research vascular endothelial growth factor...
TRANSCRIPT

3548
Published OnlineFirst July 6, 2010; DOI: 10.1158/1078-0432.CCR-09-2797
Human Cancer Biology Clinical
CancerResearch
Vascular Endothelial Growth Factor Receptors VEGFR-2 andVEGFR-3 Are Localized Primarily to the Vasculature inHuman Primary Solid Cancers
Neil R. Smith1, Dawn Baker1, Neil H. James1, Kirsty Ratcliffe1, Martin Jenkins2, Susan E. Ashton1, Graham Sproat1,Ruth Swann1, Neil Gray1, Anderson Ryan1, Juliane M. Jürgensmeier1, and Chris Womack2
Abstract
Authors' ADevelopme
Note: SuppResearch O
CorresponMereside,Kingdom. PN.R.Smith@
doi: 10.115
©2010 Am
Clin Canc
Downl
Purpose: Vascular endothelial growth factor (VEGF) signaling is key to tumor angiogenesis and is animportant target in the development of anticancer drugs. However, VEGF receptor (VEGFR) expression inhuman cancers, particularly the relative expression of VEGFR-2 and VEGFR-3 in tumor vasculature versustumor cells, is poorly defined.Experimental Design: VEGFR-2– and VEGFR-3–specific antibodies were identified and used in the
immunohistochemical analysis of human primary cancers and normal tissue. The relative vascular local-ization of both receptors in colorectal and breast cancers was determined by coimmunofluorescence withvascular markers.Results: VEGFR-2 and VEGFR-3 were expressed on vascular endothelium but not on malignant cells in
13 common human solid tumor types (n > 400, bladder, breast, colorectal, head and neck, liver, lung,skin, ovarian, pancreatic, prostate, renal, stomach, and thyroid). The signal intensity of both receptors wassignificantly greater in vessels associated with malignant colorectal, lung, and breast than adjacent non-tumor tissue. In colorectal cancers, VEGFR-2 was expressed on both intratumoral blood and lymphaticvessels, whereas VEGFR-3 was found predominantly on lymphatic vessels. In breast cancers, both recep-tors were localized to and upregulated on blood vessels.Conclusions: VEGFR-2 and VEGFR-3 are primarily localized to, and significantly upregulated on, tu-
mor vasculature (blood and/or lymphatic) supporting the majority of solid cancers. The primary clinicalmechanism of action of VEGF signaling inhibitors is likely to be through the targeting of tumor vesselsrather than tumor cells. The upregulation of VEGFR-3 on tumor blood vessels indicates a potential ad-ditional antiangiogenic effect for dual VEGFR-2/VEGFR-3–targeted therapy. Clin Cancer Res; 16(14); 3548–61.
©2010 AACR.
Vascular endothelial growth factor (VEGF) signaling iskey to physiologic and pathologic angiogenesis and lym-phangiogenesis, and is an established target in the develop-ment of anticancer therapeutics. VEGF-A is an importantdriver of the neovascular growth required to support solidtumor progression (1). The primary signaling receptor forVEGF-A is VEGF receptor (VEGFR)-2 (Flk-1, KDR; refs. 2, 3),and activation of VEGFR-2 by VEGF-A on endothelial tipand stalk cells directs the migration and extension ofsprouting vessels, respectively (4). VEGFR-3 (Flt-4) is the
ffiliations: 1Cancer Bioscience and 2Oncology Clinicalnt, AstraZeneca, Cheshire, United Kingdom
lementary data for this article are available at Clinical Cancernline (http://clincancerres.aacrjournals.org/).
ding Author: Neil R. Smith, Cancer Bioscience, AstraZeneca,Alderley Park, Macclesfield, Cheshire SK10 4TG, Unitedhone: 44-1625-233731; Fax: 44-1625-510097; E-mail: Neil.astrazeneca.com.
8/1078-0432.CCR-09-2797
erican Association for Cancer Research.
er Res; 16(14) July 15, 2010
Researcon Septembclincancerres.aacrjournals.org oaded from
receptor for VEGF-C and VEGF-D (5). Expression ofVEGFR-3 is largely confined to lymphatic endotheliumin adult tissues (6), and its activation induces lymphan-giogenesis (7). VEGFR-3 was recently also found expressedon the endothelial tip cells of angiogenic tumor vascula-ture, and blockade of the receptor in tumor models in-duces blood vessel endothelial cell sprouting and vesselbranching (8).Agents that inhibit VEGF signaling and are already reg-
istered for the treatment of cancer include the VEGF-A neu-tralizing antibody bevacizumab (9) and small-moleculemultikinase inhibitors sorafenib, sunitinib, and pazopa-nib (10, 11). Other small-molecule VEGFR inhibitors un-der evaluation in clinical trials include vandetanib andcediranib. Cediranib is a potent inhibitor of VEGF signal-ing, targeting VEGFR-1, VEGFR-2, and VEGFR-3 and an-giogenesis (12). Recent publications have shown thatVEGFR-3 inhibition by cediranib reduces lymphangiogen-esis as well as the incidence of lymphatic metastases in vivo(13, 14). The combined blockade of VEGFR-2 and VEGFR-3 has an additive antiangiogenic and antitumor effect
h. er 16, 2018. © 2010 American Association for Cancer

Translational Relevance
Vascular endothelial growth factor (VEGF) signalinginhibitors have shown clinical efficacy in a range ofsolid tumor types, including colorectal, lung, andbreast cancer. A better understanding of the expressionand localization of VEGF receptors is needed to under-stand the potential for direct antitumor effects fromVEGF signaling inhibitors in the clinic and to helpidentify patients who might gain greatest benefit fromtreatment. Our findings indicate that VEGFR-2 andVEGFR-3 are primarily localized to tumor vessels in abroad range of solid tumors, suggesting that tumor en-dothelial cells and not tumor cells are the likely prima-ry target for anti–VEGFR-2 and anti–VEGFR-3 therapyin cancer. Furthermore, the expression of VEGFR-3, aswell as VEGFR-2, on tumor blood vessels suggests thatdual VEGFR-2/VEGFR-3 inhibitors may have additionalantiangiogenic benefits in some patients.
VEGFR-2 and VEGFR-3 Status of Human Solid Tumors
Published OnlineFirst July 6, 2010; DOI: 10.1158/1078-0432.CCR-09-2797
in vivo over inhibition of VEGFR-2 alone (8). In additionto reducing lymphangiogenic metastases, dual VEGFR-2and VEGFR-3 inhibitors may improve the antiangiogenicresponse in patients.An understanding of the VEGFR-2 and VEGFR-3 status of
human tumors may help to identify patients who wouldbenefit most from treatment with dual VEGFR-2 andVEGFR-3 inhibitors and clarify the clinical mechanism ofaction of these agents. Presently, there is no clear agreementfrom the literature on the location (vasculature and/orcancer cells) or extent of expression of either VEGFR-2 orVEGFR-3 in human solid tumors. For example, using im-munohistochemistry, VEGFR-2 has been detected on tu-mor vessels supporting human colorectal (CRC; ref. 15),breast (BC; refs. 16, 17), and non–small cell lung carcino-mas (NSCLC; refs. 18, 19). However, tumor cell expressionhas also been reported in 40% to 100% of CRC (20, 21),BC (22–24), and NSCLC (18, 19, 25). Similarly, somereports confine VEGFR-3 to tumor vessels, primarilylymphatic but also blood endothelium in CRC (26), BC(27–30), and NSCLC (19, 31–33), whereas others localizethe receptor to tumor cells [CRC (34), BC (23, 35), andNSCLC (19, 25, 32, 33, 36, 37)]. Recently, Petrova et al.(38) have provided compelling evidence that poor anti-body specificity is likely to be responsible for some of theclaims that VEGFR-3 is found on human tumor cells (38).Using a validated antibody, they confirm that VEGFR-3 isconfined primarily to blood and lymphatic vessels in solidtumors (38).This report aimed to resolve the issues around the rela-
tive extent and location of VEGFR-2 and VEGFR-3 expres-sion in human primary solid tumors and, in doing so,better define the clinical mechanism of action of VEGF-targeted therapy. To do this, we determined the cross-reactivity of human VEGFR antibodies using multiple
www.aacrjournals.org
Researcon Septembclincancerres.aacrjournals.org Downloaded from
screening platforms and identified two antibodies,55B11 (VEGFR-2; Cell Signaling Technology) and AF349(VEGFR-3; R&D Systems), with credible specificity. Thesewere used in immunohistochemical and coimmunofluor-escent analyses of archival human solid tumor samples tounderstand the expression of the two receptors between(a) tumor endothelium and malignant cells and (b) tumorand normal endothelium and investigate the relative local-ization of VEGFR-2 and VEGFR-3 on tumor vasculature.
Materials and Methods
AntibodiesThe following antibodies were used: custom rabbit
polyclonal antibody raised to COOH-terminal peptide ofmouse CD31 (CHG-CD31-P1; AstraZeneca); mousemonoclonal antibodies to human CD31 (JC70A; Dako),α-smooth muscle actin (α-SMA; 1A4; Sigma), phosphotyr-osine (P-Tyr-100; Cell Signaling Technology), and rabbitglyceraldehyde-3-phosphate dehydrogenase (AM4300;Ambion); rabbit monoclonal against human VEGFR-2(55B11); goat polyclonal against human VEGFR-3(AF349); and rabbit polyclonal against human LYVE-1(102-PA50AG; Reliatech). Other VEGFR antibodies arelisted in Supplementary Table S1.
Cell linesCell lines were maintained at 37°C with 5% CO2:
HT-29 (obtained from the European Collection of Ani-mal Cell Cultures) in DMEM with 5% fetal bovine se-rum (FBS) and 1% nonessential amino acids; Colo-205(European Collection of Animal Cell Cultures) in RPMI1640/10% FBS; A549 (American Type Culture Collec-tion) in DMEM/10% FBS/10% M1; PC-9 (a kind giftfrom Dr. K Nishio, National Cancer Centre of Japan)in RPMI 1640/10% FBS/10% M1; MCF-7 (ImperialCancer Research Fund) in DMEM/10% FCS; aorticVSMC (C-12533; Promocell) in Promocell VSMC growthmedia (C-22062); M-07e (Deutsche Sammlung von Mik-roorganismen und Zellkulturen) in RPMI 1640/10%FBS/10% M1/interleukin-3 (10 ng/mL)/granulocyte mac-rophage colony-stimulating factor (10 ng/mL); PAE(Ludwig Institute for Cancer Research) in Ham's F12/10% FBS/L-glutamine; and NIH-3T3 (Jefferson CancerInstitute) in DMEM/10% FBS/L-glutamine. Full-lengthhuman cDNA for VEGFR-1, VEGFR-2, and CSF1R wasPCR amplified from nm_002019.3, nm_002253.2, andnm_005211.3, respectively, using the following primerpairs: VEGFR-1, 5′-GTTTAACTTTAAGAAGGAGATATAAC-CATGGTCAGCTACTGGGACACCGGGG-3′ (forward)and 5′-CTATAGGTCCTCCTCTGATATTAGCTTTTGCTC-GATGGGTGGGGTGGAGTACAGGACC-3′ (reverse);VEGFR-2, 5′-ATTTAACTTTAAGAAGGAGATATAACCCA-GAGCAAGGTGCTGCTGGCCGTCGCCC-3′ (forward)and 5′-TCTATAGGTCCTCCTCTGATATTAGCTTTTGCT-CAACAGGAGGAGAGCTCAGTGTGG-3′ (reverse); andC S F 1 R , 5 ′ - T C A T A G G T C C T C C T C T G A T A T -TAGCTTTTGCTCGCAGAACTGATAGTTGTTGGGCTGC-3′
Clin Cancer Res; 16(14) July 15, 2010 3549
h. er 16, 2018. © 2010 American Association for Cancer

Smith et al.
3550
Published OnlineFirst July 6, 2010; DOI: 10.1158/1078-0432.CCR-09-2797
(forward) and 5′-ATTTAACTTTAAGAAGGAGATATAAC-CATGGGCCCAGGAGTTCTGCTGCTCCTGC-3′ (reverse).VEGFR-3 (cDNA cloned into pDONR221), accession
number nm_182925, was obtained from GeneArt. DNAfragments were subcloned into the retroviral vector pBINCIP SIN via Gateway technology (Invitrogen) and cotrans-fected with constructs expressing the viral envelope pro-tein VSVG (Clontech) into a packaging cell line,Phoenix-Hek 293 (Garry P. Nolan, Stanford UniversitySchool of Medicine). Retroviral particles were used to in-fect NIH-3T3 and PAE cells, and stable lines were selectedfor resistance to puromycin.
Tumor xenograftsTumor xenograft tissue was derived from experiments
conducted with licenses issued under the UK Animals (Sci-entific Procedures) Act 1986 and after local ethical reviewand approval. Cell lines were maintained in the recom-mended growth medium and implanted s.c. into the leftflank of immunocompromised mice: nude, scid, or scid-bg (see Supplementary Table S2 for details). Tumors weregrown to ∼1 cm3 volume and then collected and fixed informalin for 24 hours before being embedded in paraffin.
Patient tissue samplesFormalin-fixed, paraffin-embedded (FFPE) human pri-
mary cancer resection samples and tissue microarrays(TMA) from patients with untreated human primary blad-der, breast, colorectal, head and neck, liver, lung, skin,ovarian, pancreatic, prostate, renal, stomach, and thyroidcancers and matched adjacent normal tissue (for colorec-tal, lung, and breast cancers) were sourced under approvedlegal contract from three commercial tissue suppliers (As-terand, Cytomyx, and TriStar Technology Group) and ahospital tissue bank (Wales Cancer Bank). Appropriateconsents, licensing, and ethical approval were obtainedfor this research. The suitability of each specimen for im-munohistochemical analyses was determined by patholo-gy assessment of tissue morphology and preservation(H&E) and the general extent of antigen preservation(CD31 and p-Tyr immunostains).
Western blot analysisSubconfluent (70-80%) cell lines were scraped into ice-
cold lysis buffer [20 mmol/L Tris (pH 7.5), 137 mmol/LNaCl, 10% glycerol, 1% NP40, 0.1% SDS, 50 mmol/LNaF, 1 mmol/L Na3VO4, 1 protease inhibitor tablet/25-mLbuffer (Boehringer Ingelheim)], incubated on ice for 30minutes, aspirated, and cleared by centrifugation. Frozentumor xenograft tissues were homogenized in ice-cold ly-sis buffer using an Ultraturrax T25 homogenizer (Jankeand Kunkel) before incubation on ice. Lysate samples(250 μg) were separated by SDS-PAGE under reducingconditions and transferred to nitrocellulose membrane(Invitrogen) by Western blotting. Membranes wereblocked in TBS containing 0.05% Tween (TBST) and5% nonfat dry milk (Marvel) for 1 hour at room temper-ature and then incubated for 16 hours at 4°C in VEGFR-2
Clin Cancer Res; 16(14) July 15, 2010
Researcon Septembclincancerres.aacrjournals.org Downloaded from
antibodies (55B11, A3, sc-504, and sc-315; diluted1:1,000 in block), VEGFR-3 antibody (AF349; 1:1,000),CD31 antibody (CHG-CD31-P1; 1:500), or glyceralde-hyde-3-phosphate dehydrogenase antibody (AM4300;1:10,000). Immunoblots were incubated in a 1:2,000 di-lution of horseradish peroxidase–conjugated anti-rabbit(New England Biolabs), anti-mouse (New England Bio-labs), or anti-goat (Dako) antibody and visualized usingthe SuperSignal West Pico Substrate method of detection(Perbio Science).
Immunohistochemistry and immunocytochemistryAll incubations were at room temperature and washes
were done with TBST. FFPE tissues sectioned at 4 μm ontoslides were dewaxed and rehydrated. Antigen retrieval wasdone in a RHS-1 microwave vacuum processor (Mile-stone) at 110°C for 5 minutes; in pH 6 retrieval buffer(S1699; Dako) for p-Tyr, CD31, and VEGFR-3; or in pH9 retrieval buffer (S2367; Dako) for VEGFR-2. Endogenousbiotin was blocked using an avidin-biotin kit (SP-2002;Vector Laboratories, Inc.), endogenous peroxidase activitywas blocked with 3% hydrogen peroxide for 10 minutes,and nonspecific binding sites were blocked with serum-free protein block (X0909; Dako) for 20 minutes. Primaryantibodies to p-Tyr (P-Tyr-100), CD31 (JC70A and CHG-CD31-P1), VEGFR-2 (55B11), and VEGFR-3 (AF349) werediluted 1:500, 1:500, 1:600, and 1:200, respectively, in an-tibody diluent (S0809; Dako) and incubated with sectionsfor 1 hour. Either mouse Envision secondary (K4007;Dako) for p-Tyr and CD31, rabbit Envision secondary(K4003; Dako) for VEGFR-2 and CD31 (CHG-CD31-P1), or biotinylated rabbit anti-goat immunoglobulinantibodies (E0466, diluted 1:400 in TBST; Dako), fol-lowed by Vectastain Elite ABC solution (PK-6100; VectorLaboratories), for VEGFR-3 were added for 30 minuteseach. Sections were developed in diaminobenzidine for10 minutes (K3466; Dako) and counterstained with Car-azzi's hematoxylin. Appropriate no primary antibody andisotype controls were done for each antibody.For immunocytochemical analyses, subconfluent cells
were transferred to 10% neutral buffered formalin for 24hours at 4°C. Cells were centrifuged to a pellet, washed in80% ethanol, processed into 1% agarose, and embeddedinto paraffin blocks. Cell pellets were sectioned at 4 μm,and 55B11 and AF349 antibodies were evaluated usingthe immunohistochemical conditions without the antigenretrieval.
ImmunofluorescenceImmunofluorescence was done on dewaxed and rehy-
drated FFPE sections. Antigen retrieval was carried out asabove in pH 9 retrieval buffer. After blocking in 20% horseserum, sections were incubated for 1 hour with antibodypairs diluted in serum: 55B11 (1:20, VEGFR-2) and AF349(1:20, VEGFR-3); 55B11 and JC70A (1:20, CD31); AF349(1:20) and JC70A (1:20); 55B11 (1:20) and 1A4 (1:1,000,α-SMA); AF349 (1:20) and 1A4 (1:1,000); and JC70A(1:20) and 102-PA50AG (1:60). Appropriate secondary
Clinical Cancer Research
h. er 16, 2018. © 2010 American Association for Cancer

VEGFR-2 and VEGFR-3 Status of Human Solid Tumors
Published OnlineFirst July 6, 2010; DOI: 10.1158/1078-0432.CCR-09-2797
antibody pairs combined 1:800 in serum were addedfor 30 minutes: 55B11 was detected with donkey anti-rabbit IgG conjugated to Alexa Fluor 555 or Alexa Fluor488 (A31572 or A21206; Molecular Probes); AF349 withdonkey anti-goat IgG Alexa Fluor 555 (A21432; MolecularProbes); NP056 with donkey anti-mouse IgG AlexaFluor 488 or Alexa Fluor 555 (A21202 or A31570; Molec-ular Probes); 1A4 with donkey anti-mouse IgG AlexaFluor 488; and 102-PA50AG with donkey anti-rabbitIgG Alexa Fluor 488. Sections were counterstained withProLong Gold antifade reagent with 4′,6-diamidino-2-phenylindole (P36931; Molecular Probes), and fluores-cent images were scanned and captured using a MIRAXscan (Carl Zeiss).
Image analysis and pathology reviewThe mean number of VEGFR-2– and CD31-positive ves-
sels per mm2 viable tumor for each xenograft tumor mod-el was quantified as described previously (39). Thepercentage of VEGFR-2–positive vessels per tumor modelwas calculated as follows: number VEGFR-2 vessels permm2/number of CD31 vessels per mm2 × 100. All hu-man tissue sections were scored by a pathologist usinga light microscope. A subjective reporting procedurewas implemented, recording location (tumor cell, normalepithelium and vessel) and intensity [− (none), + (weak),++ (medium), and +++ (high)] of VEGFR-2, VEGFR-3,and CD31 staining.
Statistical analysisFor each tissue sample, the maximum VEGFR-2 or
VEGFR-3 staining intensity was recorded on an ordinalscale (0, +, ++, +++). Separate intensities were recordedfor the epithelium and endothelium in each sample wherepossible. A Wilcoxon signed-rank test was used to comparethe relative intensity of staining between (a) tumor epithe-lium and endothelium in a sample and (b) tumor and nor-mal endothelium for matched pairs of tumor and normaltissue. The Bonferroni-Holm procedure was used to takeaccount of the multiple tests being carried out in several tis-sue types and to control the family-wise error rate at 0.05 foreach of VEGFR-2 and VEGFR-3. Those tests where the nullhypothesis (of no difference between groups) would be re-jected are highlighted in Table 1. The proportion of sampleswith positive VEGFR staining (+, ++, or +++) was also esti-mated in each tissue type, and an exact 95% binomial con-fidence interval was given.
Results
Evaluation of VEGFR antibodiesA total of 12 antibodies, raised to either human VEGFR-2
(n = 8) or VEGFR-3 (n = 4), were selected for evaluationbased on the supplier's recommendation and/or a litera-ture precedence for their use in immunohistochemistry(Supplementary Table S1).Of the eight VEGFR-2 antibodies tested, only 55B11 was
VEGFR-2 specific and qualified for use in immunohisto-
www.aacrjournals.org
Researcon Septembclincancerres.aacrjournals.org Downloaded from
chemistry (Supplementary Table S1). By Western blotanalysis, 55B11 detected a 230-kDa doublet band indica-tive of fully and partially glycosylated VEGFR-2 (40) inNIH-3T3 and PAE cells engineered to overexpress humanVEGFR-2 but detected nothing in the untransfected paren-tal cell lines (Fig. 1A). A band ∼150 kDa was also detectedin VEGFR-2–overexpressing PAEs, which is likely to beunglycosylated VEGFR-2 protein (40). These data wereconfirmed by immunocytochemical analysis of FFPE pa-rental and VEGFR-overexpressing PAE cells (Fig. 1B).55B11 did not cross-react with VEGFR-2–related humankinases [VEGFR-1, VEGFR-3, platelet-derived growth factorreceptor-β (PDGFR-β), c-Kit, or CSF1R] as determined byWestern blot analysis of PAE and NIH-3T3 cells engi-neered to overexpress human VEGFR-1 (150 kDa),VEGFR-3 (75/120/195 kDa), or CSF1R (130/150 kDa)and human aortic VSMCs and M-07e, which express highlevels of endogenous PDGFR-β (180 kDa; ref. 41) and c-Kit (145 kDa; ref. 42), respectively (Fig. 1A). Furthermore,55B11 cleanly detected a 230-kDa VEGFR-2 doublet bandby Western blotting in more complex heterogeneous pro-tein lysates (e.g., human cancer xenograft lysates; Fig. 2C).In our hands, the VEGFR-2 antibodies 55B11, A3,
sc-315, and sc-504 commonly used to investigate the ex-pression of VEGFR-2 in human tumors (18–24), did notqualify as specific. All three detected multiple bands, run-ning at different sizes to that expected for VEGFR-2, in ly-sates prepared from cell lines expressing VEGFR familymembers (Fig. 1A) and tumor xenografts and matched pa-rental tumor cell lines (Fig. 2C). Because cross-reactivity ofthese antibodies with other proteins could potentially con-found any analysis of VEGFR-2 expression by immunohis-tochemistry, they were not used in the analysis of humantumor samples.Of four antibodies evaluated, both AF349 and 9D9F9
were specific for VEGFR-3 and suitable for immunohisto-chemistry (Supplementary Table S1). AF349 was selectedfor further analyses over 9D9F9, as this was raised in goatand was therefore more suitable for use in coimmunofluor-escence experiments withmousemonoclonal antibodies toCD31 and α-SMA. AF349 detected a 195-kDa protein,representing immature (nonproteolytically cleaved)VEGFR-3, in VEGFR-3–overexpressing PAE and NIH-3T3cells, which was absent from the parental cell lines(Fig. 1A). Bands running at approximately 120 and 75kDa were also detected in VEGFR-3–overexpressing PAEs.These represent the denatured mature protein, as fully pro-cessed mature VEGFR-3 consists of 120- and 75-kDa pro-teins linked by a disulfide bridge (43). In addition, AF349did not cross-react with VEGFR-3–related kinases VEGFR-1,VEGFR-2, PDGFR-β, c-Kit, or CSF1R (Fig. 1A). The specific-ity of AF349 for VEGFR-3 was confirmed by immunocyto-chemical analysis of engineered PAE cells (Fig. 1B).
VEGFR-2 expression in human tumor xenograftsgrown in immunocompromised miceTMAs were generated from 29 histologically diverse hu-
man tumor xenografts grown to ∼1 cm3. Serial sections
Clin Cancer Res; 16(14) July 15, 2010 3551
h. er 16, 2018. © 2010 American Association for Cancer

Table 1. Maximum VEGFR-2 and VEGFR-3 histopathology intensity scores for human tumors
Tissue VEGFR-2 VEGFR-3
n T/N Endo T cells/N Epi
Endo positives T Endovs. T cell
(P)
T Endovs. N
Endo (P)
n T/N Endo T c ls/N pi
Endo positives T Endovs. T cell
(P)
T Endovs. N
Endo (P)0 + ++ +++ 0 +/++/+++
Prop+ve
Exact 95% CI 0 + ++ +++ 0 + ++/++
Prop+ve
Exact 95% CI
Bladder cancer* 10 0 0 4 6 10 0 1.000 (0.692-1.000) 0.0020† 9 0 0 0 9 9 0 1.000 (0.664-1.000) 0.0039†
BC* 36 2 2 18 14 36 0 0.944 (0.813-0.993) <0.0001† 31 1 2 23 5 31 0 0.968 (0.833-0.999) <0.0001†
CRC* 41 0 2 15 24 41 0 1.000 (0.914-1.000) <0.0001† 41 1 4 18 18 41 0 0.976 (0.871-0.999) <0.0001†
Head andneck cancer*
11 0 1 4 6 11 0 1.000 (0.715-1.000) 0.0010† 11 2 5 4 0 11 0 0.818 (0.482-0.977) 0.0039†
Liver cancer* 22 5 4 7 6 22 0 0.773 (0.546-0.922) <0.0001† 21 1 5 12 3 21 0 0.952 (0.762-0.999) <0.0001†
NSCLC* 90 1 4 46 39 90 0 0.989 (0.940-1.000) <0.0001† 90 4 9 34 43 90 0 0.956 (0.890-0.988) <0.0001†
Melanoma* 9 0 0 3 6 9 0 1.000 (0.664-1.000) 0.0039† 9 0 0 5 4 9 0 1.000 (0.664-1.000) 0.0039†
Ovarian cancer* 18 3 1 8 6 18 0 0.833 (0.586-0.964) <0.0001† 15 1 6 6 2 15 0 0.933 (0.681-0.998) 0.0001†
Pancreaticcancer*
20 2 5 5 8 20 0 0.900 (0.683-0.988) <0.0001† 16 0 4 6 6 16 0 1.000 (0.794-1.000) <0.0001†
Prostate cancer* 12 0 3 7 2 12 0 1.000 (0.735-1.000) 0.0005† 7 4 3 0 0 7 0 0.429 (0.099-0.816) 0.2500Renal cancer* 19 0 3 3 13 19 0 1.000 (0.824-1.000) <0.0001† 17 1 1 6 9 17 0 0.941 (0.713-0.999) <0.0001†
Stomach cancer* 8 0 1 3 4 8 0 1.000 (0.631-1.000) 0.0078† 8 0 0 1 7 8 0 1.000 (0.631-1.000) 0.0078†
Thyroid cancer* 9 0 0 4 5 9 0 1.000 (0.664-1.000) 0.0039† 9 0 1 1 7 9 0 1.000 (0.664-1.000) 0.0039†
BC‡ 84 1 6 22 55 84 0 0.988 (0.935-1.000) <0.0001† 56 5 9 34 8 56 0 0.911 (0.804-0.970) <0.0001†
Matched normalbreast‡
50 10 17 20 3 50 0 0.800 (0.663-0.900) <0.0001† 50 17 17 16 0 50 0 0.660 (0.512-0.788) <0.0001†
CRC‡ 65 0 2 34 29 65 0 1.000 (0.945-1.000) <0.0001† 40 0 1 13 26 40 0 1.000 (0.912-1.000) <0.0001†
Matched normalcolorectal‡
33 0 4 28 1 33 0 1.000 (0.894-1.000) 0.0059† 33 7 13 9 4 33 0 0.788 (0.611-0.910) <0.0001†
NSCLC‡ 31 0 0 9 22 31 0 1.000 (0.888-1.000) <0.0001† 31 0 3 15 13 31 0 1.000 (0.888-1.000) <0.0001†
Matched normallung‡
31 10 16 5 0 31 0 0.677 (0.486-0.833) <0.0001† 31 7 19 5 0 31 0 0.774 (0.589-0.904) <0.0001†
NOTE: VEGFR-2 and VEGFR-3 expression levels were scored in tumor endothelium (T Endo), normal endothelium (N Endo), mor cells (T cells), and normal epithelium (N Epi),where + = weak, ++ = medium, +++ = high. The highest intensity score was used when a sample exhibited a range of expres ion levels. n = number of unique samples in eachgroup.*Data were generated from TMA.†Statistically significant differences according to the Bonferroni-Holm procedure.‡Data were generated from whole tumor and matched normal samples.
Smith
etal.
Clin
Cancer
Res;
16(14)July
15,2010
Clin
icalCancerRese
arc
h3552
Research.
on Septem
ber 16, 2018. © 2010 A
merican A
ssociation for Cancer
clincancerres.aacrjournals.org D
ownloaded from
Published O
nlineFirst July 6, 2010; D
OI: 10.1158/1078-0432.C
CR
-09-2797
elE
/+
tus

VEGFR-2 and VEGFR-3 Status of Human Solid Tumors
Published OnlineFirst July 6, 2010; DOI: 10.1158/1078-0432.CCR-09-2797
were evaluated by immunohistochemistry for murineCD31 and murine and human VEGFR-2 using 55B11. Asdetermined by CD31, murine vessels were associated withall xenograft tumors. VEGFR-2 localized to a subpopula-tion of murine intratumoral vessels but could not be de-tected in any of the human xenograft tumor cells (Fig. 2B).CD31 and VEGFR-2 microvessel density analysis was done
www.aacrjournals.org
Researcon Septembclincancerres.aacrjournals.org Downloaded from
for each tumor, and the percentage of VEGFR-2–positivetumor vessels was calculated (Fig. 2A). Less than 50% ofthe supporting CD31-positive vasculature was VEGFR-2positive for the majority of tumor xenograft models. Therespective presence and absence of VEGFR-2 in tumor vas-culature and tumor cells were confirmed by comparing thereceptor expression in a selection of five xenograft tumor
Fig. 1. Specificity of VEGFR-2 and VEGFR-3 antibodies. A, the specificity of antibodies raised to VEGFR-2 (55B11, sc-315, sc-504, and sc-6251) andVEGFR-3 (AF349) was determined by Western blot analysis of protein extracts from PAE and NIH-3T3 cells engineered to overexpress VEGFR-1, VEGFR-2,VEGFR-3, and CSF1R and cells expressing endogenous PDGFR-β (VSMC) and c-Kit (M-07e). B, 55B11 and AF349 were evaluated by immunocytochemicalanalysis of FFPE PAE cells engineered to overexpress VEGFRs.
Clin Cancer Res; 16(14) July 15, 2010 3553
h. er 16, 2018. © 2010 American Association for Cancer

Smith et al.
3554
Published OnlineFirst July 6, 2010; DOI: 10.1158/1078-0432.CCR-09-2797
Fig. 2. VEGFR-2 expression in human tumor xenograft models. TMAs representing 29 different xenograft models of human tumors (n = maximum 12 coresper model) were analyzed by immunohistochemistry for VEGFR-2 and CD31. A, using an ACIS II at ×10 magnification, the mean CD31+ and VEGFR-2+
vessel number per mm2 of viable tumor [microvessel density (MVD)] and the ratio of VEGFR-2+ to CD31+ vessels for each model was calculated andplotted. B, typical VEGFR-2 immunostaining of tumor vasculature in xenograft models. C, evaluation of VEGFR-2 antibodies 55B11, A3, sc-315, and sc-504by Western blot analysis of xenograft tumor lysates compared with their matched parental cell lines.
Clin Cancer Res; 16(14) July 15, 2010 Clinical Cancer Research
Research. on September 16, 2018. © 2010 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
VEGFR-2 and VEGFR-3 Status of Human Solid Tumors
Published OnlineFirst July 6, 2010; DOI: 10.1158/1078-0432.CCR-09-2797
lysates [HT-29 and Colo-205 (colon), A549 and PC-9(lung), and MCF-7 (breast)] compared with their matchedparental cell lines (Fig. 2C). The 230-kDa VEGFR-2 dou-blet was present in each of the xenograft models testedbut could not be detected in the parental cell lines, sug-gesting that the receptor was present in murine stromabut not in the human tumor. Moreover, VEGFR-2 levelsin xenograft tumors paralleled that of CD31, indicating avascular association.The antibody AF349 reacts with only human VEGFR-3
and was therefore used to assess whether VEGFR-3 wasexpressed in human tumor xenografts. Using AF349,VEGFR-3 could not be detected in any of the human xeno-graft tumor cells (data not shown).
Quality assessment of human archival tumor tissueTo ensure that only suitable archival FFPE human tis-
sues was used for immunohistochemical analyses, tissuesamples were subjected to a series of morphologic(H&E) and immunohistochemical (CD31 and p-Tyr)checks to determine tissue quality and the extent of anti-gen preservation, respectively. Morphologic assessment ofH&E sections for each specimen was done by a patholo-
www.aacrjournals.org
Researcon Septembclincancerres.aacrjournals.org Downloaded from
gist. CD31 and total protein tyrosine phosphorylationimmunostaining were examined to assess antigen preser-vation in the vasculature and general preservation ofphosphorylated proteins, respectively. Specimens with in-sufficient material, poor morphology, or poor antigenpreservation were not used in this study (14% of cases).
Expression of VEGFR-2 and VEGFR-3 in humanprimary solid tumorsTMAs were constructed from archival whole tissue
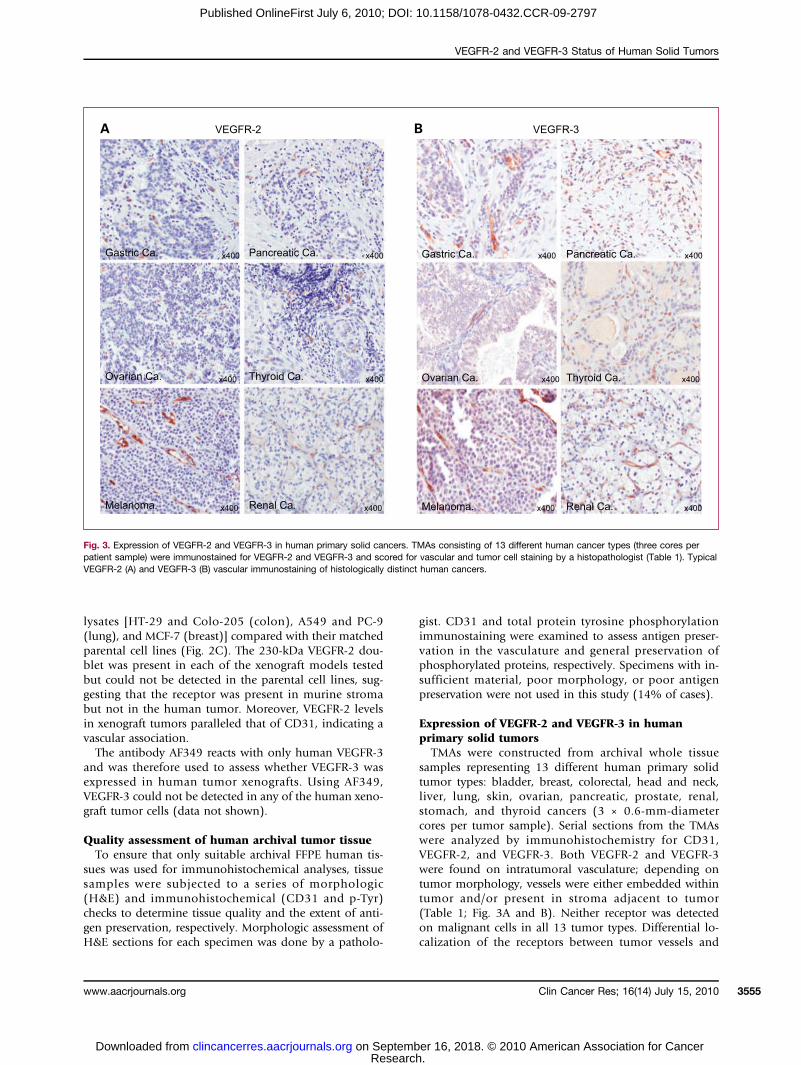
samples representing 13 different human primary solidtumor types: bladder, breast, colorectal, head and neck,liver, lung, skin, ovarian, pancreatic, prostate, renal,stomach, and thyroid cancers (3 × 0.6-mm-diametercores per tumor sample). Serial sections from the TMAswere analyzed by immunohistochemistry for CD31,VEGFR-2, and VEGFR-3. Both VEGFR-2 and VEGFR-3were found on intratumoral vasculature; depending ontumor morphology, vessels were either embedded withintumor and/or present in stroma adjacent to tumor(Table 1; Fig. 3A and B). Neither receptor was detectedon malignant cells in all 13 tumor types. Differential lo-calization of the receptors between tumor vessels and
Fig. 3. Expression of VEGFR-2 and VEGFR-3 in human primary solid cancers. TMAs consisting of 13 different human cancer types (three cores perpatient sample) were immunostained for VEGFR-2 and VEGFR-3 and scored for vascular and tumor cell staining by a histopathologist (Table 1). TypicalVEGFR-2 (A) and VEGFR-3 (B) vascular immunostaining of histologically distinct human cancers.
Clin Cancer Res; 16(14) July 15, 2010 3555
h. er 16, 2018. © 2010 American Association for Cancer

Smith et al.
3556
Published OnlineFirst July 6, 2010; DOI: 10.1158/1078-0432.CCR-09-2797
malignant cells was significant for all tumor types (P <0.05; Table 1).
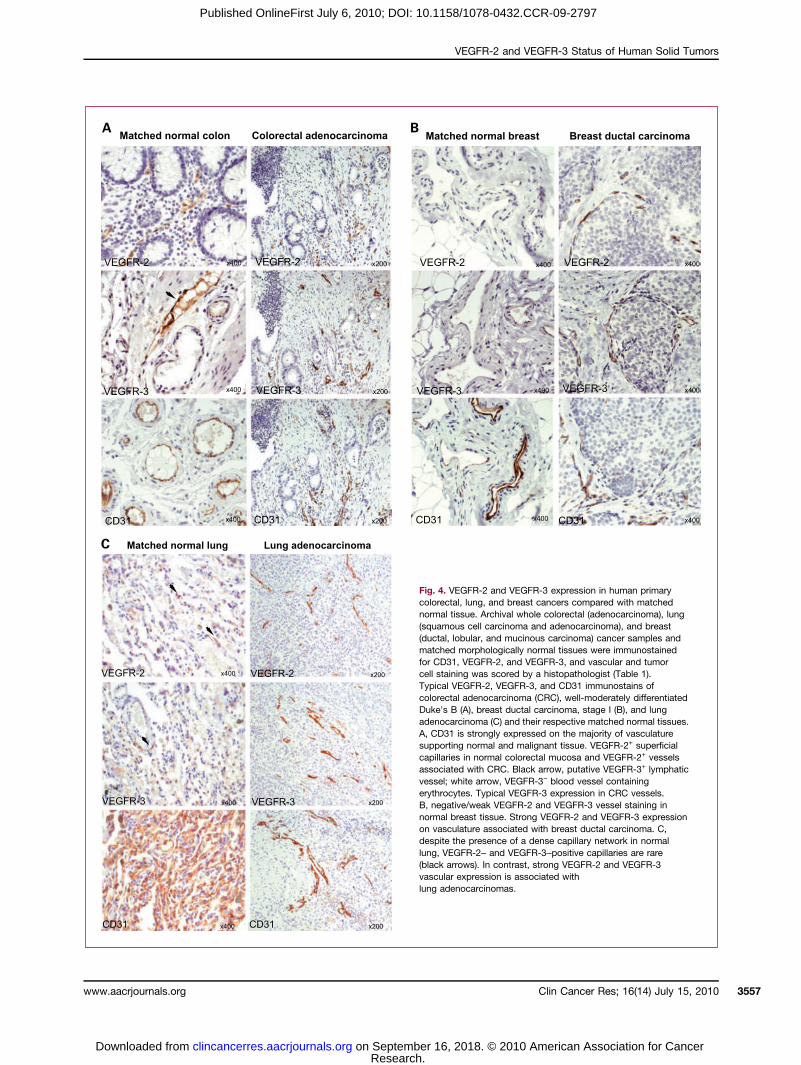
VEGFR-2 and VEGFR-3 distribution in human primarycolorectal, breast, and lung cancers compared withmatched normal tissueArchival whole samples representing CRC (adenocarci-
noma and Dukes' A-D), BC (ductal, lobular, and mucin-ous carcinomas, stage I-III), and NSCLC (squamous cellcarcinoma and adenocarcinoma, stage I-IV) and morpho-logically normal matched adjacent tissues were analyzedby immunohistochemistry for CD31, VEGFR-2, andVEGFR-3 expression (Table 1).Although not precise, vessels having thick walls and
containing erythrocytes are more likely to be blood vessels(BEC), whereas those typified by thin walls and a lack ofRBCs are more likely to be of lymphatic origin (LEC).Weak to medium VEGFR-2 expression was observed pri-marily on putative BECs, supporting mucosa (++) andsubmucosa (+ to ++) in 33 matched normal colorectalsamples (Fig. 4A). VEGFR-2 expression on putative lym-phatic vessels in the submucosa was either absent or weak.VEGFR-2 was localized to intratumoral vasculature, pri-marily with a blood vessel morphology, but was not de-tected on malignant cells in all 65 CRC samplesanalyzed (Fig. 4A). The vascular signal intensity ofVEGFR-2 was higher in tumor compared with normal tis-sue (P = 0.0059; Table 1). In normal colon, VEGFR-3 wasmost prominently expressed (+/++) on the endotheliumof vessels likely to be of lymphatic origin, whereas expres-sion on putative BECs was generally weak (+) or absent(Fig. 4A). VEGFR-3 was confined to tumor vasculature inall CRCs analyzed (n = 40, ++ to +++), with the signal in-tensity significantly greater than that of normal vasculature(P < 0.0001; Table 1). VEGFR-3 tumor cell expression wasnot evident in any CRC.A VEGFR-2 or VEGFR-3 signal was absent from many
CD31+ vessels supporting normal breast tissue (Table 1;Fig. 4B). When vascular expression for either receptorwas observed, this was weak-medium (+/++) in intensity(Table 1). VEGFR-2 and VEGFR-3 were expressed on in-tratumoral vasculature, morphologically resemblingblood vessels, in all BCs analyzed (n = 84 and 56, re-spectively). VEGFR-2 signal intensity was generally greaterthan VEGFR-3 being strong (+++) compared with medi-um (++; Table 1). The signal intensity for both recep-tors was significantly greater on vasculature associatedwith malignant breast tissue than that from matchednormal tissue (P < 0.0001; Fig. 4B). Similar to coloncancers, no tumor cell expression was observed in anyof the cases.Despite the high blood vessel content of normal lung,
as determined by CD31 analysis, VEGFR-2– and VEGFR-3–positive vessels were mainly absent or rare in 31matched normal samples (occasional weakly positivealveoli capillary; Table 1; Fig. 4C). Stronger VEGFR-2signal was observed in neovasculature around lymphoidaggregates in five normal samples. Both receptors were
Clin Cancer Res; 16(14) July 15, 2010
Researcon Septembclincancerres.aacrjournals.org Downloaded from
expressed on intratumoral vasculature (++/+++) but noton tumor cells in 31 NSCLCs, with signal levels signifi-cantly greater on tumor vessels compared with normalvasculature (P < 0.0001; Table 1; Fig. 4C). SpecificVEGFR-2 and VEGFR-3 expression was not detected inmalignant lung cells.
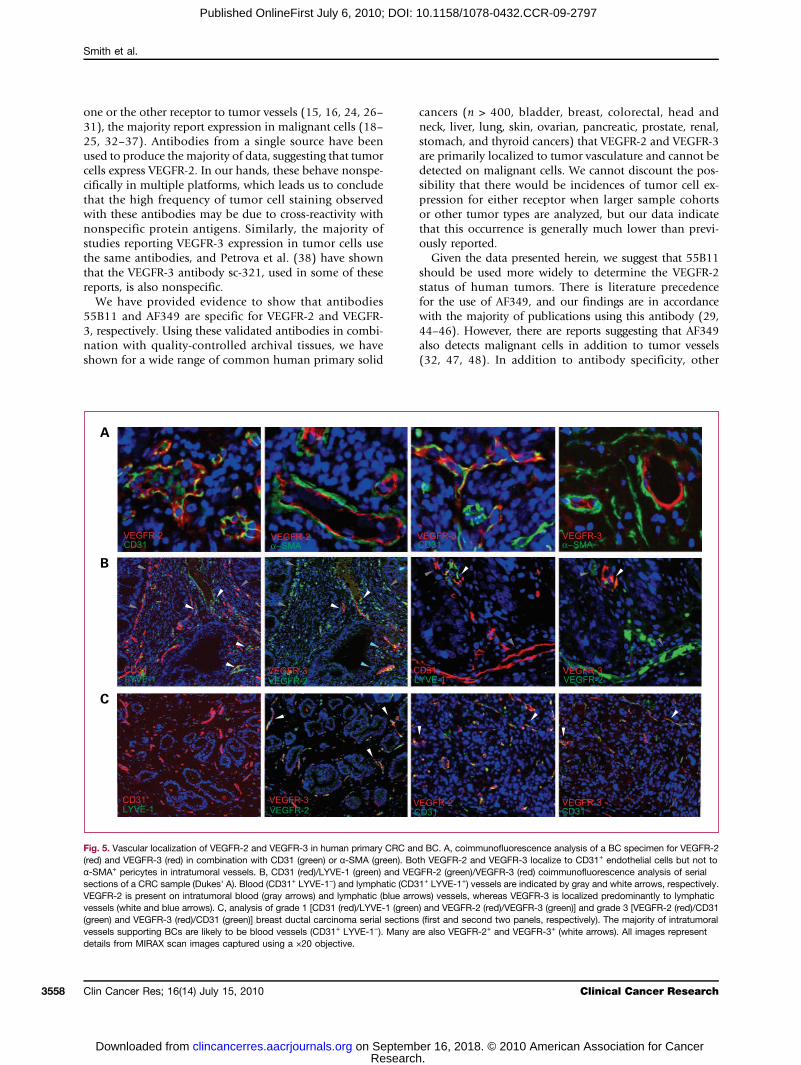
Relative vascular localization of VEGFR-2 andVEGFR-3 in primary CRC and BCTo confirm the endothelial cell localization of VEGFR-2
and VEGFR-3, serial sections from six CRC (adenocarcino-ma and Dukes' A-C) and six BC (ductal carcinoma, grade1-3) samples were immunofluorescently costained forVEGFR-2 (red) or VEGFR-3, (red) in combination withCD31 (green) and α-SMA (green). Both VEGFR-2 andVEGFR-3 were localized to CD31+ endothelial cells butwere absent from α-SMA+ pericytes associated with tumorblood vessels (Fig. 5A). The distinction between BEC andLEC based on vessel morphology alone is subjective, sotumor vessels were characterized further by combiningpan–endothelial cell marker, CD31, and LEC marker,LYVE-1, in coimmunofluorescence analyses of CRC andBC samples (n = 6 each). CD31+ LYVE-1+ and CD31+
LYVE-1− intratumoral vessels were classified as lymphaticand blood, respectively. A section, serial to each CD31/LYVE-1 section, was analyzed for VEGFR-2/VEGFR-3 andcompared with the former to determine the relative bloodand lymphatic vascular localization of VEGFR-2 andVEGFR-3.In CRC, intratumoral vasculature could be subclassified
into CD31+ LYVE-1− (blood vessels, gray arrows in Fig. 5B)and less abundant CD31+ LYVE-1+ (lymphatic vessels,white arrows). VEGFR-2 was present on both types ofvasculature, although the VEGFR-2 signal was generallyweaker for CD31+ LYVE-1+ vessels than for CD31+ LYVE-1−
(Fig. 5B, blue arrows). VEGFR-3 was localized predomi-nantly to intratumoral lymphatic vessels (Fig. 5B, whiteand blue arrows) but was also observed on the occasionalsmall CD31+ LYVE-1− vessel. In addition, VEGFR-2 wasexpressed on vessels transporting erythrocytes, whereasVEGFR-3 was not. In BC, all intratumoral vessels wereCD31+ LYVE-1− and were likely to be blood vessels(Fig. 5C). Although intratumoral lymphatic vessels(CD31+ LYVE-1+) could not be detected, peritumoral lym-phatics (distal to tumor in normal stroma) were detectedin one larger sample. A subpopulation of CD31+ LYVE-1+
vessels was also VEGFR-2+ and, to a lesser extent, VEGFR-3+,indicating that both receptors are expressed on a subsetof blood vessels in BC (Fig. 5C, white arrows). Indeed,both VEGFR-2 and VEGFR-3 were found on breast tumorvessels carrying RBCs.
Discussion
Published literature presents conflicting reports to theextent and localization of VEGFR-2 and VEGFR-3 expres-sion in human tumors. Although some studies confine
Clinical Cancer Research
h. er 16, 2018. © 2010 American Association for Cancer
VEGFR-2 and VEGFR-3 Status of Human Solid Tumors
Published OnlineFirst July 6, 2010; DOI: 10.1158/1078-0432.CCR-09-2797
Fig. 4. VEGFR-2 and VEGFR-3 expression in human primarycolorectal, lung, and breast cancers compared with matchednormal tissue. Archival whole colorectal (adenocarcinoma), lung(squamous cell carcinoma and adenocarcinoma), and breast(ductal, lobular, and mucinous carcinoma) cancer samples andmatched morphologically normal tissues were immunostainedfor CD31, VEGFR-2, and VEGFR-3, and vascular and tumorcell staining was scored by a histopathologist (Table 1).Typical VEGFR-2, VEGFR-3, and CD31 immunostains ofcolorectal adenocarcinoma (CRC), well-moderately differentiatedDuke's B (A), breast ductal carcinoma, stage I (B), and lungadenocarcinoma (C) and their respective matched normal tissues.A, CD31 is strongly expressed on the majority of vasculaturesupporting normal and malignant tissue. VEGFR-2+ superficialcapillaries in normal colorectal mucosa and VEGFR-2+ vesselsassociated with CRC. Black arrow, putative VEGFR-3+ lymphaticvessel; white arrow, VEGFR-3− blood vessel containingerythrocytes. Typical VEGFR-3 expression in CRC vessels.B, negative/weak VEGFR-2 and VEGFR-3 vessel staining innormal breast tissue. Strong VEGFR-2 and VEGFR-3 expressionon vasculature associated with breast ductal carcinoma. C,despite the presence of a dense capillary network in normallung, VEGFR-2– and VEGFR-3–positive capillaries are rare(black arrows). In contrast, strong VEGFR-2 and VEGFR-3vascular expression is associated withlung adenocarcinomas.
www.aacrjournals.org
Researcon Septembclincancerres.aacrjournals.org Downloaded from
Clin Cancer Res; 16(14) July 15, 2010 3557
h. er 16, 2018. © 2010 American Association for Cancer
Smith et al.
3558
Published OnlineFirst July 6, 2010; DOI: 10.1158/1078-0432.CCR-09-2797
one or the other receptor to tumor vessels (15, 16, 24, 26–31), the majority report expression in malignant cells (18–25, 32–37). Antibodies from a single source have beenused to produce the majority of data, suggesting that tumorcells express VEGFR-2. In our hands, these behave nonspe-cifically in multiple platforms, which leads us to concludethat the high frequency of tumor cell staining observedwith these antibodies may be due to cross-reactivity withnonspecific protein antigens. Similarly, the majority ofstudies reporting VEGFR-3 expression in tumor cells usethe same antibodies, and Petrova et al. (38) have shownthat the VEGFR-3 antibody sc-321, used in some of thesereports, is also nonspecific.We have provided evidence to show that antibodies
55B11 and AF349 are specific for VEGFR-2 and VEGFR-3, respectively. Using these validated antibodies in combi-nation with quality-controlled archival tissues, we haveshown for a wide range of common human primary solid
Clin Cancer Res; 16(14) July 15, 2010
Researcon Septembclincancerres.aacrjournals.org Downloaded from
cancers (n > 400, bladder, breast, colorectal, head andneck, liver, lung, skin, ovarian, pancreatic, prostate, renal,stomach, and thyroid cancers) that VEGFR-2 and VEGFR-3are primarily localized to tumor vasculature and cannot bedetected on malignant cells. We cannot discount the pos-sibility that there would be incidences of tumor cell ex-pression for either receptor when larger sample cohortsor other tumor types are analyzed, but our data indicatethat this occurrence is generally much lower than previ-ously reported.Given the data presented herein, we suggest that 55B11
should be used more widely to determine the VEGFR-2status of human tumors. There is literature precedencefor the use of AF349, and our findings are in accordancewith the majority of publications using this antibody (29,44–46). However, there are reports suggesting that AF349also detects malignant cells in addition to tumor vessels(32, 47, 48). In addition to antibody specificity, other
Fig. 5. Vascular localization of VEGFR-2 and VEGFR-3 in human primary CRC and BC. A, coimmunofluorescence analysis of a BC specimen for VEGFR-2(red) and VEGFR-3 (red) in combination with CD31 (green) or α-SMA (green). Both VEGFR-2 and VEGFR-3 localize to CD31+ endothelial cells but not toα-SMA+ pericytes in intratumoral vessels. B, CD31 (red)/LYVE-1 (green) and VEGFR-2 (green)/VEGFR-3 (red) coimmunofluorescence analysis of serialsections of a CRC sample (Dukes' A). Blood (CD31+ LYVE-1−) and lymphatic (CD31+ LYVE-1+) vessels are indicated by gray and white arrows, respectively.VEGFR-2 is present on intratumoral blood (gray arrows) and lymphatic (blue arrows) vessels, whereas VEGFR-3 is localized predominantly to lymphaticvessels (white and blue arrows). C, analysis of grade 1 [CD31 (red)/LYVE-1 (green) and VEGFR-2 (red)/VEGFR-3 (green)] and grade 3 [VEGFR-2 (red)/CD31(green) and VEGFR-3 (red)/CD31 (green)] breast ductal carcinoma serial sections (first and second two panels, respectively). The majority of intratumoralvessels supporting BCs are likely to be blood vessels (CD31+ LYVE-1−). Many are also VEGFR-2+ and VEGFR-3+ (white arrows). All images representdetails from MIRAX scan images captured using a ×20 objective.
Clinical Cancer Research
h. er 16, 2018. © 2010 American Association for Cancer

VEGFR-2 and VEGFR-3 Status of Human Solid Tumors
Published OnlineFirst July 6, 2010; DOI: 10.1158/1078-0432.CCR-09-2797
factors, such as tissue quality, antigen retrieval, and stringen-cy of the immunohistochemical reaction, can influence theaccurate detection of an epitope, which may explain theseobservations. Although not used in these analyses, we alsofound antibody clone 9D9F9 to be specific for VEGFR-3,which corroborates recent data by Petrova et al. (38). Similarto our findings with AF349, 9D9F9 has been shown to detectprimarily vasculature in lung (31, 38), breast (27, 30, 38),and colorectal tumors (26, 38). Using 9D9F9 inWestern blotanalysis, Morelli et al. (49) observed VEGFR-3 to be presentin gastrointestinal tumor cell lines, including colorectal linesHT-29, SW620, and HCT116, whereas we found it to beabsent from these cells grown in vivo and primary colorectaltumors. These data indicate potential differences in theexpression of VEGFR-3 between cultured tumor cell linesand cell lines grown s.c. as xenografts or primary humantumors.We have shown that VEGFR-2 and VEGFR-3 are ex-
pressed infrequently and at low levels in endothelium,supporting morphologically normal tissue adjacent tobreast and lung tumors. Angiogenesis is largely absentfrom nonpathologic adult tissues, and so, most endo-thelium is quiescent and expected to express low levelsof VEGFRs. However, we observed that VEGFR-2and VEGFR-3 were frequently expressed on blood andlymphatic vessels, respectively, in apparently normal co-lon. This may be expected, as epithelial turnover ismore rapid in normal colonic epithelium than breastand lung tumors. In addition, colorectal tumors maydiffuse angiogenic factors (e.g., VEGF and interleukin-8) into the surrounding normal tissue, which promoteangiogenesis (50).VEGFR-2 and VEGFR-3 were significantly upregulated
on vessels, supporting all colorectal, breast, and lungcancers analyzed compared with matched normal tissue.Moreover, neither receptor could be detected in tumorcells derived from these tissues. In contrast to our data,all previous NSCLC studies report that VEGFR-2 is pre-dominantly (48-100%) found on malignant cells (18,19, 25), although vessel expression is also observed insome (18, 19). Similar to our findings, VEGFR-2 haspreviously been localized to vessels, supporting BCs(16, 17); however, there is only one report of upregula-tion on vessels (16) and the majority of other studiesclaim a breast tumor cell localization (22–24). SimilarVEGFR-3 vascular expression data have been reportedfor BCs (27–30) and NSCLCs (31); however, others re-port localization of VEGFR-3 to malignant breast (23,35) or lung cells (19, 25, 32, 33, 36, 37). VEGFR-2was originally shown to be restricted to vessels associat-ed with CRCs (15). Our data agree with this original re-port but is in contradiction to subsequent work,suggesting that VEGFR-2 expression is expressed onCRC cells (20, 21). Likewise, our findings support anearlier observation that VEGFR-3 expression is restrictedto tumor vessels in the majority of CRCs (36), but con-trast with a study reporting a CRC cell localization forVEGFR-3 (34).
www.aacrjournals.org
Researcon Septembclincancerres.aacrjournals.org Downloaded from
We used vascular markers CD31 and LYVE-1 to betterclassify blood and lymphatic vessels, respectively, andconfirm our original morphologic observations that CRCand BC differ with respect to localization of VEGFR-2 andVEGFR-3 on these two vascular subpopulations. Our dataindicate that in CRC, VEGFR-2 is present on both intratu-moral blood and lymphatic vessels, whereas VEGFR-3 ispredominantly localized to lymphatic vessels. In BC, intra-tumoral lymphatic vessels are scarce and we have shownthat both VEGFR-2 and VEGFR-3 are localized to bloodvessels. This vascular localization pattern combined within vivo functional data implicating VEGFR-3 in sproutingangiogenesis (8) suggests that VEGFR-3, in combinationwith VEGFR-2, may have a stronger angiogenic role inBC than CRC. In addition, the relative balance of thetwo receptors between blood and lymphatic vessels couldinfluence the tumor response to VEGF therapy andwarrants further investigation.Our data may also provide some insights into the mech-
anism of action of VEGF signaling inhibitors in the clinic.Tumor endothelial cells, both lymphatic and blood, andnot tumor cells are the likely direct cellular targets for an-ti–VEGFR-2 and anti–VEGFR-3 therapy in many primarysolid tumors. The upregulation of VEGFR-3 on blood ves-sels suggests that dual VEGFR-2/VEGFR-3 inhibitors mayhave additional antiangiogenic benefits. This and in vivofindings indicating that VEGFR-3 can sustain low-level an-giogenesis even in the presence of VEGFR-2 inhibitors (8)suggest that tumors expressing high levels of VEGFR-3 onblood vessels may be less sensitive to VEGFR therapies thattarget VEGFR-2 alone.An accurate understanding of the expression levels
and localization of drug targets in disease is importantfor elucidating the mechanism of action of drugs in theclinic and, potentially, for identifying patients whowould gain most benefit from treatment with VEGFRsignaling inhibitors. Because immunohistochemicalmethodology has many potential pitfalls and confound-ing factors, our work suggests that greater antibody val-idation and tissue quality control measures than hashitherto generally been the case would give greaterconfidence in immunolocalization results for VEGFR-2and VEGFR-3 in clinical samples.
Disclosure of Potential Conflicts of Interest
All authors are full-time employees of AstraZeneca and hold shares inthe company.
Acknowledgments
We thank the Wales Cancer Bank for providing the human tumorsamples.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
Received 10/27/2009; revised 05/20/2010; accepted 05/20/2010;published OnlineFirst 07/06/2010.
Clin Cancer Res; 16(14) July 15, 2010 3559
h. er 16, 2018. © 2010 American Association for Cancer

Smith et al.
3560
Published OnlineFirst July 6, 2010; DOI: 10.1158/1078-0432.CCR-09-2797
References
1. Ferrara N. VEGF and the quest for tumor angiogenesis factors. NatRev Cancer 2002;2:795–803.2. Meyer M, Clauss M, Lepple-Wienhues A, et al. A novel vascular en-
dothelial growth factor encoded by Orf virus, VEGF-E, mediates an-giogenesis via signalling through VEGFR-2 (KDR) but not VEGFR-1(Flt-1) receptor tyrosine kinases. EMBO J 1999;18:363–74.
3. Gille H, Kowalski J, Li B. Analysis of biological effects and signalingproperties of Flt-1 (VEGFR-1) and KDR (VEGFR-2). A reassessmentusing novel receptor-specific vascular endothelial growth factor mu-tants. J Biol Chem 2001;276:3222–30.
4. Gerhardt H, Golding M, Fruttiger M, et al. VEGF guides angiogenicsprouting utilizing endothelial tip cell filopodia. J Cell Biol 2003;161:1163–77.
5. Veikkola T, Karkkainen M, Claesson-Welsh L, Alitalo K. Regulation ofangiogenesis via vascular endothelial growth factor receptors.Cancer Res 2000;60:203–12.
6. Kaipainen A, Korhonen J, Mustonen T, et al. Expression of the fms-like tyrosine kinase 4 gene becomes restricted to lymphatic endo-thelium during development. Proc Natl Acad Sci U S A 1995;92:3566–70.
7. Veikkola T, Jussila L, Makinen T, et al. Signalling via vascular endo-thelial growth factor receptor-3 is sufficient for lymphangiogenesis intransgenic mice. EMBO J 2001;20:1223–31.
8. Tammela T, Zarkada G, Wallgard E, Murtomäki A, Suchting S,Wirzenius M. Blocking VEGFR-3 suppresses angiogenic sproutingand vascular network formation. Nature 2008;454:656–60.
9. Grothey A, Ellis LM. Targeting angiogenesis driven by vascular endo-thelial growth factors using antibody-based therapies. Cancer J2008;14:170–7.
10. Ainsworth NL, Lee JSZ, Eisen T. Impact of anti-angiogenic treat-ments on metastatic renal cell carcinoma. Expert Rev AnticancerTher 2009;9:1793–805.
11. Bukowski RM, Yasothan U, Kirkpatrick P. Pazopanib. Nat Rev DrugDiscov 2010;9:17–8.
12. Wedge SR, Kendrew J, Hennequin LF, et al. AZD2171: a highly po-tent, orally bioavailable, vascular endothelial growth factor receptor-2 tyrosine kinase inhibitor for the treatment of cancer. Cancer Res2005;65:4389–400.
13. Padera TP, Kuo AH, Hoshida T, et al. Differential response of pri-mary tumor versus lymphatic metastasis to VEGFR-2 and VEGFR-3kinase inhibitors cediranib and vandetanib. Mol Cancer Ther 2008;7:2272–9.
14. Heckman CA, Holopainen T, Wirzenius M, et al. The tyrosine kinaseinhibitor cediranib blocks ligand-induced vascular endothelial growthfactor receptor-3 activity and lymphangiogenesis. Cancer Res 2008;68:4754–62.
15. Takahashi Y, Kitadai Y, Bucana CD, Cleary KR, Ellis LM. Expressionof vascular endothelial growth factor and its receptor, KDR, corre-lates with vascularity, metastasis, and proliferation of human coloncancer. Cancer Res 1995;55:3964–8.
16. Kranz A, Mattfeldt T, Waltenberger J. Molecular mediators of tumorangiogenesis: enhanced expression and activation of vascular endo-thelial growth factor receptor KDR in primary breast cancer. Int JCancer 1999;84:293–8.
17. Dales JP, Garcia S, Carpentier S, et al. Prediction of metastasis risk(11 year follow-up) using VEGF-R1, VEGF-R2, Tie-2/Tek and CD105expression in breast cancer (n = 905). Br J Cancer 2004;90:1216–21.
18. Seto T, Higashiyama M, Funai H, et al. Prognostic value of expres-sion of vascular endothelial growth factor and its flt-1 and KDR re-ceptors in stage I non-small-cell lung cancer. Lung Cancer 2006;53:91–6.
19. Kajita T, Ohta Y, Kimura K, et al. The expression of vascular endo-thelial growth factor C and its receptors in non-small cell lung cancer.Br J Cancer 2001;85:255–60.
20. Amaya H, Tanigawa N, Lu C, et al. Association of vascular endothe-lial growth factor expression with tumor angiogenesis, survival andthymidine phosphorylase/platelet-derived endothelial cell growthfactor expression in human colorectal cancer. Cancer Lett 1997;119:227–35.
Clin Cancer Res; 16(14) July 15, 2010
Researcon Septembclincancerres.aacrjournals.org Downloaded from
21. Duff SE, Jeziorska M, Rosa DD, et al. Vascular endothelial growthfactors and receptors in colorectal cancer: implications for anti-angiogenic therapy. Eur J Cancer 2006;42:112–7.
22. Rydén L, Linderholm B, Nielsen NH, Emdin S, Jönsson PE, LandbergG. Tumor specific VEGF-A andVEGFR2/KDRprotein are co-expressedin breast cancer. Breast Cancer Res Treat 2003;82:147–54.
23. Wülfing P, Kersting C, Buerger H, et al. Expression patterns of angio-genic and lymphangiogenic factors in ductal breast carcinomain situ. Br J Cancer 2005;92:1720–8.
24. Ghosh S, Sullivan CA, Zerkowski MP, et al. High levels of vascularendothelial growth factor and its receptors (VEGFR-1,VEGFR-2,neuropilin-1) are associated with worse outcome in breast cancer.Hum Pathol 2008;39:1835–43.
25. Donnem T, Al-Saad S, Al-Shibli K, et al. Inverse prognostic impact ofangiogenic marker expression in tumor cells versus stromal cells innon small cell lung cancer. Clin Cancer Res 2007;13:6649–57.
26. White JD, Hewett PW, Kosuge D, et al. Vascular endothelial growthfactor-D expression is an independent prognostic marker for survivalin colorectal carcinoma. Cancer Res 2002;62:1669–75.
27. Valtola R, Salven P, Heikkilä P, et al. VEGFR-3 and its ligand VEGF-Care associated with angiogenesis in breast cancer. Am J Pathol1999;154:1381–90.
28. Nathanson SD, Zarbo RJ, Wachna DL, Spence CA, AndrzejewskiTA, Abrams J. Microvessels that predict axillary lymph node me-tastases in patients with breast cancer. Arch Surg 2000;135:586–93.
29. Nakamura Y, Yasuoka H, Tsujimoto M, et al. Flt-4-positive vesseldensity correlates with vascular endothelial growth factor-d expres-sion, nodal status, and prognosis in breast cancer. Clin Cancer Res2003;9:5313–7.
30. Clarijs R, Schalkwijk L, Hofmann UB, Ruiter DJ, de Waal RM. Induc-tion of vascular endothelial growth factor receptor-3 expression ontumor microvasculature as a new progression marker in human cu-taneous melanoma. Cancer Res 2002;62:7059–65.
31. Niki T, Iba S, Yamada T, Matsuno Y, Enholm B, Hirohashi S. Expres-sion of vascular endothelial growth factor receptor 3 in blood andlymphatic vessels of lung adenocarcinoma. J Pathol 2001;193:450–7.
32. Chen F, Takenaka K, Ogawa E, et al. Flt-4-positive endothelial celldensity and its clinical significance in non-small cell lung cancer. ClinCancer Res 2004;10:8548–53.
33. Kojima H, Shijubo N, Yamada G, et al. Clinical significance of vascu-lar endothelial growth factor-C and vascular endothelial growth fac-tor receptor 3 in patients with T1 lung adenocarcinoma. Cancer2005;104:1668–77.
34. Witte D, Thomas A, Ali N, Carlson N, Younes M. Expression of thevascular endothelial growth factor receptor-3 (VEGFR-3) and its li-gand VEGF-C in human colorectal adenocarcinoma. AnticancerRes 2002;22:1463–6.
35. Mylona E, Alexandrou P, Mpakali A, et al. Clinicopathological andprognostic significance of vascular endothelial growth factors(VEGF)-C and -D and VEGF receptor 3 in invasive breast carcinoma.Eur J Surg Oncol 2007;33:294–300.
36. Arinaga M, Noguchi T, Takeno S, Chujo M, Miura T, Uchida Y. Clin-ical significance of vascular endothelial growth factor C and vascularendothelial growth factor receptor 3 in patients with non small celllung carcinoma. Cancer 2003;97:457–64.
37. Saintigny P, Kambouchner M, Ly M, et al. Vascular endothelialgrowth factor-C and its receptor VEGFR-3 in non-small-cell lungcancer: concurrent expression in cancer cells from primary tumorand metastatic lymph node. Lung Cancer 2007;58:205–13.
38. Petrova TV, Bono P, Holnthoner W, et al. VEGFR-3 expression is re-stricted to blood and lymphatic vessels in solid tumors. Cancer Cell2008;13:554–6.
39. Smith NR, James NH, Oakley I, et al. Acute pharmacodynamic andantivascular effects of the vascular endothelial growth factor signal-ing inhibitor AZD2171 in Calu-6 human lung tumor xenografts. MolCancer Ther 2007;6:2198–208.
40. Takahashi T, Shibuya M. The 230 kDa mature form of KDR/Flk-1
Clinical Cancer Research
h. er 16, 2018. © 2010 American Association for Cancer

VEGFR-2 and VEGFR-3 Status of Human Solid Tumors
Published OnlineFirst July 6, 2010; DOI: 10.1158/1078-0432.CCR-09-2797
(VEGF receptor-2) activates the PLC-γ pathway and partially inducesmitotic signals in NIH3T3 fibroblasts. Oncogene 1997;14:2079–89.
41. Chen Z, Lee FY, Bhalla KN, Wu J. Potent inhibition of platelet-derivedgrowth factor-induced responses in vascular smooth muscle cells byBMS-354825 (dasatinib). Mol Pharmacol 2006;69:1527–33.
42. Kuriu A, IkedaH, Kanakura Y, et al. Proliferation of humanmyeloid leu-kemia cell line associated with the tyrosine-phosphorylation and acti-vation of the proto-oncogene c-kit product. Blood 1991;78:2834–40.
43. Petrova TV, Makinen T, Alitalo K. Signaling via vascular endothelialgrowth factor receptors. Exp Cell Res 1999;253:117–30.
44. Neuchrist C, Erovic BM, Handisurya A, et al. Vascular endothelialgrowth factor C and vascular endothelial growth factor receptor 3expression in squamous cell carcinomas of the head and neck. HeadNeck 2003;25:464–74.
45. Zeng Y, Opeskin K, Baldwin ME, et al. Expression of vascular endo-thelial growth factor receptor-3 by lymphatic endothelial cells is as-sociated with lymph node metastasis in prostate cancer. Clin CancerRes 2004;10:5137–44.
www.aacrjournals.org
Researcon Septembclincancerres.aacrjournals.org Downloaded from
46. Yu XM, Lo CY, Chan WF, Lam KY, Leung P, Luk JM. Increased ex-pression of vascular endothelial growth factor C in papillary thyroidcarcinoma correlates with cervical lymph node metastases. ClinCancer Res 2005;11:8063–9.
47. Yokoyama Y, Charnock-Jones DS, Licence D, et al. Expression ofvascular endothelial growth factor (VEGF)-D and its receptor, VEGFreceptor 3, as a prognostic factor in endometrial carcinoma. ClinCancer Res 2003;9:1361–9.
48. Jennbacken K, Vallbo C, Wang W, Damber JE. Expression of vascu-lar endothelial growth factor C (VEGF-C) and VEGF receptor-3 inhuman prostate cancer is associated with regional lymph node me-tastasis. Prostate 2005;65:110–6.
49. Morelli MP, Brown AM, Pitts TM, et al. Targeting vascular endothelialgrowth factor receptor-1 and -3 with cediranib (AZD2171): effects onmigration and invasion of gastrointestinal cancer cell lines. Mol Can-cer Ther 2009;8:2546–58.
50. Fox SH, Whalen GF, Sanders MM, et al. Angiogenesis in normal tis-sue adjacent to colon cancer. J Surg Oncol 1998;69:230–4.
Clin Cancer Res; 16(14) July 15, 2010 3561
h. er 16, 2018. © 2010 American Association for Cancer

2010;16:3548-3561. Published OnlineFirst July 6, 2010.Clin Cancer Res Neil R. Smith, Dawn Baker, Neil H. James, et al. Primary Solid CancersVEGFR-3 Are Localized Primarily to the Vasculature in Human Vascular Endothelial Growth Factor Receptors VEGFR-2 and
Updated version
10.1158/1078-0432.CCR-09-2797doi:
Access the most recent version of this article at:
Material
Supplementary
http://clincancerres.aacrjournals.org/content/suppl/2010/08/25/1078-0432.CCR-09-2797.DC1
Access the most recent supplemental material at:
Cited articles
http://clincancerres.aacrjournals.org/content/16/14/3548.full#ref-list-1
This article cites 50 articles, 22 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/16/14/3548.full#related-urls
This article has been cited by 9 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/16/14/3548To request permission to re-use all or part of this article, use this link
Research. on September 16, 2018. © 2010 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 6, 2010; DOI: 10.1158/1078-0432.CCR-09-2797