recent advances in basic science sweet-talk: role of host ... · sweet-talk: role of host...
TRANSCRIPT

Sweet-talk: role of host glycosylationin bacterial pathogenesis of thegastrointestinal tractA P Moran,1,2,3 A Gupta,1,2 L Joshi2,3
ABSTRACTGlycosylation is a key modification of proteins and lipidsand is involved in most intermolecular and intercellularinteractions. The gastrointestinal mucus gel is continuousand can be divided into two layers: a secreted looselyassociated layer and a layer firmly attached to themucosa. In addition, the membrane-bound glycosylatedproteins and lipids create a glycocalyx, which remainsadherent on each cell and is dynamic and responsive tothe physiological state and environment of the cell. Thesecreted glycans form a mucus gel layer that serves asa physicochemical sensor and barrier network and isprimarily composed of mucins and associated peptides.These glycans protect gut epithelial cells from chemical,biological and physical insults and are continuouslyrenewed. Pathogens colonise and invade the hostepithelial cells using proteineprotein and glycanelectininteractions. During the process of colonisation andinfection, the glycosylation state of both host andpathogen change in response to the presence of theother. This complex modulation of glycan expressioncritically determines pathogenesis and the host responsein terms of structural changes and immune response. Inaddition, by influencing host immunity and gutglycosylation, the microbiota can further effect protectionagainst pathogens. In this review, the roles of hostglycosylation in interactions with two prevalent bacterialpathogens, Campylobater jejuni and Helicobacter pylori,are discussed to illustrate important concepts inpathogenesis.
INTRODUCTIONBeyond genes and proteinsFor the past 60 years, the central dogma of molec-ular biology has emphasised the importance of thetranscription of DNA to RNA and translation ofRNA into amino acids to form proteins. By theirphysical and chemical nature, DNA, RNA andproteins are linear molecules and are synthesisedusing a pre-existing template. However, with thecompletion of sequencing of several organisms’genomes, it has emerged that the complement ofgenes alone does not define the complexity of anorganism and its interactions with other organisms.Once amino acid chains are synthesised, they canundergo over 200 covalent modifications, so-calledpost-translational modifications, which determinethe biological, physical and chemical properties ofthe proteins.1 The realisation that proteins areheavily modified, and that these modifications are
critical to their structure and function, has led toa shift in emphasis from primarily studying genesand proteins to the analysis of post-translationalmodifications, such as phosphorylation, sulfationand glycosylation.
What is glycosylation?Glycosylation is one of the most prevalent modifi-cations mediated by complex enzymaticmachinery, whereby glycans (sugars) are covalentlyattached to specific amino acid sites of proteins.Mammalian glycosylation takes place on nascentprotein chains when they are passing through theendoplasmic reticulum (ER) and the Golgi complex.Subsequently, these proteins and lipids are trans-ported to different organelles in the cell, to the cellmembrane, or secreted from the cell.All cells, from bacteria to human, are covered in
glycans. Glycosylation leads to the formation ofmonomeric and multimeric glycan linkages whichare essential for cell viability, biochemical commu-nication and normal function. The resultingglycome encompasses a diverse and abundantrepertoire of glycans, which form one of the fourfundamental macromolecular components of allcells (together with nucleic acids, proteins andlipids). Glycans have important biological functionsin protein maturation and turnover, cell adhesionand trafficking, and receptor binding and activa-tion. Protein glycosylation has crucial roles in mostphysiological processes and diseases, including cellsignalling, cellular differentiation and adhesion.Dysfunction leads to pathogenesis of infectiousdiseases and development of chronic diseases, forexample, cancer, neurological disorders and severeinflammation.2
The diversity and complex characteristic ofglycans on glycoconjugates is derived from themany ways in which monosaccharides can belinked together during their synthesis to formhigher-order structures (figure 1B). Unlike nucleicacids and proteins, which are synthesised on thebasis of a template and are linear molecules,glycan synthesis is not template-based andglycans can be both linear and branched. They canproduce a large number of isomers because of thepossibility of linkage and anomeric isomers. Theseisomers have very different three-dimensionalstructures and biological activities. A mono-saccharide can be involved in more than twoglycosidic linkages, thus serving as a branch point.
1Microbiology, School of NaturalSciences, National University ofIreland, Galway, Ireland2Alimentary GlycoscienceResearch Cluster, NationalUniversity of Ireland, Galway,Ireland3National Centre for BiomedicalEngineering Science, NationalUniversity of Ireland, Galway,Ireland
Correspondence toProfessor L Joshi, NationalCentre for BiomedicalEngineering Science, NationalUniversity of Ireland, Galway,Ireland;[email protected]
Published Online First12 January 2011
1412 Gut 2011;60:1412e1425. doi:10.1136/gut.2010.212704
Recent advances in basic science
on May 21, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.2010.212704 on 12 January 2011. D
ownloaded from

The common occurrence of branched sequences isunique to glycans and contributes to their struc-tural diversity. The glycosidic linkage is the mostflexible part of a disaccharide structure. Thus,a disaccharide of well-defined primary structurecan adopt multiple conformations in solution thatdiffer in the relative orientation of the twomonosaccharides. The combination of structuralrigidity and flexibility is typical of complexcarbohydrates and, more than likely, essential totheir biological function.2 3
Glycans are linked to other biomolecules, such aslipids or amino acids within polypeptides, throughglycosidic linkages to form glycoconjugates. Theglycan may be a single monosaccharide or an oligo-saccharide. The attachment of many glycans toa polypeptide scaffold creates tremendous diversityamong glycoproteins (figure 1). Glycoconjugatestherefore make up a highly dynamic system capa-ble of participating in a wide range of biologicalinteractions.3
Nature of sugars and glycosylationdglycoproteins,proteoglycans and glycolipidsMammalian glycoconjugates are classified on thebasis of whether the glycans are attached toproteins or lipids (figure 1A). In the case of proteinglycosylation, there are two major types of glyco-protein, N-linked and O-linked (figure 1B). InN-linked glycoproteins, the glycosylation takesplace on an asparagine residue when asparagine isfound in the following consensus motif: Asn-X-Ser/Thr (where X can be any amino acid but proline).N-Linked glycoproteins go through a complex seriesof attachment events in the ER and the Golgicomplex, with the resultant glycoprotein carryinga mixture of glycan structures varying in lengthfrom five to 15 sugars. These structures areprimarily composed of N-acetyl-D-glucosamine(GlcNAc), D-mannose (Man), D-galactose (Gal),L-fucose (Fuc) and sialic acid also known as neura-minic acid (N-acetylneuraminic acid; Neu5Ac)(figure 1B). Several cell-surface-bound as well as
Figure 1 Glycosylation on the mammalian cell surface. (A) Diagrammatic representation of glycosylation on glycoconjugates of the mammalian cellsurface including glycolipid, proteoglycan, N-linked and O-linked glycoprotein molecules. (B) The types of mammalian glycans, their diversity andcomplexity. Glycan structures of the six classes of secretory glycan (N-glycans, hyaluronan, O-glycans, glycolipids, glycosylphosphatidylinositol (GPI)anchor and glycosaminoglycans) and the single intracellular glycan, O-linked b-N-acetylglucosamine (O-GlcNAc), are shown. Representative examplesof each type are indicated using the symbol nomenclature for monosaccharides (see symbols). PI is phosphatidylinositol, EteP denotesa phosphoethanolamine linkage, and es denotes the sulfation positions of the glycosaminoglycan chains.
Gut 2011;60:1412e1425. doi:10.1136/gut.2010.212704 1413
Recent advances in basic science
on May 21, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.2010.212704 on 12 January 2011. D
ownloaded from

secreted proteins are N-glycosylateddfor example,erythropoietin and immunoglobulins.3 Unlike N-linked glycosylation, O-linked glycosylation takesplace entirely in the Golgi complex where a singleN-acetyl-D-galactosamine (GalNAc) residue isenzymatically attached to the Ser/Thr residue onfolded proteins (figure 1B). In general, O-linkedglycan chains are shorter than N-linked glycanchains but are composed of the same mono-saccharides as used in N-linked glycosylation. Cell-surface-bound and secreted mucins are examples ofO-linked glycoproteins (figure 2).4 These heavilyO-glycosylated proteins carry clusters of GalNAc-based glycans in repetitive Ser- and Thr-rich VNTR(variable number of tandem repeats) domains.These complex oligosaccharide structures have beenarbitrarily assigned three main domains: the coreregion, which comprises the innermost two orthree sugars of the glycan chain proximal to thepeptide; the backbone region, which mainlycontributes to the length of the chains formed byuniform elongation; and the peripheral or terminalregion, which exhibits a high degree of structuralcomplexity (figure 2).4 Mucins are the most prom-inent host molecules involved in hostepathogen
interactions and are discussed in more detail in thefollowing sections.Proteoglycans are a subclass of glycoproteins, but
the distinction between the two is not very clear, asboth carry a protein backbone with glycansattached at specific amino acids. In the case ofproteoglycans, several acidic disaccharide-repeatingunits are attached to the protein backbone. Thesedisaccharide units (called glycosaminoglycans) arecomposed of a hexosamine and a hexose ora hexuronic acid (glucouronic acid or iduronic acid).Proteoglycans are integral components of theextracellular matrix; examples include chondroitinsulfate, dermatan sulfate and heparan sulfate.5
A glycolipid (also referred to as glyco-sphingolipid) is a glycoconjugate in which glycanchains are attached to the lipid ceramide withglucose (Glc) or Gal as the first sugar, therebyforming glucosylceramide and galactosylceramide.Glycolipids are ubiquitous components ofmembranes, and their distribution and structuresvary depending on the location in the organism.6
During the process of colonisation and infection,the glycosylation state of both host and pathogenchange in response to the presence of the other.
Figure 2 Mucin structure and glycosylation. (A) Schematic representation of structures of secreted and cell-membrane-associated mucins showing different domains. (B) Cartoon showing distribution and structure of gel-formingsecreted, non-gel-forming secreted, and membrane-bound mucins. (C) Schematic representation of the complexglycosylated structure and multimeric assembly of a gel-forming secreted mucin. Immediately after its synthesis andtranslocation into the endoplasmic reticulum, the mucin polypeptide chain is N-glycosylated and forms disulfide-bondeddimers through its C-terminal Cys-rich domains. The dimers are then transported to the Golgi complex, and the tandemrepeat domains are O-glycosylated. Once the glycosylated dimers reach the trans-Golgi compartments, they areassembled into disulfide-bonded multimers through their N-terminal D-domains. The Y-shaped structures on theN-terminal D-domains and the C-terminal Cys-rich domains represent N-linked oligosaccharides. The squiggly linesperpendicular to the polypeptide chain represent O-linked oligosaccharides in the tandem repeat domains.
1414 Gut 2011;60:1412e1425. doi:10.1136/gut.2010.212704
Recent advances in basic science
on May 21, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.2010.212704 on 12 January 2011. D
ownloaded from

This complex modulation of glycan expressioncritically determines pathogenesis and the hostresponse in terms of structural changes andimmune response. The aim of this paper is toreview the roles of glycosylation of the gastroin-testinal (GI) tract in colonisation and invasion ofthis mucosa by bacteria. In particular, the gastro-duodenal pathogen, Helicobacter pylori, and theenteric pathogen, Campylobacter jejuni, will be usedas examples of acute and chronic bacterial infec-tions, respectively, to illustrate important conceptsconcerning the role of GI glycosylation inpathogenesis.
MUCUS LAYER AND EPITHELIAL GLYCOCALYX ASBARRIERS TO BACTERIAL INFECTIONMucus layerThe mucosal surface of the GI tract, which isa complex ecosystem of resident microbiota,immune cells and an epithelium covered bya mucus layer, is the largest body surface(200e300 m2) in contact with the external envi-ronment.7 The adherent gastrointestinal mucus gelin vivo is continuous and can be divided into twolayers: a loosely adherent layer and a layer firmlyattached to the mucosa (figure 3A). It ranges inthickness from 300 mm in the stomach and 700 mmin the intestine,8 and represents the front-linedefence barrier between the external environmentand tissues of the host.9 The relative thickness of thetwo component layers of the mucus gel varies fordifferent regions of the gut. Thus, in the stomach,the thickness of the firmly adherent mucus compo-nent is 80 and 154 mm for the corpus and antrum,respectively, with an overlying layer of looselyadherent mucus of similar thickness (figure 3A). Thegastric pathogen H pylori has been shown to reducethe rate of renewal of the mucus layer, as well ascause an acid-induced increase in its renewal rate.The mucus gel is the thickest in the colon, andthe rate of mucus accumulation is highest. Inthe small intestine, the mucus layer is thin anddiscontinuous.8 9 This viscous, supramucosal gelpresents a highly hydrated, mechanical and lubrica-tive barrier, as well as a matrix for immune-relatedand antimicrobial proteins. It is not static, since theoutermost loosely adherent mucus layer is continu-ally removed by movement of the luminal contentsof the GI tract and renewed. The rate of renewal ofthe mucus layer varies in different parts of the GItract (figure 3A).8 9 Thus, the mucus layer protectsthe underlying epithelial cells against chemical,enzymatic, microbial and mechanical insult, andserves as a physical gel to inhibit and entrapinvading microbes and aid clearance. In addition, inthe stomach, mucus acts as an unstirred layer inwhich bicarbonate ions secreted by the surfaceepithelium counteract protons diffusing from thelumen into the gel, causing the pH in the mucuslayer to range from acidic in the lumen to neutral atthe cell surface which, in turn, can influence bacte-rial colonisation (figure 3B).10 For example, H pyloriusually resides within the one-fourth zone
(0e25 mm) of mucin gel above the epithelium inclose proximity to the cells where the pH rangesfrom 5 to 7 (figure 3B).10
The protective physicochemical properties ofmucus are substantially attributable to the highcarbohydrate content of mucins, which also pres-ents considerable potential for interaction betweenmicrobial lectins and O-linked glycans. In addition,glycosidases and other enzyme activities secreted orinduced by microbes have the capacity to directlymodify these glycans after the mucins have beensecreted or exported to the cell membrane.11 12 Thisstrategy is significant as a mechanism for gener-ating binding ligands and in the provision of sugarsfor bacterial metabolism.13 Bacterial interactionwith the supramucosal gel can lead to chroniccolonisation of the mucus. On one hand, themucosal microbiota blocks access to the underlyingcells, and on the other, in the case of pathogens,necessitates the engagement of gel-penetrationstrategies before cellular adhesion.9 It is thereforesurprising that the relevance of this layer to colo-nisation and invasion of the mucosal epitheliumhas not historically been of higher priority.
GlycocalyxUnderneath the mucus layer, the second line ofdefence is constituted by the epithelial glycocalyx,which is partially integrated with the overlying gel(figure 3A).14 The glycocalyx represents an array ofhighly diverse glycoproteins and glycolipidsexpressed on the membrane of epithelial cells. Theglycocalyx is highly variable from tissue to tissue aswell as in thickness (intestinal microvilli tips,100e500 nm; lateral microvilli surface, 30e60 nm)(figure 3A).8 The glycosylation machinery ofmucosal epithelia not only varies with the cell typebut is also influenced by the sub- and supra-mucosal environmentdfor example, hormonalstatus, inflammation and microbial colonisationhave all been shown to influence the glycosylationof epithelial glycoproteins.15e18 Like the mucus gel,the glycocalyx is constantly renewed; the averageturnover time of the human jejunal glycocalyx is6e12 h. In the gut lumen, the glycocalyx interactswith the overlying mucus layer, the gastric-biliaryjuices and the resident probiotic microflora to limitcolonisation by pathogens. The complex glycosy-lated structures expressed within the glycocalyxserve as receptors and form the primary site forbacterial adhesion. For example, the carcinoem-bryonic antigen-related cell adhesion molecule6 (CEACAM6) expressed on the apical surface ofileal epithelial cells serves as a receptor for adherent-invasive Escherichia coli in Crohn disease.19 Expres-sion of such glycosylated ligands can in turn bemodulated by bacterial infection and inflammation,further promoting colonisation.
Bacterial colonisation and adherenceTo colonise mucosal surfaces and invade theepithelium, microbes commonly interact withglycan structures of the host glycocalyx.11 16
Bacterial fimbriae (or pili), various outer membrane
Gut 2011;60:1412e1425. doi:10.1136/gut.2010.212704 1415
Recent advances in basic science
on May 21, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.2010.212704 on 12 January 2011. D
ownloaded from

proteins and cell wall componentsdfor example,lipopolysaccharides (LPSs)dmay all function asadhesins. Adhesion to host cells is considered to bemediated by hydrophobic interactions, cation-bridging whereby divalent cations counteract therepulsion of the negatively charged surfaces ofbacteria and host, and receptor-ligand binding.7 11
One of the most extensively studied mechanisms ofadhesion is the binding of bacterial lectins to thecorresponding glycosylated receptors associatedwith host cells.10 Bacteria may have multipleadhesins with different carbohydrate specificities,and modulation of surface receptor density, kineticparameters, or topographical distributions of thesereceptors on cell membranes regulate adhesion.After H pylori enters the stomach, infection occursin three crucial steps: establishing colonisation;evading host immunity; and invading gastricmucosa. Urease activity and motility mediated bythe flagella of H pylori are important in penetratingthe mucus gel and harbouring colonies beneath thegastric mucus in niches adjacent to the epithelium.
Several putative adhesins attach the organism tothe gastric epithelium and prompt the succeedingprocesses for evading host immunity and invadingthe mucosa.20 21 H pylori expresses at least sixlectin-like adhesins for recognition of the gastricmucosa, of which the blood group-binding adhesin(BabA), which binds to Lewis b (Leb) and H-1antigens,20 and sialic acid-binding adhesin (SabA),which binds sialyl-Lex in inflamed tissue,21 are mostimportant for colonisation (figure 3C). HumanLewis antigens represent terminal modifications onmucin-type O-glycans. Their structure is composedof two different backbones (figure 3D). The type 1chain contains Gal and GlcNAc composed as Gal-b(1,3)-GlcNAc, while the type 2 chain has Gal-b(1,4)-GlcNAc, which is also termed N-acetyl-lactosamine (LacNAc). Adding a-Fuc as a side chainto the backbone structure on the 1e4 location, onboth 1e2 and 1e4 locations, and on both 2e2 and2e4 locations gives rise to Lewis a (Lea), Leb andsialyl-Lea antigens, respectively (figure 3D). Basedon the backbone structure of the type 2 chain, the
Figure 3 The gastrointestinal mucus layer and its role in bacterial adhesion. (A) Diagram showing the thickness of the two mucus gel layers in vivo in thestomach and small and large intestine. The mucus gel layer is continuous throughout the gastrointestinal tract and does not follow the contours of the villi inthe intestine. The membrane-associated glycocalyx is also shown. (B,C) Glycan-mediated adhesion of Helicobacter pylori to gastric epithelial cells. Thefigure illustrates some of the characterised bacterialeglycan interactions that contribute to successful colonisation of the gastric mucosa. (D) Schematicrepresentation of the terminal ABH structures and Lewis epitopes present in O-glycans, which act as receptors for H pylori adhesins, BabA and SabA.
1416 Gut 2011;60:1412e1425. doi:10.1136/gut.2010.212704
Recent advances in basic science
on May 21, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.2010.212704 on 12 January 2011. D
ownloaded from

addition of a-Fuc as a side chain at the 1e3 locationon both 1e2 and 1e3 locations, or on both the 2e2and 2e3 locations, gives rise to the Lewis x (Lex),Lewis y (Ley) and sialyl-Lex antigens, respectively(figure 3D).22
The distribution of type 1 antigens such as Lea
and Leb is primarily on the surface of the gastricepithelium. This is the primary site of ligation forthe H pylori adhesion, BabA. BabA-positive statusand its binding to Leb have been associated withsevere gastric injury, high H pylori density, andsevere clinical outcomes.23 The fucosylated H-type1 and Leb antigens are naturally expressed on thegastric mucosa of secretor or Lewis-positive indi-viduals. The secretor status depends on an activeFUT2 enzyme, an a(1,2)-fucosyltransferase thatcatalyses the addition of terminal a(1,2)-fucoseresidues. Inactivating mutations in the secretorgene affect 20% of the human population (knownas non-secretors) and have been associated withreduced susceptibility to infections by H pylori.23 24
Nearly 95% of the BabA-encoding H pylori adapt toany of the Leb sub-types, and such strains aretermed ‘generalists’. A small subset of H pyloristrains express BabA, but cannot adhere to theA-Leb antigen. Such strains are called ‘specialist’.23
BabA can also act as a mediator for H pylori bindingto MUC5AC, even in non-secretors or thosewithout Leb. In contrast, type 2 antigens may belocated deeper in the glands, such as mucous, chief,or parietal cells. Leb is weakly expressed on thesurface epithelium during chronic inflammation, aswell as in gastric atrophy (atrophic gastritis) andintestinal metaplasia. Under these clinical condi-tions, there is an upward migration of Lex associ-ated with atrophy of the epithelium. Duringchronic inflammation or in atrophic gastritis,H pylori binds to Lex and sialyl-Lex via SabA adhesin.SabA production and SabA-mediated adhesion areassociated with severe disease outcomes, includinggastric atrophy, intestinal metaplasia and thedevelopment of gastric cancer. The minimal struc-ture required for SabA adhesin binding was shownto be NeuAca2-3Gal (figure 3C,D).25
H pylori CagL binds to the integrin, a5b1, to forman injection pile by which CagA translocates intothe cytosol of gastric epithelium, using a type IVsecretory system. Other known H pylori adhesinsinclude neutrophil-activating protein, which bindsto sulfated carbohydrate structures,26 27 a 25 kDaadhesin that binds the glycoprotein laminin in theextracellular matrix,28 29 as well as adherence-associated lipoprotein A and B (AlpA/B)10 30 andHorB,31 whose gastric receptors have not beenidentified. Moreover, H pylori lectins bind moleculesin the oral cavity, such as salivary agglutinin, thatmay influence re-infection of the gastric mucosafrom this secondary infection reservoir.32 At leastfour protein adhesins, which differ substantially inactivity and importance depending on the anatomicalsite within the orogastric route, the mucin-type, theenvironmental pH and the gastric disease status, areinvolved in H pylori binding to mucus-related oligo-saccharides.33 The adhesive interactions are not
limited to protein adhesins, as H pylori LPS interactsand binds with the gastric epithelium34 throughrecognition by the host lectin, galectin-3.35 As occurswith protein adhesins,36 environmental pH, particu-larly low pH, can influence expression of the LPSligand involved in the latter interaction.37 In addition,this bacterium recognises a range of glycosylatedmolecules, not only glycoproteins and oligosaccharidemoieties, but also proteoglycans and glycolipids.10 Incontrast, the nature and specificities of the lectinsexpressed by intestinal-colonising C jejuni appearless diverse,38 39 possibly reflecting the lessermucus and tissue diversity of the jejunal environ-ment (figure 3A). Nevertheless, although individualadhesionereceptor binding is usually of low affinity,clustering of adhesins and receptors can causemultivalency effects resulting in strong attachment.The O antigen of H pylori LPS shares structural
homology with the Lewis blood group antigensexpressed in human gastric mucosa, including bothtype 1 chains, namely Lea and H-type 1, and type 2chains, such as Lex and Ley. The expression of theseantigens undergoes phase variation, with differentbacterial fucosyltransferases being switched on andoff, contributing to a dynamic glycosylation even inthe same host. It has been proposed that Lex
structures expressed by the H pylori O antigen sidechain may further promote adhesion to gastricepithelial cells.37 40
Collectively, ligandereceptor binding can havedifferent consequences for bacterial pathogensduring colonisation of the GI tract and whether theresultant infection is acute or chronic. Adhesion canaffect bacteria by stimulation/inhibition of growth,as well as induction of other adhesive structures andproteins required for invasion, whereas the effects ofadhesion on host cells can be altered morphology,fluid loss, induction of cytokine release, upregulationof cell adhesion molecules, and apoptosis.7 In addi-tion, infection and associated inflammation caninduce changes in mucosal glycosylation,41 42
including the release of carbohydrates for bacterialutilisation. An example of this occurs in H pyloriinfection whereby human a-L-fucosidase is secretedwhich liberates L-Fuc from host cells, which becomesavailable for H pylori to synthesise mammalianLewis antigens,43 which contribute to pathogenesisof the bacterium.40 44
Of the range of GI barrier-related molecules withwhich bacterial pathogens interact, mucins asa common family of molecules present in secretedmucus and in the glycocalyx are receivingincreasing attention and will form the predominantfocus for further discussion in this review.
PROPERTIES AND CHARACTERISTICS OF MUCINSGeneral attributes of mucinsThe polymeric scaffolding of mucus is formed bysecreted mucin glycoproteins. However, it is impor-tant to note that certain mucins (eg, MUC1) can bemembrane-bound and hence form a major constit-uent of the epithelial glycocalyx of a variety ofmucosal tissues (figure 2B). Mucin-type moleculesconsist of a core protein moiety (apomucin) within
Gut 2011;60:1412e1425. doi:10.1136/gut.2010.212704 1417
Recent advances in basic science
on May 21, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.2010.212704 on 12 January 2011. D
ownloaded from
which a number of carbohydrate chains are attachedto Ser, Thr and Pro residues by glycosidebonds forming O- and N-linked oligosaccharides15 45
(figure 2C). Mucins have a high molecular mass(53105 to 30 3106), and more than 50% (often70e80%) of the mucin molecular mass is composedof carbohydrate.9 Each mucin carries of the order of100 different oligosaccharide structures. Thesecarbohydrate chains are often clustered into highlyglycosylated domains, giving the mucin a ‘bottle-brush’ appearance (figure 2C). In secreted mucins,these O-linked oligosaccharides are concentrated inlarge peptide domains of repeating amino acidsequences rich in Ser and Pro.15 The size and numberof repeats vary between mucin types, and in many ofthe mucin genes there is polymorphism in thenumber of repeats (VNTR) (table 1)8 9 (figure 2C).Thus, the size of such mucins can differ substantiallybetween individuals and could represent a basis fordifferences in infection susceptibility between hosts.
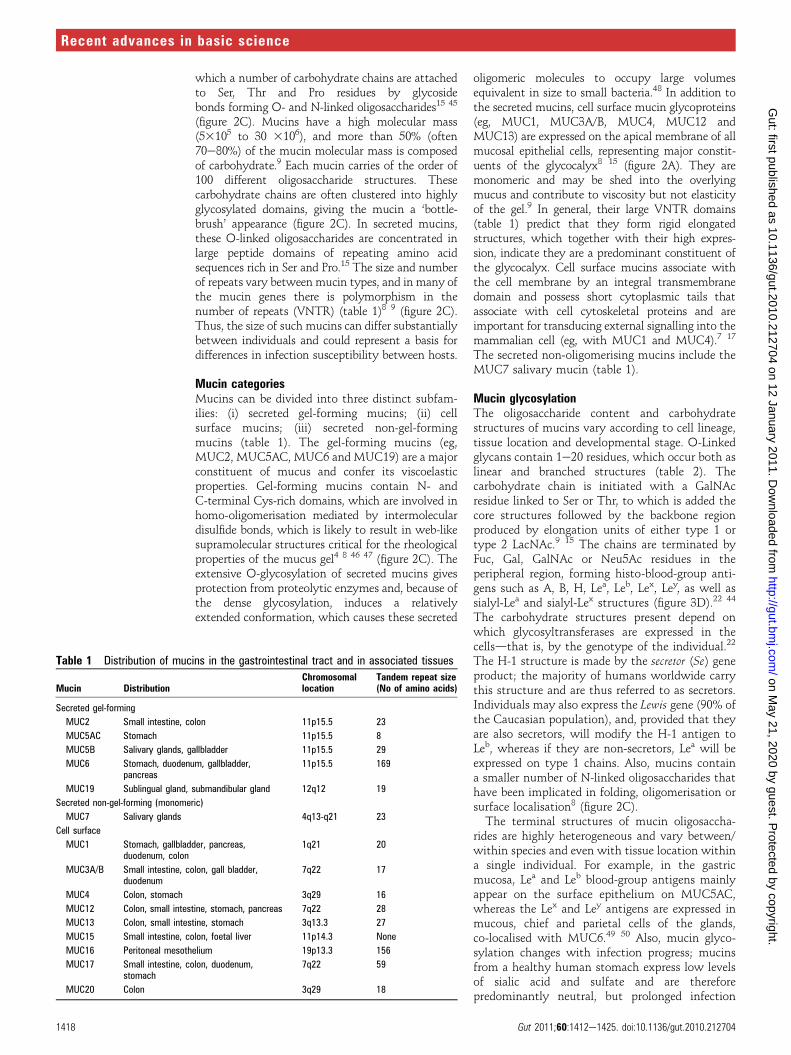
Mucin categoriesMucins can be divided into three distinct subfam-ilies: (i) secreted gel-forming mucins; (ii) cellsurface mucins; (iii) secreted non-gel-formingmucins (table 1). The gel-forming mucins (eg,MUC2, MUC5AC, MUC6 and MUC19) are a majorconstituent of mucus and confer its viscoelasticproperties. Gel-forming mucins contain N- andC-terminal Cys-rich domains, which are involved inhomo-oligomerisation mediated by intermoleculardisulfide bonds, which is likely to result in web-likesupramolecular structures critical for the rheologicalproperties of the mucus gel4 8 46 47 (figure 2C). Theextensive O-glycosylation of secreted mucins givesprotection from proteolytic enzymes and, because ofthe dense glycosylation, induces a relativelyextended conformation, which causes these secreted
oligomeric molecules to occupy large volumesequivalent in size to small bacteria.48 In addition tothe secreted mucins, cell surface mucin glycoproteins(eg, MUC1, MUC3A/B, MUC4, MUC12 andMUC13) are expressed on the apical membrane of allmucosal epithelial cells, representing major constit-uents of the glycocalyx8 15 (figure 2A). They aremonomeric and may be shed into the overlyingmucus and contribute to viscosity but not elasticityof the gel.9 In general, their large VNTR domains(table 1) predict that they form rigid elongatedstructures, which together with their high expres-sion, indicate they are a predominant constituent ofthe glycocalyx. Cell surface mucins associate withthe cell membrane by an integral transmembranedomain and possess short cytoplasmic tails thatassociate with cell cytoskeletal proteins and areimportant for transducing external signalling into themammalian cell (eg, with MUC1 and MUC4).7 17
The secreted non-oligomerising mucins include theMUC7 salivary mucin (table 1).
Mucin glycosylationThe oligosaccharide content and carbohydratestructures of mucins vary according to cell lineage,tissue location and developmental stage. O-Linkedglycans contain 1e20 residues, which occur both aslinear and branched structures (table 2). Thecarbohydrate chain is initiated with a GalNAcresidue linked to Ser or Thr, to which is added thecore structures followed by the backbone regionproduced by elongation units of either type 1 ortype 2 LacNAc.9 15 The chains are terminated byFuc, Gal, GalNAc or Neu5Ac residues in theperipheral region, forming histo-blood-group anti-gens such as A, B, H, Lea, Leb, Lex, Ley, as well assialyl-Lea and sialyl-Lex structures (figure 3D).22 44
The carbohydrate structures present depend onwhich glycosyltransferases are expressed in thecellsdthat is, by the genotype of the individual.22
The H-1 structure is made by the secretor (Se) geneproduct; the majority of humans worldwide carrythis structure and are thus referred to as secretors.Individuals may also express the Lewis gene (90% ofthe Caucasian population), and, provided that theyare also secretors, will modify the H-1 antigen toLeb, whereas if they are non-secretors, Lea will beexpressed on type 1 chains. Also, mucins containa smaller number of N-linked oligosaccharides thathave been implicated in folding, oligomerisation orsurface localisation8 (figure 2C).The terminal structures of mucin oligosaccha-
rides are highly heterogeneous and vary between/within species and even with tissue location withina single individual. For example, in the gastricmucosa, Lea and Leb blood-group antigens mainlyappear on the surface epithelium on MUC5AC,whereas the Lex and Ley antigens are expressed inmucous, chief and parietal cells of the glands,co-localised with MUC6.49 50 Also, mucin glyco-sylation changes with infection progress; mucinsfrom a healthy human stomach express low levelsof sialic acid and sulfate and are thereforepredominantly neutral, but prolonged infection
Table 1 Distribution of mucins in the gastrointestinal tract and in associated tissues
Mucin DistributionChromosomallocation
Tandem repeat size(No of amino acids)
Secreted gel-forming
MUC2 Small intestine, colon 11p15.5 23
MUC5AC Stomach 11p15.5 8
MUC5B Salivary glands, gallbladder 11p15.5 29
MUC6 Stomach, duodenum, gallbladder,pancreas
11p15.5 169
MUC19 Sublingual gland, submandibular gland 12q12 19
Secreted non-gel-forming (monomeric)
MUC7 Salivary glands 4q13-q21 23
Cell surface
MUC1 Stomach, gallbladder, pancreas,duodenum, colon
1q21 20
MUC3A/B Small intestine, colon, gall bladder,duodenum
7q22 17
MUC4 Colon, stomach 3q29 16
MUC12 Colon, small intestine, stomach, pancreas 7q22 28
MUC13 Colon, small intestine, stomach 3q13.3 27
MUC15 Small intestine, colon, foetal liver 11p14.3 None
MUC16 Peritoneal mesothelium 19p13.3 156
MUC17 Small intestine, colon, duodenum,stomach
7q22 59
MUC20 Colon 3q29 18
1418 Gut 2011;60:1412e1425. doi:10.1136/gut.2010.212704
Recent advances in basic science
on May 21, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.2010.212704 on 12 January 2011. D
ownloaded from

Table 2 Common O-linked oligosaccharide structures on mucins
Gut 2011;60:1412e1425. doi:10.1136/gut.2010.212704 1419
Recent advances in basic science
on May 21, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.2010.212704 on 12 January 2011. D
ownloaded from

with H pylori decreases fucosylation and increasessialylation of mucins.21 51 However, the sialylationpattern returns to normal after successful H pylorieradication therapy. The structural diversity ofglycosylation has been suggested to allow the hostto cope with diverse and changing pathogen chal-lenges.11 Notably, host susceptibility to a specificbacterial pathogen and subsequent disease severitycan vary between individuals expressing differenthisto-blood groups. This has been demonstrated forH pylori infection in rhesus monkeys, which sharesstrong similarities to the natural history of H pyloriinfection in humans.24 This bacterium has anability to vary and adapt its surface characteristics,including expression of glycosylation and outermembrane proteins to aid infection in humans andin animal models.35 52e55 A strong correlation hasbeen found between the adhesive properties ofH pylori endemic in specific human populations andthe mucin-related blood group structures expressedby those populations,51 as well as the ability of thebacterial adhesins to vary to aid colonisation.10 23 56
Changes in the expression levels and glycosylationpatterns of mucins have been associated with severaldiseases, including carcinomas. In gastric cancer,alterations in mucin polypeptide expression havebeen reported: loss of expression of MUC5AC,increased mucin heterogeneity, and glycosylationchanges, including exposure of simple mucin-typecarbohydrates.57 The majority of gastric carcinomasare preceded by a precancerous stage, characterisedby sequential development of atrophic gastritisleading to intestinal metaplasia, and dysplasia.Intestinal metaplasia consists of replacement of thegastric mucosa by an epithelium that histologicallyresembles the intestinal mucosa and is characterisedby the presence of absorptive cells, Paneth cells,and goblet cells secreting sialo-mucins and/orsulfo-mucins and corresponds to the small intestinephenotype.58 Altered mucin expression patternsreported in intestinal metaplasia include under-expression of MUC1, MUC5AC and MUC6 and denovo expression of MUC2.58
BACTERIAL INTERACTIONS WITH MUCINSEffects on mucins and their synthesisMucus is secreted by specialised goblet cells (alsoknown as mucin-secreting cells).59e61 Through theexpression of different mucins, variations in mucinglycosylation and the co-secretion of mucin-asso-ciated molecules, the nature of the mucus varies atdifferent anatomical locations and constitutesa responsive system that can be adapted to localphysiological requirements, including response tobacterial colonisation and infection.7e9 15 60 Themucins are produced by cells in the epithelial surfaceand/or by glands located in the submucosalconnective tissues, and secretion occurs viaboth constitutive and regulated pathways.8 15 Theconstitutive pathway continuously secretes suffi-cient mucin to maintain the mucus layer, whereasthe regulated pathway affords a massive discharge ofmucus in response to environmental and/or patho-
physiological stimuli such as bacterial infection andinflammation. Although release is regulated by acti-vation of signalling pathways by a number ofsecretagogues (including acetylcholine, vasoactiveintestinal peptide and neurotensin), activationcan occur by bacterial-induced inflammatory medi-ators (eg, interleukin (IL)-1, IL-4, IL-6, IL-9, IL-13,interferons, tumour necrosis factor a and nitricoxide).7 8 59 61 The responsiveness to cytokines andother immune regulators provides a link betweenmucin-based defence, innate mucosal immunity andmucosal inflammatory responses.8 Stimulated mucinrelease occurs rapidly and is accompanied bya hundredfold or so expansion of secretory granules.Changes in goblet cell function and in the
chemical composition of intestinal mucins havebeen detected in response to a broad range ofluminal insults, including changes in the normalmicrobiota and intrusion of enteric pathogens.41 42
Probiotic bacteria have been shown to be capable ofenhancing mucin secretion in colonic epithelialcells,62 thus potentially boosting this defensivemechanism against pathogens. There is evidencethat a variety of microbial products can stimulateincreased production of mucins by mucosal epithe-lial cells.61e63 Also, germ-free mice exhibit changesin mucin gene expression, mucus composition andmucus secretion in response to intestinal bacteria orhost-derived inflammatory mediators.64 65 There-fore, it can be surmised that alterations in mucussecretion and composition, including mucin glyco-sylation, represent important mechanisms forunfavourably changing the niche occupied bya mucosal pathogen. Successful pathogens, however,are capable of countering this mechanism. Forexample, H pylori has been shown to decreasemucin exocytosis,66 and LPS of this bacterium hasbeen shown to affect mucin synthesis67 and themacromolecular nature of the mucin secreted,68
thereby representing a potential strategy bywhich the bacterium can favourably modulatemucin composition of the mucus barrier and theglycocalyx.Specifically, H pylori causes an aberrant expres-
sion of human MUC1, MUC5AC and MUC6 ininfected gastric mucosa,69 70 in particular withdownregulation of MUC5AC and upregulation ofMUC6,71 both of which are secreted mucins. Asimilar downregulation of MUC5AC has beenfound in rhesus monkeys, which have strongsimilarities to mucin glycosylation in humans.72
The reduced MUC5AC synthesis induced byH pylori can impair the gastric mucus gel,9 but theupregulation of MUC6 may represent an antimi-crobial host response, since the O-linked glycans ofMUC6 display antimicrobial activity againstH pylori.73 The basis for this is the inhibition ofbiosynthesis of the essential bacterial membranelipid, cholesterol-a-D-glucopyranoside, by theterminal a(1/4)-linked GlcNAc of MUC6 inter-acting with the respective enzyme.74 A closeassociation with secreted MUC5AC andmembrane-bound MUC1 is considered importantfor H pylori colonisation of the gastric mucosa.75 76
1420 Gut 2011;60:1412e1425. doi:10.1136/gut.2010.212704
Recent advances in basic science
on May 21, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.2010.212704 on 12 January 2011. D
ownloaded from

H pylori is bound by secreted MUC5AC through thebacterial outer membrane protein, BabA, by recog-nising fucosylated Leb expressed in this mucin.49
Although, on the one hand, aiding H pylori coloni-sation of the mucus layer, this binding may, on theother hand, limit colonisation to the mucus gel, andhence reduce the number of H pylori bacteriainteracting with the epithelial surface by blockingBabA recognition of structures in the epithelialglycocalyx. Nevertheless, H pylori is capable ofbinding cell surface MUC1,75 77 and thus bacterialcontact with the epithelial surface can be mediatedin a mucin-dependent manner.
Other bacterial effects on mucinsIn addition to direct interactions with mucins andtheir synthesis, bacteria can use other strategies toovercome the gel-based barrier function of themucus layer. Flagellation is a shared characteristicof the vast majority of successful mucosal patho-gens that are highly motile, such as C jejuni andH pylori.78 Motility, along with the corkscrewmorphology of these bacteria, allows them toburrow through the mucus layer to reach theepithelium.79 Indeed, H pylori that have dysfunc-tional flagella have a greatly reduced ability toinfect.80 Combined with motility, degradativeenzymes, such as fucosidases, mucinases (mucin-degrading enzymes), glycosylsulfatases, sialidases,sialate O-acetylesterases and N-acetylneuraminatelyases, are produced by a broad spectrum ofbacterial pathogens that destabilise the mucus aswell as removing mucin decoy carbohydrates forbacterial adhesins.9 11 12 81 The size of the mucuspolymer is important for the formation of entan-gled gels and the viscous properties of mucus,8 15
and thus degradation of mucins will result inlocalised disintegration of mucus, thereby aidingbacterial penetration of the epithelium.Supporting the view of the mucus gel as an
effective infection barrier, avoidance of this layerhas been suggested as a further strategy used bycertain mucosal pathogens of the GI tract.8
Although M cells are only a very small percentageof mucosal epithelial cells, they are the major pointof attachment and/or entry used by a largeproportion of enteric bacteria, including C jejuni.82
Intestinal M cells, which capture and presentmicrobial antigens to the underlying lymphoidtissue, do not produce gel-forming mucins and havea thin glycocalyx.83 M cells selectively express thesialyl-Lea antigen, whereas Lea is expressed on allepithelial cells of the GI tract.84 85 Thus, M cellswith a different profile of glycocalyx glycopro-teins84 85 represent a focal point of entry formucosal pathogens. Once M cells are invaded ormucus is penetrated, a further strategy of GI tractpathogens is to disrupt the tight junctions betweenadjacent mucosal cells, thereby exposing thevulnerable lateral membranes not protected by theglycocalyx. Such a mechanism is used by H pylori,which affects the permeability of cellular tightjunctions,86 penetrates intercellular junctions, and,mediated by a bacterial lectin and LPS, binds to the
glycoprotein laminin in the extracellular matrix andbasement membrane.28 87 The latter interaction hasfurther significance since H pylori LPS inhibitsrecognition of laminin by an epithelial cell receptor(integrin), which has been hypothesised tocontribute to lack of mucosal integrity.88
Cell surface mucins in protectionAlthough the cell surface mucins can act as recog-nition molecules for bacterial adhesins, thesemucins may also show other protective effectsagainst pathogenic bacteria. For example, MUC1has been shown to protect p53-expressing epithelialcells in vitro from the effects of C jejuni cytolethaldistending toxin, a genotoxin, and, as demonstratedin Muc1�/� mice, lowers gastric colonisation byC jejuni.89 It should be noted that glycocalyxmucins are capable of initiating intracellularsignalling in response to bacteria, reflecting theirreporter function, as well as their barrier role asconstituents of the glycocalyx on the apical surfaceof mucosal epithelial cells.7 17 Importantly, thecytoplasmic domain of MUC1 can be cleaved,translocates to mitochondria, and, together withthe p53 transcription factor, modulates the cellcycle and protects against the apoptotic response togenotoxic stress.90 91 This protective effect, firstidentified in cancer research, has been suggested tohave evolved as part of the natural epithelialdefence against microbial genotoxins.8
Another protective mechanism of cell surfacemucins that has been hypothesised, again exem-plified by MUC1, involves their role as releasabledecoy ligands for mucosal pathogens attempting toanchor to the glycocalyx. Supportive evidence forsuch a role of MUC1 has been accumulated forboth C jejuni89 and H pylori.77 Although H pylori canbind MUC1, primary murine gastric epithelial cellsexpressing MUC1 bind fewer H pylori than murineMuc�/� cells.77 These, at first apparentlyconflicting, results can be explained if MUC1 actsas a releasable decoy; the bacteria bind MUC1 onnormal gastric epithelial cells, but this is then shedby the host92; however, owing to the absence ofMUC1 on Muc1�/� murine cells, the bacteriaattach to other epithelial receptors and hence arenot lost. The findings are reflected in pathology,since Muc1�/� mice develop fivefold greater H pyloricolonisation density, which is maintained for atleast 2 months, whereas heterozygous mice havea lower level of gastric MUC1 expression andintermediate colonisation densities.77 Moreover,comparing H pylori-positive patients, those withMUC1 polymorphisms exhibiting short extracel-lular mucin domains are more likely to developgastritis than those without,75 because the shorterMUC1 molecules will allow greater access of thebacterium to the cell surface. Likewise, infectionexperiments in Muc1�/� mice have shown greatersusceptibility to infection, GI barrier transit, andpathology by C jejuni than in normal mice.89
Collectively, the results are consistent with theimportance of MUC1 in shielding the epithelialsurface from pathogen contact.
Gut 2011;60:1412e1425. doi:10.1136/gut.2010.212704 1421
Recent advances in basic science
on May 21, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.2010.212704 on 12 January 2011. D
ownloaded from

Mucin-associated molecules in bacterial infectionIn addition to mucins, other mucosa-associatedmolecules that involve carbohydrateeligand inter-actions can play an important role in determiningthe success of bacterial colonisation of epithelialsurfaces.15 An example of such molecules is trefoilfactors.9 These factors, comprising TFF1e3, area group of small proteins involved in wound repairand protection of the integrity of the epithelialbarrier; they are secreted by mucus-secreting cellsand expressed in a site-specific manner in the GItract.93 TFF1 segregates with MUC5AC in thegastric foveolar epithelium, TFF2 with MUC6 inthe glandular epithelium in the distal stomach,whereas TFF3 and MUC2 are co-expressed by thegoblet cells of the large and small intestine. The roleof these factors in infection is exemplified in H pyloricolonisation of the gastric mucosa. H pylori has beenshown to bind avidly to dimeric TFF1, and thisinteraction enables binding to gastric mucin,suggesting that TFF1 acts as a linker moleculebinding to both H pylori and MUC5AC.94 95
Although the H pylorieTFF1 interaction appearswell-founded, recognition by TFF1 has been ascribedto Glc- and Man-related structures within H pyloriLPS by in vitro testing.89 However, this requiresfurther examination because the sugar Man isabsent from this LPS,40 44 but is present in bacterialcell surface mannans which co-purify with LPS.96
Another group of molecules, the surfactantproteins, have relevance to antibody-independentpathogen recognition and clearance from the GItract. Surfactant protein D (SP-D), a member of thecollectin family, is a C-type lectin that is found inmucus-secreting cells within gastric pits and withmaximal expression at the luminal surface.97 98
Levels of expression of SP-D are significantlyincreased in H pylori-associated antral gastritiscompared with normal human gastric mucosa, andco-localises with infecting H pylori bacteria.96
SP-D�/� mice are more susceptible to Helicobactercolonisation, and exhibit decreased inflammationand immune response to infection, emphasising the
important influence on the establishment of infec-tion.99 Functioning as a lectin, SP-D binds H pyloriby recognition of the carbohydrate-bearing O-chainof LPS, resulting in bacterial aggregation andimmobilisation,97 100 thus aiding eventual phago-cytosis and clearance. Nevertheless, to evade thisimportant mechanism of innate immune recogni-tion, escape variants within the infecting H pyloripopulation can arise98 100 with modifications intheir O-chain glycosylation, thus decreasing bacte-rial interaction with SP-D and aiding colonisa-tion.100 This represents an important example ofchanges in bacterial glycosylation to overcomelectin-based microbial recognition in the GI mucosa.
CONCLUSIONSGI glycosylation in health and diseaseThe GI tract and its mucosa is a complex ecosystemof resident microbiota, resident and recruitedimmune cells, and a responsive mucus layer andepithelium. In addition to the barrier function ofthe mucus gel and glycocalyx against pathogens,protection is augmented by the production ofantimicrobial peptides, including defensins, by thehost.7 101 For example, expression of the humancathelicidin LL-37102 and human b-defensin 2103 isupregulated in the gastric mucosa of H pylori-infected patients.Not to be overlooked, the microbiota may influ-
ence host antimicrobial peptide production.Angiogenin-4, a molecule with bactericidal activityproduced by Paneth cells, is induced by Bifidobacte-rium thetaiotaomicron, a dominant member of thegut microbiota.104 Also, Bifidobacterium cell wallproteins have been shown to induce b-defensin 2production from human intestinal epithelial cells.7
Moreover, Gram-negative bacterial components,such as LPS, peptidoglycan105 and protein derivedfrom flagella (FliC), can induce expression ofhuman b-defensin 2,106 and hence is likely to beinduced by resident intestinal colonisers. Althoughmembers of the resident microbiota have beendescribed as inducing inhibition of other bacterialspecies, including pathogens, by the production ofthe antimicrobial compounds microcins and coli-cins,7 107 invading pathogens can also produce suchcompounds to shape the GI tract flora. H pyloriproduces a ceropin-like peptide, Hp(2-20),108 towhich it is resistant, compared with other Gram-negative bacteria, because of modification ofa membrane lipoglycan moiety.109
In general, the gut microbiota is no longerconsidered inert and stagnant but evolving, andinfluences the homoeostasis of the host,7 110 111 forexample, by influencing the host immunitythrough the T regulatory cell response.110 111 Thecapsular polysaccharide A of the intestinalsymbiont Bacteroides fragilis induces protection fromexperimental colitis, produced by Helicobacter hepa-ticus, through induction of tolerogenic T cells.112
Also, members of the microbiota can limit anti-inflammatory responses and colonisation,113 andestablish a balanced relationship with the host.110
Overall, it has been deduced that commensal
Key points
< All cells, including those of the gastrointestinal tract, are covered ina glycocalyx composed of glycolipids, glycoproteins and proteoglycans.
< Mucins are a major class of glycoproteins that are both cell-membrane-boundand secreted to form a gel.
< Mucins are key molecules in acute and chronic infections of the gut, and theirglycosylation changes in response to microbial pathogens.
< The supramucosal gel is a physicochemical sensor responsive to theenvironment that acts as a highly hydrated, mechanical and lubricative barrierand a matrix for immune-related and antimicrobial proteins.
< Bacterial surface proteins, adhesins, bind to carbohydrate moieties on hostcells and affect the adhesion and colonisation process.
< Glycanelectin interactions are emerging as key elements in gut pathogenesis.< The glycocalyx and carbohydrate components of the mucus gel are
responsive to pathogens, which can promote or hinder adhesion andinfection.
1422 Gut 2011;60:1412e1425. doi:10.1136/gut.2010.212704
Recent advances in basic science
on May 21, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.2010.212704 on 12 January 2011. D
ownloaded from

bacteria have evolved specialised molecules, such aspolysaccharide A of B fragilis, to induce tolerogenicresponses that coordinate tolerance of the micobiotain the gut, in contrast with pathogen colonisation.111
This represents another level of communicationwith the host that can be utilised by the microbiota.By modifying GI glycosation, the gut microbiota
can further influence this barrier against pathogens.Seminal work showed that, in germ-free rodents,the mucus layer is reduced by half and is lessstable,114 and has an altered glycosylation profilecompared with conventional hosts.18 41 The use ofcDNA microarray analysis has shown the partici-pation of a well-established intestinal microbiota inregulation of the gut mucus layer and composi-tion.65 It has been suggested that the residentmicrobiota can affect goblet cell dynamics andmucin composition directly, by the release ofbioactive factors, and indirectly by activating hostcells.7 41 For example, Gram-negative LPS increasesthe expression of mRNA of MUC5AC and MUC5Band stimulates their secretion.61 A bacterialquorum-sensing signal molecule of Gram-negativebacteria may stimulate production of MUC5AC. Astrain of Lactobacillus plantarum, a Gram-positivesubdominant species of the microbiota, increasesexpression of MUC2 and MUC3 mRNA,115 whichhas been shown to inhibit the cell attachment ofenteropathogenic E coli (EPEC) strains.116 More-over, a cocktail of probiotic bacteria (Lactobacillusand Bifidobacterium spp.) has been shown to atten-uate C jejuni association with, and translocationthrough, mucus-secreting intestinal cells in vitro.117
Therefore, in considering glycosylation ascontributing to the structures and barrier functionsof the GI tract against pathogens, the influence ofthe resident microbiota promises to be a subject ofdeveloping interest and potential.
Glycosylation and glycosylated receptors as noveltherapeutic targetsBiomedical applications of glycobiology havefocused greatly on the roles of carbohydrates asreceptors for adhesion of pathogens and toxins tohost cells, appearance as tumour-associatedmarkers, the regulation of inflammation, and thecirculatory half-life of plasma glycoproteins andglycosylated therapeutic proteins. Manyapproaches have been adopted to modulate biolog-ical systems involving carbohydrates and lectininteractions. The most common involve theutilisation of glycans or glycomimetics to regulatethe activity of lectins. The other approaches includemodulation of genes and enzymes involved inglycosylation pathways and the use of glycotopesas vaccination targets.118 The success of sialic acidanalogues (zanamivir and oseltamivirdRelenza andTamiflu, respectively) as inhibitors of influenzavirus sialidase shows promise for the use of glycananalogues against pathogens.119
The increasing antibiotic resistance associatedwith H pylori eradication by antimicrobial therapyis raising the need to search for alternative strate-gies. As H pylori uses the host glycan receptors for
adhesion and infection, a strategy of carbohydrate-dependent inhibition of H pylori colonisation usingcarbohydrate-binding mimics (bio-mimics) willhave a therapeutic and/or prophylactic effect onH pylori infection. Although H pylori-inducedchanges in host glycosylation have been implicatedin the progression of gastric disease, the molecularmechanisms by which H pylori modulates host cellglycosylation is not known. In addition, thepotential of using synthetic carbohydrates toinhibit H pylori adhesion to glycan receptors ongastric epithelial cells needs to be explored. Char-acterisation of the glycosylation genes and glycanreceptors essential for H pylori adhesion will allowidentification of individuals at highest risk ofdeveloping H pylori infection-associated diseases, aswell as provide the proof of concept towards thedesign of new infection eradication strategies basedon inhibition of bacterial adhesion to gastricepithelial cells.
Acknowledgements It is with deep sadness that weacknowledge that Professor Anthony Moran passed away on 30September 2010. We dedicate this review to his memory.
Funding The authors were supported by Science Foundation Irelandgrant No 08/SRC/B1393.
Competing interests None.
Provenance and peer review Commissioned; externally peerreviewed.
REFERENCES1. Mann M, Jensen ON. Proteomic analysis of post-translational
modifications. Nat Biotechnol 2003;21:255e61.2. Varki A, Lowe JB. Biological Roles of Glycans. In: Varki A,
Cummings RD, Esko JD, et al, eds. Essentials of Glycobiology. 2ndedn. Cold Spring Harbor, New York: Cold Spring HarborLaboratory Press, 2009:75e88.
3. Walsh G, Jefferis R. Post-translational modifications in thecontext of therapeutic proteins. Nat Biotechnol2006;24:1241e52.
4. Perez-Vilar J, Hill RL. The structure and assembly of secretedmucins. J Biol Chem 1999;274:31751e4.
5. Bartlett AH, Park PW. Proteoglycans in host-pathogeninteractions: molecular mechanisms and therapeutic implications.Expert Rev Mol Med 2010;12:e5.
6. Hakomori S. Structure, organization, and function ofglycosphingolipids in membrane. Curr Opin Hematol2003;10:16e24.
7. Lievin-Le Moal V, Servin AL. The front line of enteric hostdefense against unwelcome intrusion of harmful microorganisms:mucins, antimicrobial peptides, and microbiota. Clin MicrobiolRev 2006;19:315e37.
8. Linden SK, Sutton P, Karlsson NG, et al. Mucins in the mucosalbarrier to infection. Mucosal Immunol 2008;1:183e97.
9. Carrington SD, Clyne M, Reid CJ, et al. Microbial interactionswith mucus and mucins. In: Moran AP, Holst O, Brennan PJ, vonItzstein M, eds. Microbial Glycobiology: Structures, Relevanceand Applications. London, UK: Academic Press, 2009:655e71.
10. Linden SK, Arnqvist A, Teneberg S, et al. H. pylori adhesion tothe gastric surface. In: Sutton P, Mitchell H, eds. HelicobacterPylori in the 21st Century. Wallingford, UK: CABI Publishers,2010:248e68.
11. Hooper LV, Gordon JI. Glycans as legislators of host-microbialinteractions: spanning the spectrum from symbiosis topathogenicity. Glycobiology 2001;11:1Re10R.
12. Sonnenburg JL, Xu J, Leip DD, et al. Glycan foraging in vivo byan intestine-adapted bacterial symbiont. Science2005;307:1955e9.
13. Sonnenburg JL, Angenent LT, Gordon JI. Getting a grip onthings: how do communities of bacterial symbionts becomeestablished in our intestine? Nat Immunol 2004;5:569e73.
14. Hattrup CL, Gendler SJ. Structure and function of the cellsurface (tethered) mucins. Annu Rev Physiol 2008;70:431e57.
Gut 2011;60:1412e1425. doi:10.1136/gut.2010.212704 1423
Recent advances in basic science
on May 21, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.2010.212704 on 12 January 2011. D
ownloaded from

15. Corfield AP, Carroll D, Myerscough N, et al. Mucins in thegastrointestinal tract in health and disease. Front Biosci 2001;6:D1321e57.
16. Corfield AP, Wiggins R, Edwards C, et al. A sweet coating ehow bacteria deal with sugars. Adv Exp Med Biol2003;535:3e15.
17. Singh PK, Hollingsworth MA. Cell surface-associated mucins insignal transduction. Trends Cell Biol 2006;16:467e76.
18. Sharma R, Schumacher U. Carbohydrate expression in theintestinal mucosa. Adv Anat Embryol Cell Biol 2001;160:1e91.
19. Barnich N, Carvalho FA, Glasser AL, et al. CEACAM6 actsas a receptor for adherent-invasive E. coli, supporting ilealmucosa colonization in Crohn disease. J Clin Invest2007;117:1566e74.
20. Ilver D, Arnqvist A, Ogren J, et al. Helicobacter pylori adhesinbinding fucosylated histo- blood group antigens revealed byretagging. Science 1998;279:373e7.
21. Mahdavi J, Sonden B, Hurtig M, et al. Helicobacter pyloriadhesin in persistent infection and chronic inflammation. Science2002;297:573e8.
22. Green C. The ABO, Lewis and related blood group antigens;a review of structure and biosynthesis. FEMS Microbiol Immunol1989;1:321e30.
23. Aspholm-Hurtig M, Dailide G, Lahmann M, et al. Functionaladaptation of BabA, the H. pylori ABO blood group antigenbinding adhesin. Science 2004;305:519e22.
24. Linden S, Mahdavi J, Semino-Mora C, et al. Role of ABOsecretor status in mucosal innate immunity and H. pyloriinfection. PLoS Pathogens 2008;4:e2.
25. Yamaoka Y. Roles of Helicobacter pylori BabA in gastroduodenalpathogenesis. World J Gastroenterol 2008;14:4265e72.
26. Teneberg S, Miller-Podraza H, Lampert HC, et al. Carbohydratebinding specificity of the neutrophil-activating protein ofHelicobacter pylori. J Biol Chem 1997;272:19067e71.
27. Namavar F, Sparrius M, Veerman EC, et al. Neutrophil-activatingprotein mediates adhesion of Helicobacter pylori to sulfatedcarbohydrates on high-molecular-weight salivary mucin. InfectImmun 1998;66:444e7.
28. Valkonen KH, Wadstrom T, Moran AP. Identification of theN-acetylneuraminyllactose- specific laminin-binding protein ofHelicobacter pylori. Infect Immun 1997;65:916e23.
29. Moran AP, Broaders SA, Rapa A, et al. In vivo expression of the25-kDa laminin- binding protein of Helicobacter pylori. FEMSImmunol Med Microbiol 2005;43:331e7.
30. Odenbreit S, Till M, Hofreuter D, et al. Genetic and functionalcharacterization of the alpAB gene locus essential for theadhesion of Helicobacter pylori to human gastric tissue.Mol Microbiol 1999;31:1537e48.
31. Snelling WJ, Moran AP, Ryan KA, et al. HorB (HP0127) isa gastric epithelial cell adhesin. Helicobacter 2007;12:200e9.
32. Issa S, Moran AP, Ustinov SN, et al. O-linked oligosaccharidesfrom salivary agglutinin: Helicobacter pylori binding sialyl-Lewis xand Lewis b are terminating moieties on hyperfucosylated oligo-N-acetyllactosamine. Glycobiology 2010;20:1046e57.
33. Linden SK,Wickstrom C, Lindell G, et al. Four modes of adhesionare used during Helicobcater pylori binding to human mucins inthe oral and gastric niches. Helicobacter 2008;13:81e93.
34. Edwards NJ, Monteiro MA, Faller G, et al. Lewis X structures inthe O antigen side- chain promote adhesion of Helicobacter pylorito the gastric epithelium. Mol Microbiol 2000;35:1530e9.
35. Fowler M, Thomas RJ, Atherton J, et al. Galectin-3 binds toHelicobacter pylori O- antigen: it is upregulated and rapidlysecreted by gastric epithelial cells in response to H. pyloriadhesion. Cell Microbiol 2006;8:44e54.
36. Yamaoka Y, Ojo O, Fujimoto S, et al. Helicobacter pylori outermembrane proteins and gastroduodenal disease. Gut 2006;55:775e81.
37. Moran AP, Knirel YA, Senchenkova SN, et al. Phenotypicvariation in molecular mimicry between Helicobacter pylorilipopolysaccharides and human gastric epithelial cell-surfaceglycoforms. Acid-induced phase variation in Lewisx and Lewisy
expression by H. pylori lipopolysaccharides. J Biol Chem2002;277:5785e95.
38. Ruiz-Palacios GM, Cervantes LE, Ramos P, et al. Campylobacterjejuni binds intestinal H(O) antigen (Fuca1,2Galb1,4GlcNAc), andfucosyloligosaccharides of human milk inhibit its binding andinfection. J Biol Chem 2003;278:14112e20.
39. Hu L, Kopecko DJ. Cell biology of human host cell entry byCampylobacter jejuni. In: Nachamkin I, Szymanski CM, Blaser MJ,eds. Campylobacter. 3rd edn. Washington, DC: ASM Press,2008:297e313.
40. Moran AP. Lipopolysaccharides of Helicobacter pylori:importance in gastric adaptation and pathogenesis. In: Sutton P,
Mitchell H, eds. Helicobacter Pylori in the 21st Century.Wallingford, UK: CABI Publishers, 2010:190e211.
41. Deplancke B, Gaskins HR. Microbial modulation of innatedefense: goblet cells and the intestinal mucus layer. Am J ClinNutr 2001;73:1131Se41S.
42. Linden SK, Florin THJ, McGuckin MA. Mucin dynamics inintestinal bacterial infection. PLoS One 2008;3:e3952.
43. Liu TW, Ho CW, Huang HH, et al. Role of a-l-fucosidase in thecontrol of Helicobacter pylori-infected gastric cancer cells. ProcNatl Acad Sci USA 2009;106:14581e6.
44. Moran AP. Relevance of fucosylation and Lewis antigenexpression in the bacterial gastroduodenal pathogen Helicobacterpylori. Carbohydr Res 2008;343:1952e65.
45. Seregni E, Botti C, Massaron S, et al. Structure, function andgene expression of epithelial mucins. Tumori 1997;83:625e32.
46. Perez-Vilar J, Eckhardt AE, DeLuca A, et al. Porcinesubmaxilliary mucin forms disulphide-linked multimers through itsamino-terminal D domains. J Biol Chem 1999;273:1442e9.
47. Lidell ME, Johansson ME, Morgelin M, et al. The recombinantC-terminus of the human MUC2 mucin forms dimers in Chinese-hamster ovary cells and heterodimers with full- length MUC2 inLS 174T cells. Biochem J 2003;372:335e45.
48. Jentoft N.Why are proteins O-glycosylated? Trends Biochem Sci1990;15:291e4.
49. Linden S, Nordman H, Hedenbro J, et al. Strain- and blood group-dependent binding of Helicobacter pylori to human gastric MUC5ACglycoforms. Gastroenterology 2002;123:1923e30.
50. Nordman H, Davies JR, Lindell G, et al. Gastric MUC5AC andMUC6 are large oligomeric mucins that differ in size, glycosylationand tissue distribution. Biochem J 2002;364:191e200.
51. Ota H, Nakayama J, Momose M, et al. Helicobacter pyloriinfection produces reversible glycosylation changes to gastricmucins. Virchows Arch 1998;433:419e26.
52. Moran AP, Ferris JA, Perepelov AV, et al. Adaptation of Lewisantigen expression in LPS of Helicobacter pylori from experimentallyinfected rhesus monkeys. J Endotoxin Res 2004;10:315e16.
53. Wirth HP, Yang M, Sanabria-Valentın E, et al. Host Lewisphenotype-dependent Helicobacter pylori Lewis antigenexpression in rhesus monkeys. FASEB J 2006;20:1534e6.
54. Nilsson C, Skoglund A, Moran AP, et al. An enzymatic rulermodulates Lewis antigen glycosylation of Helicobacter pylori LPS duringpersistent infection. Proc Natl Acad Sci USA 2006;103:2863e8.
55. Nilsson C, Skoglund A, Moran AP, et al. Lipopolysaccharidediversity evolving in Helicobacter pylori communities through geneticmodifications in fucosyltransferases. PLoS One 2008;3:e3811.
56. Solnick JV, Hansen LM, Salama NR, et al. Modification ofHelicobacter pylori outer membrane protein expression duringexperimental infection of rhesus macaques. Proc Natl Acad SciUSA 2004;101:2106e11.
57. Yasui W, Sentani K, Motoshita J, et al. Molecular pathobiologyof gastric cancer. Scand J Surg 2006;95:225e31.
58. Reis CA, David L, Correa P, et al. Intestinal Metaplasia of HumanStomach Displays Distinct Patterns of Mucin (MUC1, MUC2,MUC5AC, and MUC6) Expression1. Cancer Res1999;59:1003e7.
59. Forstner G. Signal transduction, packaging and secretion ofmucins. Annu Rev Physiol 1995;57:585e605.
60. Corfield AP, Myerscough N, Longman R, et al. Mucins andmucosal protection in the gastrointestinal tract: new prospectsfor mucins in the pathology of gastrointestinal disease. Gut2000;47:589e94.
61. Smirnova MG, Guo L, Birchall JP, et al. LPS up-regulates mucinand cytokine mRNA expression and stimulates mucin andcytokine secretion in goblet cells. Cell Immunol 2003;221:42e9.
62. Cabellero-Franco C, Keller K, De Simone C, et al. The VSL#3probiotic formula induces mucin gene expression and secretion incolonic epithelial cells. Am J Physiol Gastrointest Liver Physiol2007;292:G315e22.
63. Lemjabbar H, Basbaum C. Platelet-activating factor receptor andADAM10 mediate responses to Staphylococcus aureus inepithelial cells. Nat Med 2002;8:41e6.
64. Fukushima K, Sasaki L, Ggawa H, et al. Colonization ofmicroflora in mice: mucosal defence against luminal bacteria.J Gastroenterol 1999;34:54e60.
65. Comelli EM, Simmering R, Faure M, et al. Multifacetedtranscriptional regulation of the murine intestinal mucus layer byendogenous microbiota. Genomics 2008;91:70e7.
66. Micots I, Augeron C, Laboise CL, et al. Mucin exocytosis: a majortarget for Helicobacter pylori. J Clin Pathol 1993;46:241e5.
67. Slomiany BL, Slomiany A. Cytosolic phospholipase A2 activationin Helicobacter pylori lipopolysaccharide-induced interferencewith gastric mucin synthesis. IUBMB Life 2006;58:217e23.
1424 Gut 2011;60:1412e1425. doi:10.1136/gut.2010.212704
Recent advances in basic science
on May 21, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.2010.212704 on 12 January 2011. D
ownloaded from

68. Slomiany BL, Liau YH, Lopez RA, et al. Effect of Helicobacterpylori lipopolysaccharide on the synthesis of sulphated gastricmucin. Biochem Int 1992;27:687e97.
69. Byrd JC, Yunker CK, Xu QS, et al. Inhibition of gastric mucinsynthesis by Helicobacter pylori. Gastroenterology 2000;118:1072e9.
70. Morgenstern S, Koren R, Moss SF, et al. Does Helicobacterpylori affect gastric mucin expression? Relationship betweengastric antral mucin expression and H. pylori colonization.Eur J Gastroenterol Hepatol 2001;13:19e23.
71. Byrd JC, Yan P, Sternberg L, et al. Aberrant expression of gland-type gastric mucin in the surface epithelium of Helicobacterpylori-infected patients. Gastroenterology 1997;113:455e64.
72. Cooke CL, An HJ, Kim J, et al. Modification of gastric mucinoligosaccharide expression in rhesus macaques after infectionwith Helicobacter pylori. Gastroenterology 2009;137:1061e71.
73. Kawakubo M, Ito Y, Okimura Y, et al. Natural antibiotic functionof a human gastric mucin against Helicobacter pylori infection.Science 2004;305:1003e6.
74. Lee H, Wang P, Hoshino H, et al. Alpha1,4GlcNAc-cappedmucin-type O-glycan inhibits cholesterol a-glucosyltransferasefrom Helicobacter pylori and suppresses H. pylori growth.Glycobiology 2008;18:549e58.
75. Vinall LE, King M, Novelli M, et al. Altered expression and allelicassociation of the hypervariable membrane mucin MUC1 inHelicobacter pylori gastritis. Gastroenterology 2002;123:41e9.
76. Van den Bovenkamp JH,Mahdavi J, Korteland-Van Male AM, et al.The MUC5AC glycoprotein is the primary receptor for Helicobacterpylori in the human stomach. Helicobacter 2003;8:521e32.
77. McGuckin MA, Every AL, Skene CD, et al. Muc1 mucin limitsboth Helicobacter pylori colonization of the murine gastric mucosaand associated gastritis. Gastroenterology 2007;133:1210e18.
78. Josenhans C, Suerbaum S. The role of motility as a virulencefactor in bacteria. Int J Med Microbiol 2002;291:605e14.
79. Moran AP. Pathogenic properties of Helicobacter pylori. Scand JGastroenterol 1996;31(Suppl 215):22e31.
80. Ottemann KM, Lowenthal AC. Helicobacter pylori uses motilityfor initial colonization and to attain robust infection. Infect Immun2002;70:1984e90.
81. Corfield AP, Wagner SA, Clamp JR, et al. Mucin degradationin the human colon: production of sialidase, sialateO-acetylesterase, N-acetylneuraminate lyase, arylesterase, andglycosulfatase activities by strains of fecal bacteria. Infect Immun1992;60:3971e8.
82. Jones B, Pascopella L, Falkow S. Entry of microbes into the host:using M cells to break the mucosal barrier. Curr Opin Immunol1995;7:474e8.
83. Neutra MR, Mantis NJ, Frey A, et al. The composition andfunction of M cell apical membranes: implications for microbialpathogenesis. Semin Immunol 1999;11:171e81.
84. Lelouard H, Reggio H, Mangeat P, et al. Mucin-related epitopesdistinguish M cells and enterocytes in rabbit appendix andPeyer’s patches. Infect Immun 1999;67:357e67.
85. Lelouard H, Reggio H, Roy C, et al. Glycocalyx on rabbitintestinal M cells displays carbohydrate epitopes from Muc2.Infect Immun 2001;69:1061e71.
86. Terres AM, Pajares JM, Hopkins AM, et al. Helicobacter pyloridisrupts epithelial barrier function in a process inhibited by proteinkinase C activators. Infect Immun 1998;66:2943e50.
87. Valkonen KH, Wadstrom T, Moran AP. Interaction oflipopolysaccharides of Helicobacter pylori with basementmembrane protein laminin. Infect Immun 1994;62:3640e8.
88. Slomiany BL, Piotrowski J, Rajiah G, et al. Inhibition of gastricmucosal laminin receptor by Helicobacter pylori lipopolysaccharide:effect of nitecapone. Gen Pharmacol 1991;22:1063e9.
89. McAuley JL, Linden SK, Png CW, et al. MUC1 cell-surfacemucin is a critical element of the mucosal barrier to infection.J Clin Invest 2007;117:2313e24.
90. Ren J, Agata N, Chen D, et al. Human MUC1 carcinoma-associated protein confers resistance to genotoxic anticanceragents. Cancer Cell 2004;5:163e75.
91. Wei X, Xu H, Kufe D. Human MUC1 oncoprotein regulates p53-responsive gene transcription in the genotoxic stress response.Cancer Cell 2005;7:167e78.
92. Linden SK, Sheng YH, Every AL, et al. MUC1 limits Helicobacterpylori infection both by steric hindrance and by acting asa releasable decoy. PLoS Pathog 2009;5:e1000617.
93. Wright NA, Hoffman W, Otto WR, et al. Rolling in the clover:trefoil factor family (TFF)-domain peptides, cell migration andcancer. FEBS Lett 1997;408:121e3.
94. Clyne M, Dillon P, Daly S, et al. Helicobacter pylori interacts withthe human single- domain trefoil protein TFF1. Proc Natl Acad SciUSA 2004;101:7409e14.
95. Reeves EP, Ali T, Leonard P, et al. Helicobacter pylorilipopolysaccharide interacts with TFF1 in a pH-dependentmanner. Gastroenterology 2008;135:2043e54.
96. Ferreira JA, Azevedo NF, Vieeira MJ, et al. Identification of cell-surface mannans in a virulent Helicobacter pylori strain.Carbohydr Res 2010;345:830e8.
97. Murray E, Khamri W, Walker MM, et al. Expression of surfactantprotein D in the human gastric mucosa and during Helicobacterpylori infection. Infect Immun 2002;70:1481e7.
98. Moran AP, Kharmi W, Walker MM, et al. Role of surfactantprotein D (SP-D) in innate immunity in the gastric mucosa:evidence of interaction with Helicobacter pylorilipopolysaccharide. J Endotoxin Res 2005;11:357e62.
99. Khamri W,Worku ML, Anderson AE, et al. Helicobacter infectionin the surfactant protein D-deficient mouse. Helicobacter2007;12:112e23.
100. Khamri W, Moran AP, Worku ML, et al. Variations inHelicobacter pylori lipopolysaccharide to evade the innateimmune component surfactant protein D. Infect Immun2005;73:7677e86.
101. Salzman NH, Underwood MA, Bevins CL. Paneth cells,defensins, and the commensal microbiota: a hypothesis onintimate interplay at the intestinal mucosa. Semin Immunol2007;19:70e83.
102. Hase K, Murakami M, Iimura M, et al. Expression of LL-37 byhuman gastric epithelial cells as a potential host defensemechanism against Helicobacter pylori. Gastroenterology2003;125:1613e25.
103. Hamanaka Y, Nakashima M, Wada A, et al. Expression ofhuman beta-defensin 2 (hBD-2) in Helicobacter pylori inducedgastritis: antibacterial effect of hBD-2 against Helicobacter pylori.Gut 2001;49:481e7.
104. Hooper LV, Stappenback TS, Hong CV, et al. Angiogenins: a newclass of microbicidal proteins involved in innate immunity.Nat Immunol 2003;4:269e73.
105. Vora P, Youdim A, Thomas LS, et al. Beta-defensin 2 expressionis regulated by TLR signaling in intestinal epithelial cells.J Immunol 2004;173:5398e405.
106. Ogushi K, Wada A, Niidome T, et al. Gangliosides act asco-receptors for Salmonella enteritidis FliC and promote FliCinduction of human beta-defensin 2 expressionn in Caco-2 cells.J Biol Chem 2004;279:12213e19.
107. Duquesne S, Petit V, Peduzzi J, et al. Structural and functionaldiversity of microcins, gene-encoded antibacterial peptides fromenterobacteria. J Mol Microbiol Biotechnol 2007;13:200e9.
108. Putsep K, Branden CI, Boman HG, et al. Antibacterial peptidefrom H. pylori. Nature 1999;398:671e2.
109. Moran AP, Trent MS. Helicobacter pylori lipopolysaccharidesand Lewis antigens. In: Yamaoka Y, ed. Helicobacter Pylori:Molecular Genetics and Cellular Biology. Norfolk, UK: CaisterAcademic Press, 2008:7e36.
110. Round JL, Mazmanian SK. The gut microbiota shapes intestinalimmune responses during health and disease. Nat Rev Immunol2009;9:313e23.
111. Round JL, O’Connell RM, Mazmanian SK. Coordination oftolerogenic immune responses by the commensal microbiota.J Autoimmun 2010;34:J220e5.
112. Mazmanian SK, Round JL, Kasper DL. A microbial symbiosisfactor prevents intestinal inflammatory disease. Nature2008;453:620e5.
113. Chow J, Mazmanian SK. A pathobiont of the microbiotabalances host colonization and intestinal inflammation. Cell HostMicrobe 2010;7:265e76.
114. Szentkuti L, Riedesel ML, Enss K, et al. Pre-epithelial mucuslayer in the colon of conventional and germ-free rats. Histochem J1990;22:491e7.
115. Mack DR, Ahrne S, Hyde L, et al. Extracellular MUC3 mucinsecretion follows adherence of Lactobacillus strains to intestinalepithelial cells in vitro. Gut 2003;52:827e33.
116. Mack DR, Michail S, Wei S, et al. Probiotics inhibitenteropathogenic E. coli adherence in vitro by inducingintestinal mucin gene expression. Am J Physiol 1999;276:G941e50.
117. Alemka A, Clyne M, Shanahan F, et al. Probiotic colonizationof mucus-adherent HT29MTXE12 cells attenuatesCampylobacter jejuni virulence properties. Infect Immun 2010;78:2812e22.
118. Dwek RA, Butters TD, Platt FM, et al. Targeting glycosylationas a therapeutic approach. Nat Rev Drug Discov 2002;1:65e75.
119. Merry T, Astrautsova S. Alternative approaches to antiviraltreatments: focusing on glycosylation as a target for antiviraltherapy. Biotechnol Appl Biochem 2010;56:103e9.
Gut 2011;60:1412e1425. doi:10.1136/gut.2010.212704 1425
Recent advances in basic science
on May 21, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.2010.212704 on 12 January 2011. D
ownloaded from