patterns of secondary succession in a mangrove forest of southern florida
TRANSCRIPT

Oecologia (Berl.) 44, 226-235 (1980) Oecologia �9 by Springer-Verlag 1980
Patterns of Secondary Succession in a Mangrove Forest of Southern Florida
Marilyn C. Ball ~
Department of Environmental Engineering, Post, Buckley, Schuh and Jernigan, Inc., 7500 NW 52 St., Miami, Florida, USA
Summary. Successional patterns were studied in mangrove forests which had developed recently in response to salinization of areas formerly supporting freshwater marshes along Biscayne Bay in North Miami, Florida. The population structures of these Induced Forests were compared with an adjacent Historical Forest which consisted of a nearly pure stand of Rhizophora mangle. A mixed forest of Rhizophora and Laguncularia raeemosa had developed in intertidal areas, while areas above the mean high water elevation supported a scrub community dominated by Laguncularia. Maxi- mum growth of both Rhizophora and Laguncularia occurred in intertidal areas, while both species were stunted and had sparse, poorly formed canopies in drier environments above the mean high water level. Analysis of population structure suggests that Induced Forests in intertidal areas are undergoing succession to a stand of Rhizophora. Laguncularia is unable to compete effec- tively with Rhizophora in these areas and it is suggested that it eventually will be limited to the drier areas, where competition from Rhizophora will be reduced or absent.
Introduction
Mangroves are the dominant form of coastal vegetation in South- ern Florida. There are three species: Rhizophora mangle L. (red mangrove), Laguncularia racemosa Gaernt. (white mangrove) and Avicennia germinans L. (black mangrove). 1 Although the composi- tion and structure of mangrove forests may vary considerably with local physical conditions, the mangrove species chaicacteristi- cally are distributed in a banded zonation pattern. Davis (1940) recognized seven distinct zones within this pattern. Some of these zones are dominated by a single species while others consist of a mixture of species.
The significance of zonation patterns has been a controversial topic in mangrove ecology. It has been argued that zonation pat- terns are equivalent to seral stages in succession to the climatic climax, a tropical forest (Davis, 1940; Chapman, 1944, 1970). In this Clementsian interpretation, the coastal zone is the pioneer
* formerly Kimball
a Scientific nomenclature throughout follows: Long, R.W. and Lakela, O. (1971) A Flora of Tropical Florida, University of Miami Press, Coral Gables, Florida Present address: Department of Environmental Biology, Research School of Biological Sciences, Australian National University, Canberra City, ACT 2601, Australia
stage and more landward zones are progressively later stages in succession, thereby implying seaward motion of the system (Davis, 1940; Chapman, 1944, 1970). This classical view is inconsistent with geological data; the predicted patterns of sediment accumula- tion are not always realized in successive zones (Egler, 1952) and the zonation patterns of some forests have existed in situ for millenia (Thom et al., 1975). Further, studies of the distribution of mangroves in relation to coastal geomorphology have shown that vegetational change occurs in response to changes in the physical characteristics of the environment (Thorn, 1967, 1975; Thom et al., 1975; Cintron et al., 1978).
Extensive changes in the vegetation of Southern Florida have been caused by alteration of the hydrological conditions by man during the twentieth century (Alexander and Crook, 1974). Hydrological changes leading to salinization of soil and water have induced the landward expansion of mangrove vegetation along Biscayne Bay into areas which formerly were freshwater marshes (Reark, 1975; Teas et al., 1976). The present study reports on species interactions in the development of a portion of this mangrove forest. Changes in the distribution of mangroves and marshes are mapped from aerial photographs and historical data. The population structure of a Historical Forest is compared with Induced Forests of recent origin and successional trends are inferred from these data. The results emphasize the role of compe- tition in the formation of zonation patterns.
Site History
Early observations of the vegetation along Biscayne Bay were recorded on survey maps made by William de Brahm in 1770 (Chardon, 1976). A mangrove forest was indicated near Arch Creek and the Oleta River (see Fig. 1); landward vegetation was described as ~ tall grass with scattered trees, subject to inundation" (Chardon, 1976). Bowman (1917) reported a dense forest of Rhizo- phora and other trees, presumably freshwater swamp hardwoods, along the banks of the Miami River, several kilometers to the south. Rhizophora growing along the river were 7-8 m in height, whereas only dwarf forms of Rhizophora were found in adjacent freshwater marshes, dominated by the sawgrass, Cladium ja- maicensis Crantz. These observations are consistent with vegeta- tion patterns in the Southeastern Everglades (Egler, 1952), and with geological studies of the Biscayne Bay area (Wanless, 1976) and probably are representative of conditions in the study area at the same period.
A reconstruction of the vegetation of the study area based on a 1928 aerial photograph is shown in Fig. 2 and probably
0029-8549/80]0044/0226/$2.00

[ ] Historical forest,1928 . . . . . Tidal creeks TP Tidal pond - - - - - - Forest boundaries 1977
Fig. 1. Map of the boundaries of the Historical Forest in 1928 and 1977 and the Induced Forest in 1977. Only the limits of the Induced Forest B study area are shown although the forest encompasses the Oleta River
approximates closely to the conditions before 1900. The location of study sites with respect to the historical vegetation is shown in Fig. 1. The vegetation consisted principally of a mangrove swamp (the Historical Forest) and marshes. The mangrove swamp was dominated by Rhizophora, and this species appears to have been abundant along the margins of the Oleta River and lesser drainages which are outlined on the photograph by dense growth of trees and shrubs. The cabbage palm (Sabal palmetto Lodd. ex Schultes.) and the buttonwood (Conocarpus erecta L.), both of which are tolerant of brackish conditions, occurred in the man- grove forest. Crowns of the palm are visible in the photograph and trunks of dead individuals of both species were found in the present field studies. Some areas near the mouth of Arch Creek may have supported salt marshes, probably composed of either Juncus or Spartina. Three tidal ponds separated the present mangrove forest from upland freshwater marshes, probably domi- nated by sawgrass (Cladium jamaicensis). Although it is now al- most absent from the area, clumps of living Cladium were found growing on spoil piles from 1935 1936 mosquito ditch excavations. This species reproduces almost entirely by vegetative means (Alex- ander, 1971) and may be a remnant of the original marsh,
The location of these plant communities before 1900 corre- sponds to the distribution of soils (Fig. 3). The location of the Historical Forest is marked by deep peat deposits which character- istically formed to depths of 1-4 m in the swamps of Biscayne Bay (Wanless, 1976). The landward edge of the peat probably represents the historical limits of tidal influence, as accumulation of red mangrove peat generally is limited to elevations in the upper half of the tidal range (Scholl, 1964). Freshwater marshes occurred on Perrine marl, which is of freshwater origin and typi- cally accumulates in marshes with a moderate growth of sawgrass
or a sparse covering of spike rush (Eleocharis cellulosa Torr.) (Craighead, 1974). The thin layer of organic materials covering marl in some intertidal areas is probably of recent origin.
A detailed account of urban development and associated hydrological changes which have caused the expansion of man- grove vegetation along Biscayne Bay has been reported by Teas et al. (1976). The Historical Forest probably developed at a time when the salinity regime ranged from fresh to slightly brackish during the wet and dry seasons, respectively (Teas et al., 1976). The mangroves were limited to this seasonally saline environment by their intolerance of the seasonal droughts and fires typical of freshwater wetlands in Southern Florida (Egler, 1952) and by their apparent inability to compete effectively with freshwater veg- etation (Thorn, 1967). Drainage of the Everglades and diversion of runoff reduced the height of the freshwater table and allowed intrusion of saline water into wetlands along Biscayne Bay (Bu- channon and Klein, 1976). Further, the excavation of mosquito ditches in 1935-1936 reduced impediments to the flow of tidal waters and facilitated the landward advance of the effective mean high water level (Teas et al., 1976). Freshwater marsh communities underwent succession to mangrove communities following hydro- logic changes leading to salinization (Alexander and Crook, 1974; Reark, 1975; Teas et al., 1976).
Landward expansion of mangroves at the expense of freshwater marshes occurred by 1938 (Fig. 2). Mosquito ditches appear to have promoted the spread of mangroves in western areas of the site, whereas mangroves expanded laterally from the margins of creeks and ponds in other areas. The white mangrove (Laguncula- ria) can be identified from the aerial photograph as the dominant canopy component in much of this new growth. In one area, the species composition of this new mangrove growth is not clear from the aerial photograph and is shown as "mixed mangrove" on the map. Marshes persisted over large areas.
In 1945 (Fig. 2) the freshwater marshes were reduced to patches encircled by mangrove vegetation. The composition of the Induced Forest at this stage is not clear from the aerial photographs. The canopy probably consisted of a mixture of both Rhizophora and Laguncularia; the former species may have been dominant along water courses but Laguncularia probably was the dominant species in most of the forest. The tidal ponds were reduced in size. Rhizophora colonized the shallow areas, forming scattered "tree islands".
In 1958 (Fig. 2), mangrove vegetation covered the entire wet- land area. The tidal ponds were reduced further in size and the merger of Rhizophora "tree islands" nearly covered the central pond.
Present Site Characteristics
The study was conducted in three Induced Forests (A, B and C) and one Historical Forest along Biscayne Bay in North Miami, Florida (Fig. 4), latitude 25 ~ North and longitude 80 ~ West. An- nual rainfall averages 1.64 m with most rain falling from June through October; mean annual temperature is 23.5~ (U.S. Weather Bureau, 1972). The mean high water elevation is +0.325m with a tidal range of approximately 0.27 m. Details of the hydrology have been presented elsewhere (Kimball et al., 1978). The distribution of marl and peat soils on the site is sum- marized in Fig. 3 (Leighty and Henderson, 1958).
The present site configuration as shown in Fig. 4 was formed by landfilling operations and construction of NE 151 Street in 1963 and NE 135 Street in 1969. The remaining area of Historical
227

LEGEND [ ] Rhizophora [ ] Upland, disturbed [ ] Marsh (salt) [ ] Mud flats, tree islands [ ] Open water [ ] taguncularia [ ] Mixed mangrove [ ] Marsh (fresh/ TP Tidal pond
Fig. 2. Change in vegetation from 1928-1958 based on aerial photographs in the years indicated
Forest together with a contiguous region of Induced Forest A occupy 85.7 hectares. The Historical Forest and most of Induced Forest A occupy an intertidal zone; a narrow band along the landward limits of Induced Forest A is at ground elevations above the mean high water level. Two 1.5 m culverts were installed be- neath NE 135 Street to permit exchange of tidal waters with North Biscayne Bay in which the salinities approach full strength seawater (i.e. 28-320o). Tidal waters are distributed throughout the site by a network of mosquito ditches. Water movement is vigorous in the southeastern area (Historical Forest) and decreases with distance north and west of a centrally located tidal pond to almost stagnant conditions near the landward limits of the intertidal zone.
Induced Forest B is an integral part of the remaining Oleta River system. The entire site occurs within an intertidal zone and receives natural overland flow of tidal water from the Oleta River. Although this area receives pulses of freshwaters during periods of storm runoff from urban areas, the surface waters generally are brackish (i.e. 15~ Tidal flushing is vigorous, al- lowing very little leaf litter to accumulate on the forest floor.
A tidal creek marks the eastern limit of the study area (Fig. 4). Induced Forest C covers an isolated tract of 4.9 ha. It previ-
ously was drained by a southern continuation of the same creek which flows through Induced Forest B. Mosquito ditches link this forest with North Biscayne Bay; however, the culverts are nearly filled with silt and debris and permit very little exchange of tidal waters. Although the ground elevations of Induced Forest C are less than the mean high water level of +0.325 m, it is effectively an upland area because normal tidal flooding has been prevented by surrounding development.
Method
Selection of study sites was-based on preliminary reconnaissance of the hydrologic and floristic characteristics in the general area. Vegetation was surveyed by field observations and false color infrared aerial photographs taken in April, 1974. Study sites were then selected in what were considered to be representative areas of the forests. Their locations are shown in Fig. 4.
228

Table 1. Characteristics of the mangrove vegetation
Living
Height (m)
D.B.H. (cm)
Relative density (%)
Basal area (m 2 ha- l)
Dominance (% total basal area)
Dead
Sapling density (m- 2)
Tree density (m- 2)
Tree D.B.H. (cm)
(a) Areas flooded by daily tides
(1) Historical Forest
Station 1 Rhizophora Laguncularia Station 2 Rhizophora Laguncularia Station 3 Rhizophora Laguncularia Station 4 Rhizophora Laguncularia Station 5 Rhizophora Laguncularia
(2) Induced Forests
(a) Induced Forest A
Station 1 Rhizophora Laguncularia Station 2
Rhizophora Laguncularia Station 3 Rhizophora Laguncularia Station 4 Rhizophora Laguncularia (b) Induced Forest B
Station 1 Rhizophora Laguncularia Station 2 Rhizophora Laguncularia Station 3 Rhizophora Laguneularia Station 4 Rhizophora Laguncularia Station 5 Rhizophora Laguncularia Station 6 Rhizophora Laguncularia
5
13
20
24
10 10
8 10
8 12
9 11
8 12
8 11
4.2 8.0
4.9
7.8
11.1
13.2
5.0 7.0
4.5 4.5
3.2 3.1
3.4 3.7
7.5 9.5
3.4 7.2
3.5 6.7
5.9 6.0
3.5 6.7
3.9 8.9
87.5 12.5
100.0 0.0
100.0 0.0
100.0 0.0
100.0 0.0
90.0 10.0
57.1 42.9
46.5 53.5
43.9 56.1
83.3 16.7
55.0 45.0
75.0 25.0
75.0 25.0
63.2 36.8
58.3 41.7
4.2 2.0
13.3
13.7
12.5
35.7
16.4 3.1
11.7 8.7
6.8 7.0
6.9 9.9
7.1 2.8
4.1 16.0
6.5 7.1
12.2 3.9
4.9 10.8
3.7 14.7
67.5 32.5
100.0 0.0
100.0 0.0
100.0 0.0
100.0 0.0
83.9 16.1
57.4 42.6
49.4 50.6
41.1 58.9
71.3 28.7
20.4 79.6
48.1 51.9
76.0 24.0
31.0 69.0
20.0 80.0
0.04 0.0
0.01 0.0
0.0 0.01
0.0 0.0
0.0 0.0
0.04 0.04
0.04 0.0
0.08 0.44
0.16 0.52
0.0 0.0
0.24 0.12
0.96 0.0
0.0 0.0
0.28 0.20
0.12 0.60
0.0l 0.0
0.0 0.0
0.06 0.0
0.05 0.0
0.0 0.0
0.16 0.20
0.0 0.20
0.0 0.08
0.04 0.0
0.04 0.04
0.24 0.20
0.0 0.08
0.12 0.24
0.0 0.72
0.0 0.72
3.2
m
3.8
5.5
m
3.2 3.9
2.8
m
2.8
2.7
2.7 3.7
3.1 3.3
m
2.8
3.7 3.7
m
3.1
3.5
229
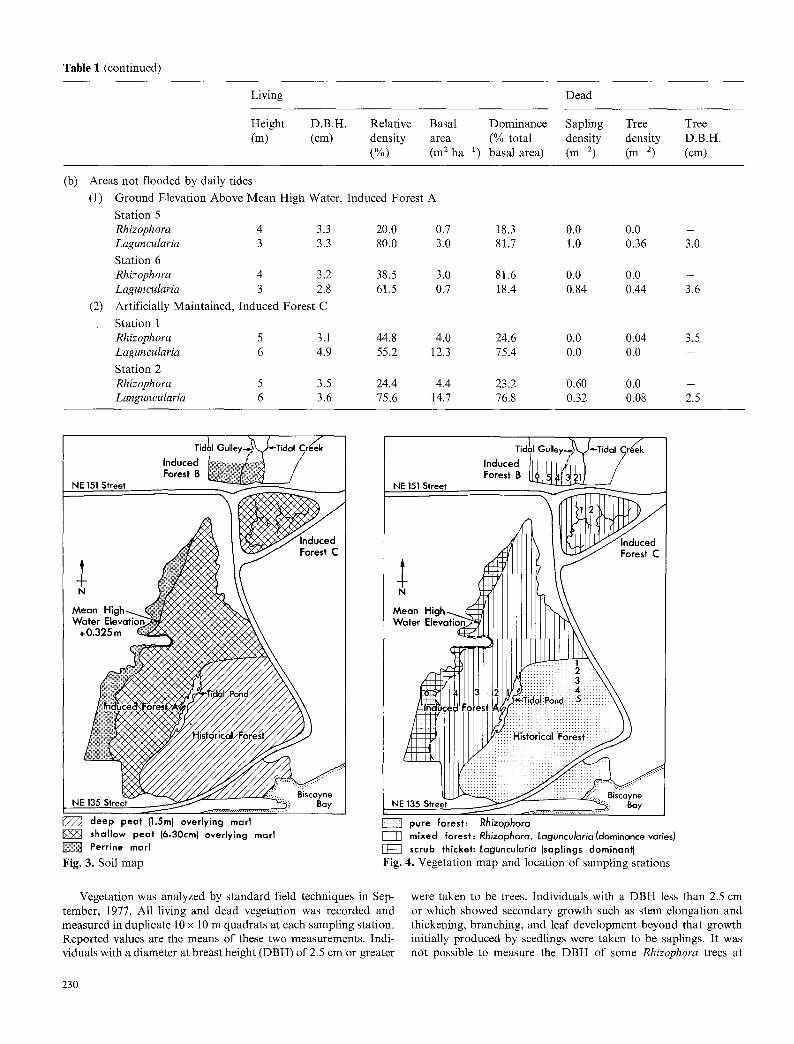
Table 1 (continued)
Living
Height D.B.H. Relative Basal (m) (cm) density area
(%) (m 2 ha *)
Dead
Dominance Sapling Tree Tree (% total density density D.B.H. basal area) (m 2) (m 2) (cm)
(b) Areas not flooded by daily tides
(1) Ground Elevation Above Mean High Water, Induced Forest A
Station 5 Rhizophora 4 3.3 20.0 0.7 Laguncularia 3 3.3 80.0 3.0
Station 6 Rhizophora 4 3.2 38.5 3.0 Laguncularia 3 2.8 61.5 0.7
(2) Artificially Maintained, Induced Forest C
Station 1 Rhizophora 5 3.1 44.8 4.0 Laguncularia 6 4.9 55.2 12.3
Station 2 Rhizophora 5 3.5 24.4 4.4 Languncularia 6 3.6 75.6 14.7
18.3 0.0 0.0 - 81.7 1.0 0.36 3.0
81.6 0.0 0.0 - 18.4 0.84 0.44 3.6
24.6 0.0 0.04 3.5 75.4 0.0 0.0 -
23.2 0.60 0.0 - 76.8 0.32 0.08 2.5
Tidal Gulley-~'~" Tidal Creek Induced l~ v.~: ~,~:~::: ,~?~ / Forest B !~i~is /
J ~ deep peat {1.5m) overlying marl shallow peat (6-30cm) overlying marl Perrine marl
Fig. 3. Soil map
, ,~, Tidal Gulley.~'~Tidal Creek Induced I ,' j Forest B ~ /
h
M~ V~
NE
pure forest: Rhizophora mixed forest: Rhizophora, Laguncularia(dominance varies)
j z ~ scrub thicket'. Laguncularia (saplings dominant) Fig. 4. Vegetation map and location of sampling stations
Vegetation was analyzed by standard field techniques in Sep- tember, 1977. All living and dead vegetation was recorded and measured in duplicate 10 x 10 m quadrats at each sampling station. Reported values are the means of these two measurements. Indi- viduals with a diameter at breast height (DBH) of 2.5 cm or greater
were taken to be trees. Individuals with a DBH less than 2.5 cm or which showed secondary growth such as stem elongation and thickening, branching, and leaf development beyond that growth initially produced by seedlings were taken to be saplings. It was not possible to measure the DBH of some Rhizophora trees at
230

1.4 m above ground because of the prop root structure. In these cases trunk diameters were measured above the insertion of the highest prop root. Calculations of Rhizophora basal area were made from these trunk measurements and do not include the area occupied by prop roots. Tree heights were estimated by trian- gulation and use of aerial photographs.
Observations
The distribution of mangrove vegetation in 1976 is shown in Fig. 4. The Historical Forest is composed of an almost pure stand of Rhizophora mangle. Mature individuals of Avieennia germinans (DBH approximately 20-30 cm) are very rare in this community, and did not occur in the study plots. The forest has a wedge shaped profile as shown by the increase in height from approxi- mately 5 m at the natural northern boundary to an average of 2(~24 m in the interior (Table 1). These trees are straight boled and well formed, averaging 13.2 cm DBH with some individuals measuring as much as 27 cm DBH. Trunks of dead buttonwood (Conoearpus erecta) with DBH values of approximately 76 cm occur near the northern periphery and stumps of dead cabbage palm (Sabal palmetto) are scattered widely in the forest. Dead mangroves and living saplings are almost entirely Rhizophora; however, seedlings of both Rhizophora and LaguncuIaria are abun- dant at the forest edge where tree growth is relatively poor, and become scarce towards the center of the forest (Fig. 5). In the following text, I shall compare the characteristics of the best devel- oped region of the Historical Forest, represented by Station 5, with those of the Induced Forests.
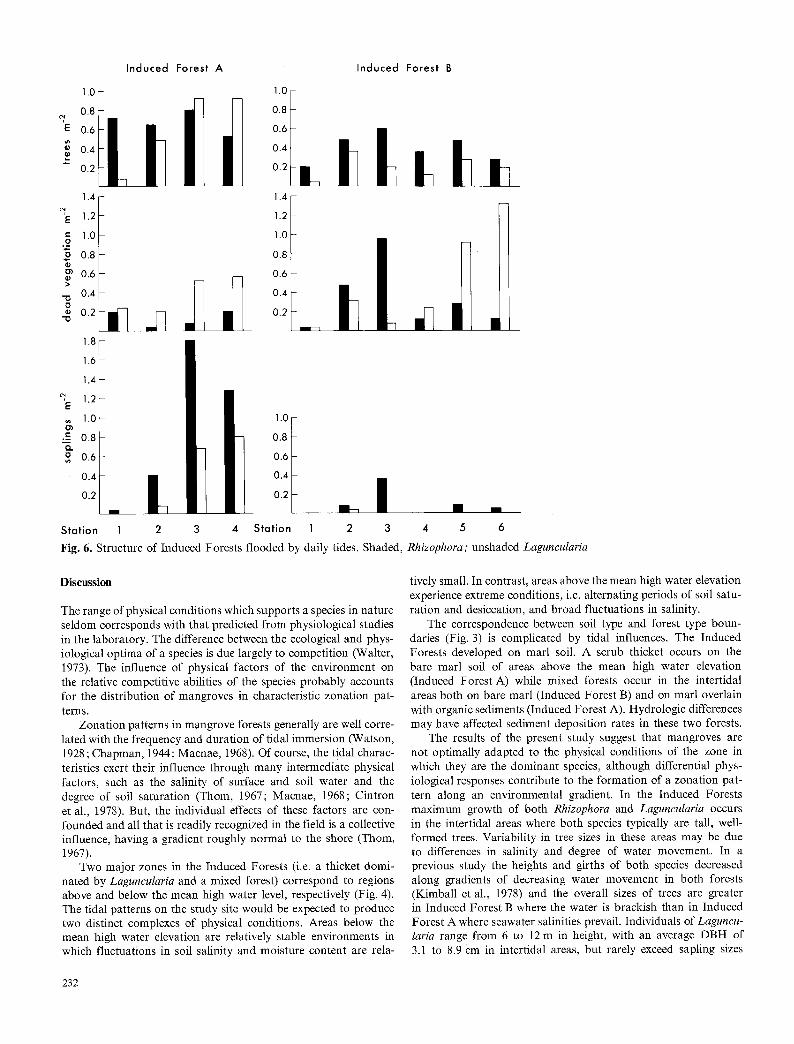
The intertidal region of Induced Forest A supports a mixed forest of Rhizophora and Laguncularia (Fig. 4). The structure and vigor of this forest is influenced by local hydrological conditions. Rhizophora generally is the most abundant species and is dominant in areas of most vigorous water movement, i.e. in the vicinity of the tidal pond (Stations 1 and 2), whereas Laguncularia generally is dominant in areas of sluggish water movement to the west. Tree density is approximately 0,8 m - z at Station 1 and increases along a gradient of decreasing water movement to approximately 1.6 m z at Station 4 (Fig. 6). These values are 3 to 6 times greater than those in the Historical Forest. There is a concomitant reduc- tion in height and DBH with increasing density (Table 1). Laguncu- laria accounts for the majority of dead vegetation (Fig. 6), most of which are trees at Stations 1 and 2 and saplings at Stations 3 and 4 (Table 1). Saplings of both species are present in the forest, with the largest populations occurring in areas dominated by La- guncularia (Fig. 6). Seedlings of both species are found throughout the forest, with densities of Rhizophora and Laguncularia ranging from 0 to 2.4 m - 2 and 0 to 3.6 m - 2, respectively.
The structure of Induced Forest B is similar to that in the intertidal region of Induced Forest A, and is affected by local hydrological differences associated with creeks and gullies. Rhizo- phora is the most abundant species in all areas of the forest (Fig. 6) but is dominant in areas of most vigorous water movement, i.e. near the tidal creek (Station 1) and near the tidal gulley (Stations 3 and 4) while Laguneularia generally is dominant in the forest interior (Table 1). Tree densities range from values similar to those in the Historical Forest (0.24 m 2) to values approximately 3 times greater. Dead vegetation consists mainly of trees of Laguncu- laria (Table 1). The sapling population consists a lmos t entirely of Rhizophora, ranging in density from 0 to 0.36 m - 2. Seedlings of both species are scattered at relatively low densities throughout the forest.
1~ i 0.8 E 0.6
0.4
- ! | 0.2 �9 |
E 1.0 I ._~ 0.8 "fi 0.6
0.4
-a| 0 " 2 ~ ~ 1 : 3 ~ ~ --a
0.8
0,6
�9 ~ 0.d
02 | 1.6-
1.4-
~, 1 . 2 -
E 1.0 o . ~ -
B
i: ~' 0 .4 -
0 .2 -
Station 1 2 3 d 5
Fig. 5. Structure of the Historical Forest. Shaded Rhizophora; unshaded Laguncularia
Areas of Induced Forest A which are not flooded by daily tides are characterized by a dense covering of scrub mangroves, dominated by Laguncularia (Fig. 4). Trees are sparse, dwarfed, gnarled and very poorly formed, reaching average heights of 3-4 m (Table 1). Most of the community consists of a dense population of saplings of Laguncularia (Fig. 7). Dead vegetation is composed almost entirely of Laguncularia (Fig. 7). The sapling density of Rhizophora decreases while that of Laguncularia increases with distance landward of the mean high water elevation. The densities of Rhizophora and Laguncularia seedlings are 1.2 and 0.08 m - 2, respectively.
Induced Forest C is composed of a mixture of Rhizophora and Laguncularia. The latter species is dominant in all areas of the forest accounting for approximately 55 to 76% of the trees near the creek (Station 1) and in the interior (Station 2) respec- tively (Table 1). Tree density is 4 to 7 times greater than that in the Historical Forest. It ranges from 1.06 m -2 near the creek to 1.80 m -2 in the interior, most of the difference being due to an increase in Laguncularia density (Fig. 7). Rhizophora accounts for most of the dead vegetation, as trees at Station 1 and as saplings at Station 2 (Table 1). Saplings of Rhizophora occur throughout the forest but saplings of Laguneularia are found only in the interior where the total sapling density is greatest. Seedlings are very sparse and consist mostly of Rhizophora (0.08 m - 2).
231
Induced Forest A Induced Forest B
1.0 I.O ~ ~ [ ~ ~ ~ 0.8 0.8
i
0.6 0.6
o �9 o.4 0.4 '- 0.2 0.2
1.4 o l
i E 1.2 g ~.o
2 0.8 m 0.6 >
0.4 O
�9 0.2 i-] "o
1 . 8 -
1 . 6 -
1 . 4 -
1.2- E 1.o-
�9 -= 0.8- 0 , , . o 0.6-
0.4 0.2
1.0
t
1.0 0.8 0.6 0.4 0.2
Stat ion 1 2 3 4 S ta t ion 1 2 3
Fig. 6. Structure of Induced Forests flooded by daily tides. Shaded,
�9 l
4 5 6
Rhizophora; unshaded Laguncularia
Discuss ion
The range of physical conditions which supports a species in nature seldom corresponds with that predicted from physiological studies in the laboratory. The difference between the ecological and phys- iological optima of a species is due largely to competition (Walter, 1973). The influence of physical factors of the environment on the relative competitive abilities of the species probably accounts for the distribution of mangroves in characteristic zonation pat- terns.
Zonation patterns in mangrove forests generally are well corre- lated with the frequency and duration of tidal immersion (Watson, 1928 ; Chapman, 1944; Macnae, 1968). Of course, the tidal charac- teristics exert their influence through many intermediate physical factors, such as the salinity of surface and soil water and the degree of soil saturation (Thom, 1967; Macnae, 1968; Cintron et al., 1978). But, the individual effects of these factors are con- founded and all that is readily recognized in the field is a collective influence, having a gradient roughly normal to the shore (Thom, 1967).
Two major zones in the Induced Forests (i.e. a thicket domi- nated by Laguncularia and a mixed forest) correspond to regions above and below the mean high water level, respectively (Fig. 4). The tidal patterns on the study site would be expected to produce two distinct complexes of physical conditions. Areas below the mean high water elevation are relatively stable environments in which fluctuations in soil salinity and moisture content are rela-
tively small. In contrast, areas above the mean high water elevation experience extreme conditions, i.e. alternating periods of soil satu- ration and desiccation, and broad fluctuations in salinity.
The correspondence between soil type and forest type boun- daries (Fig. 3) is complicated by tidal influences. The Induced Forests developed on marl soil. A scrub thicket occurs on the bare marl soil of areas above the mean high water elevation (Induced Forest A) while mixed forests occur in the intertidal areas both on bare marl (Induced Forest B) and on marl overlain with organic sediments (Induced Forest A). Hydrologic differences may have affected sediment deposition rates in these two forests.
The results of the present study suggest that mangroves are not optimally adapted to the physical conditions of the zone in which they are the dominant species, although differential phys- iological responses contribute to the formation of a zonation pat- tern along an environmental gradient. In the Induced Forests maximum growth of both Rhizophora and Laguncularia occurs in the intertidal areas where both species typically are tall, well- formed trees. Variability in tree sizes in these areas may be due to differences in salinity and degree of water movement. In a previous study the heights and girths of both species decreased along gradients of decreasing water movement in both forests (Kimball et al., 1978) and the overall sizes of trees are greater in Induced Forest B where the water is brackish than in Induced Forest A where seawater salinities prevail. Individuals of Laguncu- laria range from 6 to 12 m in height, with an average DBH of 3.1 to 8.9 cm in intertidal areas, but rarely exceed sapling sizes
232

I nduced Forest I n d u c e d Forest
A C
1.4 1.4 -
1.2 1.2 - /
, .o 1.o - I
E 0.8 0.8 -
0.4 0.4
0 . 2 ~ 0 . 2
~, 1.4 1.4 ~-
I:: 1.2 1.2
1.0 1.0 *.,=
0.8 o, 0.8 o o~ 0.6 0.6
"o 0.4 0.4 O �9 0.2 0.2
" 0
2.4 2.4~ 2.2 2.2 -
2.0 2.0
1.8 1.8
1.6 1.6
1.4 1.4
1.2 T.2
1.0 1.0
" 0.8 0.8
0.6 0.6
0.4 0.4
0.2 ~ _ ~ 0.2
Stat ion 5 6 1 2
Fig. 7. Structure of Induced Forests not flooded by daily tides. Shaded, Rhizophora ; unshaded, Laguncularia
in areas above the mean high water elevation (Induced Forest A). Trees in the latter areas are stunted (height 3 m), gnarled and have an average DBH of 3 cm. Individuals of Rhizophora in interti- dal areas average from 5 to 10 m in height and from 3.4 to 7.5 cm in DBH; however, trees at higher elevations (Induced Forest A) are dwarfed, having mean height and DBH values of approxi- mately 4 m and 3.3 cm respectively. Trees in the intertidal areas are much more vigorous in appearance than those in the adjacent upland environment. Clearly, growth of Laguncularia is not opti- mally adapted to the drier environments above the mean high water level where this species most often is dominant and Laguncu- laria would dominate both zones in the absence of competition from Rhizophora.
The effect of competition on the distribution of mangrove species along environmental gradients is suggested by the popula- tion structure of the developing forests. The intertidal mixed man- grove communities' of Induced Forest A and B appear to be under- going changes which may culminate in a nearly pure stand of Rhizophora (Fig. 6). Although Rhizophora is the most abundant
species, Laguncularia accounts for most of the dead vegetation. The dead are mostly trees (Table 1). In many areas Laguncularia appears to have been killed when overtopped by Rhizophora. Thus, live trees of Laguncularia are generally as tall, and usually taller than those of Rhizophora (Table 1). Large individuals of Laguncu- laria may remain in the forest canopy with no apparent loss of vigor but when one of these trees dies it is likely to be replaced by Rhizophora because most of the saplings are Rhizophora (Fig. 6).
Propagules are widely dispersed by the tides and are readily established everywhere, as shown by the presence of seedlings of both species throughout the intertidal forests. Propagules of both species become established in both shade and full sunlight. However, seedlings of Rhizophora tend to persist for a longer period than those of Laguncularia in deep shade (Rabinowitz, 1978), presumably because of larger storage reserves.
The number and species distribution of seedlings which develop into saplings appear to be affected by the canopy composition, possibly because of the effect of canopy structure on the intensity of light. Teas (1974) reported that the intensities of light transmit- ted through the canopies of Laguncularia and Rhizophora commu- nities along Biscayne Bay were approximately 70% and 20-30% respectively of the incident light. Therefore, the amount of light reaching the floor of mixed forests is less the greater the frequency of trees of Rhizophora. Saplings of Rhizophora are present in most areas of the forest, whereas Laguncularia saplings occur only in areas where at least 40% of the trees are Laguncularia. The density of Rhizophora saplings is also greater in these mixed stands than in pure stands of Rhizophora. Laguncularia appears to be shade- intolerant; hence, increase in canopy density reduces, and even- tually eliminates, development of Laguncularia saplings. In con- trast, Rhizophora appears to be shade-tolerant; saplings develop in all areas of the forest, thereby increasing the chance that Rhizo- phora would replace a dead resident. Thus, although propagules of both species may become established beneath the dense canopy of Rhizophora, it produces light conditions which generally are inhibitory to seedling growth in both species, but which selectively favor development of Rhizophora (refer Fig. 5).
These successional trends may lead to the development of a nearly pure stand of Rhizophora similar to the Historical Forest. This forest appears to be a stable self-maintaining community as shown by the population structure illustrated in Fig. 5. Dead vegetation consists almost entirely of Rhizophora. The dead are mostly trees (Table 1), but one dead Laguncularia sapling was found at Station 3 (Fig. 5). A dead tree is most likely to be replaced by a Rhizophora, because all the saplings found were of this species, despite an abundance of both Rhizophora and Laguneularia seed- lings throughout the forest. The characteristics of this forest are similar to those reported for mature Rhizophora forests in South- western Florida, Central America and the Caribbean (Pool et al., 1977).
The successional trends in areas above the mean high water level differ markedly from those in intertidal areas. The vegetation is characterized by a dense thicket, predominantly composed of Laguncularia saplings and interspersed with widely scattered dwarf trees of both Rhizophora and Laguncularia (Table 1). This thicket of scrub mangroves may be a stable self-maintaining community although the stability is not clear from the population structure (Fig. 7). There is an abundance of established seedlings ; however, relatively few seedlings of Rhizophora develop and the growth of Laguncularia produces a dense thicket of saplings. Thus, in spite of being a shade-tolerant species, which presumably is an advantage in a dense thicket, Rhizophora development probably is limited by the soil/water conditions. Few individuals of Laguncu-
233

laria exceed sapling size presumably due to edaphic limitations on growth, and a high mortality rate. However, establishment and growth of Laguncularia may be sufficient to maintain commu- nity composition because similar scrub Laguncularia communities characteristically occur in areas of Florida and the Caribbean which are flooded only by spring or storm tides (Chapman, 1944).
Contrast of Induced Forests B and C shows the importance of externally imposed changes in the physical environment on the composition of mangrove communities. It is evident from examination of aerial photographs that the vegetation of Induced Forest C was at the same stage of development as Induced For- est B, prior to the separation of these two forests by development in 1963. Since that time, Induced Forest C has been deprived of daily tidal flushing: in effect, it is as though ground elevations were above the mean high water level.
Induced Forest C is a mixed community dominated by Lagun- cularia. The trees are smaller and more closely spaced than those in intertidal areas; DBH values are 50-60% less than those in comparable areas (Stations 1 and 2) of Induced Forest B. Many Rhizophora near to and lining the creeks are either dead or stressed in appearance, with poorly developed canopies and numerous dead branches. Trees of Laguncularia generally are larger and appear to be healthier; however, false color infrared photographs taken in April, 1974, suggest that the entire forest is physiologically stressed.
Comparison of the floristics at Stations 1 and 2 in Induced Forest B with the comparable station in Induced Forest C shows that the composition of these two forests is not comparable in 1977. Near the creek in Induced Forest B, (Station 1), 83% of the trees are Rhizophora. Thus succession to a pure stand of this species is nearly complete. In the interior of Induced Forest B (Station 2), Rhizophora accounts for 55% of the trees. Succession seems to be proceeding at a slower rate (see Fig. 6). In contrast, Laguncularia dominates a nearly even mixture (1 : 1) of Rhizophora and Laguncularia near the creek (Station 1) in Induced Forest C. In the interior (Station 2) 75% of the trees are Laguncularia and the density of this species is 1.5 times greater than at Station 2 in Induced Forest B. The population structure illustrated in Fig. 7 does not show a clear successional trend in Induced Forest C; however, it appears that Laguneularia can retain dominance under the drier conditions because of reduced competition with Rhizo- phora while comparable areas receiving tidal flooding (Induced Forest B) are undergoing succession to a Rhizophora community.
Conclusion
A sequence of changes in vegetation at the study sites is suggested by the observations. Seedlings colonize areas made available by disturbances in hydrologic conditions. Any species is able to colo- nize, and the species composition of the seedlings that are initially established reflects that of the propagules that happen to arrive as has been shown by the rapid establishment of mangroves both on dredged materials in Southwestern Florida (Teas et al., 1975) and in clear-cut areas of Mexico (Thom, 1967) and Puerto Rico (Wadsworth, 1959). The general absence of Avicennia germinans in the Induced Forests may be attributed to physical factors affect- ing the dispersal of propagules. Sources of propagules on the site itself are limited by the rarity of mature trees in the Historical Forest. In other areas of northern Biscayne Bay, Avicennia seed- lings have established readily on dredged embankments (Teas, 1974). Thus, viable seedlings of Avicennia do occur, but the direc- tion of winds and currents may prevent substantial transport of propagules to the study sites.
Competition is not manifest during the early phases of colo- nization and mixed stands of shade-tolerant (eg Rhizophora) and intolerant (eg Laguncularia) species become established. Laguncu- laria has a greater growth rate than Rhizophora and attains heights of 1.5 m within a year (Wadsworth, 1959) whereas individuals of Rhizophora seldom exceed seedling size during the same time period (Lugo and Snedaker, 1975; Teas et al., 1975). These obser- vations of growth are supported by measurements of photosyn- thesis under field conditions along Biscayne Bay. Moore et al. (1973) found that rates of assimilation in sun leaves in mature individuals of Laguncularia exceeded those of Rhizophora growing in the same environment throughout the year. Thus shade-tolerant and intolerant species continue to coexist because individuals of the latter overtop those of the former.
Competition becomes critical as individuals reach maturity and require more space and other resources. These established individ- uals affect the development of later colonists. Conditions favoring maximum growth and development of both Rhizophora and La- guncularia occur in the intertidal zone. However, the ability of Laguncularia to maintain itself as a viable component of the com- munity is so affected by competition from Rhizophora, that the former species is eventually limited to the less desirable habitats of areas above the mean high water level. In these areas competi- tion from Rhizophora is greatly reduced or absent. These succes- sional trends are expected to culminate in a zonation pattern similar to that described by Chapman (1944) for Florida and the Caribbean in which Laguncularia generally occurs in drier environments, flooded only by spring or storm tides (approxi- mately 15~250 or d~100 .submersions per annum respectively) whereas Rhizophora inhabits the intertidal areas.
Although the above system predicts the development of mono- specific zones of Rhizophora in intertidal areas, mixed forests of Avicennia, Laguncularia and Rhizophora occur in other areas of Biscayne Bay (Teas, 1974) and Southwestern Florida (Lugo and Snedaker, 1975; Pool et al., 1977). Waterborne propagules are distributed seasonally and cotyledonary reserves allow temporary development of seedlings under conditions which otherwise are un- suitable for growth. Seedlings of shade-intolerant species may de- velop into saplings and secure space and other resources when they become established beneath a break in the forest canopy. Small breaks, such as those caused by lightning strikes, occasion- ally may enable a shade-intolerant species to become a canopy member. However, large breaks are caused periodically by hurri- canes (Craighead and Gilbert, 1962) and may be the most impor- tant factor maintaining the diversity of intertidal mangrove com- munities (Lugo and Snedaker, 1975; Pool et al., 1977). Thus an intertidal mangrove forest could be envisaged as a mosaic of differ- ent successional stages where localized disturbances promote the establishment of less shade-tolerant species, even in a mature com- munity.
Zonation patterns appear to result from the differential in- fluence of physical factors on the competitive abilities of the man- grove species. Different patterns of succession would be expected within each zone or set of physical conditions, and these patterns would be independent of biotic interactions in adjacent zones. Once the vegetation forming a zone reaches equilibrium, the com- munity is not likely to change in the absence of an externally imposed perturbation on the environment. The spatial distribution of species forming zones cannot be regarded as equivalent to the temporal sequence of species forming seres.
Acknowledgements. Technical assistance of John Buckley, P.E. and Thomas Checca, P.E., of Post, Buckley, Schuh and Jernigan, Inc.
234

is greatly appreciated. Thanks are due to John Acton and Martin Crowl for help with field collection of data, and to Said Haddad of Munisport, Inc. for support and cooperation. It is a pleasure to thank Professor Bruce Thorn and Professor Ian Cowan and Drs Eldon Ball, Graham Farquhar, Malcolm Gill, Ian Noble and Hank Shugart for their comments and helpful suggestions in the preparation of the manuscript.
References
Alexander, T.R.: Sawgrass biology related to the future of the Everglades ecosystem. Soil Crop Sci. Soc. Fla. Proc. 31, 72-74 (1971)
Alexander, T.R., Crook, A.G.: Recent vegetational changes in Southern Florida. In: Environments of South Florida: Past/ Present/Future (P.J. Gleason, ed.), pp. 61-77. Miami: Miami Geological Society 1974
Bowman, H.H.M. : Ecology and physiology of the red mangrove. Proc. Amer. Philos. Soc. 56, 589-672 (1917)
Buchanan, T.J., Klein, H. : Effects of water management on fresh- water discharge to Biscayne Bay. In: Biscayne Bay: Past/Pre- sent/Future (A. Thorhaug A. Volker, eds.), pp. 271-278. Coral Gables: University of Miami Press 1976
Chapman, V.J.: The 1939 Cambridge University expedition to Jamaica. J. Linn. Soc. London 52, 407-533 (1944)
Chapman, V.J. : Mangrove phytosociology. Trop. Ecol. 11, 1-19 (1970)
Chardon, R.E. : A geographical history of the Biscayne Bay area. In: Biscayne Bay: Past/Present/Future (A. Thorhaug A. Volker, eds.), Coral Gables: University of Miami Press 1976
Cintron, G., Lugo, A.E., Pool. D.J., Morris, G.: Mangroves of arid environments in Puerto Rico and adjacent islands. Biotropica 10, 110-121 (1978)
Craighead, F.C.: The Trees of South Florida: Volmaae 1. The Natural Environments and Their Succession. Coral Gables: University of Miami Press 1974
Craighead, F.C., Gilbert, V.C.: The effects of hurricane Donna on the vegetation of Southern Florida. Q.J. Fla. Acad. Sci. 25, 1-28 (1962)
Davis, J.H. Jr.: The ecology and geologic role of mangroves in Florida. Carnegie Inst. Wash. Pub. 32, 305 412 (1940)
Egler, F.E. : Southeast saline Everglades vegetation, Florida, and its management. Vegetatio 3, 213-265 (1952)
Kimball, M.C., Checca, T., Buckley, J.D.: Water management for preservation of a mangrove forest. In: Proceedings of the Fifth Annual Conference on Restoration of Coastal Vegetation in Florida. (R.R. Lewis, ed.), in press, Tampa: Hillsborough Community College 1978
Leighty, R.G., Henderson, J.R.: Soil survey of Dade County, Florida. Florida: U.S. Dept Agric. 1958
Long, R.W., Lakela, O.: A Flora of Tropical Florida. Coral Ga- bles: University of Miami Press 1971
Lugo, A.E., Snedaker, S.C.: Properties of a mangrove forest in Southern Florida. In : Proceedings of the International Sympo- sium on Biology and Management of Mangroves (G.E. Walsh, S.C. Snedaker, H.J. Teas, eds.), pp. 170~12. Gainesville: University of Florida Press 1975
Macnae, W.: A general account of the fauna and flora of the mangrove swamps and forests in the Indo-Pacific region. Adv. Mar. Biol. 6, 73-270 (1968)
Moore, R.T., Miller, P.C., Ehleringer, J., Lawrence, W. : Seasonal trends in gas exchange characteristics of three mangrove species. Photosynthetica 7, 387-394 (1973)
Pool, D.J., Snedaker, S.C., Lugo, A.E.: Structure of mangrove forests in Florida, Puerto Rico, Mexico and Costa Rica. Biotropica. 9, 195-212 (1977)
Rabinowitz, D. : Mortality and initial propagule size in mangrove seedlings in Panama. J. Ecol. 66, 45-52 (1978)
Reark, J.B.: A history of the colonization of mangroves on a tract of land on Biscayne Bay, Florida. In: Proceedings of the International Symposium on the Biology and Management of Mangroves. (G.E. Walsh, S.C. Snedaker, H.J. Teas, eds.), pp. 776-804. Gainesville: University of Florida Press 1975
Scholl, D.W.: Recent sedimentary record in mangrove swamps and rise in sea level over the southwestern coast of Florida. J. Mar. Geol. 1, 344-366 (1964)
Teas, H.J.: Mangroves of Biscayne Bay. Mimeo report to Dade County, Florida 1974
Teas, H.J., Jurgens, W., Kimball, M.C. : Plantings of red mangrove (Rhizophora mangle L.) in Charlotte and St Lucie countries, Florida. In: Proceedings of the Second Annual Conference on Restoration of Coastal Vegetation in Florida (R.R. Lewis, ed.), pp. 132-161. Tampa: Hillsborough Community College 1975
Teas, H.J., Wanless, H.R., Chardon, R.: Effects of man on the shore vegetation of Biscayne Bay. In : Biscayne Bay: Past/Pre- sent/Future (A. Thorhaug A. Volker, eds.), pp. 133-156. Coral Gables: University of Miami Press 1976
Thorn, B.G. : Mangrove ecology and deltaic geomorphology, Ta- basco, Mexico. J. Ecol. 55, 301-343 (1967)
Thorn, B.G. : Mangrove ecology from a geomorphic viewpoint. In: Proceedings of the International Symposium on the Biology and Management of Mangroves (G.E. Walsh, S.C. Snedaker and H.J. Teas, eds.), pp. 469-481. Gainesville: University of Florida Press 1975
Thorn, B.G., Wright, L.D., Coleman, J.M.: Mangrove ecology and deltaic-estaurine geomorphology: Cambridge Gulf-Ord River, Western Australia. J. Ecol. 63, 203-232 (1975)
U.S. Weather Bureau: Climatography of the U.S. No. 6(L8. Supt. Doc. U.S. Printing Office, Washington, D.C. (1972)
Wadsworth, F.H.: Growth and regeneration of white mangrove in Puerto Rico. Carib. For. 20, 5%71 (1959)
Walter, H. : Vegetation of the Earth in Relation to Climate and the Eco-physiological Conditions. Berlin-Heidelberg-New York: Springer-Verlag 1973
Wanless, H.R. : Geologic setting and recent sediments of the Bis- cayne Bay region. Florida. In: Biscayne Bay: Past/Present/Fu- ture (A. Thorbaug and A. Volker, eds.), pp. 1-32. Coral Ga- bles: University of Miami Press 1976
Watson, J.C. : Mangrove forests of the Malayan peninsula. Malay. For. Rec. 6, 1-275 (1928)
Received August 17, 1979
235