proteomic characterization of alternative splicing and coding polymorphism nathan edwards center for...
TRANSCRIPT

Proteomic Characterization
of Alternative Splicing and
Coding Polymorphism
Proteomic Characterization
of Alternative Splicing and
Coding PolymorphismNathan EdwardsCenter for Bioinformatics and Computational BiologyUniversity of Maryland, College Park

2
Mass Spectrometry for Proteomics
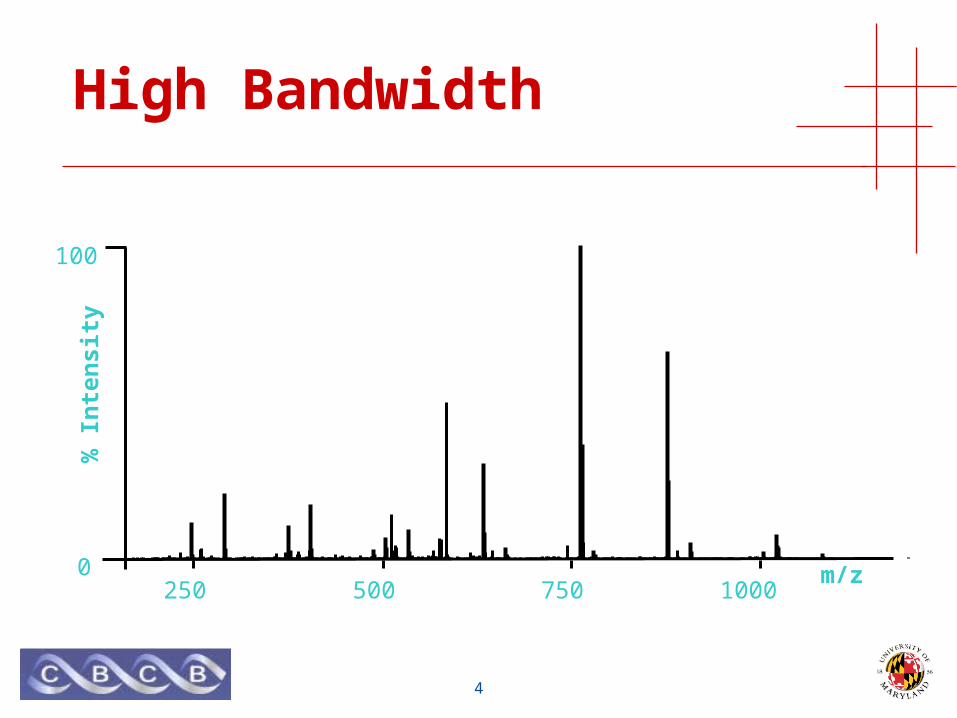
• Measure mass of many (bio)molecules simultaneously• High bandwidth
• Mass is an intrinsic property of all (bio)molecules• No prior knowledge required

3
Mass Spectrometry for Proteomics
• Measure mass of many molecules simultaneously• ...but not too many, abundance bias
• Mass is an intrinsic property of all (bio)molecules• ...but need a reference to compare to
4
High Bandwidth
100
0250 500 750 1000
m/z
% I
nte
nsit
y

5
Mass is fundamental!

6
Mass Spectrometry for Proteomics
• Mass spectrometry has been around since the turn of the century...• ...why is MS based Proteomics so new?
• Ionization methods• MALDI, Electrospray
• Protein chemistry & automation• Chromatography, Gels, Computers
• Protein sequence databases• A reference for comparison
7
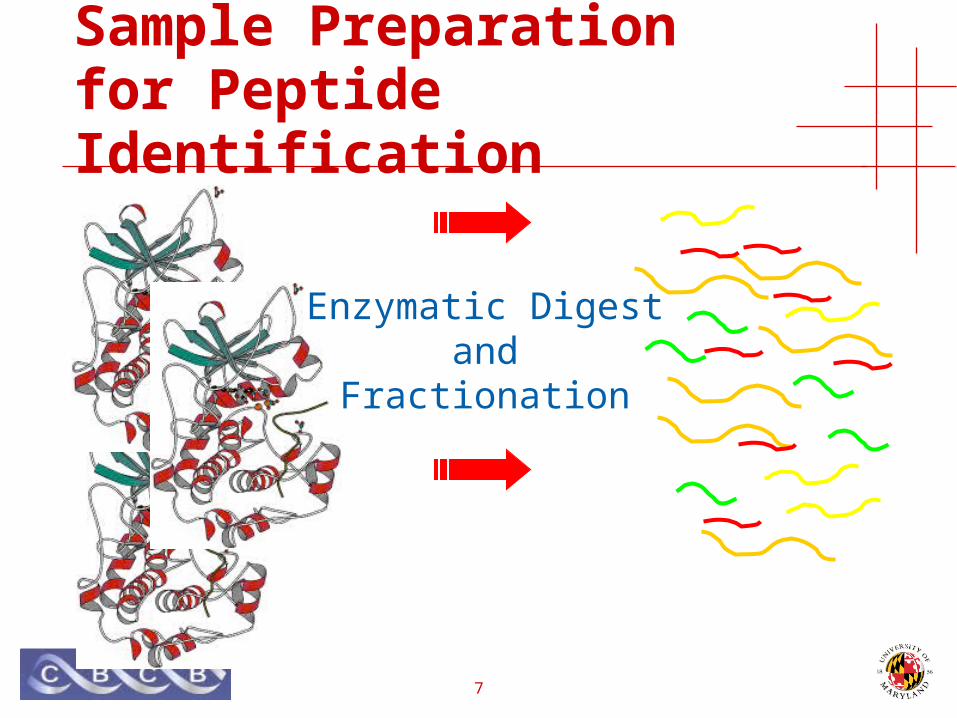
Sample Preparation for Peptide Identification
Enzymatic Digestand
Fractionation
8
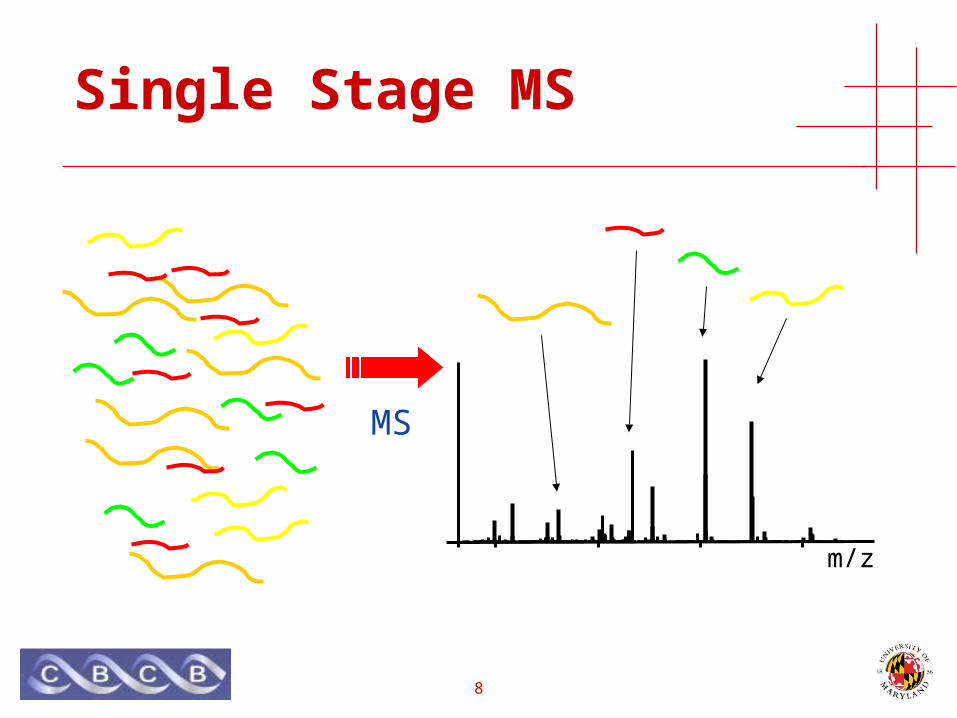
Single Stage MS
MS
m/z
9
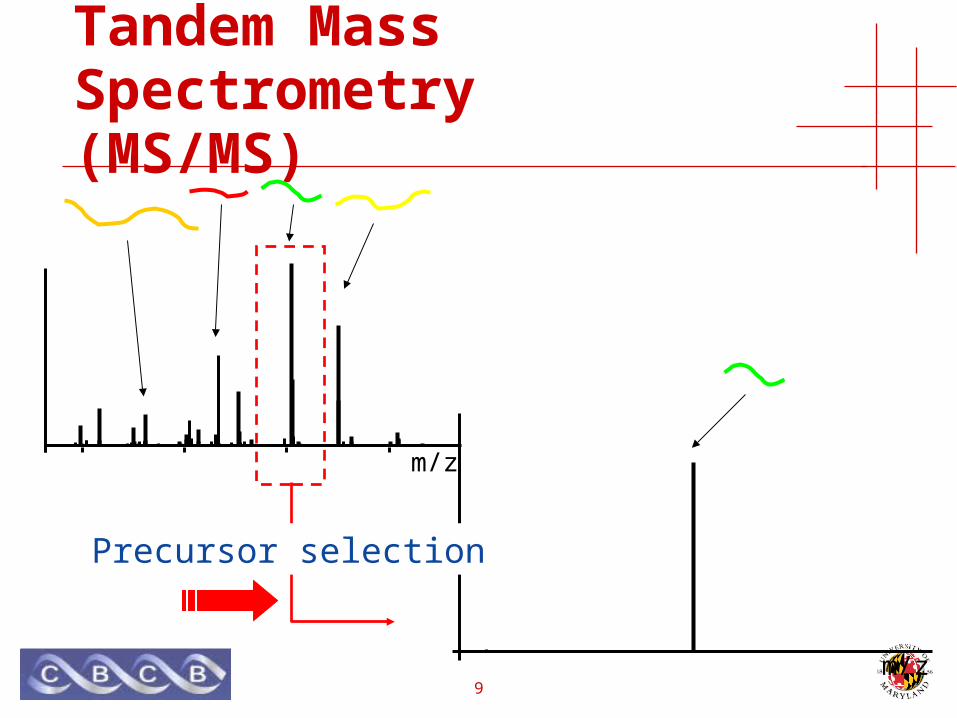
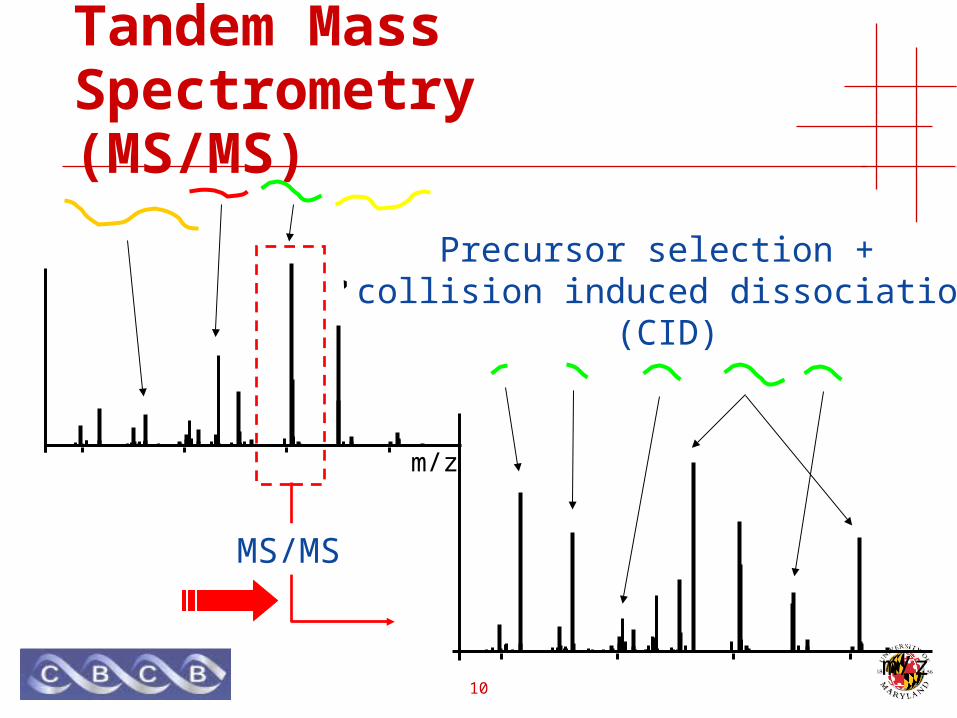
Tandem Mass Spectrometry(MS/MS)
Precursor selection
m/z
m/z
10
Tandem Mass Spectrometry(MS/MS)
Precursor selection + collision induced dissociation
(CID)
MS/MS
m/z
m/z

11
Peptide Identification
• For each (likely) peptide sequence1. Compute fragment masses2. Compare with spectrum3. Retain those that match well
• Peptide sequences from protein sequence databases• Swiss-Prot, IPI, NCBI’s nr, ...
• Automated, high-throughput peptide identification in complex mixtures

12
Why don’t we see more novel peptides?
• Tandem mass spectrometry doesn’t discriminate against novel peptides...
...but protein sequence databases do!
• Searching traditional protein sequence databases biases the results towards well-understood protein isoforms!

13
What goes missing?
• Known coding SNPs
• Novel coding mutations
• Alternative splicing isoforms
• Alternative translation start-sites
• Microexons
• Alternative translation frames

14
Why should we care?
• Alternative splicing is the norm!• Only 20-25K human genes• Each gene makes many proteins
• Proteins have clinical implications• Biomarker discovery
• Evidence for SNPs and alternative splicing stops with transcription• Genomic assays, ESTs, mRNA sequence.• Little hard evidence for translation start site

15
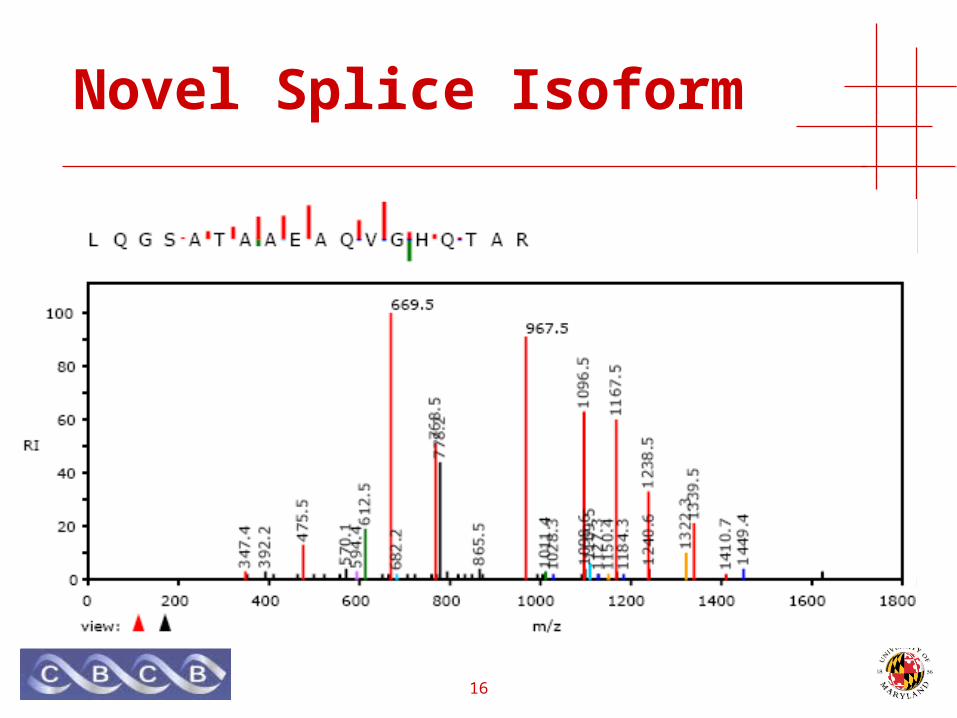
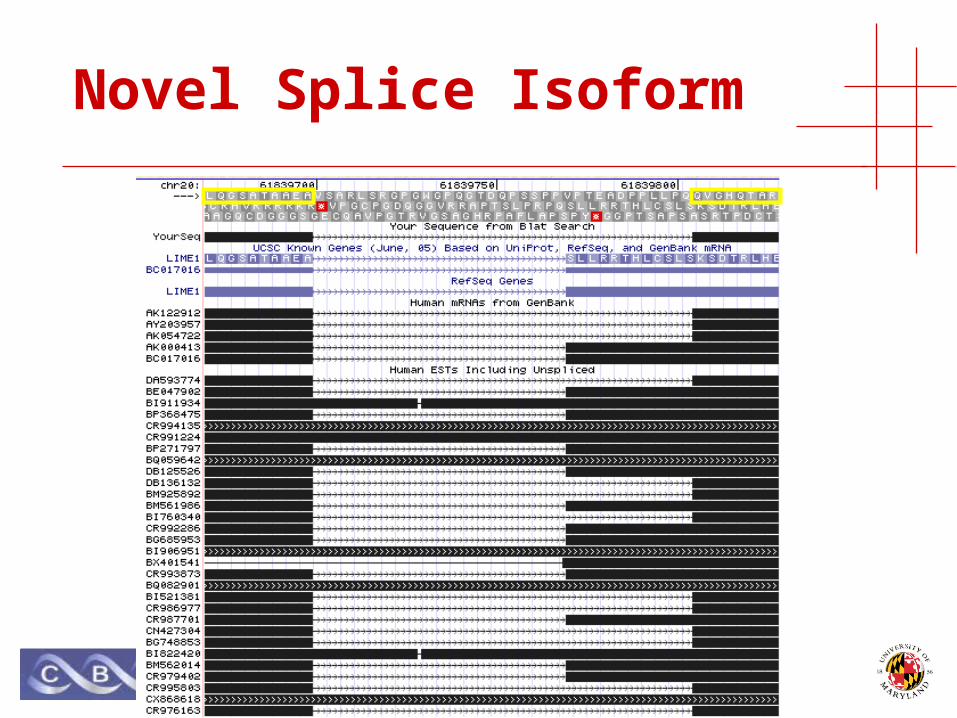
Novel Splice Isoform
• Human Jurkat leukemia cell-line• Lipid-raft extraction protocol, targeting T cells• von Haller, et al. MCP 2003.
• LIME1 gene:• LCK interacting transmembrane adaptor 1
• LCK gene:• Leukocyte-specific protein tyrosine kinase• Proto-oncogene• Chromosomal aberration involving LCK in leukemias.
• Multiple significant peptide identifications
16
Novel Splice Isoform
17
Novel Splice Isoform
18
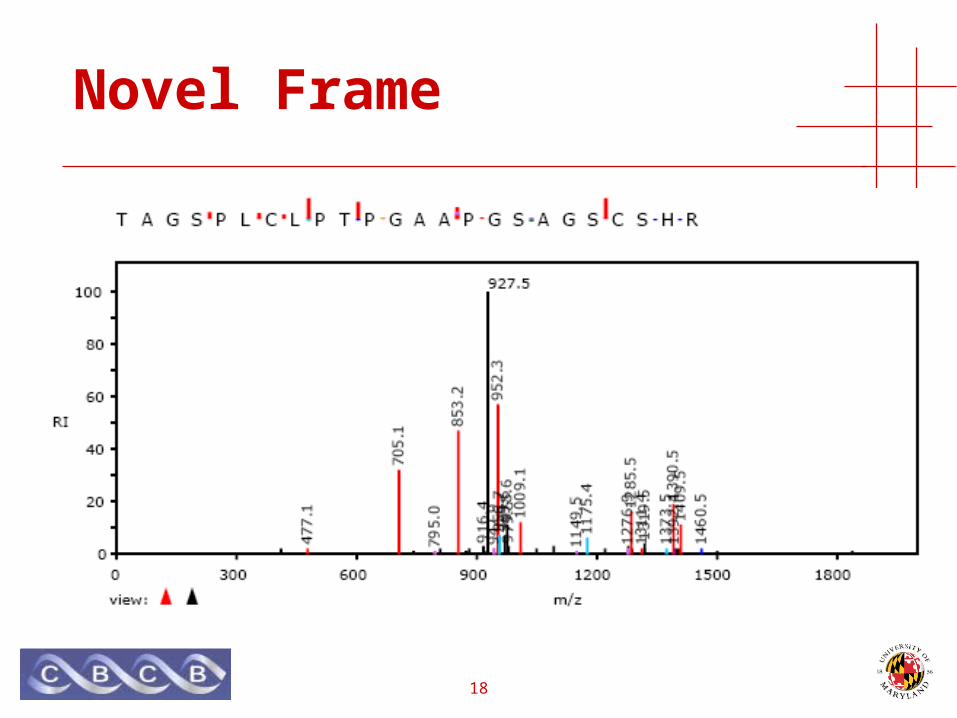
Novel Frame

19
Novel Frame

20
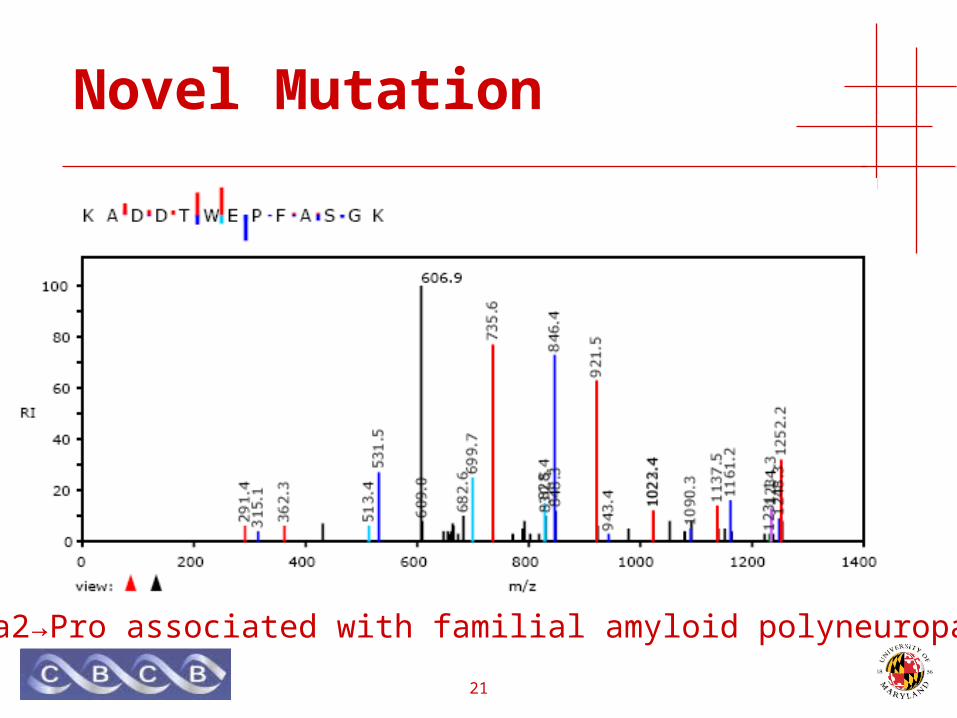
Novel Mutation
• HUPO Plasma Proteome Project• Pooled samples from 10 male & 10 female
healthy Chinese subjects• Plasma/EDTA sample protocol• Li, et al. Proteomics 2005. (Lab 29)
• TTR gene• Transthyretin (pre-albumin) • Defects in TTR are a cause of amyloidosis.• Familial amyloidotic polyneuropathy
• late-onset, dominant inheritance
21
Novel Mutation
Ala2→Pro associated with familial amyloid polyneuropathy

22
Novel Mutation

23
Searching ESTs
• Proposed long ago:• Yates, Eng, and McCormack; Anal Chem, ’95.
• Now:• Protein sequences are sufficient for protein identification• Computationally expensive/infeasible• Difficult to interpret
• Make EST searching feasible for routine searching to discover novel peptides.

24
Searching Expressed Sequence Tags (ESTs)
Pros• No introns!• Primary splicing
evidence for annotation pipelines
• Evidence for dbSNP• Often derived from
clinical cancer samples
Cons• No frame• Large (8Gb)• “Untrusted” by
annotation pipelines• Highly redundant• Nucleotide error
rate ~ 1%

25
Compressed EST Peptide Sequence Database
• For all ESTs mapped to a UniGene gene:• Six-frame translation• Eliminate ORFs < 30 amino-acids• Eliminate amino-acid 30-mers observed once• Compress to C2 FASTA database
• Complete, Correct for amino-acid 30-mers
• Gene-centric peptide sequence database:• Size: < 3% of naïve enumeration, 20774 FASTA entries• Running time: ~ 1% of naïve enumeration search• E-values: ~ 2% of naïve enumeration search results

26
Compressed EST Peptide Sequence Database
• For all ESTs mapped to a UniGene gene:• Six-frame translation• Eliminate ORFs < 30 amino-acids• Eliminate amino-acid 30-mers observed once• Compress to C2 FASTA database
• Complete, Correct for amino-acid 30-mers
• Gene-centric peptide sequence database:• Size: < 3% of naïve enumeration, 20774 FASTA entries• Running time: ~ 1% of naïve enumeration search• E-values: ~ 2% of naïve enumeration search results
27
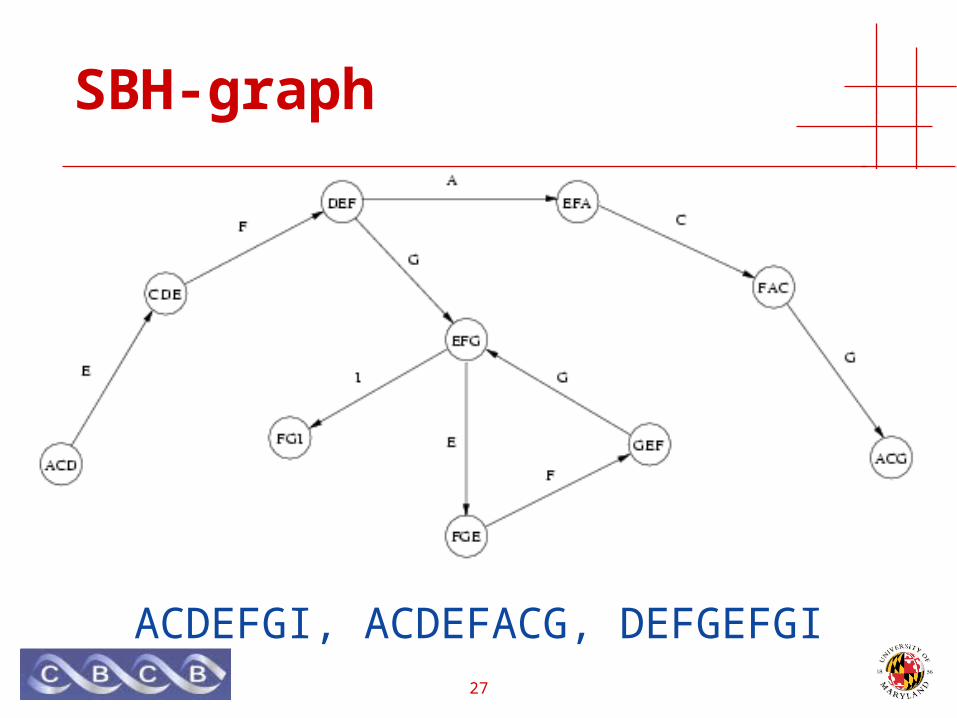
SBH-graph
ACDEFGI, ACDEFACG, DEFGEFGI
28
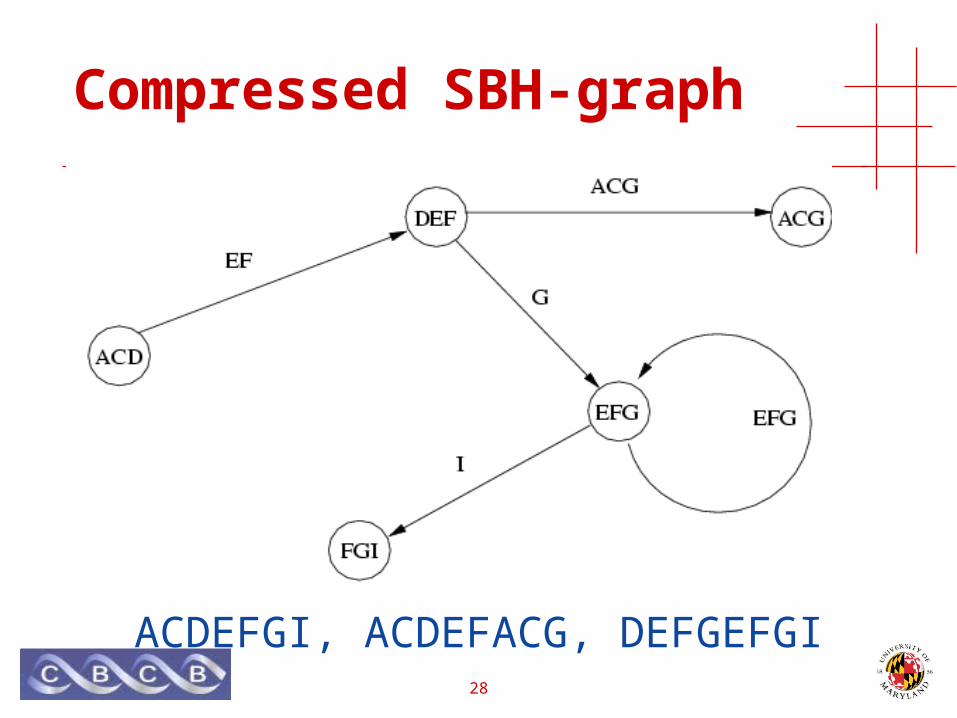
Compressed SBH-graph
ACDEFGI, ACDEFACG, DEFGEFGI
29
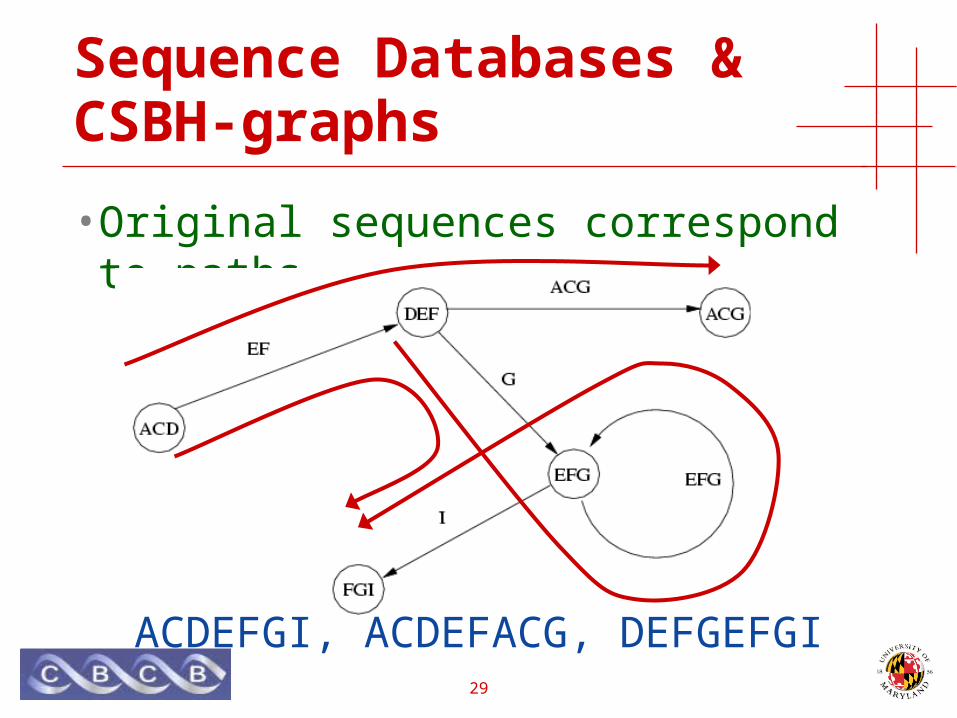
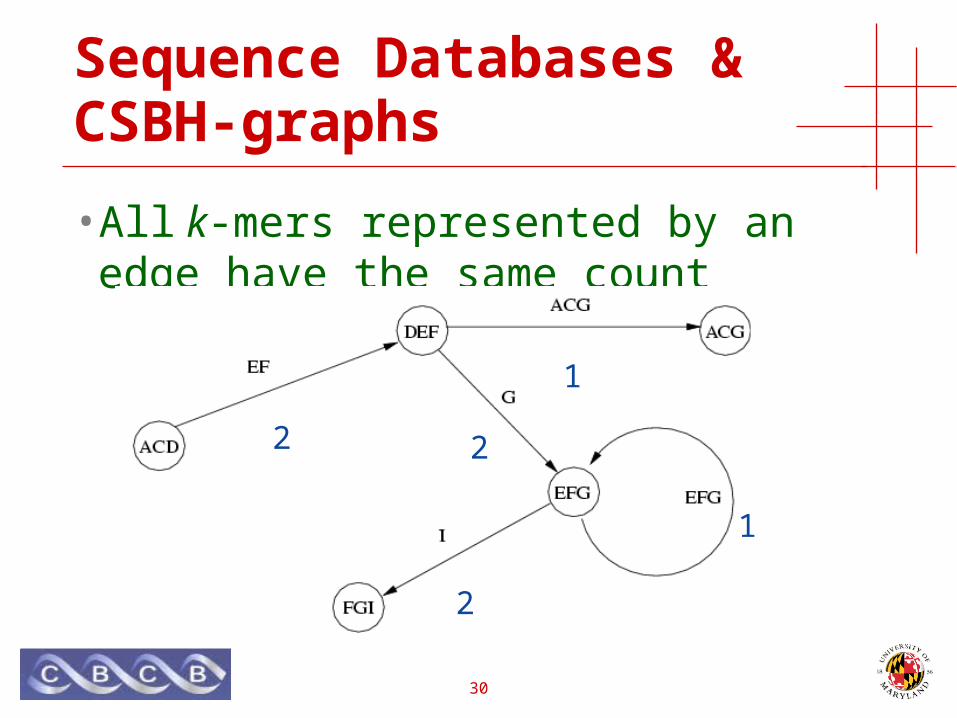
Sequence Databases & CSBH-graphs
• Original sequences correspond to paths
ACDEFGI, ACDEFACG, DEFGEFGI
30
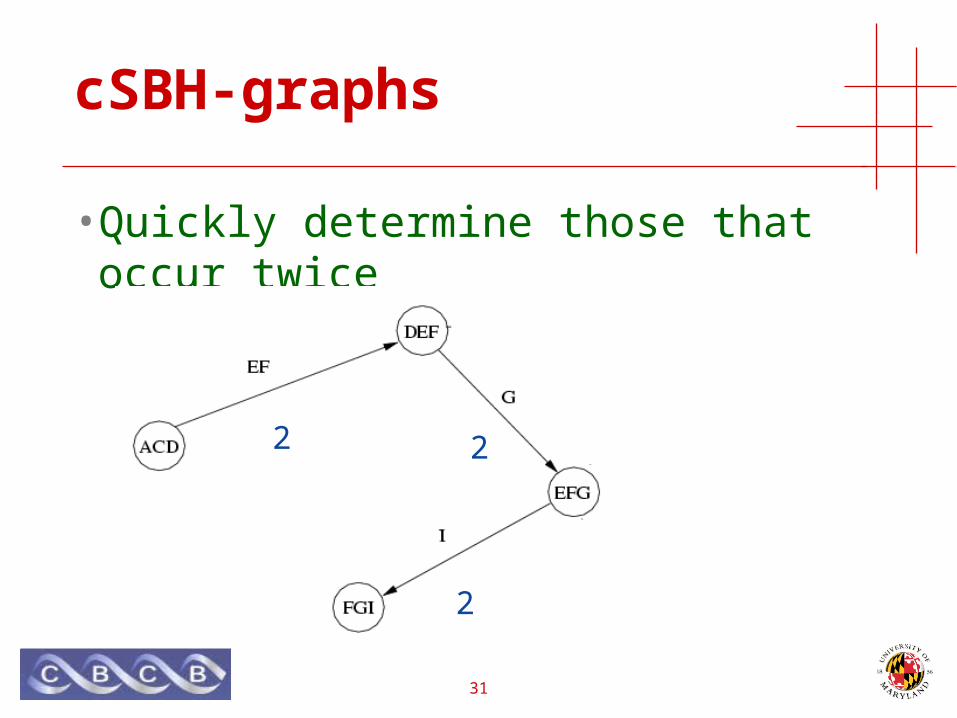
Sequence Databases & CSBH-graphs
• All k-mers represented by an edge have the same count
2 2
1
2
1
31
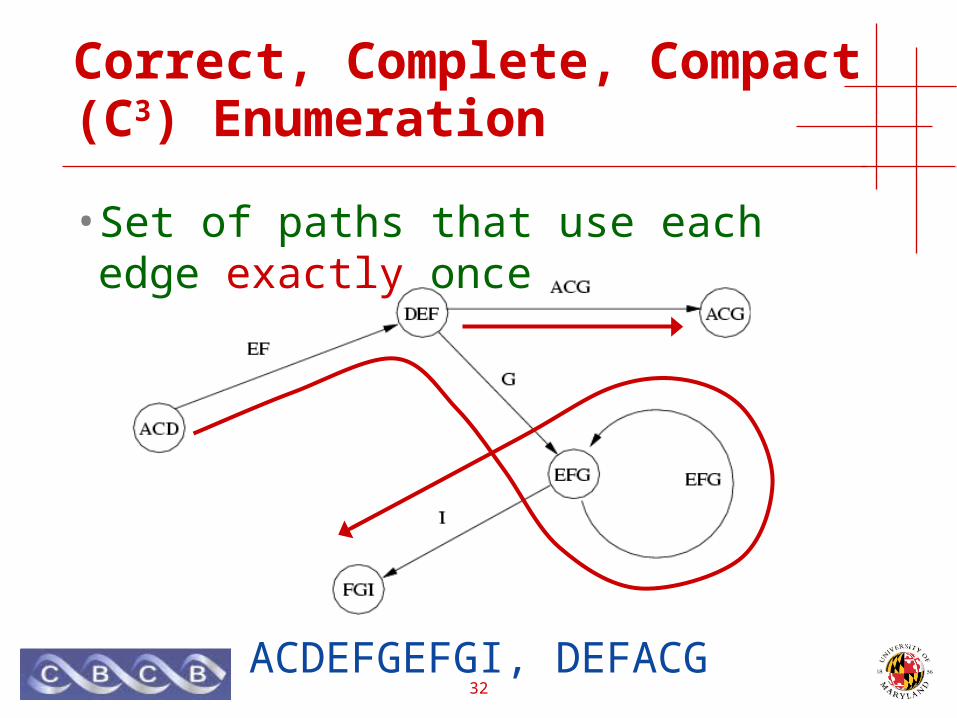
cSBH-graphs
• Quickly determine those that occur twice
2 2
1
2
32
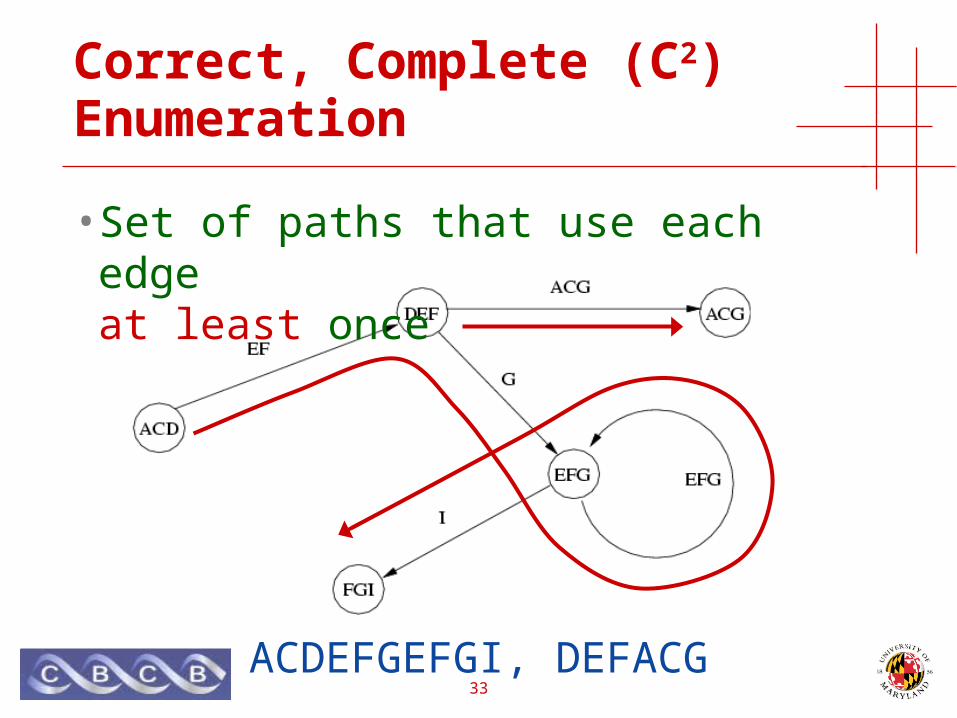
Correct, Complete, Compact (C3) Enumeration
• Set of paths that use each edge exactly once
ACDEFGEFGI, DEFACG
33
Correct, Complete (C2) Enumeration
• Set of paths that use each edge at least once
ACDEFGEFGI, DEFACG

34
Patching the CSBH-graph
• Use artificial edges to fix unbalanced nodes

35
Compressed EST Database
• Gene centric compressed EST peptide sequence database• 20,774 sequence entries• ~8Gb vs 223 Mb• ~35 fold compression
• 22 hours becomes 15 minutes• E-values improve by similar factor!
• Makes routine EST searching feasible• Search ESTs instead of IPI?

36
“Novel Peptide” Computational Infrastructure
• Binaries (C++)• cSBH-graph construction
• Condor grid-enabled• Eulerian path k-mer enumeration
• Suitable for large graphs
• Data-model for peptide identification• Spectra (>5 million)• Peptide identifications
• Mascot, SEQUEST, X!Tandem, NIST • Genomic context of peptides

37
“Novel Peptide” Computational Infrastructure
• Condor grid-enabled MS/MS search• Mascot, X!Tandem, (Inspect, OMSSA)
• TurboGears python web-stack• SQLObject Object-Relational-Manager• MVC web-application framework• Suitable for AJAX & web-services too
• Integration with UCSC genome browser• caBIG compatible web-services
• Java applet for viewing spectra
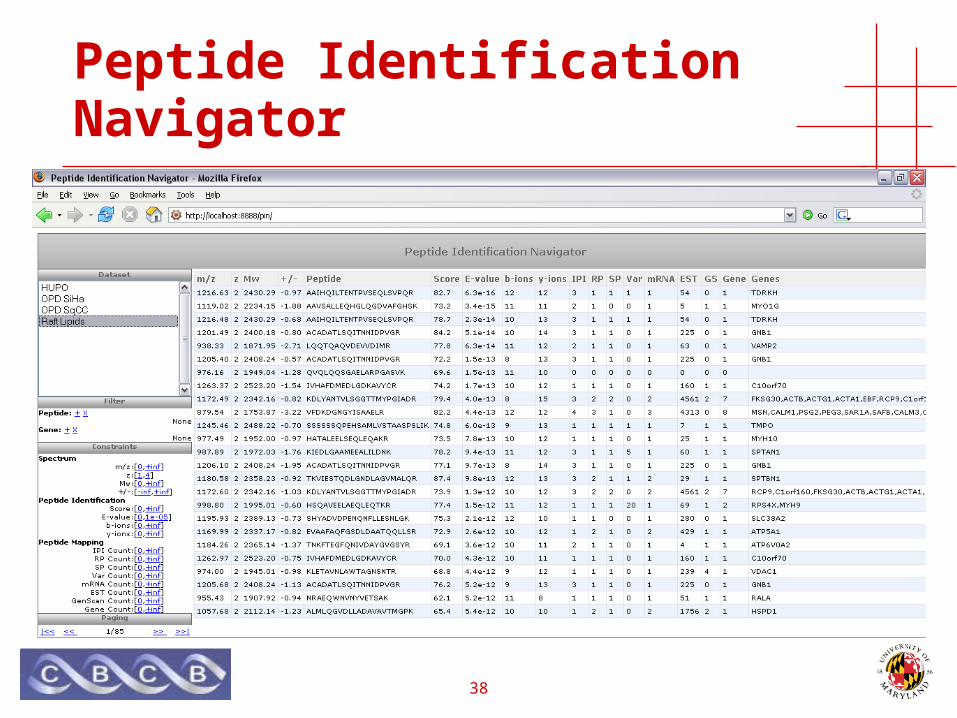
38
Peptide Identification Navigator
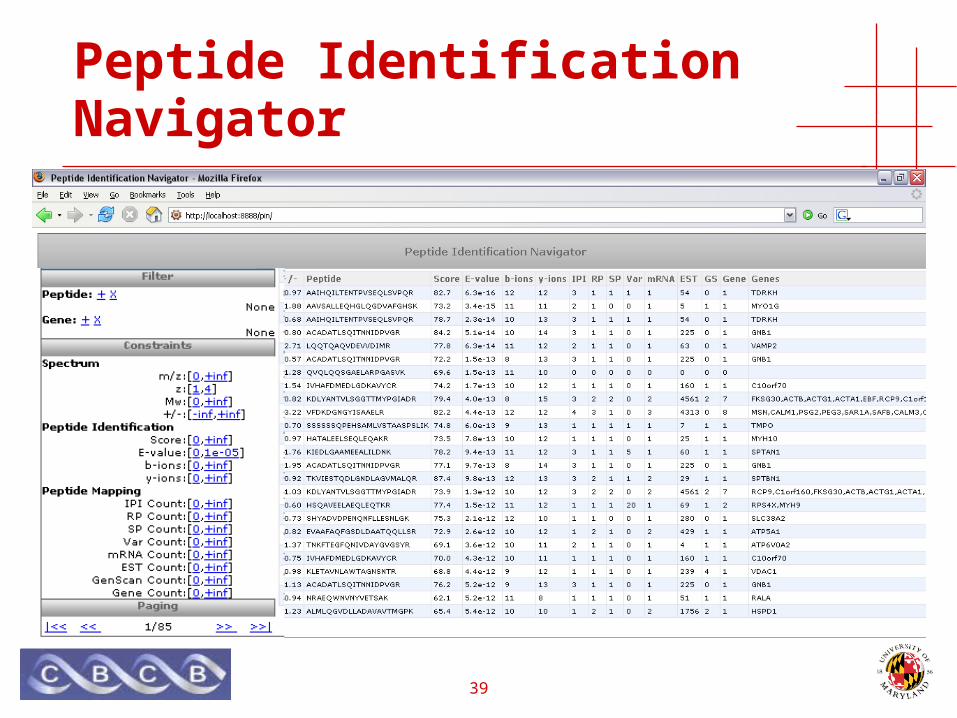
39
Peptide Identification Navigator
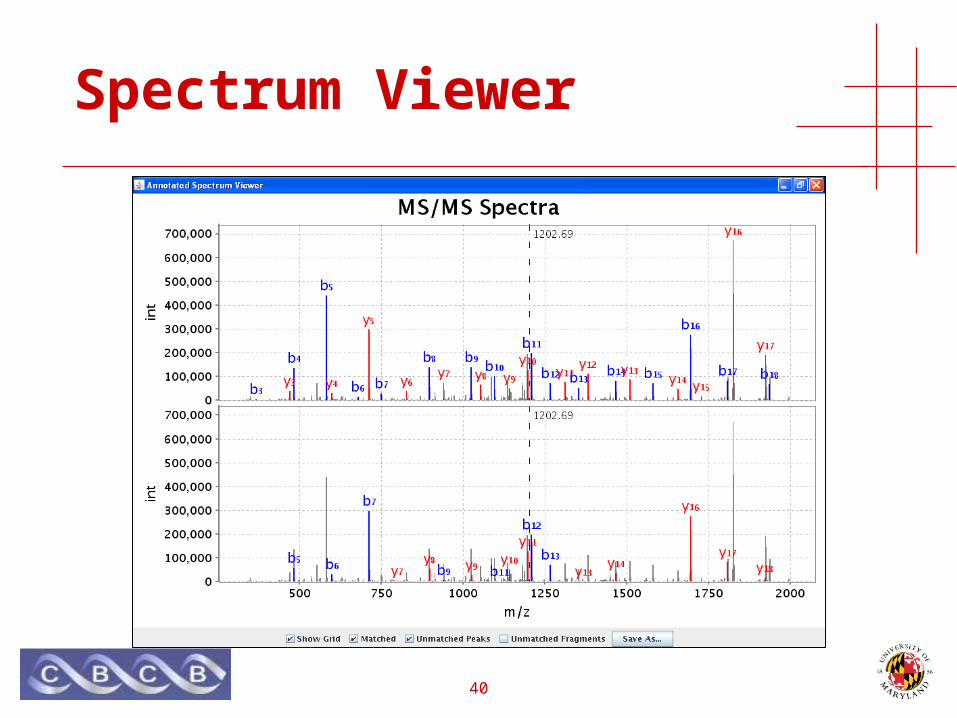
40
Spectrum Viewer
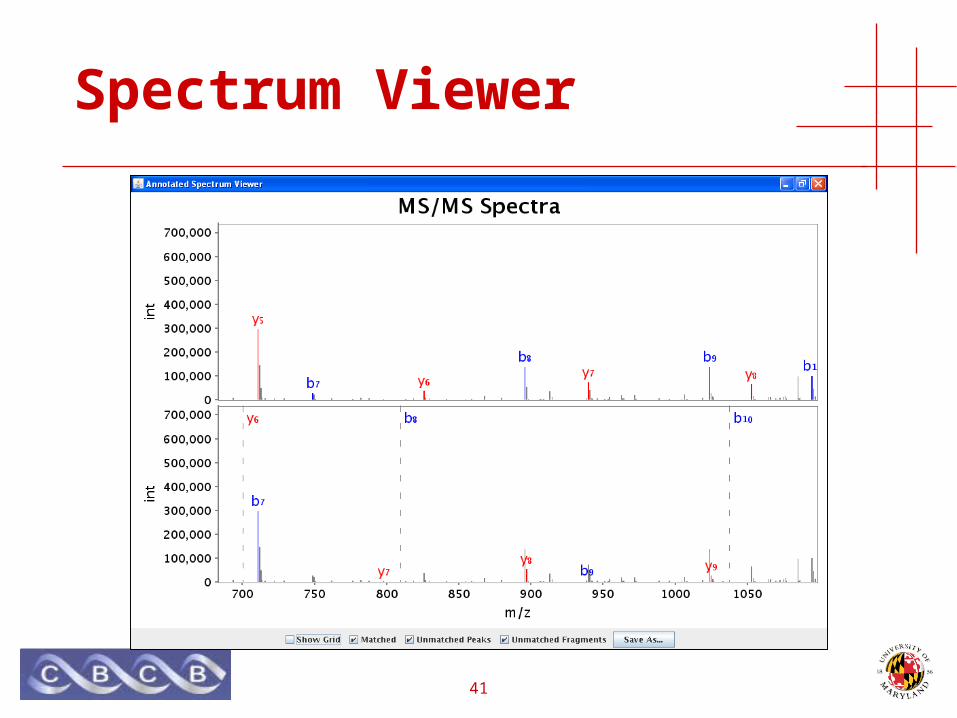
41
Spectrum Viewer

42
Back to the lab...
• Current LC/MS/MS workflows identify a few peptides per protein• ...not sufficient for protein isoforms
• Need to raise the sequence coverage to (say) 80%• ...protein separation prior to LC/MS/MS
analysis• Potential for database of splice sites of
(functional) proteins!
43
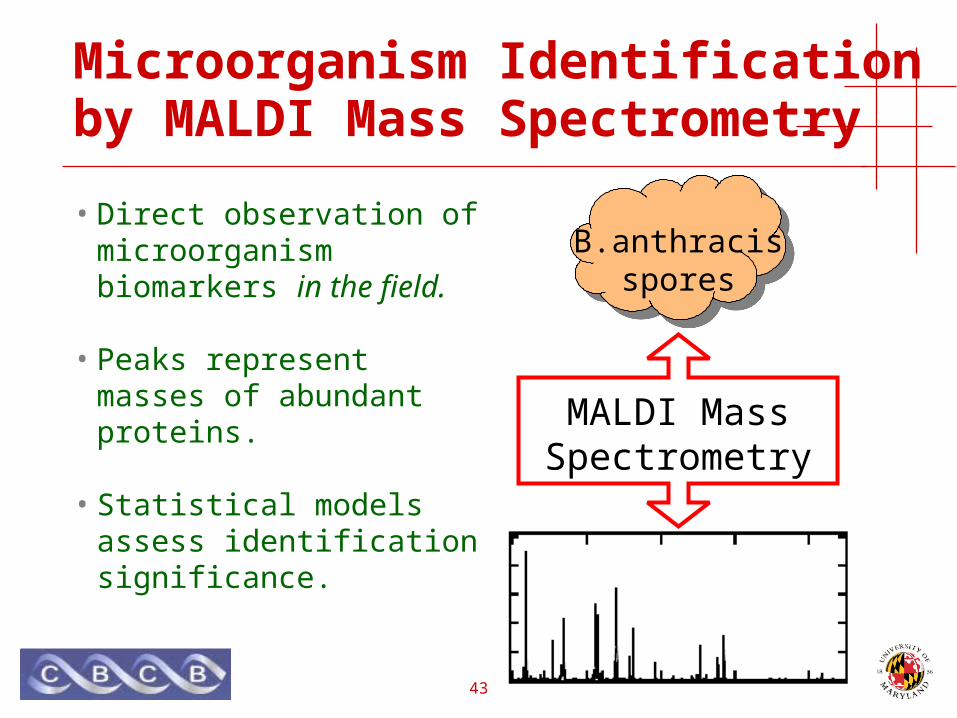
Microorganism Identification by MALDI Mass Spectrometry
• Direct observation of microorganism biomarkers in the field.
• Peaks represent masses of abundant proteins.
• Statistical models assess identification significance.
B.anthracisspores
MALDI Mass Spectrometry

44
Key Principles
• Protein mass from protein sequence• No introns, few PTMs
• Specificity of single mass is very weak• Statistical significance from many peaks
• Not all proteins are equally likely to be observed• Ribosomal proteins, SASPs
45
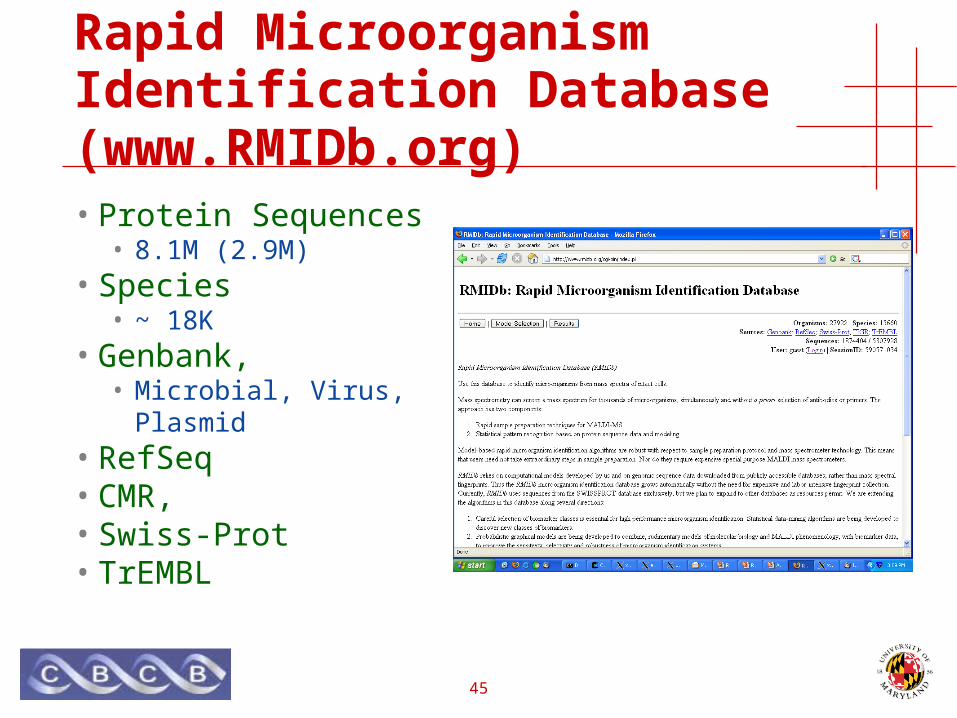
Rapid Microorganism Identification Database (www.RMIDb.org)
• Protein Sequences• 8.1M (2.9M)
• Species• ~ 18K
• Genbank,• Microbial, Virus, Plasmid
• RefSeq• CMR,• Swiss-Prot• TrEMBL
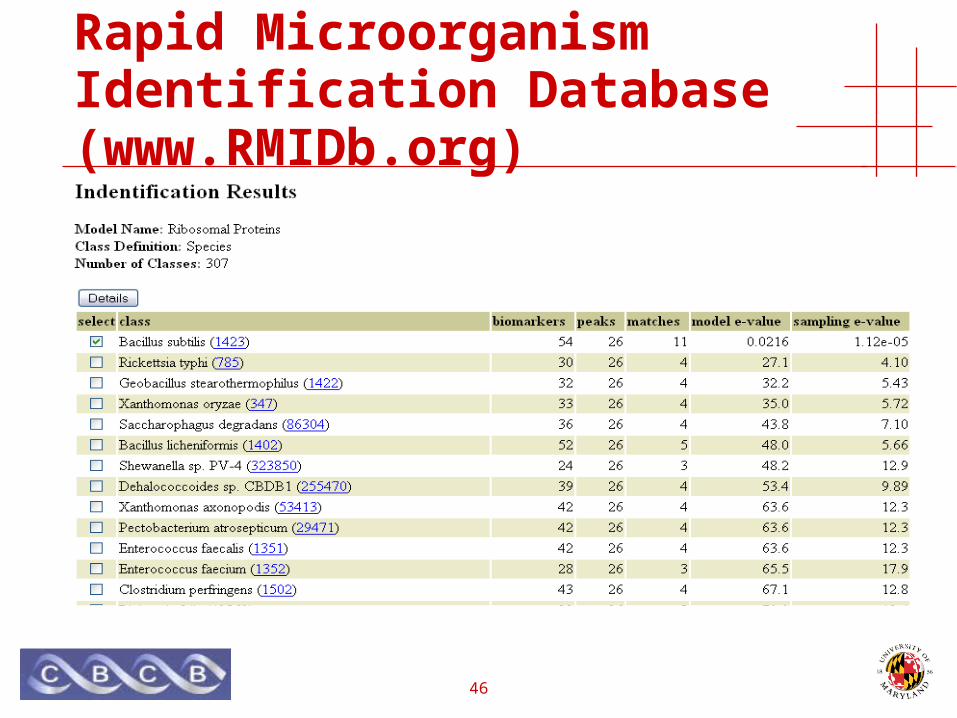
46
Rapid Microorganism Identification Database (www.RMIDb.org)

47
Informatics Issues
• Need good species / strain annotation• B.anthracis vs B.thuringiensis
• Need correct protein sequence• B.anthracis Sterne α/β SASP• RefSeq/Gb: MVMARN... (7442 Da)• CMR: MARN... (7211 Da)
• Need chemistry based protein classification

48
Conclusions
• Proteomics can inform genome annotation• Eukaryotic and prokaryotic • Functional vs silencing variants
• Peptides identify more than just proteins• Untapped source of disease biomarkers
• Compressed peptide sequence databases make routine EST searching feasible

49
Future Research Directions
• Identification of protein isoforms:• Optimize proteomics workflow for isoform
detection• Identify splice variants in cancer cell-lines
(MCF-7) and clinical brain tumor samples• Aggressive peptide sequence enumeration• dbPep for genomic annotation• Open, flexible informatics infrastructure for
peptide identification

50
Future Research Directions
• Proteomics for Microorganism Identification• Specificity of tandem mass spectra• Revamp RMIDb prototype• Incorporate spectral matching
• Primer design• k-mer sets as FASTA sequence databases• Uniqueness oracle for exact and inexact match• Integration with Primer3• Tiling, multiplexing, pooling, & tag arrays

51
Acknowledgements
• Chau-Wen Tseng, Xue Wu• UMCP Computer Science
• Catherine Fenselau, Steve Swatkoski• UMCP Biochemistry
• Calibrant Biosystems
• PeptideAtlas, HUPO PPP, X!Tandem
• Funding: National Cancer Institute