reversal of the cd4 /cd8 t-cell ratio in lymph node cells ... · rauf latif,1‡ nicole kerlero de...
TRANSCRIPT

INFECTION AND IMMUNITY,0019-9567/01/$04.0010 DOI: 10.1128/IAI.69.5.3073–3081.2001
May 2001, p. 3073–3081 Vol. 69, No. 5
Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Reversal of the CD41/CD81 T-Cell Ratio in Lymph Node Cellsupon In Vitro Mitogenic Stimulation by Highly Purified,
Water-Soluble S3-S4 Dimer of Pertussis Toxin†RAUF LATIF,1‡ NICOLE KERLERO DE ROSBO,1 TANY AMARANT,2 RINO RAPPUOLI,3
GREGOR SAPPLER,1 AND AVRAHAM BEN-NUN1*
Department of Immunology, The Weizmann Institute of Science,1 and ProSpec-TechnoGene, Weizmann Science Park,2
Rehovot, Israel, and Immunobiological Research Institute Siena (IRIS), 53100 Siena, Italy3
Received 1 December 2000/Returned for modification 4 January 2001/Accepted 19 February 2001
Pertussis toxin (PT), a holomer consisting of a catalytic S1 subunit and a B oligomer composed of S2-S4 andS3-S4 dimers, held together by the S5 subunit, exerts profound effects on immune cells, including T-cellmitogenicity. While the mitogenic activity of PT was shown to reside fully within the B oligomer, it could notbe assigned to any particular B-oligomer component. In this study, we purified the S3-S4 dimer to homogeneityunder conditions propitious to maintenance of the native conformation. In contrast to previous reports whichsuggested that both S3-S4 and S2-S4 dimers are necessary for mitogenic activity, our preparation of the highlypurified S3-S4 dimer was as strongly mitogenic as the B oligomer, suggesting that the S3-S4 dimer accountsfor the mitogenic activity of the B oligomer. Moreover, in vitro stimulation of naive lymphocytes by the S3-S4dimer resulted in reversal of the normal CD41/CD81 T-cell ratio from approximately 2:1 to 1:2. The reversalof the CD41/CD81 T-cell ratio is unlikely to be due to preferential apoptosis-necrosis of CD41 T cells, asindicated by fluorescence-activated cell sorter analysis of annexin-stained T-cell subsets, or to preferentialstimulation of CD81 T cells. The mechanism underlying the reversal requires further investigation. Never-theless, the data presented indicate that the S3-S4 dimer may have potential use in the context of diseasesamenable to immunological modulation.
Pertussis toxin (PT), the major virulence determinant ofBordetella pertussis (35), is composed of two distinct functionalunits, the A protomer, consisting of a single polypeptide (S1)which mediates adenosine diphosphate ribosylation of host Gproteins, and the B oligomer, which mediates the binding ofthe toxin to host cells and the translocation of toxic S1 to itstarget (8, 28). The B oligomer is a complex pentamer com-posed of subunits S2, S3, S4, and S5 in a respective molar ratioof 1:1:2:1, with S2 and S3 occuring as heterodimers each withS4, i.e., dimer S2-S4 and dimer S3-S4, held together by S5 (28,32). The effects of the toxin on cells of the immune system aremultiple and include induction of lymphocytosis, inhibition ofmacrophage migration, adjuvant activity, and T-cell mitogenic-ity (18). A number of the biological activities of PT, such aslymphocytosis and adjuvant activity, implicate the enzymaticactivity of PT in its toxicity and can be abrogated by inactiva-tion of the S1 subunit (1, 5, 13). In contrast, PT-associatedT-cell mitogenicity is mediated by the B oligomer (9, 31, 36)and appears to be independent of the enzymatic activity of thetoxin, as inactivation of the S1 subunit by mutation has noeffect on the mitogenic activity of PT (13, 36), while alterationsin the B oligomer can totally abrogate the mitogenic activity ofPT (15, 16, 20–22). In addition, the B oligomer devoid of S1
induces T-cell proliferation to the same extent as PT (22, 29,31, 32, 36). However, although the mitogenic effect of the Boligomer is well known, the roles of its individual componentsin the mitogenic function have not been extensively studied.Both dimers have been implicated in the binding of PT to cellsvia interaction of the B oligomer with glycoproteins and gly-colipids on many types of eukaryotic cells (10, 26, 37), seem-ingly via carbohydrate-recognizing domains on subunits S2 andS3 (11, 26, 34, 37). However, there is evidence of a differencebetween the binding specificities of the two dimers (26, 37),which may account for observations leading to the suggestionthat the B oligomer must bind to the cell surface to permittranslocation of the A protomer into the cell in a mannerdifferent from its binding leading to T-cell mitogenicity, sincethese two types of binding displayed different susceptibilities tochemical modification of the molecule (22; see Discussion).Experiments with hybrid PTs composed of various combina-tions of chemically modified dimers indicate a differential roleof the two dimers in T-cell stimulation and suggest that theS3-S4 dimer is more relevant to the binding of the B oligomerwhich results in T-cell stimulation than to the binding whichresults in translocation of the S1 subunit (20–22). A mitogeniceffect for the S3-S4 dimer as an isolated dimeric molecule wasnot demonstrated.
We have been investigating PT as an immunomodulatoryagent for experimental autoimmune encephalomyelitis (EAE),the well-accepted animal model for multiple sclerosis, andfound that the protective effect imparted by PT against EAEcould be fully attributed to the B oligomer (3). To understandthe mechanism by which the B oligomer immunomodulates
* Corresponding author. Mailing address: Dept. of Immunology,The Weizmann Institute of Science, Rehovot 76100, Israel. Phone:972-8-9342991. Fax: 972-8-9344141. E-mail: [email protected].
† This paper is dedicated to Tany Amarant, who passed away sud-denly on 6 March 2000.
‡ Present address: Mount Sinai Medical Center, New York, NY10128.
3073
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

EAE, the role of its components in the biological activity of theB oligomer should be investigated.
In the B oligomer, S2 and S3 occur naturally as parts ofheterodimers, each with S4. These dimers can only be dissoci-ated into monomers under strong denaturing conditions, sug-gesting that the dimeric conformation is functionally impor-tant. Isolation of functional dimers is necessary if we are toinvestigate their role in the biological activities of the B oli-gomer. We have now purified the S3-S4 dimer to homogeneityunder conditions which favor preservation of the native con-formation and investigated its biological activity in vitro. Weshow that the S3-S4 dimer is as strong a mitogen for T cells asis the B oligomer, in contrast to previous reports suggestingthat a combination of S2-S4 and S3-S4 dimers is absolutelynecessary to induce T-cell proliferation and that neither dimeris mitogenic by itself (31, 36). The heterodimeric moleculeappears to be essential for the mitogenic activity, as recombi-nant preparations of individual subunits could not significantlyinduce T-cell proliferation. Most striking was the in vitro mod-ulatory effect of the purified S3-S4 dimer, as well as that of theB oligomer, in reversing the CD41/CD81 T-cell ratio uponstimulation of naive lymph node (LN) cells.
MATERIALS AND METHODS
PT and recombinant subunits. PT was obtained commercially (catalog no. P9452; Sigma, St. Louis, Mo.). Mutant PT (PTmu), a genetically derived mutantof PT, PTX-9K/129G, was prepared as described previously (24). This mutantcontains two amino acid substitutions, Arg9 with Lys and Glu129 with Gly, in theS1 subunit which abolish its enzymatic activity (24); the B oligomer is unaffected,and PTmu retains its mitogenic activity (13; this study). The DNAs correspond-ing to PT subunits S2, S3, and S4 cloned into expression vector pEx31 or pEx34(19) were subcloned into expression vector pRSET (Invitrogen Corporation, SanDiego, Calif.) after amplification by PCR using specific primers in which NheIand BamHI restriction sites or NheI and BglII restriction sites had been includedto enable ligation at the 59 and 39 ends of S2 and S4 or S3, respectively. Theprimers used were as follows: 59 S2 primer, CATGGTATGGCTAGCACGCCAGGCATCGTCATTCCG; 39 S2 primer, GCAGCCGGATCCTCAGCATAAGGATGATCCAGGATT; 59 S4 primer, CATGGTATGGCTAGCGACGTTCCTTATGTGCTG; 39 S4 primer, GAGCTCGGATCCTCAGGGGCACTGCTTGCCGCT; 59 S3 primer, CATGGTATGGCTAGCGTTGCGCCAGGCATCGTCATC; 39 S3 primer, TGAGCTCAGATCTCAGCATATCGACGCTGCCGGGTT. The nucleotide sequence of the PCR product within each construct wasconfirmed by direct sequencing using primers derived from pRSET (forwardprimer, 59-ATGCGGGGTTCTCATCAT-39; reverse primer, 59-TAGCAGCCGGATCAAGCT-39) and a 373A DNA sequencer (Applied Biosystems, FosterCity, Calif.). The DNA sequences obtained confirmed for each construct an openreading frame for the relevant subunit, preceded by (Met)-Arg-Gly-Ser-(His)6-Gly-Met-Ala-Ser. Expression of the recombinant subunits was induced in thehost Escherichia coli BL21(DE3) (catalog no. C6000-03; Invitrogen Corpora-tion), and the recombinant protein was purified to homogeneity by metal chelateaffinity chromatography on Ni21 nitrilotriacetic acid-agarose (catalog no. 30230;Qiagen, Chatsworth, Calif.) according to the manufacturer’s protocol.
Preparation of antisera against PT and recombinant subunits. Antiseraagainst PT and the recombinant S2 subunit (rS2) were prepared in SJL/J mice bysubcutaneous injection of PT (500 ng of PT in incomplete Freund adjuvant) orpurified rS2 (50 mg of rS2 in complete Freund adjuvant), followed by two boosterinjections of 25 mg of rS2 administered subcutaneously at weekly intervals in theflank. The anti-PT serum which reacts predominantly with the S1 subunit (seeResults) is henceforth referred to as anti-S1 serum. Rabbit antisera were pre-pared against rS3 and rS4 by standard procedures.
Separation of PT components by HPLC. PTmu was dissociated by incubationin 5 M cold urea by the procedure of Tamura et al. (32), which yields productscorresponding to the S1 and S5 subunits and the S2-S4 and S3-S4 dimers (32).The dissociated PTmu (100 mg in 500 ml of 5 M urea) was separated by high-pressure liquid chromatography (HPLC) using a Spectra Physics SP8750 HPLCsystem and a Superdex 75 HR 10/30 gel filtration column equilibrated with 100mM phosphate buffer, pH 7.0, containing 25 mM NaCl. Elution (flow rate, 0.5
ml/min; 264 lb/in2) was carried out in the same buffer. Protein elution wasmonitored at 220 nm with a Waters model 441 detector. The protein concen-tration was estimated according to the area of the peaks.
SDS-PAGE and Western blotting. Sodium dodecyl sulfate-polyacrylamide gelelectrophoresis (SDS-PAGE) was carried out according to the procedure ofLaemmli (14) on 12% gels (henceforth referred to as “Laemmli gels”) or ac-cording to the procedure of Schagger and von Jagow (27) on 10% gels (hence-forth referred to as “Tricine gels”). All gels were stained with silver. Westernblotting onto nitrocellulose membrane was carried out according to the proce-dure of Towbin et al. (33). Dilutions of 1:500, 1:1,000, 1:8,000, and 1:10,000 wereused for anti-S1, anti-S2, anti-S3, and anti-S4 primary antisera, respectively.Reactive bands were detected by enhanced chemiluminescence (ECL) using anECL analysis system (catalog no. RPN 2108; Amersham, Amersham, England)according to the manufacturer’s protocol.
Slot blot analysis for the presence of PT dimers. A glycoprotein bindingexperiment to assay for the presence of PT subunits in the form of dimers in therelevant peak was essentially done according to the procedure of Witvliet et al.(37), with minor modifications. Briefly, proteins resuspended in phosphate-buff-ered saline (PBS) were applied directly to nitrocellulose using a Bio-Rad Bio-Dot SF microfiltration apparatus (catalog no. 170–6542). After blocking for 2 hin bovine serum albumin (BSA) solution (3% BSA in PBS), the blots wereincubated with the various preparations of PT and PT components as indicatedin Results, washed with PBS-Tween (37), and reacted with anti-S4 serum; bind-ing by anti-S4 serum was detected by ECL as described above.
In vitro assay of the mitogenic activity of PT and PT components on naive andcommitted T cells. LN cells from naive SJL/J mice were cultured in microtiterwells (5 3 105 cells/well) as described previously (2), in the presence of PT, PTcomponents, or concanavalin A (ConA), as indicated. The mitogenic effect of PTand PT components on committed T cells was tested on mouse line PL/J T cellsspecific for pMOG35-55, a myelin oligodendrocyte glycoprotein peptide encom-passing amino acids 35 to 55 of the molecule (12). The pMOG35-55-specific Tcells (104 cells/well) were cultured as described above, in the presence of irra-diated (2,500 rads) syngeneic spleen cells (5 3 105 cells/well) from naive PL/Jmice. The cells were incubated for 72 h (naive LN cells) or 48 h (pMOG35-55-specific T cells) at 37°C in humidified air containing 7.5% CO2. [3H]thymidine (1mCi/well) was added for the last 16 h of incubation, and the cultures wereharvested and counted using a Matrix 96 Direct beta counter (Packard Instru-ments, Meriden, Conn.). The proliferative response was measured as [3H]thy-midine incorporation expressed as mean counts per minute of triplicate cultures.
Cytofluorometric analysis of T-cell subsets in naive LN cells stimulated withPTmu, PT components, and other T-cell mitogens. Naive LN cells (5 3 106
cells/ml) from SJL/J mice were cultured as described previously (2), in thepresence of PTmu (250 ng/ml), peak II (see Results) (2 mg/ml), ConA (500ng/ml), or an anti-CD3 monoclonal antibody (MAb) (30 ml of a 1:2 dilution ofsupernatant from hybridoma hamster anti-mouse CD3, clone no. 145-2C11, fromthe American Type Culture Collection). After 2 to 3 days, the cells were split inmedium containing interleukin-2. At the time intervals indicated in Results, thecells were analyzed by fluorescence-activated cell sorting (FACS) as describedpreviously (17), using the following MAbs: fluorescein isothiocyanate (FITC)-conjugated anti-mouse TCRab (H57-597; catalog no. 01304D; Pharmingen, SanDiego, Calif.); FITC-conjugated anti-mouse CD8 (YTS 169.4; catalog no.RM2201; Caltag, San Francisco, Calif.), and phycoerythrin-conjugated anti-mouse CD4 (catalog no. 1447; Becton Dickinson, Mountain View, Calif.).
Cytofluorometric analysis for apoptosis-necrosis with annexin. FACS analysisof apoptotic-necrotic CD41 and CD81 cells was performed by double stainingwith FITC-conjugated anti-CD4 (catalog no. 09424D; Pharmingen) or anti-CD8(as described above) MAb and Annexin-V-Alexa 568 (catalog no. 1985 485;Boehringer Mannheim/Roche, Mannheim, Germany) according to the manufac-turer’s instructions.
Depletion of CD81 and CD41 T cells from naive mice by anti-CD81 andanti-CD41 MAbs. Naive SJL/J mice were injected intraperitoneally on threeconsecutive days, 7 to 9 days before in vitro analysis of the mitogenic effect of PT,with a rat anti-mouse CD81 (YTS-169) (6) or CD41 (GK 1–5) (7) MAb (1 mlof a 1:10 dilution of ascites fluid per injection). The hybridomas, YTS-169 andGK1-5, secreting anti-CD8 and anti-CD4 Abs respectively, were a kind gift fromL. Eisenbach of the Department of Immunology, The Weizmann Institute ofScience. Depletion of the relevant T-cell subsets was monitored on the day of invitro culture with PT or mitogens by FACS analysis of the LN cell populationwith anti-CD81 and anti-CD41 MAbs as described above; the results obtainedshowed full depletion of CD41 T cells in mice treated with the anti-CD4 MAb(0.4% CD41 and 50% CD81 TCRab1 cells in the LN cell population) and ofCD81 T cells in mice treated with the anti-CD8 MAb (0.5% CD81 and 65%CD41 TCRab1 cells in the LN cell population).
3074 LATIF ET AL. INFECT. IMMUN.
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
RESULTS
Dissociation of PT components with urea and separation byHPLC. Native S2-S4 and S3-S4 dimers can be dissociated fromthe PT holomer by incubation of the holomer with 5 M ice-coldurea (32). To isolate the dimers, PTmu was incubated for 4days at 4°C in 0.05 M phosphate buffer (pH 6.0) containing 5M urea and the dissociation products were separated by gelfiltration on HPLC. As can be seen on the typical HPLCelution profile presented in Fig. 1A, three protein peaks wereeluted, which were further analyzed for PT components andthe presence of dimers.
Characterization of HPLC gel filtration peaks: isolation ofS3-S4 dimer. (i) SDS-PAGE. Because of its greater power ofseparation, the Tricine SDS-PAGE method of Schagger andvon Jagow (27) was used to analyze the HPLC fractions ob-tained from the dissociated PT (Fig. 1B). Although PT mi-grates as five bands in the Laemmli (14) SDS-PAGE system(32) (Fig. 1C), an additional band running close to the S3subunit could be observed on a 10% Tricine gel (Fig. 1C).Interestingly, the S3 subunit was demonstrated by Westernblotting with a highly specific anti-S3 Ab (see below) to run asa doublet in this gel system but not on a 12% Laemmli gel (Fig.1C). The significance of this observation is unclear. The elec-trophoretic pattern of peak I on the 10% Tricine gel (Fig. 1B)was similar to that of undissociated PT, except that the densityof the band corresponding in molecular size to S1 was de-creased, suggesting that peak I represents a mixture of the Boligomer and whole PT. A doublet corresponding to S3 andone band corresponding to S4 were seen in peak II (Fig. 1B).Peak III contained very small amounts of protein. Upon con-centration, most of the protein aggregated (Fig. 1B).
To ascertain the presence or absence of different subunits inpeaks I and II, rabbit or mouse polyclonal Abs were raised
against preparations of rS2, rS3, and rS4 and components ofthe peaks were analyzed by Western blotting.
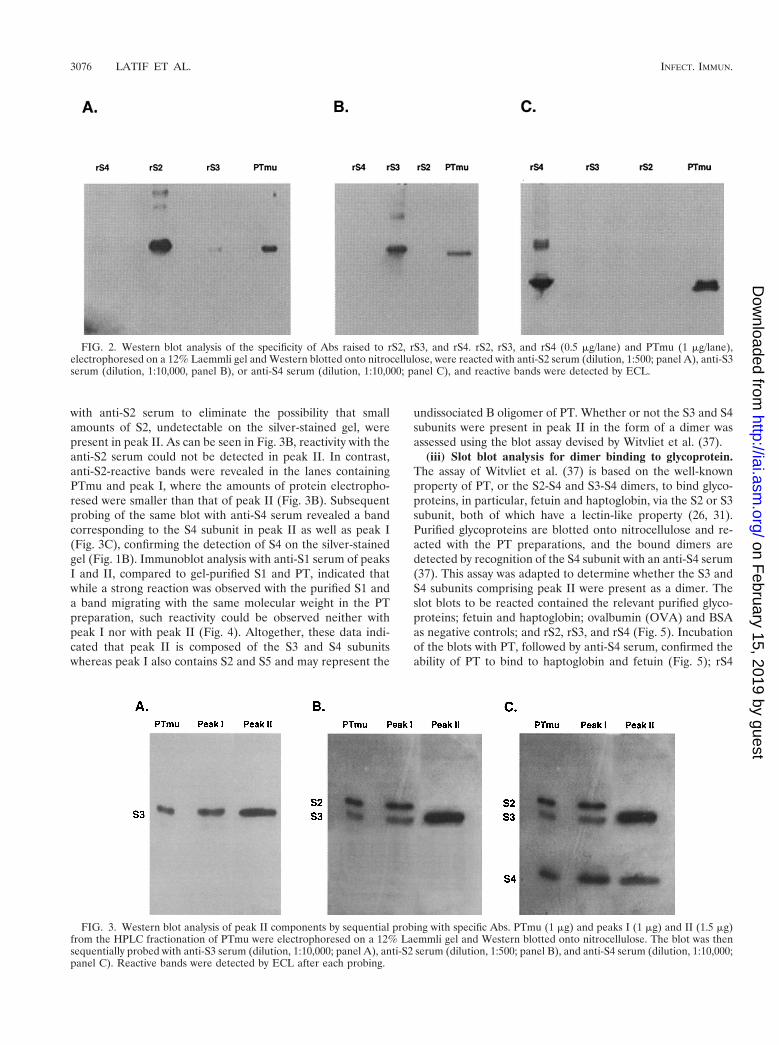
(ii) Western blotting. The specificity of the polyclonal Abs tobe used was determined by Western blotting of native subunitsof PTmu and of the recombinant proteins used for Ab produc-tion. The Laemmli SDS-PAGE system (14) was used through-out in Western blotting analyses to ascertain Ab specificity andto determine the composition of the HPLC fractions. On SDS-PAGE, rS2, rS3, and rS4 migrated with slightly higher molec-ular weights than the native subunits, due to the His tag addedto enable purification of the recombinant proteins by nickelaffinity chromatography. As can be seen in Fig. 2, single bandsreacted when native PT was analyzed with anti-S2 (Fig. 2A),anti-S3 (Fig. 2B), and anti-S4 (Fig. 2C) Abs, respectively, in-dicating that despite the high degree of homology between S2and S3 (19), the Abs raised recognize the native subunits in ahighly specific manner and can be used to analyze the HPLCpeaks for the presence or absence of the various subunits. Aslight cross-reactivity was observed when the Abs were testedagainst the recombinant subunits. Such cross-reactivity, ob-served mainly with the anti-S2 and anti-S4 Abs (see also be-low), is likely to be due to the presence, within the polyclonalantisera, of Abs which react to the common His tag portion ofthe recombinant proteins used for immunization. The presenceof S1 was tested with a mouse anti-PT serum which had showna predominant high reactivity to S1 (see below).
According to the electrophoretic profile, it is likely that peakII contains only S3 and S4, possibly in the form of a dimer. Toascertain the presence of S3, peak II, along with PTmu andpeak I, was analyzed by Western blotting with anti-S3 serum(Fig. 3A). Reaction of a single band migrating at the sameposition in all three samples confirmed the presence of S3 inpeak II, as well as in peak I. The same blot was then reprobed
FIG. 1. Fractionation of urea-dissociated PT by HPLC. Panels A, chromatographic profile; B, silver-stained 10% Tricine gel of PT and peaksI through III (protein amounts loaded: PT, 4 mg; peak I, 1 mg; peak II, 1.5 mg; peak III, 1.5 mg); C, electrophoresis profiles of PTmu on Tricineand Laemmli gels (lanes a) and corresponding Western blots (lanes b) indicating that the S3 subunit migrates as a doublet on Tricine gels but noton Laemmli gels. Lanes a are silver-stained gels, and lanes b are the corresponding Western blots probed with anti-S3 serum. Rabbit anti-S3 serumwas used at a dilution of 1:8,000, and reactive bands were detected by ECL. Two micrograms of PTmu was loaded on the gels. OD, optical density.
VOL. 69, 2001 CD41/CD81 T-CELL RATIO REVERSAL BY S3-S4 DIMER OF PT 3075
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
with anti-S2 serum to eliminate the possibility that smallamounts of S2, undetectable on the silver-stained gel, werepresent in peak II. As can be seen in Fig. 3B, reactivity with theanti-S2 serum could not be detected in peak II. In contrast,anti-S2-reactive bands were revealed in the lanes containingPTmu and peak I, where the amounts of protein electropho-resed were smaller than that of peak II (Fig. 3B). Subsequentprobing of the same blot with anti-S4 serum revealed a bandcorresponding to the S4 subunit in peak II as well as peak I(Fig. 3C), confirming the detection of S4 on the silver-stainedgel (Fig. 1B). Immunoblot analysis with anti-S1 serum of peaksI and II, compared to gel-purified S1 and PT, indicated thatwhile a strong reaction was observed with the purified S1 anda band migrating with the same molecular weight in the PTpreparation, such reactivity could be observed neither withpeak I nor with peak II (Fig. 4). Altogether, these data indi-cated that peak II is composed of the S3 and S4 subunitswhereas peak I also contains S2 and S5 and may represent the
undissociated B oligomer of PT. Whether or not the S3 and S4subunits were present in peak II in the form of a dimer wasassessed using the blot assay devised by Witvliet et al. (37).
(iii) Slot blot analysis for dimer binding to glycoprotein.The assay of Witvliet et al. (37) is based on the well-knownproperty of PT, or the S2-S4 and S3-S4 dimers, to bind glyco-proteins, in particular, fetuin and haptoglobin, via the S2 or S3subunit, both of which have a lectin-like property (26, 31).Purified glycoproteins are blotted onto nitrocellulose and re-acted with the PT preparations, and the bound dimers aredetected by recognition of the S4 subunit with an anti-S4 serum(37). This assay was adapted to determine whether the S3 andS4 subunits comprising peak II were present as a dimer. Theslot blots to be reacted contained the relevant purified glyco-proteins; fetuin and haptoglobin; ovalbumin (OVA) and BSAas negative controls; and rS2, rS3, and rS4 (Fig. 5). Incubationof the blots with PT, followed by anti-S4 serum, confirmed theability of PT to bind to haptoglobin and fetuin (Fig. 5); rS4
FIG. 2. Western blot analysis of the specificity of Abs raised to rS2, rS3, and rS4. rS2, rS3, and rS4 (0.5 mg/lane) and PTmu (1 mg/lane),electrophoresed on a 12% Laemmli gel and Western blotted onto nitrocellulose, were reacted with anti-S2 serum (dilution, 1:500; panel A), anti-S3serum (dilution, 1:10,000, panel B), or anti-S4 serum (dilution, 1:10,000; panel C), and reactive bands were detected by ECL.
FIG. 3. Western blot analysis of peak II components by sequential probing with specific Abs. PTmu (1 mg) and peaks I (1 mg) and II (1.5 mg)from the HPLC fractionation of PTmu were electrophoresed on a 12% Laemmli gel and Western blotted onto nitrocellulose. The blot was thensequentially probed with anti-S3 serum (dilution, 1:10,000; panel A), anti-S2 serum (dilution, 1:500; panel B), and anti-S4 serum (dilution, 1:10,000;panel C). Reactive bands were detected by ECL after each probing.
3076 LATIF ET AL. INFECT. IMMUN.
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
reacted strongly with the anti-S4 serum (a slight reactivity ofthe anti-S4 serum with rS2 and rS3 was observed as notedabove). No reactivity with OVA or BSA could be detected.When the blots were probed with peak II containing S3 and S4,a pattern of reactivity identical to that of blots probed with PTwas observed (Fig. 5), indicating that peak II contains the S3and S4 subunits as a dimer which binds to fetuin and hapto-globin via the S3 subunit and is detected by antigen-antibodyrecognition of the S4 subunit. The lower intensity of binding tofetuin by the S3-S4 dimer compared to PT is related to thepresence of the S2-S4 dimer in PT and its absence in peak II,as the S2-S4 dimer has a stronger affinity for fetuin than doesthe S3-S4 dimer (37). That the S4 subunit by itself does notbind to fetuin or haptoglobin and therefore could not be re-sponsible for the pattern observed with peak II is demon-strated in the blot probed with the rS4 subunit and the anti-S4Ab, which displayed a pattern commensurate only with reac-tivity of the anti-S4 Ab with rS4 (Fig. 5); no reactivity could beobserved with fetuin, haptoglobin, OVA, or BSA.
The S3-S4 dimer is mitogenic for T cells. The mitogeniceffect of PT is known to reside with the B oligomer (9, 31, 36).In a preliminary attempt to investigate the possibility that suchmitogenic activity of the B oligomer involves the S3-S4 dimer,the effect of peak II on naive and committed T cells wasassessed, compared to the effects of PTmu, the B oligomer,rS2, rS3, and rS4. As can be seen in Fig. 6, culture of naive LNcells from SJL/J mice in the presence of 250 ng of PT or the Boligomer resulted in extensive proliferation; the same amountof peak I or peak II induced an even stronger response, com-mensurate with the proliferative response to the T-cell mito-gen ConA. Naive LN cells did not proliferate in response to250 ng of rS2, rS3, or rS4 (Fig. 6) or in response to lowerconcentrations (50 and 100 ng/well) of the individual recom-binant subunits (data not shown).
The mitogenic effects of PTmu, peak I, and peak II couldalso be observed with PL/J line CD41 T cells highly specific forpMOG35-55 (Fig. 6), a myelin oligodendrocyte glycoproteinpeptide (12). The pMOG35-55-specific T cells proliferated tothe same extent in response to peak II as in response to peakI or PTmu, and such mitogenic proliferation was as strong asthe antigenic proliferation of the T cells in response to thepeptide (Fig. 6). The pMOG35-55-specific T-cells did not pro-liferate in the presence of any of the recombinant subunits.The mitogenic effect of peaks I and II on T cells was dose
dependent, reaching a plateau at a mitogen concentration of500 ng/well (data not shown).
Reversal of the CD41/CD81 T-cell ratio upon stimulation ofnaive LN cells with the S3-S4 dimer, as well as PTmu. LN cellsisolated from naive SJL/J mice were cultured (5 3 106 cells/ml)for 96 h in the presence of PTmu (250 ng/ml), the S3-S4 dimer(peak II) (2 mg/ml), ConA (500 ng/ml), or the anti-CD3 MAb(30 ml of a 1:2 dilution of culture supernatant/ml), and theproportion of TCRab1 T cells which expressed CD41, CD81,or CD41 CD81 was estimated by FACS analysis. As can beseen in Table 1, the ratio of CD41 to CD81 T cells in theTCRab1 LN cell population which were stimulated in vitrowith ConA or the anti-CD3 MAb (53 and 44% CD41 and 21and 20% CD81) did not differ significantly from the ratio ofCD41 to CD81 T cells estimated in naive TCRab1 cells (50%CD41 and 28.5% CD81), indicating that ConA and the anti-CD3 MAb have a nondiscriminatory mitogenic effect on Tcells. In contrast, the proportions of CD81 T cells and CD41
T cells changed dramatically following stimulation with PTmu(Table 1), resulting in a complete reversal of the CD41/CD81
ratio (19.9% CD41 and 65.9% CD81). A similar reversal ofthe CD41/CD81 ratio was obtained upon stimulation withnative PT or with the B oligomer (data not shown). As shownin Table 1, stimulation of naive LN cells with the S3-S4 dimer(peak II) also resulted in reversal of the CD41/CD81 ratio(24% CD41 and 58.2% CD81), indicating that the mitogenicS3-S4 dimer is sufficient to reproduce the effect of PTmu onthe CD41/CD81 ratio. Similar results were obtained whether
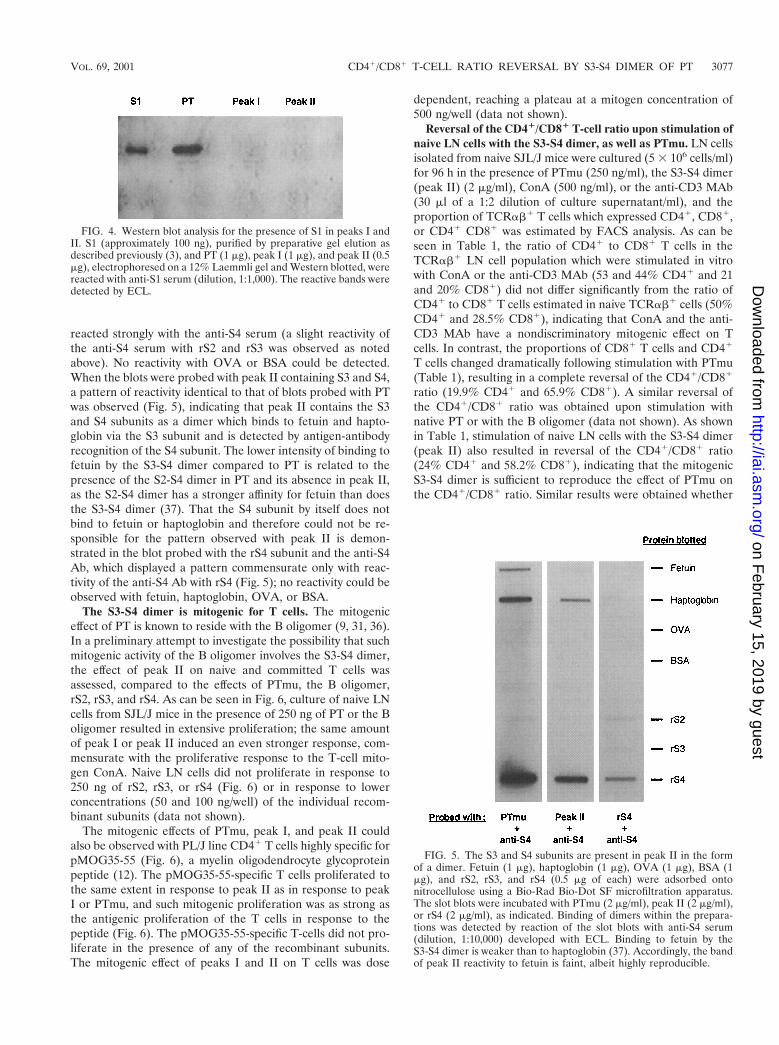
FIG. 4. Western blot analysis for the presence of S1 in peaks I andII. S1 (approximately 100 ng), purified by preparative gel elution asdescribed previously (3), and PT (1 mg), peak I (1 mg), and peak II (0.5mg), electrophoresed on a 12% Laemmli gel and Western blotted, werereacted with anti-S1 serum (dilution, 1:1,000). The reactive bands weredetected by ECL.
FIG. 5. The S3 and S4 subunits are present in peak II in the formof a dimer. Fetuin (1 mg), haptoglobin (1 mg), OVA (1 mg), BSA (1mg), and rS2, rS3, and rS4 (0.5 mg of each) were adsorbed ontonitrocellulose using a Bio-Rad Bio-Dot SF microfiltration apparatus.The slot blots were incubated with PTmu (2 mg/ml), peak II (2 mg/ml),or rS4 (2 mg/ml), as indicated. Binding of dimers within the prepara-tions was detected by reaction of the slot blots with anti-S4 serum(dilution, 1:10,000) developed with ECL. Binding to fetuin by theS3-S4 dimer is weaker than to haptoglobin (37). Accordingly, the bandof peak II reactivity to fetuin is faint, albeit highly reproducible.
VOL. 69, 2001 CD41/CD81 T-CELL RATIO REVERSAL BY S3-S4 DIMER OF PT 3077
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

interleukin-2 was present or not for the last 2 to 3 days ofculture (data not shown). To further assess the effect of PT andthe S3-S4 dimer on T-cell subsets, the proportions of CD41
and CD81 T cells in naive LN cells were determined by FACSanalysis at various intervals during incubation with PTmu, theS3-S4 dimer, the anti-CD3 MAb, and ConA. As can be seenin Fig. 7, an increase in the proportion of CD81 T cells, inparrallel with a decrease in the proportion of CD41 T cells,was observed upon incubation with PTmu or with the S3-S4dimer; the extent of the reversal of the CD41/CD81 ratiopeaked after approximately 72 h of incubation and leveled outthereafter (Fig. 7). The proportion of CD41 or CD81 T cellsdid not change at any time during incubation with the anti-CD3 MAb or ConA (Fig. 7). The reversal of the CD41/CD81
ratio in naive LN cells incubated with PTmu or the S3-S4dimer does not appear to be due to preferential stimulation ofCD81 T cells by PTmu or the S3-S4 dimer; indeed, the extentof proliferation in response to PTmu, as standardized to theproliferation in response to the anti-CD3 MAb, did not differbetween LN cells isolated from mice fully depleted of CD81 Tcells by treatment with the anti-CD8 MAb and LN cells iso-lated from mice fully depleted of CD41 T cells by treatmentwith the anti-CD4 MAb (Fig. 8). To assess the possibility thatthe increase in the proportion of CD81 T cells upon stimula-tion with PT or the S3-S4 dimer was a result of preferentialapoptosis-necrosis of CD41 T cells, LN cells from naive miceand from mice depleted of CD81 or CD41 T cells were stim-ulated with PTmu, the anti-CD3 MAb, or ConA and analyzedby FACS for annexin staining at various intervals during a5-day culture. FACS analysis of cells double stained with an-nexin and the anti-CD4 or anti-CD8 MAb did not indicate anypreferential apoptosis-necrosis of CD41 T cells over CD81 T
cells in naive LN cells incubated with PTmu (Fig. 9, Unde-pleted), nor was there any difference in the extent of apoptosis-necrosis in the naive LN cells incubated with the anti-CD3MAb or ConA, compared with the PTmu incubation (Fig. 9,Undepleted). These observations were corroborated by dataobtained with LN cells from CD41 T-cell-depleted or CD81
T-cell-depleted mice (Fig. 9); indeed, the extent of apoptosis-necrosis of the T cells was essentially the same in culturesstimulated with PTmu, the anti-CD3 MAb, or ConA, regard-less of the origin of the LN cells (Fig. 9).
DISCUSSION
The data presented in this report provide direct evidencethat the S3-S4 dimer of PT is mitogenic and suggest, moreover,that the S3-S4 dimer would be sufficient to account for themitogenic activity of the B oligomer. To the best of our knowl-
FIG. 6. The S3-S4 dimer of PT is mitogenic for naive and committed T cells. Cells were cultured as described in Materials and Methods withPT, the B oligomer, peak I, peak II, rS2, rS3, or rS4 (250 ng/well); ConA (100 ng/well); or pMOG35-55 (1 mg/well). The stimulation index (inparentheses) was calculated as the mean stimulation in the presence of the relevant protein preparation divided by the mean stimulation in theabsence of the added protein (None). n.t., not tested; n.a., not applicable; SD, standard deviation.
TABLE 1. In vitro stimulation of naive LN cells with PTmu or theS3-S4 dimer results in reversal of the CD41/CD81 T-cell ratioa
Marker
% Positive LN cellsb
NaiveStimulated in vitro with:
Anti-CD3 MAb ConA PTmu S3-S4 dimer
CD41 54 6 9 49.5 6 5.5 48 6 4.5 19 6 3 25 6 4CD81 27 6 5 20.5 6 0.5 25 6 3 58 6 12 53 6 6CD41 CD81 0.2 6 0.1 3 6 2 3.5 6 0.5 3.5 6 1.5 2 6 0.3
a Naive LN cells were incubated for 96 h in the presence of the anti-CD3 MAb,ConA, PTmu, or the S3-S4 dimer and FACS analyzed for CD4 and CD8 markersas described in Materials and Methods. Data are presented as the mean 6 thestandard deviation of three experiments.
b The percentage of marker-positive cells among TCRab1 cells is shown.
3078 LATIF ET AL. INFECT. IMMUN.
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
edge, this study is also the first to demonstrate that in vitrostimulation with PT or its S3-S4 dimer component, of naivecells derived from a mature lymphoid organ, results in a re-versal of the CD41/CD81 T-cell ratio.
The best-described property of the B oligomer is its mito-genic activity; the catalytic subunit S1 does not play a role inthe mitogenicity of PT, as PTmu was as strongly mitogenic asthe B oligomer (13; this study). Some of the biological effectsand biochemical properties of the B oligomer have been shownto be more strongly associated with the S2-S4 or S3-S4 het-erodimer of PT, rather than with the S2 or S3 subunit alone,suggesting that conformation of the subunit as a dimer with S4is important for activity. Along these lines, we observed a verystrong mitogenic effect of the isolated S3-S4 dimer (peak II),on both naive and committed antigen-specific T cells, whereasthe recombinant subunits had no such effect. These results arein contrast to previous reports indicating that the S2-S4 andS3-S4 dimers are not mitogenic by themselves (31, 36). Theonly obvious difference between our study and those previouslyreported (31, 36) is the absence of urea in our S3-S4 dimerpreparation. The possibility that the mitogenic activity of peakII is, in fact, due to contamination with small amounts of the
FIG. 7. Time course analysis of reversal of the CD41/CD81 T-cell ratio upon in vitro stimulation of naive LN cells with PT and the S3-S4 dimer(peak II). Naive LN cells were cultured as described in Materials and Methods in the presence of the indicated mitogens. At the time intervalsindicated, 0.2 3 106 cells were tested for CD4 and CD8 expression by double-staining FACS analysis. The data shown are means 6 standarddeviations of three separate experiments. An asterisk indicates CD41 or CD81 cells among TCRab1 cells.
FIG. 8. Reversal of the CD41/CD81 ratio in LN cells stimulatedwith PTmu is not due to preferential stimulation of CD81 T cells. Theproliferative response to the anti-CD3 Ab (5 ml/ml) or PTmu (250ng/ml) by LN cells isolated from normal mice, mice depleted of CD41
T cells, or mice depleted of CD81 T cells was assayed as described inMaterials and Methods and is expressed as a mean stimulation index(SI) 6 the standard deviation (SD). Standardization of PTmu stimu-lation to anti-CD3 Ab stimulation equals 0.85, 0.77, and 1.02 fornormal, CD4-depleted, and CD8-depleted mice, respectively.
VOL. 69, 2001 CD41/CD81 T-CELL RATIO REVERSAL BY S3-S4 DIMER OF PT 3079
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

S2-S4 dimer, and therefore to divalent binding on T cells by theS2-S4 and S3-S4 dimers, is highly unlikely, as S2 could not bedetected by very sensitive Western blotting of relatively largeamounts of peak II, where S3 and S4 appear as very stronglyreacting, broad bands. It is more likely that an appropriateconformation of the S3-S4 dimer is necessary for mitogenicactivity. Hence, the previous failure to demonstrate the mito-genic activity of the dimer itself may be due simply to the factthat all previous attempts were performed with dimers thatwere isolated and stored in urea and therefore did not presentan appropriate conformation for the biological activity. Isola-tion of the S3-S4 dimer in the absence of urea in our studywould have allowed adequate refolding of the dimer. It ishighly relevant to point out the circumstantial evidence sug-gesting that the S3-S4 dimer is most likely implicated in themitogenicity of the B oligomer, whereas the S2-S4 dimer mayonly play a more minor role in this function. Studies have usedmethylated PT to analyze the involvement of the various PTcomponents in the biological activity of PT (20, 21). Chemicalmodification of PT by methylation of lysine residues does notsignificantly interfere with the A protomer-transporting activ-ity of the B oligomer but results in loss of the mitogenic activityof PT (21). Subsequent studies with hybrid, differentially meth-ylated PT holomers indicated that the two dimers play differ-ential roles in the mitogenic activity of the B oligomer; thus,hybrid PT formed with a methylated S3-S4 dimer lost its abilityto stimulate T lymphocytes, whereas hybrid PT formed with amethylated S2-S4 dimer was as potent a mitogen as unmodifiedPT (22). Unfortunately, the direct role of the S2-S4 dimercould not be evaluated in our study, as we were not able topurify the S2-S4 dimer under the chromatography conditionsused, possibly as a result of its strong binding to Sephadex.
How stimulation by PT or the S3-S4 dimer results in reversalof the CD41/CD81 T-cell ratio is unclear. From our data, it
does not appear to be a result of preferential CD41 T-celldeath or preferential stimulation of CD81 T cells. Recentstudies suggest the possibility that PTmu or the S3-S4 dimercan selectively induce the generation of CD81 T cells fromCD41 CD81 T-cell precursors. Thus, using ZAP-702 mutantthymus organ cultures in which T-cell development is arrestedat the CD41 CD81 thymocyte stage, Takahama et al. (30)showed that PT or, more specifically, the B oligomer selectivelyinduced the generation of CD42 CD81 TCRhigh cells, possiblyby up-regulating Notch expression (30). Notch has been impli-cated as a participant in the CD4 versus CD8 lineage decision,whereby expression of an activated form of Notch in develop-ing murine T cells was demonstrated to lead to both an in-crease in CD8 lineage T cells and a decrease in CD4 lineage Tcells (25). It is interesting that an increase in the proportion ofCD81 T cells was also observed in ex vivo LN cells obtainedfrom mice injected with PT (our unpublished results) andsimilar observations of sustained increases in CD81 T-celllevels were noted in rhesus macaques as long as 53 days afterPT injection (23). The possibility that in vitro PT or the S3-S4dimer may act on CD41 CD81 T cells to preferentially inducedifferentiation into CD81 T cells is being further investigated.
Hence, we have demonstrated that the mitogenic activity ofPT does not necessarily require the whole B oligomer or diva-lent binding by the S2-S4 and S3-S4 heterodimers, as shownpreviously, but that the S3-S4 heterodimer alone can be fullyfunctional in stimulating T cells. In addition, in contrast toother T-cell mitogens, PT or the S3-S4 dimer may exert animmunomodulating effect on the T-cell population by stimu-lating an increase in the overall proportion of CD81 T cells.We are considering this aspect further in the context of ourinvestigation of the protective effect of PT on autoimmunediseases such as EAE (3, 4).
FIG. 9. Time course analysis of apoptosis-necrosis in CD41 and CD81-expressing cells upon in vitro stimulation with different mitogens of LNcells from naive (Undepleted), CD41 T-cell-depleted, and CD81 T-cell-depleted mice. LN cells from naive (Undepleted), CD41 T-cell-depleted,and CD81 T-cell-depleted mice were cultured as described in Materials and Methods in the presence of PTmu, the anti-CD3 MAb and ConA.At the time intervals indicated, 0.2 3 106 cells were tested by double-staining FACS analysis for the proportion of CD41 or CD81 apoptotic-necrotic T cells in the cultures. The mean data of three experiments are shown for each time point; the standard deviation (not shown) variedbetween 5 and 15% of the mean. An asterisk indicates the percentage of CD41 or CD81 T cells which were also stained with annexin.
3080 LATIF ET AL. INFECT. IMMUN.
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

ACKNOWLEDGMENTS
This work was partially supported by the Minerva Foundation, Mu-nich, Germany. A. Ben-Nun is the incumbent of the Eugene andMarcia Appelbaum Professorial Chair.
REFERENCES
1. Bartley, T. D., D. W. Whiteley, V. L., D. L. Mar Burns, and W. N. Burnette.1989. Pertussis holotoxoid formed in vitro with a genetically deactivated S1subunit. Proc. Natl. Acad. Sci. USA 86:8353–8357.
2. Ben-Nun, A., and Z. Lando. 1983. Detection of cells responding to myelinbasic protein by proliferation and selection of T cell lines functional inmediating experimental autoimmune encephalomyelitis in mice. J. Immunol.130:1205–1209.
3. Ben-Nun, A., I. Mendel, and N. Kerlero de Rosbo. 1997. Immunomodulationof murine experimental autoimmune encephalomyelitis by pertussis toxin:the protective activity, but not the disease-enhancing activity, can be attrib-uted to the nontoxic B-oligomer. Proc. Assoc. Am. Phys. 109:120–125.
4. Ben-Nun, A., S. Yossefi, and D. Lehman. 1993. Protection against autoim-mune disease by bacterial agents. II. PPD and pertussis toxin as proteinsactive in protecting mice against experimental autoimmune encephalomyeli-tis. Eur. J. Immunol. 23:689–696.
5. Black, W. J., J. J. Munoz, M. J. Peacock, P. A. Schad, J. L. Cowell, J. J.Burchall, M. Lim, A. Kent, L. Steinman, and S. Falkow. 1988. ADP-ribo-syltransferase activity of pertussis toxin and immunomodulation by Borde-tella pertussis. Science 240:656–659.
6. Cobbold, S. P., A. Jayasuriya, A. Nash, T. D. Prospero, and H. Waldmann.1984. Therapy with monoclonal antibodies by elimination of T-cell subsets invivo. Nature (London) 312:548–551.
7. Dialynas, D. P., Z. S. Quan, K. A. Wall, A. Pierres, J. Quintans, M. R. Loker,M. Pierres, and F. W. Fitch. 1983. Characterization of the murine T cellsurface molecule, designated L3T4, identified by monoclonal antibodyGK1.5: similarity of L3T4 to the human Leu-3/T4 molecule. J. Immunol.131:2445–2451.
8. Gierschik, P. 1992. ADP-ribosylation of signal-transducing guanine nucle-otide-binding proteins by pertussis toxin. Curr. Top. Microbiol. Immunol.175:69–96.
9. Gray, L. S., K. S. Huber, M. C. Gray, E. L. Hewlett, and V. H. Engelhard.1989. Pertussis toxin effects on T lymphocytes are mediated through CD3and not by pertussis toxin catalyzed modification of a G protein. J. Immunol.142:1631–1638.
10. Hausman, S. Z., and D. L. Burns. 1993. Binding of pertussis toxin to lipidvesicles containing glycolipids. Infect. Immun. 61:335–337.
11. Heerze, L. D., P. C. S. Chong, and G. D. Armstrong. 1992. Investigation ofthe lectin-like binding domains in pertussis toxin using synthetic peptidesequences. J. Biol. Chem. 267:25810–25815.
12. Kerlero de Rosbo, N., I. Mendel, and A. Ben-Nun. 1995. Chronic relapsingexperimental autoimmune encephalomyelitis with a delayed onset and anatypical clinical course, induced in PL/J mice by myelin oligodendrocyteglycoprotein (MOG)-derived peptide: preliminary analysis of MOG T cellepitopes. Eur. J. Immunol. 25:985–993.
13. Kimura, A., K. T. Mountzouros, P. A. Schad, W. Cieplak, and J. L. Cowell.1990. Pertussis toxin analog with reduced enzymatic and biological activitiesis a protective immunogen. Infect. Immun. 58:3337–3347.
14. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature (London) 227:680–685.
15. Lobet, Y., C. Feron, G. Dequesne, E. Simoen, P. Hauser, and C. Locht. 1993.Site-specific alterations in the B-oligomer that affect receptor-binding activ-ities and mitogenicity of pertussis toxin. J. Exp. Med. 177:79–87.
16. Loosmore, S., G. Zealey, S. Cockle, H. Boux, P. Chong, R. Yacoob, and M.Klein. 1993. Characterization of pertussis toxin analogs containing mutationsin B-oligomer subunits. Infect. Immun. 61:2316–2324.
17. Mendel, I., N. Kerlero de Rosbo, and A. Ben-Nun. 1995. A myelin-oligoden-drocyte glycoprotein (MOG) peptide induces typical chronic experimentalautoimmune encephalomyelitis in H-2b mice. Fine specificity and T cellreceptor Vb expression of encephalitogenic T cells. Eur. J. Immunol. 25:1951–1959.
18. Munoz, J. J. 1985. Biological activities of pertussigen (pertussis toxin), p.1–18. In R. D. Sekura, J. Moss, and M. Vaughan (ed.), Pertussis toxin.Academic Press, Inc. New York, N.Y.
19. Nicosia, A., A. Bartoloni, M. Perugini, and R. Rappuoli. 1987. Expressionand immunological properties of the five subunits of pertussis toxin. Infect.Immun. 55:963–967.
20. Nogimori, K., K. Ito, M. Tamura, S. Satoh, S. Ishii, and M. Ui. 1984.Chemical modification of islet-activating protein, pertussis toxin. Essentialrole of free amino groups in its lymphocytosis-promoting activity. Biochim.Biophys. Acta 801:220–231.
21. Nogimori, K., M. Tamura, M. Yajima, K. Ito, T. Nakamura, N. Kajikawa, Y.Maruyama, and M. Ui. 1984. Dual mechanisms involved in development ofdiverse biological activities of islet-activating protein, pertussis toxin, asrevealed by chemical modifications of lysine residues in the toxin molecule.Biochim. Biophys. Acta 801:232–243.
22. Nogimori, K., M. Tamura, M. Yajima, N. Hashimura, S. Ishii, and M. Ui.1986. Structure-function relationship of islet-activating protein, pertussistoxin: biological activities of hybrid toxins reconstituted from native andmethylated subunits. Biochemistry 25:1355–1363.
23. Pauza, C. D., P. W. Hinds, C. Yin, T. S. McKechnie, S. B. Hinds, and M. S.Salvato. 1997. The lymphocytosis-promoting agent pertussis toxin affectsvirus burden and lymphocyte distribution in the SIV-infected rhesus ma-caque. AIDS Res. Hum. Retrovir. 13:87–95.
24. Pizza, M., A. Covacci, A. Bartoloni, M. Perugini, L. Nencioni, M. T. DeMagistratis, L. Villa, D. Nucci, R. Manetti, M. Bugnoli, F. Giovanni, R.Olivieri, J. T. Barbieri, H. Sato, and R. Rappuoli. 1989. Mutants of pertussistoxin suitable for vaccine development. Science 246:497–500.
25. Robey, E., D. Chang, A. Itano, D. Cado, H. Alexander, D. Lans, G. Wein-master, and P. Salmon. 1996. An activated form of Notch influences thechoice between CD4 and CD8 T cell lineages. Cell 87:483–492.
26. Saukkonen, K., W. N. Burnette, V. L. Mar, H. R. Masure, and E. I. Tuo-manen. 1992. Pertussis toxin has eukaryotic-like carbohydrate recognitiondomains. Proc. Natl. Acad. Sci. USA 89:118–122.
27. Schagger, H., and G. von Jagow. 1987. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the rangefrom 1 to 100 kDa. Anal. Biochem. 166:368–379.
28. Stein, P. E., A. Boodhoo, G. D. Armstrong, S. A. Cockle, M. H. Klein, andR. J. Read. 1994. The crystal structure of pertussis toxin. Structure 2:45–57.
29. Strnad, C., and R. A. Carchman. 1987. Human T lymphocyte mitogenesis inresponse to the B oligomer of pertussis toxin is associated with an earlyelevation in cytosolic calcium concentrations. FEBS Lett. 223:16–20.
30. Takahama, Y., Y. Tokoro, T. Sugawara, I. Negishi, and H. Nakauchi. 1997.Pertussis toxin can replace T cell receptor signals that induce positive selec-tion of CD8 T cells. Eur. J. Immunol. 27:3318–3331.
31. Tamura, M., K. Nogimori, M. Yajima, K. Ase, and M. Ui. 1983. A role of theB-oligomer moiety of islet-activating protein, pertussis toxin, in developmentof the biological effects on intact cells. J. Biol. Chem. 258:6756–6761.
32. Tamura, M., K. Nogimori, S. Murai, M. Yajima, K. Ito, T. Katada, M. Ui,and S. Ishii. 1982. Subunit structure of islet-activating protein, pertussistoxin, in conformity with the A-B model. Biochemistry 21:5516–5522.
33. Towbin, H., T. Staehelin, and J. Gordon. 1979. Electrophoretic transfer ofproteins from polyacrylamide gels to nitrocellulose sheets: procedure andsome applications. Proc. Natl. Acad. Sci. USA 76:4350–4354.
34. van’t Wout, J., W. N. Burnette, V. L. Mar, E. Rozdzinski, S. D. Wright, andE. I. Tuomanen. 1992. Role of carbohydrate recognition domains of pertussistoxin in adherence of Bordetella pertussis to human macrophages. Infect.Immun. 60:3303–3308.
35. Weiss, A. A., and E. L. Hewlett. 1986. Virulence factors of Bordetella pertus-sis. Annu. Rev. Microbiol. 40:661–686.
36. Witvliet, M. H., M. L. Vogel, E. J. H. J. Wiertz, and J. T. Poolman. 1992.Interaction of pertussis toxin with human T lymphocytes. Infect. Immun.60:5085–5090.
37. Witvliet, M. H., D. L. Burns, M. J. Brennan, J. T. Poolman, and C. R.Manclark. 1989. Binding of pertussis toxin to eucaryotic cells and glycopro-teins. Infect. Immun. 57:3324–3330.
Editor: J. D. Clements
VOL. 69, 2001 CD41/CD81 T-CELL RATIO REVERSAL BY S3-S4 DIMER OF PT 3081
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from