příčně pruhované
DESCRIPTION
Příčně pruhované. svaly:. Kontrakce. Jak to tedy funguje ???. Nervosvalové spojení. podněty vyvolávající kontrakci svalu přicházejí po motoneuronu a končí na svalovém vlákně ve speciálním útvaru nervosvalová ploténka ta má stavbu a vlastnosti jednoduché synapse - PowerPoint PPT PresentationTRANSCRIPT



podněty vyvolávající kontrakci svalu přicházejí po motoneuronu a končí na svalovém vlákně ve speciálním útvaru nervosvalová ploténka
ta má stavbu a vlastnosti jednoduché synapse
akční potenciály uvolňují na ploténce acetylcholin
vazba acetylcholinu na N-cholinové receptory (= zároveň chemicky řízené iontové kanály) v místech záhybů sarkolemy otevření iontového kanálu (na 1ms) depolarizace vznik ploténkového potenciálu
vyprázdnění 1 vezikuly s Ach aktivace několika tisíců N-cholinových receptorů; pro vybavení akčního potenciálu (AP) je však nutné vyprázdnit asi 100 vezikul (aktivace asi 200 000 kanálů)
akční potenciál (AP) se šíří podél sarkolemy na celé svalové vlákno
acetylcholin v synaptické štěrbině velmi rychle štěpen acetylcholinesterázou to umožňuje rychlou repolarizaci a účinný přenos všech po sobě jdoucích podnětů
ACh
nervosvalové spojení

axon motoneuronu
nervosvalová ploténka
svalové vlákno

terminální část axonu motoneuronu obsahující vezikuly
s mediáterem (actylcholinem)
T-tubulus
= speciální typ spojení mezi nervem a svalem
sarkoplazma
SVALOVÁ BUŇKA
sarkoplazmatické retikulum (L-tubulus)
kontraktilní aparát buňky
sarkolema

N-cholinové receptory (zároveň iontové kanály)
Ca 2+
vezikula s mediátorem
(ACh)
vápenatý kanál
AP

ACh
depolarizace membránydepolarizace membrány
- 90mV
+ 30mVN-cholinový receptor (zároveň iontový kanál)
sarkolemyK+K+ K+
K+
Na+
Na+
Na+
Na+
Na+
Na+
Na+
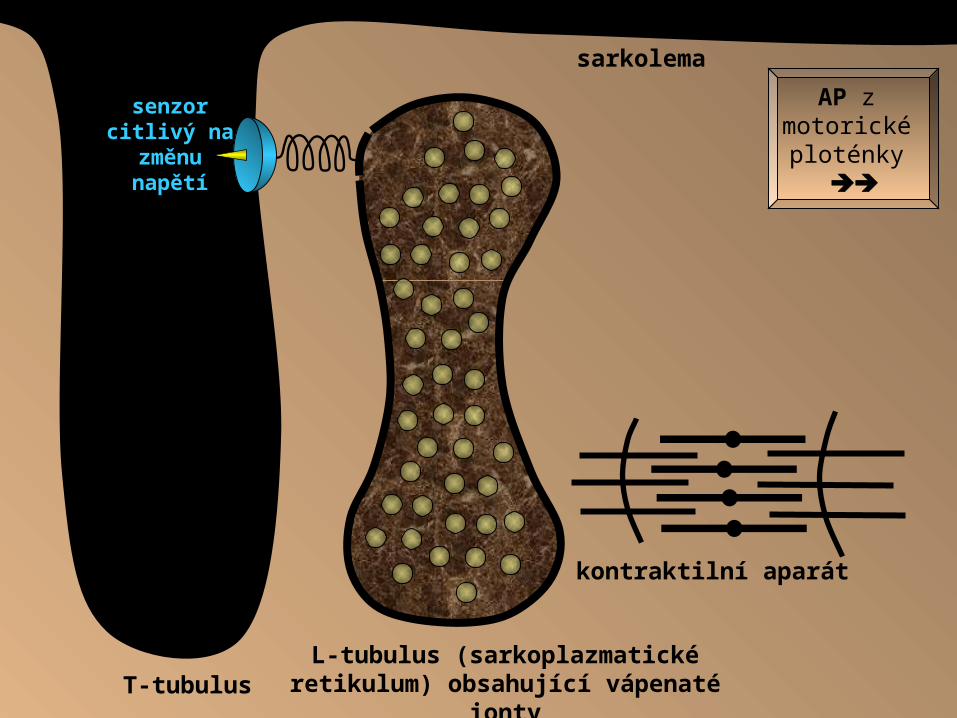
L-tubulus (sarkoplazmatické retikulum)
obsahující vápenaté iontyT-tubulus
sarkolema
AP z motorické ploténky
senzor citlivý na
změnu napětí
kontraktilní aparát

Akční potenciál se šíří po membráně motoneuronu.
Když dorazí AP k synaptickému knoflíku, otevře iontové kanály pro Ca 2+ a vápenaté ionty putují do terminální části axonu.
Ca 2+ ionty (přes několik dalších dějů) uvolňují vezikuly s mediátory ze synaptického zakončení.
Acetylcholin se váže na N-cholinové receptory=kanály, tím se kanály na okamžik otevřou a po koncentračním spádu jsou ionty Na+ čerpány do svalové
buňky a naopak ionty K+ ven z buňky (draselných iontů se dostane z buňky méně, než kolik se do buňky dostalo sodných iontů) membrána se
depolarizuje.
Vlna depolarizace (AP) opouští nervosvalovou ploténku, šíří se po sarkolemě a v membráně T-tubulu tuto změnu membránového potenciálu zaznamená citlivý
receptor. Tento receptor je mechanicky spřažen s vrátky (Ca 2+ kanály) sarkoplazmatického retikula. Po projití AP membránou receptor tuto změnu
napětí zaznamenává a otvírá vrátka sarkoplazmatického retikula, odkud mohou být po koncentračním spádu čerpány Ca 2+ ionty do sarkoplazmy k myofibrilám,
které je využijí k zahájení vlastní svalové kontrakce.

nervosvalové spojení je velmi citlivé, může být ovlivňováno různými látkami
vybavení svalového vzruchu lze zabránit podáním kurare (alkaloid), který se pevně váže na acetylcholinové receptory postsynaptické membrány ireverzibilní blokace receptorů
podobně působí také hadí jedy
blokace možná také inhibicí acetylcholinesterázy (tak působí některé látky obsažené v pesticidech)
toxin botulin blokuje uvolňování acetylcholinu při akčním potenciálu a brání tak excitaci sarkolemy
Strychnos toxifera rostlina, ze které se
získává jed kurare

molekula MYOZINU je tvořena dvojšroubovicí polypeptidových
řetězců, na konec každého řetězce nasedá globulární část - hlavice
MYOZINOVÉ FILAMENTUM je tvořené svazkem molekul myozinu (150 - 300)
Molekula myozinu má dvojdílnou kloubovitě napojenou hlavou, která obsahuje enzym adenozintrifosfatázu ( štěpící ATP). Každá hlavice má vazebné místo
pro ATP a aktin.
Klidový stav = hlavice myzinu nejsou ve spojení s molekulami aktinu.
Kloubovitá pohyblivost hlavice + její reverzibilní vazba na aktin vzájemný posuv filament.
ocas hlavy

molekuly aktinu
Aktinové vlákno je dvojšroubovice tvořená globulárními monomery proteinu aktinu. Vždy 2 takovéto vzájemně spirálně stočené řetězce
tvoří aktinové filamentum.
Dvojšroubovicí se táhne vlákno tropomyozinu (tenká polypeptidová dvojitá šroubovice), na němž jsou v pravidelných odstupech
navázané molekuly troponinu (1 molekula troponinu je tvořena komplexem 3 proteinů).
vlákno tropomyozinutroponin

Ca 2+
Úloha Ca 2+ ve svalové kontrakci
Vápenaté ionty způsobí konformační změnu troponinu vedoucí k zasunutí
tropomyozinového řetězce hlouběji do šroubovice. Obnaží se tak vazebná
místa pro myozinovou hlavici.
vazebné místo pro myozin
volno
troponin

primární podnět pro svalový stah = akční potenciál (AP)
spojovací článek mezi AP a kontraktilním aparátem = ionty vápníku
v sarkolemně svalového vlákna – složitý systém příčních (transverzálních) tubulů = T-tubulů – vychlípeniny sarkolemy zasahující hluboko do nitra svalového vlákna
AP depolarizace sarkolemy depolarizace T-tubulů
uvnitř svalového vlákna podélné (longitudinální) tubuly = L-tubuly; fungují jako rezervoár Ca 2+ iontů, které uvolňují do cytoplazmy jako reakci na depolarizaci sarkolemy …

klidová vysoká koncentrace Ca 2+ v tubulech musí být udržována výkonnými pumpami, které je čerpají ze sarkoplazmy (antiport s Mg 2+)
koncentrace vápníku je v sarkoplazmě v klidovém stavu nízká
AP vylití Ca 2+ iontů do sarkoplazmy navázání Ca 2+ iontů na molekuly troponinu konformační změna troponinu, zasunutí
tropomyozinového vlákna hlouběji do aktinové šroubovice, změna polohy tropomyozinu odhalení vazebných míst aktinu
pro hlavice myozinu
uvolněné Ca 2+ ionty jsou okamžitě čerpány zpět do L-tubulů; na dva ionty Ca 2+ se spotřebuje jedna molekula ATP

obě hlavy jedné molekuly myozinu váží po jednom ADP a anorganický fosfor Pi
v této formě (komplex myozin-ADP-Pi) svírají s myozinovým vláknem úhel 90°
při vysoké intracelulární koncentraci Ca 2+ se hlavy myozinu spojují s aktinem vzniká komplex aktin-myozin-ADP-Pi
uvolní-li se z tohoto komplexu anorganický fosfor Pi, myozinové hlavy využijí této vzniklé energie a překlopí se z polohy 90° na 50° filamenta se proti sobě posunou
po uvolnění i ADP se hlavy dostanou do polohy 45°, tím se posuv ukončí
zbývající komplex aktin-myozin tvoří stabilní „rigorový komplex“, který může být uvolněn pouze vazbou ATP
snadná protažitelnost svalu v klidu je důležitá např. při plnění srdce nebo pro snadnou poddajnost natahovačů

vazba myozinu s novým ATP uvolňuje vazbu myozinové hlavice s aktinovým vláknem
ATP se přitom štěpí myozinovou ATPázou, ale na hlavici zůstává navázáno; uvolněnou energii myozin spotřebuje na narovnání hlavice do pozice 90°
spotřeba ATP tedy provází relaxaci, nikoli kontrakci svalu
pokud je intracelulární koncentrace ATP a Ca 2+ nadále vysoká (závislost na frekvenci přicházejících vzruchů), začíná celý cyklus znovu, hlavice se opět připojí, ale už v jiném bodě POSUV
délka posuvu aktinových vláken při jedné kontrakci velmi malá (5-10 mikronů); proto opakované připojení hlavic v dalších bodech
všechny hlavy myozinového vlákna se pohybují asynchronně kontrakce je plynulá
pokles intracelulárního Ca 2+ nakonec ukončí cyklus posuvu

.
.
Ca
CaCa
Ca Ca
myozin s hlavicí
aktinové vlákno
tropomyozinové vlákno
troponinADP
Pi
sark.retikulum

50°
45°
dvě možnosti pokračování akce:
s ATP – uvolnění stahu
bez ATP – „rigor mortis“

ve svalech mrtvého organismu (3-6 hodin po zastavení dodávky kyslíku) se ATP již netvoří
Ca 2+ nemůže být čerpáno zpět do L-tubulů
není k dispozici ATP pro rozštěpení stabilního komplexu aktin-myozin
nastává rigor mortis = mrtvolná ztuhlost
Trvá dokud nejsou svalové proteiny zničeny autolýzou (15-25 hodin).

ATP
.

CaCa
Ca CaCa

… úloha proteinů, ATP a iontů při svalové kontrakci
Myozin
Aktin
Tropomyozin
Troponin
ATP
ADP Mg 2+
Ca 2+

svaly jsou stále v mírné kontrakci, kterou označujeme jako svalové napětí (svalový tonus)
svalové napětí představuje základní nervosvalový děj
není energeticky náročné, nevyvolává únavu
mizí jen po ztrátě inervace nebo smrtí
během života není klidové napětí svalů stále stejné (snížení ve spánku, bezvědomí, atd.)
mění v závislosti na informacích z periferních receptorů a na informacích z CNS
rozdíl je i v klidovém napětí jednotlivých svalů
PORUCHY SVALOVÉHO NAPĚTÍ
Hypotonie – snížení svalového napětí
Hypertonie – zvýšení svalového napětí
během spánku nelze stát

svaly pro svou činnost potřebují přívod vzruchů
svaly jsou inervovány mozkovými a míšními nervy
MOTORICKÁ N. VLÁKNA
umožňují aktivní volní pohyb
vycházejí z:
motorických jader mozkových nervů pro svaly obličejové a krční
předních rohů míšních pro svaly trupu a končetin
SENZORICKÁ N. VLÁKNA
podávají informace o stavu natažení či smrštění svalů
vycházejí ze:
svalových vřetének a šlachových tělísek (= proprioreceptory)
Aktivní svalový pohyb vyvolán na základě nervových vzruchů přicházejících do svalu motorickými vlákny. Nejdůležitější úlohu mají
vzruchy přiváděné prostřednictvím pyramidové dráhy z motorické oblasti mozkové kůry. Při porušení části této dráhy ztrácejí svaly schopnost
aktivních vědomých pohybů.

Axony motoneuronů
Nervosvalové ploténky
= všechna svalová vlákna inervovaná 1 motoneuronem
Svalová vlákna
MOTONEURON = nervová buňka končící na
svalovém vláknu

Příčně pruhovaný sval je řízen motorickými nervovými vlákny z předních rohů
míšních. Jeden motoneuron inervuje několik svalových vláken a vytváří motorickou jednotku.
MALÉ MOTORICKÉ JEDNOTKY
Tvořeny pouze několika vlákny. Ve svalech vykonávajících přesné a jemné pohyby (př. okohybné svaly, drobné svaly ruky). V okohybných svalech jsou motorické jednotky tvořeny 8-12 svalovými vlákny.
VELKÉ MOTORICKÉ JEDNOTKY
Obsahují stovky až tisíce vláken. Vyskytují se tam, kde se jedná především o dlouhodobé udržení napětí k zajištění vzpřímené polohy těla (antigravitační svaly). V hýžďových svalech připadá na 1 MJ asi 500 vláken.
Většina svalů obsahuje oba typy motorických
jednotek, ale podle funkce svalu vždy jeden typ
převládá.

Jeden sval může mít mnoho různých
motorických jednotek.
Ne při každé kontrakci musí být aktivovány všechny motorické
jednotky daného svalu.
mícha
Motorická jednotka 1
Motorická jednotka 2
Motorická jednotka 3
motorický nerv
Všechna svalová vlákna dané
motorické jednotky jsou stejného typu.

Chceš více světla ? Rozsviť více žárovek !
Chceš větší svalovou kontrakci ? Aktivuj více motorických jednotek !
Čím více motorických jednotek je aktivováno (prostřednictvím povelů vydaných mozkovou kůrou, procházejících míchou a končících na zakončeních motoneuronů na motorických ploténkách), tím větší je svalová kontrakce.

myograf – přístroj k registraci svalových stahů
myogram – časový záznam kontrakce izolovaného svalu
myograf
elementárním svalovým úkonem je jednotlivé svalové trhnutí (kontrakce) = reakce svalu na jediný podnět
již při vybavení tohoto jednoduchého svalového stahu můžeme zaznamenat dobu latence (doba potřebná k cestě vzruchu nervovým vláknem a zdržení na nervosvalové ploténce; zlomek vteřiny)
moderní myograf

na zápisu svalových stahů = na myografické křivce rozlišujeme části vzestupné (krescenty) a sestupné (dekrescenty)
nervový impulz
intenzita svalového stahu
čas

1. Padne-li druhý podnět do doby absolutní refrakterní fáze, kladoucí odpor, je neúčinný.
Výsledná křivka je totožná s křivkou prvního podnětu.
V době velmi krátké absolutní refrakterní fáze je sval nedráždivý.
2. Padne-li druhý podnět do krescenty prvního trhnutí, oba stahy splynou v 1 trhnutí o větší amplitudě.
Dochází k sečtení obou podráždění - tzv. sumace.
3. Padne-li druhý podnět do dekrescenty prvního trhnutí, získáme myogram se dvěma vrcholy - tzv. superpozice.
podráždíme-li sval 2 podněty, jdoucími po sobě ve velmi krátkém časovém intervalu,
můžeme zaznamenat tyto výsledky:
nervový impulz
nervový impulz nervový
impulz
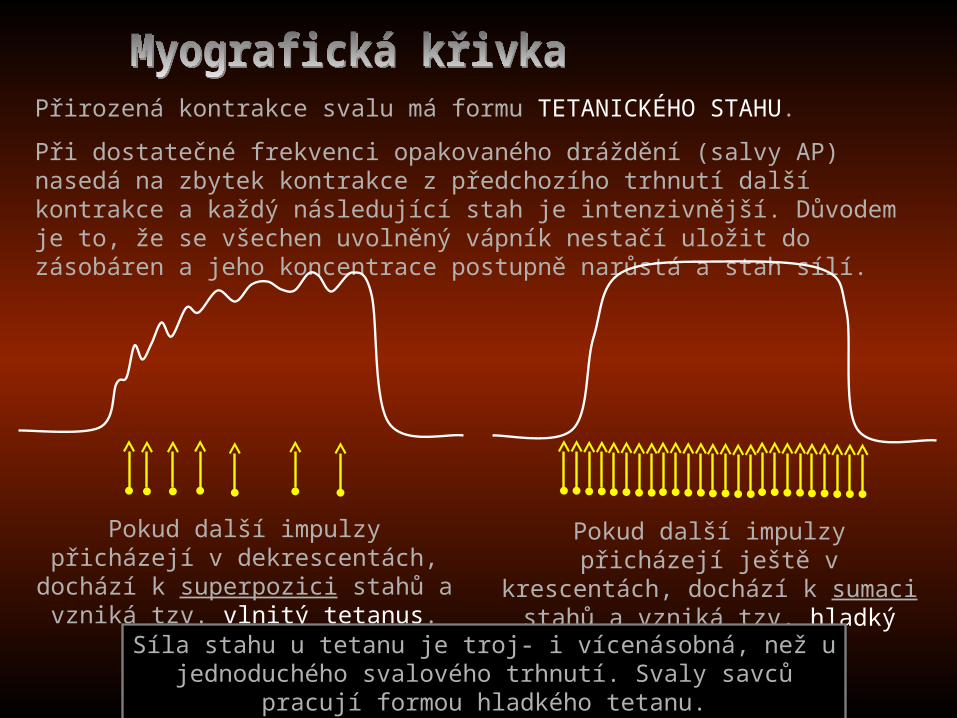
Přirozená kontrakce svalu má formu TETANICKÉHO STAHU.
Při dostatečné frekvenci opakovaného dráždění (salvy AP) nasedá na zbytek kontrakce z předchozího trhnutí další kontrakce a každý následující stah je intenzivnější. Důvodem je to, že se všechen uvolněný vápník nestačí uložit do zásobáren a jeho koncentrace postupně narůstá a stah sílí.
Pokud další impulzy přicházejí v dekrescentách, dochází k superpozici
stahů a vzniká tzv. vlnitý tetanus.
Pokud další impulzy přicházejí ještě v krescentách, dochází k sumaci stahů a
vzniká tzv. hladký tetanus.
Síla stahu u tetanu je troj- i vícenásobná, než u jednoduchého svalového trhnutí. Svaly savců pracují formou hladkého tetanu.

svalový stah
sumace super- pozice
vlnitý tetanus hladký tetanus

přirozená kontrakce kosterního svalu má podobu hl. tetanického stahu
stupňování svalové aktivity je možné díky tomu, že někdy je aktivováno více, někdy méně motorických jednotek (MJ) svalu = prostorová sumace
jeden sval může mít 100 MJ, ale i 2 000 MJ (př. okohybné svaly = jedny z nejjemnějších)
čím větší počet MJ, tím jemnější odstupňování kontrakce
síla každé MJ může být navíc stupňována zvyšováním frekvence nervových impulzů = časová sumace

IZOTONICKÁ IZOMETRICKÁ
Svalové napětí zůstává stejné.
Sval mění svoji délku.
Sval vykonává práci.
Svalové napětí se zvyšuje.
Sval nemění svoji délku.
Svalová práce je nulová.

svaly převádějí energii vázanou chemicky přímo na energii mechanickou
zdroj E = ATP - - - jeho štěpení na ADP a Pi (může probíhat i anaerobně)
spotřebovaný ATP je ihned regenerován
ZPŮSOBY ZISKU ENERGIE
• 1. štěpení kreatinfosfátu
• 2. anaerobní glykolýza
• 3. aerobní spalování glukózy a tuků na CO2
• ve svalu je obsaženo ATP asi na 10 kontrakcí
• štěpením kreatinfosfátu se získá E na dalších asi 50 kontrakcí, než je i tato zásoba vyčerpána (rychle využitelná energetická rezerva – s využitím jeho E lze dosahovat krátkodobých špičkových výkonů)

anaerobní glykolýza se rozběhne s malým zpožděním
glukóza z krve a glykogen z jater ve svalu odbourávány na kyselinu mléčnou
při lehké práci je tato energeticky málo výhodná produkce ATP vystřídána asi po 1 minutě aerobním odbouráváním glukózy (pokud to ale při déletrvající práci nestačí, anaerobní glykolýza probíhá současně s aerobní glykolýzou)
anaerobní glykolýza však nemůže probíhat dlouhodobě, a to díky hromadění kyseliny mléčné a reakcím vedoucím k únavě svalu
déletrvající svalové výkony možné pouze díky aerobního uvolňování E z glukózy a tuků

RYCHLE UNAVITELNÉ
relativně malé množství myglobinu
relativně chudé prokrvení
krátká, rychlá kontrakce
specializace na anaerobní metabolismus
rozsáhlé sarkoplazmatické retikulum, méně mitochondrií
spotřebují veliké množství energie a velice snadno se unaví
př. ve svalech horních končetin, které často zvedají těžké objekty na krátkou dobu
VYTRVALÉ
velké množství myoglobinu
bohaté prokrvení
dlouhá, pomalá kontrakce
specializace na aerobní metabilismus (energeticky výhodnější než anaerobní)
šetří energii, ale pracují pomalu, zato vytrvale
př. ve svalech zabezpečujících držení těla
podle síly, rychlosti a stálosti kontrakce

kromě červených a bílých svalů existují ještě vlákna přechodná
využívají aerobní metabolismus, ale nejsou odolná proti únavě tolik, jako červené svaly
př. ve svalech dolní končetiny, které musí dlouhou dobu pohybovat tělem (př. při chůzi nebo běhu)
Většina kosterních svalů v těle obsahuje směs všech typů vláken, některé však mají výraznou převahu vláken určitého typu. Př. šíjová vlákna - červené svaly,
okohybná vlákna - bílé svaly.
bílé vlákno
přechodné vlákno
červené vlákno
zvětšeno 600x
Někteří lidé mají relativně větší počet vláken určitého typu (genetické příčiny).
př. LÝTKOVÉ SVALY
Bílá vlákna (krátký sprint)
Přechodná vlákna (dlouhý vytrvalostní běh)
Červená vlákna (držení postavení těla)

energetické potřeby svalu při práci jsou závislé na dostatečném zásobení kyslíkem – tedy na prokrvení svalů, na srdečním výkonu a dýchání
určitou rezervu O2 přímo ve svalech poskytuje barvivo MYOGLOBIN
ale i tak vzniká během namáhavé svalové práce díky anaerobní fázi KYSLÍKOVÝ DLUH (= nadspotřeba kyslíku po svalovém výkonu)
vyrovnání kyslíkového dluhu pomocí zrychleného dýchání umožňuje opětovné obnovení energetických rezerv a odbourání nahromaděné kyseliny mléčné (proto je jeho splátka často vyšší, než původní výpůjčka)
Při výkonech delších než cca 30 sekund se kyselina mléčná nahromadí takovým
způsobem, že se prudce zvyšuje čas na zotavení. Běžci na 400 metrů proto
potřebují k zopakování kvalitního výkonu mnohem delší odpočinek než sprinteři na
100 metrů.
Kyslíkový dluh zahrnuje:
odbourání kyseliny mléčné
doplnění zásob ATP, kreatinfosfátu, glykogenu, kyslíku vázaného na myoglobin

klidový objem O2
pracovní objem
O2
čas
objem O2 „kyslíkový deficit“
„kyslíkový dluh“
intenzita dýchání
náročná svalová práce
Kyslíkový deficit se splácí formou kyslíkového dluhu.

= síla, kterou vyvíjí sval při stahu
je vyjádřena hmotností závaží, které sval právě ještě udrží v rovnováze proti gravitaci
měří se různými typy dynamometrů (siloměrů) a udává se v [kg/cm2], tzn. v kilogramech zvednuté hmotnosti na jeden centimetr čtvereční příčného průřezu svalem
největší je na počátku stahu a postupně se zmenšuje
svalová síla závisí na počtu motorických jednotek
- v ontogenetickém vývoji se zvětšuje až do 25 let- vrchol svalové síly je mezi 25. a 30. rokem, pak postupně klesá
- ženy mají asi o 30-50 % menší sval. sílu než muži- trénováním se síla svalů zvětšuje
svalová práce = svalová síla působící po určité dráze, měří se ergometry, v J
svalový výkon = svalová práce vykonaná za časovou jednotku

je vyvolána dlouhou a silnou nebo opakovanou svalovou kontrakcí
příčiny únavy:
vyčerpání zdrojů energie (hlavně ATP)
snížení zásob glykogenu
nahromadění kyseliny mléčné ( snížení pH ve tkáni, bolesti, otoky)
útlum nervosvalového přenosu
svalová únava je signál pro přerušení práce, než dojde u úplnému vyčerpání a případně poškození svalu – má tedy pro organismus ochranný význam
odolnost proti svalové únavě se dá zvyšovat tréninkem
dráždění sympatiku může svalovou únavu zmenšit a oddálit (tzv. Orbeliho fenomén)
únavu odstraňuje odpočinek
akutní (mizí do 24 hod) x chronická (až úplná vyčerpanost)

V rychlosti nemá člověk proti některým zvířatům šanci…
Nejrychlejší lidský závodník dosáhne ve sprinterském závodě pouhých 37 km/hod.

nejrychlejší tvor naší planety
pasivně až 350 km/hod, aktivně až 180 km/hod
Sokol stěhovavý (Falco peregrinus)
Plachetník atlantský (Istiophorus platypterus)
nejrychlejší obyvatel oceánů
rychlost až 120 km/hod
Gepard (Acynonyx jubatus)
nejrychlejší suchozemský
živočich
rychlost až 120 km/hod
za 3 s zrychlení z 0 na 90 km/hod

Hlemýžď by urazil sprinterskou „stovku“ za necelých dvanáct hodin …