rhythmic growth rings of wood and their relationship with the foliage in oak seedlings grown in a...
TRANSCRIPT

Annals of Botany 82 : 89–96, 1998
Rhythmic Growth Rings of Wood and Their Relationship with the Foliage in Oak
Seedlings Grown in a Favourable Environment
ANAS MANSOUR and ELISABETH de FAY$ §
Uni�ersiteU Henri PoincareU , Nancy I, Laboratoire de Biologie Forestie[ re, AssocieU INRA, BP 239,
54506 Vandœu�re-le[ s-Nancy Cedex, France
Received: 5 December 1997 Returned for revision: 20 January 1998 Accepted: 3 April 1998
The anatomy of wood was studied in Quercus robur L. and Quercus suber L. seedlings exhibiting 3–8 units of extension,each with a tier of photosynthesizing leaves in their upper parts, generated as the result of rhythmic shoot growth underfavourable conditions. At all the axis levels examined (i.e. the base of each of the different units of extension, fourother equidistant levels within the first unit of extension and the upper part of the taproot), the wood displayed ringswhen treated with Wiesner reagents. This indicated cinnamaldehyde groups present in lignins. No rings appearedwhen the Mau$ le reaction was used for specific detection of syringyl subunits in lignins. A trend towards a periodicalarrangement of xylem parenchyma bands was also found when sections were treated with I
#}KI. The number of rings
coincided with the number of leaf tiers above the level of measurement, and did not vary inside the first unit ofextension. Thus, the rings are called rhythmic growth rings. In sections of the first and the second units of extension,and in the taproot, the area and width of a given rhythmic growth ring were highly correlated with the total area ofleaves present above the level of measurement at the presumed time of growth ring formation. Moreover, stemdiameter at the base of the units of extension was highly correlated with the leaf area above. These results indicatethat differentiation of xylem, particularly its lignification, varies rhythmically in oak seedlings. They imply that woodproduction is linked to the photosynthesizing and}or transpiring area of the plant. Thus, during a growth cycle ofQ. robur and Q. suber seedlings, there appears to be integration of the primary metabolic activities with the layingdown of rhythmic growth rings. # 1998 Annals of Botany Company
Key words : Allometric relationship, juvenile wood, leaf area, lignification, oak seedling, periodic structure, Quercusrobur L., Quercus suber L., rhythmic growth ring, unit of extension.
INTRODUCTION
The wood of temperate-zone trees shows annual growthrings, that is to say, annual increments consisting of initiallayers with large conducting elements (early wood) and laterlayers with smaller ones (late wood). Occasionally, ad-ditional rings are formed within a growing season. Whenthis occurs, the usual number of multiple (intra-annual)rings is two, but quite often more are found, as reported byKozlowski (1971). According to this author, the formationof multiple rings, also called false rings, is promoted byabrupt environmental changes as well as various injuries ;moreover, production of lammas shoots is usually ac-companied by formation of multiple rings, at least near thetip of the shoot, but exceptions occur in several species ofoak growing under natural conditions (Wight, 1930;Grudzinskaya, 1957; Guard and Postlethwait, 1958;Longman and Coutts, 1974). In seedlings of the flushingspecies, He�ea brasiliensis (the commercial rubber tree)grown under natural conditions, and Quercus robur, grownin controlled conditions, multiple rings were found and theirnumber was related to the order of appearance of the unitof extension considered (Halle! and Martin, 1968; Payan,1982; de Fay$ , 1985). The histological characteristics of these
§ For correspondence.
false rings have not been studied in detail. Alternation oflayers rich in vessels with layers rich in fibres, withoutflattening of the cells, was found in the wood of Quercusrobur seedlings, but false rings of this kind were oftendiscontinuous and poorly distinguished (Payan, 1982). InHe�ea brasiliensis seedlings, false rings were, above all, theresult of periodic variation in the spacing of axial paren-chyma bands, and their boundaries were gradual (de Fay$ ,1985).
This paper describes a qualitative and a quantitativestudy of the wood of Quercus robur L. and Quercus suber L.seedlings, and of the external structure of the plants (unit ofextension, tier of leaves). It is designed to clarify the natureand origin of the false rings of flushing species.
MATERIALS AND METHODS
Acorns (with pericarp and testa removed) were germinatedin moist vermiculite. Fifteen (³2) d later, when the primaryroots were 7–8 cm long, the plantlets were placed in-dividually in pots filled with potting compost of shreddedsoil and rotted manure. The seedlings were grown in agrowth chamber with a 16 h photoperiod, at 20–27 °C,and 60–70% relative humidity, and watered approx. twicea week. Sodium lamps yielding a photon flux density of
0305-7364}98}07008908 $30.00}0 bo980648 # 1998 Annals of Botany Company

90 Mansour and de FayX —Rhythmic Growth Rings and Allometric Relationships in Oak Seedlings
F 1–5. For legends see facing page.

Mansour and de FayX —Rhythmic Growth Rings and Allometric Relationships in Oak Seedlings 91
F. 6–8. Transverse sections of the stem of a Q. suber seedling of four complete UE, within the first UE. At the base, showing four rings (Fig.6) ; in the middle, showing four rings (Fig. 7) ; at the top, showing four rings (Fig. 8). Bars¯ 0±5 mm. All sections stained with Wiesner reagents.
F 9–13. Transverse sections of the axis of a Q. robur seedling of four complete UE, at five levels. Upper part of the taproot, showing four rings(Fig. 9) ; base of the first UE, showing four rings (Fig. 10) ; base of the second UE, showing three rings (Fig. 11) ; base of the third UE, showing
two rings (Fig. 12) ; base of the fourth UE, showing one ring (Fig. 13). Bars¯ 0±5 mm. All sections stained with Wiesner reagents.
130 mmol m−# s−" PAR (rich in orange radiation) were used.Under these conditions, at 7 months of age the seedlings hada mean height of 1±30 m in Q. robur and 0±75 m in Q. suber.Moreover, the plants produced repeated flushes and restingperiods: they flushed, on average, every 26 d in Q. robur and
F 1–2. Transverse sections of the stem of a Q. robur seedling of eight complete units of extension (UE) at the base of the first UE, treatedwith Wiesner reagents (Fig. 1, note rings) ; and Mau$ le reagents (Fig. 2, rings not shown). Bars¯ 0±5 mm.
F 3–5. Transverse sections of the stem of a Q. robur seedling of three complete UE at the base of the first UE. Fig. 3. Detail of wood stainedwith Wiesner reagents : H, Highly-reactive layer ; P, poorly-reactive layer ; V, vessel. Bar¯ 0±1 mm. Fig. 4. General view of the section stained withI#}KI. F, Wood layer rich in fibres ; Pa, wood layer rich in parenchyma bands; Pi, pith. Bar¯ 0±5 mm. Fig. 5. The same section stained with
Wiesner reagents. Note the three rings. Bar¯ 0±5 mm.
every 40 d in Q. suber. This growth pattern was firstdescribed by Lavarenne-Allary (1965) in common oakseedlings in controlled, as well as natural, environmentsand in coppice shoots. It is common in the tropics and sub-tropics (e.g. He�ea, Thea, cacao) where the genus Quercus
92 Mansour and de FayX —Rhythmic Growth Rings and Allometric Relationships in Oak Seedlings
may have evolved (Longman and Coutts, 1974; Halle! ,Oldeman and Tomlinson, 1978; Tomlinson and Longman,1981). The oak seedling stems had an articulate morphology,as previously observed in Q. robur under similar conditionsof growth by Payan (1982), and by Alatou (1990) in Q.robur and Q. suber. Q. robur built typical units of extension(UE): cataphylls were produced in relatively great numberand foliage leaves were thus arranged in well-separatedtiers. In contrast, the UE of Q. suber appeared less distinctinitially, because of a less marked heteroblasty and a greaternumber of foliage leaves, but, after a preliminary study ofthe growth, they could be delimited accurately.
For the anatomical and histological study of wood, atleast ten plants of each species were examined at the end ofthe fourth interflush, and several more at the end of thethird or eighth interflush. The seedlings were sampled: (1) at3 cm from the base of each UE, i.e. always below the firstleaf of the UE; (2) at four other equidistant levels within thefirst UE; and (3) in the upper part of the taproot at 3 cmfrom the root collar. Samples were fixed in CRAF I (Sass,1958) ; 40 µm-thick transverse sections were made witha freezing microtome and stained for observation by lightmicroscopy with one of the three following reagent stains :(1) Wiesner reagents (Sass, 1958) which mainly indicatecinnamaldehyde groups present in lignins (Monties, 1980;Vallet et al., 1996). Fresh sections were left for 3 min in 1%phloroglucinol in 95% ethanol and mounted in 50%HCl; (2) Mau$ le reagents (Dop and Gautie! , 1928; Locquinand Langeron, 1978), which are specific for syringyl subunitsin lignins (di-methoxylated residues). The treatment con-sisted of a 5 min staining in 1% KMnO
%, followed by
washing in water and then transfer of sections to 50% HCluntil partially decoloured. Finally, the sections were washedthoroughly in water and mounted in 50% NH
%OH; (3)
Lugol reagents, (Sass, 1958) which identify starch, and thusshowed starch-rich parenchyma at low magnification. Freshsections were left for 10 min in solution of I
#}KI (1 g of each
in 100 ml distilled water), and washed quickly.For the allometric study, nine seedlings (five Q. robur and
four Q. suber) were used. At the end of the eighth interflush,stem diameter was measured at 3 cm from the base of eachUE, twice at right angles, and the mean recorded. Thefoliage leaves were collected from each UE, and quicklyphotocopied, with the leaf blades arranged on the samesheet (typing paper of uniform thickness). The photocopiedsurfaces were carefully cut out ; the weight of the cutpieces of paper indicated the leaf area. In addition, for fiveUE, a second photocopy was made on graph paper,permitting direct calculation of leaf area by counting squaremillimetres. Since the results of the graph paper methoddiffered from those of the weighing method by only 1% onaverage, the former, which was the faster method, wassystematically adopted. The axis was sampled at threelevels : the base of the first UE, the base of the second UE(both at 3 cm from the base) and the upper part of thetaproot (at 3 cm below the root collar). For each sample,transverse sections were made and stained with phloro-glucinol-HCl, as above. Width of the observed rings wasmeasured with an ocular micrometer under a stereomicro-scope along four radii at right angles, and the mean used.
Ring area was calculated as the difference between the areasof two concentric circles.
RESULTS
Presence and nature of periodic structures in xylem
At all levels of the axis examined, and in all seedlings ofboth Q. robur and Q. suber, the wood displayed rings underthe stereomicroscope when treated with Wiesner reagents(Fig. 1). In contrast, Mau$ le reagents showed no rings (Fig.2). The alternation of layers poorly- and highly-reactive tophloroglucinol-HCl was regular. The poorly-reactive layerswere generally wider than the highly-reactive layers. Never-theless, in all cases, the first-formed layer (adjacent to thepith) was highly-reactive and wide. At higher magnifications(under a standard optical microscope), differences in in-tensity of staining were less clear and were not caused bysignificant concentric variations in wall thickness, radialdimensions of xylem fibres and parenchyma cells, or size andnumber of vessel pores (Fig. 3).
The I#}KI reaction at low magnification showed alterna-
ting layers rich in axial parenchyma bands and rich in fibres(Fig. 4). These successive layers merged frequently in somesectors of the cross-sectional area of the xylem. Layers richin fibres coincided fairly well with layers highly-reactive tophloroglucinol-HCl (Fig. 5).
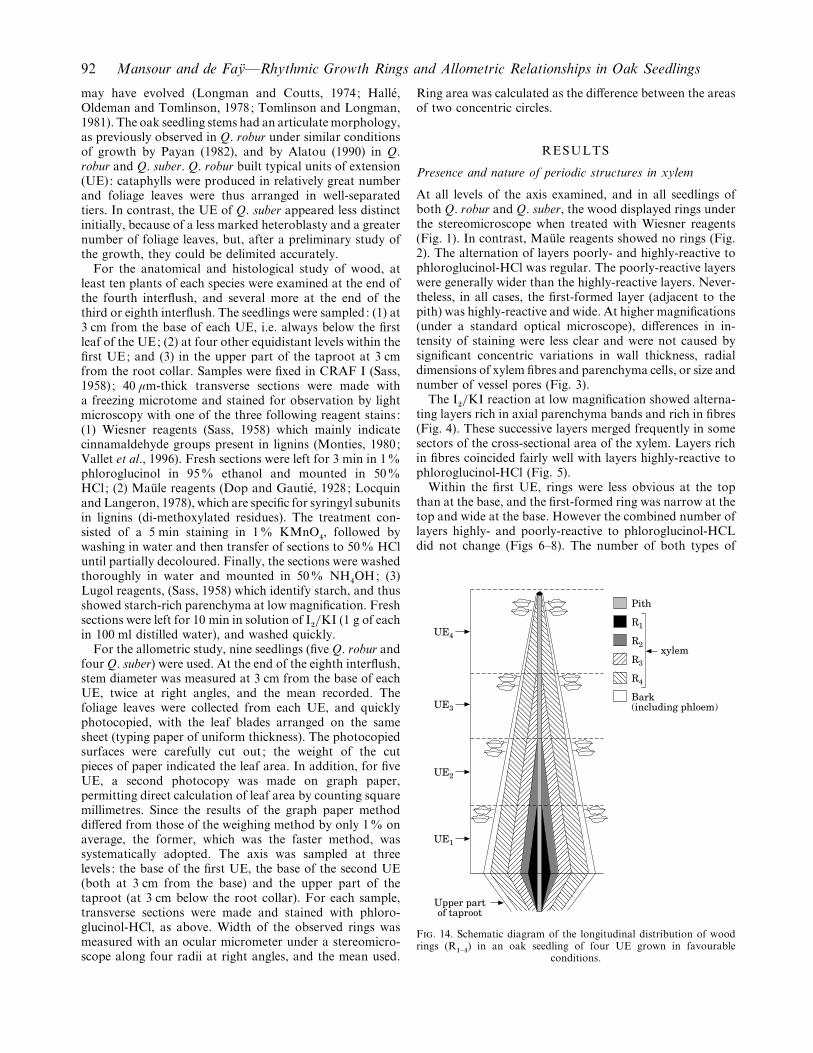
Within the first UE, rings were less obvious at the topthan at the base, and the first-formed ring was narrow at thetop and wide at the base. However the combined number oflayers highly- and poorly-reactive to phloroglucinol-HCLdid not change (Figs 6–8). The number of both types of
Upper partof taproot
Bark(including phloem)
UE2
UE1
UE3
UE4
R4
R3
R2
R1
Pith
xylem
F. 14. Schematic diagram of the longitudinal distribution of woodrings (R
"–%) in an oak seedling of four UE grown in favourable
conditions.

Mansour and de FayX —Rhythmic Growth Rings and Allometric Relationships in Oak Seedlings 93
T 1. Values of correlation coefficients (per indi�idual) between leaf area abo�e an axis le�el and stem diameter, ring widthand ring area at that le�el, in Q. robur and Q. suber seedlings of eight UE grown in fa�ourable conditions. Note that leaf area
corresponding to ring width and ring area was that abo�e the le�el gi�en at the presumed time of formation of the ring
Above-level leaf area to
Stem diameter Ring width Ring area
Levels examinedBase of
different UEBase offirst UE
Base ofsecond UE
Upper partof taproot
Base offirst UE
Base ofsecond UE
Q. robur seedling 1 0±971** 0±970** 0±961** 0±986** 0±946** 0±998**2 0±967** 0±722* 0±880** 0±992** 0±978** 0±993**3 0±977** 0±871** 0±980** 0±825* 0±988** 0±991**4 0±974** 0±699 0±964** 0±975** 0±950** 0±947**5 0±966** 0±839** 0±986** 0±981** 0±989** 0±994**
Q. suber seedling 1 0±962** 0±954** 0±977** 0±918** 0±931** 0±939**2 0±953** 0±909** 0±922** 0±975** 0±992** 0±999**3 0±982** 0±920** 0±956** 0±979** 0±950** 0±961**4 0±994** 0±646 0±808* 0±941** 0±975** 0±996**
Levels of significance of the difference from zero: **P! 0±01, *P! 0±05.
L1
R4R3R2R1Pith
xylem
Bark
T Time
I
II
III
IV
L2
L3
L4
UE4
UE3
UE2
UE1
F. 15. Schematic diagram of the evolution of the external and internal structures of the axis in an oak seedling. Longitudinal profiles andtransverse sections at the base of the first UE (or in the upper part of the taproot). R
", R
#, R
$, R
%, Cross sectional area of wood rings from pith
to bark; L", L
#, L
$, L
%, Leaf area for UE
", UE
#, UE
$, UE
%, respectively ; T, I, II, III, IV, Levels of observation of the upper part of the taproot
and base of the UE", UE
#, UE
$, UE
%, respectively.
layers was always the same at the upper part of taprootsand in the first-formed out of four UE, with four layershighly reactive to phloroglucinol-HCl. There were three atthe base of the second UE, two at the base of the thirdUE, and one at the base of the fourth and last-formedUE (Figs 9–13). The distribution of wood rings withinthe axis of an oak seedling of four UE can thus berepresented schematically as in Fig. 14.
Allometric relationships in the plant
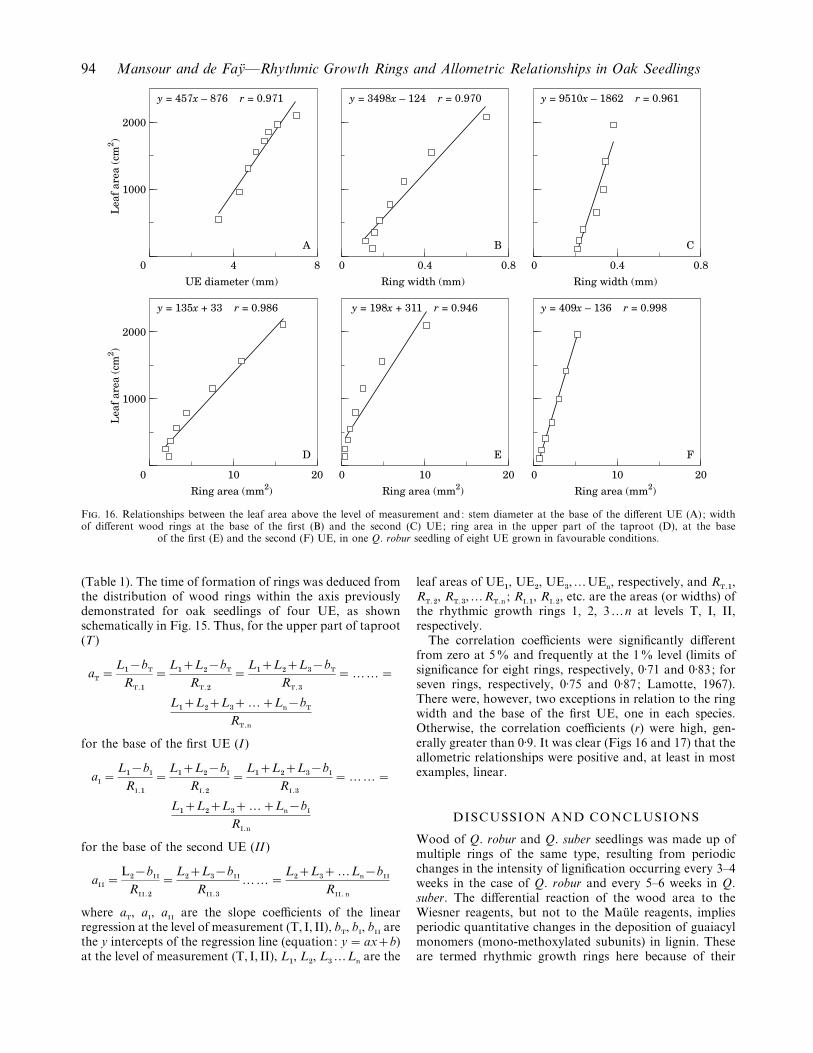
In seedlings of Q. robur and Q. suber, stem diameter wasfound to be highly correlated with the leaf area above thelevel of measurement (Table 1). Moreover at the three levelsof the axis studied, the width and area of a given ring werefound to be highly correlated with the leaf area above thelevel of measurement at the time of formation of the ring
94 Mansour and de FayX —Rhythmic Growth Rings and Allometric Relationships in Oak Seedlings
20
2000
Ring area (mm2)
Lea
f ar
ea (
cm2 )
1000
100
y = 135x + 33 r = 0.986
D
20
Ring area (mm2)
100
y = 198x + 311 r = 0.946
E
20
Ring area (mm2)
100
y = 409x – 136 r = 0.998
F
8
2000
UE diameter (mm)
Lea
f ar
ea (
cm2 )
1000
40
y = 457x – 876 r = 0.971
A
0.8
Ring width (mm)
0.40
y = 3498x – 124 r = 0.970
B
0.8
Ring width (mm)
0.40
y = 9510x – 1862 r = 0.961
C
F. 16. Relationships between the leaf area above the level of measurement and: stem diameter at the base of the different UE (A); widthof different wood rings at the base of the first (B) and the second (C) UE; ring area in the upper part of the taproot (D), at the base
of the first (E) and the second (F) UE, in one Q. robur seedling of eight UE grown in favourable conditions.
(Table 1). The time of formation of rings was deduced fromthe distribution of wood rings within the axis previouslydemonstrated for oak seedlings of four UE, as shownschematically in Fig. 15. Thus, for the upper part of taproot(T )
aT¯
L"®b
T
RT."
¯L
"L
#®b
T
RT.#
¯L
"L
#L
$®b
T
RT.$
¯……¯
L"L
#L
$…L
n®b
T
RT.n
for the base of the first UE (I )
aI¯
L"®b
I
RI."
¯L
"L
#®b
I
RI.#
¯L
"L
#L
$®b
I
RI.$
¯……¯
L"L
#L
$…L
n®b
I
RI.n
for the base of the second UE (II )
aII
¯L
#®b
II
RII.#
¯L
#L
$®b
II
RII.$
……¯L
#L
$…L
n®b
II
RII. n
where aT, a
I, a
IIare the slope coefficients of the linear
regression at the level of measurement (T, I, II), bT, b
I, b
IIare
the y intercepts of the regression line (equation: y¯ axb)at the level of measurement (T, I, II), L
", L
#, L
$…L
nare the
leaf areas of UE", UE
#, UE
$,…UE
n, respectively, and R
T.",
RT.#
, RT.$
,…RT.n
; RI."
, RI.#
, etc. are the areas (or widths) ofthe rhythmic growth rings 1, 2, 3…n at levels T, I, II,respectively.
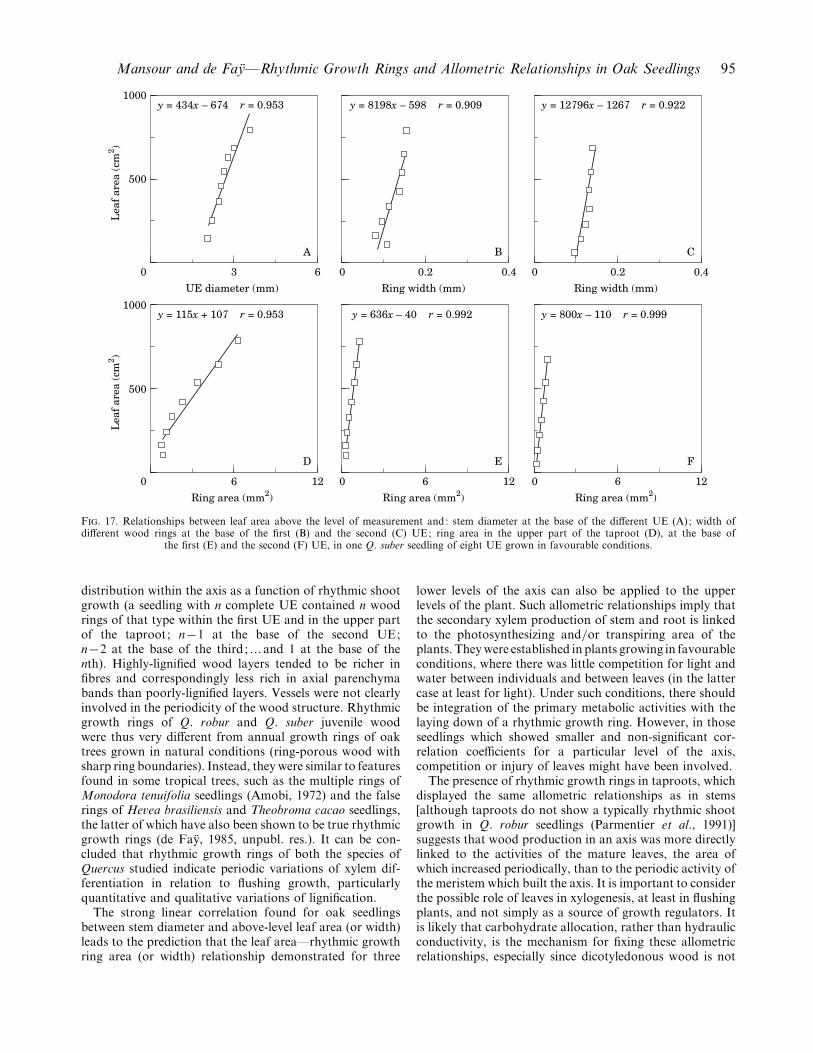
The correlation coefficients were significantly differentfrom zero at 5% and frequently at the 1% level (limits ofsignificance for eight rings, respectively, 0±71 and 0±83; forseven rings, respectively, 0±75 and 0±87; Lamotte, 1967).There were, however, two exceptions in relation to the ringwidth and the base of the first UE, one in each species.Otherwise, the correlation coefficients (r) were high, gen-erally greater than 0±9. It was clear (Figs 16 and 17) that theallometric relationships were positive and, at least in mostexamples, linear.
DISCUSSION AND CONCLUSIONS
Wood of Q. robur and Q. suber seedlings was made up ofmultiple rings of the same type, resulting from periodicchanges in the intensity of lignification occurring every 3–4weeks in the case of Q. robur and every 5–6 weeks in Q.suber. The differential reaction of the wood area to theWiesner reagents, but not to the Mau$ le reagents, impliesperiodic quantitative changes in the deposition of guaiacylmonomers (mono-methoxylated subunits) in lignin. Theseare termed rhythmic growth rings here because of their
Mansour and de FayX —Rhythmic Growth Rings and Allometric Relationships in Oak Seedlings 95
12
1000
Ring area (mm2)
Lea
f ar
ea (
cm2 )
500
60
y = 115x + 107 r = 0.953
D
12
Ring area (mm2)
60
y = 636x – 40 r = 0.992
E
12
Ring area (mm2)
60
y = 800x – 110 r = 0.999
F
6
1000
UE diameter (mm)
Lea
f ar
ea (
cm2 )
500
30
y = 434x – 674 r = 0.953
A
0.4
Ring width (mm)
0.20
y = 8198x – 598 r = 0.909
B
0.4
Ring width (mm)
0.20
y = 12796x – 1267 r = 0.922
C
F. 17. Relationships between leaf area above the level of measurement and: stem diameter at the base of the different UE (A); width ofdifferent wood rings at the base of the first (B) and the second (C) UE; ring area in the upper part of the taproot (D), at the base of
the first (E) and the second (F) UE, in one Q. suber seedling of eight UE grown in favourable conditions.
distribution within the axis as a function of rhythmic shootgrowth (a seedling with n complete UE contained n woodrings of that type within the first UE and in the upper partof the taproot; n®1 at the base of the second UE;n®2 at the base of the third;…and 1 at the base of thenth). Highly-lignified wood layers tended to be richer infibres and correspondingly less rich in axial parenchymabands than poorly-lignified layers. Vessels were not clearlyinvolved in the periodicity of the wood structure. Rhythmicgrowth rings of Q. robur and Q. suber juvenile woodwere thus very different from annual growth rings of oaktrees grown in natural conditions (ring-porous wood withsharp ring boundaries). Instead, they were similar to featuresfound in some tropical trees, such as the multiple rings ofMonodora tenuifolia seedlings (Amobi, 1972) and the falserings of He�ea brasiliensis and Theobroma cacao seedlings,the latter of which have also been shown to be true rhythmicgrowth rings (de Fay$ , 1985, unpubl. res.). It can be con-cluded that rhythmic growth rings of both the species ofQuercus studied indicate periodic variations of xylem dif-ferentiation in relation to flushing growth, particularlyquantitative and qualitative variations of lignification.
The strong linear correlation found for oak seedlingsbetween stem diameter and above-level leaf area (or width)leads to the prediction that the leaf area—rhythmic growthring area (or width) relationship demonstrated for three
lower levels of the axis can also be applied to the upperlevels of the plant. Such allometric relationships imply thatthe secondary xylem production of stem and root is linkedto the photosynthesizing and}or transpiring area of theplants. Theywere established in plants growing in favourableconditions, where there was little competition for light andwater between individuals and between leaves (in the lattercase at least for light). Under such conditions, there shouldbe integration of the primary metabolic activities with thelaying down of a rhythmic growth ring. However, in thoseseedlings which showed smaller and non-significant cor-relation coefficients for a particular level of the axis,competition or injury of leaves might have been involved.
The presence of rhythmic growth rings in taproots, whichdisplayed the same allometric relationships as in stems[although taproots do not show a typically rhythmic shootgrowth in Q. robur seedlings (Parmentier et al., 1991)]suggests that wood production in an axis was more directlylinked to the activities of the mature leaves, the area ofwhich increased periodically, than to the periodic activity ofthe meristem which built the axis. It is important to considerthe possible role of leaves in xylogenesis, at least in flushingplants, and not simply as a source of growth regulators. Itis likely that carbohydrate allocation, rather than hydraulicconductivity, is the mechanism for fixing these allometricrelationships, especially since dicotyledonous wood is not

96 Mansour and de FayX —Rhythmic Growth Rings and Allometric Relationships in Oak Seedlings
homogeneous in structure (conducting elements are presentnext to fibres and parenchyma cells).
Several allometric relationships have been studied inforest trees (Pressler’s law, Huber value, Pipe model, LeafArea—Sapwood Area relationship, in Larson, 1963;Shinozaki et al., 1964; Waring, Schroeder and Oren, 1982;Marchand 1984; Dean and Long, 1986; Espinosa Bancalari,Perry and Marshall, 1987; Keane and Weetman, 1987;Deleuze, 1996; Bartelink, 1997; Hatsh, 1997). These relate toannual rings of branched individuals and are often empirical,or not proved, in all species, notably in broad-leaved trees.It would be useful to know whether allometric relationships,analogous with those established in seedlings of flushingspecies, exist in forest trees.
LITERATURE CITED
Alatou D. 1990. Recherches sur le deU terminisme de la croissance
rythmique du cheW ne : Quercus pedunculata Ehrh—Quercus
mirbeckii Durieu—Quercus suber L.—eU tude morphologique et
eU cophysiologique. PhD. Thesis. University of Constantine,
Constantine, Algeria.
Amobi CC. 1972. Multiple bud growth and multiple wood formation in
the seedlings of Monodora tenuifolia Benth. Annals of Botany 36 :
199–205.
Bartelink HH. 1997. Allometric relationships for biomass and leaf area
of beech (Fagus syl�atica L.). Annales des Sciences Forestie[ res 54 :
39–50.
de Fay$ E. 1985. Recherche de marqueurs du mode de croissance
cambiale chez He�ea brasiliensis (Euphorbiaceae) : les anneaux
de croissance rythmique du bois juve!nile. Bulletin du MuseUum
national d ’Histoire naturelle, Paris, 4 e[ m serie 7 : 139–154.
Dean TJ, Long JN. 1986. Variations in sapwood area—leaf area
relations within two stands of lodgepole pine. Forest Science 32 :
749–758.
Deleuze C. 1996. Pour une dendromeU trie fonctionnelle : essai sur
l’inteUgration des connaissances eU cophysiologiques dans les mode[ lesde production ligneuse. PhD Thesis. University of Lyon, I, Lyon,
France.
Dop P, Gautie! A. 1928. Manuel de technique botanique histologique et
microbie. Second edition. Paris : J Lamarre.
Espinosa Bancalari MA, Perry DA, Marshall JD. 1987. Leaf area—
sapwood area relationships in adjacent young Douglas-fir stands
with different early growth rates. Canadian Journal of Forest
Research 17 : 174–180.
Grudzinskaya IA. 1957. The formation of false annual rings in Quercus
robur L., as affected by summer sprouts. Doklady Akademii Nauk
SSSR 115 : 392–395.
Guard AT, Postlethwait SN. 1958. Relation of the formation of annual
rings to multiple flushes of growth in several species of Quercus.
Indiana Academy of Science 67 : 104–106.
Halle! F, Martin R. 1968. Etude de la croissance rythmique chez l’he! ve! a(He�ea brasiliensis Mu$ ll-Arg. Euphorbiace! es-crotonoı$de! es).Adansonia se! rie 2, 8 : 475–503.
Halle! F, Oldeman RAA, Tomlinson PB. 1978. Tropical trees and forests:an architectural analysis. Heidelberg: Springer-Verlag.
Hatsch E. 1997. ReUpartition de l’aubier et acquisition de la forme de latige chez le cheW ne sessile (Quercus petraea) : analyse, modeU lisation etrelation a�ec le deU �eloppement du houppier. PhD Thesis ENGREF-Nancy, Nancy, France.
Keane MG, Weetman GF. 1987. Leaf area—sapwood cross sectionalarea relationships in repressed stands of lodgepole pine. CanadianJournal of Forest Research 17 : 205–209.
Kozlowski TT. 1971. Growth and de�elopment of trees. Vol. II: Cambialgrowth, root growth and reproducti�e growth. New York, London:Academic Press.
Lamotte M. 1967. Initiation aux meU thodes statistiques en biologie.Second edition. Paris : Masson et Cie.
Larson PR. 1963. Stem form development of forest trees. ForestScience-Monograph 5 : 1–41.
Lavarenne-Allary S. 1965. Recherches sur la croissance des bourgeonsde Che# ne et de quelques autres espe' ces ligneuses. Annales desSciences Forestie[ res 22 : 1–203.
Locquin M, Langeron M. 1978. Manuel de microscopie. Paris : Masson.Longman KA, Coutts MP. 1974. Physiology of the oak tree. In: Morris
MG, Perring FH, eds. The British oak. Faringdon, England:BSBI}E. W. Classey Ltd, 194–221.
Marchand PJ. 1984. Sapwood area as an estimator of foliage biomassand projected leaf area for Abies balsamea and Picea rubens.Canadian Journal of Forest Research 14 : 85–87.
Monties B. 1980. Les polyme[ res �eUgeU taux. Polyme[ res parieU taux etalimentaires non azoteU s. Paris : Bordas.
Parmentier C, Barnola P, Maillard P, Lavarenne S. 1991. E! tude de lacroissance rythmique du che# ne pe!doncule! . Influence du syste' meracinaire. Colloque International ‘‘ l’arbre’’, Montpellier, France.Naturalia monspeliensa, ‘‘l’arbre, biologie et de! veloppement ’’Edelin C, ed: 327–343.
Payan E. 1982. Contribution a[ l’eU tude de la croissance rythmique desjeunes cheW nes peUdonculeU s (Quercus pedunculata Ehrh). PhD Thesis.University of Clermont-Ferrand II, Clermont-Ferrand, France.
Sass JE. 1958. Botanical microtechnique. Third edition. Ames, Iowa:The Iowa State University Press.
Shinozaki K, Yoda K, Hozumi K, Kira T. 1964. A quantitative analysisof plant form—the pipe model theory—I. Basic analyses. JapaneseJournal of Ecology 14 : 97–105.
Tomlinson PB, Longman, KA. 1981. Growth phenology of tropical treesin relation to cambial activity. In: Bormann FH, Berlyn G, eds.Age and growth rate of tropical trees: new directions for research.Yale University, New Haven: School of Forestry and EnvironmentStudies, Bulletin 94 : 7–19.
Vallet C, Chabbert B, Czaninski Y, Monties B. 1996. Histochemistry oflignin deposition during sclerenchyma differentiation in alfalfastems. Annals of Botany 78 : 625–632.
Waring RH, Schroeder PE, Oren R. 1982. Application of the pipemodel theory to predict canopy leaf area. Canadian Journal ofForest Research 12 : 556–560.
Wight W. 1930. Secondary elongation growth in oaks, 1929. Naturalist,Hull: 65–70.