prenatal immune challenge induces developmental changes in the morphology of pyramidal neurons of...
TRANSCRIPT

Schizophrenia Research 107 (2009) 99–109
Contents lists available at ScienceDirect
Schizophrenia Research
j ourna l homepage: www.e lsev ie r.com/ locate /schres
Prenatal immune challenge induces developmental changes in themorphology of pyramidal neurons of the prefrontal cortex andhippocampus in rats
Moogeh Baharnoori a, Wayne G. Brake b, Lalit K. Srivastava a,⁎a Douglas Mental Health University Institute, Department of Psychiatry, McGill University, 6875 LaSalle Blvd, Montreal, Canada H4H 1R3 (QC)b Centre for Studies in Behavioral Neurobiology (CSBN) Department of Psychology, Concordia University Montreal, Canada H4B 1R6 (QC)
a r t i c l e i n f o
⁎ Corresponding author. Tel.: +514 761 6131x2936;E-mail address: [email protected] (L.K. Sriv
0920-9964/$ – see front matter © 2008 Elsevier B.V.doi:10.1016/j.schres.2008.10.003
a b s t r a c t
Article history:Received 21 July 2008Received in revised form 1 October 2008Accepted 2 October 2008Available online 12 November 2008
The neural mechanisms by which maternal infections increase the risk for schizophrenia arepoorly understood; however, animal models using maternal administration of immuneactivators suggest a role for cytokine imbalance in maternal/fetal compartments. As cytokinescan potentially affect multiple aspects of neuronal development and the neuropathology ofschizophrenia is believed to involve subtle temporo-limbic neurodevelopmental alterations, weinvestigated morphological development of the pyramidal neurons of the medial prefrontalcortex (mPFC) and hippocampus in rats that were prenatally challenged with the immuneactivator lipopolysaccharide (LPS).Pregnant Sprague-Dawley rats were administered with LPS (at E15- E16) or saline. The brains ofoffspring were processed for Golgi-Cox staining at postnatal days 10, 35 and 60. Dendriticlength, branching, spine density and structure were quantified using Neurolucida software. Atall ages, dendritic arbor was significantly reduced in mPFC and CA1 neurons of LPS-treatedanimals. Dendritic length was significantly reduced in the mPFC neurons of LPS group at P10and 35 but returned to control values at P60. Opposite pattern was observed in CA1 region ofLPS animals (normal values at P10 and 35, but a reduction at P60). LPS treatment significantlyaltered the structure of CA1 dendritic spines at P10. Spine density was found to be significantlylower only in layer V mPFC of P60 LPS rats. The study provides the first evidence that prenatalexposure to an immune activator dynamically affects spatio-temporal development ofpyramidal neurons in mPFC and hippocampal that can potentially lead to aberrant neuronalconnectivity and functions of these structures.
© 2008 Elsevier B.V. All rights reserved.
Keywords:SchizophreniaNeurodevelopmentPrenatal infectionDendrite arbor and spine density
1. Introduction
Evidence from epidemiology, imaging and neuropatholo-gical studies led to the notion that early developmentalaberrations within the central nervous system may form thestarting point for schizophrenia (Heinz et al., 2003; Marencoand Weinberger, 2000; McDonald and Murray, 2000). Inaddition, epidemiological data indicate enhanced risk of otherpsychiatric disorders with possible neurodevelopmental
fax: +514 762 3034.astava).
All rights reserved.
origin like autism following prenatal and early postnatalinfections. The complex mode of transmission of schizophre-nia suggests important roles of both genetic and environ-mental factors (Cannon and Clarke, 2005). Indeed, geneticapproaches have identified a cluster of candidate genesassociated with schizophrenia that may have potential rolesin neural development and function (Harrison and Weinber-ger, 2005). Possible contribution of early environmental factorto the origins of psychiatric disorders was recognized as earlyas by Kraepelin who proposed childhood inflammation of thebrain and head injury as potential risk factors, as cited byDean and Murray (2005). Over the years, a number of earlylife environmental risk factors, e.g., obstetric complications

100 M. Baharnoori et al. / Schizophrenia Research 107 (2009) 99–109
(Boksa and El-Khodor, 2003) and maternal stress (Khashanet al., 2008), malnutrition (McClellan et al., 2006) andinfection (Brown et al., 2000; Penner and Brown, 2007)have been investigated. Substantial evidence has accumu-lated in recent years suggesting that pre- or early postnatalinfections enhance the risk of psychiatric disorders withneurodevelopmental origin like schizophrenia and autism(Fatemi et al., 2002a; Libbey et al., 2005; Meyer et al., 2007)due to their potential to interfere with brain developmentprocesses (Patterson, 2002) . Animal studies support thishypothesis as rodent offspring prenatally exposed to influ-enza virus (Shi et al., 2003) , viral mimic PolyI:C (Meyer et al.,2008b; Ozawa et al., 2006; Zuckerman and Weiner, 2005) orbacterial endotoxin LPS (Fortier et al., 2004, 2007; Golan et al.,2005; Hava et al., 2006; Romero et al., 2007, 2008) displaybehavioral deficits parallel to those reported in schizophrenia.For example LPS treated adult offspring show increase inamphetamine-induced locomotion (Fortier et al., 2004) andanxiety response (Hava et al., 2006), deficits in prepulseinhibition to acoustic startle (PPI) (Fortier et al., 2007; Romeroet al., 2007, 2008), reduced social interaction (Hava et al.,2006) and alteration in distinct forms of learning andmemory(Golan et al., 2005).
The mechanisms by which maternal immune activationleads to behavioral changes arenot fullyunderstood. However,it is well-documented that prenatal LPS administration, asother immune activators, alters the level of pro inflammatorycytokines in maternal and fetal compartments (Ashdownet al., 2006; Cai et al., 2000; Urakubo et al., 2001). Cytokinescan potentially affect multiple aspects of neuronal develop-ment (Bauer et al., 2007; Jonakait, 2007) Particularly relevantin the present context are the findings that suggests cytokinesdetrimentally affect the development of hippocampal andcortical neurons (Gilmore et al., 2004; Marx et al., 2001;Neumann et al., 2002) In addition, severe neuronal loss andastrogliosis along with defects in corticogenesis and neuronalmigration have been reported following prenatal immuneactivation (Fatemi et al., 1999,2002a,b; Samuelsson et al.,2006; Zuckerman et al., 2003). Maternal LPS treatmentspecifically increases the number of pyramidal and granularcells in the hippocampus alongwith the shrinkage of pyramidalneurons in hippocampus of adult offspring. In CA1 region, LPStreatment changes the pyramidal layer thickness at early agesbut not in adulthood (Golan et al., 2005). In addition to neuronalchanges, there is also evidence for neuroanatomical changes inthe central dopaminergic, GABAergic andglutamatergic sys-tems following prenatal immune activation (Meyer et al.,2008a,b; Nyffeler et al., 2006).
With regard to brain anatomical changes reported inschizophrenia, it is notable that alterations in pyramidalneuron morphology in prefrontal cortical and limbic struc-tures is a replicated finding in post-mortem studies (Glantzand Lewis, 2000; Kolluri et al., 2005).
The aim of the present study was to make a quantitativeassessment of pyramidal neuronal morphology at definedneonatal and pre- and post-pubertal periods in mPFC (layerIII/V) and hippocampus CA1 region, following moderate LPS(100 μg/kg) challenge at embryonic days 15 and 16, a periodwhich approximates mid to late pregnancy in human.
Our data showed that prenatal LPS exposure is able toinduce significant age-dependent cytoarchitectural changes
in pyramidal neurons of mPFC and hippocampus that mayaffect the proper connectivity and neuronal transmission inthese regions leading to cognitive and behavioral impair-ments reported in prenatally infected offspring.
2. Methods
2.1. Animals
Timed Pregnant Sprague-Dawley rats (Charles RiverLaboratory, Quebec) were individually housed in temperatureand humidity-controlled cages on a 12 h light/dark cycle withad libitum access to food and water. All procedures wereperformed in strict accordance with the guidelines estab-lished by the Canadian Council on Animal Care and wereapproved by the McGill University Animal Care Committee.The animals were injected intraperitoneally (ip) with 100 μg/kg of LPS or saline (from E. coli serotype 0111:B4, L-2630,Sigma, Canada) once daily, at E15 and E16 (n=5 per group).
On the day of birth, the pups were cross-fostered withsurrogate dams in mixed litters. Male pups were retained forthe study. At Postnatal 10 (P10) 5 pups per treatment (onefrom each dam) were anesthetized with Ketamine–Xylazinemixture (100 mg/kg Ketamine, 0.8 mg/kg xylazine) andtranscardially perfused with 0.9% saline and their brainprocessed for Golgi staining. At P35 and P60, 7 animals pertreatment (1–2 pups per dam) were perfused and their brainsprocessed as above for Golgi staining.
2.2. Modified Golgi-Cox staining
The brains were processed for Golgi-Cox staining asdescribed in (Gibb and Kolb,1998). The brains were immersedin 20 ml of Golgi-Cox solution and stored for 21 days in thedark. Then the solution was replaced with 30% sucrose andkept for 5more days. Brains were cut at coronal plane (200 μmthickness) using a Vibrotome (VT1000S, Leica). The sectionswere placed on microslides (Snow-coat extra, Surgipath) andthe blotted slides kept in a humid chamberovernight. The nextday, the sections were developed in ammonium hydroxide for30min and then placed in Kodak film fixer for another 30min.Sections were washed in water and dehydrated in gradedconcentration of ethanol (70%, 95%, 100%, and 100%) andmounted using Permount (Fisher, SP15-100).
2.3. Morphological analysis
Basilar dendrites of pyramidal neurons in layer III and V ofmPFC (Cg1 and Prelimbic cortex), plate 8-10 and pyramidalneurons in the CA1 region of the hippocampus, plate 58-63(Paxinos and Watson, 2007) were selected for analysis. Aperson blind to the conditions of the study performed neuronselection and tracing. Pyramidal neurons were identified bytheir specific triangular shape of the soma and their apicalextensions toward pial surface. From each animal, 5 sectionswere selected per region and in each section; one neuron perhemisphere was reconstructed three-dimensionally usingcomputer based neuron tracing system (NEUROLUCIDA,MicroBrightField, Williston, VT). A Leica microscope (LeicaDM5000B) together with a motor stage equipped withtransducers on the XYZ-axes was used to trace each neuron
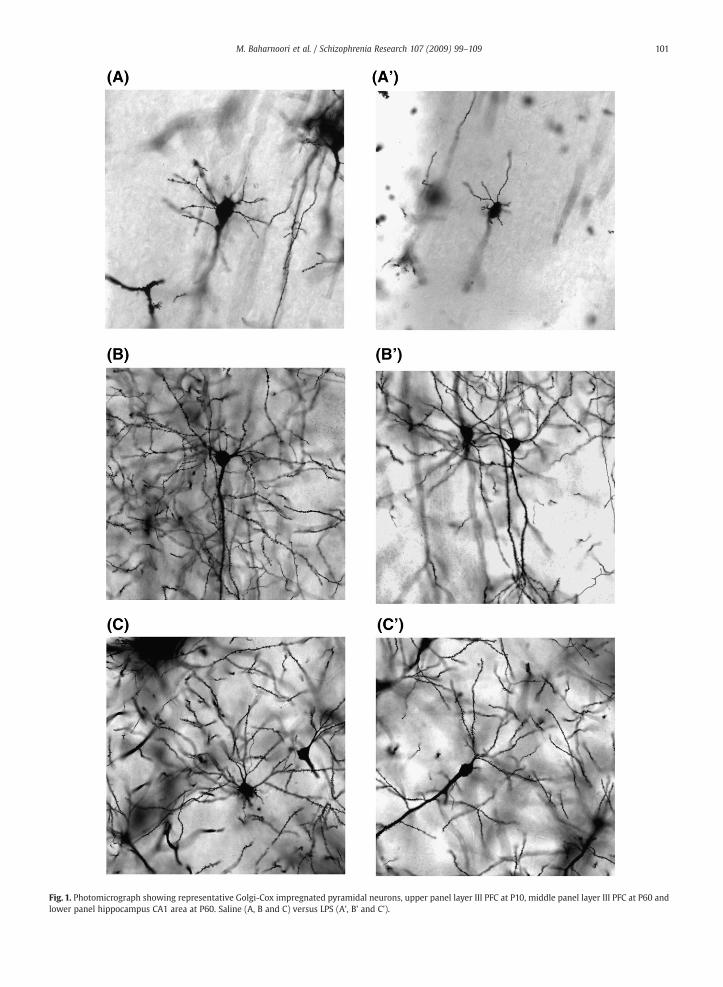
Fig. 1. Photomicrograph showing representative Golgi-Cox impregnated pyramidal neurons, upper panel layer III PFC at P10, middle panel layer III PFC at P60 andlower panel hippocampus CA1 area at P60. Saline (A, B and C) versus LPS (A', B' and C').
101M. Baharnoori et al. / Schizophrenia Research 107 (2009) 99–109
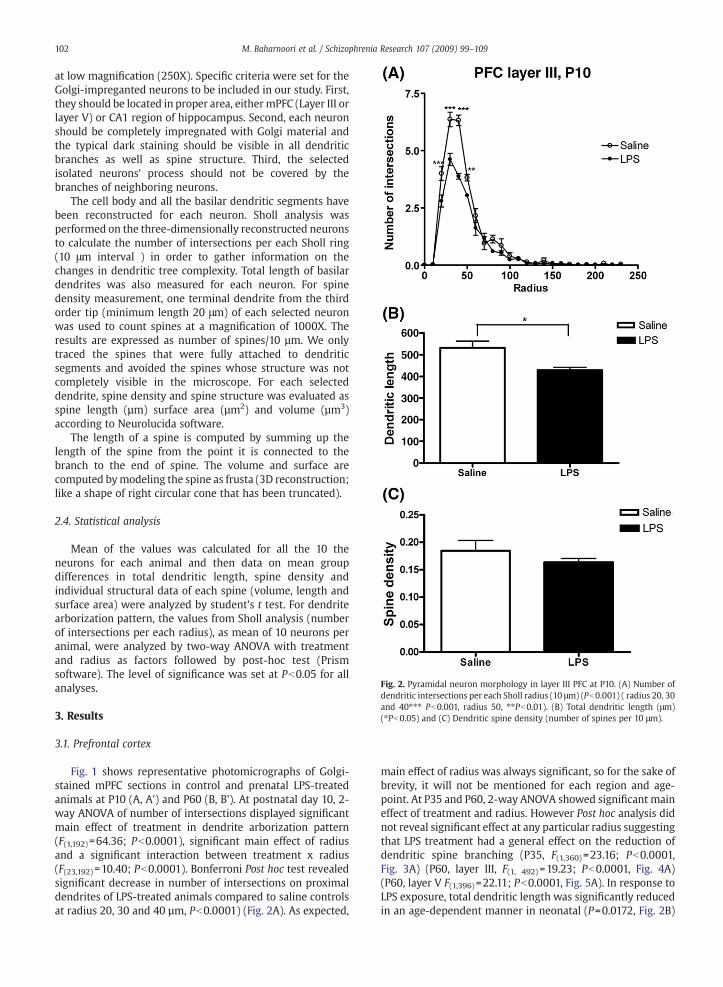
Fig. 2. Pyramidal neuron morphology in layer III PFC at P10. (A) Number ofdendritic intersections per each Sholl radius (10 μm) (Pb0.001) ( radius 20, 30and 40⁎⁎⁎ Pb0.001, radius 50, ⁎⁎Pb0.01). (B) Total dendritic length (μm)(⁎Pb0.05) and (C) Dendritic spine density (number of spines per 10 μm).
102 M. Baharnoori et al. / Schizophrenia Research 107 (2009) 99–109
at low magnification (250X). Specific criteria were set for theGolgi-impreganted neurons to be included in our study. First,they should be located in proper area, eithermPFC (Layer III orlayer V) or CA1 region of hippocampus. Second, each neuronshould be completely impregnated with Golgi material andthe typical dark staining should be visible in all dendriticbranches as well as spine structure. Third, the selectedisolated neurons' process should not be covered by thebranches of neighboring neurons.
The cell body and all the basilar dendritic segments havebeen reconstructed for each neuron. Sholl analysis wasperformed on the three-dimensionally reconstructed neuronsto calculate the number of intersections per each Sholl ring(10 μm interval ) in order to gather information on thechanges in dendritic tree complexity. Total length of basilardendrites was also measured for each neuron. For spinedensity measurement, one terminal dendrite from the thirdorder tip (minimum length 20 μm) of each selected neuronwas used to count spines at a magnification of 1000X. Theresults are expressed as number of spines/10 µm. We onlytraced the spines that were fully attached to dendriticsegments and avoided the spines whose structure was notcompletely visible in the microscope. For each selecteddendrite, spine density and spine structure was evaluated asspine length (µm) surface area (µm2) and volume (µm3)according to Neurolucida software.
The length of a spine is computed by summing up thelength of the spine from the point it is connected to thebranch to the end of spine. The volume and surface arecomputed bymodeling the spine as frusta (3D reconstruction;like a shape of right circular cone that has been truncated).
2.4. Statistical analysis
Mean of the values was calculated for all the 10 theneurons for each animal and then data on mean groupdifferences in total dendritic length, spine density andindividual structural data of each spine (volume, length andsurface area) were analyzed by student's t test. For dendritearborization pattern, the values from Sholl analysis (numberof intersections per each radius), as mean of 10 neurons peranimal, were analyzed by two-way ANOVA with treatmentand radius as factors followed by post-hoc test (Prismsoftware). The level of significance was set at Pb0.05 for allanalyses.
3. Results
3.1. Prefrontal cortex
Fig. 1 shows representative photomicrographs of Golgi-stained mPFC sections in control and prenatal LPS-treatedanimals at P10 (A, A') and P60 (B, B'). At postnatal day 10, 2-way ANOVA of number of intersections displayed significantmain effect of treatment in dendrite arborization pattern(F(1,192)=64.36; Pb0.0001), significant main effect of radiusand a significant interaction between treatment x radius(F(23,192)=10.40; Pb0.0001). Bonferroni Post hoc test revealedsignificant decrease in number of intersections on proximaldendrites of LPS-treated animals compared to saline controlsat radius 20, 30 and 40 µm, Pb0.0001) (Fig. 2A). As expected,
main effect of radius was always significant, so for the sake ofbrevity, it will not be mentioned for each region and age-point. At P35 and P60, 2-way ANOVA showed significant maineffect of treatment and radius. However Post hoc analysis didnot reveal significant effect at any particular radius suggestingthat LPS treatment had a general effect on the reduction ofdendritic spine branching (P35, F(1,360)=23.16; Pb0.0001,Fig. 3A) (P60, layer III, F(1, 492)=19.23; Pb0.0001, Fig. 4A)(P60, layer V F(1,396)=22.11; Pb0.0001, Fig. 5A). In response toLPS exposure, total dendritic length was significantly reducedin an age-dependent manner in neonatal (P=0.0172, Fig. 2B)
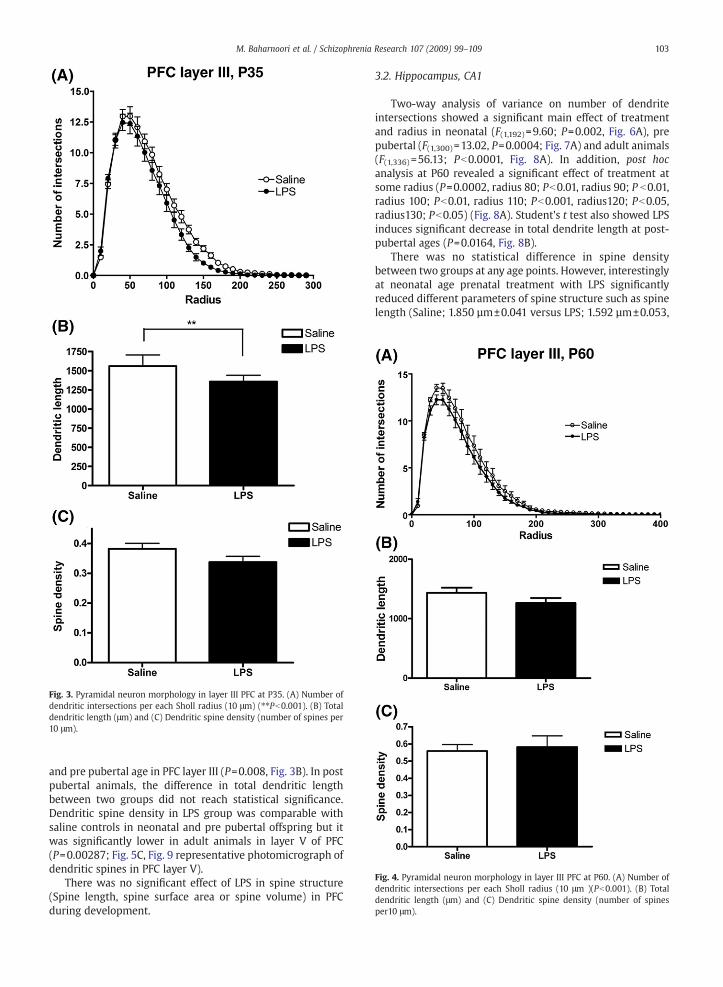
Fig. 4. Pyramidal neuron morphology in layer III PFC at P60. (A) Number ofdendritic intersections per each Sholl radius (10 μm )(Pb0.001). (B) Totaldendritic length (μm) and (C) Dendritic spine density (number of spinesper10 μm).
Fig. 3. Pyramidal neuron morphology in layer III PFC at P35. (A) Number ofdendritic intersections per each Sholl radius (10 μm) (⁎⁎Pb0.001). (B) Totaldendritic length (μm) and (C) Dendritic spine density (number of spines per10 μm).
103M. Baharnoori et al. / Schizophrenia Research 107 (2009) 99–109
and pre pubertal age in PFC layer III (P=0.008, Fig. 3B). In postpubertal animals, the difference in total dendritic lengthbetween two groups did not reach statistical significance.Dendritic spine density in LPS group was comparable withsaline controls in neonatal and pre pubertal offspring but itwas significantly lower in adult animals in layer V of PFC(P=0.00287; Fig. 5C, Fig. 9 representative photomicrograph ofdendritic spines in PFC layer V).
There was no significant effect of LPS in spine structure(Spine length, spine surface area or spine volume) in PFCduring development.
3.2. Hippocampus, CA1
Two-way analysis of variance on number of dendriteintersections showed a significant main effect of treatmentand radius in neonatal (F(1,192)=9.60; P=0.002, Fig. 6A), prepubertal (F(1,300)=13.02, P=0.0004; Fig. 7A) and adult animals(F(1,336) =56.13; Pb0.0001, Fig. 8A). In addition, post hocanalysis at P60 revealed a significant effect of treatment atsome radius (P=0.0002, radius 80; Pb0.01, radius 90; P b0.01,radius 100; Pb0.01, radius 110; Pb0.001, radius120; Pb0.05,radius130; Pb0.05) (Fig. 8A). Student's t test also showed LPSinduces significant decrease in total dendrite length at post-pubertal ages (P=0.0164, Fig. 8B).
There was no statistical difference in spine densitybetween two groups at any age points. However, interestinglyat neonatal age prenatal treatment with LPS significantlyreduced different parameters of spine structure such as spinelength (Saline; 1.850 µm±0.041 versus LPS; 1.592 µm±0.053,
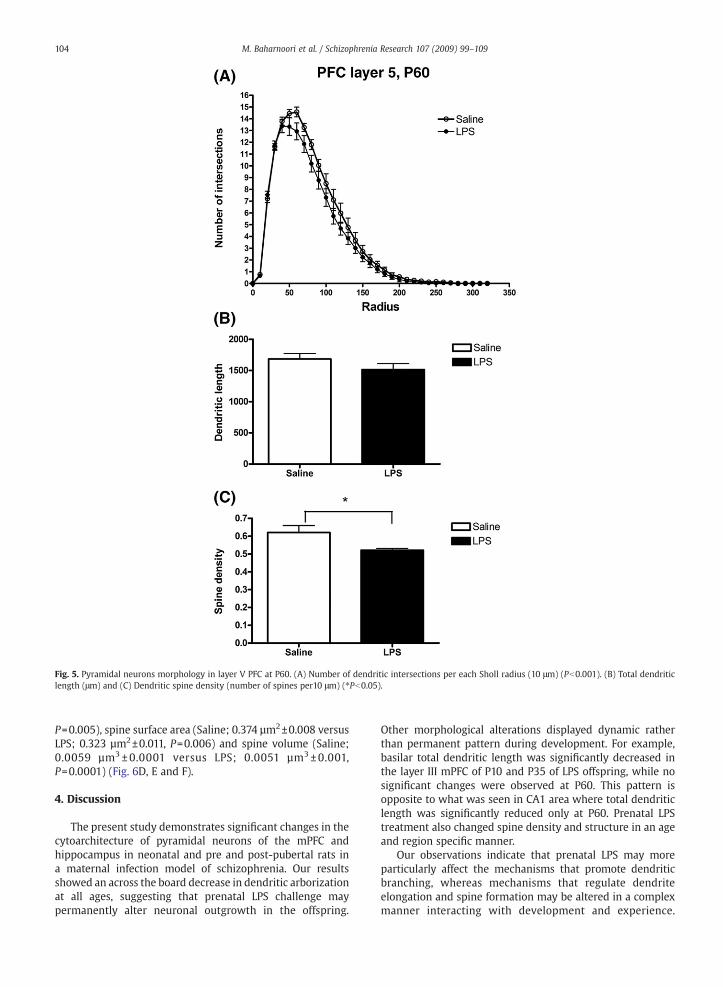
Fig. 5. Pyramidal neurons morphology in layer V PFC at P60. (A) Number of dendritic intersections per each Sholl radius (10 μm) (Pb0.001). (B) Total dendriticlength (μm) and (C) Dendritic spine density (number of spines per10 μm) (⁎Pb0.05).
104 M. Baharnoori et al. / Schizophrenia Research 107 (2009) 99–109
P=0.005), spine surface area (Saline; 0.374 µm2±0.008 versusLPS; 0.323 µm2±0.011, P=0.006) and spine volume (Saline;0.0059 µm3 ± 0.0001 versus LPS; 0.0051 µm3± 0.001,P=0.0001) (Fig. 6D, E and F).
4. Discussion
The present study demonstrates significant changes in thecytoarchitecture of pyramidal neurons of the mPFC andhippocampus in neonatal and pre and post-pubertal rats ina maternal infection model of schizophrenia. Our resultsshowed an across the board decrease in dendritic arborizationat all ages, suggesting that prenatal LPS challenge maypermanently alter neuronal outgrowth in the offspring.
Other morphological alterations displayed dynamic ratherthan permanent pattern during development. For example,basilar total dendritic length was significantly decreased inthe layer III mPFC of P10 and P35 of LPS offspring, while nosignificant changes were observed at P60. This pattern isopposite to what was seen in CA1 area where total dendriticlength was significantly reduced only at P60. Prenatal LPStreatment also changed spine density and structure in an ageand region specific manner.
Our observations indicate that prenatal LPS may moreparticularly affect the mechanisms that promote dendriticbranching, whereas mechanisms that regulate dendriteelongation and spine formation may be altered in a complexmanner interacting with development and experience.
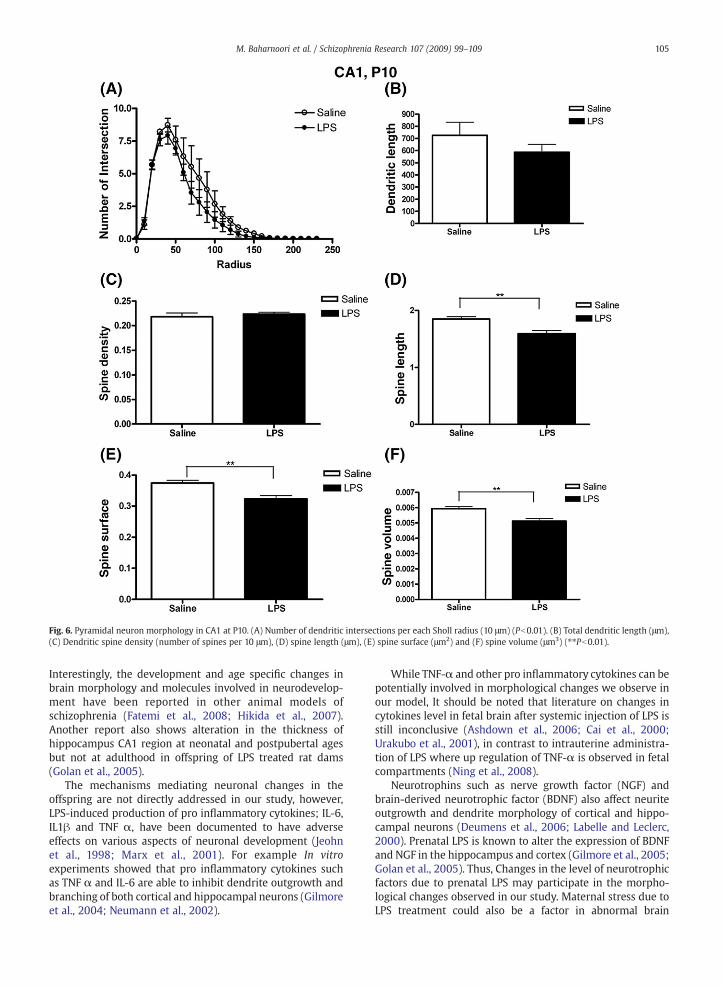
Fig. 6. Pyramidal neuron morphology in CA1 at P10. (A) Number of dendritic intersections per each Sholl radius (10 μm) (Pb0.01). (B) Total dendritic length (μm),(C) Dendritic spine density (number of spines per 10 μm), (D) spine length (μm), (E) spine surface (μm2) and (F) spine volume (μm3) (⁎⁎Pb0.01).
105M. Baharnoori et al. / Schizophrenia Research 107 (2009) 99–109
Interestingly, the development and age specific changes inbrain morphology and molecules involved in neurodevelop-ment have been reported in other animal models ofschizophrenia (Fatemi et al., 2008; Hikida et al., 2007).Another report also shows alteration in the thickness ofhippocampus CA1 region at neonatal and postpubertal agesbut not at adulthood in offspring of LPS treated rat dams(Golan et al., 2005).
The mechanisms mediating neuronal changes in theoffspring are not directly addressed in our study, however,LPS-induced production of pro inflammatory cytokines; IL-6,IL1β and TNF α, have been documented to have adverseeffects on various aspects of neuronal development (Jeohnet al., 1998; Marx et al., 2001). For example In vitroexperiments showed that pro inflammatory cytokines suchas TNF α and IL-6 are able to inhibit dendrite outgrowth andbranching of both cortical and hippocampal neurons (Gilmoreet al., 2004; Neumann et al., 2002).
While TNF-α and other pro inflammatory cytokines can bepotentially involved in morphological changes we observe inour model, It should be noted that literature on changes incytokines level in fetal brain after systemic injection of LPS isstill inconclusive (Ashdown et al., 2006; Cai et al., 2000;Urakubo et al., 2001), in contrast to intrauterine administra-tion of LPS where up regulation of TNF-α is observed in fetalcompartments (Ning et al., 2008).
Neurotrophins such as nerve growth factor (NGF) andbrain-derived neurotrophic factor (BDNF) also affect neuriteoutgrowth and dendrite morphology of cortical and hippo-campal neurons (Deumens et al., 2006; Labelle and Leclerc,2000). Prenatal LPS is known to alter the expression of BDNFand NGF in the hippocampus and cortex (Gilmore et al., 2005;Golan et al., 2005). Thus, Changes in the level of neurotrophicfactors due to prenatal LPS may participate in the morpho-logical changes observed in our study. Maternal stress due toLPS treatment could also be a factor in abnormal brain
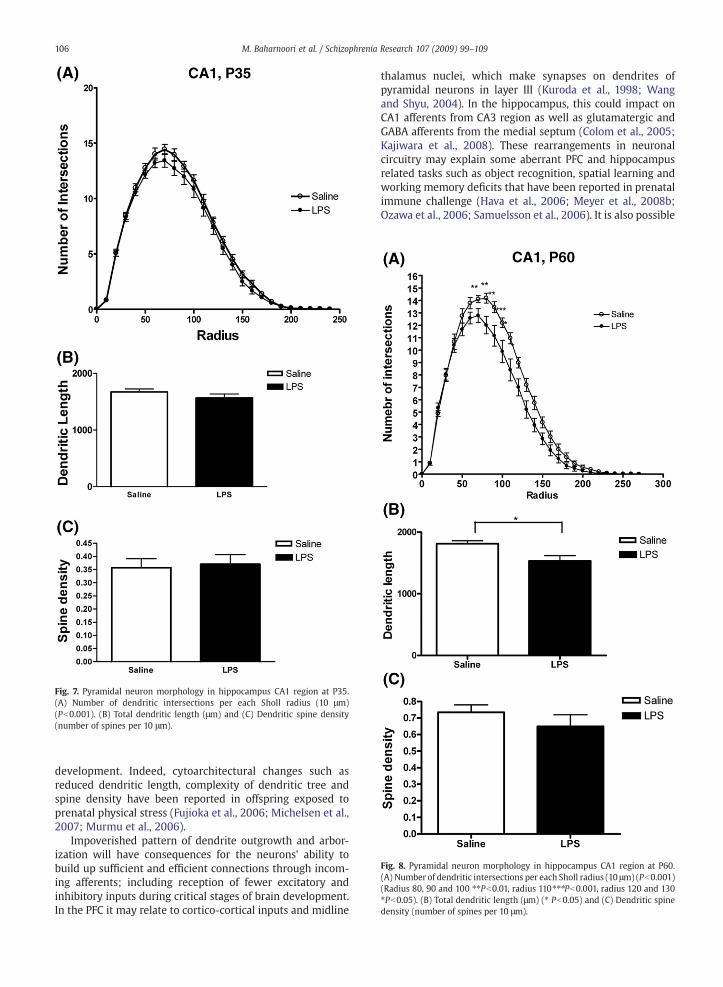
Fig. 7. Pyramidal neuron morphology in hippocampus CA1 region at P35.(A) Number of dendritic intersections per each Sholl radius (10 μm)(Pb0.001). (B) Total dendritic length (μm) and (C) Dendritic spine density(number of spines per 10 μm).
Fig. 8. Pyramidal neuron morphology in hippocampus CA1 region at P60.(A) Number of dendritic intersections per each Sholl radius (10 μm) (Pb0.001)(Radius 80, 90 and 100 ⁎⁎Pb0.01, radius 110⁎⁎⁎Pb0.001, radius 120 and 130⁎Pb0.05). (B) Total dendritic length (μm) (⁎ Pb0.05) and (C) Dendritic spinedensity (number of spines per 10 μm).
106 M. Baharnoori et al. / Schizophrenia Research 107 (2009) 99–109
development. Indeed, cytoarchitectural changes such asreduced dendritic length, complexity of dendritic tree andspine density have been reported in offspring exposed toprenatal physical stress (Fujioka et al., 2006; Michelsen et al.,2007; Murmu et al., 2006).
Impoverished pattern of dendrite outgrowth and arbor-ization will have consequences for the neurons' ability tobuild up sufficient and efficient connections through incom-ing afferents; including reception of fewer excitatory andinhibitory inputs during critical stages of brain development.In the PFC it may relate to cortico-cortical inputs and midline
thalamus nuclei, which make synapses on dendrites ofpyramidal neurons in layer III (Kuroda et al., 1998; Wangand Shyu, 2004). In the hippocampus, this could impact onCA1 afferents from CA3 region as well as glutamatergic andGABA afferents from the medial septum (Colom et al., 2005;Kajiwara et al., 2008). These rearrangements in neuronalcircuitry may explain some aberrant PFC and hippocampusrelated tasks such as object recognition, spatial learning andworking memory deficits that have been reported in prenatalimmune challenge (Hava et al., 2006; Meyer et al., 2008b;Ozawa et al., 2006; Samuelsson et al., 2006). It is also possible

Fig. 9. Photomicrograph showing representative Golgi-Cox impregnated basilar dendrite from pyramidal neuron of layer V PFC at P60 in (A) Saline versus (B) LPStreated animals.
107M. Baharnoori et al. / Schizophrenia Research 107 (2009) 99–109
that the changes in prefrontal and hippocampal morphologymay underlie the prefrontal and hippocampal GABAergic andglutamatergic dysfunctions reported to be induced byprenatal immune challenges (Meyer et al., 2008b; Nyffeleret al., 2006).
Interestingly, at P10 there is significant decrease in thenumber of perisomal and proximal dendritic branches in PFCcompared with distal segments. In neonatal period, theproximal dendritic segments specifically receive inhibitoryinputs from fast-spiking, GABAergic interneurons (Chatto-padhyaya et al., 2004). It is noteworthy that neurodevelop-mental deficits of GABAergic interneurons along with gammaoscillation disruption have been reported in PFC of schizo-phrenia subjects and in animal models (bdul-Monim et al.,2007; Beasley and Reynolds, 1997; Spencer et al., 2003).Prenatal LPS treatment also resulted in significant decrease inspine structure (e.g. spine volume, surface area, length) in CA1at P10, the time of maximum spinogenesis in hippocampus(Fiala et al., 1998; Harris, 1999). Significant decrease in spinevolume implies a very early aberration in spine formation andsynaptogenesis in hippocampus. Decreased spine volumemight mean that the excitatory connections that are made atthis time are probably immature and unable to drive futuredevelopment of hippocampus connectivity that may explainthe persistent decrease in dendrite arborization and branch-ing during development.
In post-pubertal rats, analysis showed significant decreasein spine density of pyramidal neurons in layer V PFC in LPSgroup. Alterations in spine density may affect interactionsbetween glutamatergic and dopaminergic transmission inPFC and output of pyramidal neurons to sub-cortical targetssuch as mesolimbic dopamine areas (Ding et al., 2001). It isnoteworthy that DA depletion of PFC following ventraltegmental area lesions causes decrease in spine density inneurons of layer V mPFC (Wang and Deutch, 2007). Andconsistent with mesolimbic DA system deregulation, severalstudies in prenatal infection model show dopamine-relatedbehavioral deficits in psychostimulant-induced locomotion,PPI and latent inhibition (Fortier et al., 2007; Meyer et al.,2006; Ozawa et al., 2006; Romero et al., 2008; Zuckerman andWeiner, 2005).
The different pattern of morphological changes acrossages in the two brain structures induced by prenatal LPS couldbe due to the fact that the pyramidal neurons in the prefrontalcortex and CA1 region represent two distinct neuronal
populations in term of their embryonic site of origin in dorsaltelencephalon; neocortex originates from precursors in theanterior and lateral sites whereas posterior and medial sitesgive rise to hippocampus. Genetic regulation of the formationof these structures is also different; for example, Wnt3 andLEF1 gene expressions are necessary for hippocampus devel-opment while neocortex requires Pax6 expression for areaspecific gene expression and proper neurogenesis (Hebertand Fishell, 2008; Takahashi and Liu, 2006). Exposure todistinct neuronal input and extrinsic signaling moleculesthrough different local and long distance neuronal connec-tions may also generate significantly different pattern ofneuronal development and they may interact with LPS toproduce differential morphological alterations in the tworegions (Cenquizca and Swanson, 2007; Leone et al., 2008).Differences in the ontogenetic expression of brain derivedneurotrophic factor in neocortex compared to forebrainstructures like hippocampus also suggests different timecourse for neuronal maturations in the two structures(Friedman et al., 1991). It is also interesting to note thatwhile in both hippocampus and cortex, the sensitivity toinhibitory transmitter GABA appears prior to the sensitivity tothe excitatory transmitter glutamate, the time course ofreceptor development is significantly different in culturedembryonic neurons (Koller et al., 1990).
The cytoarchitectural changes reported here are reminis-cent of morphological changes reported in putative animalmodels for schizophrenia as well as in the brains ofschizophrenia patients. Post-mortem studies in schizophreniahave demonstrated altered number of dendritic branches inPFC layer III/V and decreased dendritic spine density in thedorsolateral PFC (Black et al., 2004; Broadbelt et al., 2002;Garey et al., 1998; Glantz and Lewis, 2000; Kalus et al., 2000).Significant changes in spine density and dendritic arbor andlength of pyramidal neurons have also been reported in somewidely-used animal models for schizophrenia such asneonatal ventral hippocampus-lesioned rats (Flores et al.,2005, 2007; Wedzony et al., 2005).
In conclusion, our results show that prenatal LPS challengeleads to morphological abnormality in pyramidal neuronsthat starts at very early postnatal days, with some abnormalfeatures persisting through to postnatal days while othersshowing a more dynamic pattern. Taking into account thatprenatal immune-challenged animals display significantdeficits in schizophrenia-related behaviors, we suggest that

108 M. Baharnoori et al. / Schizophrenia Research 107 (2009) 99–109
alterations in pyramidal neuronal morphology may be animportant mechanism underlying such behavioral changes.
Role of funding source
The research was funded by a team grant from the Canadian Institutes ofHealth Research (CIHR, ELA 80228) to L.K.S.
Contributors
Baharnoori M:Performed the experiments and data analysis and wrote the manuscript.Brake W:Helped with data analysis and comment on manuscript.Srivastava L.K:Designed the experiments and wrote the manuscript.
Conflict of interest
No conflict declared.
References
Ashdown, H., Dumont, Y., Ng, M., Poole, S., Boksa, P., Luheshi, G.N., 2006. Therole of cytokines in mediating effects of prenatal infection on the fetus:implications for schizophrenia. Mol. Psychiatry 11, 47–55.
Bauer, S., Kerr, B.J., Patterson, P.H., 2007. The neuropoietic cytokine family indevelopment, plasticity, disease and injury. Nat. Rev. Neurosci. 8, 221–232.
bdul-Monim, Z., Neill, J.C., Reynolds, G.P., 2007. Sub-chronic psychotomimeticphencyclidine induces deficits in reversal learning and alterations inparvalbumin-immunoreactive expression in the rat. J. Psychopharmacol.21, 198–205.
Beasley, C.L., Reynolds, G.P., 1997. Parvalbumin-immunoreactive neurons arereduced in the prefrontal cortex of schizophrenics. Schizophr. Res. 24,349–355.
Black, J.E., Kodish, I.M., Grossman, A.W., Klintsova, A.Y., Orlovskaya, D.,Vostrikov, V., Uranova, N., Greenough, W.T., 2004. Pathology of layer Vpyramidal neurons in the prefrontal cortex of patients with schizo-phrenia. Am. J. Psychiatry 161, 742–744.
Boksa, P., El-Khodor, B.F., 2003. Birth insult interacts with stress atadulthood to alter dopaminergic function in animal models: possibleimplications for schizophrenia and other disorders. Neurosci. Biobehav.Rev. 27, 91–101.
Broadbelt, K., Byne, W., Jones, L.B., 2002. Evidence for a decrease in basilardendrites of pyramidal cells in schizophrenic medial prefrontal cortex.Schizophr. Res. 58, 75–81.
Brown, A.S., Schaefer, C.A., Wyatt, R.J., Goetz, R., Begg, M.D., Gorman, J.M.,Susser, E.S., 2000. Maternal exposure to respiratory infections and adultschizophrenia spectrum disorders: a prospective birth cohort study.Schizophr. Bull. 26, 287–295.
Cai, Z.W., Pan, Z.L., Pang, Y., Evans, O.B., Rhodes, P.G., 2000. Cytokine inductionin fetal rat brains and brain injury in neonatal rats after maternallipopolysaccharide administration. Pediatr. Res. 47, 64–72.
Cannon, M., Clarke, M.C., 2005. Risk for schizophrenia–broadening theconcepts, pushing back the boundaries. Schizophr. Res. 79, 5–13.
Cenquizca, L.A., Swanson, L.W., 2007. Spatial organization of directhippocampal field CA1 axonal projections to the rest of the cerebralcortex. Brain Res. Rev. 56, 1–26.
Chattopadhyaya, B., Di, C.G., Higashiyama, H., Knott, G.W., Kuhlman, S.J.,Welker, E., Huang, Z.J., 2004. Experience and activity-dependentmaturation of perisomatic GABAergic innervation in primary visualcortex during a postnatal critical period. J. Neurosci. 24, 9598–9611.
Colom, L.V., Castaneda, M.T., Reyna, T., Hernandez, S., Garrido-Sanabria, E.,2005. Characterization of medial septal glutamatergic neurons and theirprojection to the hippocampus. Synapse 58, 151–164.
Dean, K., Murray, R.M., 2005. Environmental risk factors for psychosis.Dialogues Clin. Neurosci. 7, 69–80.
Deumens, R., Koopmans, G.C., Jaken, R.J., Morren, K., Comhair, T., Kosar, S.,Steinbusch, H.W., Den Bakker, C.G., Joosten, E.A., 2006. Stimulation ofneurite outgrowth on neonatal cerebral astrocytes is enhanced in thepresence of BDNF. Neurosci. Lett. 407, 268–273.
Ding, D.C., Gabbott, P.L., Totterdell, S., 2001. Differences in the laminar originof projections from the medial prefrontal cortex to the nucleusaccumbens shell and core regions in the rat. Brain Res. 917, 81–89.
Fatemi, S.H., Earle, J., Kanodia, R., Kist, D., Emamian, E.S., Patterson, P.H., Shi, L.,Sidwell, R., 2002a. Prenatal viral infection leads to pyramidal cell atrophyand macrocephaly in adulthood: implications for genesis of autism andschizophrenia. Cell Mol. Neurobiol. 22, 25–33.
Fatemi, S.H., Emamian, E.S., Kist, D., Sidwell, R.W., Nakajima, K., Akhter, P.,Shier, A., Sheikh, S., Bailey, K., 1999. Defective corticogenesis andreduction in Reelin immunoreactivity in cortex and hippocampus ofprenatally infected neonatal mice. Mol. Psychiatry 4, 145–154.
Fatemi, S.H., Emamian, E.S., Sidwell, R.W., Kist, D.A., Stary, J.M., Earle, J.A.,Thuras, P., 2002b. Human influenza viral infection in utero alters glialfibrillary acidic protein immunoreactivity in the developing brains ofneonatal mice. Mol. Psychiatry 7, 633–640.
Fatemi, S.H., Reutiman, T.J., Folsom, T.D., Huang, H., Oishi, K., Mori, S., Smee, D.F.,Pearce, D.A., Winter, C., Sohr, R., Juckel, G., 2008.Maternal infection leads toabnormal gene regulation and brain atrophy in mouse offspring: implica-tions for genesis of neurodevelopmental disorders. Schizophr. Res. 99,56–70.
Fiala, J.C., Feinberg, M., Popov, V., Harris, K.M., 1998. Synaptogenesis viadendritic filopodia in developing hippocampal area CA1. J. Neurosci. 18,8900–8911.
Flores, C., Wen, X., Labelle-Dumais, C., Kolb, B., 2007. Chronic phencyclidinetreatment increases dendritic spine density in prefrontal cortex andnucleus accumbens neurons. Synapse 61, 978–984.
Flores, G., Alquicer, G., Silva-Gomez, A.B., Zaldivar, G., Stewart, J., Quirion, R.,Srivastava, L.K., 2005. Alterations in dendritic morphology of prefrontalcortical and nucleus accumbens neurons in post-pubertal rats afterneonatal excitotoxic lesions of the ventral hippocampus. Neuroscience133, 463–470.
Fortier, M.E., Joober, R., Luheshi, G.N., Boksa, P., 2004. Maternal exposure tobacterial endotoxin during pregnancy enhances amphetamine-inducedlocomotion and startle responses in adult rat offspring. J. Psychiatr. Res.38, 335–345.
Fortier, M.E., Luheshi, G.N., Boksa, P., 2007. Effects of prenatal infection onprepulse inhibition in the rat depend on the nature of the infectiousagent and the stage of pregnancy. Behav. Brain Res. 181, 270–277.
Friedman, W.J., Olson, L., Persson, H., 1991. Cells that Express Brain-DerivedNeurotrophic Factor mRNA in the Developing Postnatal Rat Brain. Eur. J.Neurosci. 3, 688–697.
Fujioka, A., Fujioka, T., Ishida, Y., Maekawa, T., Nakamura, S., 2006. Differentialeffects of prenatal stress on the morphological maturation of hippo-campal neurons. Neuroscience 141, 907–915.
Garey, L.J., Ong, W.Y., Patel, T.S., Kanani, M., Davis, A., Mortimer, A.M., Barnes,T.R., Hirsch, S.R., 1998. Reduced dendritic spine density on cerebralcortical pyramidal neurons in schizophrenia. J. Neurol. Neurosurg.Psychiatry 65, 446–453.
Gibb, R., Kolb, B., 1998. A method for vibratome sectioning of Golgi-Coxstained whole rat brain. J. Neurosci. Methods 79, 1–4.
Gilmore, J.H., Jarskog, L.F., Swarooparani,V., Lauder, J.M., 2004.Prenatal infectionand risk for schizophrenia: IL-1?, IL-6, and TNF? Inhibit cortical neurondendrite development. Neuropsychopharmacology 29, 1221–1229.
Gilmore, J.H., Jarskog, L.F., Vadlamudi, S., 2005. Maternal poly I: C exposureduring pregnancy regulates TNF alpha BDNF, and NGF expression inneonatal brain and the maternal-fetal unit of the rat. J. Neuroimmunol.159, 106–112.
Glantz, L.A., Lewis, D.A., 2000. Decreased dendritic spine density onprefrontal cortical pyramidal neurons in schizophrenia. Arch. Gen.Psychiatry 57, 65–73.
Golan, H.M., Lev, V., Hallak, M., Sorokin, Y., Huleihel, M., 2005. Specificneurodevelopmental damage in mice offspring following maternalinflammation during pregnancy. Neuropharmacology 48, 903–917.
Harris, K.M., 1999. Structure, development, and plasticity of dendritic spines.Curr. Opin. Neurobiol. 9, 343–348.
Harrison, P.J., Weinberger, D.R., 2005. Schizophrenia genes, gene expression,and neuropathology: on the matter of their convergence. Mol. Psychiatry10, 40–68.
Hava, G., Vered, L., Yael, M., Mordechai, H., Mahoud, H., 2006. Alterations inbehavior in adult offspring mice followingmaternal inflammation duringpregnancy. Dev. Psychobiol. 48, 162–168.
Hebert, J.M., Fishell, G., 2008. The genetics of early telencephalon patterning:some assembly required. Nat. Rev. Neurosci.
Heinz, A., Romero, B., Gallinat, J., Juckel, G., Weinberger, D.R., 2003. Molecularbrain imaging and the neurobiology and genetics of schizophrenia.Pharmacopsychiatry 36 (Suppl 3), S152–S157.
Hikida, T., Jaaro-Peled, H., Seshadri, S., Oishi, K., Hookway, C., Kong, S., Wu, D.,Xue, R., Andrade, M., Tankou, S., Mori, S., Gallagher, M., Ishizuka, K.,Pletnikov, M., Kida, S., Sawa, A., 2007. Dominant-negative DISC1transgenic mice display schizophrenia-associated phenotypes detectedby measures translatable to humans. Proc. Natl. Acad. Sci. U. S. A. 104,14501–14506.
Jeohn, G.H., Kong, L.Y., Wilson, B., Hudson, P., Hong, J.S., 1998. Synergisticneurotoxic effects of combined treatments with cytokines in murineprimary mixed neuron/glia cultures. J. Neuroimmunol. 85, 1–10.
Jonakait, G.M., 2007. The effects of maternal inflammation on neuronaldevelopment: possible mechanisms. Int. J. Dev. Neurosci. 25, 415–425.

109M. Baharnoori et al. / Schizophrenia Research 107 (2009) 99–109
Kajiwara, R., Wouterlood, F.G., Sah, A., Boekel, A.J., Baks-te Bulte, L.T., Witter,M.P., 2008. Convergence of entorhinal and CA3 inputs onto pyramidalneurons and interneurons in hippocampal area CA1 — an anatomicalstudy in the rat. Hippocampus 18, 266–280.
Kalus, P., Muller, T.J., Zuschratter, W., Senitz, D., 2000. The dendriticarchitecture of prefrontal pyramidal neurons in schizophrenic patients.Neuroreport 11, 3621–3625.
Khashan, A.S., Abel, K.M., McNamee, R., Pedersen, M.G., Webb, R.T., Baker, P.N.,Kenny, L.C., Mortensen, P.B., 2008. Higher risk of offspring schizophreniafollowing antenatal maternal exposure to severe adverse life events. Arch.Gen. Psychiatry 65, 146–152.
Koller, H., Siebler, M., Schmalenbach, C., Muller, H.W., 1990. GABA andglutamate receptor development of cultured neurons from rat hippo-campus, septal region, and neocortex. Synapse 5, 59–64.
Kolluri, N., Sun, Z., Sampson, A.R., Lewis, D.A., 2005. Lamina-specificreductions in dendritic spine density in the prefrontal cortex of subjectswith schizophrenia. Am. J. Psychiatry 162, 1200–1202.
Kuroda, M., Yokofujita, J., Murakami, K., 1998. An ultrastructural study of theneural circuit between the prefrontal cortex and the mediodorsalnucleus of the thalamus. Prog. Neurobiol. 54, 417–458.
Labelle, C., Leclerc, N., 2000. Exogenous BDNF, NT-3 and NT-4 differentiallyregulate neurite outgrowth in cultured hippocampal neurons. Brain Res.Dev. Brain Res. 123, 1–11.
Leone, D.P., Srinivasan, K., Chen, B., Alcamo, E., McConnell, S.K., 2008. Thedetermination of projection neuron identity in the developing cerebralcortex. Curr. Opin. Neurobiol. 18, 28–35.
Libbey, J.E., Sweeten, T.L., McMahon, W.M., Fujinami, R.S., 2005. Autisticdisorder and viral infections. J. Neurovirol. 11, 1–10.
Marenco, S., Weinberger, D.R., 2000. The neurodevelopmental hypothesis ofschizophrenia: following a trail of evidence from cradle to grave. Dev.Psychopathol. 12, 501–527.
Marx, C.E., Jarskog, L.F., Lauder, J.M., Lieberman, J.A., Gilmore, J.H., 2001.Cytokine effects on cortical neuron MAP-2 immunoreactivity: Implica-tions for schizophrenia. Biol. Psychiatry 50, 743–749.
McClellan, J.M., Susser, E., King, M.C., 2006. Maternal famine, de novomutations, and schizophrenia. JAMA 296, 582–584.
McDonald, C., Murray, R.M., 2000. Early and late environmental risk factorsfor schizophrenia. Brain Res. Brain Res. Rev. 31, 130–137.
Meyer, U., Feldon, J., Schedlowski, M., Yee, B.K., 2006. Immunological stress atthe maternal-foetal interface: a link between neurodevelopment andadult psychopathology. Brain Behav. Immun. 20, 378–388.
Meyer, U., Yee, B.K., Feldon, J., 2007. The neurodevelopmental impact ofprenatal infections at different times of pregnancy: the earlier theworse? Neuroscientist 13, 241–256.
Meyer, U., Nyffeler, M., Schwendener, S., Knuesel, I., Yee, B.K., Feldon, J., 2008a.Relative prenatal and postnatal maternal contributions to schizophrenia-related neurochemical dysfunction after in utero immune challenge.Neuropsychopharmacology 33, 441–456.
Meyer, U., Nyffeler, M., Yee, B.K., Knuesel, I., Feldon, J., 2008b. Adult brain andbehavioral pathological markers of prenatal immune challenge duringearly/middle and late fetal development in mice. Brain Behav. Immun.22, 469–486.
Michelsen, K.A., Van den Hove, D.L., Schmitz, C., Segers, O., Prickaerts, J.,Steinbusch, H.W., 2007. Prenatal stress and subsequent exposure tochronic mild stress influence dendritic spine density and morphology inthe rat medial prefrontal cortex. BMC Neurosci. 8, 107.
Murmu, M.S., Salomon, S., Biala, Y., Weinstock, M., Braun, K., Bock, J., 2006.Changes of spine density and dendritic complexity in the prefrontalcortex in offspring of mothers exposed to stress during pregnancy. Eur. J.Neurosci. 24, 1477–1487.
Neumann, H., Schweigreiter, R., Yamashita, T., Rosenkranz, K., Wekerle, H.,Barde, Y.A., 2002. Tumor necrosis factor inhibits neurite outgrowth and
branching of hippocampal neurons by a rho-dependent mechanism.J. Neurosci. 22, 854–862.
Ning, H., Wang, H., Zhao, L., Zhang, C., Li, X.Y., Chen, Y.H., Xu, D.X., 2008.Maternally-administered lipopolysaccharide (LPS) increases tumornecrosis factor alpha in fetal liver and fetal brain: its suppression bylow-dose LPS pretreatment. Toxicol. Lett. 176, 13–19.
Nyffeler, M., Meyer, U., Yee, B.K., Feldon, J., Knuesel, I., 2006. Maternalimmune activation during pregnancy increases limbic GABAA receptorimmunoreactivity in the adult offspring: implications for schizophrenia.Neuroscience 143, 51–62.
Ozawa, K., Hashimoto, K., Kishimoto, T., Shimizu, E., Ishikura, H., Iyo, M., 2006.Immune activation during pregnancy in mice leads to dopaminergichyperfunction and cognitive impairment in the offspring: a neurodeve-lopmental animal model of schizophrenia. Biol. Psychiatry 59, 546–554.
Patterson, P.H., 2002. Maternal infection: window on neuroimmune interac-tions in fetal brain development and mental illness. Curr. Opin. Neurobiol.12, 115–118.
Paxinos, G., Watson, C., 2007. The rat brain in stereotaxic coordinates.Academic Press, New York.
Penner, J.D., Brown, A.S., 2007. Prenatal infectious and nutritional factors andrisk of adult schizophrenia. Expert Rev. Neurother. 7, 797–805.
Romero, E., Ali, C., Molina-Holgado, E., Castellano, B., Guaza, C., Borrell, J.,2007. Neurobehavioral and immunological consequences of prenatalimmune activation in rats. Influence of antipsychotics. Neuropsycho-pharmacology 32, 1791–1804.
Romero, E., Guaza, C., Castellano, B., Borrell, J., 2008. Ontogeny ofsensorimotor gating and immune impairment induced by prenatalimmune challenge in rats: implications for the etiopathology ofschizophrenia. Mol. Psychiatry.
Samuelsson, A.M., Jennische, E., Hansson, H.A., Holmang, A., 2006. Prenatalexposure to interleukin-6 results in inflammatory neurodegeneration inhippocampus with NMDA/GABA(A) dysregulation and impaired spatiallearning. Am. J. Physiol. Regul. Integr. Comp. Physiol. 290, R1345–R1356.
Shi, L., Fatemi, S.H., Sidwell, R.W., Patterson, P.H., 2003. Maternal influenzainfection causes marked behavioral and pharmacological changes in theoffspring. J. Neurosci. 23, 297–302.
Spencer, K.M., Nestor, P.G., Niznikiewicz, M.A., Salisbury, D.F., Shenton, M.E.,McCarley, R.W., 2003. Abnormal neural synchrony in schizophrenia.J. Neurosci. 23, 7407–7411.
Takahashi, H., Liu, F.C., 2006. Genetic patterning of the mammaliantelencephalon by morphogenetic molecules and transcription factors.Birth Defects Res C Embryo Today 78, 256–266.
Urakubo, A., Jarskog, L.F., Lieberman, J.A., Gilmore, J.H., 2001. Prenatalexposure to maternal infection alters cytokine expression in theplacenta, amniotic fluid, and fetal brain. Schizophr. Res. 47, 27–36.
Wang, C.C., Shyu, B.C., 2004. Differential projections from the mediodorsaland centrolateral thalamic nuclei to the frontal cortex in rats. Brain Res.995, 226–235.
Wang, H.D., Deutch, A.Y., 2007. Dopamine depletion of the prefrontal cortexinduces dendritic spine loss: reversal by atypical antipsychotic drugtreatment. Neuropsychopharmacology.
Wedzony, K., Fijal, K., Mackowiak, M., 2005. Alterations in the dendriticmorphology of prefrontal pyramidal neurons in adult rats after blockadeof NMDA receptors in the postnatal period. Brain Res. 1062, 166–170.
Zuckerman, L., Rehavi, M., Nachman, R., Weiner, I., 2003. Immune activationduring pregnancy in rats leads to a postpubertal emergence of disruptedlatent inhibition, dopaminergic hyperfunction, and altered limbicmorphology in the offspring: a novel neurodevelopmental model ofschizophrenia. Neuropsychopharmacology 28, 1778–1789.
Zuckerman, L, Weiner, I, 2005. Maternal immune activation leads to behavioraland pharmacological changes in the adult offspring. J. Psychiatr. Res. 39,311–323.