildet. Sporencortex (C),innere (ISPM) und äussere (ASPM) Sporen¬membran und Exosporium (EX) sind deutlich
erkennbar.
Elektronenmikroskopische Darstellung nach
Verfahren II.
X 32*000
18

Abb. 13 Zelle nach 20 Std. Bebrütung. Die Einschluss-
korper (E) erscheinen kantig. Sporencortex
(C), äussere Sporenmembran (ASPM), innere
Sporenmembran (ISPM), Exosponum (EX).
Elektronenmikroskopische Darstellung nach
Verfahren II.
X 40*000
19

Abb. 14 Zelle nach 20 Std. Bebrutung. Einschlusskor-
per (E), Cortex (C), Exosponum (EX).
Elektronenmikroskopische Darstellung nach
Verfahren I.
X 30'000
20

II. BIOCHEMISCHE UND PHYSIOLOGISCHE EIGENSCHAFTEN
1. Material und Methoden
a) Medien
Die Herstellung der Medien für die physiologischen Untersuchungen und die Prüfungdes Reaktionsausfalles erfolgten nach den jeweils angegebenen Autoren.
Lecithinase-Reaktion auf Eigelb-Agar, Eigelb-Salz und Eigelb-Bouillon.McGAUGHEY and CHU (1948).
Phosphatase-Reaktion auf Natriumphenolphthaleindiphosphat.
Die Zone des durch die Phosphatase abgespaltenen Phenolphthalein wurde in mm ge¬
messen. BÖTTCHER (1959).
HarnstoffSpaltung auf Agar. CHRISTENSEN (1946)
Verflüssigung von koaguliertem Serum nach LÖFFLER.
MACKIE and McCARTNEY (1960)
Salicin-Test auf Fleischextraktbouillon mit 0,5 und 1 % Salicin.
DIFCO-MANUAL (1953)
Pigment-Bildung auf koaguliertem Eidotter. MÜLLER-KÖGLER (1959).
Caseinhydrolyse auf Milchagar. HASTINGS (1903).
Die Temperaturabhängigkeit des Wachstums wurde auf Nährbouillon und Glukose-
Agar geprüft.
Die Ausnützung von L-Glutaminsäure wurde auf folgendem Medium nach KNIGHT
and PROOM (1950) geprüft :
KH2P04 1,5 g; MgS04. 7H20 0,5 g; FeSO4.7H20 0,025 g; CaCl2.2H20 0,3 g;
MnS04.4H20 0,04 g; Ammoniummolybdat 0,002 g; H20 dest. ad 1000 ml. pH 7.
+ L-Glutaminsäure als einzige N- und C-Quelle 6 g/L.
Folgende Teste wurden nach HALLMANN (1961) durchgeführt:
Verhalten gegenüber Q2 auf Schrägagar mit WRIGHT-BURRI-Verschluss.
Schwefelwasserstoff-Bildung auf Bleiacetat-Agar mit 0,02 % Bleiacetat.
Aesculin-Spaltung auf Fleischextraktbouillon mit 0,1 % Aesculln.
Fettspaltung auf Agar mit 3 % Erdnussöl.
HarnstoffSpaltung auf GlukosepeptonboulUon. Modifiziertes CHRISTENSEN-Medium.
Peroxydase-Reaktion auf Agar, Als Reagens wurde Tetramethyl-p-phenylendiamin-hydrochlorid in 1% iger Lösung verwendet.
Folgende Teste wurden nach dem Manual of Microbiological Methods (1957) durch¬
geführt :
Zuckervergärung auf Grundmedium + 1 % der in Tab. 2 angeführten Kohlehydrate.Grundmedium: (NH4)H2P04 1 g (einzige N-Quelle); KCl 0,2 g; MgS04. 7H200,2 g; H2Odest. ad 1000 ml. pH 7.
Voges-Proskauer-Reaktion auf Glukose-Pepton-Bouillon.
Nitratreduktion auf Nitratbouillon.
21

Indolbildung auf Tryptophan-Bouillon.
Proteolytische Aktivität auf alkalischem Eimedium.
Stärkehydrolyse auf Agar mit 0,2% löslicher Stärke.
Katalase-Reaktion auf Agar. Prtlfreagens : 3 % H202.
Gelatine-Verflüssigung auf Nährgelatine.
Gelatine-Ausnützung auf Agar mit 0,4% Gelatine.
Peptonisierung von Milch auf neutralem Lackmusmilch-Medium.
Haemolyse auf Blutagar.
Zitrat-Ausnützung auf synthetischem Medium nach KOSER.
Wachstum auf Nährbouillon und auf Nährbouillon + 4 % NaCl.
b) pH-Bestimmung, Indikatoren
Die pH-Werte wurden unter Verwendung einer Glaselektrode eingestellt. Die Säure-,
bezw. Alkalibildung bei den Gärungstesten wurde mit folgenden Indikatoren, herge¬stellt nach SÖRENSEN, CLARK, LUBS und COHEN (HALLMANN 1961), nachgewie¬sen:
Bromthymolblau pH-Bereich 6 -7,6
Bromkresolpurpur pH-Bereich 5,2 - 6,8
Bromkresolgrün pH-Bereich 3,8 - 5,4
c) Glaswaren Und Sterilisation
Für die Urease-Reaktion und die L-Glutaminsäure-Ausnützung wurden sterile 100-
ml-Erlenmeyerkolben, für alle übrigen Teste sterile Reagenzgläser und Petrischa¬
len verwendet. Die Medien wurden im Autoklav bei 120° C, 1 atü, während 10 - 20
Minuten sterilisiert.
d) Impfmaterial und Bebrütung
Als Ausgangsmaterial zur Herstellung von Impfsuspensionen dienten neben den Ba¬
cillus cereus Alt dorf-Stämmen lyophilisierte Präparate der in Tab. 1 angegebenenStämme. Geimpft wurde auf Nährbouillon und anschliessend auf Schrägagar, der
dann bei + 4° C aufbewahrt wurde. Auf diese Stammkulturen wurde für die Untersu¬
chungen Jeweils zurückgegriffen. Die oben definierten Medien wurden in mindestens
drei parallelen Versuchsreihen mit einer Oese voll 24 Std. alter Agarkultur, bzw.
einem Tropfen (0,05 ml) 18 Std. alter Nährbouillon-Schüttelkultur (100 Ausschläge/Min. ) beimpft. Die Bebrütung auf Nährgelatine und in Nitrat-Bouillon erfolgte bei
20° C, für alle übrigen Teste bei 30° C. Die Reaktion wurde nach 5, 16, 24 und 36
Std. bis 30 Tagen geprüft.
e) Stämme
Ausser den Bacillus cereus Altdorf-Stämmen wurden zwei Bacillus cereus Samm¬
lungs-Stämme und zwei Bacillus thuringiensis var. thuringiensis Stämme -
wovon
je ein Stamm auch aus Melolontha isoliert -, sowie Bacillus thuringiensis var. ales-
ü, Bacillus thuringiensis var. sotto, Bacillus megaterium, Bacillus subtilis und
Bacillus apthracis als Vergleichsstämme in die Untersuchungen einbezogen. Ferner
wurde als typischer Urease-Bildner Proteus vulgaris als Vergleichsstamm verwen¬
det. Tab. 1 gibt nähere Informationen über diese Stämme.
22

2. Versuchsergebnisse
Die Resultate sind in Tab. 2 und 3 zusammengefasst. Wie daraus hervorgeht, sind alle
Bacillus cereus Altdorf Stamme mesophil. Sie sind für folgende Reaktionen positiv :
Voges-Proskauer, Zitrat, Peroxydase, Katalase, Phosphatase, Haemolyse, Lecithina-
se (s. Abb. 15 und 16). Sie besitzen starke proteolytische Aktivität auf einfachen und
zusammengesetzten Eiweiss-Stoffen (s. Abb. 17), bilden organische Säuren ohne Gas
von Glukose, Fruktose, Saccharose, Dextrin, Aesculin und Salicin auf einem Grund¬
medium mit Ammonphosphat als einziger Stickstoffquelle und verwenden auf dem Salz¬
medium L-Glutaminsäure als einzige N- und C-Quelle, wobei sie auch sporulieren.
Diese Resultate treffen mit gewissen Abweichungen auch für Bacillus thuringiensis var.
thuringiensis, Bacillus thuringiensis var. alesti, Bacillus thuringiensis var. sotto und
Bacillus cereus LBG B 4061 zu. Unterschiede zwischen diesen Arten und Varietäten be¬
stehen hinsichtlich der Intensität der Lecithinase- und Phosphatase-Reaktionen, der
Vergärung von Saccharose, Aesculin und Salicin und der proteolytischen Aktivität auf
einfachen und zusammengesetzten Eiweiss-Stoffen, sowie in bezug auf die Temperatur¬
abhängigkeit des Wachstums.
In Tab. 3 wurden die untersuchten Stämme von Bacillus cereus und Bacillus thuringien¬
sis aufgrund der unterschiedlichen biochemischen und physiologischen Merkmale ein¬
ander gegenüber gestellt und in Gruppen eingeteilt.
Wie aus Tab. 2 und 3 hervorgeht, sind alle Bacillus cereus Altdorf Stämme, die aus
verschiedenen Wirtsindividuen derselben Art (Melolontha melolontha L. ) isoliert wur¬
den, in bezug auf ihren biochemischen und physiologischen Charakter homogen und wur¬
den deshalb derselben Gruppe zugeteilt.
Die Bacillus thuringiensis var. thuringiensis Stämme, die aus verschiedenen Wirtsgat¬tungen, nämlich Ephestia und Melolontha, isoliert wurden, zeigen ebenfalls grosse
Uebereinstimmung in ihren Eigenschaften und wurden einer zweiten Gruppe zugeteilt.Sie unterscheiden sich von den Bacillus cereus Altdorf Stämmen unter anderem darin,
dass sie Proteide (alkalisches Eimedium) nicht spalten können, dass sie bei der Pepto-
nisierung von Milch, bei der SerumverflUssigung und der Phosphatase-Reaktion eine
geringere Aktivität aufweisen und dass ihr Temperaturminimum für das Wachstum
höher liegt.
Bacillus cereus B 4126, der von KRIEG 1955 aus Melolontha sp. isoliert wurde, weist
im Gegensatz zu Stamm B 4061 nicht den typischen Bacillus cereus Charakter auf. Er
weicht davon vor allem in bezug auf die intensive Ureasebildung ab ; dies ist jedochnicht als Einzelfall zu betrachten, da BUCHER (1960) aus Heuschrecken ebenfalls urea-
sepositive Bacillus cereus Stämme isolierte. Die weiteren Unterschiede liegen in der
sehr schwachen Lecithinase-Reaktion und der negativen proteolytischen Aktivität auf
alkalischem Eimedium. Diese beiden Bacillus cereus Stämme wurden in separate Grup¬pen (III und IV) eingeordnet. Von den Bacillus cereus Altdorf Stämmen und Bacillus
thuringiensis var. thuringiensis Stämmen unterscheiden sie sich darin, dass sie aus
Aesculin, Saccharose und Salicin keine Säure bilden und dass sie geringere Aktivität
aufweisen in der Phosphatase- und Lecithinase-Reaktion, der SerumverflUssigung und
Peptonisierung von Milch ; ferner liegen ihre Temperatur-Maxima höher.
23
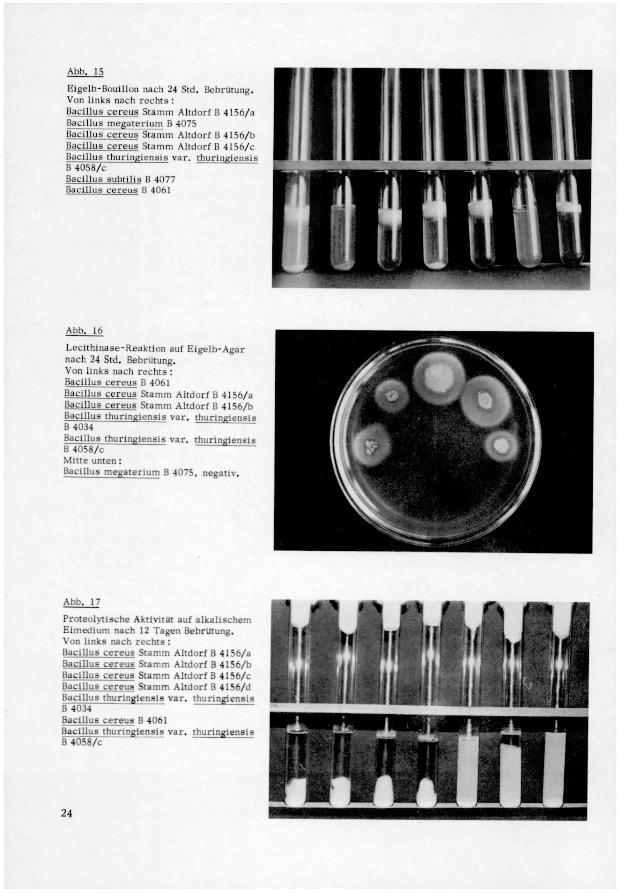
Ahh. 15
bigclb-Bouillon nach 24 Std. Bebrutung.Von links nach rechts •
Bacillus cereus Stamm Altdorf B 4156/jBacillus megatertum B 4075
Bacillus cereus Stamm Altdorf B 4156/hBacillus cereus Stamm Altdorf ß 41 -m/Bacillus thunnglensis var. thurinKiB 4058/cBacillus subtihs B 4077
Bacillus cereus B 4061
Abb. 16
Leothinase-Reaktion auf Eigelb-Agarnach 24 Std. Bebrutung.Von links nach rechts •
Bacillus cereus B 4061
Bacillus cereus Stamm Altdorf B 4156/aBacillus cereus Stamm Altdorf B 4156/bBacillus thunnglensis var. thunngiensi-B 4034
Bacillus thunngiensis var. thunnglensisB 4058/cMitte unten :
Bacillus megaterium B 4075, negativ.
Abb. 17
Proteolytische Aktivität auf alkalischem
Eimedium nach 12 Tagen Bebrlltung.Von links nach rechts :
Bacillus cereus Stamm Altdorf B 4156/aBacillus cereus Stamm Altdorf B 4156/bBacillus cereus Stamm Altdorf B 4156/cBacillus cereus Stamm Altdorf B 4156/dBacillus thuringlensls var. thunngiensisB4034
Bacillus cereus B 4061
Bacillus thunngiensis var. thuringlenslsB 4058/c
24

vulgaris
Proteus
anthracis
Bacillus
ssubtil!
Bacillus
megaterium
Bacillus
cereus
Bacillus
cereus
Bacillus
sotto
var.
thuringiensis
Bacillus
alesti
var.
thuringiensis
Bacillus
ZUrich
Institut
Hygiene
ZUrich
Tierspital
ZUrich
Kantonsspital
2193
CN
Kent.
Lab.
Res.
Welcome
Darmstadt
Bundesanstalt,
Biol.
Krieg.
A.
1955
10702
ATCC
Randall
W.A.
1949
CCEB
Ishiwata
1902
CCEB
Vago
und
Toumanoff
1951
10792
ATCC
Steinhaus
E.A.
1950
ntt
ti
tt
tt
tl
il
it
It
If
ff
(1
tl
ff
ff
ETH
Institut
Mikrobiologisches
1961
L.
melolontha
Melolontha
L.
mori
Bombyx
L.
mori
Bombyx
(Zel
l)kUhniella
Ephestia
hii
ii
ii
mti
tt
ii
L.
melolontha
Melolontha
4003
BLBG
217
LBGB
4077
BLBG
4075
BLBG
4126
BLBG
4061
BLBG
4084
BLBG
4089
BLBG
4034
BLBG
8/c
405
BLBG
4156/e
BLBG
4156/d
BLBG
4156/c
BLBG
4156/b
BLBG
4156/a
BLBG
thuringiensis
var.
thuringiensis
Bacillus
thuringiensis
var.
thuringiensis
Bacillus
Altdorf
Stamm
cereus
Bacillus
Altdorf
Stamm
cereus
Bacillus
Altdorf
Stamm
cereus
Bacillus
Altdorf
Stamm
cereus
Bacillus
Altdorf
Stamm
cereus
Bacillus
VON
AUS
ISOLIERT
Inst.
Mikrobiol.
ETH
NO.
REF.
SPEZIES
Mikroorganismen
verwendeten
Untersuchungen
physiologischen
den
In
die
liber
Angaben
1Tab.

Tab. 2
Zusammenstellung der Resultate der
biochemischen und physiologischen
Untersuchungen.
Stämme
Lecithinase
a<L)
SP3
3OCQl
ap,bpS
n
I
a
bo•1-4
N
cd
cd «^
&E
Oh •O-
cd CO
D. M
e cd4-*
0) «
h w
cda
I<uDl,
Bacillus cereus Stamm Altdorf B 4156/a
Bacillus cereus Stamm Altdorf B 4156/b
Bacillus cereus Stamm Altdorf B 4156/c
Bacillus cereus Stamm Altdorf B 4156/d
Bacillus cereus Stamm Altdorf B 4156/e
Bacillus thuringiensis var.
thuringiensis B 405 8/c
Bacillus thuringiensis var.
thuringiensis B 4034/ATCC 10792
Bacillus muringiensis var. alesti B 4089
Bacillus thuringiensis var. sotto
B 4084/CCEB Prag
Bacillus cereus B 4061/ATCC 10702
Bacillus cereus B 4126
Bacillus megaterium B 4075
Bacillus subtilis B 4077
Bacillus anthracis B 217
Proteus vulgaris B 4003
+-H-
-H+
-m-
+++
1,5-1
1,5-1
1,5-1
1,5-1
1,5-1
0,9
0,9
0,2
1,8-2,0
0,5
0,5
Zeichenerklärung :
Wo nichts Spezielles erwähnt, gilt : + positive Reaktion, - negative Reaktion
Lecithinase
Eigelb-Salz und -Bouillon: +++ Flockenbildung innert 5 Std.
Koloniebildung innert 16 Std.
++ Flockenbildung innert 16 Std.
Koloniebildung innert 24 Std.
+ Flockenbildung innert 24 Std.
Koloniebildung innert 48 Std.
- keine Reaktion
+++ üppige Koloniebildung innert 16 Std.
++ Üppige Koloniebildung innert 24 Std.
+ dünne Koloniebildung innert 48 Std.- keine Reaktion
Eigelb-Agar :
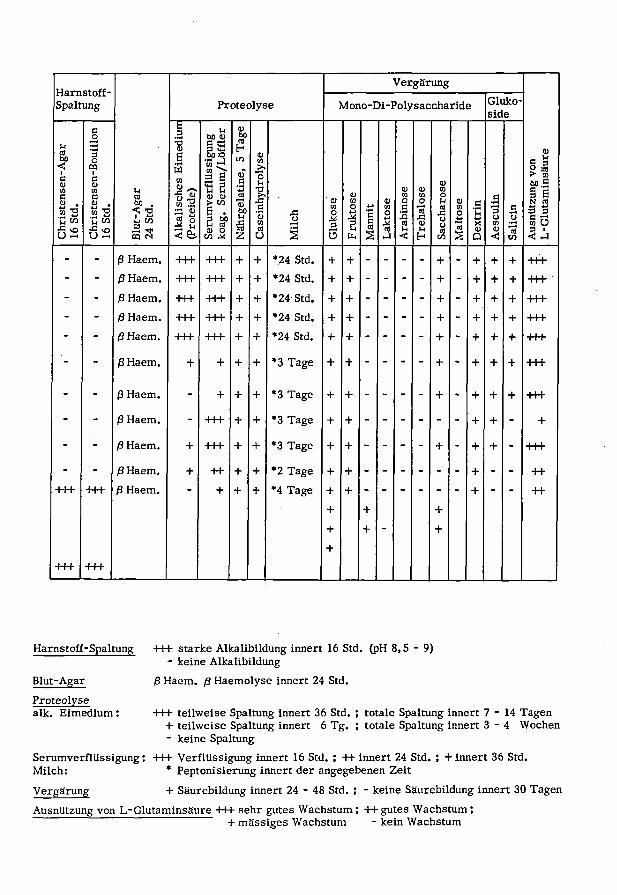
Harnstoff-
Spaltung
Vergärung
Proteolyse Mono-Di-PolysaccharideGluko¬
side
o3 u
bo <u
eu
bo
Agar Bouill Eimed ssigun /Löffl 0)CO
eu
ß 39 a
sen- sen-i-i
CO 3 ß tine dro <D eu CO ung\
minsf5 eu ^ CO CO o
8) CO"° Ti J2 <u VI (!) O o u 9) n cd
S v. tt-d < -o "H Q) fi.
bo .y _eVi ü
•t-iVi C
cdcd CO 3 CS 3 3
Chri 16
St
Chri 16
St
Blut- 24
St
Alka (Prot Seru koag Nähr Case Mile Gluk Fruk cd Lakt Arab Treh Sacc Malt Dext Aesc Salic Ausn L-Gl- - ß Haem. +++ +++ + + *24 Std. + + - - - - + - + + + +H-
- - ß Haem. +++ +++ + + 24 Std. + + - - - - + - + + + +H-•
- - ß Haem. +++ +H- + + 24 Std. + + - - - - + - + + + +++
- - ß Haem. +++ +++ + + *24 Std. + + - - - - + - + + + -H+
- - ßHaem. -H+ -H+ + + *24 Std. + + - - - - + - + + + H+
- - ß Haem. + + + + *3 Tage + + - - - - + - + + + +++
- - ß Haem. - + + + *3 Tage + + - - - - + - + + + +H-
- - ß Haem. - +++ + + *3 Tage + + + + - +
- - ß Haem. + -H+ + + *3 Tage + + - - - - + - + + - +++
- - ß Haem. + ++ + + *2 Tage + + - - - - - - + - - -H-
+++ -H+ ß Haem. + + + *4 Tage +
+
+
+
+
+
+ -
+
+
+ -H-
+++ -H+
Harnstoff-Spaltung
Blut-Agar
Proteolyse
alk. Eimedium :
+++ starke Alkalibildung innert 16 Std. (pH 8,5-9)- keine Alkalibildung
ß Haem. ß Haemolyse innert 24 Std.
+++• teilweise Spaltung innert 36 Std. ; totale Spaltung innert 7-14 Tagen+ teilweise Spaltung innert 6 Tg. ; totale Spaltung innert 3-4 Wochen
- keine Spaltung
SerumverflUssigung: +++ Verflüssigung innert 16 Std. ; ++ innert 24 Std. ; + innert 36 Std.
Milch: *
Peptonisierung innert der angegebenen Zeit
Vergärung + Säurebildung innert 24 - 48 Std. ; - keine Säurebildung innert 30 Tagen
Ausnutzung von L-Glutaminsäure +++ sehr gutes Wachstum ; ++ gutes Wachstum ;
+ massiges Wachstum - kein Wachstum

IV
C18-45°
"96
++
0,5
--
--
4126
Bcereus
Bacillus
in
C18-50°
"
48
++
++
0,5
+-
--
4061
Bcereus
Bacillus
II
C18-40°
C18-40°
"
72
"
72
++
+++
+++
0.9
0,9
-++
++
++
4034
Bth
urin
giensis
var.
thuringiensis
Bacillus
4058/c
Bthuringiensis
var.
thur
ingi
ensi
sBacillus
I
C10-40°
C10-40°
C10-40°
C10-40°
C10-40°
"
24
"
24
"
24
"
24
St.
24
-H+
+++
+++
+++
-H+
1,5-1,8
1,5-1,8
1,5-1,8
1,5-1,8
1,5-1,8
+++++
+++++
+++++
4156/e
BAltdorf
Stamm
cereus
Bacillus
4156/d
BAltdorf
Stamm
cereus
Bacillus
4156/c
BAltdorf
Stamm
cereus
Bacillus
4156/b
BAltdorf
Stamm
cereus
Bacillus
4156/a
BAltdorf
Stamm
cereus
Bacillus
SalicinAesculinSaccharose
Eint
eilu
ngGruppen-
Wachstum
fürTemperaturgrenzen) Std. ... (nach Milch
von
PeptonisierungSerum-Verflüssigung Lecithinase-Reaktionmm in
Phosphatase-ReaktionEimedium
alk. auf Proteolyse
Vergärung
Stämme
)wurden.
untersucht
serologisch
auch
die
einbezogen,
Stämme
die
nur
wurden
Uebersicht
diese
(In
Stämme.
thuringiensis
Bacillus
und
cereus
Bacillus
untersuchten
der
Merkmalen
physiologischen
und
biochemischen
den
in
Unterschiede
die
über
Uebersicht
3Tab.

m. IMMUNOBIOLOGISCHE EIGENSCHAFTEN
1. Material und Methoden
a) Stämme
Bei diesen Untersuchungen wurden folgende Organismen geprüft :
Bacillus cereus Stamm Altdorf B 4156/aBacillus cereus Stamm Altdorf B 4156/bBacillus cereus Stamm Altdorf B 4156/cBacillus cereus Stamm Altdorf B 4156/dBacillus cereus Stamm Altdorf B 4156/eBacillus thuringiensis var. thuringiensis B 4058/cBacillus thuringiensis var. thuringiensis B 4034
Bacillus cereus B 4126
Bacillus cereus B 4061
b) Herstellung des Immunserums
Herstellung der Muttersuspensionen
Da bei den zu prüfenden Stämmen die beweglichen Zellen nur je ca. 50 % der Ge¬
samtpopulation ausmachten, wurde mit allen Stämmen eine künstliche Selektion der
begeisselten Formen durchgeführt. Hierzu wurde eine im Labor leicht herzustellen¬
de, einfache Einrichtung konstruiert, die auf dem Prinzip der Wanderung der Mi¬
kroorganismen von halbstarrem in flüssiges Medium beruht.
Ein selbst hergestelltes U-Röhrchen (Abb. 18) von 5 mm
0 wurde in einen 200-ml-Breithals-Erlenmeyerkolben ge¬
stellt, dieser bis ca. 1 cm über das untere Ende des U-
Röhrchens mit Schwärmagar beschickt (Schwärmagar =
wie gew. Nähragar, jedoch mit 0,4 % Agar) und autokla-
viert. Nach dem Erstarren wurden 10 ml Nährbouillon
über die Agarschicht im Erlenmeyerkolben pipettiertund gleichzeitig das Niveau im U-Röhrchen zur Verhin¬
derung einer Kommunikation ausgeglichen. Die Impfungerfolgte durch die obere Oeffnung des U-Röhrchens,
worauf bei 37° C bebrütet wurde.
Die beweglichen Mikroorganismen wuchsen durch den
Agar in die Nährbouillon, aus der dann weitergeimpft Abb. 18
wurde. (Abb. 19) Die ersten beweglichen Zellen er¬
reichten die Nährbouillon nach ca. 8-14 Stunden. Durch 3 solche Passagen konnte
die Beweglichkeit aller untersuchten Stämme praktisch auf 100 % erhöht werden.
Bei der Durchführung dieser Selektion muss darauf geachtet werden, dass die Wei-
terimpfung früh genug erfolgt, da die Beweglichkeit mit zunehmendem Alter stark
abnimmt.
Die selektionierten Stämme wurden in 100 ml Nährbouillon in 1-Liter-Erlenmeyer-kolben übergeimpft, 7 Std. bei 30° C auf Schüttelmaschinen mit 100 Ausschlägenpro Minute bebrütet, dann mit 5 Vol. °/oo Formalin versetzt und bei +2° C im
Kühlschrank aufbewahrt.
Dieses Material stellte die Muttersuspensionen für die Immunisierung der Kanin¬
chen, sowie für die Durchführung der Agglutinations-Teste dar. Die Konzentration
betrug ca. 2 x 10^ vegetative Zellen pro cc.
25

beimpft unbeimpft
Abb. 19
Herstellung der lebenden Suspensionen
Fur das Sattigungs-Verfahren wurden lebende Organismen benützt, die in Kolle-
Schalen auf Agar bei 37° C gezüchtet und nach 24 Std. mit physiologischem NaCl ab¬
geschwemmt wurden.
Immunisierung
Vor der Immunisierung wurden die Kaninchen 3 Wochen in Quarantäne gehalten;ferner wurden Blutproben aller Kaninchen mit den zu untersuchenden Bakterienstäm-
men getestet, um eine eventuelle, bereits auf naturlichem Weg erworbene Immuni¬
tät festzustellen. Diese Teste fielen alle negativ aus.
Die Antigen-Applikation erfolgte in allen Versuchen gleich. Pro Stamm wurden 2 Ka¬
ninchen immunisiert (mit Ausnahme der Bacillus cereus Altdorf Stämme B 4156/dund B 4156/e, mit denen nur je 1 Kaninchen geimpft wurde). Die erste Injektion von
0,5 cc wurde subcutan und die vier folgenden intraperitoneal verabreicht, mit Ab¬
ständen von 4-5 Tagen und steigender Dosis : 0, 5 - 1 - 2 und 4 cc.
Um die Antikörper-Produktion zu verfolgen, wurden wahrend der Impfperiode 2 mal
Blutproben genommen und dabei ein ansteigender Agglutinations-Titer festgestellt.5 Tage nach der letzten Injektion wurden von jedem Kaninchen aus der Ohrvene ca.
80 cc Blut entnommen, das Serum 2-cc-weise in Reagenzröhrchen abgefüllt und bei- 30° C aufbewahrt.
c) Agglutinations-Technik
Der Agglutinations-Titer wurde nach dem üblichen Verdünnungsverfahren (1/10 bis
1/25*600) mit anschliessender Bebrutung (2 Std. bei 37° C) festgestellt. Als Agglu¬tinations-Titer galt die letzte Verdünnung mit totaler Agglutination. (Totale Agglu¬tination = positive Agglutination : das Agglutinât ist eine flockige Masse am Boden
des Fläschchens, die darüber liegende Flüssigkeit ist ganz klar. Teil-Agglutination:das Agglutinât ist weniger ausgeprägt in einer getrübten Flüssigkeit. NegativeAgglutination : keine Veränderung).
d) Sättigungs-Technik
Wenn das gleiche Serum verschiedene Stämme agglutiniert, besitzen diese minde¬
stens einen gemeinsamen Antigen-Faktor. Um festzustellen, ob die einzelnen Stäm¬
me ausser dem gemeinsamen Faktor (oder den gemeinsamen Faktoren) noch einen
26

andern, nicht gemeinsamen Faktor haben, wurde das Sättigungsverfahren von
DeBARJAC und BONNEFOI (1962) angewandt, und zwar nach folgendem Schema :
Hypothese : Stamm A besitzt 2 Faktoren : a) und b)
Serum anti-A
+ Suspension B
Agglutination+ Suspension A
keine zusätzliche Agglutination :
Stamm B besitzt beide Faktoren
a) und b) (d. h. Stamm A besitzt
keine bei Stamm B nicht vorhan¬
denen Faktoren)
zusätzliche Agglutination :
Stamm B besitzt nur einen
gemeinsamen Faktor (b)
Serum anti-B
+ Suspension A
Agglutination+ Suspension B
keine zusätzliche Agglutination :
Stamm B besitzt nur einen
Faktor (b)
zusätzliche Agglutination :
Stamm B besitzt einen bei
Stamm A nicht vorhande¬
nen dritten Faktor (c)
2. Versuchsergebnisse
Die Resultate der serologischen Untersuchungen sind in Tab. 4 zusammengefasst. Dar¬
aus wurden folgende Schlüsse gezogen :
Alle Bacillus cereus Altdorf Stämme zeigten bei den kreuzweise durchgeführten Agglu¬tinationstesten positive Reaktion. Mit dem Sättigungsverfahren wurde bei keinem der
5 Stämme ein bei den übrigen Stämmen nicht vorhandener serologischer Faktor gefun¬den. Folglich sind die Bacillus cereus Altdorf Stämme, die aus unterschiedlichen Wirts¬
individuen der gleichen Gattung isoliert wurden, in bezug auf die untersuchten antigenenEigenschaften völlig gleich und wurden deshalb in der zusammenfassenden Tabelle der
gleichen serologischen Gruppe, nämlich I, zugeteilt.
Die zwei untersuchten Bacillus thuringiensis Stämme zeigten keine Unterschiede in
ihren antigenen Eigenschaften, obwohl sie aus unterschiedlichen Wirtsgattungen, näm¬
lich Ephestia und Melolontha isoliert waren. Gegenüber Bacillus cereus Stamm Altdorf
fielen die Teste negativ aus. Die zwei Bacillus thuringiensis Stämme wurden deshalb
der serologischen Gruppe II zugeteilt.
27

Die Agglutinations-Teste mit Bacillus cereus Stamm B 4061 und Stamm B 4126 fielen
gegenüber allen Bacillus cereus Altdorf Stämmen und Bacillus thuringiensis Stämmen
negativ aus. Auch unter sich zeigten die Bacillus cereus Stämme B 4061 und B 4126
keine Verwandtschaft in bezug auf die untersuchte Antigen-, bzw. Partialantigenstruk-tur, weshalb sie in zwei verschiedene Gruppen, nämlich III und IV klassiert wurden.
Die serologische Gruppeneinteilung der geprüften Mikroorganismen wird von den bio¬
chemischen Eigenschaften derselben Gruppen weitgehend gestützt. D. h., Mikroorga¬nismen, die zur selben serologischen Gruppe gehören, zeigen auch weitgehende Ueber-
einstimmung in bezug auf ihren biochemischen Charakter.
SYSTEMATIK
Auf Grund der morphologischen, physiologischen und immunobiologischen Untersuchun¬
gen wurden die neu isolierten Stämme als Bacillus cereus identifiziert. Als Grundlagedieser Einordnung diente der in BERGEY's Manual VII. Aufl. (1957) angewandte Bestim-
mungsschlüssel von GORDON & CLARK (1946) und SMITH, GORDON & CLARK (1952),

B) ZUECHTUNG
1. Problemstellung
Dieses Kapitel befasst sich mit der Abklärung der optimalen Sporulationsbedingungen für
Bacillus cereus Stamm Altdorf und insbesondere mit der Zusammenstellung eines geeignetenMediums für die Massenherstellung eines Sporenpräparates zur Prüfung als allfälliges biolo¬
gisches Schädlingsbekämpfungsmittel in Labor- und Freilandversuchen.
2. Material und Methoden
a) ZUchtungsgefässe
Für SchUttelkulturen wurden 100 ml und 1000 ml Erlenmeyerkolben mit 25, resp. 100
ml Nährlösung, und rotierende Kolben mit 250 ml Nährlösung verwendet. Die Gärtank¬
kulturen wurden mit einem New Brunswick Apparat (Modell FS 307) in 7-Liter-Pyrex-
Glasgefässen mit 4 - 5 lt Nährlösung und in einem Bertrams 150-Liter-Gärtank (mit
Zentrifugal-TurborUhrer mit ca. 400 UD/Min., ringförmigem Luftverteiler und Prall¬
platte) mit 100 lt Nährlösung gezüchtet.
b) Medien
Medium 1) : Kartoffelextrakt mit und ohne 0,5 % Glukose
Ca. 150 g geschälte Kartoffeln wurden in 1000 ml Leitungswasser 10 Minuten bei 120°C
autoklaviert, anschliessend zu einem Brei zerstossen, durch Watte filtriert und mit
H20 dest. wieder auf 1000 ml aufgefüllt. pH 7.
Medium 2) : Leberbouillon + Kartoffelextrakt + 0,5% Glukose
50 g Schweineleber wurden in 1000 ml Leitungswasser 10 Minuten bei 120° C autokla¬
viert, filtriert und mit Kartoffelextrakt im Verhältnis 1 : 4 gemischt. pH 7.
Medium 3) : Nährbouillon
Fleischextrakt (Oxo Lab Lemco) 10 g; Pepton (Bacto) 10 g; NaCl 5 g; H2O dest. ad
1000 ml. pH 7.
Medium 4) : Medium nach LEMOIGNE et al. (1956) (für die Züchtung von Bacillus thu-
ringiensis var. thuringiensis)
KH2PC>4 6,8g; MgS04.7H20 0,123 g ; MnS04.4H2<D 0,00223 g ; ZnS04.7H20 0,014 g;
Fe2(S04)3 0,020 g; CaCl2.4H20 0,183 g; Pepton 7,5 g; Saccharose 20 g ; H20 dest.
ad 1000 ml. pH 7.
Medium 5 : Medium nach WILLE und WIKEN (1953) (für die Züchtung eines sporen¬
bildenden, nicht näher beschriebenen Bacillus A^)
Glukose 2 g; L(+)Glutaminsäure 4 g; DL-Cystin 0,06 g ; K2HP04 0,5 g; KH2P04
0,5 g; MgSO4.7H2O0,2g; NaCl 0,01g; MnS04.4H20 0,01 g; FeS04.7H20 0,01 g;
Ca(H2P04)2.H20 2 cc bei Zimmertemperatur gesättigter Lösung; Aneurin 0,5 mg;
(+)Biotin25y;H2Odest. ad 1000 ml. pH 7.
Medium 6) : Modifiziertes Medium von WILLE und WIKEN (1953)
Glukose 2 g; L(+)Glutaminsäure 4 g; DL-Cystin 0,06 g ; Aneurin 0,5 mg; (+)Biotin25 Y ; Kartoffelextrakt 250 ml; H20 dest. ad 1000 ml. pH 7.
29

Medium 7) : Medium nach HOWIE und CRUICKSHANK (1940) (für die Züchtung von
Bacillus cereus)
Natrium-Asparaginat 1 g; Glukose 1 g; MnS04.7H20 0,2 g; NH4H2P04 1,5 g; CaCl2
0,1 g; KCl 0,1 g; FeCl3 Spur; Agar 20 g; H20 dest. ad 1000 ml. pH 7.
Medium 8) : Medium nach YOUNG and FITZ-TAMES (1959) (für die Züchtung von Bacil¬
lus thuringiensis var. alesti)
Grundmedium : 40 % Beef Papain Digest Bouillon (Herstellung des Papain Digest nach
ASHESHOV (1941); 0,5 % Proteosepepton 2 (DIFCO); 0,5 % Proteosepepton 3 (DIFCO);dieses Grundmedium wurde im Verhältnis 1 : 4 mit.Medium,nach GRELET (1951) in
H20 dest. verdünnt pH 6,5.
Medium nach GRELET für die Züchtung von Bacillus megaterium : Glukose
45 g; KNO3 10 g ; K2S04 0,17 g; KH2P04 6, 8 g; MgS04> 7H20 0,123 g; MnS04.4H20
0,00223 g; Fe2(S04)3 0,02 g; ZnS04. 7H20 0,0144 g ; CaCl2.4H20 0,183 g; KOH
0,330 g ; H20 dest. ad 1000 ml. pH 7.
Medium 9) : Glukose-Glutaminsäure-Medium, selbst zusammengestellt für die Züch¬
tung der Bacillus cereus Altdorf Stämme
Hefeextrakt 0,5 g; Glukose 2 g; L(+)Glutaminsäure 4 g; Kartoffelextrakt 250 ml;
H2O dest. ad 1000 ml. pH 7.
Medium 10) : wie Medium 9), jedoch ohne Kartoffelextrakt
Medium 11): Glukose-Glutaminsäure-Salz-Medium, selbst zusammengestellt für die
Züchtung der Bacillus cereus Altdorf Stämme .
Glukose 6 g ; L(+)Glutaminsäure 6 g ; Hefeextrakt 0,5g; Proteosepepton 3 (DIFCO)0,4 g; MgS04.7H20 0,012 g; ZnS04. 7H20 0,014 g; CaCl2.4H20 0,090 g; KH2P04
0,680 g; Fe2(S04)3 Spur; MnS04.4H20 0,001 g; Kartoffelextrakt 50 ml. Für festen
Nährboden 1,5 - 2 % Agar. H20 dest. ad 1000 ml. pH 6,2.
c) Bestimmung der Wasserstoffionenkonzentration
Alle pH-Werte wurden mit einer Glaselektrode gemessen.
d) Sterilisation der Medien
Die Medien wurden im Autoklav bei 120° C und 1 atü während 20 Minuten sterilisiert.
Glukose und Silikon-Antifoam wurden separat sterilisiert und unter sterilen Bedingun¬
gen zum Medium gegeben.
e) Impfmaterial
Das Impfmaterial wurde für alle Sporulationskulturen auf Medium 3) hergestellt. Für
die Beimpfung der 100 und 1000 ml Erlenmeyerkolben, sowie des 7-Liter-New-Bruns-
wick-Fermenters wurde 1 Vorkultur, für den 150-Liter-Gärtank 2 Vorkulturen zube¬
reitet.
1. Vorkultur. Eine Oese voll Kultur von Schrägagarröhrchen wurde auf Nährbouillon
in Schüttelkolben abgeimpft und während 18 Std. bei 30° C bebrütet. Die 100 ml Erlen¬
meyerkolben wurden mit 0,05 ml, die 1000 ml Erlenmeyerkolben mit 0,25 ml und die
7-Liter-Fermenter mit 50 ml dieser Vorkultur beimpft. (Konzentration der Vorkultur :
108 veg. Zellen/ml).
2. Vorkultur. Der so beimpfte New Brunswick Fermenter (mit 5 Liter Inhalt) wur¬
de während 18 Std. bei 30° C mit 2 Liter Luft/Min. belüftet und mit 300 UD/Min. ge¬
rührt. Mit 4 Liter dieser zweiten Vorkultur wurde hierauf der Bertrams-Gärtank beimpft.
30

Tab. 4 Resultate der Agglutinationsteste
Muttersuspension H
Serum
Gruppen¬
einteilung
anti B 4156/a anti B 4156/b anti B 4156/c anti B 4156/d anti B 4156/e anti B 4058/c anti B 4034 anti B 4126 anti B 4061
Tier Nr. 1 Tier Nr. 2 Tier Nr. 1 Tier Nr. 2 Tier Nr. 1 Tier Nr. 2 Tier Nr. 1 Tier Nr. 1 Tier Nr. 1 Tier Nr. 2 Tier Nr. 1 Tier Nr. 2 Tier Nr. 1 Tier Nr. 2 Tier Nr. 1 Tier Nr. 2
Bac. cereus St. Altdorf
B 4156/a
Bac. cereus St. Altdorf
B 4156/b
Bac. cereus St. Altdorf
B 4156/c
Bac. cereus St. Altdorf
B 4156/d
Bac. cereus St. Altdorf
B 4156/e
1 : 9600
1 : 9600
1 :12800
1 : 9600
1 : 9600
1 : 9600
1 : 9600
1 : 12800
1 : 9600
1 : 9600
1 : 9600
1 : 9600
1 : 9600
1 : 9600
1 : 9600
1 : 6400
1 : 6400
1 : 9600
1 : 9600
1 : 9600
1 : 9600
1 : 9600
1 : 12800
1 : 9600
1 : 9600
1 : 9600
1 : 9600
1 : 12800
1 : 9600
1 : 9600
1 : 9600
1 : 9600
1 : 12800
1 : 9600
1 : 9600
1 : 9600
1 : 9600
1 : 12800
1 : 9600
1 : 12800
- - tiit -
|
-
-
i
i
i
i
i
I
Bac. thuringiensis var.
thuringiensis B 4058/c
Bac. thuringiensis var.
thuringiensis B 4034- -
: : :-
:-
1 : 6400
1 : 3200
1 : 6400
1 : 6400
1 : 3200
1 : 6400
1 : 3200
1 : 6400: :
- -
II
Bac. cereus B 4126 - - - - - - - - - - - - 1 : 3200 1 : 3200 - - III
Bac. cereus B 4061 - " - - - - - - - - - - - - 1 : 3200 1 : 3200 IV

f) Belüftung
Im Bertrams-Gärtank wurde eine durchschnittliche Belüftungsintensität von ca. 0,4-
0,5 Luftvolumen auf 1 Volumen Nährlösung pro Minute eingestellt. Im New-Brunswick-
Fermenter wurden 2 Liter Luft pro Minute durchgeleitet bei einer Ruhrgeschwindig-keit von 300 UD/Min. Bei dieser Einstellung wurde pro Liter und Stunde ein Sauerstoff-
Übergang von 30 mMol O2 gemessen.
g) Sauerstoffbestimmung
Die Bestimmung des Sauerstoffübergangs im New-Brunswick-Fermenter erfolgte nach
der Sulfit-Oxydations-Methode von COOPER et al. (1944). Mit einer vorher mit Stick¬
stoff begasten Pipette wurden jeweils 4-ml-Proben entnommen, blasenfrei in 50 ml
0,1 n Iodlösung gegeben und das restliche Sulfit iodometrisch titriert.
h) Schaumbekämpfung
Bei Beginn der Gärung und während der Sporulation erfolgt eine intensive Schaumbil¬
dung. Diese wurde mit Silikon-Antifoam in 1 : 100 Verdünnung mit H2O dest. unter¬
drückt. Bei der angegebenen Belüftungsintensität wurden im grossen Gärtank 1-2 Li¬
ter, im New-Brunswick-Fermenter 25 - 50 ml verdünntes Silikon insgesamt benötigt.
i) Glutaminsäurebestimmung
Die Bestimmungen wurden mit der DUnnschichtchromatographie-Technik unter Anwen¬
dung des eindimensionalen Trennverfahrens durchgeführt. Methodik und Auswertungmit Flächenvermessung s. STAHL (1962).
k) Glukosebestimmung
Die Glukose wurde nach der Methode von SUMMER (1925) bestimmt. Die durch die re¬
duzierende Gruppe der Glukose bewirkte Aenderung der optischen Dichte der 3,5-
Dinitrosalicylsäure wurde bei 546 m ji gemessen. Der Glukosegehalt der Proben wurde
von einer Eichkurve abgelesen.
1) Photometrische Messungen
Diese wurden mit einem EPPENDORF-Photometer in Glasküvetten mit 1 cm Lichtwegdurchgeführt.
m) Konzentrationsbestimmung
Die Zell- und Sporenbildung wurde in verschiedenen Zeitabständen in feuchten Präpa¬raten phasenkontrastmikroskopisch untersucht und die Konzentration mit einer PE -
TROFF-HAUSSERschen Zählkammer bestimmt. Bakterien-Suspensionen wurden mit
physiologischer Kochsalzlösung oder Formaldehydsalzlösung (0,9 % NaCl mit 1 % For¬
maldehydgehalt) so verdünnt, dass die Keimzahl pro grosses Quadrat zwischen 20 und
80 lag. Es wurden jeweils mindestens 6 grosse Quadrate in dreimaliger Wiederholungausgezählt. Die Unterschiede in der Keimzahl zwischen den drei Auszählungen lagenzwischen 5 und 10 %.
n) Abernten des Sporenmaterials
Das Sporulationsmedium wurde mit einer Christ-Zentrifuge Typ Junior II bei 9000 g
oder mit einer Westfalia-Zentrifuge Typ LVA 205 Trommel UD 1,2 x 104/Min. zen-
trifugiert, das Sediment bei Zimmertemperatur getrocknet und mit einem Mörser ho¬
mogenisiert.
31

3. Versuchsergebnisse
a) Vorversuche
Die erste Aufgabe war, ein Medium zu finden, auf dem Bacillus cereus Stamm Altdorf
nicht nur vegetativ wächst, sondern auch sporuliert. In Vorversuchen wurden deshalb
15 Medien, teils Komplexe, teils halb- oder vollsynthetische, getestet. Dabei erwie¬
sen sich Nährböden, die Kleieextrakt, Melasse, Casein-Totalhydrolysat oder Papain Di¬
gest enthalten, ftir die Sporulation als ungeeignet. Auf dem Medium nach HOWIE and
CRUICKSHANK (1940) wurde lediglich bei Kultivierung auf festem Nährboden eine Spo¬renproduktion erreicht ; in Submerskulturen war sie total gehemmt.
Auf 8 der 15 Medien kam Bacillus cereus Stamm Altdorf mehr oder weniger gut zur
Sporulation und zwar sowohl auf festem Nährboden wie in Submerskultivierung. Diese
8 Medien sind in Tab. 5 aufgeführt.
Eine gute Sporulation zeigte sich auf Medien, die Kartoffelextrakt, Glukose, Glutamin¬
säure und Hefeextrakt enthalten. Besonders günstig ist Kartoffelextrakt für die Sporen¬produktion. Auf dem synthetischen Medium nach WILLE und WIKEN (1953) wurde nur
eine sehr spärliche Sporulation beobachtet (s. Tab. 5) ; durch Zugabe von Kartoffelex¬
trakt konnte die Sporenkonzentration wesentlich erhöht werden. Eine sporulationsför-dernde Wirkung des Kartoffelextraktes wurde auch beim Medium 9) gegenüber Medium
10) festgestellt.
Bei Zugabe von 0,3 % Fleischextrakt (Oxo Lab Lemco) zu einem Sporulationsmedium(z.B. Medium 9), wurden die Einschlusskörper nicht ausgebildet, während die Sporu¬lation bei dieser Konzentration nicht gehemmt wurde.
Wie aus Tab. 5 hervorgeht, wurde in allen Medien bei belüfteter Kultivierung eine hö¬
here Sporulationsgeschwindigkeit als in Schüttelkulturen beobachtet. Dies bestätigt die
bekannte Tatsache, dass die Sporulationsgeschwindigkeit in beträchtlichem Masse von
der Sauerstoffsättigung des Mediums abhängt.
Medien, die für Bacillus thuringiensis und dessen Varietäten beschrieben wurden (Me¬dium 4 und 8), eignen sich nicht für die Sporenproduktion von Bacillus cereus Stamm
Altdorf. Auf dem Medium nach LEMOIGNE (1956) wächst Bacillus cereus Stamm Alt¬
dorf sehr gut vegetativ (s. Tab. 5), kommt jedoch nicht richtig zur Sporulation. Zu
Beginn der stationären Phase (d. h. also bei einem geeigneten Medium zur Zeit der Spo¬rulation), bilden sich im Sporangium mehrere runde refraktile Körperchen von einer
Grösse bis zu 1 u. Nach Auflösung des Sporangiums werden diese Körperchen freige¬setzt. In diesem Stadium sind sie mikroskopisch wegen ihrer Formähnlichkeit von Spo¬ren nicht leicht zu unterscheiden. Eine rasche und sichere Identifizierung ist jedochdadurch möglich, dass sie nicht keimungsfähig sind.
In einem Gegenversuch wurde interessanterweise festgestellt, dass Bacillus thuringien¬
sis var. thuringiensis auf dem für Bacillus cereus Stamm Altdorf sehr guten Sporula¬tionsmedium 11) (Glukose-Glutaminsäure-Salz) keine bis sehr wenige Sporen, dafür
massenweise die beschriebenen Körperchen bildet.
b) Sporulation auf einem selbst zusammengestellten Glukose-Glutaminsäure-Salz-Medium
(Medium 11)
Wie aus den Voruntersuchungen (Tab. 5) hervorgeht, wurde auf Medium 9) im New
Brunswick Fermenter eine 80 %-ige Sporulation bei einer Ausbeute von 900 mg Trok-
kensubstanz pro Liter erzielt. Um dieses Resultat noch zu verbessern, wurde Medium
9) zu Medium 11) ergänzt (s. Tab. 6>. Dieses enthält Glukose als Kohlenstoffquelle,Glutaminsäure, Pepton und Hefextrakt als Stickstoff- und Wuchsstoffquellen, ferner
Phosphat, Mineralsalze und Kartoffelextrakt.
32

Auf diesem Medium wurden die die Sporulation beeinflussenden physikalischen Fakto¬
ren, nämlich Wasserstoffionenkonzentration, Temperatur und Belüftung weiter geprüft.Ferner wurde neben der Sporenproduktion die Glukose- und Glutaminsäureaufnahme
gemessen.
Wasserstoffionenkonzentration
Wie aus Tab. 7 hervorgeht, beeinflusst das Anfangs-pH des Mediums die Endausbeute
ganz beträchtlich. Das optimale Anfangs-pH für Wachstum, Sporulation und Ausbeute
liegt zwischen 6 und 7. Die beste Sporenproduktion wurde bei Anfangs-pH 6,5 erzielt.
Die pH-Grenzen liegen für Wachstum, Sporulation und Sporenkeimung gleich, nämlich
bei pH 4,5 und pH 10.
Temperatur
Die optimale Temperatur für die Sporenproduktion liegt bei 30° C (Tab. 8). Die Tem¬
peraturgrenzen liegen für Wachstum und Sporulation bei 10° C und 40° C (s. Tab. 3).
Belüftung und Schaumbekämpfung
Im New Brunswick Fermenter wurde die vollständige Sporulation und Freisetzung der
Sporen nach 72 Std., im Bertrams Fermenter bereits nach 48 Std. erreicht, wobei die
Trockenausbeute im Bertrams Fermenter um 900 mg/L höher lag. Diese schnellere
und bessere Entwicklung der Kultur wird auf die günstigeren BelUftungs- und Durch¬
mischungsverhältnisse im grösseren Gärtank zurückgeführt. Beim New Brunswick
System wurde eine positive Beziehung zwischen Tourenzahl des RUhrers und Sauerstoff¬
absorption festgestellt. (Tab. 9). Bei Durchlüftung mit 2 L Luft/Min. und einer Tou¬
renzahl von 300 UD/Min. wurde ein höherer C^-Uebergang gemessen als bei einer
Durchlüftung mit 4 L/Min. und einer Tourenzahl von 200 UD/Min.
Das Schäumen liess sich mit Silikon-Antifoam unterdrücken. Es wurde jedoch die
Beobachtung gemacht, dass sich die Silikonzugabe auf die Sporenausbeute ungünstigauswirkte (Tab. 9). Es ist anzunehmen, dass dies auf eine durch Silikonzugabe verur¬
sachte Verminderung des O^-Uebergangs im Medium zurückzuführen ist,
Glukose-Aufnahme
Bei einer Anfangskonzentration von 6 mg/ml wurde die Glukose innert 24 Std. praktischtotal aufgenommen. 74 - 82 % wurden während des Wachstums innert 12-14 Std.
(s. Abb. 20 - 21) und Tab. 10), also vor Anfang der Sporulation ausgenützt. Dieser
Prozess wurde begleitet von der Produktion organischer Säuren, deren Gipfelpunktnach 8 Std. bei pH 5,2 lag. Die Bedeutung der Glukose als C-Quelle für das Wachstum
(vegetative Vermehrung) ist besonders bei Weglassen aus Medium 11) augenfällig(Tab. 6). Das Weglassen der Glukose verursachte eine starke Verminderung der
Trockenausbeute, ohne jedoch den prozentualen Sporenanteil wesentlich zu beeinflus¬
sen.
Glutaminsäure-Aufnahme
Der zeitliche Verlauf der Ausnutzung von Glutaminsäure als N-Quelle ist grundsätzlichverschieden von dem der Glukoseaufnahme. Nach 18 Std. Bebrütung (s. Tab. 10 und
Abb. 20), also bei Einsetzen der Massensporulation (intrazellulär), wurde die Auf¬
nahme beschleunigt. Nach 24 Std. waren 16 %, nach 48 Std. 66 % und nach 72 Std.
100 % der beim Start vorhandenen 6 mg/ml Glutaminsäure verbraucht. Bei Weglassender Glutaminsäure aus Medium 11) wird die Trockenausbeute nur wenig vermindert,wohl aber der prozentuale Sporenanteil (Tab. 6). Daraus wurde die Folgerung gezogen,dass Glutaminsäure weniger für das vegetative Wachstum von Bacillus cereus Stamm
Altdorf als vielmehr zur Erreichung einer maximalen Sporulation erforderlich ist.
33

Verlauf der Sporulation auf Medium 11)
In der logarithmischen Wachstumsphase wurde eine zunehmende Kettenformation mit
bis zu maximal 6-7 vegetativen Zellen pro Kette beobachtet. Diese Kettenbildung er¬
reicht ihren Maximalwert, sobald die ersten Sporen ausgebildet werden (nach 12 - 14
Stunden). Mit zunehmender Sporulation brechen die Ketten auf, so dass nach 24 Std.die durchschnittliche Länge bei 2-3 Zellen liegt. Eine weitere Abnahme läuft parallelmit der Freisetzung der Sporen und der Lysis der Sporangien. (S. Abb. 22).
Die beste Sporulation aller Versuche wurde auf dem vollständigen Glukose-Glutamin¬
säure-Salz-Medium (Medium 11) erzielt (s. Tab. 6). Nach 72 Std. BebrUtung im New
Brunswick Fermenter, resp. 48 Std. im Bertrams Fermenter sporulierten 95 % der
Zellen, bei einer Konzentration von 2 x 10.„
Sporen pro Gramm Trockensubstanz im
New Brunswick Fermenter, resp. 2, 8 x 10 Sporen pro Gramm im Bertrams Fermen¬
ter.
Abb. 20
mg/ml * L (+) Glutaminsäure
X Glukose
5 -
clo
4 -
t-i4-»
e
n 3co
«-
72 Std.
Inkubationszeit
34

oH3
ön>I—'
H—
n>
N
w4cl
<:
PNs>
esETo
5-
o6CD,
trOHc
a
7-•
SporangienderLysis%1t
Sporulation%X*
KetteproZellenveg.AnzahlDurchschnittliche°°
22Abb.
Std.242016
11T
onszeitInkubati
128
48
T
»o—o.
100
80
60
40
20
6§
UÄ4-1T335-1
u
S*o
co
to
boMtu
21Abb.

%100
=Zellen)
vegetative
und
Sporangien
Sporen
intrazelluläre
%:
Nenner
(Sporen,
Elemente
zellulären
der
Gesamtzahl
die
auf
berechnet
|Sporen
freie
%:
Zähler
minsäure
850
xlO7
160/10
50/20
5/50
0/10
106
5x
10/30
5/20
0/10
0/5
Gluta¬
+Glukose
+Hefeextrakt
900
107
x2
70/10
60/20
5/60
0/20
106
6x
25/25
5/30
0/15
0/5
Kartoffelextrakt
+minsäure
Gluta¬
+Glukose
+Hefeextrakt
700
107
x2
70/10
55/20
5/60
0/20
106
6x
30/20
5/40
0/20
0/10
Glukose
0,5%
+
Kartoffelextrakt
+Leberbouillon
500
106
x2
70/10
60/20
10/50
0/20
9x.l05
40/10
10/40
0/30
0/10
Glukose
0,5%
+Kartoffelextrakt
3500
103
x5
25/5
1/12
0/4
0/0
104
3x
5/20
0/10
0/0
0/0
Lemoigne
nach
Medium
-10
3x
35/10
0/10
0/5
0/0
103
2x
0/5
0/5
0/0
0/0
Fitz-James
u.
Young
nach
Medium
800
107
x3
60/20
50/30
5/60
0/25
106
8x
30/10
10/30
0/20
0/0
Kartoffelextrakt
-1-
Wikén
u.
Wille
nach
Medium
300
104
x3,4
10/2
10/2
0/5
0/0
104
Ix
5/5
0/5
0/0
0/0
*Wikén
u.
Wille
nach
Medium
mgTS/L
Std.
96
nach
ren/ccSpo-
+freie
intrazell.
Std.
96
Std.
48
Std.
24
Std.
16
Std.
96
nach
ren/cc
+freieSpo-
intrazell.
Std.
96
Std.
48
Std.
24
Std.
16
L)
5Brunswick
(New
Gärtank-Kultur
in
Sporulation
ml)
100
(Erlenm.
SchUttelkult.
in
Sporulation
Nährboden
Sporulationsmedien
verschiedenen
mit
Voruntersuchungen
der
Resultate
5Tab.

Tab. 6 Sporulation auf Medium 11) in Gärtankkulturen
Nährboden
New Brunswick Fermenter mit 5 L Medium
16 Std. 24 Std. 48 Std. 72 Std. gTS/L Anzahl Sporen/g TS
Medium 11) vollständig
Medium 11) ohne Glukose
Medium 11) ohne Glutaminsäure
Medium 11) ohne Hefeextrakt
Medium 11) ohne Kartoffelextrakt
Medium 11) ohne Pepton
* 0/50
0/30
0/20
0/50
0/50
0/50
10/70
5/70
5/50
10/60
10/70
10/70
80/5
50/20
20/40
70/10
70/10
70/10
90/5
60/20
30/35
80/5
80/5
80/5
1.9
1,2
1.7
1.8
1.8
1,8
2 x 1012
3 x 1011
2x IQ9
2 x 1012
2 x 1012
2 x 1012
Medium 11) vollständig
Bertrams Gärtank mit 100 L Medium
0/50 20/60 90/5 2,8 2,8x 1012
* Zähler : % freie Sporen berechnet auf die Gesamtzahl der zellulären Elemente
Nenner : % intrazelluläre Sporen I (Sporen, Sporangien und vegetative Zellen) = 100 %

Tab. 7 Einfluss des pH auf die Sporulation von vegetativen Zellen und auf die Sporenkeimung.
Geprüft auf Medium 11) in 1000 ml Erlenmeyerkolben mit 100 ml Medium in Schtlttel-
kulturen.
Anfangs-pH
Impfmaterial vegetative ZellenImpfmaterialreine Sporen
24 Std. 48 Std. 72 Std.mg TS/100 ml
Medium nach
72 Std.
12 Std. 24 Std.
4,0 * 0 0 0 0 neg. neg.
4,5 0 0 0 0 neg. neg.
5,0 10 40 40 55 pos. pos.
5,5 10 50 50 60•
6,0 30 80 80 92r
6,5 30 80 85 95t
7,0 30 80 80 90•
7,5 15 50 70 83t
8,0 0 15 15 80•
8,5 0 10 10 59•
9,0 0 5 5 46i
9,5 0 5 5 20i
10,0 0 0 0 0 neg. neg.
* Angabe in % Sporulation (intrazelluläre + freie Sporen) auf die Gesamtzahl der zellulären Ele¬
mente (vegetative Zellen, Sporangien, freie Sporen) = 100 %.
neg. = keine Sporenkeimung, pos. = Sporenkeimung.

Tab. 8 Einfluss der Temperatur auf die Sporulation (geprüft auf Medium 11)
Temperatur°C
Wachstum und Sporulati on
Fester Nährboden (Sehrägagar-Röhrchen)
Submerskultur
New Brunswick Gärtank
mit 5 L Medium
24 Std. 48 Std. 72 Std. 96 Std. 1 Woche 24 Std. 48 Std. 72 Std.
0 * 0 0 0 0 0 - - -
3 0 0 0 0 0 - - -
6 0 0 0 0 0 - - -
9 0 0 0 0 0 - - -
12 W W W 50 70 - - -
15 W W W 50 70 - - -
18 W W 40 80 80 - - -
20 W W 50 80 90 - - -
25 w 50 70 90 90 60 80 90
30 20 80 90 95 95 80 95 95
35 20 80 90 95 95 80 95 95
*angegeben in % Sporulation (intrazelluläre + freie Sporen) auf die Gesamtzahl der zellu¬
lären Elemente (vegetative Zellen, Sporangien, freie Sporen) = 100 %.
W = vegetatives Wachstum
- = nicht geprüft

Tab. 9 SauerstoffUbergang in New Brunswick Fermenter mit 5 Liter lonenaustauschwasser mit
und ohne Silikonzugabe
UD/Min. Luft
L/Min.
O2 in mMol pro Liter und Stunde
Probeentnahme
nach Minuten
Belüftungohne Silikon 50 ml Silikon 250 ml Silikon
300
ii
il
il
2
II
II
28
35
32
30
20
29
30
28
12
21
24
22
10 Min.
20"
40"
60 "
200
II
II
II
4
II
tl
II
21
29
25
27
nicht geprüft nicht geprüft
10 Min.
20"
40"
60"
Tab. 10 Glukose- und Glutaminsäure-Aufnahme, sowie pH-Verlauf auf Medium 11).
BelUftungskultur in New Brunswick Fermenter.
Zeit in Std. GlukosepH
L (+) Glutaminsäure
ab Impfung mg/ml mg/ml
0 (Start) 6,0 6,20 6,00
6 5,6 5,35 6,00
8 4,6 5,20 5,80
10 3,5 5,55 5,80
12 1,6 6,50 5,80
14 1.1 7,00 5,75
16 0,6 7,25 5,70
18 0,4 7,45 5,70
24 0,25 7,70 5,00
48 0,19 8,00 2,00
72 0,15 8,20 Spuren

OWIRKSAMKEIT VON SPORENPRÄPARATEN
I. IM LABORATORIUM DURCHGEFÜHRTE INFEKTIONSVERSUCHE
Diese Versuche hatten zum Zweck, die Wirksamkeit von Bacillus cereus Stamm Altdorf
gegenüber Melolontha-Larven in verschiedenen Entwicklungsstadien, sowie gegenüber an¬
deren Insekten zu prüfen.
1. Material und Methoden
a) Herkunft und Haltung der Larven
Die Melolontha - Larven wurden in mehreren Grabungen in Altdorf, Kt. Uri, aus Frei-
landpopulationen im ersten, zweiten und dritten Entwicklungsstadium gesammelt.Die labormassige Haltung und Fütterung der Larven erfolgte nach der Methode von
HORBER (1959).
Während der Haltung aufgetretene Krankheiten, resp. identifizierte Krankheitserre-
srer :
Die angegebenen Prozentzahlen basieren auf 1000 Melolontha-Larven.
Mykosen - Metharrhizium anisopliae. Abgestorben ca. 1%.Beauveria tenella. Abgestorben ca. 10 %.
Aspergillus niger. Abgestorben ca. 0,2 %.
Bakteriosen - Maladie transparente HURPIN & VAGO (1958).
Abgestorben ca. 3 %.
Pseudomonas sp. ,„
. , „ m
Bacillus sp.abgestorben ca. 1.8 %
b) Stämme
Neben Bacillus cereus Stamm Altdorf wurden mit folgenden Vergleichsstämmen In¬
fektionsversuche durchgeführt :
Bacillus thuringiensis var. thuringiensis B 4034
Bacillus thuringiensis var. thuringiensis B 4058/cBacillus thuringiensis var. alesti B 4089
Bacillus thuringiensis var. sotto B 4084
c) Herstellung des Sporenmaterials
Das Sporenmaterial von Bacillus cereus Stamm Altdorf wurde auf Medium 11)
(s. Kapitel "Züchtung") hergestellt. Die Vergleichs stamme wurden auf gewöhnlichem
Agar in Kolle-Schalen gezüchtet. Nach 4 Tagen Bebrütung bei 30 C wurde der Agarmit steriler physiologischer Kochsalzlösung abgeschwemmt und diese Sporensuspen¬sion für die Infektionsversuche verwendet.
d) Verabreichung des Sporenmaterials und Kontrolle des Versuchs-Ablaufs
Das Sporenmaterial wurde peroral direkt durch MikrofUtterung nach MARTIGNONI &
STEINHAUS (1961) und indirekt durch Erdbeimpfung verabreicht (pro Zuchtdose wur¬
den 20 g sterile Erde -3x1 Std. bei 160° C sterilisiert - abgewogen und mit der
wässerigen Sporensuspension gut vermischt).Alle Infektionsversuche wurden durch parallel eingestellte Kontrollversuche über¬
wacht, wobei die entsprechende Kontrollgruppe je nach Art der Verabreichung des
Sporenmaterials mit der gleichen Menge sterilem, destilliertem H2O gefüttert, resp.
die Erde beimpft wurde. Alle Versuche wurden täglich kontrolliert und die abgestor¬benen Larven auf die Todesursache untersucht.
35

2. Versuchsergebnisse
Durch die Versuche mit Mikrofütterung wurde experimentell festgestellt, dass Bacilluscereus Stamm Altdorf fähig ist, nach der Invasion der Haemolvmphe eine Septicaemiezu verursachen Mit einer Dosis von 6 x 10^ Sporen/Larve wurde innerhalb 5-6 Tageneine Mortalität von 80 - 92 % erzielt ; bei allen Vergleichsstammen blieb die Mortalitätbei denselben Voraussetzungen unter 30 %. (S. Tab. 11).
Die Ergebnisse der Versuche mit Erdbeimpfung (s. Tab. 12) zeigen, dass der Infektions¬
vorgang, resp. die Inkubationszeit und Wirksamkeit von Bacillus cereus Stamm Altdorfwesentlich vom Entwicklungsstadium der Melolontha-Larven abhangt. Für Lj-Larvenwurde bei einer Konzentration von 3 x 10^ Sporen/Zuchtdose innerhalb 55 Tagen eine
Mortalität von 94 %, fur I_2-Larven bei der gleichen Konzentration innerhalb 80 Tagenvon 84 % und für L^-Larven bei einer Konzentration von 1,5 x 10 Sporen/Zuchtdoseinnerhalb 100 - 110 Tagen eine solche von 70 - 74 % erreicht. (Das erbte Absterben trat
nach ca. 3 Wochen auf. ) (Spatere Wiederholungen dieser Versuche (LÜTHI 1964, unpu-bhziert) ergaben allerdings auch bei den Kontrollgruppen Mortalitäten bis zu 50 %. )
3. Inf ekti on s ver s uche mit Bacillus cereus Stamm Altdorf gegen Rau¬
pen der grossen Wachsmotte (Galleria mellonella L. )
Das benutzte Sporenpraparat enthielt 500 mg Sporen auf 2,5 g Talkpulver als Trager¬stoff. Je 10 dm' Bienenwabe wurden mit 50 mg dieses Präparates mit einem selbst her¬
gestellten Gerat gleichmassig bestaubt und 50 Lj-Larven darauf ausgesetzt. Bei dieser
Dosierung wurde innerhalb 3 Wochen eine Mortalität von 82 % erzielt. (S. Abb. 23).
Abb. 23
Wabe zerstört Wabe grösstenteils unbeschädigt
4. Inf ekti on s ver suche mit Bacillus cereus Stamm Altdorf gegenRaupen des Kohlweisslings (Pieris brassicae L. )
Gegenüber Pieris brassicae L. erwies sich Bacillus cereus Stamm Altdorf als unwirk¬
sam. Bei einer Konzentration von 10? Sporen pro cm2 Blattflache wurden nur 20 %Mortalität erzielt.
36

Tab. 11 Ergebnisse der Infektionsversuche mit verschiedenen Mikroorganismen. Perorale Verabreichung des Sporenmaterials
durch MikrofUtterung. Versuchstemperatur : 23° C.
Anzahl infizierte
Engerlinge. Lar¬
venstadium.
Inoculum
pro
Larve
MortalitätVersuchsdauer
in Tagen
Kontrolle
innert
Tagen%
Anzahl Engerlinge/Stadium
Mortalität
%
Bacillus thuringiensis var.
25 L2
25 L2
25 L2
25 L2
25 L2
25 L2
25 L2
6x 105
6x 105
6x 105
6x 105
6x 105
6x 105
6x 105
4
4
6
6
6
5
6
28
12
28
24
80
92
84
20
25
25
20
25
20
25
25 L2
25 L2
25 L2
25 L2
25 L2
25 L2
25 L2
4
4
8
12
4
12
8
thuringiensis B 4058/c
Bacillus thuringiensis var.
thuringiensis B 4034
Bacillus thuringiensis var.
alesti B 4089
Bacillus thuringiensis var.
sotto B 4084
Bacillus cereus Stamm Altdorf B 4156
Bacillus cereus Stamm Altdorf B 4156
Bacillus cereus Stamm Altdorf B 4156

Tab. 12 Ergebnisse der Infektionsversuche mit Bacillus cereus Stamm Altdorf. Infektion der Erde der Zuchtdosen. Versuchs¬
temperatur : 22° C
Anzahl infizierte Engerlinge
Larvenstadium
Konzentration
pro Zuchtdose
Mortalität Versuchsdauer in Tagen Kontrolle
innert
Tagen% Anzahl Engerlinge - Stadium % Mortalität
50 L: 3 x 106 55 94 63 50 Lx 14
25 L2 3 x 106 80 84 85 25 L2 12
50 L2 1 x 107 78 80 86 50 L2 14
50 L2 1 xlO8 65 86 72 50 L2 12
50 L2 5 x 108 89 90 100 50 L2 20
50 L2 1 x 109 85 94 96 50 L2 16
50 L2 1 x 1010 85 90 92 50 L2 12
50 L3 1,5 x 1010 110 70 120 50 L2 12
50 L3 1,5 x 1010 100 74 112 50 L2 20

II. FREILANDVERSUCH
1. Problemstellung
Im Jahre 1963 wurde in Collonges (bei Evionnaz, Kt. VS) in Zusammenarbeit mit der Eidr
genössischen Landwirtschaftlichen Versuchsanstalt *, Lausanne, ein Feldversuch durchge¬führt, um
a) die Wirksamkeit des selbsthergestellten Bacillus cereus Stamm Altdorf-Präparates unter
natürlichen Verhältnissen,
b) die Verwendbarkeit des Präparates in Form einer Suspension zu prüfen. Im Sinne der
Aufgabestellung wurde bei diesem ersten Feldversuch auf mehrfache Wiederholung mit
verschiedenen Konzentrationen verzichtet und absichtlich eine hohe Dosierung angewandt.
2. Material und Methoden
Das Bacillus cereus Stamm Altdorf Präparat wurde auf Medium 11) hergestellt (s. unter
Kapitel "Züchtung"). Als Vergleichsstamm diente Bacillus fribourgensis B 4204, dessen
Vermehrung nach der Methode von DUTKY (1942) durch intrazoelomale Injektion in 900
L3-Larven von Melolontha melolontha erfolgte.
Das Engerlingsmaterial im Versuchsgebiet (Naturwiese) befand sich während der Behand¬
lung im zweiten Larvenstadium. Die behandelten Parzellen wurden mit 10 L wässerigerSporensuspension pro m2, bzw. die unbehandelten Kontrollparzellen mit der entsprechen¬den Menge Wasser bespritzt. Die Konzentration beider Präparate wurde auf ca. 2 x 10*2
Sporen/m2 eingestellt.
Der Versuchsplan entspricht dem Lateinischen Quadratsystem. Die unbehandelten Kon¬
trollparzellen lagen unmittelbar neben den behandelten Parzellen. Die totale Grösse des
Versuchsgebietes betrug 45 m . Davon wurden 2x5 m mit Bacillus cereus Stamm Alt¬
dorf Präparat und 1x5 m2 mit Bacillus fribourgensis Präparat behandelt und die restli¬
chen 6x5 m2 dienten als unbehandelte Kontrollen. Die Dimensionen der einzelnen Par¬
zellen betrugen 2 x 2,5 m. Auf jeder Parzelle wurde mit Hilfe zufällig angeordneter Zah¬
len (LINDER 1961) der Ort der Probeentnahme bestimmt. Bei der ersten und zweiten
Kontrolle wurden pro Parzelle 8 Proben von je Vß m2, bei der dritten Kontrolle 4 Proben
von je V8 m2 genommen.
Die Resultate wurden mit Hilfe einer Streuungszerlegung (LINDER 1960) für die Ermitt¬
lung der F- und t-Werte ausgewertet.
Bei der Auswertung der Resultate der dritten Kontrolle wurde die Quadratwurzel-Trans-
.
formation für die Aufhebung der Nullwerte angewandt, wobei statt mit x = Einzelwerte,mit x' =V x + 1 gerechnet wurde.
Die Wirksamkeit des Präparates wurde nach der Abott'schen Formel (ABBOT 1925) be¬
stimmt (siehe auch FINNEY 1952).
W = 100. ^, wobei W = Wirksamkeit in %
a = Zahl der lebenden Tiere pro Flächeneinheit unmittelbar
vor der Behandlung
k = Zahl der in einer gewissen Zeitspanne pro Flächeneinheit
der unbehandelten Parzellen abgetöteten Tiere ( = Bezugs¬mortalität)
b = Zahl der in der gleichen Zeitspanne pro Flächeneinheit
der behandelten Parzellen abgetöteten Tiere.
* Herrn R. Murbach, Dipl. Ing. Agr. ETH, Leiter der Abteilung für Pflanzenschutz, sei an dieser
Stelle für seine wertvolle Hilfe herzlich gedankt.
37

3. Versuchsergebnisse
Temperatur, Niederschlags- und Feuchtigkeitsverhältnisse sind in Tab. 13 aufgeführt.Wie ersichtlich, war die Versuchszeit durch feuchte und eher kühle Witterung gekenn¬zeichnet. Das Versuchsgebiet wies einen besonders starken Engerlingsbefall auf ; wie
aus Tab. 14 hervorgeht, wurde bei der ersten Kontrolle am 21.5.63 (1 Tag vor der Be¬
handlung) ein durchschnittlicher Befall von über 100 Engerlingen/m2 in den 9 Parzellen
festgestellt. Der berechnete F-Wert für die 9 Durchschnittswerte liegt innerhalb der
Sicherheitsgrenzen für P = 0,05 (s. Tab. 15) ; die Engerlinge waren also auf dem gan¬zen Versuchsgebiet homogen verteilt, und die Befallsunterschiede zwischen den 9 Par¬
zellen müssen als rein zufällig betrachtet werden. Bei der zweiten Kontrolle (am 28.6.
63, 5 Wochen nach der Behandlung) konnte auf den mit Bacillus cereus Stamm Altdorf
behandelten Parzellen eine hoch signifikante Wirksamkeit von 50,47 % festgestellt wer¬den. Wie aus Tab. 14 hervorgeht, blieb der Befall in der mit Bacillus fribourgensis be¬
handelten Parzelle und in allen 6 unbehandelten Kontrollparzellen in diesem Versuchs¬
abschnitt praktisch gleich (keine Abnahme). Bei der dritten Kontrolle (am 4.10. 63, 19
Wochen nach der Behandlung) wurde für die mit Bacillus cereus Stamm Altdorf behandel¬
ten Parzellen eine Wirksamkeit von total 81 % berechnet. Da die Bezugsmortalität wäh¬
rend der Zeit zwischen zweiter und dritter Kontrolle sehr hoch war, wurde der Befalls¬
unterschied zwischen den mit Bacillus cereus Stamm Altdorf behandelten und den 6 un¬
behandelten Parzellen geprüft. Dabei wurde berechnet, dass der Befallsunterschied zwi¬
schen den mit Bacillus cereus Stamm Altdorf behandelten Parzellen und 3 unbehandelten
Parzellen gesichert ist; bei einer Parzelle liegt dieser Wert an der Grenze und bei zwei¬
en ist er nicht gesichert und muss als zufällig betrachtet werden. Bacillus fribourgensiszeigt bei der letzten Kontrolle 12,5 % berechnete Wirksamkeit. Der Befallsunterschied
zwischen der mit Bacillus fribourgensis behandelten Parzelle und den 6 Kontrollparzel¬len ist nicht gesichert.
Betreffend der Wirkung des Bacillus fribourgensis Präparates muss darauf hingewiesenwerden, dass die effektive Befallsabnahme bei Versuchsabschluss 71 % beträgt. Die be¬
rechneten 12,5 % sind ein rein theoretischer Wert, der durch die hohe Bezugsmortalitätbedingt ist. Die hohe Bezugsmortalität kann durch eine Verbreitung der Krankheit auf
die Kontrollparzellen erklärt werden, die ja unmittelbar neben den behandelten Parzellen
lagen. Diese Vermutung wird durch die Beobachtung gestützt, dass in den Kontrollpar¬zellen einige an Bacillus cereus Stamm Altdorf erkrankte Engerlinge gefunden wurden.
Der Anteil dieses additiven Effektes an der Bezugsmortalität ist jedoch nicht abschätz¬
bar.
Es ist hier noch zu bemerken, dass bei einer Nachkontrolle ein Jahr später ein Aus¬
gleich im Befall zwischen behandelten und unbehandelten Parzellen festgestellt wurde.
38

Tab. 13 Temperatur, Niederschlag und Feuchtigkeitsverhältnisse im Versuchsgebiet während der Zeit von Mai - Oktober 1963
(Diese Werte wurden von der meteorologischen Station Martigny gemessen. )
Mai Juni Juli August September Oktober
Mittlere monatl. Lufttemperatur 12,74°C 15,82°C 19,00° C 15,98° C 14,07° C 9,63° C
Monatliche Niederschlagsmenge 41,7 mm 80,6 mm 43,0 mm 81,9 mm 27,5 mm 36,2 mm
Anzahl Niederschlags-Tage 8 Tage 18 Tage 8 Tage 16 Tage 7 Tage 4 Tage
Relative Luftfeuchtigkeit 65,4% 70,7% 68,4% 73,5% 76,5% 73,7%
Tab. 14 Ergebnisse des in Collonges durchgeführten Freilandversuchs mit Bacillus cereus Stamm Altdorf und Bacillus fribourgen-
sis.
Kontrollparzellenmit Bacillus cereus Stamm Altdorf
behandelte Parzellen
mit Bacillus fribourgensis
behandelte Parzelle
s- N XX
N X s-X
N
Engerlings-Befall pro 0,25 m2
am 21.5.63 (1 Tag vor Behandlung)26 14,28 48 31,7 16,9 16 23,5 12,17 8
Engerlings-Befall pro 0,25 m2
am 26. 6.63 (5 Wochen nach Behandlung) 26,3 9,08 48 16,0 7,58 16 25 3,0 8
Engerlings-Befall pro 0,25 m2am 4.10.63 (19 Wochen nach Behandlung)
10,5 4,84 24 3,0 2,82 8 7 2,0 4
Symbole : x = Mittelwert s- = Standardabweichung des Mittelwertes x N = Anzahl Proben

Tab. 15 Statistische Auswertung der Unterschiede im Engerlings-Befall zwischen behandelten und unbehandelten Par¬
zellen mit Hilfe der F- und t-Verteilung
Versuchsparzelle
Datum der Kontrolle,
resp. Zeitspanne zwischen den
Kontrollen
F "F
berechnet'
Tabelle
resp.
berechnet'
Tabelle
Wirksamkeit
in%
Alle 9 Parzellen 21.5.63
(1 Tag vor Behandlung)
1.1503 : n = 8P=°'05 : F=2'097
P=0,01 :F=2,823
n2=63P=0,001 : F=3,865
0
Mit Bacillus cereus Stamm Altdorf
behandelte Parzellen
21.5.'63 -
26.6.63
(5 Wochen nach Behandlung)
11,780** : n = lP=°'05 :F=4'183
n=29P=0,01 :F=7,5972
P=0,001 : F=13,391
50,47
Vergleich der mit Bacillus cereus
Stamm Altdorf behandelten Par¬
zellen mit den 6 einzelnen Kontroll¬
parzellen
4.10. 63
(19 Wochen nach Behandlung)(14 Wochen nach Behandlung)
Kx 0,4802
K2 3,0065* P=0,05 :t=2,228
g Ä : n=10P=0.01 : t =3,169
K5 2,3059* P=0,001 :t =4,587
K6 4,1734**
81,00
Vergleich der mit Bacillus cereus
Stamm Altdorf behandelten Par¬
zellen mit der mit Bacillus fri-
bourgensis behandelten Parzelle
4.10.63
P=0,05 :t =2,228
2,4197* : n=10p=001 . t^lß9
P=0,001 : t =4,587
Vergleich der mit Bacillus fribour-
gensis behandelten Parzelle mit
den 6 einzelnen Kontrollparzellen
4.10.63Ki 0,7639
K2 0,9623 P=0,05 :t =2,447
Il S:îSî = n=6 p=°'01 :t=3-707
K5 0,4690 P=0,001 : t=5,959
K6 2,1323
12,5
Verhältnis der berechneten Durchschnittsquadrate
Tabellenwert der berechneten Durchschnittsquadrate bei Frequenz n. und n,
Berechneter Unterschied zwischen zwei Durchschnitten
Tabellenwert der berechneten Unterschiede zwischen zwei Durchschnitten
bei Frequenz n
*= signifikant bei 5% Symbole: Fberechnet
significant bei 1 % F„
berechnet
Tabelle
t
1Tabelle

D) UNTERSUCHUNGEN DER HAEMOLYMPHE VON INFIZIERTEN UND GE¬
SUNDEN MELOLONTHA-LARVEN
I. BESTIMMUNGEN DES GEHALTES AN FREIEN AMINOSÄUREN IN DER HAEMOLYMPHE
GESUNDER UND KRANKER MELOLONTHA-LARVEN
1. Material und Methoden
Für die Blutuntersuchungen wurden Melolontha-Larven im dritten Stadium verwendet,die in bezug auf Gewicht (1,5 g), Grösse und physiologischen Zustand streng einheitlich
waren.
Es wurde das Blut von mit folgenden Mikroorganismen infizierten Larven untersucht :
Bacillus cereus Stamm .Altdorf LBG B 4156/a und Bacillus fribourgensis LBG B 4204.
Die Larven wurden nach der Methode von MARTIGNONI (1957) durch perorale Verab¬
reichung des Impfmaterials mit präparierter Glasnadel.(Mikrofütterung) infiziert.
a) Blutentnahme und Präparation des Blutes
Das Blut wurde den kranken Larven vor dem Absterben entnommen, wobei die Lar¬
ven noch schwache Reflexe zeigten.
Der Femur der anästhesierten Melolontha-Larven wurde durchgeschnitten und das
Blut mit einer Kapillarpipette aufgefangen. Zur Fällung der Proteine im Blut wurde
nach der Methode von WYATT (1956) vorgegangen, indem 7,5 % ige Trichloressig-säure im Verhältnis von 2 : 1 zum Blut gegeben, dieses während 3 Std. im Eisschrank
aufbewahrt und dann bei 6000 g abzentrifugiert wurde. Das wasserklare, proteinfreieBlut wurde vom Sediment getrennt und für die anschliessende Aminosäurebestimmungverwendet.
b) DUnnschichtchromatographie-Technik
Die 200 x 200 mm grossen DC-Platten wurden mit Kieselgel G nach der Üblichen La¬
bormethode von STAHL (1962) beschichtet und bei Zimmertemperatur luftgetrocknet.Auf jede Platte wurde 3 mm^ zu untersuchendes Material mit Hilfe einer Agla-Mikro-meterspritze aufgetragen. Die Trenneffekte wurden mit der zweidimensionalen Tech¬
nik erzielt, wobei als Fliessmittel für die erste Phase Chloroform-Methanol-17 %-ige Ammoniaklösung im Gewichtsverhältnis von 2:2:1, und für die zweite Phase
Phenol-Wasser im Verhältnis von 3 : 1 diente.
Zum Nachweis der Aminosäuren auf dem Chromatogramm diente Ninhydrin-Sprüh-
lösung: 0,3 g Ninhydrin wurde in 100 ml n-Butanol gelöst und mit 3 ml Eisessig ver¬
setzt. Nach gleichmässigem Besprühen der Platten und anschliessender Hitzebehand-
lung während 10 Minuten bei 110° C wurden die Resultate abgelesen und ausgewertet.
c) Auswertung
Die Auswertung erfolgte halbquantitativ mit Flächenmessung. Zur Gewinnung einer
Schätzungsbasis wurde 3 mm' gesundes Blut aufgetragen. Die daraus ausfallenden
Flecken, resp. Flächen wurden bei der Schätzung der Aminosäureabnahmen im kran¬
ken Blut als 100 % angenommen. (Fehlerquelle: ca. + 10 %). Alle angegebenen Re¬
sultate stützen sich auf dreimalige Wiederholung mit stets gleichen Ergebnissen.
2. Versuchsergebnisse
In der gesunden Haemolymphe wurden 17 freie Aminosäuren nachgewiesen (s. Abb. 24
und Tabelle 16), nämlich: Tryptophan, Leucin, Isoleucin, Valin, Tyrosin, Methionin,
Histidin, Threonin, Alanin, Prolin, Glycin, Serin, ß -Alanin, Glutaminsäure, Hydroxy-
lysin, Lysin, Arginin.
39

In der Haemolymphe der mit Bacillus cereus Stamm Altdorf, resp. Bacillus fribourgen-
sis infizierten Larven wurden Aminosäuren-Abnahmen bis zu 100 % festgestellt
(s. Abb. 25 - 26 und Tab. 16). Keiner der Mikroorganismen griff Leucin, Isoleucin
und Valin an. Nirgends wurde eine Zunahme freier Aminosäuren in kranker gegenüber
gesunder Haemolymphe beobachtet.
Zweidimensionale Aminosäure-Chromatogramme der gesunden, mit Bacillus cereus
Stamm Altdorf und Bacillus fribourgensis infizierten Haemolymphe von Melolontha-Lar¬
ven.
Val £
r\ Leu + Heu
Hist
Tyr
Q uUet
Gly OoßAla0GluO HylysU Lys
OStart _o<3
Abb. 24 Gesunde Haemolymphe
Start
nOTryU Leu + Heu
Val 0
f) Tyr
0 Hy!ys
0 Lvs
ÛArg°o 2
• Q f
Abb. 25 Mit Bacillus cereus Stamm
Altdorf infizierte Haemolymphe40

-, Leu + lieu
Val00 Tyr
Histn
Thre
Se d0o Glu
O HylysLys O o Arg
OStart A
0 Ala o Prol
Û jSAla
Abb. 26 Mit Bacillus fribourgensis
infizierte Haemolymphe
IL BESTIMMUNGEN DES PROTEINGEHALTES IN DER HAEMOLYMPHE GESUNDER UND
KRANKER MELOLONTHA-LARVEN UND UNTERSUCHUNGEN ZUM NACHWEIS HUMO¬
RALER IMMUNITÄTSFAKTOREN
1. Material und Methoden
Es wurde das Blut von mit folgenden Mikroorganismen infizierten Larven untersucht :
Bacillus cereus Stamm Altdorf LBG B 4156/a und Bacillus fribourgensis LBG B 4204.
Die Untersuchungen an infizierten und gesunden Larven wurden gleichzeitig durchge¬führt. Um eine eventuelle altersbedingte Fehlerquelle auszuschalten, wurden in bezugauf Alter, Gewicht (1,5 g) und physiologischen Zustand gleiche L3"Larven verwendet.
Als Vergleichsbasis wurden die Normalwerte der gesunden Haemolymphe mit der glei¬chen Methode ermittelt. Es wurde dabei die Haemolymphe von drei gesunden Larven
getrennt untersucht und der prozentuale Anteil der einzelnen Proteinfraktionen am Ge-
samteiweiss ermittelt.
a) Behandlung des Blutes
Das Blut der kranken Larven wurde im fortgeschrittenen Krankheitsstadium entnom¬
men und bei 12'000 g zentrifugiert.Wie bekannt, bildet sich im Insekten-Blut beim Kontakt mit Luft Melanin (LERNERand PATRICK, 1950). Bei Vorversuchen wurde festgestellt, dass dieses schwarze
Pigment von den wandernden Eiweissen mitgeschleppt wird und eine klare Auftren¬
nung der Proteinfraktionen erschwert. Das Blut wurde deshalb 2 Std. lang mit Cya¬nid behandelt (PRYOR 1955) und dadurch das katalytische Ferment Tyrosinase (ein
Kupfer-Protein-Komplex) inaktiviert.
41

b) Papierelektrophorese-Technik
Die elektrophoretische Auftrennung der Haemolymphe-Proteine erfolgte auf WHAT¬
MAN Nr. 1 Papierstreifen (7 x 30 cm) und wurde in einer Kunststoff-Trennkammer,
wie sie von GRASSMANN und Mitarbeitern (1950) beschrieben worden ist, durchge¬führt. Auf jeden mit Pufferlösung angefeuchteten Papierstreifen wurden mit Hilfe
einer Mikropipette 80 p 1 der zu untersuchenden Haemolymphe quer zur Laufrichtungin einem 3 cm langen Strich gleichmässig aufgetragen. Die Auftrennung erfolgte bei
Zimmertemperatur bei einer Gleichstromspannung von 250 V und einer Stromstärke
von 8-10 mA während 4 Stunden. Als Pufferlösung diente ein Natrium-Veronal-Puf-
fer nach MICHAELIS mit einem pH von 8,6 und einer Ionenstärke von u = 0,1, in
folgender Zusammensetzung: 29,43 g Veronal und 11, 71 g Natriumacetat anhydr.in dest. H2O gelöst. Nach Zufügen von 180 cc 0,1 n Salzsäure wurde mit Wasser auf
3 Liter aufgefüllt.
Nach der Auftrennung wurden die Papierstreifen bei 100° C freihängend 10 Minuten
lang getrocknet und anschliessend 15 Minuten zur Färbung in eine kalt gesättigte
Lösung von Amidoschwarz 10 B in Methanol-Eisessig 10 : 1 eingelegt. Entfärbt wur¬
de nach der üblichen Methode mit Methanol-Eisessig 10 : 1.
Zur Auswertung diente ein vollautomatischer Extinktionsschreiber Typ ZEISS.
c) Gesamtstickstoffbestimmung
Diese Bestimmung wurde nach der Mikro-Kjeldahl-Methbde (A. O. A. C. 1955) durch¬
geführt. Zur Analyse wurden jeweils 270 ji 1 Haemolymphe (= ca. 12 mg TS) verwen¬
det.
d) Versuche zum Nachweis humoraler Immunitätsfaktoren in der kranken Haemolymphe
Agglutinationsteste zum Nachweis spezifischer Antikörper wurden nach den üblichen
Labormethoden, Sensibilitätsteste zum Nachweis von Inhibinen mit dem Plattentest-
verfahren nach HALLMANN (1961) durchgeführt.
2. Versuchsergebnisse
Wie aus Tab. 17 und Abb. 27 hervorgeht, lassen sich die Haemolymphe-Proteine der
Melolontha-Larven in 4 Fraktionen aufteilen, wobei die Fraktionen Bj + Bu eine etwa
1,6 x höhere Konzentration aufweisen als die Fraktionen Ai + Ajj. Der isoelektrische
Punkt für die Fraktionen Aj und Ay liegt bei pH 6,1 und für die Fraktionen Bi und Bjjbei 7,0. Dies stimmt annähernd mit den Beobachtungen von KRIEG (1956) Uberein :
"Hauptfraktion" A bei pH 6,5, "Hauptfraktion" B bei pH 6,8.
In bezug auf Wanderungsgeschwindigkeiten, resp. isoelektrische Punkte zeigen die Pro¬
teinfraktionen in der Melolontha-Haemolymphe Aehnlichkeit mit den Globulin-Fraktio-
nen der Wirbeltiere, deren isoelektrische Punkte zwischen pH 5 und pH 7,3 liegen. In
der Melolontha -Haemolymphe konnte jedoch keine Fraktion registriert werden, die
der Albumin-Fraktion bei den Wirbeltieren entspräche, deren isoelektrischer Punkt
bei pH 4,2 liegt.
Die Infektion mit Bacillus cereus Stamm Altdorf bewirkt einen starken Konzentrations¬
anstieg der beiden B-Fraktionen und möglicherweise eine relative Abnahme der Frak¬
tion Aj. Wie aus Abb. 28 hervorgeht, wurde die Fraktion Ajj durch die zwei B-Fraktio¬
nen überdeckt, so dass sich über ihren Anteil nichts aussagen lässt.
Die mit Bacillus fribourgensis infizierte Haemolymphe zeigt ebenfalls eine deutliche
Konzentrationszunahme der Bj - und Bjr-Fraktionen und eine starke Abnahme der Apund Ajj-Fraktionen (s. Abb. 29).
42

Tab. 16 Abnahme des Gehaltes an freien Aminosäuren in der Haemolymphe von Melolontha-
Larven im L3-Stadium bei Infektion mit Bacillus cereus Stamm Altdorf und Bacillus
fribourgensis
Gesunde Haemolymphe Mit Bacillus cereus Stamm Altdorf Mit Bacillus friboureensis
infizierte Haemolymphe infizierte Haemolymphe
Aminosäure Abkürzung Abnahme in % Abnahme in %
Tryptophan Try 100 100
Leucin Leu - -
Isoleucin Heu - -
Valin Val - -
Tyrosin Tyr 50 -
Methionin Met - 100
Histidin His 30 10-20
Threonin Thre 100 60
Alanin Ala 100 -
Prolin Pro 100 -
Glycin Gly - 10-20
Serin Se 100 -
ß-Alanin /SAla 50 30
Glutaminsäure Glu 10-20 40
Hydroxylysin Hylys 10-20 60
Lysin Lys - 20
Arginin Arg 100 60

Tab. 17
Prozentualer Anteil der Protein-Fraktionen am
Gesamt-Protein in der gesunden Haemolymphe
Larve No. Ai AII Bi BII A : B
20
21
22
28,126
27
10,9
9
13
40,744
38
20,321
22
39 : 61
35 : 65
40 : 60
x = 27 x = 10,9 x = 40,9 x = 21,1 x = 38 : 62
Tab. 18
Prozentualer Anteil der Protein-Fraktionen am Gesamt-Protein in
der mit Bacillus cereus Stamm Altdorf infizierten Haemolvmphe
Larve No. Ai An Bi + Bu A : B
23
24
10
8
•90
92
10 : 90
8 : 92
x = 9 •x = 91 x = 9 : 91
Tab. 19
Prozentualer Anteil der Protein-Fraktionen am Gesamt-Protein in
der mit Bacillus fribourgensis infizierten Haemolymphe
Larve No. AI AII BI BII A : B
25
26
5,5
6,5
4,0
3,5
43
42
47,548
9,5 : 90,5
10,0 : 90
x = 6,0 x = 3, 75 x = 42,5 x = 47,75 x = 9,75 : 90,25
Tab. 20 Gesamtstickstoffgehalt in gesunder und mit Bacillus cereus Stamm Altdorf infizierter,
bakterienfrei zentrifugierter Haemolymphe
Total-N-Gehalt in mg % TS
Wiederholungen
Mittelwert1. 2. 3. 4. 5.
Gesamt-N-Gehalt in gesunderHaemolymphe
10,126 10,126 10,103 10,150 10,126 10,126
Gesamt-N-Gehalt in mit
Bacillus cereus Stamm Altdorf
infizierter Haemolymphe
14,186 14,280 14,233 14,140 14,140 14,186

Die Konzentrationszunahme zugunsten der B-Fraktionen steht beim mit Bacillus fribour-
gensis infizierten Blut im Verhältnis 1 : 8-9 (Fraktion A : B) (s. Tab. 19). Beim mit Ba¬
cillus cereus Stamm Altdorf infizierten Blut lässt sich das Verhältnis der Konzentra¬
tionszunahme wegen der Ueberdeckung der An-Fraktion nicht abschätzen (s. Tab. 18).
Wie aus den Gesamtstickstoff-Bestimmungen (Tab. 20) hervorgeht, rührt die Zunahme
der B-Fraktionen von einer effektiven Proteinzunahme um ca. 40 % des Gesamtstick¬
stoffgehaltes gegenüber normalem Blut her.
Um festzustellen, ob diese Proteinzunahme auf die Bildung von spezifischen Abwehr¬stoffen zurückgeführt werden kann, wurden Agglutinationsteste durchgeführt. Es konn¬
ten jedoch keine Antikörper nachgewiesen werden.
In diesem Zusammenhang sei bemerkt, dass mit Plattendiffusionstesten auch keine In-
hibine mit bakteriostatischer oder bakterizider Wirkung (wie beschrieben von BRIGGS,
1958, STEPHENS, 1959, 1962a, 1962b, und STEPHENS-MARSHALL 1962) nachgewie¬sen werden konnten.
Ext,
B. B.II I II I
Abb. 27 Elektrophorese-Diagram einer normalen Haemolymphe
43

Ext.
Abb. 28 Elektrophorese-Diagramm einer mit Bacillus
cereus Stamm Altdorf infizierten Haemo-
lymphe
Ext.
Abb. 29 Elektrophorese-Diagramm einer mit Bacillus44 fribourgensis infizierten Haemolymphe

Abb. 30 keine Agglutination
Abb. 31 positive AgglutinationAgglutinationstiter 1 : 20,
3 Wochen nach der Immunisierung
45

III.PASSIVE IMMUNISIERUNG
1. Material und Methoden
Es wurden total 100 Lß-Larven je 3 mm^ Kaninchenserum anti-Bacillus cereus Stamm
Altdorf B 4156/a, mit einem Titer von 1 : 6400, nach der üblichen Methode injiziert.Nach der Injektion entspricht dies einer Serumkonzentration und einem Titer von ca.
1 : 80, da der Blutgehalt einer L3 -Larve ca. 250 mm3 beträgt.
24 Std. nach der Immunisierung und anschliessend mit Intervallen von 4-5 Tagen wur¬de der jeweilige Agglutinationstiter der Haemolymphe bestimmt, und zwar mit dem Ob¬
jektträger-Agglutinationsverfahren mit gleichzeitiger mikroskopischer Kontrolle. Die
positive Reaktion wird durch eine sofortige Verklumpung der Bacillen gekennzeichnet,während bei negativer Reaktion keine Veränderung eintritt, d. h. die Mikroorganismennicht verklumpen, sondern nach wie vor einzeln erkennbar sind (s. Abb. 30 - 31). Die¬
ser passiv erworbene Schutz wurde durch Injektionen von lebenden Kulturen von Bacillus
cereus Stamm Altdorf B 4156/a überwacht, und die Mortalität der immunisierten Lar¬
ven, bzw. der nicht immunisierten Kontroll-Larven erfasst (s. Tab. 22).
2. Versuchsergebnisse
Die passive Immunisierung durch Verabreichung von 3 mm3 Immun-Kaninchenserum
wurde von den Melolontha-Larven ohne den geringsten Schaden ertragen. Wie aus Tab.
21 hervorgeht, konnte der berechnete Agglutinationstiter von 1 : 80 noch 24 Std. nach
der Injektion nachgewiesen werden. Der Wirtsorganismus brauchte 5 Wochen bis zur
totalen Eliminierung der erworbenen Antikörper-Moleküle, und da er selbst keine Anti¬
körper bilden kann, ging gleichzeitig auch die Schutzwirkung verloren. Wie die Morta¬
litätszahlen zeigen, konnten die immunisierten Larven nach 5 Wochen einer Infektion
von Bacillus cereus Stamm Altdorf genau so wenig entgehen wie die nicht immunisier¬
ten Kontroll-Larven.
Zusammenfassung
Im Jahre 1961 wurden in Altdorf (Kt. Uri) aus kranken Melolontha-Larven sporenbildendeAngehörige des Genus Bacillus isoliert, deren morphologische, physiologische und immu-
nobiologische Eigenschaften in der vorliegenden Arbeit eingehend untersucht wurden. Auf
Grund dieser Untersuchungen wurden die isolierten Stämme als Bacillus cereus identifiziert.
Für die Züchtung von Bacillus cereus Stamm Altdorf in belüfteten Submerskulturen zur Her¬
stellung eines Sporenpräparates bewährte sich ein selbst zusammengestelltes Medium.
Dieses enthält Glukose als Kohlenstoffquelle, L (+) Glutaminsäure, Pepton und Hefeextrakt
als Stickstoff- und Wuchsstoffquellen, ferner Phosphat, Mineralsalze und Kartoffelextrakt.
Die beste Sporenausbeute wurde bei einem Anfangs-pH von 6,5 und bei einer Temperaturvon 30° C erzielt.
Glukose als Kohlenstoffquelle wurde wahrend des Wachstums innerhalb 12 - 14 Std. prak¬tisch total ausgenützt. Ein Weglassen der Glukose aus dem Medium bewirkte eine starke
Abnahme der Endausbeute, ohne den prozentualen Sporenanteil wesentlich zu beeinflussen.
Ein Weglassen der L (+) Glutaminsäure hingegen verminderte die Trockenausbeute nur
wenig, den prozentualen Sporenanteil aber ganz beträchtlich.
Die Wirksamkeit des selbst hergestellten Sporenpräparates gegen Melolontha-Larven wur¬
de in Labor- und Freilandversuchen geprüft. In den Laborversuchen wurden mit Mikro-
fütterung und Erdbeimpfung Mortalitäten bis zu 94 % erzielt, wobei sich vor allem das er¬
ste und zweite Entwicklungsstadium der Melolontha-Larven für die Behandlung als günstigerwiesen. Bei Infektionsversuchen mit Bacillus cereus Stamm Altdorf gegenüber Raupenvon Galleria Mellonella L. wurde eine Mortalität von 82 % erzielt. Gegenüber Raupen von
Pieris brassicae L. erwies sich Bacillus cereus Stamm Altdorf als nicht wirksam.

Tab. 21 Versuchsergebnisse der passiven Immunisierung von Melolontha-Larven mit Bacillus cereus Stamm Altdorf
Blutentnahme nach Anzahl
Tagen nach ImmunisierungAgglutination Anzahl inf. Larven/
Anzahl Kontroll-Larven
Inoculum/veg.Zellen pro
Larve
Mortalität
Reaktion Titer Immunis.
Larven
Kontroll-
Larven
24 Std. + 1 : 80 5/5 2x 104 0 5
5 Tage + 1 :40 5/5 2x 104 0 5
10 Tage + 1 :40 5/5 2x 104 0 5
15 Tage + 1 :40 5/5 2x 104 0 5
20 Tage + 1 : 30 5/5 2x 104 0 5
25 Tage + 1 : 20 5/5 2x 104 0 5
30 Tage + 1 : 1 5 /5 2x 104 3 5
34 Tage - unverdünnt 5/5 2x 104 5 5

Im Freilandversuch wurde nach 5 Wochen eine signifikante Wirksamkeit von 50,4 % festge¬stellt. Während dieses Versuchsabschnittes blieb der Befall in den Kontrollparzellen unver¬
ändert (keine Abnahme). 19 Wochen nach der Behandlung lag die Wirksamkeit bei 81 % ;
dieser Wert ist jedoch wegen der relativ hohen Bezugsmortalität nicht gegenüber allen Kon¬
trollparzellen gesichert. Im zweiten Jahr war keine Wirkung mehr festzustellen.
In der Haemolymphe von gesunden Melolontha-Larven wurden 17 freie Aminosäuren und 4
Proteinfraktionen nachgewiesen. Nach einer Infektion mit Bacillus cereus Stamm Altdorf
wurden von den 17 freien Aminosäuren 6 total und weitere 5 teilweise abgebaut. Ebenfalls
änderte sich das Proteinspektrum der Haemolymphe mit eindeutigem Anstieg der B-Fraktio-
nen und einer Gesamtproteinzunahme um ca. 40 %. Antikörper oder Inhibine konnten nicht
nachgewiesen werden.
48

Literatur
ABBOT, W.S.
A.O.A.C.
ASHESHOV, J.N.BERGEY's Manual of Determinative
Bacteriology,BONNEFOI, A., et BEGUIN, S.
BÖTTCHER, K.
BRIGGS, J.D.BUCHER, E.G.
BURDON, K.L.
CHAPMAN, B.G.
COOPER, CM., FERNSTRÖM, G.A.,
and MILLER, S.A.
CHRISTENSEN, W.B.
DE BARJAC, H., et BONNEFOI, A.
DIFCO MANUAL
DUTKY, S.R.
DUTKY, S.R.
ETTLINGER, L., LEISINGER, Th.,
und KABAY, A.
FINNEY, D.J.
GRASSMANN, W., u. HANNIG, K.
GRELET, N.
HALLMANN, L.
HASTINGS, E.G.
HEIMPEL, A.M.
HORBER, E.
HOWIE, J.W., and CRUICKSHANK, J.HURPIN, B.
HURPIN, B., etVAGO, C.
KELLENBERGER, E., RYTER, A.,
and SECHAUD, J.
KERN, F.
KNIGHT, B.C.J.G., and PROOM, H.
KRIEG, A.
KRIEG, A.
KUHNER, J., u. HEIMPEL, A.M.
LEMOIGNE, M.A., BONNEFOI, A.,
BEGUIN, S., GRISON, P., MARTOU-
RET, D., SCHENK, A., u. VAGO, C.
: J. Econ. Entomol. 18, 265 - 267 (1925): Official Methods of Analysis of the Association
of Official Agricultural Chemists. 8th Edition
Washington (1955): Can. Publ. Health J. 32, 468 - 471 (1941)
: 7th Edition, Baltimore (1957): Entomophaga ±, 194 - 199 (1959): Naturwiss. 46, 630 - 631 (1959): J. Exptl. Zool. 138, 155 - 158 (1958): J. Insect Pathol. 2, 172 - 195 (1960): J. Bacteriol. 52, 665 - 678 (1946): J. Bacteriol. 71, 348 - 355 (1956)
: Industr. Eng. Chemistry 36, 504 - 508 (1944): J. Bacteriol. 52, 461 - 466 (1946):' Entomophaga 7, 5-31 (1962): 9th Edition, Detroit (1953): J. Agr. Res. 61 (1) (1940): U. S. Dept. Agr. Bur. Entomol. Plant Quarantine,
ET 192, 10 pp
: Bacillen der thuringiensis-Gruppe als Parasiten
von Melolontha. Internationales Kolloquium über
Insektenpathologie und mikrobiologische Bekämp¬
fung, Paris 1962, Entomophaga, Mémoire hors
série No. 2, 345 - 350 (1964): Statistical Methods in Biological Assay, London
(1952)Naturwiss. 37, 396 (1950)
: Ann. Inst. Pasteur 81, 430 - 440 (1951): Bakteriologie und Serologie, 3. Auflage, Stutt¬
gart (1961): Zbl. Bakteriol. II. Abt., 10,
384 (1903): Can. J. Zool. 33, 311 - 326 (1955): Landw. Jahrbuch der Schweiz 73, 361 - 369 (1959): J. Pathol. 50, 235 - 242 (1940): Compt. rend. Soc. Biol. 149, 1966 - 1967 (1955): Entomophaga 3_, 285 - 330 (1958)
: J. Biophys. and Biochem. Cytol. 4, 671 - 678
(1958): Promotionsarbeit Eidgenössische Technische
Hochschule (1950): J. General Microbiol. 13, 474 - 480 (1950): Naturwiss. 43, 60 (1956): Unveröffentlicht (1957): Can. J. Microbiol. 1, 551 - 574 (1957)
Entomophaga J_, 19-34 (1956)
49

LEIFSON, E. :
LERNER, A.B, and FRITZPATRICK,
T.B. :
LINDER, A. :
LINDER, A. :
MACKIE and McCARTNEY :
Manual of Microbiological Methods. :
MARTIGNONI, M. E. :
MARTIGNONI, M.E., and STEINHAUS,E.A. :
McGAUGHEY and CHU :
MOOR, H., and MÜHLETHALER, K. :
MÜLLER-KÖGLER, E. :
NORDBERC, B.K. :
NORDBERG, B.K. :
PRYOR, M.G.M. :
ROBINOW, CF. :
SMITH, N.R., GORDON, R.E. and
CLARK, F.E. :
SUMMER, J. B. :
STAHL, E. :
Staining Procedures used by the
Biological Stain Commission :
STEPHENS, J.M. :
STEPHENS, J.M. :
STEPHENS, J. M. :
STEPHENS, J. M., and MARSHALL,
J.H. :
TOUMANOFF, C. :
WIKEN, T., und WILLE, H. :
WIKEN, T., BOVEY, P., WILLE, H.,
u. WILDBOLZ, T. :
WILDBOLZ, T. :
WILLE, H. :
WILLE, H., WIKEN, T., u. BOVEY, P. :
WYATT, G.R., LOUGHHEED, T.C.,
andWYATT, S.S. :
YOUNG, E.I., and FITZ-JAMES, Ph.C:
YOUNG, E.I., and FITZ-JAMES, Ph.C:
Atlas of Bacterial Flagellation, New York and
London (1960)
Physiol. Revs. 30, 91 - 126 (1957)Mathematische Statistik, Basel und Stuttgart (1960)Handliche Sammlung mathematisch-statistischer
Tafeln, Basel und Stuttgart (1961)Handbook of Bacteriology. 10th Edition, Edinburgand London (1960)
Society of American Bacteriologists, Committee
on Bacteriological Technic, New York (1957)
J. Insect. Pathol. J_, 294 - 296 (1957)
Laboratory exercises in insect microbiology and
insect pathology. Minneapolis (1961)J. General Microbiol. 2, 334 - 340 (1948)
J. Cell Biol. 17, 609 - 628 (1963)
Entomophaga ±, 261 - 274 (1959)Studies of Bacillus anthracis in regard to its pro¬
perties of diagnostic and pathogenic importance,Stockholm (1951)Nord. Veterinärmed., 5, 915 - 924 (1953)
J. Exp. Biol. 32, 468 - 484 (1955)Bacterial Anatomy Sixth Symp. Soc. for General
Microbiology, 181 - 201 (1956)
US Dept. Agric. misc. Publ. 559 (1946)J. biol. Chem. 62, 287 (1925)
DUnnschichtchromatographie Berlin (1962)
2nd Edition ; Baltimore (1960)Can. J. Microbiol. _5_, 203 - 228 (1959)Can. J. Microbiol. .8, 491 - 499 (1962a)Can. J. Microbiol. _8, 597 - 602 (1962b)
Can. J. Microbiol. _8, 719 - 725 (1962)Ann. Inst. Pasteur J55_, 90 - 98 (1953)Zbl. Bakteriol. II. Abt. 107, 259 - 271 (1953)
Z. angew. Entom. 36, 1 - 19, (1954)Mitteil. Schweiz. Entomol. Ges., ,27, 240 (1954)Mitteil. Schweiz. Entomol. Ges., 29, 271 - 282
(1956)
Entomophaga _7, 161 - 174 (1962)
J. General Physiol. 39, 853 - 868 (1956)J. Biophys. Biochem. Cytol. 6, 467 - 482 (1959a)
J. Biophys. Biochem. Cytol. 6, 483 - 498 (1959b)
50

DANK
Herrn Professor Dr. L. Ettlinger, Vorstand des Mikrobiologischen Institutes, möchte ich für
seine persönliche Anteilnahme und seine allseitige Unterstützung bei der Ausführung dieser Ar¬
beit meinen vorzüglichen Dank aussprechen.
Herrn Professor Dr. K. Mühlethaler, Leiter des Elektronenmikroskopischen Laboratoriums
der E.T. H., danke ich bestens für die Zurverfügungstellung des Elektronenmikroskopes, sowie
für seine wertvollen Ratschläge.
Ferner gilt mein Dank Herrn Dr. E. Horber, Leiter der Abteilung für Pflanzenschutz der Eid¬
genössischen Landwirtschaftlichen Versuchsanstalt, Zürich-Oerlikon, für die grosszügige Ueber-
lassung von Versuchsmaterial und für seine wertvollen Hinweise.
Allen Institutsmitgliedern, die mir in irgendeiner Weise behilflich waren, danke ich be¬
stens.
An die Druckkosten dieser Arbeit wurde vom Laur-Fonds ein Beitrag geleistet, wofür ich
auch an dieser Stelle herzlich danken möchte.

Curriculum Vitae
von
Antal Kabay
1932 22. November, geboren in Budapest.
1939 - 1943 Elementarschule inSzbllösgyörbk (Ungarn) .
1943 - 1951 Humangymnasium Sopron (Ungarn). Matura Typ B.
1951 - 1956 Studium an der Agrarwissenschaftllchen Universität Budapest-Gödöl-1 ö. Neun abgeschlossene Semester mit allen regulären Prüfungen.
1957 Einreise in die Schweiz.
1957 - 1958 Assistent am Veterinär-Bakteriologischen Institut der Universität
Bern unter Herrn PD Dr. K. Klingler (Direktor: Herr Prof. Dr.
G. Schrriid -))
1958 Sommersemester. Studium an der Abteilung für Landwirtschaft an
der Eidgenössischen Technischen Hochschule.
1959 März: 2. Vordiplomprüfung.
1960 Studiumabschluss. Dipl. Ing.-Agr. E.T.H.
1960 - 1961 Wissenschaftlicher Mitarbeiter am Institut für Tierzucht der Eidge¬nössischen Technischen Hochschule, unter Leitung von Herrn Prof. Dr.
H. Lörtscher. Arbeit im Gebiet der Biometrik.
1961- 1964 Wissenschaftlicher Mitarbeiter am Mikrobiologischen Institut der Eid-
genössichen Technischen Hochschule, unter Leitung von Herrn Prof. Dr.
L. Ettlinger. Ausführung der Promotionsarbeit.