photosynthese i „lichtreaktionen“ - uni-muenster.de · durch absorption von licht kann ein...
TRANSCRIPT

12.05.2016
1
Vorlesung „Zellbiologie Physiologie und Genetik“
SoSe 2016
Photosynthese I„Lichtreaktionen“
Prof. Dr. Iris Finkemeier
Institut für Biologie und Biotechnologie der Pflanzen
© 2015 American Society of Plant Biologists
Photosynthese I
• Photosynthese Einführung: Bedeutung, Evolution und Diversität
• Plastiden und Chloroplasten
• Licht und Pigmente
• Struktur und Funktion der Photosynthesekomplexe
• Linearer Elektronentransport und Wasserspaltung

12.05.2016
2
Joseph Priestley (1772)
erkannte, dass die Atmung
von Tieren und Menschen
die Luft “verbraucht”..und
Pflanzen die Fähigkeit
besitzen, Luft wieder
regenerieren zu können.
Prinzip der Photosynthese

12.05.2016
3
Weltweiter Vegetationsindex Mai 2015
Quelle: NEO NASA Earth Observations
Pflanzen bilden die Grundlage unseres Lebens auf der Erde
99% des Sauerstoffs in der Atmosphäre stammt aus der Photosynthese

12.05.2016
4
Pflanzen bilden die Grundlage unseres Lebens auf der Erde
Quelle: NEO NASA Earth Observations
Chlorophyll im Phytoplankton Mai 2015
pro Jahr werden ca. 258 *109 Tonnen Kohlendioxid durch Photosynthese gebunden
© 2015 American Society of Plant Biologists
Photonenfluss in microEinstein (µE = µmol Quanten m-2 s-1)
Die Photosynthese besteht aus zweimiteinander verbundenen Reaktionen

12.05.2016
5
© 2015 American Society of Plant Biologists
Die Photosynthese besteht aus zweimiteinander verbundenen Reaktionen
Adapted from Kramer, D.M., and Evans, J. R. (2010). The importance of energy balance in improving photosynthetic productivity. Plant Physiol.155: 70–78.
Chloroplast2 H2O O2 + 2 H+ + 2 ADPH+
ATP
Die Lichtreaktionen in der Thylakoidmembran
Die Kohlenstofffixierungim Stroma
2 NADP+
2 NADPH
2 H+e−
e−
• Photosynthese liefert Triosephosphate, die in andere Kohlenhydrate umgewandelt und in andere Pflanzenteile transportiert werden müssen.
• Photosynthese ist naturbedingt diurnal mit wichtigen Konsequenzen für den Kohlenhydratmetabolismus bei Tag und Nacht.
Photosynthese

12.05.2016
6
Photosynthetische Organismen
Höhere Pflanzen
Niedere Pflanzen
Vielzellige Algen
Einzellige Algen
Cyanobakterien
PurpurbakterienPurpurbakterienCyanobakterienEinzellige Grünalge
Vielzellige BraunalgeNiedere (Moose, Farne) und Höhere Pflanzen (z.B. Klee)
25 µm Eckhard Völker, 2012

12.05.2016
7
Cyanobakterien als Vorgänger der Chloroplasten
Primäre Endosymbiose

12.05.2016
8

12.05.2016
9
ein Organismus lebt in einem anderen
VorticellaGlockentierchen
HeterotrophieAbhängigkeit von externen organischen Substanzen, die von anderen Lebewesen produziert werden.Tiere, Pilze, viele Bakterien
AutotrophieFähigkeit organische Substanzenaus Anorganischen zu synthetisierenPflanzen, Algen, Cyanobakterien

12.05.2016
10
© 2015 American Society of Plant Biologists
Photosynthetische Tiere:Flecken-Querzahnmolch
Kerney, R., Kim, E., Hangarter, R.P., Heiss, A.A., Bishop, C.D. and Hall, B.K. (2011). Intracellular invasion of green algae in a salamander host. Proc. Natl. Acad. Sci. USA 108: 6497-6502. Camazine
Egg capsule
Ambystoma maculatum
Die Alge liefert Sauerstoffund Kohlenhydrate und profitiert selber vomStickstoffabfall des Molchs
Die einzellige AlgeOophila amblystomatis lebtin den Eikapseln des Molchs
Die Alge (rote Spots) wurde sogarin Zellen des Salamanders gefunden. Dies weist auf eineEndosymbiose mit dem Molch hin
Photosynthetische Tiere:Plastiden-stehlende Schnecke
Pelletreau, K.N., Bhattacharya, D., Price, D.C., Worful, J.M., Moustafa, A. and Rumpho, M.E. (2011). Sea slug kleptoplasty and plastid maintenance in a metazoan. Plant Physiol. 155: 1561-1565. Rumpho, M.E., Pelletreau, K.N., Moustafa, A. and Bhattacharya, D. (2011). The making of a photosynthetic animal. J. Exp. Biol. 214: 303-311.
Vaucheria litorea (algae)
Elysia chlorotica (lebt imMeer, nordamerikanischeAtlantikküste)
Plastiden überlebenmehrer Monate in den Zellen der Schnecke –den “Plastidendiebstahl” nennt man KleptoplastyChloroplasts in sea slug

12.05.2016
11
© 2015 American Society of Plant Biologists
Photosynthese I
• Photosynthese Einführung: Bedeutung, Evolution und Diversität
• Plastiden und Chloroplasten
• Licht und Pigmente
• Struktur und Funktion der Photosynthesekomplexe
• Linearer Elektronentransport und Wasserspaltung
• Ein Kompartiment (und circa 4000 zusätzliche Gene), das Tiere nicht haben.
• Viele metabolische Reaktionen finden in der Plastide statt und Metaboliten müssen importiert und exportiert werden.
Plastide

12.05.2016
12
Differenzierung von Plastiden
Immage credit LadyofHats; see also Sakamoto W., Miyagishima S., and Jarvis P. (2008). Chloroplast Biogenesis: Control of Plastid Development, Protein Import, Division and Inheritance. The Arabidopsis Book 6:e0110. doi:10.1199/tab.0110
LeafBlüten und Früchten
In dunklenphotosynthetischenGeweben
Samen, Embryo, Meristem und reproduktivenGeweben
Speicherung von Stärke, Öl und Proteinen
Licht induziert Umwandlung von Etioplastenzu Chloroplasten
Pribil, M., Labs, M. and Leister, D. (2014). Structure and dynamics of thylakoids in land plants. J. Exp. Bot. 65: 1955-1972 by permission of Oxford University Press . Von Wettstein, D., Gough, S. and Kannangara, C.G. (1995). Chlorophyll biosynthesis. Plant Cell. 7: 1039-1057..
Prolamellar Körper in Etioplast
Bildung von Lamellen
Primäre Lagenvon Lamellen
Granastapeln
Dunkel
Licht

12.05.2016
13
Plastiden in Pflanzenzellen
Aufbau Chloroplast

12.05.2016
14
Grana- und Stromathylakoide habenverschiedene Funktionen
Daum, B., et al., (2010). Arrangement of Photosystem II and ATP synthase in chloroplast membranes of spinach and pea. Plant Cell. 22: 1299-1312; Nagy, G., et al. and Minagawa, J. (2014). Chloroplast remodeling during state transitions in Chlamydomonas reinhardtii as revealed by noninvasive techniques in vivo. Proc. Natl. Acad. Sci. USA. 111: 5042-5047.
Lumen
Grana
PSII gepackt zwischen zwei Membranen
State 1
ATP synthase
PSI-LHCI supercomplex
Cyt b6f
LHCIICyt b6f
PSII
PSII findet man bevorzugt in Granathylakoiden, ATP synthase and PSIin StromalamellenCyt b6f überall in der Thylakoide
© 2015 American Society of Plant Biologists
Photosynthese I
• Photosynthese Einführung: Bedeutung, Evolution und Diversität
• Plastiden und Chloroplasten
• Licht und Pigmente
• Struktur und Funktion der Photosynthesekomplexe
• Linearer Elektronentransport

12.05.2016
15
© 2015 American Society of Plant Biologists
Wellenlängen des Lichts
Gammastrahlen
Röntgenstrahlen
IR
Visible
UV
Mikrowellen
Radiowellen
1 nm 1 μm 1 mm 1 mWellenlängen
500nm
400nm
600nm
Wellenlängen
Viel EnergieHohe FrequenzKurze Wellenlänge
Geringe EnergieGeringe FrequenzLange Wellenlänge
LichtenergieDie Energie des Lichts verhält sich umgekehrt proportionalzu seiner Wellenlänge:
blaues Licht ~ 300 kJ/mol Quantenrotes Licht ~ 170 kJ/mol Quanten
Zum Vergleich: eine C-C Bindung hat einen Energiegehalt von 346 kJ/mol
E = hc / lE = Energie eines Lichtquants (Joule photon-1)
h = Planck‘sches Wirkungsquantum (Proportionalitätskonstante)
c = Lichtgeschwindigkeit (3 x 108 m s-1)
l = Wellenlänge
Internat. Einheit des „Lichtflusses“: Mol Quanten m-2 s-1 (Einstein) Max. Sonnenlicht: ca. 2000 Mol Quanten m-2 s-1 (Hochlicht!)Sichtbarer Bereich: 400 – 700 nm (blau bis rot)

12.05.2016
16
© 2015 American Society of Plant Biologists
Das kurzwellige Licht wird durch die Erdatmosphäre herausgefiltert
NASA
300 400 500 600 700 800
Wellenlänge (nm)
Lich
tspe
ktru
mde
s S
onne
nlic
hts
Erd
ober
fläch
e(μ
mol
phot
os /
m2 -
sec) Abhängig von der
Wellenlänge wird die Sonnenstrahlung von der Atmosphäre mehr oder weniger stark absorbiert. Die an der Erdoberfläche eintreffende Intensität hängt zudem stark von der Bewölkung und vom Sonnenstand ab.
Ultraviolett
Die Erdatmosphäreblockiert kurzeWellenlängen
Was ist Farbe?
1. Weißes Licht ist eine Mischung aus Licht verschiedener Farben
2. Farbiger Gegenstand absorbiert einige Wellenlängen (rot: absorbiert alles bis auf rot) und besitzt ein Absorptionsspektrum
3. Substanz die Licht absorbieren kann: Pigment
4. Wichtigstes Pigment der Photosynthese: Chlorophyll (grüne Farbe): absorbiert im blauen und roten Bereich und reflektiert grünes Licht

12.05.2016
17
© 2015 American Society of Plant Biologists
Wichtige Pigmente der Photosynthese
300 400 500 600 700 800
Wellenlänge (nm)
Abs
orpt
ions
spek
trum
phot
osyn
thet
isch
erP
igm
ente Chlorophyll a
Chlorophyll bβ-CarotinPhycoerythrinPhycocyanin
Akz
ess
oris
che
Pig
men
te
Phycocyanobilin(lineares Tetrapyrrol)
β-Carotin
Chlorophyll a
Tetrapyrrolring mitMg im Zentrum
© 2015 American Society of Plant Biologists
Absorptionsspektrum der photosynthetischen Pigmente
300 400 500 600 700 800
Wellenlänge (nm)
Abs
orpt
ions
spek
trum
phot
osyn
thet
isch
erP
igm
ente Chlorophyll a
Chlorophyll bβ-CarotinPhycoerythrinPhycocyanin
Akz
ess
oris
che
Pig
men
te
Alle Chlorophyll-basiertenPhotosynthesesystemebenutzen Chlorophyll a
Akzessorischen Pigmente:• Chlorophyll b (Landpflanzen,
Grünalgen und Cyanobakterien)
• Carotenoide (in allenChlorophyll-basiertenPhotosynthesesystemen)
• Phycoerythrin(Cyanobakterien und nicht-grünen Algen)
• Phycocyanin(Cyanobakterien)
12.05.2016
18
© 2015 American Society of Plant Biologists
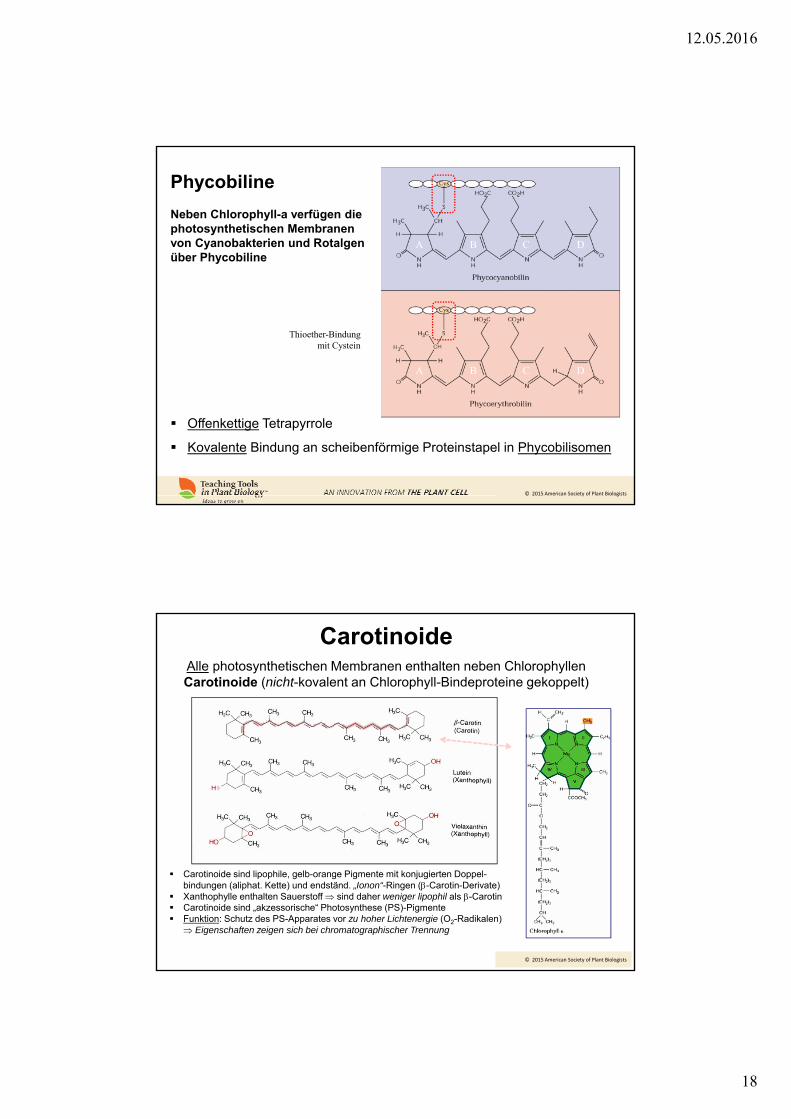
Phycobiline
Neben Chlorophyll-a verfügen die photosynthetischen Membranen von Cyanobakterien und Rotalgen über Phycobiline
Offenkettige Tetrapyrrole
Kovalente Bindung an scheibenförmige Proteinstapel in Phycobilisomen
A B C D
A B C D
Thioether-Bindung mit Cystein
© 2015 American Society of Plant Biologists
CarotinoideAlle photosynthetischen Membranen enthalten neben ChlorophyllenCarotinoide (nicht-kovalent an Chlorophyll-Bindeproteine gekoppelt)
Carotinoide sind lipophile, gelb-orange Pigmente mit konjugierten Doppel-bindungen (aliphat. Kette) und endständ. „Ionon“-Ringen (-Carotin-Derivate)
Xanthophylle enthalten Sauerstoff sind daher weniger lipophil als -Carotin Carotinoide sind „akzessorische“ Photosynthese (PS)-Pigmente Funktion: Schutz des PS-Apparates vor zu hoher Lichtenergie (O2-Radikalen)
Eigenschaften zeigen sich bei chromatographischer Trennung

12.05.2016
19
Photosynthetische Organismen enthalten Chlorophyll als Photosynthese-Pigmente. WICHTIG: Diese sind immer an Membranproteine gebunden!
Bei Bakterien sind die Pigmente an die Zellmembran oder an spezielle Photosynthese-Membranen gebunden, bei Algen und Pflanzen fungieren die Thylakoide (s. oben) als Träger der Chlorophyll-Proteinkomplexe.
Abbr.: st, stroma thylacoid; pg, plastoglobuli; gt, grana thalycoid.
polarem Tetrapyrrol-Ring („farbig“, durch konjugierte Doppelbindungen)
zentralem Magnesium-Atom (Mg2+-freies „Chlorophyll“ = Phaeophytin, griech. hässlich)
hydrophobem Phytolschwanz (dient der Membran-Verankerung)
nicht-kovalenter Assoziation mit sogen. „chlorophyll-binding proteins“ (CBPs)
Chlorophylle unterscheiden sich in ihren Seitengruppen und im Tetrapyrrolring (Abb. ob.)
Seitengruppen (Methyl-, Aldehyd-, etc.), bedingen die Position der Absorptionsbanden
AldehydMethylChlorophyllePorphyrin-Ringsystem mit zentralem Metall-Atom (Magnesium)
bestehen aus:

12.05.2016
20
Die Absorptionsspektren der verschiedenen Chlorophylleunterscheiden sich in der Lage von je zwei Hauptbanden
„Grün-Lücke“
Eigenschaft der Pigmente:
Wenn Chlorophylle oder andere Pigmente Licht absorbieren, werden Elektronen auf ein höheres Energieniveau gehoben
Pigment + Licht = Pigment*

12.05.2016
21
Eigenschaften eines Pigments Pigmente („farbige“ Moleküle) verfügen über ein konjugiertes pi-Elektronen-
system
Durch Absorption von Licht kann ein Pigment von seinem Grundzustand (P) in einen „angeregten“ Zustand (P*) überführt werden, sofern der Energiegehalt der absorbierten Lichtquanten der Energiedifferenz von P und P* entspricht.
Dabei werden konjugierte pi-Elektronen auf ein energetisch höheres Energie-Niveau gehoben (bei gleichbleibendem Elektronenspin).
Das Energieniveau von P* wird durch die molekularen Eigenschaften des Pigments bestimmt und äußert sich in charakteristischen Absorptionsbanden.
P P*
Grundzustand
angeregter Zustand
© 2015 American Society of Plant Biologists
Der angeregte Zustand von Chlorophyll ist kurzfristig. Rückkehr zum Grundzustand
kann auf vier Wegen erfolgen:
Chl
Chl*Photon
4. Elektronenübertragung (Photochemie): Energie wird als energiereiches Elektron auf benachbartes Molekül übertragen (Elektronenakzeptor) und hinterlässt Elektronenlücke im ursprünglichen Molekül (Ladungs-trennung).
1. Relaxation (Nicht-photochemisches quenchen; Energie wird in Wärme umgewandelt)
2. Fluoreszenz (Energie wird in Form von langwelligerem Licht abgestrahlt)
3. Resonanzenergieübertragung (Excitonentransfer):Energie wird auf anderes Pigmentmolekül übertragen und regt dieses an.

12.05.2016
22
UV 400 500 600 700 nm IR
Chl a
Chlorophylle haben zwei Haupt-Absorptionsbanden im blauen und roten Spektralbereich
Abs
orpt
ion
Die zwei Haupt-Absorptionsbanden entsprechen den zwei Haupt-Anregungszuständen der Chlorophylle
400 500 600 700 nm
Ch
l
2. Anregungszustand
1. Anregungszustand
Grundzustand
Rel
axat
ion(
en)
Fluo
resz
enz
Abs
orpt
ion
eine
s bl
auen
Lic
htqu
ants
Abs
. e. r
oten
Lic
htqu
ants
Energie
der 1. Anregungszustand entspricht der Energie eines roten Lichtquants
der 2. Anregungszustand entspricht der Energie eines blauen Lichtquants
der energiereichere 2. Anregungszustand ist aber instabil und relaxiert spontan zum energieärmeren 1. Anregungszustand
die Energiedifferenz geht als Wärme verloren
der stabilere 1. Anregungszustand kann unter Abgabe eines dunkelroten Lichtquants relaxieren ( Fluoreszenz)
MERKE: Für Photosynthese ist nur der stabilere, 1. Anregungszustand nutzbar!
12.05.2016
23
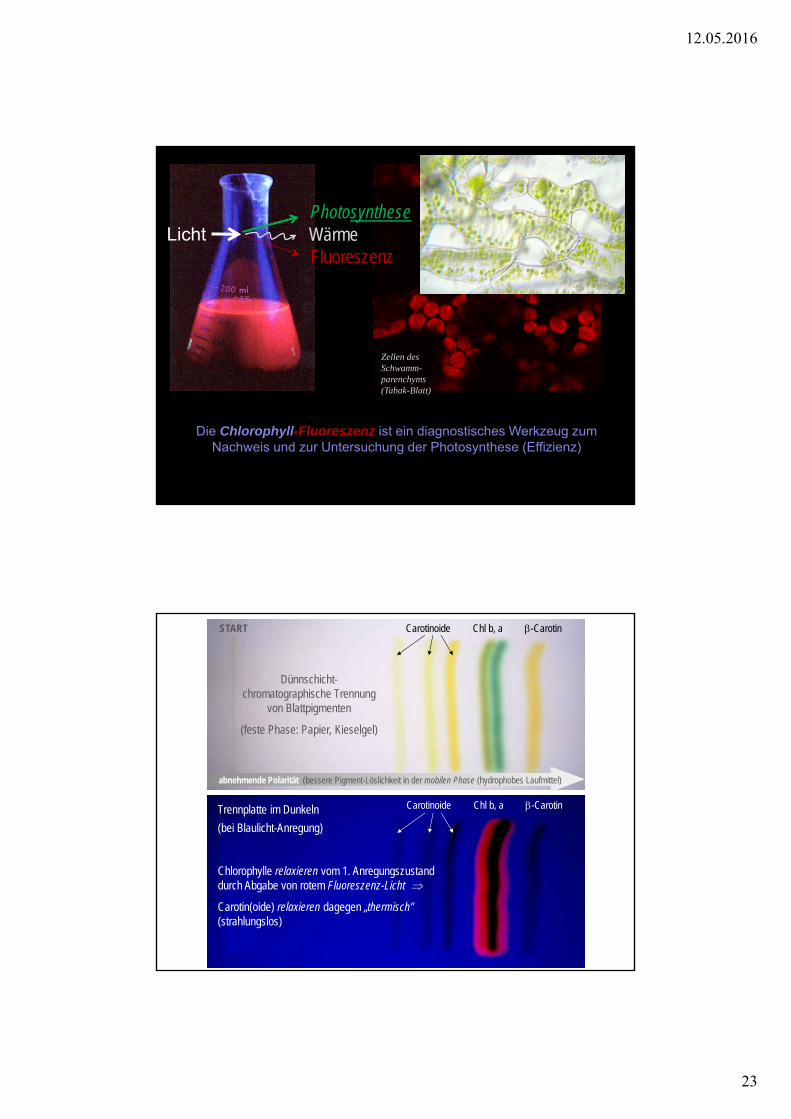
Die Chlorophyll-Fluoreszenz ist ein diagnostisches Werkzeug zum Nachweis und zur Untersuchung der Photosynthese (Effizienz)
PhotosyntheseLicht
Fluoreszenz Wärme
Zellen des Schwamm-parenchyms(Tabak-Blatt)
Trennplatte im Dunkeln
(bei Blaulicht-Anregung)
Chlorophylle relaxieren vom 1. Anregungszustand durch Abgabe von rotem Fluoreszenz-Licht
Carotin(oide) relaxieren dagegen „thermisch“ (strahlungslos)
Carotinoide Chl b, a -Carotin
abnehmende Polarität (bessere Pigment-Löslichkeit in der mobilen Phase (hydrophobes Laufmittel)
Dünnschicht-chromatographische Trennung
von Blattpigmenten
(feste Phase: Papier, Kieselgel)
START
Carotinoide Chl b, a -Carotin

12.05.2016
24
Zusammenfassung• Chlorophylle (Chl) bestehen aus einem polaren, „farbigen“ Tetra-
pyrrolring mit zentralem Magnesium (Mg2+)-Atom und einem hydrophoben Phytolschwanz. Sie sind nicht-kovalent an Chl-Binde-proteine (in der Membran) gebunden.
• Phycobiline sind offenkettige Tetrapyrrole niederer Organismen, die kovalent an ihre Trägerproteine gebunden sind.
• Chlorophylle und Phycobiline dienen der Aufnahme und Weiterleitungvon Lichtenergie in photosynthetischen Antennen.
• Carotinoide sind langkettige Pigmente (40 C-Atome), die ebenfalls nicht-kovalent an Chlorophyll-Bindeproteine gebunden sind. Sie üben eine Schutzfunktion bei übermäßiger Lichtanregung aus.
• Alle photosynthetischen Organismen enthalten Chlorophylle und Carotinoide, Cyanobakterien und Rotalgen enthalten zusätzlichPhycobiline.
© 2015 American Society of Plant Biologists
Photosynthese I
• Photosynthese Einführung: Bedeutung, Evolution und Diversität
• Plastiden und Chloroplasten
• Licht und Pigmente
• Struktur und Funktion der Photosynthesekomplexe
• Linearer Elektronentransport und Wasserspaltung
12.05.2016
25
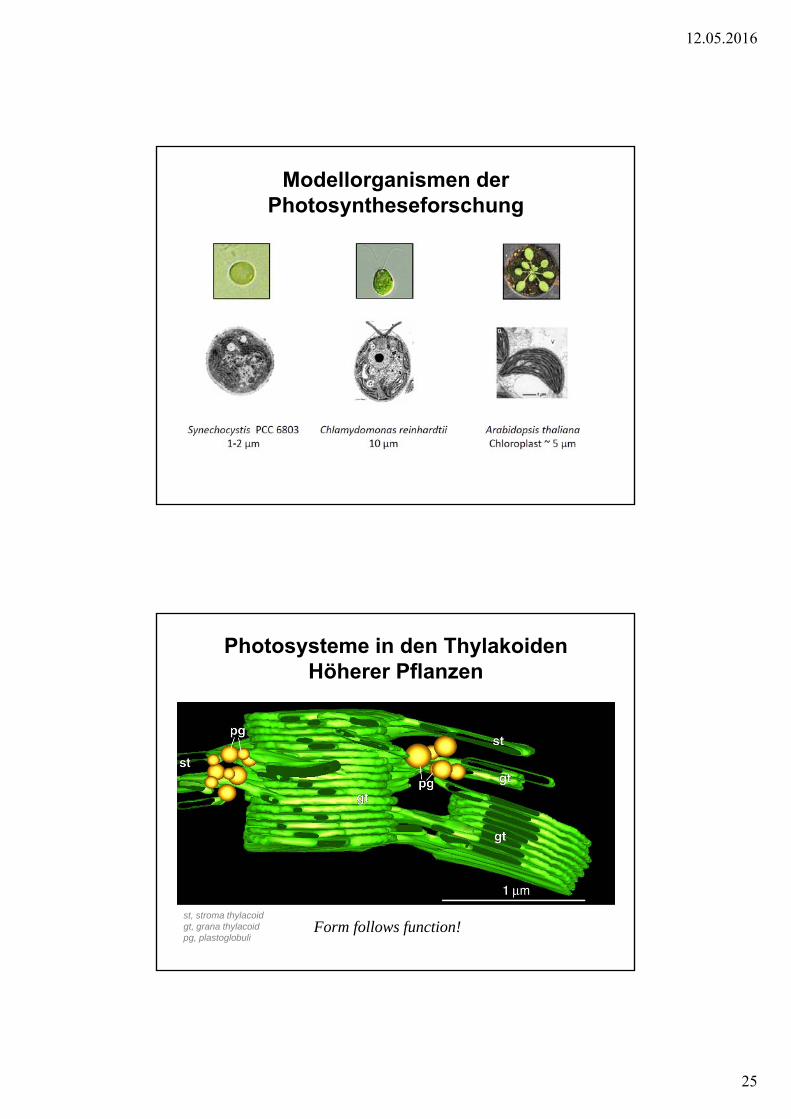
Modellorganismen der Photosyntheseforschung
Photosysteme in den ThylakoidenHöherer Pflanzen
st, stroma thylacoidgt, grana thylacoidpg, plastoglobuli
Form follows function!

12.05.2016
26
Photosysteme Höherer Pflanzen (schematisch, Seitenansicht)
Light Harvesting Complexes(LHCs)
core Antenna
Reaction Center (RC)
Thylakoid-Membran
RC, core Antennen und LHCs
sind Komplexe aus transmembranen, Chlorophyll-bindenden Proteinen
Chlorophylle sind in die hydrophoben Taschen der transmembranen Proteine eingesenkt.
Spiralen: a-Helices, grüne Ringsysteme: Chlorophyll a (Porphyrine, hier ohne Phytolschwanz)
Seitenansicht
Molekulare Struktur eines PS Höherer Pflanzencore Antenne (gelbe Helices) und Reaktionszentrum (blaue und rote Helices)
12.05.2016
27
LHC
LHC
LHC
LHCLHC
LHC
LHC
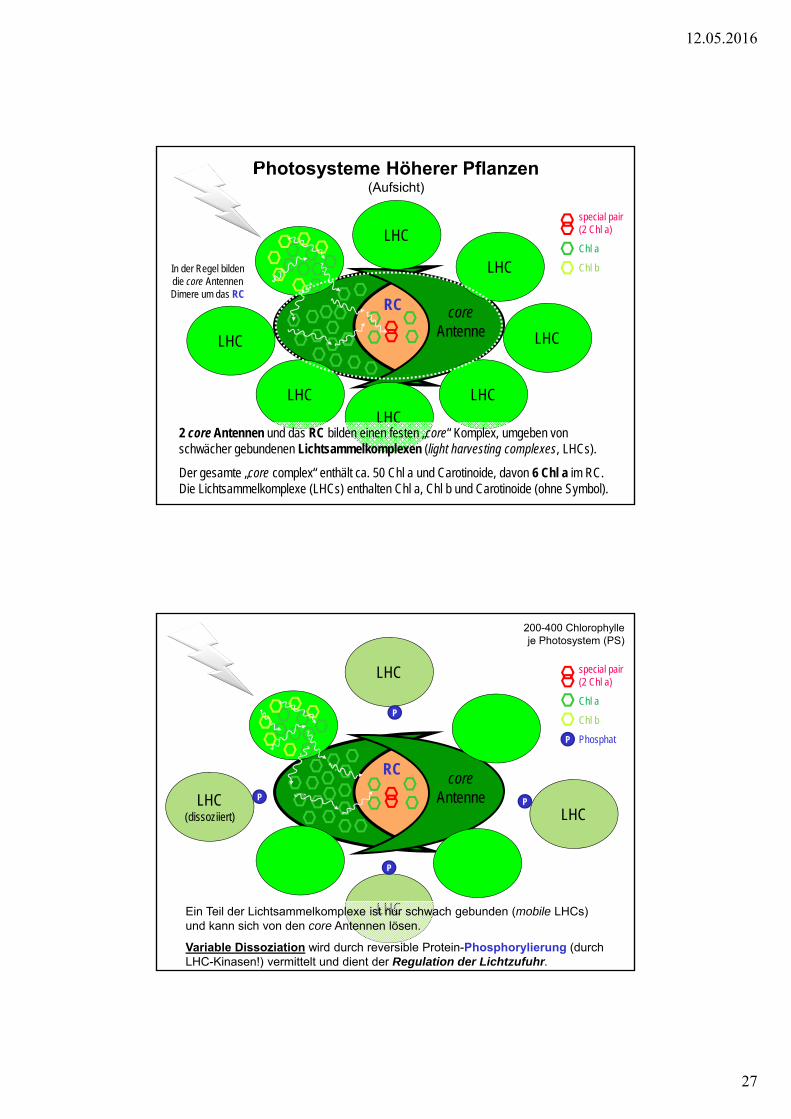
Photosysteme Höherer Pflanzen (Aufsicht)
coreAntenne
special pair (2 Chl a)
Chl a
Chl b
2 core Antennen und das RC bilden einen festen „core“ Komplex, umgeben von schwächer gebundenen Lichtsammelkomplexen (light harvesting complexes, LHCs).
Der gesamte „core complex“ enthält ca. 50 Chl a und Carotinoide, davon 6 Chl a im RC. Die Lichtsammelkomplexe (LHCs) enthalten Chl a, Chl b und Carotinoide (ohne Symbol).
RC
In der Regel bilden die core Antennen Dimere um das RC
LHC
LHC
LHC(dissoziiert)
200-400 Chlorophylle je Photosystem (PS)
coreAntenne
special pair (2 Chl a)
Chl a
Chl b
Phosphat
RC
LHC
P
P P
P
Ein Teil der Lichtsammelkomplexe ist nur schwach gebunden (mobile LHCs) und kann sich von den core Antennen lösen.
Variable Dissoziation wird durch reversible Protein-Phosphorylierung (durch LHC-Kinasen!) vermittelt und dient der Regulation der Lichtzufuhr.
P

12.05.2016
28
LHC
Einfangen von Lichtquanten und Energietransfer zum Reaktionszentrum (RC)
Chlorphyll a und b in den peripheren LHCs fangen die Lichtquanten ein und leiten diese Anregungsenergie mittels Resonanzschwingungenüber die core Antennen zum „special pair“ im Reaktionszentrum (RC) weiter, bis die Energie für Ladungstrennung ausreicht…
RCcore
Antenne
Durch Absorption eines Lichtquants entsteht im Chlein „angeregter“ Zustand
Chl b Chl b*
LichtquantLichtquant

12.05.2016
29
Chl b* Chl b
Angeregte Zustände (‚Exitonen‘) können zu benachbarten Chlorophyllen transferiert werden
Chl b Chl b*
Chl b* Chl b Chl a Chl a
Exitonentransfer in der Antenne

12.05.2016
30
Chl b Chl b* Chl a Chl a
Exitonentransfer in der Antenne
Chl b Chl b Chl a* Chl a
Exitonentransfer in der Antenne

12.05.2016
31
Chl b Chl b Chl a Chl a*
Exitonentransfer in der Antenne
kein Netto-Transfer von Elektronen !
Transfer zwischen Chlorophyllen ist extrem schnell: 10-12 sec !
Energietransfer bevorzugt zwischen eng benachbarter Pigmenten mit überlappenden -Orbitalen.
Aus energetischen Gründen bevorzugte Transferrichtung: Chl.b Chl.a special pair
Chl b Chl b Chl a Chl a*
Exitonentransfer in der Antenne

12.05.2016
32
D [Chl a]2* A D [Chl a]2+ A- D+ [Chl a]2 A-
Ladungstrennung im Reaktionszentrum
Durch Exitonentransfer wird die Lichtenergie aus den Antennen zumspecial pair [Chl a]2 im Reaktionszentrum übertragen.
Das angeregte special pair gibt ein Elektron an einen benachbarten Akzeptor ab (A A-).
Das dadurch entstandene „Elektronenloch“ im special pair wird durch Elektronentransfer von einem benachbarten Donor (D) aufgefüllt.
Es entsteht ein „arbeitsfähiges“ Redoxpaar, nämlich: D+/ A-
Die Thylakoidmembran ist dicht mit Photosystemengefüllt (200 - 400 Photosysteme pro Grana „Etage“).
Die Lichtsammelkomplexe stehen z.T. in engem Kontakt, so dass Lichtenergie zwischen den Photosystemenausgetauscht werden kann.
Die Gesamtheit der Photosysteme einer Membran-Ebene bilden daher eine Funktionseinheit.
PSII- Komplexe(Aufsicht)
Granastapel (Seitenansicht)

12.05.2016
33
Bakterielle Photosystemevon Purpurbakterien und Cyanobakterien
Während das photosynthetische Reaktionszentrum hoch konserviert ist,
haben sich in verschiedenen Organismen sehr unterschiedliche Antennensysteme entwickelt
Struktur des Reaktionzentrums von Purpurbakterien Nobelpreis für Chemie 1988, ging an drei deutsche Wissenschaftler:
Johann Deisenhofer — Robert Huber — Hartmut Michel am Max-Planck-Instituts für Biochemie in Planegg-Martinsried (bei München).

12.05.2016
34
Die Reaktionszentren von Höheren Pflanzen und photosynthetischen Bakterien sind homolog
RC höherer Pflanzen
bakterielles RC
Vielfalt von photosynthetischen MembranenLH
Die Reaktionszentren der Purpurbakterien sind von ringförmigen Chlorophyll-haltigen Antennen umgeben (Aufsicht)

12.05.2016
35
Vielfalt von photosynthetischen Membranen
RC
RC
Die Reaktionszentren der Purpurbakterien sind von ringförmigen chlorophyllhaltigen Antennen umgeben (Aufsicht)
schematisch
Vielfalt von photosynthetischen MembranenDie unterschiedliche Organisation der Lichtsammelkomplexe in Höheren Pflanzen (links) und
Purpurbakterien (rechts)
RCRC

12.05.2016
36
Phycobilisomen: Lichtsammelkomplexe der Cyanobakterien und Rotalgen (Seitenansicht)
Die Phycobilisomen können sich ablösen ( Regulation der Lichtzufuhr) und enthalten nur Phycobiline
Der Lichtsammelkomplex(Phycobilisom) sitzt auf dem core Komplex und ragt aus der Membran
Der transmembrane core Komplex (RC + core Antennen) ist homolog zu dem in Höheren Pflanzen
Membran
Phycobiline
Chlorophylle
Geldrollen-artige Strukturen Stapel von scheibenförmigen Phycobilin-Bindeproteinen
RC
Exitonentransfer in den Phycobilisomen

12.05.2016
37
Die unterschiedliche Architektur der Lichtsammelkomplexe in Cyanobakterien (oben) und Pflanzen (unten)
Membran
Membran
Seitenansicht
Antennen-Dissoziation
vertikal
Antennen-Dissoziation
horizontal
ZusammenfassungDie Reaktionszentren (RCs) aller photosynthetischen Organismen enthalten Chl a und sind homolog (hoch konserviert).
Die Lichtsammelkomplexe (LHCs) der verschiedenen Organismen unterscheiden sich dagegen grundlegend:
ringförmige LHCs bei Purpurbakterien (Pigmente: B-Chl)
Chlorosomen bei grünen Schwefelbakterien (phototroph obligate Anaerobier) auf der cytoplasmatischen Membranseite!
Phycobilisomen bei Cyanobakterien und Rotalgen (Pigmente: Phycobiline)
Periphere, mehr oder weniger mobile LHCs bei Höheren Pflanzen (Pigmente: Chl a und Chl b, nur mit Bindeproteinen assoziiert).
Struktur von Chlorobacten

12.05.2016
38
© 2015 American Society of Plant Biologists
Photosynthese I
• Photosynthese Einführung: Bedeutung, Evolution und Diversität
• Plastiden und Chloroplasten
• Licht und Pigmente
• Struktur und Funktion der Photosynthesekomplexe
• Linearer Elektronentransport und Wasserspaltung
Der Emerson-Effekt (oder Enhancement-Effekt, entdeckt 1957)

12.05.2016
39
Der Emerson-Effekt
•„Enhancement effect“: Überadditivität von rotem (R) und fern-rotem (FR) Licht bei der Photosynthese
Der Emerson-Effekt lässt sich nur durch die Existenz zweier Photosysteme erklären:
• P680 und P700 „Geburt des Z-Schema“, d.h. zwei Photosysteme arbeiten in Serie
• Der Cytochrom b6f-Komplex koppelt die beiden Photosysteme
• Mobile Elektronencarrier(Plastochinon und Plastocyanin) gewährleisten den Transport zwischen den Komplexen
Sta
ndar
dred
oxpo
tent
ial

12.05.2016
40
• Eigentliche endotherme Redoxreaktion, die durch Sonnenenergie ermöglicht wird.
• Elektronentransport über mehrere Komplexe und Redox-Carrier• Elektronen werden am OEC („oxygen-evolving complex“) vom PSII aus
Wasser „extrahiert)
Photosynthetischer linearer ElektronentransportOxidation von Wasser und Reduktion von NADPH
Der Cyt b6f -Komplex vermittelt beim Elektronentransport zwischen PSII und PSI
Damit fungiert er gleichzeitig als Protonenpumpe ( ATP-Synthese)
In Pflanzen und Cyanobakterien sind zwei Photosysteme(PSII und PSI) in Serie geschaltet
PSII entzieht dem Wasser Elektronen (e-)
PSI überträgt sie letztendlich auf NADP+ NADPH + H+
Abb. aus: Buchanan et al. „Biochemistry & Molecular Biology of Plants“; Amer. Soc. of Plant Physiol.
Fdx = FerredoxinFNR = Fdx-NADP-Reduktase

12.05.2016
41
© 2015 American Society of Plant Biologists
Struktur und Funktion von Photosystem II – LHCII complex
Reprinted by permission from Calderone, V., Trabucco, M., Vujičić, A., Battistutta, R., Giacometti, G.M., Andreucci, F., Barbato, R. and Zanotti, G. (2003). Crystal structure of the PsbQ protein of Photosystem II from higher plants. EMBO reports. 4: 900-905; Reprinted with permission © Annual Reviews Nickelsen, J. and Rengstl, B. (2013).Photosystem II assembly: From cyanobacteria to plants. Annu. Rev. Plant Biol. 64: 609 – 634.
Conserved reaction center core
PSII ist einMultiproteinkomplex der alsDimer funktioniert. Monomer:
D1D2
CP43 CP47
Dimer Struktur von PSIIDas konservierteReaktionszentrumbesteht aus D1 and D2, und den internenAntennenproteinenCP43 und CP47
Der SauerstoffbildendeKomplex besteht auseinem Mn4CaO5 cluster befindet sich auf der Seite des Lumens
H2O
1/2 O2 + 2 H+
© 2015 American Society of Plant Biologists
Die Proteinzusammensetzung von PSII kann man mittels SGC, SDS-PAGE und EM untersuchen
Reprinted with permission from Caffarri, S., Kouřil, R., Kereïche, S., Boekema, E.J. and Croce, R. (2009). Functional architecture of higher plant Photosystem II supercomplexes. EMBO J. 28: 3052-3063.
Sucrose gradient centrifugation (SGC)
He
avi
er
Lig
hte
r
Trennung der Proteinkomplexe mittelsSDS-PAGE
EM Untersuchung von PSII

12.05.2016
42
© 2015 American Society of Plant Biologists
Molekulare Struktur von PSII: Proteinepositionieren Pigmentmoleküle
Neveu,Curtis
© 2015 American Society of Plant Biologists
Elektronentransfer in PSII
(1) Licht regt Reaktionszentrum Chlorophyll (P680) an P680*(2) Elektron verlässt P680*, oxidiertes P680+
(Photooxidation, Ladungstrennung) (3) Pheophytin (Pheo) wird reduziert Pheo−
(4) Pheo− reduziert QA zu QA−
(5) QA− reduziert QB zu QB
−
P680 → P680* → P680+
Pheo → Pheo−
e-e−
(1) (2)
(3)
QA → QA−
e-e−
(4)
PSII
(4)
(3)
STROMA
LUMEN
(5)
QB → QB−
(5)
e-e−
12.05.2016
43
© 2015 American Society of Plant Biologists
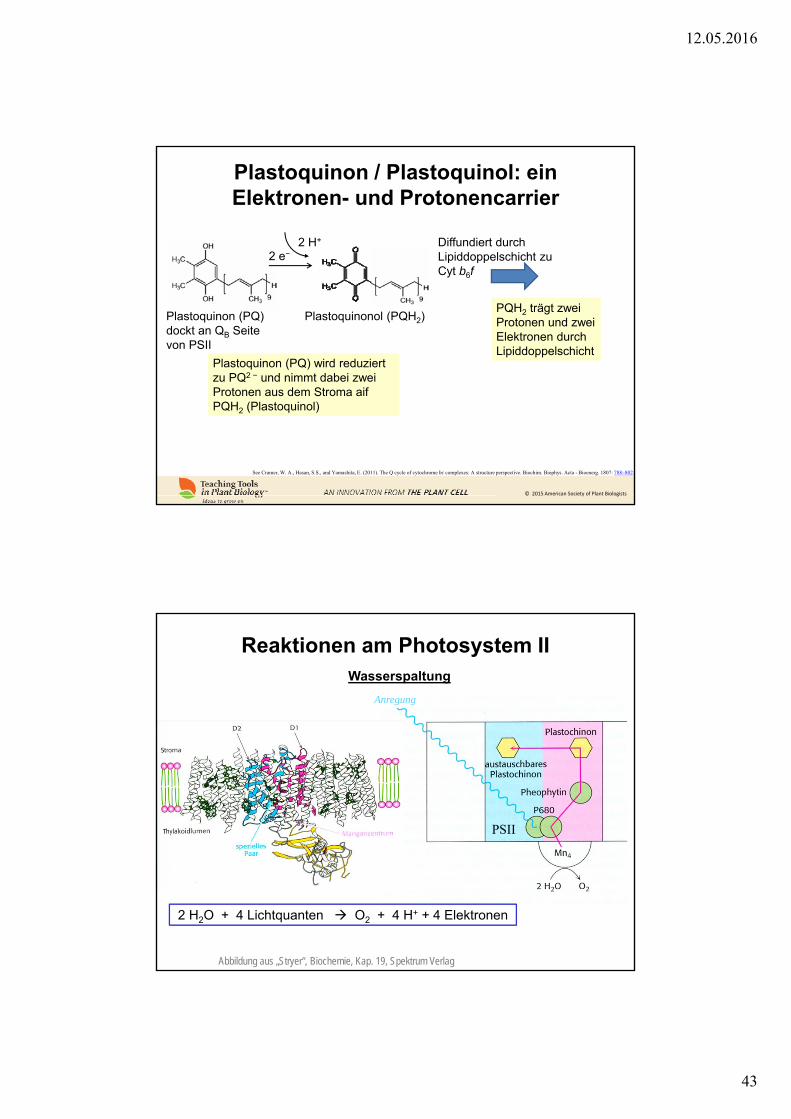
Plastoquinon / Plastoquinol: einElektronen- und Protonencarrier
See Cramer, W. A., Hasan, S.S., and Yamashita, E. (2011). The Q cycle of cytochrome bc complexes: A structure perspective. Biochim. Biophys. Acta - Bioenerg. 1807: 788–802.
Plastoquinon (PQ) wird reduziertzu PQ2 − und nimmt dabei zweiProtonen aus dem Stroma aifPQH2 (Plastoquinol)
Plastoquinon (PQ) dockt an QB Seitevon PSII
Plastoquinonol (PQH2)
2 e−2 H+ Diffundiert durch
Lipiddoppelschicht zuCyt b6f
PQH2 trägt zweiProtonen und zweiElektronen durchLipiddoppelschicht
Reaktionen am Photosystem II
Abbildung aus „Stryer“, Biochemie, Kap. 19, Spektrum Verlag
2 H2O + 4 Lichtquanten O2 + 4 H+ + 4 Elektronen
Wasserspaltung
PSII
Anregung

12.05.2016
44
Der Mn4-Komplex fungiert als Ladungs- bzw. „Redox“-Akkumulator (d.h. er akkumuliert zunächst selbst 4 „Elektronenlöcher“).
Durch die konzertierte Oxidation von 2 Mol H2O (Entzug von 4 Elektronen) wird die Bildung von destruktiven Sauerstoffradikalen stark gemindert.
2 H2O + 4 Lichtquanten 1 O2 + 4 H+Lumen + 4 e-
2 Moleküle H2O werden an einen Mangan-Komplex (Mn4) gebunden.
Nach Entzug von 4 Elektronen aus dem Wasser-Mn4-Komplex werden 4 H+
(Protonen) und ein O2 frei.
Wasserspaltung
Reaktionen am Photosystem II
ET
© 2015 American Society of Plant Biologists
Q Zyklus und Cytochrome b6f Komplex
Stroma(electro-
negative side)
Lumen(electro-
positive side)
PSII Reaction Center
Cytochromb6f complex
PSI Reaction Center
Mobiles PQH2
transportiert Elektronenzum Cyt b6f
Reprinted by permission from Baniulis, D., Yamashita, E., Zhang, H., Hasan, S.S. and Cramer, W.A. (2008). Structure–Function of the Cytochrome b6f Complex†. Photochem. Photobiol 84: 1349-1358.

12.05.2016
45
Q-Zyklus am Cytochrom b6f-Komplex
Bilanz: 1 PQH2 + 2 PCox + 4 H+(Stroma) 1 PQ + 2 PCred + 4 H+
(Lumen)Bilanz: 1 PQH2 + 2 PCox + 4 H+(Stroma) 1 PQ + 2 PCred + 4 H+
(Lumen)
Abb. aus: Buchanan et al. „Biochemistry & Molecular Biology of Plants“; Amer. Soc. of Plant Physiol.
2 e- 1 e-
Übergang2 e- 1 e-
Übergang
Die Reduktion von Plastochinon (PQ, 1) zu Plastochinol (PQH2, 3) erfolgt über das Semichinon-Radikal (PQH, 2).
2 mobile redox-Carrier: - PQ (Plastochinon): PSII Cyt b6f (in der Lipidphase) - PC (Plastocyanin): Cyt b6f PSI (im Thylakoid-Lumen)
Cytochrom b6f-Komplex: - vermittelt Elektronentransport (PSIIPQ und PCPSI) - fungiert als Protonenpumpe (StromaLumen)- homolog zum Cytochrom bc-Komplex der Mitochondrien (Atmungskette)
Photosynthetischer Elektronentransport (PET)
FNR = Ferredoxin-NADP-Reduktase
Abb. aus: Buchanan et al. „Biochemistry & Molecular Biology of Plants“; Amer. Soc. of Plant Physiol.
Fdx = FerredoxinFNR = Fdx-NADP-Reduktase

12.05.2016
46
Reaktionen am Photosystem IDonor: Plastocyanin (mobil im Thylakoid-Lumen, ca. 10 kDa),
kleines Protein mit redox-aktiver Kupfer-(Cu)-Gruppe
Akzeptor: Ferredoxin (auf Stroma-Seite von PSI, ca. 10 kDa),
kleines Protein mit redox-aktivem Fe/S-Cluster
4 PCred + 4 Quanten + 4 Fdox 4 PCox + 4 Fdred
PSI
Anregung
Abbildung aus „Stryer“, Biochemie, Kap. 19, Spektrum Verlag
Molekulare Struktur von PS IPSI besteht aus 17 Proteinuntereinheiten, vorallem PsaA and PsaB, and 178 prosthetischenGruppen, vor allemChlorophyll

12.05.2016
47
© 2015 American Society of Plant Biologists
In Pflanzen ist PSI halbmondförmig von LHCI Komplexen umgeben
Reprinted from Mazor, Y., Borovikova, A. and Nelson, N. (2015). The structure of plant photosystem I super-complex at 2.8 Å resolution. eLife. 4: e07433. Minagawa, J. (2013). Dynamic reorganization of photosynthetic supercomplexes during environmental acclimation. Front. Plant Sci. 4: 513.
PSI core
LHCI
Proteins Pigments
Sicht vom Lumen auf die Membran
ChlamydomonasTwo rows of LHCI
Pea complexSingle row of LHCI
© 2015 American Society of Plant Biologists
Ferredoxin transferiert Elektronen via Ferredoxin:NADP+ Reduktase (FNR)
Reprinted from Mulo, P. (2011). Chloroplast-targeted ferredoxin-NADP+ oxidoreductase (FNR): Structure, function and location. Biochim. Biophy. Acta (BBA) - Bioenergetics. 1807: 927-934 with permission from Elsevier.
From PSI
e-
NADP+ NADPH
e−H+ + 2
Ferredoxin: kleines Fe2S2
Protein
Ferredoxin (Fd) Ferredoxin: NADP+ reductase
Fe2S2
12.05.2016
48
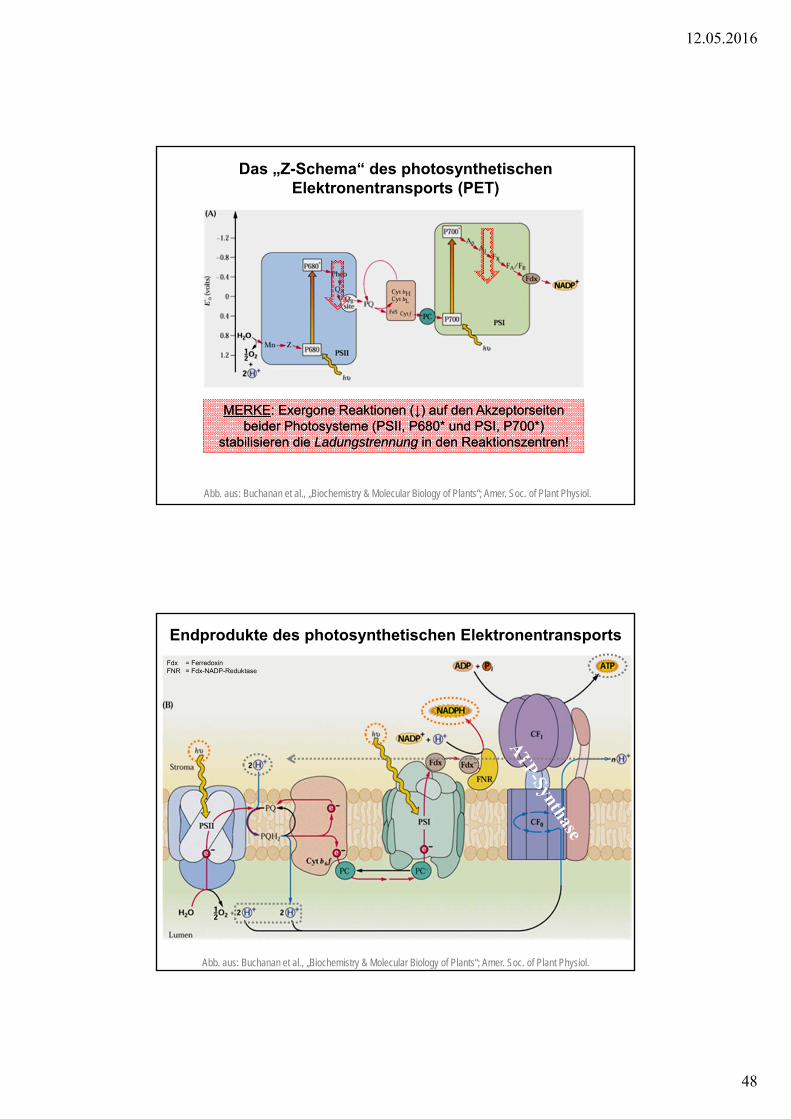
Das „Z-Schema“ des photosynthetischenElektronentransports (PET)
Abb. aus: Buchanan et al., „Biochemistry & Molecular Biology of Plants“; Amer. Soc. of Plant Physiol.
MERKE: Exergone Reaktionen (↓) auf den Akzeptorseitenbeider Photosysteme (PSII, P680* und PSI, P700*)
stabilisieren die Ladungstrennung in den Reaktionszentren!
MERKE: Exergone Reaktionen (↓) auf den Akzeptorseitenbeider Photosysteme (PSII, P680* und PSI, P700*)
stabilisieren die Ladungstrennung in den Reaktionszentren!
Abb. aus: Buchanan et al., „Biochemistry & Molecular Biology of Plants“; Amer. Soc. of Plant Physiol.
Fdx = FerredoxinFNR = Fdx-NADP-Reduktase
Endprodukte des photosynthetischen Elektronentransports

12.05.2016
49
Zusammenfassung
Die Photosysteme PSII und PSI sind in Serie geschaltet (bei Cyanobakterien und Pflanzen).
Photochemische Ladungstrennung am PSII führt zur Spaltung von Wasser.
Die konzertierte Spaltung von 2 Mol H2O wird durch einen Mangan-Komplex vermittelt.
Die dem Wasser entzogenen Elektronen werden auf Plastochinon (PQ); einen mobilen lipophilen Redox-Carrier in der Thylakoidmembran übertragen (2 e- + PQ + 2 H+ PQH2).
Photochemische Ladungstrennung am PSI führt zur Reduktion von NADP+ (NADPH + H+), Elektronendonor ist der mobile Redox-Carrier Plastocyanin (PC) im Lumen der Thylakoide.
Der Cytochrom b6f-Komplex vermittelt bei der Elektronenübertragung zwischen PQ (2 e-) und PC (1 e-) und fungiert gleichzeitig als Protonenpumpe (H+) Q-Zyklus.
Endprodukte des Licht-getriebenen Elektronentransports sind (neben O2 aus der Wasser-spaltung) NADPH und ATP, die den so genannten „Dunkel-Reaktionen“ (z.B. Calvin-Zyklus) im Chloroplasten-Stroma zur Verfügung stehen.