demography, competitive interactions and grazing effects of intertidal limpets in southern new...
TRANSCRIPT

Demography, competitive interactions and grazing
effects of intertidal limpets in southern New Zealand
R.A. Dunmore*, D.R. Schiel
Marine Ecology Research Group, Zoology Department, University of Canterbury,
Private Bag 4800, Christchurch, New Zealand
Received 14 September 2001; received in revised form 30 September 2002; accepted 27 November 2002
Abstract
Population dynamics and the effects of intraspecific competition on limpet growth and
maintenance of bare patches were investigated for the intertidal limpet Cellana ornata (Dillwyn) at
a boulder-dominated site and on a rocky platform near Kaikoura (South Island), New Zealand.
Distribution and abundance patterns of C. ornata were described in relation to other biota and tidal
height. C. ornata occurred almost exclusively in patches devoid of macroalgae, particularly in the
mid-tidal zone. Both adult and juvenile limpets were most abundant on the tops of boulders, where
their numbers were positively correlated with barnacle cover, which averaged 77%. The size
structure and growth patterns of C. ornata were different between populations. Mark-recapture
studies showed that the slopes of annual growth increments regressed on initial sizes were similar
at both sites but that the annual increments on the platform were about 6 mm greater than on
boulders. Growth virtually ceased at 27 mm for limpets at the barnacle-dominated boulder site and
at 40 mm at the platform site. Recruiting cohorts had 20% survival on boulders and 37% on the
platform during their first year. The largest size classes at both sites had around 57% annual
survival. To test the effects of varying limpet densities on the growth and mortality of limpets and
the maintenance of bare patches, densities of C. ornata were experimentally increased at both
sites. Beyond a density of 4 per 0.25 m2, sizes and survival of limpets were reduced at both sites,
but the effect was more pronounced at the boulder site. Limpets at the boulder site were more
effective at maintaining bare space than those on the reef platform. Enclosing limpets in plots with
and without barnacles showed that C. ornata and a co-occurring species (Cellana denticulata
(Martyn)) grazed more effectively and had greater growth in cleared plots. Overall, there was
0022-0981/02/$ - see front matter D 2002 Elsevier Science B.V. All rights reserved.
doi:10.1016/S0022-0981(02)00579-8
* Corresponding author. Tel.: +64-3-3197700; fax: +64-3-3197701.
E-mail addresses: [email protected] (R.A. Dunmore), [email protected]
(D.R. Schiel).
www.elsevier.com/locate/jembe
Journal of Experimental Marine Biology and Ecology
288 (2003) 17–38

considerable variation in the demographics of C. ornata between populations driven by site-
specific characteristics.
D 2002 Elsevier Science B.V. All rights reserved.
Keywords: Cellana ornata; Limpets; Barnacles; New Zealand; Rocky intertidal; Grazing
1. Introduction
Understanding the life histories of grazers is important to understanding the dynamic
processes of intertidal communities. Numerous studies have shown that grazers can
greatly affect the distribution and abundance patterns of algae and barnacles, and also
provide and maintain much of the bare space in the intertidal zone (see reviews by
Branch, 1981; Lubchenco and Gaines, 1981; Hawkins and Hartnoll, 1983; Creese, 1988;
Underwood and Kennelly, 1990). Limpets are the dominant grazers on many temperate
intertidal shores (reviews by Underwood, 1979; Branch, 1981) and their varying grazing
intensities are a major component of models concerning community structure (Lub-
chenco and Gaines, 1981; Farrell, 1991; Raffaelli and Hawkins, 1996). The demographic
features of limpet populations can therefore have a direct bearing on community
dynamics.
Information on distribution, population structure, growth, mortality and reproduction
of grazers is required to clarify their role in the dynamics of intertidal shores. Because
these characteristics can vary spatially and temporally, comparisons at different shores
through time allow a greater understanding of the demography of species and variability
between populations. For example, the structure of limpet populations and associations
with other species can vary considerably among localized sites (Johnson et al., 1997).
Growth rates can be highly variable and are affected by the densities of conspecifics and
other co-occurring invertebrates, the seasonal supply of food, shore height and sub-
stratum complexity (Lewis and Bowman, 1975; Underwood, 1979; Branch, 1981;
Creese, 1988). Growth rates and maximum sizes can be reduced where barnacles and
other sessile organisms are abundant (Lewis and Bowman, 1975; Choat, 1977;
Thompson, 1980). Reproduction and recruitment are usually highly seasonal but this
varies considerably among species (Branch, 1981). Mortality rates also vary widely
within and between species. They are related not only to species’ life histories (Choat
and Black, 1979) but also to the characteristics of habitats, especially the presence of
sessile organisms in conjunction with competitive interactions (Lewis and Bowman,
1975; Branch, 1976; Choat, 1977; Underwood, 1978; Underwood and Jernakoff, 1981;
Creese and Underwood, 1982; Marshall and Keough, 1994). The integration of life
histories with habitat characteristics, therefore, determines demographic responses to the
local environment.
In New Zealand, there is a relatively diverse fauna of limpets, which are prominent on
most rocky intertidal shores (Morton and Miller, 1973; Powell, 1979). However, the role
they play in intertidal community structure and the demographic features of even the most
common species have barely been investigated. As on rocky shores worldwide, limpets in
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–3818

southern New Zealand are usually associated with algal films, crusts and filamentous algae,
and there is considerable variation in species composition and abundances among sites
(Raffaelli, 1979; Creese, 1988). The limpet fauna is especially rich in southern NewZealand,
where several species of patellid limpets of the genus Cellana co-occur in the less-vegetated
portions of the intertidal zone (Morton and Miller, 1973; Powell, 1979). The ornate limpet
Cellana ornata is one of the most common and widely distributed limpets and is found along
the length of the country. It is greatly abundant along the shores of the central South Island
where it occurs on open reefs and boulder habitats and is the only abundant patellid limpet
species commonly associated with barnacles (cf., Choat and Black, 1979).
This work was prompted by the observations that C. ornata is the most abundant grazer
in many intertidal areas, particularly in patches with few macroalgae, and that it tended to
be smaller on boulders than on open reefs. We wished to understand its population
dynamics and variability between sites, and how it affects wider community processes
through maintenance of bare patches. To achieve this, the distribution, population
structure, growth and mortality of C. ornata were described at two sites. We experimen-
tally tested for variation in intraspecific competition and grazing ability of C. ornata by
manipulating densities of limpets at the two sites. The effect of barnacle cover on limpets
was also tested, using C. ornata and a co-occurring limpet, Cellana denticulata. It was
hypothesized that the limpets were not able to feed effectively in barnacle-covered areas,
resulting in decreased sizes.
2. Methods
2.1. Study sites
Studies on the distribution, abundance, growth, mortality and conspecific grazing of C.
ornata were done at two sites, First Bay and Blue Duck, situated near Kaikoura on the east
coast of the South Island, New Zealand (42j25VS, 173j42VE; Fig. 1). The barnacle/grazingexperiment involving both C. ornata and C. denticulata was done only at Blue Duck,
where barnacle cover was dense. First Bay and Blue Duck are 20 km apart and differ in
several characteristics. First Bay is a moderately exposed siltstone platform located on the
Kaikoura Peninsula. It is generally protected from severe swells by rocky outcrops
projecting from the peninsula. The platform is ca. 50 m from the low to high tide marks
and ca. 70 m wide. Blue Duck is moderately exposed at most times but is subjected to
strong wave action during southerly storms. It is composed mostly of large greywacke
boulders, extends ca. 35 m from the low to high tide marks and is several hundred meters
long. A major difference biologically between the sites was the presence of dense patches
of barnacles on the boulder tops at the Blue Duck site, while few barnacles occurred at the
First Bay reef site.
2.2. Distribution and abundance
To determine the abundance patterns seasonally of C. ornata, stratified random
sampling was done in three tidal zones (0.2–0.7, 0.7–1.2 and 1.2–2.0 m above chart
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–38 19

datum), characterized by abrupt changes in habitat at similar tidal positions in both
sites. At both sites, limpets were entirely confined to patches where rock surfaces were
predominantly bare of foliose macroalgae, although barnacles, algal films and crusts
were common. Most of these patches looked superficially to be bare and all were >1
m2 in area. Randomly placed 0.25 m2 quadrats were sampled within these patches in
winter (July 1995), spring (October 1995), summer (January 1996) and autumn (April
1996). At First Bay, 10 quadrats were sampled within each zone. At Blue Duck, the
tops and the sides of boulders (n = 10 quadrats each) were sampled separately within
each tidal zone. The sampling quadrat was divided into a 100-square grid, which was
used to estimate the percentage cover of algal species and barnacles. Juvenile (< 10-
mm shell length; Dunmore and Schiel, 2000) and adult C. ornata were counted
separately to identify recruitment patterns. The other common limpet in the study sites,
C. denticulata, was also counted within quadrats. Analyses of variance were used to
test the various factors on limpet abundances. All data sets were first tested for
homogeneity of variances using Cochran’s test, and data transformations were done if
necessary to stabilize variances. For ANOVAs of limpet abundances testing site
(n= 2), season (n = 4) and tidal height (n = 3), only data from the tops of boulders
at Blue Duck were used and compared to the flat reef at First Bay. Site was nested
within season for analysis, and both were treated as random factors. In this analysis,
therefore, ‘‘season’’ simply represents four different times of sampling spaced through
Fig. 1. Map of the Kaikoura Peninsula showing study sites and location within New Zealand.
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–3820

the year. Correlation analysis between C. ornata and the other main organisms was
done.
2.3. Growth and mortality
To examine the population size structure through time, C. ornata were measured bi-
monthly from February 1995 to February 1996 at both sites. A 0.25-m2 quadrat was used
to sample randomly and all limpets within each quadrat were measured to the nearest 0.5
mm using Vernier calipers. Quadrats were repeated across all shore heights until 500
limpets had been measured. Data were plotted as histograms to show the size structure of
each population through time and to identify the arrival of recruits.
To determine growth rates, 200 C. ornata were tagged in February 1995. Individuals
were chosen from the full size range across all shore heights at each site. A small plastic
numbered tag was glued with superglue to each shell and then covered with clear epoxy
resin for further protection. C. ornata is a homing species, and the location of each limpet
was mapped to facilitate later recovery. Tagged limpets were re-measured bi-monthly from
February 1995 to February 1996. Missing tags were replaced. Individuals that had tags
missing were easily identified from their mapped position and from a residual glue mark
left on the shell from the previous tag.
Annual growth rates were calculated using only those limpets that survived the whole
year. Size-specific survival was calculated for the year, based on the recovery of tagged
and mapped limpets.
2.4. Intraspecific effects
The null hypothesis that limpet density had no effect on growth and mortality of C.
ornata and on algal abundance was tested at First Bay and Blue Duck. The experiment
was initiated in October 1995 and continued for 7 months until May 1996. There were five
experimental treatments and three controls, each with four replicates. All plots were 0.25
m2 (50� 50 cm). The perimeter of experimental plots was painted with a copper-based
paint (5 cm wide) to include and exclude limpets. These plots were scraped to the bare
substratum using a chisel and wire brush.
The average densities of C. ornata in 20 random quadrats in midshore areas were
15.9F 0.36 per 0.25 m2 at Blue Duck and 7.6F 0.11 per 0.25 m2 at First Bay. Using these
densities as a guide, treatments were set at 0, 4, 8, 16 and 24 limpets per 0.25 m2. Limpets
were collected from the same tidal height from areas adjacent to each experimental area
and were 19–24 mm in size. Only limpets that were introduced to plots in October 1995
were used in analyses. Limpet densities were checked and maintained at least once every 3
weeks.
Three control treatments were used. Unmanipulated controls were untouched and were
indicative of the natural state of the substratum during the experiment. Scraped control
plots were cleared in the same way as the experimental plots, allowing the effects of the
scraping on subsequent recruitment to be identified. These two control treatments were not
bordered by paint, but had their corners marked with plastic tags nailed to the substrate.
The third controls (paint controls) were scraped and were bordered with dashed paint,
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–38 21

allowing the movement of limpets in and out of the plots while the plot was still subjected
to any effects of the paint. Algal coverage and chlorophyll concentration were measured
each month as described below.
The percentage cover of algae and bare space was estimated monthly using a 0.25-m2
quadrat sub-divided into 100 squares. Analysis of variance (ANOVA) was used to test for
treatment effects 2, 4 and 7 months after the initiation of the experiment. Chlorophyll a
was analysed as a measure of microalgal concentration on what appeared to be bare rock
(Hill and Hawkins, 1990). Three replicate 1-cm2 rock scrapings were taken at random
from each plot monthly using a flat-edged chisel. The scrapings were ground with 10 ml of
90% acetone, and left in a darkened fridge for 24 h for the chlorophyll to be extracted. The
mixture was then put through a glass filter and the absorption level read using a Kontron
Uvikon Spectrophotometer set at 665- and 750-nm wavelengths. Chlorophyll a concen-
Fig. 2. C. ornata adults (A), juveniles (B) and C. denticulata (total, C) on the tops (black bars) and sides (open
bars) of boulders at Blue Duck; and C. ornata adults (D), juveniles (E) and C. denticulata (total, F) on the reef at
First Bay. Mean number per quadrat (F S.E.) averaged over one year (n= 40).
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–3822

tration per cm2 per treatment for each month was then obtained using the formula (HMSO,
1986):
chlorophyll a ðmg=cm2Þ ¼ 11:9� Absorption reading� 10 ml
3 cm2
Average chlorophyll a concentrations per treatment were plotted against time. ANOVA
was used to test the treatment effects at 2, 4 and 7 months. Data were log-transformed to
homogenize variances.
To assess intraspecific effects, original limpets in each of the density treatment plots
were marked with a fast-drying paint and measured at the start and the end of the
experiment. Survival of these limpets was monitored monthly to investigate density-
dependent mortality. ANOVA was used to test site (treated as a random factor) and
treatment effects on shell length.
At the end of the experiment, dry weights of the original limpets were obtained by
dissecting the visceral masses from the shells and drying them to a constant weight.
Twenty limpets within the 19–24-mm size range were also collected at random from
around the experimental area at the start (October 1995) and end (May 1996) of the
experiment and dried to a constant weight. These were used for comparison with the
treatment limpets. Differences in limpet sizes were standardized by dividing the limpets’
dry weight by shell length (Fletcher and Creese, 1985). ANOVAwas used to test the effect
of site and treatment on the dry weight/shell length ratio. Data were square root-
transformed to homogenize variances.
2.5. Limpet–barnacle interactions
To test the null hypothesis that barnacles have no effect on the growth of C. ornata
and the co-occurring limpet C. denticulata, an experiment was done at Blue Duck at
Table 1
Mean (S.D.) abundances of attached organisms and bare space in the limpet-dominated patches at three shore
heights at both study sites (n= 40)
Blue Duck First Bay
Boulder tops Boulder sides High Mid Low
High Mid Low High Mid Low
Barnacles 8 (10.6) 77 (22.5) 9 (14.2) 3 (5.8) 48 (30.4) 4 (5.4) 6 (7.4) 11 (13.8) 1 (5.4)
Algae 0 (0.2) 0 (0.5) 3 (7.5) 0 (0.2) 1 (3.3) 5 (12.0) 1 (1.3) 3 (5.3) 9 (13.4)
Bare space 92 (4.7) 25 (14.5) 76 (9.2) 97 (2.6) 48 (11.6) 71 (12.0) 93 (3.4) 86 (5.9) 90 (6.3)
Littorinids 45 (53.1) 0 (0) 0 (0) 10 (25.8) 0 (0) 0 (0) 90 (27.5) 21 (38.1) 0 (0)
Other grazers 25 (18.1) 47 (25.0) 40 (25.0) 22 (12.1) 48 (22.6) 34 (17.7) 9 (5.3) 20 (11.2) 15 (8.2)
Data are pooled across seasons. Barnacles, algae and bare space are % cover, littorinids and other grazers are
number per 0.25 m2.
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–38 23

Fig. 3. C. ornata. Size frequency histograms at Blue Duck and First Bay from February 1995 to February 1996
(n= 500 per sample).
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–3824

mid-tidal level on small boulders from December 1995 until April 1996. There were
two treatments: bare (scraped plots) and natural plots that had initially between 60%
and 90% coverage of barnacles. Experimental plots were 0.0625 m2 (25� 25 cm), and
a barrier of copper-based paint (5 cm wide) was put around the outside of the plots to
include and exclude limpets. There were no paint controls used in this experiment
because there was no effect in any of our previous experiments. The average densities
of limpets in the area were 3.4F 0.33 of C. ornata and 2.1F 0.17 of C. denticulata
per 0.0625 m2 in 20 randomly thrown quadrats. Using these densities as a guide,
treatment densities were set at three limpets per plot, each with four replicates. The
limpets used were collected from around the experimental area and were between 19
and 24 mm. Only limpets that were introduced to plots in the first 4 weeks were used
in analyses.
At the end of the experiment, original limpets were removed and dry weights were
obtained by dissecting the visceral masses from the shells and drying them to a constant
weight. For analyses, this measurement was standardized by dividing the dry visceral
weight by the shell length (Fletcher and Creese, 1985). ANOVAwas used to test the effect
of treatment on the dry weight/shell length ratios for each species. Data were square root-
transformed to homogenize variances.
Fig. 4. C. ornata. Annual growth increment (mm) plotted on initial sizes at Blue Duck (n= 48) and First Bay
(n= 86), with regression equations inset.
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–38 25

3. Results
3.1. Distribution and abundance
Over an entire year, C. ornata adults and juveniles were more abundant in the boulder
habitat of Blue Duck than on the platform at First Bay (Fig. 2A,B,D,E). C. ornata reached
their greatest average densities of 20 adults and 6 juveniles per 0.25 m2 at Blue Duck (Fig.
2A,B), and 6 adults and 3 juveniles per 0.25 m2 at First Bay (Fig. 2D,E), in the mid-tidal
zone. Within the boulder habitat at Blue Duck adults were 1.5 times more abundant on the
tops than on the sides of boulders (Fig. 2A) and juveniles were twice as abundant on tops
(Fig. 2B). C. denticulata were more abundant at Blue Duck (mean = 6.3, S.E. = 0.38; Fig.
2C) than at First Bay (mean = 4.7, S.E. = 0.35; Fig. 2F). At Blue Duck, C. denticulata
reached their greatest average densities in the high tidal zone on the sides of boulders
Fig. 5. C. ornata. Percentage survival of 10-mm size classes of initial tagged cohorts through 1 year at Blue Duck
(A) and First Bay (B).
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–3826

(mean = 10.7, S.E. = 1.22, Fig. 2C). Overall, C. denticulata were more abundant on the
sides of boulders, at least in the mid and upper zones. On the platform at First Bay (Fig.
2F), C. denticulata were most abundant at the low shore (mean = 6.5, S.E. = 0.35).
These general patterns were consistent throughout the four sampling periods of the year
(i.e., ‘‘season’’ was ns for all variables) and the spatial factors were most important. The
abundance of C. ornata adults varied between sites within seasons across tidal heights
(site(season)� tidal height: F8,216 = 6.03, p < 0.001) as did the abundance of C. ornata
juveniles (site(season)� tidal height: F8,216 = 5.68, p < 0.001) and C. denticulata (adults
and juveniles combined: site(season)� tidal height, F8,216 = 3.16, p < 0.01).
Within the limpet-dominated patches, the most abundant attached organisms were
barnacles (Table 1), mostly Chamaesipho brunnea and C. columna, and they persisted
throughout the year (unpublished data). These averaged up to 77% cover at Blue Duck and
were particularly abundant in the mid-tidal zone, where C. ornata was most abundant.
Barnacles were less abundant at First Bay. Macroalgae were sparse except for encrusting
forms, and most rock surfaces were bare at all tidal heights at both sites. Small littorinid
snails were abundant in the high tidal zone, while a variety of other gastropods (mostly
trochids and small limpets) occurred at all tidal levels. These varied in abundance between
sites and tidal position. For example, Siphonaria zelandica (Quoy and Gaimard) occurred
at average densities of 15–21 per 0.25 m2 on the tops and sides of boulders at Blue Duck
in the mid to low tidal zones but at only 1–2 per 0.25 m2 in the high zone. At First Bay,
however, this species occurred at only 6–7 per 0.25 m2 in the mid to low zones. Cellana
radians (Gmelin), Cellana flava (Hutton) and Melagraphia aethiops (Gmelin) generally
occurred at densities less than 1 per 0.25 m2.
The densities of adult C. ornata were significantly correlated with C. ornata juvenile
densities both at Blue Duck (r240 = 0.44, p < 0.001) and First Bay (r120 = 0.61, p < 0.001).
At Blue Duck, both adult (r240 = 0.52, p < 0.001) and juvenile (r240 = 0.44, p < 0.001)
densities were positively correlated with barnacle cover while C. denticulata were
negatively correlated with barnacle cover (r240 =� 0.21, p = 0.002). These correlations
are a reflection of the greater abundances of C. ornata on boulder tops, where barnacles
Table 2
Summary of ANOVA tests (Mean square and F values) of bare space in December 1995, February 1996 and May
1996
Source of variation df December February May
MS F MS F MS F
Site 1 0.27 13.24*** 2182.32 7.28** 2019.68 14.21***
Treatment 7 1.20 41.74*** 10,588.47 15.38*** 9910.53 24.69***
Density treatment vs. controls 1,7 4.65 161.56*** 41,833.95 60.83*** 30,149.75 75.12***
Among density treatments 4,7 0.98 33.99*** 8671.67 12.59** 10,167.17 25.33***
Site� treatment 7 0.29 1.44 688.52 2.30* 401.34 2.82*
Residual 40 0.20 299.69 142.16
December data were square root arcsine transformed to stabilize variances; May variances could not be stabilized
and raw data were used.
*P< 0.05.
**P< 0.01.
***P< 0.001.
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–38 27
had a relatively high cover, and the greater abundances of C. denticulata on boulder sides,
where barnacles are sparse.
3.2. Growth and mortality
There were considerable differences in the size structure of the populations of C. ornata
at the two sites throughout the year (Fig. 3). The largest size recorded at Blue Duck was 36
mm but limpets reached 50 mm at First Bay. The major size mode at Blue Duck throughout
the year was 18–20 mm. There were probably two modes at Blue Duck in February 1995,
one at around 12 mm and the other at 18 mm. During the same month, the major modes at
First Bay were at 16, 26 and 34 mm. Recruits first appeared in the June 1995 sample at
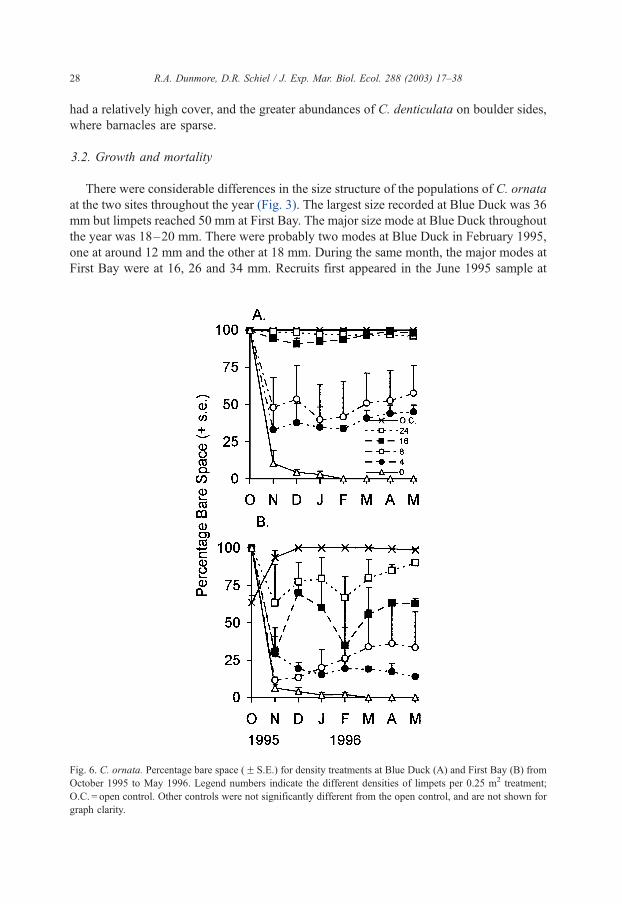
Fig. 6. C. ornata. Percentage bare space (F S.E.) for density treatments at Blue Duck (A) and First Bay (B) from
October 1995 to May 1996. Legend numbers indicate the different densities of limpets per 0.25 m2 treatment;
O.C. = open control. Other controls were not significantly different from the open control, and are not shown for
graph clarity.
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–3828

both sites, but recruitment was greater at Blue Duck than at First Bay. There was evidence
of faster growth at First Bay. By August 1995, the major mode of recruits at Blue Duck was
9–10 mm while it was 12–14 mm at First Bay. By February 1996, both sites were similar
to their size structure a year earlier. There were two major modes at Blue Duck, but at least
three and possibly four at First Bay, with the largest mode at 42 mm.
The mark-recapture study showed that C. ornata at Blue Duck had lower growth
increments in all size classes than those at First Bay (Fig. 4). The slopes of the incremental
growth regressions were parallel (F1,130 = 0.97, p = 0.326) indicating that the rate of
change in length was similar at the two sites. However, for any given size, limpets at Blue
Duck had a lower growth increment than those at First Bay (F1,131 = 127.3, p < 0.001). For
example, a limpet of 10-mm initial size grew an average of around 6 mm of linear shell
growth annually at Blue Duck, while a similarly sized limpet at First Bay grew around 13
mm. Growth ceased at around 27 mm at Blue Duck and at around 40 mm at First Bay.
Fig. 7. C. ornata. Chlorophyll a levels (Ag/cm2) (F S.E.) for limpet density treatments at Blue Duck (A) and First
Bay (B) from November 1995 to May 1996. Legend numbers indicate the different densities of limpets per 0.25
m2 treatment; O.C. = open control. Other controls were not significantly different from the open control, and are
not shown for graph clarity.
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–38 29

Growth rates were quite variable among limpets at First Bay, particularly between 10 and
15 mm, for which some limpets added only 8 mm during the year while others added as
much as 18 mm. These growth rates are consistent with the population structure
throughout the year (Fig. 3). At Blue Duck, limpets of 18 mm on average added only
another 3 mm of growth, so the major mode did not change much throughout the year.
However, similarly sized limpets at First Bay added around 9 mm, so the major modes
progressed in size throughout the year.
Because of variation in individual limpet growth, the slow growth of older limpets and
the coalescing of cohorts, it is not possible to estimate longevity accurately. However, by
comparing the size-frequency data and annual growth increments, cohort ages are identifi-
able and estimates of age-at-length can be made. Blue Duck limpets had attained mean
lengths of around 14–16 mm after 1 year but only around 22 mm after their second year.
The largest limpets, at 25 mm or more, grew little and were at least 3 years old. First Bay
limpets attained average lengths of 20–22 mm after their first year on the shore, 32–34
mm after their second year, and 38–40 mm after 3 years. There was, however,
considerable variation among limpets in the time taken to achieve these sizes. The largest
limpets at First Bay, at ca. 40 mm, grew very little and were at least 3–4 years old.
There were differences among sites and cohorts in survival rates (Fig. 5). At Blue Duck,
limpets in the 4–13-mm size class had a sharp drop in percentage survival in the first 4
months, but mortality was more gradual after this (Fig. 5A). Survival was 20% after 1 year.
This smallest size class showed a more steady decline at First Bay and reached a low of
37% survival after 1 year (Fig. 5B). The decrease in survival of the middle size class (14–
23 mm initial size) was gradual at both sites, reaching 38% at Blue Duck and 42% at First
Bay. The largest size classes at both sites had generally similar trends in survival and were
around 57% after 1 year. The large sizes had relatively good annual survival providing
further evidence, in addition to the small incremental growth of these limpets, that there is
coalescence of age groups in the largest size classes.
Table 3
Summary of ANOVA tests (Mean square and F values) of chlorophyll concentration (mg/cm2) in December 1995,
February 1996 and May 1996
Source of variation df December February May
MS F MS F MS F
Site 1 147.84 4.43* 0.25 1.15 5.50 0.022
Treatment 7 421.88 11.12** 10.68 19.68*** 2586.74 8.55**
Density treatment vs. controls 1,7 748.41 19.67** 37.99 70.00*** 6104.01 20.18**
Among density treatments 4,7 606.02 15.97** 9.01 16.60** 3148.62 10.41**
Among controls 2,7 1.62 0.04 0.16 0.30 69.82 0.04
Site� treatment 7 37.94 1.14 0.54 2.53* 302.54 1.23
Residual 40 33.40 0.21 245.31
December and May variances could not be stabilized and raw data were used; February data were log-transformed
to stabilize variances.
*P < 0.05.
**P < 0.01.
***P < 0.001.
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–3830

3.3. Intraspecific effects
Increasing the densities of C. ornata had great effects on the amount of bare space
throughout the year. Most site and density factors were significant but by February the
treatment effects varied significantly between the sites (Table 2). At Blue Duck, limpet
densitiesz 16 per 0.25 m2 maintained ca. 100% bare space, which was comparable to the
open control areas (Fig. 6A). On the reef at First Bay, however, only the density 24
treatment had bare space approaching that of controls, but this occurred only towards the
end of the experiment (Fig. 6B). The density 16 treatment maintained 90–98% bare space
at Blue Duck but only ca. 60% at First Bay. At lower limpet densities at both sites, ulvoids
contributed most of the macroalgal cover but Porphyra spp. and Ectocarpus spp. were also
present seasonally. At the start of the experiment, First Bay control plots contained
Porphyra spp. but the algae disappeared with the onset of warmer weather (Fig. 6B).
Treatments without limpets had virtually no bare space after February. At the end of the
experiment, treatments with 0, 4 and 8 limpets had less than 50% bare space while those
with 16 and 24 limpets, and the open controls, had >50% bare space.
The chlorophyll a concentrations, which are an indication of microalgal abundance, in
the density treatments mostly reflected the macroalgal cover (Fig. 7, Table 3). Chlorophyll
Fig. 8. C. ornata. Mean shell lengths (F S.E.) at the start (October) and end (May) of the density experiment, at
Blue Duck (A) and First Bay (B).
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–38 31

a concentrations were greater for most of the experiment in densities 4 and 8 at First Bay
than at Blue Duck. The overall greater chlorophyll a levels at First Bay (Fig. 7B) indicated
either greater production at this reef site or else a more intense grazing effect at the boulder
site of Blue Duck.
Greater densities of limpets resulted in decreased shell growth at both sites (Fig. 8A,B),
but the effect was more pronounced at Blue Duck (site� density interaction at end of
experiment: F3,163 = 4.95, p = 0.026). At densities of 8–24 per treatment, final lengths
were greater at First Bay than at Blue Duck. However, at Blue Duck, the limpets in all
treatments had much greater shell growth than in the natural population (cf., Fig. 4). For
example, the annual growth in shell length of limpets between 19 and 24 mm at Blue Duck
was only 1–2 mm in the natural population but in the experiment, similarly sized limpets
grew an average of 4.0–9.1 mm in only 7 months (Fig. 8A). Blue Duck Limpets at density
4 reached mean lengths of ca. 32 mm, which is close to the maximum size in the natural
population. First Bay experimental limpets were closer in growth to the natural population,
but attained 5.5–7-mm shell increments in 7 months (Fig. 8B) compared to 5–7-mm
annual increments in the natural population (Fig. 4).
Fig. 9. C. ornata. Mean dry weight of limpet tissue/shell length (F S.E.) at the end of the experiment, at Blue
Duck (A) and First Bay (B). The mean weight of 20 randomly sampled limpets at the start (N1) and end (N2) of
the 7-month experiment are also shown.
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–3832

To test for overall differences at the end of the experiment, dry weights of limpets were
adjusted for shell length. There were significant differences among sites (F1,163 = 7.03,
p = 0.009), density treatments (F1,163 = 28.58, p < 0.001) and in the interaction between
sites and treatments (F1,163 = 8.27, p < 0.001). Greater densities resulted in decreased
weights but, as for shell growth, the effect was more pronounced at Blue Duck. At this
site, however, all treatments had greater average weights than the limpets in natural
populations at the start (Fig. 9A, N1) and end (Fig. 9A, N2) of the experiment. At First
Bay, only the density 4 treatment had a much higher mean weight than the other treatments
and natural population (Fig. 9B).
Survival was also density-dependent, especially between 8 and 24 limpets per 0.25 m2
(Fig. 10). Across all densities, survival was greater at Blue Duck than at First Bay. Within
the density 4 treatment, there was 94% survival at Blue Duck but only 75% at First Bay.
By the end of the experiment, all of the higher densities had 40–45% survival at Blue
Duck and only 29–31% survival at First Bay.
Fig. 10. C. ornata. Percent survival through time in experimental treatments of 4, 8, 16 and 24 limpets per 0.25
m2 at Blue Duck (A) and First Bay (B).
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–38 33

3.4. Limpet–barnacle interactions
Where barnacles were experimentally removed, both C. ornata and C. denticulata were
larger than in barnacle-covered treatments after 4 months (Fig. 11). The treatment effect
varied between the species (treatment� species: F1,42 = 11.86, p= 0.001). C. denticulata
had similar sizes in open and barnacle-covered areas, but C. ornata was much more
affected by the presence of barnacles. C. ornata was ca. 50% larger in areas where
barnacles were removed. Although we did not set out to test for effects on algal cover in
this experiment, related treatments in which limpets were excluded from barnacle-covered
and cleared areas were completely smothered by algae (mostly Enteromorpha and Ulva)
after 4 months (Dunmore, personal observation).
4. Discussion
The population dynamics of C. ornata are closely linked to their local environment.
There were distinct differences between the two sites in their population structure, density,
growth and reproduction. While this is not unusual for grazing molluscs, there are
Fig. 11. C. ornata, C. denticulata. Mean dry weights (F S.E.) of limpet tissue/shell length in plots with and
without barnacles at the end of the experiment at Blue Duck. Density of limpets was 3 per 0.0625 m2 quadrat.
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–3834

important ramifications for understanding the structuring processes of intertidal shores in
southern New Zealand.
C. ornata is considered a mid to high shore species and generally occurs above the
other limpet species (Smith, 1969; Morton and Miller, 1973). Our study showed, however,
that this depends on site characteristics. On the reef site, C. denticulata dominated the low
shore bare patches while C. ornata was more abundant on the mid-shore. In the more
exposed boulder site, both species were abundant in the mid and upper shore zones but C.
ornata dominated the boulder tops where barnacles were abundant while C. denticulata
was more numerous on the sides of boulders. C. ornata is clearly able to live in a wide
range of conditions. Smith (1975) determined that C. ornata had variable desiccation
resistance and metabolic rates according to exposure on the shore and tidal height. Limpets
at high tidal levels on sheltered shores were more desiccation-resistant and had lower
metabolic rates than those at low tidal levels. The variability in physiological response to
environmental stresses enables C. ornata to inhabit a broad range of conditions but it
reaches its greatest densities in barnacle-dominated patches on boulders at semi-exposed
sites throughout the eastern coast of the South Island. No other large species of
prosobranch limpet occurs in high densities in barnacles in southern New Zealand, but
some small species of acmaeid limpets can be common.
Distributions of many limpet species worldwide are positively correlated with those of
barnacles (e.g., Patella granularis, Branch, 1976; Acmaea digitalis, Choat, 1977; Patella
latistrigata, Creese, 1982). This association can result in both positive and negative effects
on barnacle and limpet populations. Limpets directly affect barnacles by crushing newly
settled cyprids and bulldozing juveniles (Branch, 1981; Underwood et al., 1983; Hawkins,
1983). Indirectly, they affect barnacles by grazing algae off barnacle tests and the
surrounding rock, thus preventing smothering and allowing settlement to occur on the bare
rock (Hawkins, 1983; Hawkins and Hartnoll, 1983; Underwood et al., 1983). This indirect
effect was observed in our limpet–barnacle experiment. In plots where all limpets had died,
increased barnacle mortality due to algal overgrowth was observed (personal observation).
Barnacles can have great effects on limpet growth, reproduction, survival and com-
petitive interactions. While the heterogeneous surface provided by barnacles can reduce
the physical stresses of desiccation and wave action, the presence of barnacles also reduces
the area available for grazing by occupying space. They may also filter potential food
before it settles on the substratum (Branch, 1976; Choat, 1977; Underwood et al., 1983).
Barnacles can provide a refuge for small limpets from competition with larger individuals
(Creese, 1982), which may be unable to forage effectively in areas of dense barnacle cover
(Choat, 1977; Geller, 1991). The trade-off for limpets, however, may be reduced growth
rates, as found in our study (and see Lewis and Bowman, 1975; Thompson, 1980;
Hawkins and Hartnoll, 1982). Nonetheless, larger size is not always beneficial because
once limpets reach larger sizes, they may undergo abrupt mortality. For example, Choat
(1977) found that small A. digitalis fed within cracks and interstices of barnacles, while
larger individuals could not and underwent heavy mortality. In our study, all size classes of
C. ornata were more abundant on the tops of boulders, where barnacle cover was greatest.
C. ornata of all sizes tended to stay in the localized area to which they recruited, regardless
of habitat. No movement of tagged C. ornata was seen either away from or into barnacle
patches (personal observation).
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–38 35

Both C. ornata and C. denticulata recruited into barnacles, but large C. denticulata
were rarely found there. In contrast to C. ornata, it seems likely that C. denticulata migrate
with increasing size to the sides of boulders. Small C. denticulata survive and grow well
among barnacles (Fig. 11) but it is unlikely that they can do so as adults. C. denticulata
can reach shell lengths >70 mm (Powell, 1979; Walters, 1994) which is almost double the
maximum length reached by C. ornata. The interstices among barnacles on boulder tops
are not suitable habitat for such large limpets.
Although C. ornata was more numerous in the boulder site, it is unclear how this
relates to overall settlement patterns of larvae, exposure, differences in substratum, and
varying biological characteristics within and between sites. It remains unknown whether
C. ornata larvae settle preferentially in particular habitats or in areas of high adult density,
or if settlement is haphazard among habitats and densities and post-settlement mortality
sets distribution patterns. Newly settled juveniles are very small and often cryptic.
Judging from the timing of reproduction and population size-frequency measurements,
limpets are at least 4 months old by the time they become readily visible on the shore.
One potential mechanism of differential mortality among habitats is bulldozing of C.
ornata settlers on the sides of boulders by large C. denticulata, which may out-compete
C. ornata for space.
Most life history parameters of C. ornata at the barnacle site were negatively affected by
the high densities of the barnacles. Survival in the first year of life was much poorer in the
barnacle/boulder site than on the open reef site (18% vs. 31%). Gonad development was ca.
32% less at the barnacle/boulder site (Dunmore and Schiel, 2000), growth increments were
47% lower and the maximum size attained was 28% lower. These findings reflected those
found on British shores (Lewis and Bowman, 1975; Thompson, 1980; Hawkins and
Hartnoll, 1982) for Patella vulgata, which had reduced growth and smaller maximum sizes
in habitats with dense barnacle cover. The population structure was also affected. Limpets in
barnacle habitats had a greater overlap in size ranges of year classes, making it difficult to
separate cohorts. This structure is similar to the C. ornata population at Blue Duck.
However, P. vulgata populations on bare rock habitats had bi- or tri-modal size-frequency
distributions, similar to the structure of the C. ornata population at First Bay.
Although wave action, topography, substrate type, and available food may have
affected C. ornata populations, the experiments clearly indicate the strong effects that
barnacle cover and limpet density have on C. ornata. C. ornata was capable of
maintaining essentially bare space, especially at higher densities where there was reduced
limpet and algal growth. Experimentally, there were clear intraspecific effects on both
growth and survival of limpets at both sites. Survival at both sites was relatively poor at
densities beyond 4 per 0.25 m2, which is below the natural densities seen at both sites.
These experimental results may not be an accurate reflection of mortality at the natural
sites where limpets are free to move around over greater areas. In terms of growth,
however, most experimental limpets had greater shell and tissue growth than seen in the
natural populations, especially at Blue Duck (Fig. 9). Two factors could account for this.
First, the experimental plots were cleared of barnacles, which enabled the limpets to feed
more effectively and increased the available grazing space. Second, the natural populations
had high numbers of other small grazers, especially at Blue Duck. Interspecific competi-
tion may therefore have had stronger effects on limpets in the control plots.
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–3836

Limpets at the boulder site appeared to be more effective grazers than those at the reef
site, illustrated by the greater amount of bare space in experimental plots at Blue Duck.
This may have been due to greater productivity on the reef, more foraging time for limpets
at Blue Duck due to increased wave exposure, or a response to localized nutrient addition.
Even at a density of 24 limpets per 0.25 m2 at First Bay, limpets were not successful in
maintaining plots clear of macroalgae, even though this density was greater than that of the
total grazers at this site. Overall, therefore, C. ornata alone does not appear to be capable
of maintaining the bare space normally seen on this reef. Nutrient addition is possible at
this site, however, because of the large numbers of New Zealand fur seals (Arctocephalus
fosteri) that reside in the area for much of the year.
C. ornata is clearly a generalist in its ability to recruit, grow and reproduce in intertidal
habitats. It is also one of the few limpets widely distributed along the length of New
Zealand. On the low shore, it is probably out-competed by the larger Cellana species,
particularly the abundant and ubiquitous southern species C. denticulata. C. ornata,
however, effectively exploits the mid and upper shore and barnacle-dominated areas where
few other molluscs (except small littorinids) live. Its population dynamics are shaped by
highly localized conditions, producing strong spatial and temporal variation in demo-
graphic characteristics. C. ornata are able to prevent the growth of macroalgae, and are
clearly a major agent in the maintenance of the bare patches seen throughout the mid-tidal
zone in southern New Zealand. However, densities required to maintain such patches also
vary between sites.
Acknowledgements
We thank Jack van Berkel and the University of Canterbury for the logistic support and
the Foundation for Research, Science and Technology (Grants UOC511 and UOC610) for
the financial support. [AU]
References
Branch, G.M., 1976. Interspecific competition experienced by South African Patella species. J. Anim. Ecol. 45,
507–529.
Branch, G.M., 1981. The biology of limpets: physical factors, energy flow, and ecological interactions. Oceanogr.
Mar. Biol. Ann. Rev. 19, 235–380.
Choat, J.H., 1977. The influence of sessile organisms on the population biology of three species of acmaeid
limpets. J. Exp. Mar. Biol. Ecol. 26, 1–26.
Choat, J.H., Black, R., 1979. Life histories of limpets and the limpet– laminarian relationship. J. Exp. Mar. Biol.
Ecol. 41, 25–50.
Creese, R.G., 1982. Distribution and abundance of the acmaeid limpet, Patelloida latistrigata, and its interaction
with barnacles. Oecologia 52, 85–96.
Creese, R.G., 1988. Ecology of molluscan grazers and their interactions with marine algae in north-eastern New
Zealand: a review. N.Z. J. Mar. Freshw. Res. 22, 427–494.
Creese, R.G., Underwood, A.J., 1982. Analysis of inter- and intra-specific competition amongst intertidal limpets
with different methods of feeding. Oecologia 53, 337–346.
Dunmore, R.A., Schiel, D.R., 2000. Reproduction in the intertidal limpet Cellana ornata in Southern New
Zealand. N.Z. J. Mar. Freshw. Res. 34, 653–660.
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–38 37

Farrell, T.M., 1991. Models and mechanisms of succession: an example from a rocky intertidal community. Ecol.
Monogr. 61 (1), 95–113.
Fletcher, W.J., Creese, R.G., 1985. Competitive interactions between co-occurring herbivorous gastropods. Mar.
Biol. 86, 183–191.
Geller, J.B., 1991. Gastropod grazers and algal colonization on a rocky shore in northern California: the
importance of the body size of grazers. J. Exp. Mar. Biol. Ecol. 150, 1–17.
Hawkins, S.J., 1983. Interactions of Patella and macroalgae with settling Semibalanus balanoides (L.). J. Exp.
Mar. Biol. Ecol. 71, 55–72.
Hawkins, S.J., Hartnoll, R.G., 1982. The influence of barnacle cover on the numbers, growth and behaviour of
Patella vulgata on a vertical pier. J. Mar. Biol. Assoc. U.K. 62, 855–867.
Hawkins, S.J., Hartnoll, R.G., 1983. Grazing of intertidal algae by marine invertebrates. Oceanogr. Mar. Biol.
Ann. Rev. 21, 195–282.
Hill, A.S., Hawkins, S.J., 1990. An investigation of methods for sampling microbial films on rocky shores. J.
Mar. Biol. Assoc. U.K. 70, 77–88.
HMSO, 1986. The determination of chlorophyll a in aquatic environments, vol. 4. Methods for the Examination
of Water and Associated Materials, Section 2. HMSO, London.
Johnson, M.P., Burrows, M.T., Hartnoll, R.G., Hawkins, S.J., 1997. Spatial structure on moderately exposed
rocky shores: patch scales and the interactions between limpets and algae. Mar. Ecol., Prog. Ser. 160,
209–215.
Lewis, J.R., Bowman, R.S., 1975. Local habitat-induced variations in the population dynamics of Patella vulgata
L. J. Exp. Mar. Biol. Ecol. 17, 165–203.
Lubchenco, J., Gaines, S., 1981. A unified approach to marine plant–animal interactions: I. Populations and
communities. Ann. Rev. Ecolog. Syst. 12, 405–436.
Marshall, P.A., Keough, M.J., 1994. Asymmetry in intraspecific competition in the limpet Cellana tramoserica
(Sowerby). J. Exp. Mar. Biol. Ecol. 177, 121–138.
Morton, J., Miller, M.C., 1973. The New Zealand Sea Shore, 2nd ed. Collins, Auckland.
Powell, A.W.B., 1979. New Zealand Mollusca: Marine, Land and Freshwater Shells. Collins, Auckland.
Raffaelli, D., 1979. The grazer–algae interaction in the intertidal zone on New Zealand rocky shores. J. Exp.
Mar. Biol. Ecol. 38, 81–199.
Raffaelli, D., Hawkins, S., 1996. Community dynamics. In: Raffaelli, D., Hawkins, S. (Eds.), Intertidal Ecology.
Chapman & Hall, London, pp. 98–125.
Smith, S.L., 1969. Distribution and abundance of eulittoral grazing molluscs on rocky shores. Unpub. MSc
Thesis, University of Auckland.
Smith, S.L., 1975. Physiological ecology of the limpet Cellana ornata (Dillwyn). N.Z. J. Mar. Freshw. Res. 9 (3),
395–402.
Thompson, G.B., 1980. Distribution and population dynamics of the limpet Patella vulgata in Bantry Bay. J.
Exp. Mar. Biol. Ecol. 45, 173–217.
Underwood, A.J., 1978. An experimental evaluation of competition between three species of intertidal proso-
branch gastropods. Oecologia 33, 185–202.
Underwood, A.J., 1979. The ecology of intertidal gastropods. Adv. Mar. Biol. 16, 111–210.
Underwood, A.J., Jernakoff, P., 1981. Effects of interactions between algae and grazing gastropods on the
structure of low-shore intertidal algal community. Oecologia 48, 221–233.
Underwood, A.J., Kennelly, S.J., 1990. Ecology of marine algae on rocky shores and subtidal reefs in temperate
Australia. Hydrobiology 192, 3–20.
Underwood, A.J., Denley, E.J., Moran, M.J., 1983. Experimental analyses of the structure and dynamics of mid-
shore rocky intertidal communities in New South Wales. Oecologia 56, 202–219.
Walters, T.L., 1994. Comparative life histories of two species of prosobranch limpets, Cellana flava and C.
denticulata. Unpublished MSc thesis, University of Canterbury, New Zealand.
R.A. Dunmore, D.R. Schiel / J. Exp. Mar. Biol. Ecol. 288 (2003) 17–3838