production of β-carotene and acetate in recombinant...
TRANSCRIPT

Biotechnology and Bioprocess Engineering 17: 1196-1204 (2012)
DOI 10.1007/s12257-012-0272-2
Production of β-carotene and Acetate in Recombinant Escherichia
coli with or without Mevalonate Pathway at Different Culture
Temperature or pH
Anh Do Quynh Nguyen, Seon-Won Kim, Sung Bae Kim, Yang-Gon Seo, In-Young Chung, Dae Hwan Kim, and
Chang-Joon Kim
Received: 24 April 2012 / Revised: 21 June 2012 / Accepted: 30 June 2012
© The Korean Society for Biotechnology and Bioengineering and Springer 2012
Abstract Natural β-carotene has received much attention
as consumers have become more health conscious. Its
production by various microorganisms including metabolically
engineered Escherichia coli or Saccharomyces cerevisiae
has been attempted. We successfully created a recombinant
E. coli with an engineered whole mevalonate pathway in
addition to β-carotene biosynthetic genes and evaluated the
engineered cells from the aspects of metabolic balance
between central metabolism and β-carotene production
by comparison with conventional β-carotene producing
recombinant E. coli (control) utilizing a native methyl-
erythritol phosphate (MEP) pathway using bioreactor
cultures generated at different temperatures or pHs. Better
production of β-carotene was obtained in E. coli cultured at
37oC than at 25oC. A two-fold higher titer and 2.9-fold
higher volumetric productivity were obtained in engineered
cells compared with control cells. Notably, a marginal amount
of acetate was produced in actively growing engineered
cells, whereas more than 8 g/L of acetate was produced in
control cells with reduced cell growth at 37oC. The data
indicated that the artificial operon of the whole mevalonate
pathway operated efficiently in redirecting acetyl-CoA into
isopentenyl pyrophosphate (IPP), thereby improving
production of β-carotene, whereas the native MEP pathway
did not convert a sufficient amount of pyruvate into IPP
due to endogenous feedback regulation. Engineered cells
also produced lycopene with a reduced amount of β-
carotene in weak alkaline cultures, consistent with the
inhibition of lycopene cyclase.
Keywords: recombinant Escherichia coli, engineered
whole mevalonate pathway, bioreactor culture, β-carotene,
acetate
1. Introduction
β-carotene functions as provitamin A and plays an important
role as a pharmaceutical, nutraceutical, animal feed additive,
compound in cosmetic formulations and food colorants [1].
Recently, natural β-carotene has received much attention
because a chemically synthesized version cannot meet the
increased consumer demand for health and well-being.
Traditionally, natural β-carotene is obtained by extraction
from plants or vegetables. However, the natural β-carotene
yield is extremely low because it is present only in trace
amounts and because its extraction recovery is low due to
the rigid cell-wall structure of plants and vegetables [2].
Furthermore, β-carotene production varies seasonally and
geographically. To overcome these yield problems, microbial
production of β-carotene has been tried using green alga
(Dunaliella salina), fungi (Blakeslea trispora and Rhodotorula
Anh Do Quynh Nguyen, Sung Bae Kim, Yang-Gon Seo, Chang-Joon Kim*
Department of Chemical & Biological Engineering and ERI, GyeongsangNational University, Jinju 660-701, KoreaTel: +82-55-772-1787; Fax: +82-55-772-1789E-mail: [email protected]
Seon-Won KimDivision of Applied Life Science (BK21), PMBBRC, GyeongsangNational University, Jinju 660-701, Korea
In-Young ChungDepartment of Electronic and Communications Engineering, KwangwoonUniversity, Seoul 139-701, Korea
Dae Hwan KimSchool of Electrical Engineering, Kookmin University, Seoul 136-702,Korea
RESEARCH PAPER

Production of β-carotene and Acetate in Recombinant Escherichia coli with or without Mevalonate Pathway at… 1197
glutinis), and metabolically engineered yeast (Saccharomyces
cerevisiae) and bacteria (Escherichia coli) [3-8]. Recently,
we successfully created a recombinant E. coli with an
engineered whole mevalonate (MVA) pathway in addition
to β-carotene biosynthetic genes. This recombinant
bacterium produced a higher amount of β-carotene than the
conventional β-carotene producing recombinant E. coli
utilizing only the native methyl-erythritol phosphate
(MEP) pathway [9]. Production was further enhanced by
optimizing the feeding medium in fed-batch cultures of the
engineered E. coli [10]. As will be subsequently described
in detail, a big difference between these novel and
conventional cells concerns the biosynthesis of isopentenyl
pyrophosphate (IPP), a key intermediate metabolite for β-
carotene biosynthesis. The former cells were engineered to
maximize the biosynthesis of IPP by the expressed foreign
mevalonate (MVA) pathway as well as the native MEP
pathway, whereas conventional E. coli utilizes only the
native MEP pathway. The engineered cells utilize acetyl-
CoA in addition to pyruvate and glyceraldehyde-3-phosphate
(G3P), whereas conventional cells utilize pyruvate and G3P
for IPP biosynthesis [11]. The aforementioned metabolites
are glycolytic intermediates that are also used for cell
growth. In particular, pyruvate is a key component that
serves as the major switching point between respiratory
and fermentative metabolism. Intracellular concentration of
pyruvate is elevated in actively growing E. coli in an
aerobic environment, in which carbon flux in the glycolytic
pathway exceeds the required level for the synthesis of
cellular constituents as the threshold growth rate is
approached. In turn, excess pyruvate promotes the formation
of acetate directly or via acetyl-CoA [11-14]. It is desirable
if excess pyruvate is utilized for IPP biosynthesis because
β-carotene production may be improved and acetate
formation may be minimized. However, it is questionable
whether the MEP pathway operates efficiently for the
utilization of pyruvate because it is under the feedback
regulated control in the native host [16]. On the other hand,
it is expected that expression of heterologous MVA pathway
genes results in the redirection of a large amount of excess
pyruvate to IPP via acetyl-CoA. However, excessive
draining of pyruvate may restrict the carbon flux or deplete
this resource necessary for cell growth. These metabolic
imbalances may lead to reduced cell growth and production
[17].
In this study, we aimed to evaluate the aforementioned
engineered E. coli concerning the metabolic balance
between central metabolism and β-carotene production
by comparison with conventional β-carotene producing
recombinant E. coli cells. Bioreactor cultures were generated
at growth-promoting or growth-retarding temperatures.
The engineered cells were also cultured at different culture
pH. Responses were examined by measuring cell growth
and acetate production as an indicator of change in the
central metabolism and β-carotene production. Cell growth,
acetate metabolism, and β-carotene production of these
cells showed different responses to culture temperature, in
which the engineered cells excreted a modest amount of
acetate without changed cell growth, even at the growth-
promoting temperature of 37oC. Notably, accumulation of
lycopene was observed in weak alkaline cultures, whereas
only β-carotene was produced at neutral or weak acidic
cultures. These physiological characteristics of the novel
cells have not been demonstrated previously.
2. Materials and Methods
2.1. Bacterial strains and culture conditions
Two kinds of β-carotene producing recombinant E. coli
cells were used. Control cells were E. coli DH5α containing
the plasmid pT-DHB, which harbors the genes for β-
carotene biosynthesis. These cells represent conventional
recombinant E. coli producing β-carotene. Plasmid pT-
DHB was constructed by cloning crtE, crtB, and crtI of
Pantoea agglomerans, ipiHP1 of Haematococcus pluvialis,
crtY of Pantoea ananatis, and dxs of E. coli into pTrc99A
(Amersham Bioscience, Piscataway, NJ, USA). Engineered
cells (strain 1) were novel to this study, which are E. coli
DH5α containing an additional plasmid pS-NA bearing
genes encoding the enzymes of the whole MVA pathway
beside plasmid pT-DHB. pS-NA was constructed by
cloning mvaE and mvaS from Enterococcus faecalis
ATCC14508; mvaK1, mvaK2, and mvaD from Streptococcus
pneumoniae; and idi from E. coli) into pSTV28 (TaKaRa
Bio, Shiga, Japan). The detailed procedures for construction
of the β-carotene producing cells are described elsewhere
[9]. The blank pSTV28 vector was also introduced into
control cells for comparative study.
2.2. Fermentation conditions
All flask cultures were performed in 500 mL baffled flasks
using a shaking incubator (Jeio Tech, Seoul, Republic of
Korea) at 200 rpm. Glycerol stocks of recombinant E. coli
were inoculated into 50 mL of Luria-Bertani (LB) broth
[18] supplemented with antibiotics (100 mg/L ampicillin,
50 mg/L chloramphenicol) and was grown at 37oC until
the optical density at 600 nm (OD600) reached 0.6. The
seed culture (2.5 mL) was then transferred to 50 mL of
2 × YT medium (16 g/L tryptone, 10 g/L yeast extract, and
5 g/L NaCl) supplemented with antibiotics (100 mg/L
ampicillin, 50 mg/L chloramphenicol), and 30 g/L glycerol
for the main culture.
For bioreactor cultures, 75 mL of seed culture was added

1198 Biotechnology and Bioprocess Engineering 17: 1196-1204 (2012)
to a 3.4 L fermentor (KoBiotech, Incheon, Korea) containing
1.5 L of 2 × YT medium supplemented with 30 g/L
glycerol as well as antibiotics. Cell growth and β-carotene
production were compared by performing bioreactor
fermentations at 37 and 25oC at neutral pH. To investigate
the sole pH effect, the pH was controlled to maintain at 6.8
± 0.1 (neutral pH), 5.9 ± 0.1 (weakly acidic pH), or 7.9 ±
0.1 (weakly alkaline pH) using 2 N H2SO4 and 25%
NH4OH. The dissolved oxygen level was maintained at
greater than 30% air saturation by the addition of pure
oxygen into the air stream and/or manually controlling the
agitation speed.
2.3. Analyses
The optical density at 600 nm (OD600) was measured by an
ultraviolet (UV)-Visible spectrophotometer (Hewlett-Packard,
Palo Alto, CA, USA) to monitor cell growth. The
corresponding dry-cell weight was determined by a
calibration curve relating OD600 to dry-cell weight (g/L).
The β-carotene content in the cells was also determined by
UV-Visible spectrophotometry; 1 mL of culture broth was
taken and appropriately diluted. An orange-colored cell pellet
was obtained by centrifugation at 9,800 × g for 10 min.
After washing once with distilled water, the cell pellet was
resuspended in 1 mL of acetone and incubated at 55oC for
15 min in the dark. During this process, all β-carotene
accumulated in cells was extracted into the acetone and the
resulting cell pellet obtained by centrifugation at 9,800 × g
for 10 min was nearly colorless. β-carotene in the acetone
extract was quantified by measuring the absorbance at
454 nm.
Analytical UV-Visible spectrophotometry is not capable
of determining β-carotene quantitatively in the presence of
lycopene due to interference [19,20]. Because both β-
carotene and lycopene were produced in strain1 cultured at
a weak alkaline pH, quantitative analysis of β-carotene or
lycopene in cell extracts was carried out using a high
performance liquid chromatography (HPLC) system consisting
of a model 515 HPLC pump and a model 486 UV detector
(Waters, Milford, MA, USA). Cell pellets were collected
and then disrupted prior to extraction. Isobutyl acetate was
used instead of acetone as extraction solvent for improved
extraction. The procedure was carried out as follows. Cell
pellets were collected by centrifugation at 9,800 × g for
10 min from 1 mL culture and washed once with 1 mL of
85% methanol. Cell pellets were resuspended in 0.5 mL of
a lysozyme solution (10 mg/mL) in an STE buffer (pH 8.0)
and then incubated at 37oC for 1 h. After centrifugation at
9,800 × g for 10 min, the supernatant was discarded and
the pellet was resuspended in 1.0 or 0.5 mL of isobutyl
acetate depending on the color of the pellet. β-carotene was
completely extracted into the isobutyl acetate. The isocratic
mobile phase of ethanol/methanol/tetrahydrofuran (75:20:5,
v/v/v) for β-carotene analysis or methanol/tert-butyl-metyl-
ether/water (42:54:4, v/v/v) for lycopene analysis was
used. A 20 µL filtered sample obtained using a DISMIC-
13HP 0.2 µm disposable syringe filter (ADVENTEC,
Tokyo, Japan) was injected into a YMC 30 column (250 mm
× 4.6 mm; YMC, Kyoto, Japan) and then eluted with the
mobile phase at 1.0 mL/min at room temperature. β-
carotene and lycopene were detected at a wavelength of
454 and 472 nm, respectively. Authentic β-carotene and
lycopene (Sigma-Aldrich, St. Louis, MO, USA) were used
as standards. Acetic acid and residual glycerol concentration
were determined using a commercial kit (Megazyme,
Wicklow, Ireland).
3. Results and Discussion
3.1. Biosynthesis of IPP and β-carotene in recombinant
E. coli with or without foreign MVA pathways
Essential genes for β-carotene biosynthesis (idi, ispA, crtE,
crtB, crtI, and crtY) as well as two native genes in the MEP
pathway of E. coli (dxs and dxr) were expressed in control
cells and strain 1. Additionally, the IPP biosynthetic pathway
was reconstructed by introducing the MVA pathway in
strain 1. Therefore, the difference between the two cell
types lay in the mechanics of IPP biosynthesis. The foreign
MVA pathway in E. coli can evade endogenous feedback
regulation, whereas the endogenous MEP pathway is under
feedback regulation control. Therefore, IPP biosynthesis
may be maximized in strain 1, whereas there is limited
biosynthesis of IPP in control cells. One of the rate-limiting
steps in the biosynthesis of carotenoids is the supply of IPP
[11,16]. Accordingly, it was anticipated that β-carotene
production would be higher in strain 1 than in control cells,
possibly due to the increased supply of IPP as the building
block for β-carotene production. The biosynthesis of IPP
and β-carotene is shown in Fig. 1 [11].
Acetate metabolism could also be potentially different
between the control and strain 1. The condensation reaction
between pyruvate and G3P leads to IPP and dimethyl allyl
pyrophosphate (DMAPP) in the MEP pathway. The MVA
pathway begins with the conversion of three molecules of
acetyl-CoA to MVA, ultimately leading to IPP. The starting
materials of MVA or MEP pathway are acetyl-CoA,
pyruvate, and G3P, which are intermediates of glycolysis.
Generally, a high growth rate increases the glycolytic flux
to pyruvate in wild-type E. coli cultures at the optimal
growth temperature of 37oC, which results in the imbalance
between glycolytic and tricarboxylic acid (TCA) flux,
resulting in the accumulation of copious amounts of acetate
[12]. Activation of the MEP pathway drains G3P and
Production of β-carotene and Acetate in Recombinant Escherichia coli with or without Mevalonate Pathway at… 1199
pyruvate, and may reduce flux to acetic acid. Furthermore,
the carbon flux from acetyl-CoA to acetate can be
redirected to MVA by expressing the MVA pathway.
Therefore, it is reasonable to anticipate that the amount of
acetate generated by these cells would be lower than that
by wild type E. coli and the production of acetic acid in
strain 1 would be much lower than that in control cells.
Based on these theoretical expectations, the influence of
the metabolic characteristics on the production of β-carotene
and acetic acids at growth-promoting or growth-retarding
conditions was investigated by controlling temperature or
culture pH.
3.2. Preliminary flask culture experiments
Initially, flask cultures of strain 1 were generated to
investigate the effect of culture temperature on β-carotene
production and cell growth. Temperature of 37 and 25oC
was selected as the growth-promoting and growth-retarding
conditions, respectively. The culture pH rapidly dropped
from neutral to 4.5 after 27 h at 37oC. The same pattern of
pH decrease occurred at 25oC but at a slower speed (Data
not shown). Cell growth and β-carotene production stopped
concomitantly at pH <4.5 in both cultures. β-carotene
production appeared to be enhanced during culture at 25oC.
However, this result may have been influenced by the pH
of the culture. We hypothesized that the beneficial effect of
culture at 25oC was due to the slower rate of pH decrease
than at 37oC. Thus, cells had more time to grow and
accumulate β-carotene before succumbing to the acidic pH.
If so, eliminating the influence of pH was needed to
accurately examine the temperature effect on β-carotene
production and cell growth. However, it was impossible to
maintain the pH of these cultures at neutral, despite the
incorporation of several buffer components in flask
experiments. Generally, culture acidification is caused by
secretion of acetic acid in E. coli cultures [12-14]. Therefore,
acetic acid was measured in the next experiment.
3.3. Effect of culture temperature on the biosynthesis of
acetic acid and β-carotene
A bioreactor system can control the cultural temperature or
pH to maintain a suitable point. Thus, the individual effect
of these factors was investigated in bioreactor cultures.
Bioreactor cultures were performed to evaluate the performance
of strain 1 at two different culture temperatures by
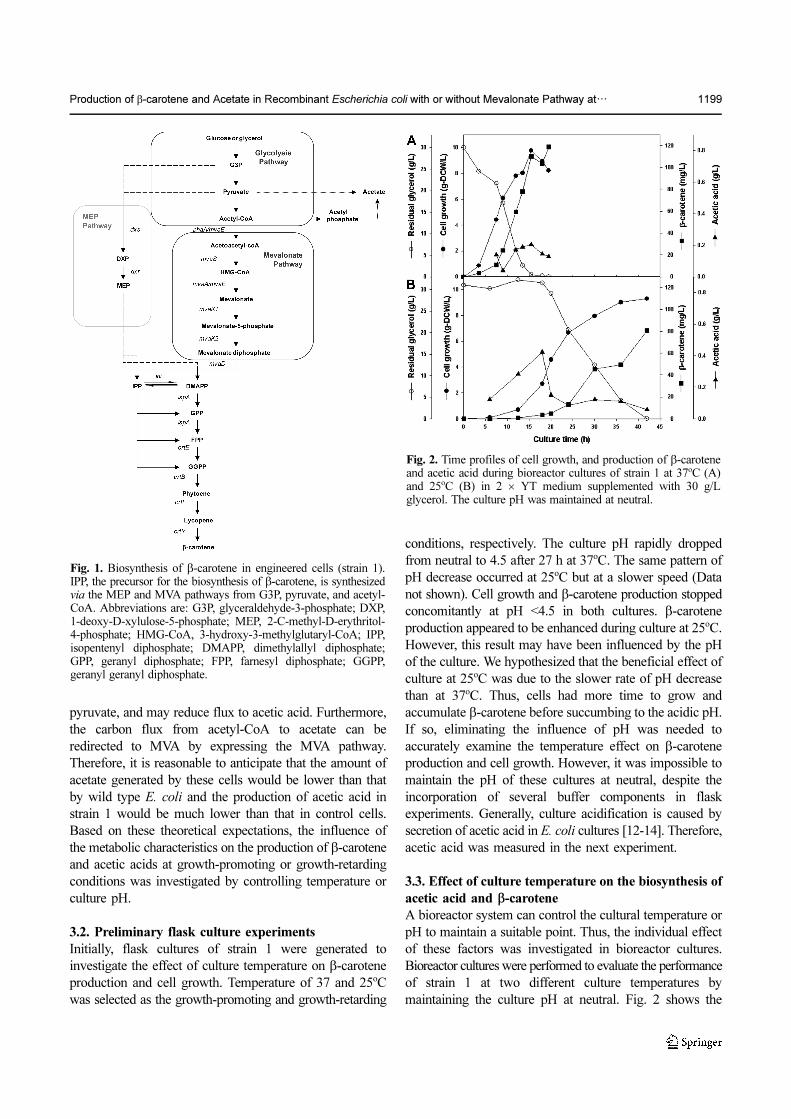
maintaining the culture pH at neutral. Fig. 2 shows the
Fig. 1. Biosynthesis of β-carotene in engineered cells (strain 1).IPP, the precursor for the biosynthesis of β-carotene, is synthesizedvia the MEP and MVA pathways from G3P, pyruvate, and acetyl-CoA. Abbreviations are: G3P, glyceraldehyde-3-phosphate; DXP,1-deoxy-D-xylulose-5-phosphate; MEP, 2-C-methyl-D-erythritol-4-phosphate; HMG-CoA, 3-hydroxy-3-methylglutaryl-CoA; IPP,isopentenyl diphosphate; DMAPP, dimethylallyl diphosphate;GPP, geranyl diphosphate; FPP, farnesyl diphosphate; GGPP,geranyl geranyl diphosphate.
Fig. 2. Time profiles of cell growth, and production of β-caroteneand acetic acid during bioreactor cultures of strain 1 at 37oC (A)and 25oC (B) in 2 × YT medium supplemented with 30 g/Lglycerol. The culture pH was maintained at neutral.

1200 Biotechnology and Bioprocess Engineering 17: 1196-1204 (2012)
profiles of cell growth and β-carotene production in
cultures of strain 1 at 37 and 25oC. β-carotene production
began when cell growth commenced and ceased after cell
growth stopped at 37oC, whereas the rate of β-carotene
production increased after the cell growth rate slowed after
24 h. Furthermore, production continued even after cell
growth had stopped at 25oC. As shown in Table 1, cells
grown at 37oC produced 119 mg/L of β-carotene after only
20 h, whereas 81 mg/L of β-carotene was produced for
42 h cultivation at 25oC. The result obtained at 37oC was
3.2-fold greater increase in volumetric productivity (Qp)
and a 1.4-fold higher increase in the specific growth rate
(µ) than that at 25oC. These results clearly indicated that β-
carotene production in strain 1 was favored in the actively
growing condition at a temperature of 37oC. This result
was contradictory to previous reports demonstrating a
temperature effect on the production of β-carotene by
recombinant E. coli, in which enhanced production of
carotenoids or β-carotene was obtained during slow growth
at 28oC, rather than active growth at 37oC in flask culture
[21] or bioreactor culture [4]. The main differences between
our system and those in the two previous studies were the
cell type and culture method. We evaluated cultures of
recombinant E. coli with additional MVA pathway (i.e.,
strain 1) in controlled bioreactor culture, whereas the
previous study utilized flask cultures of conventional β-
carotene producing recombinant E. coli utilizing native
MEP pathways only (our control cells). As described
above, we could not evaluate strain 1 in flask cultures at
the two different temperatures because acidification of
culturing medium occurred concurrently. Thus, we suspect
that the dichotomous results of the present and prior studies
are attributable to cell differences.
Bioreactor cultures of control cells were also performed
for comparison. As shown in Fig. 3, biphasic growth was
noted at 37oC, while the rates of glycerol consumption and
β-carotene production remained constant. Cultures generated
at 37oC were almost 2-fold greater in volumetric productivity
(Qp) than that at 25oC, while no significant differences in
the maximum cell growth and β-carotene titer were observed
at the two temperatures (Table 1). This result clearly
showed that 37oC was beneficial for cell growth and
production in the culture of control cells. The findings
supported the conclusion that β-carotene production in both
types of recombinant E. coli was favored during active
growth. Change in culture temperature also influenced the
acetate metabolism of control cells. An elevated amount of
acetate was produced at 37oC – 2 g/L at 6 h and increased
up to 8 g/L thereafter. Once glycerol had been consumed at
18 h, cells reutilized the excreted acetate as a complementary
carbon source. However, acetic acid production was not
notable at 25oC, perhaps due to the slower nutrient uptake
and growth rate. E. coli cells growing at 37oC produce a
high amount of acetic acid as an extracellular product, even
under fully aerobic conditions, because the cells surpass a
threshold-specific rate of nutrient consumption [12]. The
formation of acetate at this growth-promoting temperature
has been suggested to be caused by an imbalance between
glucose metabolism and respiration, a condition in which
the influx of carbon into cells exceeds the biosynthetic
Table 1. Cell growth and β-carotene production in engineered and control cells at different culture temperatures and pH
CellsTemp. (oC)
pHCulture time
(h)µ max
(/h)Xmax
(g/L)Pmax
(mg/L)Qp
(mg/L/h)Yp/x, max
(mg/g-cells)Y p/s
(mg/g-carbon sources)
Strain1 37 7 20 0.37 10 119 6.1 14.5 3.9
Strain1 25 7 42 0.26 9 81 1.9 8.7 2.7
Control 37 7 33 0.34 8 57 2.1 7.1 1.9
Control 25 7 47 0.29 9 47 1.1 5.3 1.6
Strain1 37 8 21 0.47 8 93 4.4 12.8 3.1
Strain1 37 6 24 0.34 12 112 6.2 9.3 3.8
Fig. 3. Time profiles of cell growth, and productions of β-carotene and acetic acid during the bioreactor cultures of controlcells at 37oC (A) and 25oC (B) in 2 × YT medium supplementedwith 30 g/L glycerol. The culture pH was maintained at neutral.

Production of β-carotene and Acetate in Recombinant Escherichia coli with or without Mevalonate Pathway at… 1201
demands and the capacity for energy generation within the
cell; saturation of the TCA cycle and/or the electron
transport chain may be the main cause. At 25oC, the
nutrient uptake rate and oxygen demands are reduced,
thereby reducing the formation of acetic acid [12,22,23].
Acetate is undesirable because it is toxic and inhibits cell
growth, even at concentrations as low as 8 mM (0.5 g/L)
[24]. The toxic effects of acetic acid derive from several
possible processes due to proton or anion accumulation.
The non-dissociated form of acetate can move freely
through the cell membrane and accumulate in the medium.
Some of this extracellular, non-dissociated form re-enters
the cells and dissociates due to the higher pH within the
cytoplasm. Acetate thus acts as a proton conductor and the
process causes a reduction in proton motive force [25].
Furthermore, accumulation of excess free protons may
damage ribosomal RNA as well as DNA, and also
denatures essential enzymes [26]. Therefore, the presently-
observed decrease in the cell growth rate of the control
cells from 0.34 to 0.05/h at 37oC was very likely caused by
the toxicity of the accumulated acetate. The result also
indicated that the MEP pathway does not operate efficiently
and, thus, most pyruvate is directed to the glycolytic
pathways, thereby producing acetate in control cells.
Notably, a small and comparable amount of acetic acid
was produced by strain 1 cultured at 37 or 25oC (Fig. 2). It
was demonstrated previously that strain 1 can biosynthesize
a considerable amount of MVA [8]. The reduced production
of acetate in E. coli over-producing MVA by expression of
heterologous MVA pathway genes was previously reported
[27]. Together with these reports, we suggest that all MVA
pathway enzymes are well expressed and, thus, that the
carbon flux from acetyl-CoA to acetate may have been
redirected to MVA by the expressed enzymes in strain 1
(Fig. 1). If so, why did strain 1 produce a high amount of
acetate in flask cultures? Significant differences in cell
growth and acetate production were observed in flasks and
bioreactor cultures of strain 1. During bioreactor growth, all
of the glycerol, 30 g/L, was exhausted, while approximately
16 g/L glycerol remained in the flasks when growth ceased
at either temperature. Severe oxygen limitation occurs in
shake flasks at the biomass level well below the level
achieved in bioreactors due to the lack of aeration [28,29].
Therefore, prolific production of acetate may lead to the
denaturation of the enzymes for cell growth as well as
MVA pathways, thereby resulting in low production of β-
carotene in flask cultures.
3.4. Lycopen production by strain 1 under weak
alkaline condition
The performance of strain 1 was evaluated in bioreactor
cultures at 37oC using different pH. Maximum concentration
of β-carotene, acetic acid, and cells in culture under slightly
acidic condition (pH 6) was 112 mg/L, 0.3 g/L, and 12 g/
L, respectively, which were not appreciably different from
the corresponding values obtained during culture under the
neutral pH condition (pH 7) (Table 1). However, of note,
the culture broth appeared darker with a stronger orange
color at the weakly alkaline pH of 8 than at a neutral pH.
To clarify the reason for this change, colored compounds
were extracted from cells cultured at both pH conditions
and then UV-VIS spectrophotometric analyses were
performed. The extract from cells cultured at the neutral
pH demonstrated three broad absorptions at 430, 454, and
480 nm, which represented the typical absorption spectrum
of β-carotene [30]. In contrast, absorptions at 454, 472, and
504 nm were evident in extracts from cells cultured under
the weak alkaline condition (Fig. 4). The absorption peaks
at 472 and 504 nm are typical of lycopene [30]. The
metabolites present in these extracts were identified using
HPLC; Fig. 5 shows representative chromatograms of
standards and extracts. The authentic β-carotene and
lycopene eluted at 10 and 95 min, respectively. Extracts
from cells cultured under the weak alkaline condition
revealed two peaks at the retention time of β-carotene and
lycopene, whereas only one peak was observed at the
retention time of β-carotene peak in extracts from cells
cultured under a neutral condition. These results clearly
indicated that lycopene was produced in addition to β-
carotene during culture at weak alkaline pH, whereas
growth at neutral pH resulted in the production of only β-
carotene.
Fig. 6 shows the time profiles of cell growth and productions
of β-carotene and lycopene in cultures generated at the
weak alkaline pH. Cells grew with a concomitant increase
in the production of β-carotene. No further increases in cell
growth and β-carotene production were observed at 12 h,
even though 8.9 g/L of glycerol remained at this time.
Subsequently, lycopene production began and product
Fig. 4. UV/Visible spectra of extracts from strain 1 cultured atweak alkaline pH (A) and neutral pH (B) at 37oC in 2 × YTmedium supplemented with 30 g/L glycerol.

1202 Biotechnology and Bioprocess Engineering 17: 1196-1204 (2012)
continued to increase until the glycerol was used up.
Notably, cells produced acetic acid from the early log
phase, reaching 4.5 g/L at 15 h, but cells were not able to
utilize acetate for biomass production in spite of the
exhaustion of glycerol at 18 h. This contrasted with the
result that this strain produced only a marginal amount of
acetic acid (<0.1 g/L) and the knowledge that E. coli
generally consumes acetic acid after exhaustion of the
available carbon source in neutral or weakly acidic cultures.
Alkaline pH induces deaminases such as tryptophan
deaminase (Tna A) and serine deaminase producing
pyruvic acids, which can be further degraded to acetic acid
[31,32]. Furthermore, growth of E. coli at external pHs
exceeding 7.8 generates an inverted pH gradient, which
drains the proton potential and effectively excludes acids
from cells [32,33]. This can fully explain the reason why
strain 1 could produce a high amount of acetic acid with no
consumption under the weak alkaline condition. In this
environment, the maximum cell concentration was 7.6 g/L,
which was lower than that obtained in cultures generated at
neutral or weak acidic pH. This provided further evidence
that cells can direct their overall metabolism into pathways
that channel metabolites towards acetic acid instead of
building biomass under weak alkaline condition.
At weak alkaline culture, the obtained total concentration
of carotenoids including β-carotene and lycopene was
150 µM, which corresponded to 68% of the β-carotene
titer (222 µM) obtained in neutral culture. This indicated
that carotenoid production decreased when cells were
cultured at the weak alkaline condition. It is obvious that
alkaline pH stimulated lycopene accumulation, as has been
reported previously [34]. This was probably due to the
decreased activity of lycopene cyclase, which catalyzes the
reaction transferring lycopene into β-carotene. The decreased
enzyme activity would retard the cyclization reaction,
leading to the accumulation of lycopene.
Fig. 5. HPLC chromatograms of extracts from strain 1 cultured at neutral or alkaline pH. (A) Standard lycopene (20 mg/mL); (B)standard β-carotene (60 µg/mL); (C) cell extract from neutral culture; (D) cell extract from weak alkaline culture; (E) mixture containingstandard lycopene and cell extract from weak alkaline culture.

Production of β-carotene and Acetate in Recombinant Escherichia coli with or without Mevalonate Pathway at… 1203
4. Conclusion
We evaluated a recombinant E. coli with an engineered
whole MVA pathway in addition to β-carotene biosynthetic
genes with regard to a metabolic balance between central
metabolism and β-carotene production in bioreactor cultures.
The performance of these novel cells was compared with
that of conventional β-carotene producing recombinant
E. coli. A small amount of acetic acid was produced with
no change in cell growth, even during active growth. In
contrast to this unique characteristic of the engineered cells,
control cells showed the same characteristic of central
metabolism as wild-type E. coli. The highest amount of
β-carotene, 119 mg/L, was obtained in engineered cells
cultured at 37oC, which was 2-folds higher than the
production by control cells, which also corresponded to a
2.9-fold greater increase in volumetric productivity.
The present findings are consistent with the conclusion
that the MVA pathway operates efficiently, redirecting
acetyl-CoA into IPP and thereby producing β-carotene,
whereas the native MEP pathway does not efficiently
convert pyruvate to IPP due to endogenous feedback
regulation. Furthermore, the data supports the view that the
carbon flux between cell growth and β-carotene production
are well-balanced in the engineered cells. The culture
pH was also important for maximizing the production of
β-carotene because engineered cells produced only β-
carotene at neutral pH, whereas some of the intermediate,
lycopene, could not be converted into β-carotene at weak
alkaline pH.
Acknowledgements
This work was supported by Pioneering Research Center
for Nano-morphic Biological Energy Conversion and Storage,
a Korea Science and Engineering Foundation (KOSEF)
grant funded by the Korea government (MEST) (R01-
2008-000-20835-0), and partially supported by the second
stage of the Brain Korea 21 project in 2011.
References
1. Burri, B. J. (1997) Beta-carotene and human health: A review ofcurrent research. Neutri. Res. 17: 547-580.
2. Jaramillo-Flores, M. E., J. J. Lugo-Martinez, E. Ramirez-San-juan, H. Montellano-Rosales, L. Dorantes-Alvarez, and H. Her-nandez-Sanchez (2005) Effect of sodium chloride, acetic acid,and enzymes on carotene extraction in carrots (Daccus carotaL.). J. Food Sci. 70: 136-142.
3. Garcia-Gonzalez, M., J. Moreno, J. C. Manzano, F. J. Florencio,and M. G. Guerrero (2005) Production of Dunaliella salina bio-mass rich in 9-cis-β-carotene and lutein in a closed tubular pho-tobioreactor. J. Biotechnol. 115: 81-90.
4. Kim, S. -W., J. -B. Kim, W. -H. Jung, J. -H. Kim, and J. -K. Jung(2006) Over-production of β-carotene from metabolically engi-neered Escherichia coli. Biotechnol. Lett. 28: 897-904.
5. Mantzouridou, F., T. Roukas, and P. Kotzekidou (2005) Produc-tion of beta-carotene from synthetic medium by Blakesleatrispora in fed-batch culture. Food Biotechnol. 18: 343-361.
6. Malisorn, C. and W. Suntornsuk (2008) Optimization of β-caro-tene production by Rhodotorula glutinis DM28 in fermented rad-ish brine. Biores. Technol. 99: 2281-2287.
7. Saenge, C., B. Cheirsilp, T. T. Suksaroge, and T. Bourtoom(2011) Efficient concomitant production of lipids and carotenoidsby oleaginous red yeast Rhodotorula glutinis cultured in palm oilmill effluent and application of lipids for biodiesel production.Biotechnol. Bioproc. Eng. 16: 23-33.
8. Verwaal, R., J. Wang, J. -P. Meijnen, H. Visser, G. Sandmann, J.A. V. D. Berg, and A. J. J. V. Ooyen (2007) High-level produc-tion of beta-carotene in Saccharomyces cerevisiae by successivetransformation with carotenogenic genes from Xanthophyllomy-ces dendrorhous. Appl. Environ. Microbiol. 73: 4342-4350.
9. Yoon, S. -H, S. -H. Lee, A. Das, H. -K. Ryu, H. -J. Jang, J. -Y.Kim, D. -K. Oh, J. D. Keasling, and S. -W. Kim (2009) Combi-natorial expression of bacterial whole mevalonate pathway forthe production of β-carotene in E. coli. J. Biotechnol. 140: 218-226.
10. Kim, J. H., S. -W. Kim, D. Q. A. Nguyen, H. Li, S. B. Kim, Y. -G.Seo, J. -K. Yang, I. -Y. Chung, D. H. Kim, and C. -J. Kim (2009)Production of β-carotene by recombinant Escherichia coli withengineered whole mevalonate pathway in batch and fed-batchcultures. Biotechnol. Bioproc. Eng. 14: 559-564.
11. Das, A., S. -H. Yoon, S. -H. Lee, J. -Y. Kim, D. -K. Oh, and S. -W.Kim (2007) An update on microbial carotenoid production:Application of recent metabolic engineering tools. Appl. Micro-
Fig. 6. Time profiles of cell growth (A), and productions of β-carotene and lycopene (B) during the bioreactor cultures of strain 1at weak alkaline pH in 2 × YT medium supplemented with 30 g/Lglycerol. The culture temperature was maintained at 37oC.

1204 Biotechnology and Bioprocess Engineering 17: 1196-1204 (2012)
biol. Biotechnol. 77: 505-512.12. Eiteman, M. A. and E. Altman (2006) Overcoming acetate in
Escherichia coli recombinant protein fermentations. Trends Bio-technol. 24: 530-536.
13. Selvarasu, S., D. S. -W. Ow, S. Y. Lee, M. M. Lee, S. K. -W. Oh,I. A. Karimi, and D. -Y. Lee (2009) Characterizing Escherichiacoli DH5α growth and metabolism in a complex medium usinggenome-scale flux analysis. Biotechnol. Bioeng. 102: 923-934.
14. Vemuri, G. N., T. A. Mining, E. Altman, and M. A. Eiteman(2005) Physiological response of central metabolism in Escher-ichia coli to deletion of pyruvate oxidase and introduction of het-erologous pyruvate carboxylase. Biotechnol. Bioeng. 90: 64-76.
15. Won, W., C. Park, C. Park, S. Y. Lee, K. S. Lee, and J. Lee (2011)Parameter estimation and dynamic control analysis of central car-bon metabolism in Escherichia coli. Biotechnol. Bioproc. Eng.16: 216-228.
16. Martin, V. J. J., D. J. Pitera, S. T. Withers, J. D. Newman, and J.D. Keasling (2003) Engineering a mevalonate pathway inEscherichia coli for production of terpenoids. Nat. Biotechnol.21: 796-802.
17. Pitera, D. J., C. J. Paddon, J. D. Newman, and J. D. Keasling(2007) Balancing a heterologous mevalonate pathway forimproved isoprenoid production in Escherichia coli. Metab. Eng.9: 193-207.
18. Sambrook, J. and D. W. Russel (2010) Molecular Cloning: ALaboratory Manual. 3rd ed. Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, NY, USA.
19. Schierle, J., B. Pietsch, A. Ceresa, and C. Fizet (2004) Methodfor the determination of β-carotene in supplements and raw mate-rials by reversed-phase liquid chromatography: Single laboratoryvalidation. J. AOAC Int. 87: 1070-1082.
20. Torrecilla, J. S., M. Camara, V. Fernandez-Ruiz, G. Piera, and J.O. Caceres (2008) Solving the spectroscopy interference effectsof β-carotene and lycopene by neural networks. J. Agric. Food.Chem. 56: 6261-6266.
21. Sandmann, G., M. Albrecht, G. Schnurr, O. Knorzer, and P. Boger(1999) The biotechnological potential and design of novel caro-tenoids by gene combination in Escherichia coli. Trends Biotech-nol. 17: 233-237.
22. Vemuri, G. N., E. Altman, D. P. Sangurdekar, A. B. Khodursky,and M. A. Eiteman (2006) Overflow metabolism in Escherichiacoli during steady-state growth: Transcriptional regulation and
effect of redox ratio. Appl. Env. Microbiol. 72: 3653-3661.23. Mey, M. D., S. D. Maeseneire, W. Soetaert, and E. Vandamme
(2007) Minimizing acetate formation in E. coli fermentations. J.Ind. Microbiol. Biotechnol. 34: 689-700.
24. Roe, A. J., C. O’Byrns, D. Mclaggan, and I. R. Booth (2002)Inhibition of Escherichia coli growth by acetic acid: A problemwith methionine biosynthesis and homocystein toxicity. Micro-biol. 148: 2215-2222.
25. Warnecke, T. and R. T. Gill (2005) Organic acid toxicity, toler-ance, and production in Escherichia coli biorefining applications.Microb. Cell Fact. 4: 25.
26. Raja, N., M. Goodson, D. G. Smith, and R. J. Rowbury (1991)Decrease DNA damage by acid and increased repair of acid-damaged DNA in acid-habituated Escherichia coli. J. Appl. Bac-teriol. 70: 507-511.
27. Tabata, K. and S. -I. Hashimoto (2004) Production of mevalonateby a metabolically-engineered Escherichia coli. Biotechnol. Lett.26: 1487-1491.
28. Dahlgren, M. E., A. L. Powell, R. L. Greasham, and H. A.George (1993) Development of scale-down techniques for inves-tigation of recombinant Escherichia coli fermentations: Acidmetabolites in shake flasks and stirred bioreactors. Biotechnol.Prog. 9: 580-586.
29. Soini, J., K. Ukkonen, and P. Neubauer (2008) High cell densitymedia for Escherichia coli are generally designed for aerobic cul-tivations-consequences for large-scale bioprocesses and shakeflask cultures. Microb. Cell Fact. 7: 26.
30. Anguelova, T. and J. Warthesen (2000) Lycopene stability intomato powders. J. Food. Sci. 65: 67-70.
31. Stancik, L. M., D. M. Stancik, B. Schmidt, D. M. Barnhart, Y. N.Yoncheva, and J. L. Slonczewski (2002) pH-dependent expres-sion of periplasmic proteins and amino acid catabolism inEscherichia coli. J. Bacteriol. 184: 4246-4258.
32. Krulwich, T. A., G. Sachs, and E. Padan (2011) Molecularaspects of bacterial pH sensing and homeostasis. Nat. Rev. Micro.9: 330-343.
33. Zilberstein, D., V. Agmon, S. Schuldiner, and E. Padan (1984)Escherichia coli intracellular pH, membrane potential, and cellgrowth. J. Bacteriol. 158: 246-252.
34. Alper, H., K. Miyaoku, and G. Stephanopoulos (2006) Charac-terization of lycopene-overproducing E. coli strains in high celldensity fermentations. Appl. Microbiol. Biotechnol.72: 968-974.