potentiation of 5-fluorouracil-leucovorin activity by a2a ... · interfaced with an ibm/ps2...
TRANSCRIPT

Vol. 1, 33-40, January 1995 Clinical Cancer Research 33
Potentiation of 5-Fluorouracil-Leucovorin Activity by
a2a-Interferon in Colon Adenocarcinoma Xenografts1
Janet A Houghton,2 Pamela J. Cheshire,
Christopher L. Morton, and Clinton F. StewartDepartment of Molecular Pharmacology [J. A. H., P. J. C., C. L. M.]
and Pharmaceutical Department [C. F. S.], St. Jude Children’s
Research Hospital, Memphis, Tennessee 38101
ABSTRACT
Previous studies using cultured colon adenocarcinoma
cells demonstrated that a mixture of the diastereoiosomers ofthe biologically active (6S) and inactive (6R) forms of (6RS)
leucovorin or 5-formyl-H4PteGlu (LV) and recombinant hu-man a2a-interferon (rLFN-a2a) in combination significantly
increased the cytotoxicity of 5-fluorouradil (FUra) (by 10-14-
fold) whereas FUra combined with single modulators was less
potentiated (3-fold). Maximum cytotoxicity was achieved with48-h drug exposures when drugs were applied continuously,and modulatory rIFN-a2a concentrations were obtained at�50 International units (IU)/ml. We therefore examined
whether such interactions could occur in vivo using HXGCI
clTKc3 colon adenocarcinoma xenografts, deficient in thy-
midine salvage. Potentiation of FUra activity was significantly
greater when FUra was combined with both LV and rIFN-a2a
in comparison to the use of single modulators using a 5-dayschedule for 3 courses. In mice receiving LV, the maximum
level of potentiation of FUra-induced growth inhibition was
independent of the rIFN-a2a dose between 25,000 and 600,000
lu examined in contrast to rIFN-a2a used as a single modu-lator. After administration of 25,000 IU rWN-a2a, plasmarIFN-ot2a concentrations �50 lU/mI were maintained for 6-8h, comparable to exposure times achievable clinically. Dataindicate that intennittent rWN-a2a exposure potentiatesFUra-LV activity in vivo. The efficacy of FUra combined with
dual versus single modulators will thus be of importance to
evaluate in randomized phase III clinical trials in patients withcolorectal cancer.
INTRODUCTIONWe have previously shown significant potentiation (10-
14-fold) of the cytotoxic activity of FUra3 against human colon
adenocarcinoma cell lines using dual modulation with LV and
rIFN-a2a (1). Cytotoxic effects were considerably greater than
with the use of single modulators, where a 3-fold enhancement
of FUra efficacy was obtained. Potentiation of FUra-LV cyto-
toxicity was influenced by the concentration of rIFN-a2a, and
modulation was obtained at clinically achievable rIFN-a2a con-
centrations (�50 IU/ml). Maximum interaction among FUra,
Received 6/24/94; accepted 7/26/94.
t Supported by National Cancer Institute Awards CA-23099 and CA-
21765 (Cancer Center Support Grant) and by the American Lebanese
Syrian Associated Charities.
LV, and nIFN-a2a required a minimum period of 48 h of
continuous drug exposure (2).
The mechanism of FUra-LV interaction is well understood
in the human colon adenocarcinoma cell lines and xenografts
used in this laboratory (3-6). Under conditions of exposure to
physiological folate, LV enhances FUra cytotoxicity by target-
ing FUra to the TS locus. Leucovonin has induced elevation of
critical intracellular pools of 5,10-methylenetetrahydrofolate
that enhance binding of the FUra metabolite FdUMP to TS
within a covalent ternary complex. Enhanced complex stability
mediated by elevated reduced folate pools further contribute to
potentiation of the degree and duration of TS inhibition and to
depleted dTFP pools, while drug resistance has been associated,
in pant, to inadequate pools of 5,10-methylenetetrahydrofolate
(3).
Mechanistically, the locus of the interaction among FUra,
LV, and rIFN-a2a has been identified in the GC3/cl colon
adenocarcinoma cell line (2). FUra cytotoxicity in the presence
of modulating agents was reversible by dThd. However, TS was
determined not to be the primary drug interaction site since (a)
direct assays of TS indicated no significant difference in the
level, activity, or accumulation among FUra ± LV ± rIFN-a2a
treatment groups; (b) nIFN-a2a did not potentiate the cytotoxic
activity of 10-pnopargyl 5,8-dideazafolate, a TS inhibitory
quinazoline; and (c) in TS-deficient cells rIFN-a2a cytotoxicity
was not influenced by the concentration of dThd. Data thus
indicated the requirement for the nucleotide analogue (i.e.,
5-fluoropyrimidine) in the interaction, and this suggested that
potentiation of FUra by rIFN-a2a may be a consequence of
increased incorporation of fraudulent nucleotide into DNA. The
level of DNA single-strand and double-strand breaks obtained
with FUra that were enhanced when combined with single
modulators and further increased in the presence of dual mod-
ulators paralleled the cytotoxic activity of FUra-LV-rIFN-a2a
combinations. Thus, rIFN-a2a may exert its effects by enhance-
ment of FUra base excision or incorporation into DNA, events
that subsequently become influenced by TS inhibition and
dThd-less stress and are further potentiated by LV (2).
Clinically, FUra-LV combinations have demonstrated sig-
nificantly increased response rates (33-48%) in comparison to
2 To whom requests for reprints should be addressed, at Department of
Molecular Pharmacology, St. Jude Children’s Research Hospital, P.O.Box 318, 322 North Lauderdale, Memphis, TN 38101.3 The abbreviations used are: FUra, 5-fluorouracil; LV, a mixture of the
diastereoiosomers of the biologically active (65) and inactive (6R) forms
of (6R5) leucovorin or 5-formyl-H4PteGlu; rIFN-a2a, recombinant hu-
man cr2a-interferon; IFN, interferon; FdUMP, 5-fluoro-2’ -deoxyunidy-
late; TS, thymidylate synthase; TK, thymidine kinase; dThd, deoxythy-
midine; AUC, area under curve; CR, complete response; PR, partial
response of �50% reduction in tumor volume that also includes CRs; C,
no regrowth during the observation period; CLs, clearance; IU, Inter-
national units.
Research. on April 25, 2020. © 1995 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

34 FUra-LV Activity in Colon Adenocarcinoma Xenografts
the use of FUra administered alone (7-15%) when evaluated in
phase III randomized clinical trials conducted in patients pre-
senting with colon cancer (7-10). In addition, significant in-
creases in time to disease progression (8-10) and in patient
survival (8, 9) have been indicated. Thus, by building on this
combination with additional therapeutic approaches, response
rates and survivorship may be improved. rIFN-a2a has been
evaluated for this purpose, commencing in single institutional
trials. Initially, FUra and rIFN-a2a combinations were evalu-
ated in previously untreated colon carcinoma patients and re-
sponse rates (26-63%) greater than those anticipated for FUra
treatment alone, and equal to those obtained with FUra-LV
combinations, were reported (1 1-13). In a subsequent phase II
multiinstitutional trial, Wadler et al. (14) confirmed a high
response rate of 42%. The potential for the interaction of FUra
and rIFN-a2a at a locus distinct from that of FUra and LV,
along with their nonoverlapping toxicities, suggested that rIFN-
a2a would be of importance to evaluate in combination with
FUra-LV. Subsequently, Grem et al. (15) evaluated the triple
combination in previously untreated colon carcinoma patients
and determined a response rate of 45%.
As demonstrated from both preclinical and clinical studies,
the use of rIFN-a2a in combination with FUra-LV clearly has
potential for improving therapeutic outcome in patients with
colon carcinoma. However, several key issues remain to be
addressed that will determine the utility of rIFN-a2a as a mod-
ulator of FUra-LV action. The minimum dose of rIFN-a2a that
can be used to achieve modulatory plasma concentrations must
be determined. In colorectal carcinoma patients, the rIFN-a2a
dose may be limited by fatigue and neurological toxicities (16,
1 7) and also by elevation in the plasma concentrations of FUra
(15). The duration of rlFN-a2a exposure must be sufficient to
enhance FUra-LV action, and it is evident from preclinical
studies that modulation has not been achieved with single, short
(2-h) exposures (2). The interdependence of these two parame-
tens on FUra-LV modulation at clinically achievable plasma
concentrations of rIFN-a2a is of importance for clinical therapy.
Finally, whether the effect of dual modulation versus the effect
of single modulators significantly enhances the therapeutic ac-
tivity of FUra will impact on the application of rIFN-a2a in
clinical trials.
Previously, we have extensively characterized the thera-
peutic activity of FUra-LV combinations in human colon ade-
nocarcinoma xenografts maintained in immune-deprived mice
(18). We have therefore used these models to address some
of the key issues outlined above regarding the influence of
rIFN-a2a action on the in vito activity of FUra or FUra-LV
combinations. In the studies presented, we have examined the
therapeutic utility of FUra, FUra-LV, FUra-rIFN-a2a, and
FUra-LV-rIFN-a2a combinations. The xenognaft models used
were the parental HxGC3/cl colon adenocarcinoma that sal-
vages dThd and HxGC3/clTKc3, a subline selected for its
inability to salvage dThd because of a deficiency in TK
HxGC3/clTKc3 tumors may be more representative of the
situation in humans, where reduced dThd salvage capability due
to considerably lower plasma dThd concentrations has been
demonstrated (18, 19). Specifically, we have examined (a) the
influence of dose of rIFN-a2a on FUra or FUra-LV-induced
antitumor activity in mice; (b) pharmacokinetics of nIFN-a2a
given to mice; and (c) duration of systemic exposure to rIFN-
a2a at modulatory dose levels.
MATERIALS AND METHODS
Immune Deprivation of Mice. Female 4-week-oldCBA/CaJ mice (The Jackson Laboratory, Bar Harbor, ME) were
immune deprived by thymectomy and subjected 3 weeks later to
whole-body irradiation (950 cGy) using a ‘37Cs source. Mice
received 3 X 106 nucleated bone marrow cells within 6-8 h of
irradiation (18).
Tumor Lines. HxGC�/c1 was originally derived from a
poorly differentiated adenocarcinoma of the colon that was
established as a xenograft in immune-deprived mice (HxGC3).
Subsequently, the line was cultured and cloned (GC3/cl; Refs.
20-22) and reestablished as a xenograft in mice (HxGC3/cl).
HxGC3/clTKc3, a TK-deficient variant of HxGC�/cl was
initially selected from GC3/cl in tissue culture (21, 22), and
inoculated s.c. into immune-deprived mice to produce the xc-
nografted line. All tumors were maintained as xenografts by
passage in the s.c. space of female CBA/CaJ immune-deprived
mice.
Growth Inhibition Studies. Mice bearing bilateral s.c.
tumors were randomized and received drugs when the tumors
had reached a diameter of “0.5 cm. Tumor response was
determined at 7-day intervals using digital calipers (Maxcal)
interfaced with an IBM/PS2 microcomputer. Two perpendicular
diameters were used to compute volumes (23). Growth inhibi-
tion was calculated from the difference in days required for
treated tumors to grow to four times their volume at the start of
treatment as compared with untreated controls. Each treatment
group comprised six tumor-bearing mice. Relative tumor vol-
umes were calculated using the formula RTV = V,JV�, where V�
represents the tumor volume on day X and V(, indicates the
tumor volume at the initiation of treatment. Volume regressions
were also determined and expressed as CR, PR or C, and
maximum percentage of treated/control ratio.
Formulation and Administration of Drugs. FUra (50
mg/ml), obtained from Solo-Pak Laboratories (Franklin Park,
IL) as a pharmaceutical preparation, was diluted to a concen-
tration of 15 mg/mI in sterile 0.9% saline. It was given to mice
by iv. bolus injection weekly via the tail vein for three courses
(days 1, 8, and 15) after randomization of the animals. The dose
used was 75 mg/kg (“�225 mg/m2) given in a volume of 0.1
ml/20 g body weight and constituted the maximum tolerated
dose on that schedule as described previously (18).
Leucovonin, purchased from Sigma Chemical Co. (St.
Louis, MO), was given to mice by i.v. bolus injection daily at a
dose of 500 mg/m2 on days 1-5, 8-12, and 15-19 immediately
before FUra commencing on the first day of FUra injection as
published previously (18). Leucovorin was dissolved in 0.9%
sterile saline at a concentration of 23.6 mg/ml, and after addi-
tional filtration was injected at volumes of 0.17-0.27 ml/20 g
body w as described previously (18). The dose relationship
between mg/kg and mg/m2 body-surface area was determined
for individual mice as reported previously (5, 6, 24).
rIFN-a2a was obtained, in part, as a generous gift from
Hoffmann La Roche Inc., Nutley, NJ. The lyophilized powder
representing 18 X 10� IU rIFN-a2a was initially dissolved in 1
Research. on April 25, 2020. © 1995 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

Clinical Cancer Research 35
ml sterile water. Alternatively the pharmaceutical formulation
(Roferon A; 6 X 106 IU/ml) was purchased. Further dilutions
were made in sterile 0.9% saline, and the agent was given i.p. to
mice on days 1-5, 8-12, and 15-19 in volumes of 0.1 ml!
10 g body weight at dose levels ranging from 25,000 to 600,000
lU/mouse. A 25,000-lU dose of nIFN-a2a in mice was equiva-
lent to 3.3 X 106 IU/m2 or 5.7 X 106 IU in humans. a2a-
Interferon was given immediately before leucovonin. Because of
the lack of interaction between high concentrations of rIFN-a2a
and LV in cultured cells in the absence of FUra (1, 2) and the
lack of antitumor activity or host toxicity of the combination in
initial studies in mice, the agents were examined in combination
in vivo as a control group.
Statistical Analyses of Antitumor Activity. The results
of studies were evaluated and compared by one-way analysis of
variance using the number of days required to reach 4 times the
original tumor volume as the dependent variable followed by the
Newman-Keuls range test for differences between means (18,
25). Only tumors from mice that survived the entire study were
included in the analyses, and any tumor that failed to reach 4
times its original volume was assigned a default value of 140
days for the maximum duration of the study. The percentage of
tumors showing partial and/or complete regressions, along with
the extent of regression, were also analyzed. Data are presented
as the mean and SE.
Pharmacokinetic Analyses. Plasma concentrations of
nIFN-a2a were determined following i.p. injection of non-tu-
mon-beaning immune-deprived mice at dose levels of 25,000 and
200,000 lU/dose. Time points, using 4 mice at each point,
ranged from 0.25 to 6 h following nIFN-a2a administration.
Additionally, groups of mice receiving 25,000 IU nIFN-a2a on
day 1 were injected with the same dose level of rIFN-cz2a at
24-h intervals over a period of 5 days. On day 5, samples were
obtained between time 0 (just before injection) and 6 h (4
mice/point) after rIFN-a2a administration. Blood was drawn by
cardiac puncture into hepaninized syringes and plasma prepared
(18).
Plasma concentrations of rIFN-a2a were measured by an
adaptation of an enzyme immunoradiometnic assay (Celltech
Limited, Berkshire, United Kingdom; Ref. 26). Calibrators were
prepared using pure rIFN-a2a (Hoffman La Roche Inc.) in
pooled mouse plasma over a concentration range of 25-2500
IU,’ml. The lower limit of sensitivity of the assay was 15 IU!ml.
A one-compartment model was fit to the nIFN-ot2a plasma
concentration-time data by the use of maximum likelihood
estimation (27). Parameters estimated included volume of the
central compartment (Vd�) and elimination rate constant (Ke).
Calculated parameters included t112, AUC versus time curve,
maximum drug concentration, systemic CLs, and length of time
plasma concentrations of nIFN-cz2a exceeded 50 IU/ml.
For purposes of estimating systemic exposure to rIFN-a2a
in humans from existing clinical dosing schedules, a simulation
analysis was performed. A one-compartment model was fit to
concentration versus time data from a previously published
study in humans (28). The parameters of Vd� and Ke were used
to simulate cumulative nIFN-a2a systemic exposure for two
clinically relevant dosage schedules [i.e., 5 X 106 IU!m2 daily
for S consecutive days (15) and 9 X 106 IU on alternate days for
three doses; (Ref 17)].
RESULTSPotentiation of FUra-LV Activity by rIFN-a2a in Vivo.
Previous reports in the literature indicated that a standard dose
of 200,000 IU IFN-a in mice induced growth inhibition of
xenognafted tumors (29, 30) or was modulatory to the action of
other agents (30, 31). Consequently, this dose level of rIFN-a2a
was initially evaluated for its modulatory effects on FUra or
FUra-LV activity in mice beaning HxGC3/clTKc3 xenografts,
deficient in dThd salvage (Fig. 1, left). In mice receiving no
treatment or LV-rIFN-a2a only, tumors grew at similar rates
and reached four times their pretreatment size at similar times
(35 and 38 days, respectively), indicating no direct effect of
LV-rIFN-a2a on tumor growth. When FUra was given once
weekly for 3 weeks, volume regressions were obtained and
growth was delayed by 33.5 days relative to controls. When
FUra was combined with rIFN-a2a or LV as single modulators,
volume regressions increased and further increases in growth
delay were obtained. However, when FUra was combined with
both modulators, additional and significant decreases in tumor
volume and further enhancement of growth inhibition (P <
0.001) was evident, effects considerably greater than those
achieved with single modulators (Fig. 1).
Since data obtained with cultured GC/cl cells had mdi-
cated that FUra-LV-rIFN-a2a cytotoxicity was reversible by
dThd (1, 2), modulation of FUra activity was subsequently
determined in HxGC�Jc1 xenografts capable of dThd salvage
(Fig. 1, right). In this experiment, the dose level of rIFN-a2a
was increased by 3-fold to 600,000 IU, although drug adminis-
tration schedules remained the same. In contrast to the HxGC/
clTKc3 xenognafts, no volume regressions or growth inhibi-
tion were induced in HxGC3!cl tumors when FUra was
combined with dual modulators, confirming results obtained
with cultured cells.
Influence of rIFN-a2a Dose on FUra ± LV Modulation.Since significant potentiation of the in vivo activity of FUra had
been obtained in HxGC3!clTKc3 xenografts by dual modula-
tion with rIFN-a2a administered at a dose of 200,000 IU, the
influence of nIFN-a2a dose (systemic exposure) on therapeutic
activity was examined. In these experiments, the rIFN-a2a dose
was increased to 600,000 IU to examine whether FUra or
FUra-LV activity could be potentiated further. In addition, the
dose level of rIFN-a2a was reduced to 25,000 IU (considered to
be a low dose level in mice; Ref. 29) to determine the minimum
dose exhibiting a modulatory effect. Data for rIFN-ct2a com-
bined with FUra alone as a single modulator, or combined with
LV in dual modulation studies, are shown in Tables 1 and 2 and
Fig. 2.
Tumor Growth Inhibition. The time required in days
for tumors to reach four times the volume at the initiation of
treatment has been compared for control and all drug-treated
groups (Table 1). The differences between untreated (42.1 days)
and FUra-treated (69.6 days) tumors, or between FUra-treated
and FUra-LV-treated (88.6 days) groups were significantly dif-
fenent (P < 0.05). When the efficacy of FUra treatment was
compared with on without rIFN-a2a, the time required to reach
the value at four times the volume ranged from 77.1 to 118.8
days, significantly different at rIFN-a2a doses of 25,000 and
600,000 IU (P < 0.001). Modulation of FUra activity by rIFN-
Research. on April 25, 2020. © 1995 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

HxGC3 /clTKc3
ci,E:�
0
E
ci,>
ccici)
a:
Group
ControlFUra
FUra-nIFN-a2a
Dose of rIFN-ct2a (lU) Days to 4X ± SE
P
<0.001
<0.05
36 FUra-LV Activity in Colon Adenocarcinoma Xenografts
0.1
0 2 4 6 8101214161820 0123456789
Weeks Weeks
Fig. I Modulation by rIFN-a2a of the activity of FUra or FUra-LV against HxGC3/cITK c3 (left) or HxGC3/cl (rig/it) xenografts maintained in
immune-deprived mice. 0, control; #{149},LV-rlFN-a2a; V. FUra; V, FUra-rIFN-a2a; 0, FUra-LV; #{149},FUra-LV-rIFN-a2a. Six mice bearing bilateral
tumors were randomized to each group, and results represent the mean ± SD for tumor volumes determined at each time point. Drug treatments were
conducted as follows: FUra, 75 mg/kg iv. days 1,8, and 15; LV, SOt) mg/rn2 iv. daily on days 1-5, 8-12, and 15-19: rIFN-#{128}s2a, 2(X),0(X) lU
(HxGC�/TK c3) or 600,0()() IU (HxGC�/cl) i.p. daily on days 1-5, 8-12, and 15-19. Tumor volumes were determined at weekly intervals.
Table I Delay in tumor growth induced by FUra-LV-rlFN-x2a combinations in HxGC�/clTK c3 xenografts”
Days to 4X” � ri � Group Days_to 4X ± SE N
42.1 ± 3.5 50 FUra 69.6 ± 3.2 29
69.6 ± 3.2 29 FUra-LV 88.6 ± 4.1 52
FUra-LV-rIFN-a2a
�1 Days to 4X ± SE n P
25,0(R) 96.5 �: 5.4 35 1 15.0 ± 4.7 35 <0.05
20(),0()() 77.1 � 5.4 10 1 1 1.2 ± 8.3 8 <0.001
600,0(X) 118.8 ± I 1.5 10 124.2 ± 7.9 12 >0.05
(‘ FUra vs. FUra-rlFN-u2a, P < 0.001 at 25,0(X)-lU and 60(),0()0-IU doses; FUra-LV i’s. FUra-rlFN-a2a, P < (1.05 at 600,000-lU dose; FUra-LV
vs. FUra-LV-rlFN-a2a (all doses), P < 0.05.
I, Days to 4X, time required to reach 4 times the pretreatment volume.
� P, comparison of FUra-rlFN-a2a with FUra-LV-rIFN-a2a at equivalent rIFN-cs2a dose levels.
cs2a alone appeared to be dependent on dose and was greatest at
the highest dose level of rIFN-a2a used. Data for FUra-LV
activity compared with FUra-rlFN-a2a responses were similar
(P > 0.05) at the 25,000- and 200,000-lU dose levels of rIFN-
a2a (P > 0.05), but were also greater at the high rIFN-a2a dose
level (P < 0.05). When FUra was combined with both modu-
lators however, days required to reach four times the volume
increased (range, I I I .2-1 24.2 days) and was independent of the
rIFN-a2a dose. Comparison of FUra-LV-rIFN-a2a responses
with the response to FUra-LV indicated significant differences
over the entire dose range of rIFN-a2a examined (P < 0.05).
Similarly, a comparison of FUra-LV-rIFN-a2a responses with
the responses to FUra-rIFN-a2a at equivalent rIFN-a2a dose
levels indicated increased FUra activity in the presence of dual
modulators at doses of rIFN-a2a � 200,000 IU (P < 0.05), but
not at the 600,000-lU rIFN-a2a dose.
Volume Regressions. The maximum percentage of TIC
for all treatment groups is shown in Fig. 2, and the percentage
of PR and CR responses for tumors that did not regrow during
C are presented in Table 2. FUra treatment alone induced 62%
PR that was increased significantly (P < 0.01) to 83% PR, 12%
CR, and 10% C in the presence of LV. When FUra was corn-
bined with rIFN-a2a, volume regressions were dependent on the
rIFN-a2a dose, being significantly greater at the 600,000-lU
dose of rIFN-a2a only (P < 0.001) in comparison to FUra or
FUra-LV treatment groups. Additionally, FUra-LV-rIFN-a2a-
treated tumors were smaller and demonstrated less rIFN-a2a
dose dependency, but were significantly smaller than in the
FUra-LV-treated group, only at the highest dose level of rIFN-
a2a (P < 0.01). However, when FUra-rIFN-a2a activity was
compared either in the absence or in the presence of LV at
equivalent rIFN-a2a dose levels, tumor volume regressions
Research. on April 25, 2020. © 1995 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
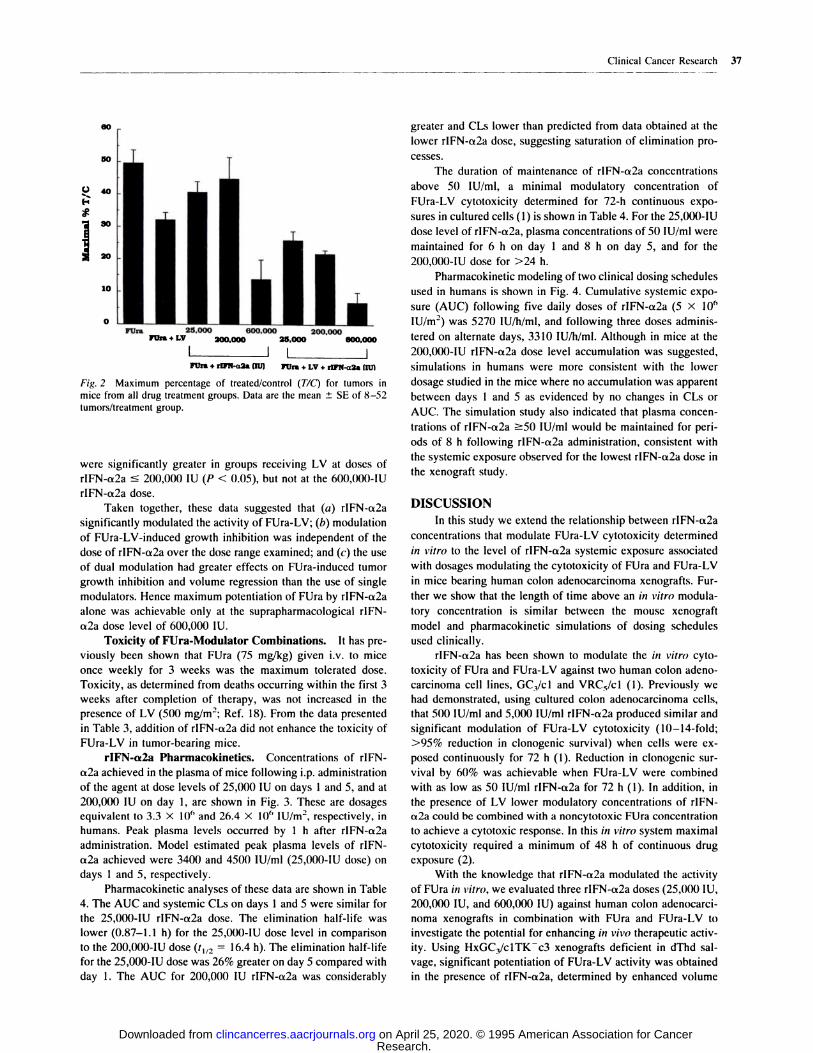
00
501
40
IC.)
I 20
1�
10
0 nFUrs
RhI � 200.000
200.000 25.000 000.000
I IL IFUrs + rlPN-a2a (flJ) FUrs + LV + riFN-u2a Intl
Fig. 2 Maximum percentage of treated/control (TIC) for tumors in
mice from all drug treatment groups. Data are the mean ± SE of 8-52
tumors/treatment group.
Clinical Cancer Research 37
were significantly greater in groups receiving LV at doses of
rIFN-ct2a � 200,000 IU (P < 0.05), but not at the 600,000-lU
rIFN-a2a dose.
Taken together, these data suggested that (a) rIFN-a2a
significantly modulated the activity of FUra-LV; (b) modulation
of FUra-LV-induced growth inhibition was independent of the
dose of rIFN-a2a over the dose range examined; and (c) the use
of dual modulation had greater effects on FUra-induced tumor
growth inhibition and volume regression than the use of single
modulators. Hence maximum potentiation of FUra by rIFN-a2a
alone was achievable only at the suprapharmacological rIFN-
a2a dose level of 600,000 IU.
Toxicity of FUra-Modulator Combinations. It has pre-
viously been shown that FUra (75 mg/kg) given i.v. to mice
once weekly for 3 weeks was the maximum tolerated dose.
Toxicity, as determined from deaths occurring within the first 3
weeks after completion of therapy, was not increased in the
presence of LV (500 mg/rn2; Ref. 18). From the data presented
in Table 3, addition of rlFN-a2a did not enhance the toxicity of
FUna-LV in tumor-bearing mice.
rIFN-a2a Pharmacokinetics. Concentrations of rIFN-
a2a achieved in the plasma of mice following i.p. administration
of the agent at dose levels of 25,000 IU on days I and 5, and at
200,000 IU on day 1, are shown in Fig. 3. These are dosages
equivalent to 3.3 x l0� and 26.4 X 106 IU/m2, respectively, in
humans. Peak plasma levels occurred by 1 h after rIFN-a2a
administration. Model estimated peak plasma levels of rIFN-
a2a achieved were 3400 and 4500 IU/ml (25,000-lU dose) on
days 1 and 5, respectively.
Pharmacokinetic analyses of these data are shown in Table
4. The AUC and systemic CLs on days 1 and S were similar for
the 25,000-lU rIFN-ct2a dose. The elimination half-life was
lower (0.87-1.1 h) for the 25,000-lU dose level in comparison
to the 200,000-lU dose (t11, = 16.4 h). The elimination half-life
for the 25,000-lU dose was 26% greater on day S compared with
day I . The AUC for 200,000 IU rIFN-a2a was considerably
greater and CLs lower than predicted from data obtained at the
lower rIFN-a2a dose, suggesting saturation of elimination pro-
cesses.
The duration of maintenance of rIFN-a2a concentrations
above 50 lU/ml, a minimal modulatory concentration of
FUra-LV cytotoxicity determined for 72-h continuous expo-
sures in cultured cells (1) is shown in Table 4. For the 25,000-lU
dose level of rIFN-a2a, plasma concentrations of 50 lUImI were
maintained for 6 h on day 1 and 8 h on day 5, and for the
200,000-lU dose for >24 h.
Pharmacokinetic modeling oftwo clinical dosing schedules
used in humans is shown in Fig. 4. Cumulative systemic expo-
sure (AUC) following five daily doses of nIFN-a2a (5 X l0�
IU/m2) was 5270 lU!h/ml, and following three doses adminis-
tered on alternate days, 3310 lU/h/mI. Although in mice at the
200,000-lU rIFN-a2a dose level accumulation was suggested,
simulations in humans were more consistent with the lower
dosage studied in the mice where no accumulation was apparent
between days 1 and 5 as evidenced by no changes in CLs or
AUC. The simulation study also indicated that plasma concen-
trations of rIFN-a2a �50 IU/ml would be maintained for pen-
ods of 8 h following nIFN-ct2a administration, consistent with
the systemic exposure observed for the lowest rIFN-a2a dose in
the xenograft study.
DISCUSSION
In this study we extend the relationship between rIFN-a2a
concentrations that modulate FUra-LV cytotoxicity determined
in vitro to the level of rIFN-a2a systemic exposure associated
with dosages modulating the cytotoxicity of FUra and FUna-LV
in mice bearing human colon adenocancinoma xenografts. Fun-
then we show that the length of time above an in vitro modula-
tory concentration is similar between the mouse xenognaft
model and pharmacokinetic simulations of dosing schedules
used clinically.
rIFN-a2a has been shown to modulate the in vitro cyto-
toxicity of FUna and FUna-LV against two human colon adeno-
carcinoma cell lines, GC/cl and VRC5/cl (I). Previously we
had demonstrated, using cultured colon adenocancinoma cells,
that 500 lU/mI and 5,000 lU/mI nIFN-a2a produced similar and
significant modulation of FUra-LV cytotoxicity (10-14-fold;
>95% reduction in clonogenic survival) when cells were cx-
posed continuously for 72 h (1). Reduction in clonogenic sun-
vival by 60% was achievable when FUra-LV were combined
with as low as 50 IU/ml rIFN-a2a for 72 h (1). In addition, in
the presence of LV lower modulatory concentrations of rIFN-
a2a could be combined with a noncytotoxic FUra concentration
to achieve a cytotoxic response. In this in vitro system maximal
cytotoxicity required a minimum of 48 h of continuous drug
exposure (2).
With the knowledge that rlFN-a2a modulated the activity
of FUra in vitro, we evaluated three rIFN-a2a doses (25,000 IU,
200,000 IU, and 600,000 IU) against human colon adenocanci-
noma xenografts in combination with FUra and FUna-LV to
investigate the potential for enhancing in vivo thenapeutic activ-
ity. Using HxGC3/clTKc3 xenografts deficient in dThd sal-
vage, significant potentiation of FUna-LV activity was obtained
in the presence of nIFN-a2a, determined by enhanced volume
Research. on April 25, 2020. © 1995 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

10�
10�
10�I10
0 1 2 S 4 5 0
Tim. �4
38 FUra-LV Activity in Colon Adenocarcinoma Xenognafts
Table 2 Volume regressions induced by FUra-LV-nIFN-a2a combinations in HxGC�/c1TKc3 xenognafts’�
Group %PR %CR %C n
Control 0 0 0 50
FUra 62 0 0 29
FUra-LV 83 12 10 52
FUna-nIFN-a2a FUna-LV-nIFN-a2a
Dose of nIFN-a2a (IU) % PR % CR % C n % PR % CR % C n
25,000 31 9 3 35 94 23 20 35 <.05
200,000 67 0 0 10 100 17 8 8 <.05600,0(X) 100 60 60 10 100 67 67 12 >05
a FUra-LV > FUra, P < 0.01; FUra-nIFN-a2a > FUna, P < 0.001 at 600,000 lU only; FUna-nIFN-a2a > FUna-LV, P < 0.05 at 600,000 IU
only; FUna-LV-nlFN-a2a > FUna-LV, P < 0.01 at 600,000 IU only. The PR group contains all tumors that achieved at least 50% reduction in tumorvolume and hence also includes CRs.
S p comparison of FUna-nIFN-a2a with FUra-LV-rIFN-ct2a at equivalent nIFN-cs2a dose levels.
Table 3 Toxicity of FUra-LV-nIFN-cc2a combinations in tumor-
bearing mice
Group % Survival” n
Control 100 30FUra 97 30
FUra-LV 100 30
FUna-LV-FUna-nIFN-a2a rIFN-a2a
Dose of rlFN-a2a (lU) % Survival n % Survival n
25,000 100 18 100 18
200,000 83 6 100 6
600,()00 100 6 100 6
“Survival was determined within the first 3 weeks after completion
of therapy.
regressions and inhibition of tumor growth. Responses were
considerably greaten than those achieved with FUra combined
with either LV on nIFN-a2a as single modulators. The drug
interaction among FUra, LV, and nIFN-a2a was dependent on
reduced ability to salvage dThd, thereby confirming previous
data obtained using cultured GC3/cl cells (1, 2).
We then evaluated the influence of the rIFN-a2a dose or
systemic exposure on FUra activity. When FUra was combined
with rIFN-a2a as a single modulator, the greatest effect was
seen at the 600,000-lU dose level. However, this would consti-
tute a suprapharmacological exposure not achievable for clinical
studies. In contrast to results with FUra-rIFN-a2a, potentiation
of FUna-LV-induced growth inhibition was independent of the
nIFN-a2a dose oven the dose range examined, as was the extent
of volume regressions at the 25,000- and 200,000-lU dose
levels. The lowest nIFN-a2a dose (25,000 IU) yielded signifi-
cant potentiation of FUna-LV-induced growth inhibition, as high
as achieved with the 600,000-lU rIFN-a2a combined with FUra
± LV. Thus, in combination with LV, lower pharmacologically
relevant doses of nIFN-a2a were effective in modulating FUra.
As recently reviewed by Galpin and Evans (32), many
studies have shown systemic exposure to anticancer drugs to be
nelated to both toxicity and efficacy. Different approaches have
been used to estimate systemic exposure, including AUC, peak
plasma concentration, and time above a minimum effective
Fig. 3 Plasma concentrations of nIFN-cx2a on day 1 of administrationon on day 5 following daily i.p. injections. Dose levels of nIFN-a2awere: 0, 25,000 IU and A, 200,000 IU on day 1, and #{149},25,000 IU onday 5. Data represent the mean ± SD of 4 determinations at each time
point. Solid line, ‘ ‘best-fit’ ‘ curve generated from maximum likelihoodestimates of pharmacokinetic parameters; dashed line, limit of assay
sensitivity (15 IU/ml).
concentration. Studies of the antiviral properties of nIFN-a2a
showed that 2’,5’-oligoadenylate synthetase activity was related
to dose and route of nIFN-a2a administration (33). At similar
dosages, 2’,5’-oligoadenylate synthetase activity in peripheral
blood mononuclear cells was greaten when nIFN-a2a was given
i.m. compared with i.v. administration, paralleling prolonged
systemic exposure at low concentration from the i.m. route and
short exposure with an initial high peak level from the i.v. route.
This would suggest antiviral activity is associated with prolong-
ing rIFN-a2a exposure above a minimum effective concentra-
tion for some period of time rather than association with a high
peak concentration. Additionally, Wills et al. (28), studying the
pharmacokinetics of rIFN-a2a in volunteers, observed that ad-
verse effects from rIFN-a2a were related to exceeding and
maintaining serum concentrations above a threshold level.
For comparison with nIFN-a2a systemic exposure deter-
mined in mice, we simulated the rIFN-a2a plasma concentra-
Research. on April 25, 2020. © 1995 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

100
10
.�
:a
Co
CCo0C0
0
0 24 48 72 96 120 144
Clinical Cancer Research 39
Table 4 Phanmaco kinetics of nIFN-a2a in mice
Apparent DurationDose Vd� CLs AUC0 . . above
(IU) �t/2 (h) (ml/m2) (mi/him2) (IU-mlih) 50 lUimI (h)
25,000, 0.87 501 399 8,536 6
day 1
25,000, 1.1 712 448 7,533 8
day 5
200,000, 16.4 410 164 88,762 >24.0day I
tions achievable in humans following s.c. administration at the
nIFN-a2a dose levels used in clinical tnials of FUra-rIFN-a2a
(1 1-14) on FUna-LV-nIFN-a2a (15). Simulations were con-
structed for 5 X l0� IU/m2 nIFN-cx2a administered daily for S
days (15) or 9 X 10� IU administered on alternate days for 3
doses (1 1-14). The total AUC measured in mice following
25,000 IU rIFN-a2a administered daily for 5 consecutive days
was 41 and 66% of that determined from phanmacokinetic
simulations of the two clinical dosing schedules. Similarly,
estimated peak plasma concentrations of rIFN-a2a were lower
in both clinical studies (-90 lU/mi) than determined in mice
(�‘4000 lU/mi). The reduced effect of nIFN-ct2a dose on tumor
volume regressions and the lack of a dose-response relationship
for nIFN-a2a when combined with FUra and LV in inducing
growth inhibition in the xenograft studies suggested that time
above a minimum effective concentration may be more impon-
tant than the peak plasma concentration in inducing a modula-
tory response to FUna-LV activity.
Simulations of dosing schedules used clinically indicated
plasma rIFN-a2a concentrations would be maintained above SO
lU/mi each day for a period of 8 h, and with the alternate day
schedule, for 8 h three times per week. Of interest from this
study was that the 25,000-lU dose level in mice maintained a
nIFN-a2a plasma concentration of �50 lU/mI for 6-8 h. This
concentration of rIFN-cs2a for the durations obtained provided
in vito modulation of FUra-LV activity in mice. Thus, when
combined with LV it may be possible that intermittent and
repeated exposure to lower dosages (i.e. , concentrations) of
rIFN-cr2a is sufficient to modulate and potentiate the activity of
FUna.
Clearly, intermittent exposure to rIFN-a2a has been mod-
ulatony in the xenograft studies, and maintenance of critical
rIFN-a2a levels for 6- to 8-h periods daily appears to be suffi-
cient to modulate the activity of FUna in the presence of LV.
Since the window of intermittent nIFN-a2a exposure is very
similar between mice and humans, it is conceivable from the
simulations presented that the rIFN-a2a doses used clinically
will modulate FUra-LV activity. It is also evident from previous
studies with cultured cells (1, 2) and from the xenograft studies
that lower rIFN-cx2a doses may be used to modulate FUra
activity in the presence of LV. However, the optimal schedule
remains to be elucidated.
In patients, an additional effect of intenfenons has been to
increase FUna systemic exposure, thereby incneasing FUra-as-
sociated toxicity. Most of these investigations have been con-
ducted with an alternative IFN-a subtype (IFN-a2b; Ref. 34),
although effects appear to be similar with rIFN-a2a (15). In
0.1
Time (hr)
Fig. 4 Plasma concentration versus time plot for pharmacokinetic sim-
ulation of two clinical dosing schedules for rlFN-a2a as described in
‘ ‘ Materials and Methods.”
these studies pharmacokinetic analyses indicated that FUra CLs
decreased by 20-35% (34). Grem et al. (15) determined that
reduction in FUna CLs was related to the dose of nIFN-a2a,
resulting in an increase of FUna AUC of 30 and 50% in patients
receiving S x 10� and 10 X 10� lU/rn2, respectively, of nIFN-
a2a. However, the effects of intenferons are species specific, and
for the xenognaft studies, a human IFN-a subtype was used in a
munine model system bearing human tumors. Thus, as antici-
pated, no increase in FUna-associated toxicity was observed in
these studies. It is unlikely therefore that suitable data on the
influence of rIFN-a2a on FUna toxicity will be forthcoming
from studies conducted in mice. In this regard, it is noteworthy
that even at the highest rIFN-a2a dose, the toxicity of FUra on
FUna-LV was not enhanced. Consequently, this type of infon-
mation will be more readily gained from clinical studies.
Modulation of the activity of FUra by rIFN-a2a has been
possible in an in vito model system, and has been considerably
greaten at a more clinically relevant lower dose level of nIFN-
ct2a in the presence of LV. Plasma levels of nIFN-a2a achieved
in mice have been maintained above critical concentrations for
periods of time achievable clinically. It is feasible that dual
modulation may enhance the clinical activity of FUna in patients
with colorectal cancer, although whether responses and survi-
vonship are increased significantly oven the use of single mod-
ulatons will await results from randomized phase III clinical
trials in this population.
REFERENCES
I. Houghton, J. A., Adkins, D. A., Rahman, A., and Houghton, P. J.Interaction between 5-fluonounacil, [6R5]leucovonin, and recombinant
human interferon-a2a in cultured colon adenocarcinoma cells. Cancer
Commun., 3: 225-231, 1991.
2. Houghton, J. A., Morton, C. L., Adkins, D. A., and Rahman, A.
Locus of the interaction among 5-fluonounacil, leucovonin, and interfen-on-a2a in colon carcinoma cells. Cancer Res., 53: 4243-4250, 1993.
3. Houghton, J. A., Manoda, S. J., Phillips, J. 0., Houghton, P. J.
Biochemical determinants of responsiveness to 5-fluorounacil and its
derivatives in human colonectal adenocarcinoma xenognafts. Cancer
Res., 41: 144-149, 1981.
4. Radparvan, S., Houghton, P. J., and Houghton, J. A. Effect of
polyglutamylation of 5,10-methylenetetnahydnofolate on the binding of
5-fluoro-2’-deoxyunidylate to thymidylate synthase purified from a hu-man colon adenocarcinoma xenograft. Biochem. Pharmacol., 38: 335-
342, 1989.
Research. on April 25, 2020. © 1995 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

40 FUra-LV Activity in Colon Adenocarcinoma Xenografts
5. Houghton, J. A., Williams, L. G., de Graaf, S. S. N., Cheshire, P. J.,
Rodman, J. H., Maneval, D. C., Wainer, I. W., Jadaud, P., and Hough-
ton, P. J. Relationship between dose rate of [6R5]leucovonin adminis-
tration, plastna concentrations of reduced folates, and pools of 5,10-
methylenetetrahydrofolates and tetnahydnofolates in human colon
adenocancinoma xenografts. Cancer Res., 50: 3493-3502, 1990.
6. Houghton, J. A., Williams, L. G., Cheshire, P. J., Wainen, I. W.,
Jadaud, P., and Houghton, P. J. Influence of dose of [6RS]leucovonin onreduced folate pools and 5-fluonounacil-mediated thymidylate synthase
inhibition in human colon adenocancinoma xenognafts. Cancer Res., 50:
3940-3946, 1990.
7. Petrelli, N., Herrera, L., Rustum, Y., Burke, P., Creaven, P., Stulc, J.,Emnich, L. J., and Mittelman, A. A prospective randomized trial of
5-fluorounacil versus 5-fluorouracil and high-dose leucovonin versus
5-fluorounacil and methotrexate in previously untreated patients with
advanced colorectal carcinoma. J. Clin. Oncol., 5: 1559-1565, 1987.
8. Erlichman, C., Fine, S., Wong, A., and Elhakim, T. A randomized
trial of fluorouracil and folinic acid in patients with metastatic colonectal
carcinoma. J. Clin. Oncol., 6: 469-475, 1988.
9. Poon, M. A., O’Connell, M. J., Moertel, C. G., Wieand, H. S.,Cullinan, S. A., Everson, L. K., Krook, J. E., Mailland, J. A., Laurie,J. A., Tschetten, L. K., and Weisenfeld M. Biochemical modulation offluorouracil: evidence of significant improvement of survival and qual-
ity of life in patients with advanced colorectal carcinoma. J. Clin.
Oncol., 7: 1407-1418, 1989.
10. Doroshow, J. H., Multhauf, P., Leong, L., Margolin, K., Litchfield,T., Akman, S., Cam, B., Bertrand, M., Goldberg, D., Blayney, D.,Odujinnin, 0., DeLap, R., Shuster, J., and Newman, E. Prospective
randomized comparison of fluorounacil versus fluonouracil and high-
dose continuous infusion leucovonin calcium for the treatment of ad-vanced measurable colorectal cancer in patients previously unexposed tochemotherapy. J. Clin. Oncol., 8: 491-501, 1990.
1 1. Wadlen, S., Schwartz, E. L., Goldman, M., Lyven, A., Raden, M.,
Zimmerman, M., Itni, L., Weinberg, V., and Wiernik, P. H. 5-fluonou-
racil and recombinant cx2a-interferon: an active regimen against cob-
rectal carcinoma. J. Clin. Oncol., 7: 1769-1775, 1989.
12. Kemeny, N., Younes, A., Seiten, K., Kelsen, D., Sammanco, P.,
Adams, L., Derby, S., Murray, P., and Houston, C. Interferon a-2a and5-fluorounacil for advanced colorectal carcinoma. Cancer (Phila.), 66:
2470-2475, 1990.
13. Pazdun, R., Ajani, J. A., Patt, Y. Z., Winn, R., Jackson, D., Shep-hard, B., DuBrow, R., Campos, L., Quanaishi, M., Faintuch, J., Abbruzz-
ese, J. L., Gutterman, J., and Levin, B. Phase II study of fluonouracil and
recombinant interferon ct-2a in previously untreated advanced colorectal
carcinoma. J. Clin. Oncol., 8: 2027-2031, 1990.
14. Wadler, S., Lembersky, B., Atkins, M., Kirkwood, J., and Petrelli,N. Phase II trial of fluorounacil and recombinant interferon alfa-2a inpatients with advanced colorectal carcinoma: an Eastern Cooperative
Oncology Group Study. J. Clin. Oncol., 9: 1806-1810, 1991.
15. Grem, J. L., McAtee, N., Murphy, R. F., Balis, F. M., Steinberg,
S. M., Hamilton, J. M. Sorensen, J. M., Santon, 0., Kramer, B. S.,
Goldstein, L. J., Gay, L. M., Caubo, K. M., Goldspiel, B., and Allegra,C. J. A pilot study of interferon alfa-2a in combination with fluorouracil
plus high-dose leucovonin in metastatic gastrointestinal carcinoma. J.Clin. Oncol., 9: 1811-1820, 1991.
16. Wadler, S., and Wiernik, P. H. Clinical update on the role of
fluorouracil and recombinant interferon alpha-2a in the treatment of
colonectal carcinoma. Semin. Oncol., 17: 16-21, 1990.
17. Wadler, S., Goldman, M., Lyver, A., and Wiennik, P. H. Phase I trial
of 5-fluorouracil and recombinant cx2a-interfenon in patients with ad-
vanced colonectal carcinoma. Cancer Res., 50: 2056-2059, 1990.
18. Houghton, J. A., Williams, L. G., Loftin, S. K., Cheshire, P. J.,
Morton, C. L., Houghton, P. J., Dayan, A., and Jolivet, J. Factors that
influence the therapeutic activity of 5-fluorouracil [6RSjleucovonin
combinations in colon adenocarcinoma xenognafts. Cancer Chemothen.
Pharmacol., 30: 423-432, 1992.
19. Howell, S. B., Mansfield, S. J., and Tactic, R. Thymidine andhypoxanthine requirements of normal and malignant human cells for
protection against methotrexate cytotoxicity. Cancer Res., 41: 945-950,
1981.
20. Woodman, P. W., Williams, D. L., and Edwards, H. H. Heteroge-
neity in cell populations established in vitro from a human colonadenocarcinoma xenograft. In Vitro, 16: 21 1, 1980.
21. Houghton, P. J., Houghton, J. A., Germain, G., and Torrance, P. M.Development and characterization of a human colon adenocarciomaxenograft deficient in thymidine salvage. Cancer Res., 47: 2117-2122,
1987.
22. Radparvar, S., Houghton, P. J., Germain, G., Pennington, J., Rah-man, A., and Houghton, J. A. Cellular pharmacology of 5-fluonouracil in
a human colon adenocarcinoma cell line selected for thymidine kinasedeficiency. Biochem. Pharmacol., 39: 1759-1765, 1990.
23. Houghton, P. J., Houghton, J. A., Myers, L., Cheshire, P., Howbert,J. J., and Gnindey, G. B. Evaluation of N-(5-indanylsulfonyl)-N’-(4-chborophenyl)-urea against xenografts of pediatric rhabdomyosarcoma.
Cancer Chemothen. Pharmacol., 25: 84-88, 1989.
24. Freireich, E. J., Gehan, E. A., Rail, D. P., Schmidt, L. H., and
Skipper, H. E. Quantitative comparison of toxicity of anticancer agentsin mouse, rat, hamster, dog, monkey, and man. Cancer Chemother. Rep.,
50: 219-244, 1966.
25. Winer, B. J., Brown, D. R., and Michels, K. M. Statistical Principles inExperimental Design, 3rd Ed, p. 182. New York: McGraw-Hill, 1991.
26. Protzman, W. P., Minnicozi, M., Jacobs, S. L., Surprenant, D. I.,
Schwartz, J., and Oden, E. M. Immunoradiometnic assay of a recombi-
nant human alpha-2-interferon (SCH 30500). J. Clin. Microbiol., 22:
596-599, 1985.
27. D’Angenio, D. Z., and Schumitzky, A. ADAPT II User’s Guide. Los
Angeles: Biomedical Simulations Resource, 1990.
28. Wills, R. J., Dennis, S., Spiegel, H. E., Gibson, D. M., and Nadlen,
P. I. Interferon kinetics and adverse reactions after intravenous, intra-
muscular, and subcutaneous injection. Clin. Pharmacol. Then., 35: 722-
727, 1984.
29. Balkwill, F. R., Goldstein, L., and Stebbing, N. Differential action
of six human interfenons against two human carcinomas growing innude mice. Br. J. Cancer, 35: 613-617, 1985.
30. Carmichael, J., Fengusson, R. J., Wolf, C. R., Balkwill, F. R., andSmyth, J. F. Augmentation of cytotoxicity of chemotherapy by humana-interfenons in human non-small cell lung cancer xenografts. Cancer
Res., 46: 4916-4920, 1986.
31. Balkwill, F. R., and Moodie, E. M. Positive interactions between
human interferon and cyclophosphamide or adniamycin in a humantumor model system. Cancer Res., 44: 904-908, 1984.
32. Galpin, A. J., and Evans, W. E. Therapeutic drug monitoring incancer management. Clin Chem., 39: 2419-2430, 1993.
33. Witter, F., Barouki, F., Griffin, D., Nadler, P., Woods, A., Wood,D., and Lietman, P. Biologic response (antiviral) to recombinant humaninterferon alpha 2a as a function of dose and route of administration in
healthy volunteers. Clin. Pharmacol. Then., 42: 567-575, 1987.
34. Danhauser, L. L., Fneimann, J. H., Gilchnist, T. L., Gutterman, J. U.,
Hunter, C. Y., Yeomans, A. C., and Markowitz, A. B. Phase I andplasma pharmacokinetic study of infusional fluonounacil combined with
recombinant interferon alfa-2b in patients with advanced cancer. J. Clin.
Oncol., Ii: 751-761, 1993.
Research. on April 25, 2020. © 1995 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

1995;1:33-40. Clin Cancer Res J A Houghton, P J Cheshire, C L Morton, et al. alpha2a-interferon in colon adenocarcinoma xenografts.Potentiation of 5-fluorouracil-leucovorin activity by
Updated version
http://clincancerres.aacrjournals.org/content/1/1/33
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/1/1/33To request permission to re-use all or part of this article, use this link
Research. on April 25, 2020. © 1995 American Association for Cancerclincancerres.aacrjournals.org Downloaded from